KAITAN DENGAN KAPASITAS DAN AKTIVITAS
SOURCE DAN SINK
HENI PURNAMAWATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa dalam disertasi yang berjudul Analisis
Potensi Hasil Kacang Tanahdalam Kaitan dengan Kapasitas dan Aktivitas Source
dan Sink adalah benar-benar karya saya sendiri dengan arahan komisi
pembimbing, bukan hasil jiplakan atau tiruan serta belum pernah diajukan dalam
bentuk apapun untuk memperoleh gelar program sejenis di perguruan tinggi
manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan
dan tidak diterbitkan dari penulis lain telah dituliskan dalam teks dan dicantumkan
dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Desember 2011
Heni Purnamawati
ABSTRACT
HENI PURNAMAWATI. Potential yield analysis of groundnut in term of capacity and activity of source and sink. Under the direction of ROEDHY POERWANTO as the chairman, ISKANDAR LUBIS, YUDIWANTI WAHYU E.K, and A. GHOZI MANSHURI as the member of advisory committee.
The research was conducted with the aim to get the source and sink character of groundnuts that affect yield and percentage of full pods. The study was conducted by performing two experiments. Twelve varieties released from the years 1950-2003 are used. The 12 varieties were planted on two planting seasons (2007 and 2010) in two different locations. The observations made include source and sink capacity and activities, sink strenght, harvest index and yield. In the second experiment eight varieties were used to observe the non-structural carbohydrates (TNC) (2007). Two varieties of groundnuts were used to observe the translocation of carbon by using carbon 13 (13C) (2009). Characters of source and sink that positively and directly influence pod yield/plant of groundnut were number of pod/plant, weight of 100 seed and number of gynophores at 70 DAP. Characters which positively and directly affects harvest index were stem and leaf dry weight at 42 DAP, seed yield/plant and the number of pod/plant. Source and sink characters directly and positively affects the percentage of full pods were number of pod/plant, weight of 100 seed and LAI 70 DAP. There is an indication that most of assimilate for pod filling was obtained from the photosynthetic activities during the pod filling. There were no differences between varieties with different growth patterns. There is also an indication that some varieties do remobilisation of assimilate during pod filling period. There are seven patterns of source and sink relationship in groundnut varieties, based on the source and sink characters. Based on the source and sink characters that influence pod poduction and quality, an ideotype of groundnut plant with high productivity and high filling percentage was contructed.
RINGKASAN
HENI PURNAMAWATI. Analisis Potensi Hasil Kacang Tanah dalam Kaitan dengan Kapasitas dan Aktivitas Source dan Sink. Komisi Pembimbing: ROEDHY POERWANTO (Ketua), ISKANDAR LUBIS, YUDIWANTI WAHYU E.K. dan A. GHOZI MANSHURI (Anggota).
Penelitian ini dilaksanakan dengan tujuan untuk mendapatkan karakter
source dan sink kacang tanah yang mempengaruhi produksi dan persentase polong penuh. Berdasarkan karakter source dan sink tadi kemudian disusun ideotype
tanaman kacang tanah yang berpotensi menghasilkan bobot polong tinggi dengan kualitas polong yang baik. Penelitian dilakukan dengan melakukan dua percobaan. Percobaan pertama untuk mendapatkan informasi mengenai kapasitas dan aktivitas source dan sink dengan mengamati beberapa varietas kacang tanah. Percobaan kedua dimaksudkan untuk mengetahui pergerakan aliran karbon dalam tanaman kacang tanah yang berbeda pola pertumbuhannya.
Dua belas varietas nasional yang dilepas dari tahun 1950-2003 digunakan sebagai bahan tanam dalam percobaan pertama. Keduabelas varietas ditanam pada dua musim tanam (2007 dan 2010) di dua lokasi yang berbeda. Pengamatan yang dilakukan meliputi karakter kapasitas dan aktivitas source dan sink serta kekuatan sink, indeks panen dan produksi polong dan biji. Pada percobaan kedua digunakan delapan varietas kacang tanah untuk mengamati kandungan karbohidrat non-struktural (TNC) (MT-2007). Dua varietas kacang tanah digunakan untuk mengamati translokasi karbon dengan menggunakan penjejak isotop karbon 13 (13C) (MT-2009).
Terdapat karakter-karakter source dan sink yang berpengaruh langsung positif meningkatkan hasil polong/tanaman kacang tanah yaitu jumlah polong/tanaman, bobot 100 biji dan jumlah ginofor 70 HST. Karakter-karakter
source dan sink yang berpengaruh langsung menaikkan indeks panen yaitu bobot kering batang dan daun pada 42 HST, bobot biji/tanaman dan jumlah polong/tanaman. Karakter-karakter source dan sink yang berpengaruh langsung meningkatkan persentase polong penuh yaitu jumlah polong/tanaman, bobot 100 biji dan ILD 70HST
Didapatkan bahwa asimilat untuk pengisian biji lebih banyak diperoleh dari kegiatan fotosintesis pada periode pengisian polong/biji. Tidak ditemukan adanya perbedaan antar varietas dengan pola pertumbuhan berbeda dalam memenuhi kebutuhan asimilat saat pengisian. Ada indikasi beberapa varietas melakukan remobilisasi asimilat.
Berdasarkan perbandingan karakter-karakter source dan sink didapatkan ada tujuh hubungan source-sink pada varietas kacang tanah yang diuji yaitu:
a. Source tinggi dan Sink tinggi dengan Indeks Panen rendah
b. Source tinggi dan Sink rendah dengan Indeks Panen sedang dan rendah
Berdasarkan karakter source dan sink yang berpengaruh langsung terhadap produksi polong dan kualitas polong disusun ideotype tanaman kacang tanah dengan produktivitas polong tinggi dan persentase pengisian tinggi yaitu :
a. Menghasilkan kapasitas dan aktifitas source tinggi sehingga mampu menghasilkan bahan kering yang besar dengan bobot 100 biji > 50 gram b. ILD pada fase awal generatif (42HST) mencapai 2, memasuki fase
pengisian (56HST) mencapai 3-4, dan pada akhir fase pengisian ILD mencapai 5-6. ILD pada fase-fase selanjutnya dipertahankan 5-6 untuk pemasakan polong.
c. Membentuk percabangan (maksimal 5-6 cabang) pada 42-56HST sehingga dapat menopang banyak bunga dan ginofor pada awal fase generatif. Tinggi batang utama saat panen tidak terlalu tinggi ±70cm. d. Bunga muncul serempak pada 26-28HST dan dalam waktu 2-3 hari 50%
populasi berbunga tercapai. Enam puluh persen bunga dari total bunga sudah muncul sebelum 40HST. Jumlah ginofor 10 hari setelah berbunga mencapai 15-20 ginofor untuk menjamin jumlah polong/tanaman lebih dari 20 polong pada saat panen.
e. Indeks panen tanaman mencapai ± 0.40-0.50
Kata Kunci: kapasitas dan aktivitas source dan sink, kekuatan sink, indeks panen, 13
@Hak Cipta milik Institut Pertanian Bogor, tahun 2011 Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk
kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan
laporan, penulisan kritik atau tinjauan suatu masalah; dan pengutipan
tersebut tidak merugikan kepentingan yang wajar IPB.
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
ANALISIS POTENSI HASIL KACANG TANAH DALAM
KAITAN DENGAN KAPASITAS DAN AKTIVITAS
SOURCE DAN SINK
HENI PURNAMAWATI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup :
Dr. Ir. Trikoesoemaningtyas, MSc
Dr Ir Sugiyanta, MSi
Penguji Luar Komisi pada Ujian Disertasi:
Prof. Dr. Ir. M.A. Chozin, M.Agr
Judul Disertasi : Analisis Potensi Hasil Kacang Tanah dalam Kaitan
dengan Kapasitas dan Aktivitas Source dan Sink
Nama : Heni Purnamawati
Nomor Pokok : A 361040011
Program Studi : Agronomi
Disetujui,
Komisi Pembimbing
Prof. Dr. Ir. Roedhy Poerwanto, M.Sc. Dr. Ir. Iskandar Lubis, M.S.
Ketua Anggota
Dr. Ir. Yudiwanti Wahyu E.K, M.S. Dr. Drs. A.Ghozi Manshuri, M.S.
Anggota Anggota
Diketahui,
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Institut Pertanian Bogor
Prof. Dr. Ir. Munif Ghulamahdi, M.S. Dr. Ir. Dahrulsyah M.Sc.Agr.
PRAKATA
Peningkatan produksi kacang-kacangan di Indonesia berperan penting tidak
hanya dalam mencukupi kebutuhan protein dan energi masyarakat tetapi juga
dalam meningkatkan perekonomian petani dan perekonomian Indonesia. Upaya
yang perlu dilakukan tidak hanya berupa perbaikan teknologi produksi dan
mendapatkan genotipe-genotipe baru yang dapat berproduksi tinggi, tetapi juga
perbaikan kualitas produk sehingga mampu bersaing dalam perdagangan global.
Dalam upaya meningkatkan produksi dan produktivitas kacang tanah di
Indonesia, maka dilakukan serangkaian penelitian berjudul: Analisis potensi hasil
kacang tanah dalam kaitan dengan kapasitas dan aktivitas source dan sink. Dengan
rahmat Allah SWT, penelitian dan penulisan disertasi ini dapat diselesaikan
dengan baik. Disertasi ini merupakan salah satu syarat untuk menyelesaikan studi
Program Doktor (S3) di Sekolah Pascasarjana, IPB.
Penulis mengucapkan terima kasih dan penghargaan yang sebesar-besarnya
kepada Prof. Dr. Ir. Roedhy Poerwanto, M.Sc sebagai Ketua Komisi Pembimbing,
Dr. Ir. Iskandar Lubis, MS, Dr. Ir. Yudiwanti Wahyu E.K, MS dan Dr. Drs. A.
Ghozi Manshuri, MS sebagai Anggota Komisi yang telah banyak membantu
dalam membimbing dan mengarahkan penelitian ini.
Ungkapan terima kasih juga penulis sampaikan kepada:
1. Departemen Pendidikan Nasional yang telah memberikan beasiswa BPPS
pada tahun 2004-2007.
2. Badan Penelitian dan Pengembangan Departemen Pertanian dan Direktorat
Jendral Pendidikan Tinggi yang telah mendanai penelitian ini melalui program
KKP3T tahun anggaran 2007-2009 dan Hibah Bersaing Dikti tahun anggaran
2008-2009.
3. Rektor Institut Pertanian Bogor , Wakil Rektor IPB Bidang Akademik, Dekan
dan Wakil Dekan Fakultas Pertanian IPB, Dekan Sekolah Pascasarjana IPB,
Ketua Departemen Agronomi dan Hortikultura IPB, Kepala Bagian
Laboratorium Produksi Departemen Agronomi dan Hortikultura IPB atas
pemberian ijin dan dukungannya sehingga penulis dapat menyelesaikan
4. Staf Pengajar Program Studi Agronomi Sekolah Pascasarjana IPB yang telah
memberikan ilmu dan masukan selama penulis kuliah untuk Program Doktor
di IPB.
5. Prof. Dr. Ir. Satriyas Ilyas, MSc yang telah menguji penulis pada Ujian
Prakualifikasi Program Doktor di IPB.
6. Dr. Ir. Trikoesoemaningtyas, MSc dan Dr. Ir. Sugiyanta, MSi yang telah
menguji penulis pada Ujian Tertutup Program Doktor di IPB.
7. Prof.(R.) Dr. Sumarno, M.Sc dan Prof. Dr. Ir M.A. Chozin, MAgr yang telah
menguji penulis pada Ujian Terbuka Program Doktor di IPB.
8. Staf Pengajar serta Staf Kependidikan Departemen Agronomi dan Hortikultura
IPB yang telah banyak memberikan dukungan, bantuan dan kerjasama dalam
pelaksanaan penelitian ini.
9. Uswatul Khasanah, Inne Ratnapuri, Angga Yudho, Shofiyatul Mas’udah,
Ambar Prasetyaningrum Oentari, V. Susirani Kusumaputri dan Seriulina N Br.
S. Keloko yang telah membantu penelitian penulis.
10.Kedua orang tua penulis, Bapak Ir. Soetarwi Soerowinoto, MS (almarhum)
dan Ibu Mukarti (almarhumah) yang semasa hidup beliau berdua telah
mendidik, mengenalkan penulis pada tanaman dan senantiasa mendoakan
penulis.
11.Suami tercinta, Dr. Ir. Ahmad Budiaman, M.Sc.F.Trop dan ananda Inas Suci
Rahmawati serta Anisah Dyah Rahmawati, atas segala pengertian, dorongan
dan doanya sehingga penulis dapat menyelesaikan studi Program Doktor ini.
Semoga Allah SWTmembalas budi baik yang telah diberikan, dan semoga
disertasi ini dapat memberikan manfaat dalam pengembangan pertanian,
khususnya kacang tanah di Indonesia.
Bogor, Desember 2011
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 6 April 1966 di Bogor sebagai anak kedua
Bapak Ir. Soetarwi Soerowinoto, MS dan Ibu Mukarti. Tahun 1973 penulis lulus
dari SD Negeri Pengadilan II Bogor. Sekolah Menengah diselesaikan pada tahun
1976 di SMP Negeri I Bogor, dan pada tahun 1984 di SMA Negeri II Bogor.
Sarjana Pertanian diperoleh penulis dari Institut Pertanian Bogor pada tahun 1989.
Pada bulan Oktober 1992 penulis menikah dengan Ahmad Budiaman dan
telah dikaruniai dua orang putri, Inas Suci Rahmawati dan Anisah Dyah
Rahmawati.
Penulis diterima menjadi pegawai negeri sipil pada bulan Januari tahun 1990
dan sampai saat ini bekerja sebagai staf pengajar di Departemen Agronomi dan
Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor. Pada bulan Agustus
1990 penulis mendapat beasiswa dari GTZ untuk melanjutkan pendidikan
Magister di Universitas Goettingen, Jerman dan memperoleh gelar Magister
Science Agrariarum dari Universitas tersebut pada tahun 1993.
Karya ilmiah yang merupakan bagian dari disertasi program S3 penulis yang
telah diterbitkan dalam jurnal ilmiah berjudul Akumulasi dan Distribusi Bahan
Kering pada Beberapa Kultivar Kacang Tanah (Jurnal Agronomi Indonesia tahun
2010, vol.38(2): 100-106, Terakreditasi B). Karya ilmiah yang merupakan bagian
disertasi S3 penulis juga pernah disajikan dalam bentuk poster berjudul
Karakteristik Pengisian Biji Kacang Tanah yang ditampilkan pada Seminar
Nasional Inovasi Teknologi Padi dan Palawija bagi Keberlanjutan Ketahanan
GLOSSARY
Aktifitas sink kemampuan/laju sink tersebut dalam mengumpulkan asimilat/bahan kering
Aktifitas source laju/kecepatan tanaman menghasilkan asimilat
Asimilat/ fotosintat hasil fotosintesis tanaman
Bahan kering atau Biomassa
hasil akhir akumulasi asimilat yang telah menjadi bagian struktural suatu organ tanaman
Carbon Exchange Rate
(CER)
Laju pertukaran (gas) CO2 yang masuk dan keluar daun
Determinate pola pertumbuhan tanaman dimana pertumbuhan tanaman diakhiri dengan munculnya bunga
Feeding Meningkatkan konsentrasi suatu unsur isotop yang
memasuki tubuh tanaman
Ginofor Bagian tanaman kacang tanah yang merupakan
perpanjangan dasar bunga, terbentuk setelah terjadinya penyerbukan
Ideotype Keragaan tanaman ideal
Indeterminate pola pertumbuhan tanaman dimana pertumbuhan tanaman masih terus berlangsung walaupun bunga sudah muncul.
Isotop Unsur-unsur yang memiliki nomor atom sama tetapi bobot massa atom berbeda
Kapasitas sink banyaknya bagian tanaman yang berfungsi sebagai sink
Kapasitas source banyaknya bagian tanaman yang mampu berfotosintesis
Photosynthetic Active Radiation (PAR)
Radiasi matahari yang jatuh diatas permukaan daun, yang dapat digunakan untuk fotosintesis
Polong cipo atau cipo Polong dimana saat setelah dikeringkan biji masih sangat kecil atau bentuk polong berubah mengerut
Polong penuh Polong dimana saat setelah dikeringkan biji mengisi penuh ruang dalam polong
Remobilisasi asimilat Pemindahan asimilat dari sink temporer ke sink
produktif
Source semua organ tanaman yang melakukan fotosintesis
Sink Semua organ tanaman yang tidak berfotosintesis (
non-fotosintetik) yang mendapat suplai asimilat dari source
Sink vegetatif bagian tanaman yang mendapat suplai asimilat tetapi
bukan merupakan bagian yang dipanen (contoh tunas, daun muda)
Sink reproduktif sink yang potensial untuk membentuk sink produktif (contoh bunga, ginofor)
Sink produktif sink yang dipanen (misal polong, biji)
Sink temporer sink penyimpan sementara asimilat, mampu
mengalihkan asimilat yang disimpan ke sink lain yang membutuhkan. Asimilat ini dapat digunakan untuk pengisian biji. Contoh sink temporer adalah batang dan daun
Sink terminal Sink yang tidak dapat mengalihkan asimilat kebagian
lain karena asimilat menjadi bagian strukturalnya.
Semi determinate pola pertumbuhan tanaman seperti indeterminate tetapi dengan laju pertumbuhan sedikit tertekan setelah bunga muncul
δ13
C Peningkatan rasio karbon 13 (13C) terhadap karbon 12 (12C) dalam satuan ‰
DAFTAR ISI
1.2. Tujuan Penelitian ………..
1.3. Hipotesis ……….……..
1.4. Kerangka Pemikiran ……….…… 1
2
3
3
2. TINJAUAN PUSTAKA
2.1. Produktivitas Tanaman ……….
2.1.1. Produksi Bahan kering ………...….…..
3.1. Percobaan Kapasitas Source dan Sink Pada Beberapa Varietas
Kacang tanah ………
3.2. Percobaan Translokasi Karbon Pada Dua Varietas Kacang
Tanah Menggunakan Penjejak Isotop 13C ……… 23
32
4. HASIL DAN PEMBAHASAN
4.1. Indeks Panen dan Produksi tanaman ………....
4.2.1.3. Kandungan Klorofil dan Kerapatan Stomata ……..
4.2.1.4. Tinggi Batang Utama dan Percabangan ………...
4.2.2. Aktivitas Source ………...
4.2.2.1. Laju Pertambahan Luas Daun ……….
4.2.2.2. Laju Asimilasi Bersih (LAB)………...
4.2.2.3. Laju Akumulasi Bahan Kering ………...
4.3. Sink ………...
4.3.1. Sink Reproduktif…..………...
4.3.1.1. Bunga ………...……..
4.3.1.2. Ginofor ………….………...
4.3.2. Kapasitas Sink ……….
4.3.2.1. Jumlah dan Bobot Polong …...
4.3.2.2. Bobot 100 butir ...
4.3.3. Aktivitas Sink ………..
4.3.4. Kekuatan Sink ((Sink Strength)………..………....
4.3.4.1. Partition Coefficient ...
4.3.4.2. Persentase Pengisian Polong ...
4.4. Translokasi Asimilat ……….
4.4.1. Kadar Total Non-structural Carbohydrate (TNC) ...
4.4.2. Translokasi 13C ...
4.5. Peran Source-Sink dalam Mendukung Pertumbuhan dan
Hasil Tanaman ………...……….………..…
4.5.1. Perbandingan Varietas Berdasarkan Bobot Polong
Per Tanaman ………..…
4.5.2. Perbandingan Varietas Berdasarkan Indeks Panen ……….
4.5.3. Perbandingan Varietas Berdasarkan Persentase
Polong Penuh Per Tanaman ..………...
4.5.4. Matrik Perbandingan Varietas ………....
4.6. Sidik Lintas ………...
4.6.1. Sidik Lintas Bobot Polong Per Tanaman ……….
4.6.2. Sidik Lintas Indeks Panen ………...
4.7. Pembahasan Umum ……….. 93
5. KESIMPULAN ………... 101
DAFTAR PUSTAKA ………... 103
DAFTAR TABEL
No Judul Halaman
1. Perbandingan morfologi varietas botani Arachis hypogaea …………... 18
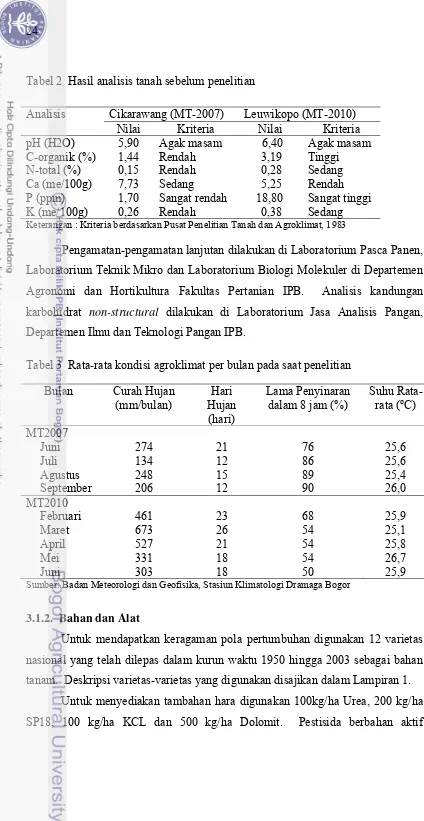
2. Hasil analisis tanah sebelum penelitian…..……….………….………... 24
3. Rata-rata kondisi agroklimat per bulan saat penelitian …..………. 24
4. Analisis ragam pada tiap musim tanam ………... 30
5. Analisis ragam gabungan dua musim tanam ……….. 30
6. Nilai indeks panen kacang tanah pada dua musim tanam ………... 35
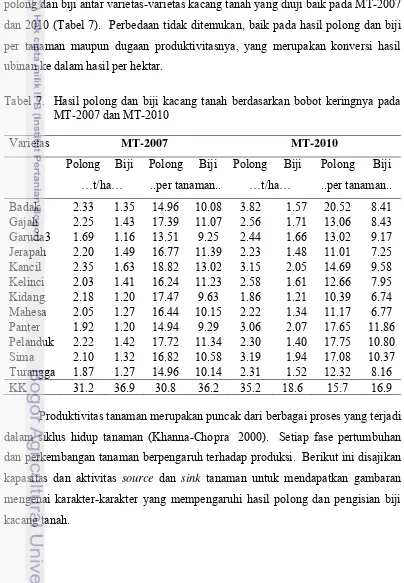
7. Hasil polong dan biji kacang tanah berdasarkan bobot keringnya
pada MT-2007 dan MT-2010………... 36
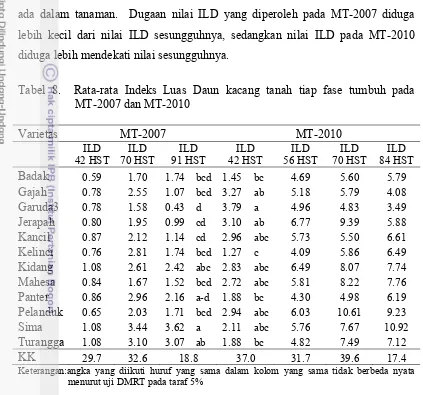
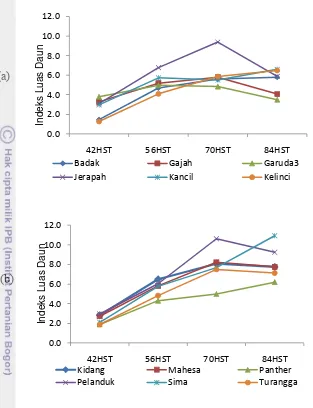
8. Rata-rata Indeks Luas Daun kacang tanah tiap fase tumbuh
pada MT-2007 dan MT-2010……….. 38
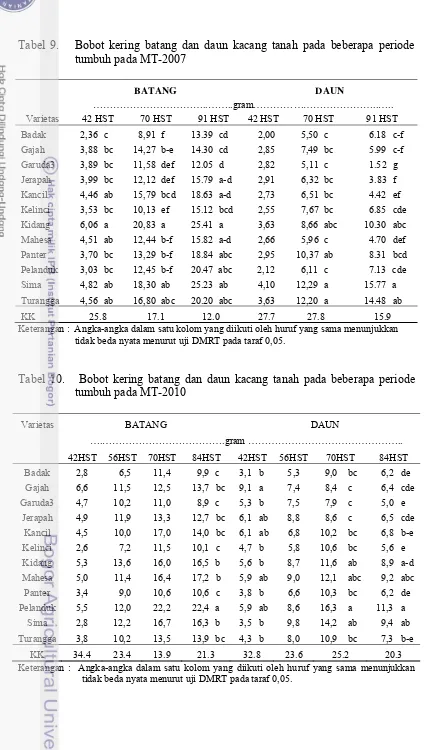
9. Bobot kering batang dan daun kacang tanah pada beberapa periode
tumbuh pada MT-2007 ……….…... 40
10. Bobot kering batang dan daun kacang tanah pada beberapa periode
tumbuh pada MT-2010 ……….... 40
11 Kadar Klorofil dan kerapatan stomata kacang tanah ….………. 43
12. Rata-rata jumlah cabang dan tinggi batang utama kacang tanah
pada MT-2010 …...………...…... 44
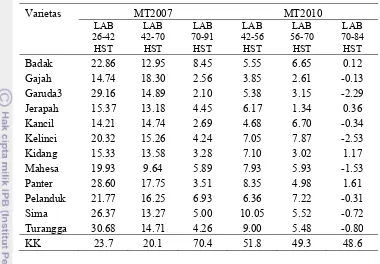
13. Rata-rata Laju Asimilasi Bersih kacang tanah pada MT-2007
dan MT-2010……….... 47
14. Laju Tumbuh Tanaman pada dua periode tumbuh kacang tanah
pada MT-2007 dan MT-2010 ………... 48
15. Waktu bunga muncul, waktu 50% populasi berbunga, periode reproduktif dan persentase polong penuh kacang
tanah pada MT-2010
………...
5016. Rata-rata jumlah bunga tiap periode, persentase bunga yang
menjadi polong dan jumlah polong per tanaman pada MT-2010 ……... 52
No Judul Halaman
18. Rataan jumlah ginofor, jumlah polong dan bobot polong per
tanaman pada MT-2007 ……….. 56
19 Rata-rata jumlah polong total, polong penuh, cipo dan persentase
polong penuh per tanaman kacang tanah pada MT-2007…………... 57
20. Rataan jumlah ginofor, jumlah polong dan bobot polong per
tanaman pada MT-2010 ……….. 58
21. Rata-rata jumlah polong total, polong penuh, polong ½ penuh, cipo, persentase polong penuh dan polong setengah penuh per
tanaman kacang tanah pada MT-2010……….……….... 59
22. Bobot 100 biji kacang tanah pada dua musim tanam ……….... 60
23. Laju Tumbuh Polong kacang tanah pada MT-2007 dan
MT-2010 ………..………... 61
24. Nilai koefisien partisi (PC) kacang tanah pada MT-2007
(PC42-panen) dan MT-2010 (PC56-panen)………. 63
25. Persentase polong penuh kacang tanah pada 2 musim tanam ………... 64
26. Kadar TNC dalam daun dan batang pada 42 dan 70 HST………... 65
27. Rata-rata hasil pengukuran kondisi umum tanaman kacang
Tanah pada fase reproduktif ……….……….…….. 68
28. Nilai 13C % atom excess dan selisih perubahannya dalam
tiap bagian tanaman ………..……….. 71
29. Rata-rata nilai ILD 42 HST, jumlah polong/tanaman,
bobot polong dan bobot biji per tanaman dari 2 MT ..……….………... 73
30. Rata-rata bobot kering tajuk (batang dan daun) 70 HST, ILD
70 HST dan Indeks Panen kacang tanah pada dua musim tanam ...……. 78
31. Rata-rata nilai bobot batang pada 42 HST, jumlah ginofor pada 70HST dan persentase polong penuh kacang tanah dari 2 musim tanam ………...
80
DAFTAR GAMBAR
No Judul Halaman
1. Alur kerangka pemikiran penelitian ……..……….. 5
2. Rak tempat feedingdengan isotop 13C. ……….…………..… 33
3. Perbandingan bobot kering polong, daun dan batang pada
(a) 70 dan 91 HST (MT-2007) dan (b) 70 dan 84 HST (MT-2010)……. 41
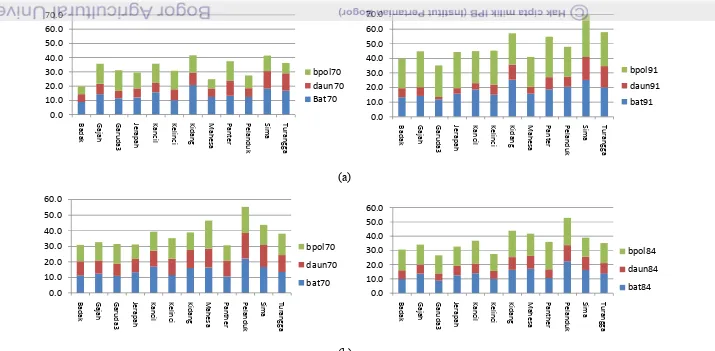
4. Pertambahan luas daun pada duabelas varietas kacang tanah (a dan b)
pada MT-2010.………...……….. 46
5. Pertambahan jumlah bunga dubelas varietas kacang tanah pada
MT-2010 ……….……….... 51
6. Pertambahan jumlah ginofor per tanaman pada MT-2007…... 53
7. Pertambahan jumlah ginofor per tanaman pada MT-2010…….……….. 54
8. Hubungan meningkatnya suhu daun (T) dengan laju CER………. 68
9. NIlai % 13C atom excess dalam tanaman kacang tanah varietas Sima
(a) dan Jerapah (b).……….………. 70
10 .
Kandungan 13C (g) dalam bagian tanaman kacang tanah varietas
Sima (a) dan Jerapah (b)…….….………... 72
11. Perbandingan varietas berdasarkan bobot dan jumlah polong per tanaman.…………...……… .
74
12 .
Perbandingan varietas berdasarkan ILD 42HST dan jumlah polong
per tanaman.………..………... 75
13 .
Perbandingan varietas berdasarkan ILD 42 HST dan bobot kering
tajuk (batang dan daun) per tanaman 42 HST………... 76
14 .
Perbandingan varietas berdasarkan bobot polong per tanaman dan
bobot biji per tanaman………... 77
15 .
Perbandingan varietas berdasarkan ILD 70 HST dan nilai
indeks panen………. 79
16 .
Perbandingan varietas berdasarkan bobot kering batang 42 HST
dan persentase polong penuh per tanaman………...……… 81
17 .
Perbandingan varietas berdasarkan jumlah ginofor 70 HST
No Judul Halaman
18 .
Karakter-karakter yang bepengaruh langsung dan tidak langsung
terhadap bobot polong/tanaman kacang tanah………..………... 88
19 .
Karakter-karakter yang berpengaruh langsung dan tidak langsung
terhadap Indeks Panen kacang tanah………..………. 90
20 .
Karakter-karakter yang berpengaruh langsung dan tidak langsung
DAFTAR LAMPIRAN
No Judul Halaman
1. Deskripsi duabelas varietas kacang tanah ………...………. 110
2. Tiga Kategori Polong ……… 116
3. Rataan bunga yang muncul pada MT-2010 dari duabelas varietas
kacang tanah ……….……… 117
4. Hasil analisis ragam karakter pengamatan pada MT-2007……… 119
5. Hasil analisis ragam karakter pengamatan pada MT-2010……… 120
6. Hasil analisis ragam gabungan MT-2007 dan MT-2010………..…….. 122
7. Rataan nilai karakter dari dua musimtanam ……….. 123
8. Korelasi antar karakter kacang tanah pada MT-2007 ……… 124
9. Korelasi antar karakter kacang tanah pada MT-2010 ……… 126
10. Korelasi antar karakter kacang tanah pada dua musim tanam ……….. 129
11. Korelasi nilai LTT dan LTP pada MT-2007 dengan bobot
polong/tanaman, persentase polong penuh dan Indeks Panen………... 131
12. Korelasi nilai LTT dan LTP pada MT-2010 dengan bobot
polong/tanaman, persentase polong penuh dan Indeks Panen ……….. 132
13. Karakter-karakter yang mempengaruhi langsung dan tidak
langsung terhadap bobot polong/tanaman kacang tanah……… 133
14. Karakter-karakter yang mempengaruhi langsung dan tidak
langsung terhadap indeks panen kacang tanah………... 134
15. Karakter-karakter yang mempengaruhi langsung dan tidak
1. PENDAHULUAN
1.1. Latar Belakang
Kacang tanah merupakan komoditas kacang-kacangan kedua yang ditanam
secara luas di Indonesia setelah kedelai. Produktivitas kacang tanah di Indonesia
tahun 1986 tercatat 0,7 ton/ha meningkat menjadi 1,1 ton/ha pada tahun 2000 dan
1,2 ton/ha pada tahun 2010 (Kasno 2004; BPS 2009). Permintaan akan kacang
tanah di Indonesia terus meningkat dari tahun ke tahun. Kenaikan permintaan ini
tidak dapat dipenuhi seluruhnya oleh produksi dalam negeri. Indonesia mulai
mengimpor kacang tanah sejak tahun 1978. Volume impor kacang tanah
meningkat sebesar 0,95% per tahun (BPS 2009), tetapi luas lahan dan produksi
sejak 2005 terus menurun.
Produktivitas tanaman kacang tanah dipengaruhi oleh agroklimat, teknik
produksi dan karakteristik varietas. Berbagai varietas baru berdaya hasil mencapai
2 - 2,5 ton/ha, bahkan lebih, telah dilepas, tetapi produktivitas petani hanya
mencapai 50 – 60 % (Kasno 2004). Kasno (2006) juga menemukan bahwa pada
MT-2004/2005 varietas lokal masih dominan ditanam petani dengan luas tanam
78,02%. Varietas Gajah, Macan dan Kelinci masing-masing luas tanamnya
10,80%, 6,54% dan 4,64%. Selain produktivitas yang rendah, masalah lain yang
sering ditemui di lapangan adalah persentase polong hampa (cipo) dan polong
yang terisi kurang maksimal yang cukup besar. Polong yang kurang terisi atau
setengah penuh menghasilkan biji keriput dan tidak mencapai kualitas yang
diharapkan pasar.
Bell dan Wright (1998) menemukan bahwa walaupun populasi tanaman
kacang tanah di Indonesia tergolong tinggi ternyata polong yang dihasilkan
banyak yang tidak berisi atau terisi tidak maksimum, yang mengakibatkan
produktivitasnya tetap dibawah 2,5 ton/ha. Hal ini dapat mengindikasikan
rendahnya partisi asimilat ke bagian yang dimanfaatkan atau dipanen. Kondisi ini
merugikan dipandang dari adanya pemborosan fotosintat/asimilat ke bagian yang
tidak produktif.
Permasalahan produktivitas dan pengisian biji/polong yang tidak optimal ini
dicoba untuk dipecahkan melalui pengamatan pada hubungan source dan sink
adalah organ tanaman yang melakukan fotosintesis, sedangkan sink adalah organ
tanaman dimana hasil fotosintesis (asimilat) disimpan. Hubungan source dan sink
pada tanaman ditentukan oleh kapasitas dan aktivitas source serta kapasitas,
aktivitas dan kompetisi diantara sink. Produksi tanaman ditentukan oleh
banyaknya akumulasi bahan kering dan partisi atau pembagian bahan kering
tersebut ke bagian yang akan dipanen. Oleh karena itu, peningkatan hasil
tanaman dapat dilakukan dengan meningkatkan akumulasi bahan kering dan/atau
meningkatkan indeks panen.
Varietas-varietas kacang tanah yang umum ditanam di Indonesia tergolong
varietas kacang tanah subtipe Spanish dan Valencia yang bertipe pertumbuhan
tegak dan umur panen berkisar 100-105 hari. Varietas-varietas kacang tanah
nasional mempunyai potensi jumlah polong diatas 15 polong per tanaman, bahkan
sejak tahun 2000 potensi varietas-varietas baru yang dilepas mencapai 20
polong/tanaman. Ginofor yang dihasilkan selama hidup tanaman dapat mencapai
lebih dari 100 ginofor, tetapi dari beberapa penelitian didapat hanya 10 – 15 %
saja yang berkembang menjadi polong berisi penuh (Lukitas 2005).
Informasi mengenai hubungan source dan sink yang berhubungan dengan
pengisian biji pada varietas-varietas kacang tanah di Indonesia masih sangat
terbatas. Pengamatan terhadap kapasitas dan aktivitas source dan sink,
hubungannya dengan hasil polong dan pengisian biji, distribusi asimilat serta
sumber asimilat untuk pengisian biji akan dapat memudahkan pemahaman
karakter-karakter penting yang berpengaruh terhadap produksi dan pengisian biji
kacang tanah serta mekanisme pembagian asimilat. Pemahaman yang lebih baik
akan karakter-karakter ini akan mampu mendorong pengembangan teknologi
budidaya tanaman kacang tanah sehingga memberikan keuntungan ekonomis
yang lebih baik, serta mendorong perbaikan genetik tanaman untuk menghasilkan
varietas kacang tanah berproduksi tinggi dengan sifat-sifat unggul yang
diharapkan pasar.
1.2. Tujuan Penelitian
Penelitian ini dilakukan dengan tujuan mendapatkan karakter source dan
Berdasarkan karakter source dan sink tadi kemudian disusun ideotype tanaman
kacang tanah.
1.3. Hipotesis
Hipotesis yang diajukan dalam penelitian ini adalah :
1. Terdapat perbedaan kapasitas dan aktivitas source dan sink pada
kultivar-kultivar kacang tanah yang mempengaruhi produksi dan pengisian polong/biji.
2. Terdapat satu atau lebih karakter source dan sink yang berpengaruh langsung
maupun tidak langsung terhadap produksi polong dan pengisian polong/biji
kacang tanah.
3. Terdapat perbedaan dalam pola translokasi asimilat pada kultivar kacang tanah
dengan pola pertumbuhan yang berbeda.
1.4. Kerangka Pemikiran
Produksi tanaman (yield) ditentukan oleh kemampuan tanaman
menghasilkan asimilat (biomassa) dan pengalokasian sebagian besar asimilat ke
bagian yang bernilai ekonomi. Total produksi biomassa tanaman tergantung pada
keseimbangan proses fotosintesis dan respirasi. Kemampuan genetik tanaman dan
kondisi lingkungan tumbuh tanaman mengendalikan kedua proses ini. Tanaman
mempunyai kemampuan berbeda dalam mengatur distribusi asimilat apabila
terjadi perubahan lingkungan tumbuh. Oleh karena itu perlu diusahakan teknik
budidaya maupun perbaikan genetik dengan memanfaatkan karakter yang
berpengaruh terhadap produksi sehingga sebanyak mungkin biomass/bahan kering
dapat dihasilkan tanaman dan sebanyak mungkin dapat ditransfer ke bagian
tanaman yang bernilai ekonomis.
Asimilat dihasilkan daun dan semua bagian tanaman yang berfotosintesis,
bagian ini disebut source. Source menghasilkan asimilat yang disamping untuk
mempertahankan metabolisme source juga untuk menyuplai kebutuhan bagian
tanaman yang non-fotosintetik. Bagian tanaman yang non-fotosintetik ini disebut
sink. Sink ada yang bersifat temporer dan/atau terminal. Sink temporer dapat
mengalihkan asimilat yang disimpannya kebagian sink lain yang membutuhkan,
Distribusi asimilat atau partisi asimilat menjadi penting dalam menentukan hasil
akhir tanaman. Banyaknya asimilat yang dihasilkan tanaman sulit untuk diukur
langsung akan tetapi dapat diukur secara tidak langsung dengan pengukuran bobot
kering yang merupakan hasil akhir akumulasi asimilat pada suatu organ tanaman.
Pada kacang tanah pembentukan bunga, ginofor dan polong yang adalah
tahap kritis yang menentukan produktivitas. Bunga, ginofor dan polong muda
merupakan sink reproduktif yang berpotensi untuk berkembang menjadi polong
yang dapat dipanen. Tiap varietas diduga mempunyai kemampuan tertentu dalam
menghasilkan sink reproduktif. Sejumlah sink reproduktif kemungkinan tidak
dapat berkembang menjadi sink produktif sebagai akibat dari pengaruh
lingkungan.
Berbeda dengan kedelai yang determinate, kacang tanah yang umumnya
dibudidayakan di Indonesia mempunyai tipe pertumbuhan semi-determinate.
Pada pola pertumbuhan yang semi-determinate, terjadi kompetisi antara sink
reproduktif dan produktif (bunga, ginofor, biji/polong) dengan sink vegetatif
(daun muda, cabang, akar, bintil akar). Kompetisi ini mempengaruhi banyaknya
asimilat untuk perkembangan sink reproduktif selanjutnya, dan apabila
kebutuhannya tidak tercukupi, mengakibatkan sebagian sink reproduktif berhenti
berkembang atau gugur. Kompetisi juga dapat mengakibatkan sink reproduktif
terlambat tumbuh dan berkembang.
Biji dapat memperoleh asimilat dari fotosintesis langsung selama fase
pengisian biji dan/atau dari remobilisasi asimilat dari sink temporer (yang berada
di daun dan batang). Mempertahankan daun hijau selama periode pengisian biji
dapat menjamin berlangsungnya fotosintesis untuk menyuplai asimilat ke biji
yang sedang berkembang. Kebutuhan asimilat untuk pengisian biji dapat pula
diperoleh dari hasil remobilisasi asimilat yang tersimpan dalam batang dan daun.
Adanya remobilisasi asimilat ini dapat menjadi merugikan karena mengakibatkan
laju fotosintesis daun terganggu dan memicu senesens (Sinclair dan deWitt 1978),
yang akibat selanjutnya menurunkan laju serapan hara akar, dan pada akhirnya
mempengaruhi pengisian biji. Gambar 1 menunjukkan alur kerangka pemikiran
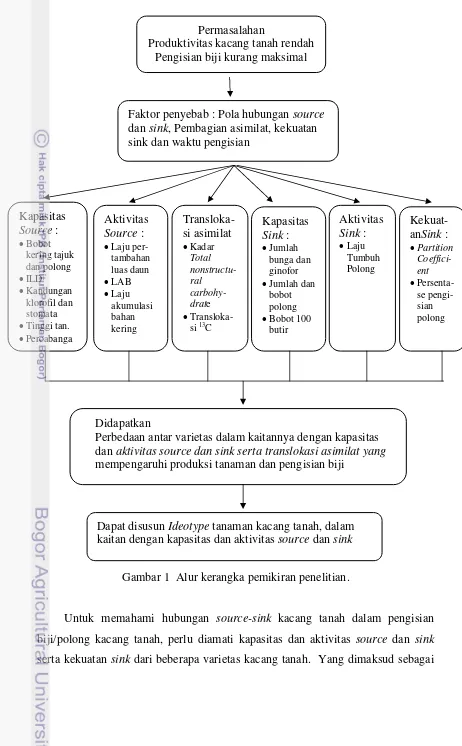
Gambar 1 Alur kerangka pemikiran penelitian.
Untuk memahami hubungan source-sink kacang tanah dalam pengisian
biji/polong kacang tanah, perlu diamati kapasitas dan aktivitas source dan sink
serta kekuatan sink dari beberapa varietas kacang tanah. Yang dimaksud sebagai
Permasalahan
Produktivitas kacang tanah rendah Pengisian biji kurang maksimal
Faktor penyebab : Pola hubungan source
dan sink, Pembagian asimilat, kekuatan sink dan waktu pengisian
Kapasitas
Perbedaan antar varietas dalam kaitannya dengan kapasitas dan aktivitas source dan sink serta translokasi asimilat yang mempengaruhi produksi tanaman dan pengisian biji
Aktivitas
Dapat disusun Ideotype tanaman kacang tanah, dalam
kapasitas source adalah banyaknya bagian tanaman yang mampu berfotosintesis,
sedangkan aktifitas source adalah laju/kecepatan tanaman menghasilkan asimilat
yang kemudian disimpan atau terukur dalam bobot keringnya. Dalam Disertasi
ini sink yang diamati hanya sink reproduktif dan sink produktif yaitu sink-sink
yang langsung berhubungan dengan produksi tanaman. Sink reproduktif adalah
sink yang kemudian dapat berkembang menjadi sink produktif, sedangkan sink
produktif adalah sink yang dipanen (polong dan biji). Kapasitas sink adalah
banyaknya bagian tanaman yang berfungsi sebagai sink reproduktif dan produktif,
sedangkan aktifitas sink adalah kemampuan/laju sink tersebut dalam
mengumpulkan asimilat/bahan kering. Persaingan untuk mendapatkan asimilat
terjadi antara sink-sink reproduktif dan produktif maupun sink reproduktif dan
produktif dengan sink-sink vegetatif. Makin kuat sink tersebut maka makin
banyak asimilat didapatkan oleh sink tersebut bahkan, kemungkinan mampu
menekan pertumbuhan dan perkembangan sink-sink lain.
Pengamatan dilakukan terhadap sebanyak mungkin varietas kacang tanah
lokal dan nasional untuk mendapatkan pola hubungan source dan sink yang
berbeda. Kondisi lingkungan tumbuh dapat mempengaruhi distribusi asimilat
yang berdampak pada produktivitas tanaman. Oleh karena itu, penelitian ini
dilakukan di dua lokasi dan tahun/musim yang berbeda untuk mendapatkan
informasi pengaruh lingkungan terhadap karakter-karakter tersebut.
Asimilat yang dihasilkan tanaman harus ditranslokasikan dari source ke
dalam sink. Perubahan dalam translokasi asimilat akan berakibat pada
ketersediaan asimilat yang dapat diakumulasikan tanaman di dalam sink produktif.
Pengamatan translokasi asimilat dilakukan terhadap translokasi karbohidrat
non-struktural pada periode pengisian biji dan penggunaan penjejak isotop karbon-13
untuk mengamati perpindahan karbon dalam varietas-varietas kacang tanah yang
memiliki pola pertumbuhan yang berbeda.
Dari pengamatan kapasitas dan aktivitas source-sink diharapkan dapat
diperoleh keragaman antar varietas kacang tanah berdasarkan karakter kapasitas
dan aktivitas source-sink serta translokasi asimilat yang mempengaruhi produksi
kemudian dilanjutkan dengan penyusunan suatu “ideotype” tanaman kacang tanah
2. TINJAUAN PUSTAKA
2.1. Produktivitas Tanaman
Produksi tanaman adalah puncak dari berbagai proses yang terjadi dalam
suatu siklus hidup tanaman (Khanna-Chopra 2000). Setiap fase pertumbuhan dan
perkembangan tanaman berpengaruh terhadap produksi. Produktivitas tanaman
(yield) ditentukan oleh kemampuan tanaman berfotosintesis dan pengalokasian
sebagian besar hasil fotosintesis ke bagian yang bernilai ekonomi.
Yoshida (1972) mengekspresikan produktivitas tanaman sebagai :
Produktivitas = Biomas x Indeks Panen.
Biomas adalah total bahan kering hasil fotosintesis yang diakumulasi
tanaman, sedangkan produktivitas adalah bagian biomas yang dipanen dan indeks
panen melambangkan koefisien partisi. Dari persamaan tersebut, produktivitas
tanaman dapat ditingkatkan dengan menghasilkan lebih banyak biomas (bahan
kering) atau meningkatkan indeks panen atau meningkatkan keduanya (Kumudini
et al. 2001). Peningkatan produktivitas pada varietas-varietas baru gandum,
jagung dan kedelai dunia lebih banyak disumbangkan dari adanya peningkatan
lebih banyak bahan kering kedalam biji atau peningkatan indeks panen (Egli
1999; Snyder dan Carlson 1983). Peningkatan indeks panen ini tidak dapat
dilakukan terus menerus karena hanya dapat dilakukan hingga batas tertentu
sehingga peningkatan selanjutnya dilakukan dengan meningkatkan akumulasi
bahan kering (Gifford dan Evans 1981).
2.1.1 Produksi Bahan Kering
Hasil fotosintesis tanaman (asimilat) diukur secara tidak langsung dengan
mengukur produksi bahan keringnya. Produksi bahan kering merupakan dasar
dari produksi tanaman. Yoshida (1972) menyatakan bahwa laju pertambahan
bahan kering atau laju pertumbuhan tanaman ditentukan oleh indeks luas daun
(ILD), tingkat fotosintesis bersih daun dan sudut daun.
Dale et al., dalam Edoka (2006) menyebutkan bahwa pertumbuhan dan
lamanya daun hijau suatu tanaman menentukan persentase radiasi matahari yang
dan hasil akhir tanaman. Indeks Luas Daun (ILD) merupakan rasio antara luas
daun yang hijau dengan luas permukaan tanah dimana tanaman tumbuh. Kiniry et
al. (2005) menggunakan nilai ILD, koefisien light extinction (k) hukum Beer dan
indeks panen untuk membandingkan produksi bahan kering (biomass) dan hasil
berbagai kultivar kacang tanah.
Produksi bahan kering sendiri merupakan perimbangan dari proses
fotosintesis dan respirasi, dimana laju pertambahan bahan kering tanaman
meningkat secara asimtot dengan peningkatan ILD sehingga hampir tidak ada
optimum ILD untuk produksi bahan kering (Yoshida 1972). Walaupun demikian
definisi titik kritis ILD dapat diartikan sebagai suatu nilai ILD dimana
peningkatan ILD tidak lagi meningkatkan laju pertumbuhan tanaman atau terjadi
peningkatan tetapi kecil. Kiniry et al. (2005) menemukan ILD kacang tanah pada
berbagai lokasi di Texas USA pada satu musim berkisar antara 5 – 6 dengan nilai
koefisien extinction cahaya (k) berkisar 0,60 – 0,65. Untuk mendapatkan
penetrasi cahaya yang lebih baik maka diharapkan daun lebih tegak dan sudut
daun kecil sehingga nilai k menjadi lebih kecil, lebih banyak daun terpapar sinar
matahari pada PAR yang lebih kecil sehingga laju pertukaran karbon (CER=
Carbon Exchange Rate) kanopi meningkat.
Banyak penelitian menunjukkan bahwa akumulasi bahan kering yang
terjadi menjelang dan saat pengisian biji amat menentukan hasil. Shiraiwa et al.
(2004) menemukan besarnya akumulasi bahan kering pada fase periode awal
pengisian biji merupakan karakteristik yang menentukan perbedaan hasil antara
genotipe-genotipe kedelai. Perbedaan hasil antara padi berdaya hasil tinggi dan
padi berdaya hasil rendah terletak pada kemampuan mengakumulasi bahan kering
sebelum heading dan translokasi asimilat selama pengisian biji (Miah et al. 1996).
Lubis et al. (2003) menyatakan bahwa berat kering padi saat pengisian biji lebih
mempengaruhi hasil daripada karbohidrat non-struktur (non structural
carbohydrate = NSC) saat berbunga penuh
Kemampuan fotosintesis dapat diamati dengan mengukur tingkat
pertukaran karbondioksida (CER) pada tajuk. Nilai CER dihitung berdasarkan
laju CO2 yang memasuki stomata. Tanaman dengan CER tinggi diharapkan
memiliki potensi produksi yang juga tinggi. Nilai CER sendiri sangat dipengaruhi
oleh konduktansi stomata (KS). Konduktansi stomata diukur dengan mengamati
jumlah CO2 yang masuk melalui hambatan stomata. Tanaman dengan nilai CER
tinggi umumnya didukung pula oleh KS yang juga tinggi (Santosa, 2000).
Semakin kecil hambatan stomata, semakin besar nilai konduktansinya sehingga
semakin banyak CO2 yang dapat masuk melalui stomata ke dalam daun. Semakin
banyak CO2 yang masuk memungkinkan terjadinya fiksasi CO2 untuk fotosintesis
yang lebih banyak.
Semakin banyak jumlah stomata, konduktansi per satuan luas daun akan
semakin tinggi (Mohr dan Schopfer 1995). Watanabe et al., dalam Evans dan
Fischer (1999) juga menemukan adanya peningkatan laju pertukaran karbon
(carbon exchange rate, CER), peningkatan N-daun, aktivitas Rubisco, kandungan
klorofil dan kapasitas transport elektron serta penurunan rasio klorofil a/b saat
membandingkan hasil beberapa kultivar gandum lama dan baru.
Terdapat korelasi yang erat antara laju fotosintesis dengan kandungan
N-daun karena enzim Rubisco merupakan 25 - 30 % dari total N-N-daun (Evans 1989).
Dalam fase pengisian terdapat dua sumber N untuk pertumbuhan biji yaitu N yang
diabsorpsi akar dan N yang berasal dari remobilisasi jaringan vegetatif (Ta dan
Weiland 1992). Penundaan remobilisasi N dari daun dapat mempertahankan
kapasitas fotosintesis lebih lama dan kemungkinan dapat meningkatkan hasil biji.
Kemampuan padi cv. Akenoshi untuk mempertahankan kandungan
N-daun yang tinggi pada periode pemasakan, sehingga laju fotosintesis tetap tinggi
pada fase tersebut, mengakibatkan pengisian biji menjadi optimal dan produksi
biji lebih tinggi dibanding padi cv Nipponbare (Ookawa et al. 2003).
Kemampuan tanaman dalam menangkap dan menggunakan radiasi cahaya
matahari untuk fotosintesis dipengaruhi pula oleh faktor morfologis, anatomis dan
fisiologis daun. Peningkatan luas daun, pengurangan trikoma, pengurangan
ketebalan daun, dan peningkatan kandungan klorofil sehingga memungkinkan
penangkapan cahaya menjadi lebih efisien (Taiz dan Zeiger 2002).
Energi cahaya yang jatuh ke daun pertama kali diserap oleh pigmen
klorofil. Tipe klorofil yang banyak terdapat dalam tanaman adalah tipe a dan b.
gugus aldehid. Fungsi kedua klorofil ini berbeda dimana klorofil b bersama
dengan pigmen lain seperti karoten berperan sebagai penangkap foton cahaya.
Foton ini kemudian diteruskan secara berantai ke klorofil a yang berfungsi sebagai
antena pusat reaksi. Rasio antara klorofil a dan b menentukan keefisienan
penangkapan cahaya. Tanaman yang tumbuh cepat dan berproduksi tinggi
membutuhkan respirasi yang juga tinggi. Dengan melakukan respirasi, energi
yang tersimpan dalam substrat organik akan dilepas dan diubah dalam bentuk
senyawa ATP yang digunakan untuk berbagai proses metabolisme. Dari proses
respirasi dapat dihasilkan banyak reduktan, berbagai substrat dan karbon skeleton
serta CO2 dan panas sebagai produk sampingannya (Taiz dan Zeiger 2003).
Pertukaran gas CO2 dan O2 dapat digunakan untuk mengukur tingkat
respirasi suatu jaringan (Moss, 1986). Pada kondisi tersedia cukup cahaya untuk
fotosintesis maka gas CO2 yang terukur pada daun (kloroplas) merupakan hasil
dari fiksasi karbon dan respirasi, sedangkan pada kondisi gelap atau cahaya
rendah maka CO2 yang terukur menunjukkan tingkat respirasi daun tersebut.
Tingkat respirasi tanaman dipengaruhi oleh faktor eksternal dan internal
(Mohr dan Schopfer 1995). Usia jaringan, adanya pelukaan merupakan contoh
faktor internal yang mempengaruhi laju respirasi. Stress air, serangan hama
penyakit sering memicu peningkatan respirasi tanaman.
Kacang tanah membutuhkan energi jauh lebih besar dalam memproduksi
polong daripada organ vegetatif (Duncan et al. 1978). Khanna-Chopra (2000)
menyatakan bahwa dari studi-studi tentang efisiensi konversi gula menjadi
karbohidrat, lipid dan protein menunjukkan bahwa untuk mensintesa lipid dan
protein dibutuhkan lebih banyak heksosa daripada untuk membentuk selulosa atau
pati. Konsekuensinya, efisiensi konversi gula menjadi bahan kering pada biji
yang kaya kandungan karbohidrat lebih tinggi daripada biji yang banyak
kandungan lipidnya. Duncan et al. (1978) dan Wright et al. (1991) menggunakan
faktor koreksi energi untuk mengukur nilai biomas kacang tanah yaitu bobot
biomas tajuk ditambah bobot polong x 1,65.
2.1.2. Distribusi Asimilat
Egli (1999) menyatakan bahwa produktivitas (yield) tanaman dibatasi oleh
yang dihasilkan source. Oleh karena itu terjadinya perubahan akumulasi bahan
kering atau perubahan indeks panen (partisi asimilat) atau keduanya, yang dapat
terjadi akibat perubahan faktor-faktor produksi, dapat mempengaruhi hasil biji.
Pembagian karbon dalam tanaman dipengaruhi oleh banyak faktor antara
lain perubahan suplai dan kebutuhan karbon selama pertumbuhan dan
perkembangan tanaman, adanya kontrol hormon atau nutrisi antar organ,
hambatan jaringan pembuluh, efek buffer dalam organ penyimpan diberbagai
lokasi dalam tanaman, laju fotosintesis (aktivitas source) dan laju penggunaan
karbon (aktivitas sink) (Wardlaw 1990). Distribusi atau partisi asimilat
dikendalikan berbagai proses mulai dari transpor sel ke sel, transfer antara xilem
dan floem, loading dan unloading dalam jaringan pembuluh, translokasi
long-distance dalam floem, hubungan jaringan pembuluh antara source dan sink
(Hendrix 2000). Distribusi asimilat menjadi penting dalam menentukan hasil
akhir tanaman.
Kekuatan sink dalam menarik asimilat berbeda-beda, sink yang kuat akan
mendapat bagian asimilat lebih cepat dan lebih banyak dibandingkan sink yang
tidak terlalu kuat. Dasar bagi kekuatan sink (sink strenght) adalah kemampuan
sink untuk secara efektif mengurangi konsentrasi asimilat dalam jaringan
pembuluh yang berhubungan dengan sink tersebut untuk menghasilkan gradien
konsentrasi yang terbaik antara source dan sink (Wardlaw 1991). Kekuatan sink
ini ditentukan oleh ukuran, aktifitas, stadia pertumbuhan, jarak sink tersebut
terhadap source dan hubungannya dengan jaringan pembuluh (Taiz and Zeiger
2003). Faktor yang paling menentukan aktifitas sink menurut Gifford dan Evans
(1981) adalah suply asimilat pada tahap ontogenik paling awal (stadia dimana
terjadi perubahan tunas vegetatif menjadi tunas generatif).
Inanaga dan Yoshihara (1997) menyatakan bahwa cabang merupakan
sumber karbon untuk bintil akar kacang tanah, sedangkan batang utama sebagai
sumber karbon bagi polong. Sebagian besar karbon untuk polong dan biji
merupakan hasil fotosintesis pada fase reproduktif.
Pada saat faktor lingkungan tumbuh terbatas, seperti kekeringan dan
naungan, maka pengaruh buruk kondisi ini diminimalisir oleh tanaman dengan
(1993) juga menyatakan kerapatan tanaman (populasi) dan ketersediaan hara
dapat mempengaruhi partisi bahan kering (Squire 1993). Perubahan alokasi
karbon (fotosintat) dalam tanaman yang mengalami stress tumbuh dapat
disebabkan adanya hambatan dalam “floem loading” sukrosa atau rendahnya
kapasitas sink (Khanna-Chopra 2000).
2.2. Hubungan Source dan Sink
Menurut definisi Snyder dan Carlson (1983), daun dan semua jaringan
tanaman yang berfotosintesis adalah source. Organ atau jaringan tanaman yang
menjadi tempat akumulasi sementara bahan kering untuk kemudian
melepaskannya kebagian yang memanfaatkan bahan kering juga termasuk source.
Bahan kering hasil fotosintesis kemudian ditranslokasikan melalui floem ke
bagian tanaman yang membutuhkannya (sink). Sink menggunakan asimilat untuk
pertumbuhannya dan sebagian lagi untuk disimpan.
Sink merupakan semua bagian tanaman yang tidak berfotosintesis atau ber
fotosintesis tetapi tidak maksimum sehingga sebagian kebutuhan karbohidratnya
disediakan oleh source (Taiz dan Zeiger 2003). Sink dapat berupa jaringan
meristematik, jaringan yang sedang mengalami pemanjangan, “respiratory sink”
dan jaringan penyimpanan (storage sink) (Gifford dan Evans 1981). Antara
sink-sink yang ada akan saling berkompetisi dalam mendapatkan asimilat yang
dihasilkan source.
Sink dapat dibagi menjadi sink vegetatif dan sink reproduktif. Sink
vegetatif ada yang bersifat temporer dan ada yang bersifat terminal, sedangkan
sink reproduktif adalah sink terminal. Sink temporer artinya asimilat yang
disimpan dapat dialihkan ke bagian sink lain apabila dibutuhkan, sedangkan sink
terminal berarti asimilat tidak dapat diremobilisasi dari bagian ini karena menjadi
bagian struktural.
2.2.1. Kekuatan Sink
Hubungan antara kapasitas source dari bagian atas daun aktif dan kapasitas
sink mempengaruhi produksi bahan kering dan menentukan produksi padi (Kato
menentukan laju fotosintesis, disamping faktor lingkungan (Gifford dan Evans
1981). Setelah tajuk berkembang penuh, CER masih dapat meningkat atau
menurun sejalan dengan perubahan kebutuhan sink. Apabila sink kuat menyerap
asimilat mengakibatkan gradien karbohidrat antara source dan sink makin tinggi,
hal ini merangsang source untuk lebih produktif. Akan tetapi apabila biji/buah
yang ada tidak terlalu kuat, asimilat akan lebih banyak dialokasikan kebagian lain
yang akhirnya dapat mengakibatkan aborsi (bunga, buah/polong). Apabila sink
berkompetisi dengan daun/source untuk nitrogen maka hal ini akan mendorong
penurunan CER dan senesens daun.
Pada awal pertumbuhan vegetatif daun muda dan akar merupakan sink
utama dimana pada sebagian tanaman, tajuk lebih mendominasi akar dalam
memperoleh asimilat. Pada fase reproduktif pertumbuhan dan perkembangan
buah dan biji mendominasi pertumbuhan tajuk (Wardlaw 1991). Hasil polong
merupakan hasil akhir dari proses-proses yang berlanjut sejak pembentukan
bunga, inisiasi ginofor, perubahan ginofor menjadi polong dan pengisian polong
(Songsri et al. 2008). Adanya bunga pada fase pembentukan dan pengisian biji
menjadi pesaing kuat bagi biji pada kondisi source terbatas.
2.2.2. Translokasi Asimilat
Pada prinsipnya asimilat yang ditranslokasikan dari source ke sink adalah
karbon dan nitrogen (Atkins dan Smith 2007). Hara K memang bukan
pembentuk senyawa organik dalam tanaman tetapi unsur K sangat penting dalam
proses pembentukan biji kacang tanah bersama hara P disamping juga penting
sebagai pengatur berbagai mekanisme dalam proses metabolik seperti fotosintesis,
transportasi hara dari akar ke daun, translokasi asimlat dari daun ke seluruh
jaringan tanaman (Sumarno 1986). Kalium berperan penting dalam translokasi
asimilat baik dalam phloem loading maupun dalam aliran asimilat dari source ke
sink (Marschner 1995). Penelitian yang telah dilakukan pada castorbean
menunjukkan bahwa banyaknya fotosintat yang ditranslokasikan dipengaruhi oleh
suplay K+ yaitu, kandungan K+ yang lebih tinggi memberikan hasil fotosintesis
mempengaruhi kapasitas source sink dengan mempengaruhi transpor floem
(Mengel 1996)
Pada tanaman leguminose, tanaman yang kekurangan kalium lebih peka
terhadap penyakit dan menunjukkan kualitas produksi yang rendah karena biji
yang dihasilkan banyak yang keriput (Leiwakabessy dan Sutandi 2004). Ispandi
(2004) menyatakan bahwa pada lahan kering alfisol pemupukan 100 kg KCl/ha
meningkatkan hasil kacang tanah secara nyata daripada yang dipupuk 50
kgKCl/ha.
Umumnya jenis karbohidrat yang ditranslokasikan dalam jaringan
pembuluh adalah jenis gula non-reduksi (nonreducing sugars) karena substrat ini
tidak sereaktif gula reduksi/pereduksi (reducing sugars). Gula reduksi/pereduksi
merupakan gula dengan gugus keton atau aldehid. Sukrosa adalah jenis gula
non-reduksi yang umumnya ditranslokasikan dalam tanaman. Beberapa tanaman ada
yang mentranslokasikan raffinosa, stachyosa, verbascosa, manitol dan sorbitol
(Taiz dan Zeiger 2002). Zimmermann dan Ziegler (1975) menyatakan sukrosa
adalah gula dominan pada tanaman legum tetapi Zheng et al. (2001) menemukan
bahwa bentuk fotosintat yang diekspor daun kacang tanah adalah fruktosa,
sukrosa mungkin disintesis di batang, akar dan polong.
Translokasi fotosintat dari sumber (source) ke pengguna (sink) diatur oleh
senyawa pengendali pertumbuhan tanaman yang disebut plant growth substances,
jika senyawa buatan yang diberikan secara eksogen disebut plant growth
regulator (Sumardi et al. 2007). Paclobutrazol secara signifikan mampu
mempengaruhi karakteristik fotosintesis dan anatomi tanaman Catharanthus
roseus (L.) G. Don, meningkatkan kandungan klorofil dan parameter yang
berhubungan dengan fotosintesis seperti laju fotosintesis bersih dan konsentrasi
CO2 internal tanaman dan mengurangi transpirasi (Jaleed et al. 2007).
Paclobutrazol juga meningkatkan ketebalan daun, epidermis dan kutikula,
jaringan palisade dan jaringan bunga karang tetapi mengurangi diameter xylem.
Paclobutrazol cenderung meningkatkan fotosintesis daun (Gonzales et al. 2003).
Senoo dan Isoda (2003) menemukan bahwa aplikasi 100 dan 200 ppm
paclobutrazol pada fase awal pembentukan polong dan fase awal pengisian biji
Pada awal pertumbuhannya kandungan pati, fruktosa dan glukosa pada
daun dan batang menurun cepat karena pertumbuhan organ-organ vegetatif dan
pembentukan polong. Pada fase pembesaran polong kandungan ketiganya tetap
tinggi di daun dan batang menunjukkan bahwa tidak semua karbohidrat yang ada
digunakan untuk pembentukan biji (Zheng et al. 2001).
Inanaga dan Yoshihara (1997) menemukan bahwa cabang merupakan
sumber karbohidrat untuk akar dan bintil akar, sedangkan batang utama adalah
sumber karbohidrat untuk pengisian biji. Mereka juga menemukan bahwa
sebagian besar sumber karbon untuk pertumbuhan buah kacang tanah tergantung
pada fotoasimilat saat fase reproduktif.
Kacang tanah diduga tidak memiliki kemampuan untuk
meretranslokasikan asimilat sehingga produksi bahan kering pada saat pengisian
biji merupakan total source untuk pertumbuhan biji (Duncan et al. 1978). Oleh
karena itu kacang tanah membutuhkan intersepsi cahaya (yang diartikan sebagai
mempertahankan luasan daun) dan fotosintesis untuk mengisi biji (Chapman et al.
1993).
2.3. Kacang Tanah (Arachis hypogaea L.)
2.3.1. Botani dan Morfologi
Kacang tanah yang banyak dibudidayakan di dunia tergolong dalam
spesies Arachis hypogaea dari famili Fabaceae. Spesies Arachis hypogaea terdiri
dari dua subspesies utama yaitu ssp. hypogaea dan ssp. fastigiata. Masing-masing
subspesies memiliki dua varietas botani (Singh dan Oswalt 1995). Empat tipe
spesies kacang tanah budidaya tersebut adalah :
1. Arachis hypogaeahypogaea hypogaea Linn.
2. Arachis hypogaea hypogaea hirsuta Kohler.
3. Arachis hypogaea fastigiata fastigiata Waldron.
4. Arachis hypogaeafastigiata vulgaris Harz.
Varietas kacang tanah yang dibudidayakan di Indonesia umumnya
tergolong ataupun merupakan hasil persilangan antar ssp. hypogaea (tipe
Virginia), fastigiata (tipe Valencia) dan vulgaris (tipe Spanish). A.hypogaea ssp
tanah berbeda dalam morfologi dan fenologi. Tipe Spanish menghasilkan polong
lebih cepat dan serempak dibandingkan tipe Virginia. Pola percabangan dan
keserempakan tumbuh menentukan respon varietas terhadap periode defisit air
(Chapman et al. 1993).
Perbandingan morfologi dari ketiga varietas botani utama dari kacang
tanah dapat dilihat pada tabel berikut (Gibbons et al. 1972 ; Rao 1988):
Tabel 1 Perbandingan morfologi varietas botani Arachis hypogaea
Karakter hypogaea
Tipe pertumbuhan Procumbent
(sedikit rebah)
Erect (tegak) Erect (tegak)
Percabangan dari
Pembungaan Satu buku satu
bunga
Keterangan* N+1 cabang tumbuh dari batang utama (cabang primer), N+2 cabang tumbuh dari cabang primer (sekunder), N+3 cabang tumbuh dari cabang sekunder (tersier)
Daun kacang tanah terdiri dari 4 hingga 5 anak daun. Frekueni stomata
atas dan bawah sebanding ± 300-400 stomata per mm2. Indeks Luas Daun kanopi
kacang tanah dapat mencapai maksimum nilai 4 – 7 pada awal periode pengisian
polong. Mc Cloud et al. (1980) menyatakan bahwa tanaman kacang tanah dapat
meng-intersepsi 95% cahaya matahari pada saat ILD mencapai 3. Hal ini berarti
laju pertumbuhan tanaman mungkin mencapai maksimal pada ILD 3 dan kenaikan
ILD tidak dapat meningkatkan atau hanya meningkat sedikit pada ILD > 3
(Yoshida 1972).
Laju pertukaran CO2 per luasan lahan per hari (CER) pada saat kanopi
berkembang penuh rata-rata mencapai 6 – 8 g CO2/m2/jam (Boote et al. 1980).
CER meningkat dengan makin berkembangnya daun. Daun pada buku ketiga
menghasilkan laju CER lebih tinggi daripada daun yang tumbuh pada buku-buku
dibawahnya. Maksimum CER tercapai saat ukuran daun masimum yang tercapai
Laju peningkatan berat kering tanaman (CGR) mencapai maksimum ±
pada 60 – 90 HST dengan rata-rata nilai CGR 19,6 ± 4,2 g/m2/hari (Ketring et al.
1982). Pertambahan berat kering daun dan batang meningkat mengikuti kurva
sigmoid hingga maksimum pada 90 – 100 HST. Berat kering akar terhadap berat
kering tanaman hanya signifikan pada 2 minggu pertama pertumbuhan. Pada 80
HST berat akar hanya 2-4% dari berat kering tanaman.
Bunga kacang tanah berwarna kuning dan merupakan bunga sempurna.
Bunga muncul pada 25 – 30 HST, jumlahnya mencapai maksimum 2 – 4 minggu
kemudian untuk kemudian menurun pada periode pengisian biji. Pada bagian
pangkal bunga terdapat ovari (bakal biji). Setelah terjadi penyerbukan dari dasar
bunga muncul suatu struktur yang disebut ginofor. Ginofor tumbuh memanjang
secara “geonasti”menembus permukaan tanah. Pada kedalaman ± 3 – 8 cm dari
permukaan tanah ginofor berhenti tumbuh, mulai menyerap air dan hara, dan
mengembang membentuk polong. Pada kondisi awal bobot kering polong lebih
didominasi oleh kulit polong. Biji mulai berkembang setelah polong mencapai
ukuran maksimumnya.
Pertambahan jumlah polong awalnya lambat untuk kemudian meningkat
secara linier. Menurut Ketring et al. (1982), laju pertambahan berat kering polong
(PGR) rata-rata dapat mencapai 8,3 ± 2,1 g/m2/hari.
2.3.2. Tahapan Pertumbuhan Kacang Tanah
Kacang tanah mampu menghasilkan 100 – 200 bunga/tanaman (Oentari
2008), tetapi ternyata hanya ± 19 % saja yang berkembang menjadi polong. Pada
saat panen sebagian besar polong berada dalam berbagai tahapan dan ukurannya
terlalu kecil untuk dipasarkan. Periode pertumbuhan kacang tanah secara garis
besar dapat dibagi menjadi beberapa fase pertumbuhan yang saling overlapping
yaitu fase perkecambahan, fase pertumbuhan vegetatif, fase awal berbunga, fase
pembentukan ginofor, fase pembentukan polong, fase pengisian biji, fase
pemasakan (Maria 2000).
Famili kacangan (Leguminosae) merupakan penimbun protein dan lignin
dalam bijinya sehingga peningkatan hasilnya berhubungan dengan perbaikan laju
menyatakan legum mempunyai sifat “self destruktif” saat sebagian besar N harus
ditranslokasikan untuk memenuhi kebutuhan perkembangan biji. Hal ini juga
akan memperpendek periode pengisian biji. Kacang tanah termasuk tanaman
dengan respon plastis terhadap lingkungan. Suatu genotipe plastis
menguntungkan saat kondisi stress karena mampu berproduksi walaupun total
bahan kering sangat sedikit, tapi saat kondisi optimum kebutuhan organ vegetatif
akan bahan kering menguat (Squire 1993).
Pertumbuhan kacang tanah dapat dibagi menjadi fase vegetatif dan fase
reproduktif. Fase vegetatif dibagi menjadi 3 stadia yaitu perkecambahan,
pembukaan kotiledon dan perkembangan daun hingga awal pembungaan
(26-30HST). Fase reproduktif dibagi menjadi 9 stadia yaitu stadia pembungaan (R1),
stadia pembentukan ginofor (R2), stadia pembentukan polong (R3), stadia ukuran
polong penuh/maksimum (R4), stadia pembentukan biji (R5), stadia biji penuh
(R6), stadia pemasakan biji (R7), stadia masak panen (R8) dan stadia polong
lewat masak (R9) (Trustinah 1993).
2.3.3. Variasi Hasil
Produktivitas tanaman dapat meningkat dengan memproduksi lebih
banyak buah per unit area atau memperbesar ukuran buah. Pada kacang tanah
jumlah polong dan ukuran polong (boot 100 biji) adalah faktor penentu hasil
(Howell, 2001). Karakter jumlah polong amat dipengaruhi oleh lingkungan dan
manajemen sedangkan ukuran buah dipengaruhi oleh sifat genetik (Cahaner dan
Ashri 1974).
Jumlah polong merupakan fungsi dari faktor lingkungan dan manajemen
termasuk populasi tanaman, jarak tanam, produksi bunga dan ginofor (Howell
2001). Kondisi yang dapat meningkatkan jumlah bunga pada awal periode
pembungaan dapat mempengaruhi hasil. Akan tetapi masalahnya adalah
rendahnya persentase bunga yang menjadi polong dan jumlah bunga yang
dihasilkan tiap-tiap hari sangat bervariasi sehingga banyaknya bunga yang
dihasilkan tidak banyak mempengaruhi hasil.
Jumlah ginofor tinggi juga tidak menjamin jumlah polong tinggi karena
malam hari. Maksimum pembentukan polong terjadi pada suhu 30 – 33oC sama
seperti CGR.
Duncan et al. (1978) menerangkan bahwa adanya variasi hasil antara
empat kultivar kacang tanah di Amerika Serikat dengan tipe pertumbuhan
merambat (runner) dan tegak ditentukan oleh tiga proses fisiologi yaitu
pembagian asimilat antara bagian vegetatif dan reproduktif (faktor partisi),
lamanya periode pengisian biji dan kecepatan pembentukan polong. Makin besar
faktor partisi, makin lama periode pengisian biji, dan semakin cepat serta
serempaknya pembentukan polong maka makin tinggi hasil yang ditunjukkan
kultivar tersebut. Ketring et al. (1982) menyatakan bahwa karakteristik tanaman
kacang tanah yang menentukan hasil adalah jumlah polong yang terbentuk, partisi
asimilat selama periode pengisian polong, dan lamanya pengisian polong.
Duncan et al. (1978) menyatakan bahwa karakter fisiologi yang
menentukan hasil kacang tanah adalah jumlah polong, distribusi asimilat ke
polong tinggi sehingga nilai PGR (Pod Growth Rate) tinggi, dan lamanya periode
pengisian biji. Kultivar kacang tanah dengan peningkatan jumlah polong tertinggi
memberikan hasil polong tertinggi. Dengan tidak adanya perbedaan laju
peningkatan berat kering tanaman, sehingga fotosintesis potensial diasumsikan
relatif sama maka pembagian asimilat yang banyak ke dalam polong dapat
mempengaruhi hasil. Peningkatan berat kering polong (PGR) yang relatif sama
dapat meningkatkan hasil secara nyata dengan periode pengisian yang lebih lama.
Jumlah polong per unit area menentukan perbedaan hasil polong antar
genotipe kacang tanah dan perbedaaan ini dipengaruhi oleh Crop Growth Rate
pada fase R6 – R7 (Phakamas et al. 2008). Proses produksi bahan kering
bervariasi tergantung pada genotipe, kondisi lingkungan dan teknik budidaya yang
dilakukan. Howell (2001) menyatakan bahwa jumlah polong merupakan faktor
penentu hasil kacang tanah yang sangat dipengaruhi oleh lingkungan (iklim dan
manajemen) sedangkan ukuran polong lebih dipengaruhi sifat genetik. Perubahan
alokasi karbon (fotosintat) dalam tanaman yang mengalami stress tumbuh dapat
disebabkan adanya hambatan dalam “floem loading” sukrosa atau rendahnya
kapasitas sink (Khanna-Chopra 2000). Pemahaman tentang perbedaan produksi
penting dalam upaya mengembangkan kultivar berdaya hasil tinggi yang stabil