TUMBUFIAN INANG, PARASITOID
DAN
HPERPARASITOID
KUTUDAUN
MYZUS PEMICAE
(SULZER) (HOMOPTERA:
APHIDIDAE) DI
SEKITAR BOGOR DAN CIANJtTR,
JAWA
BARAT
SEKOLAH
PASCASARJANA
ABSTRAK
CHANDRA
IRS
AN.
Twnbuhan Inang, Parasi toid dan Hiperparasitoid Kutudaun&zlrspersicae (Sulzer) (Hamoptera: Aphididae) di Sekitar Bogor dan Cianjur, Jaws Barat. Dibimbing oleh UTOMO KARTOSUWONDO sebagai ketua komisi pembimbing, dan SYAFRIDA MANUWOTO, SOEMARTQNO SOSROMARSONO,
NINA MARYANA serta HERMANU TRIWIDODO berturut-tmt sebagai anggota komisi pembimbing.
Kutudaun A@us persicae (Sulzer) merupakan spesies yang kosmopol i t, polifag, di daerah tropis berkembangbiak partenogenetik dan siklus hidupnya pendek
sem &pat berperan sebagai hama dan vektor penyakit virus tanaman. Predator, parasitoid dan patogen merupakan musuh alami penting kutudaun. Secara mum penelitian ini bertujuan mempelajari tumbuhan inang, parasitoid dan hiperparasitoid
kutudaun
M.
persicae di sekitar Bogor dan Cianjur. Tujuan khusus penelitian ini I Jahmem peIajari keanekaragarnan t umbuhan inang, parasitoid, hi perpami toid dan sebaran vertikalnya serta penggunaan insekti sida terhadap parasi toid dan hlperparasitoid pada kutudaun
M
persicae. Penelitian laboratorium dilakukan untuk mernpelajari potensi parasitoid sebagai agens hayati pengendali kutudaun. Hasil penelitian menunjukkan bahwa keanekaragaman spesies tumbuhan inangM.
persicae tinggi, dan tumbuhan yang berkerabat secara taksonomi berpotensi sebagai inang M. persicae. Tumbuhan inang yang dominan tergolong ke &lam fami 1 i Solanaceae dan Brassicaceae. Ada enam spesies parasi toid yang memarasi t M. persicae, yaitu Aphiditis rosae Haliday,Aphidius delicat us Baker, Aph idius sp., DiaeretieIla r a p e (M' intosh) (Aphidi idae), Aphelinus asychis (Walker), dm Aphelinw sp. (Aphelinidae). Empat spesies hyperparasitoid ditemukan memarasit parasitoid tersebut, yaitu Aphidenqrfus sp., Coagem sp., Ooencyrtw sp. (Encyrtidae), dan Ckarps sp. (C hari pidae). Kelimpahan populasi dan persebaran parasitoid di pertanaman di pengaruhi oleh spesies tumbuhan inang, kerapatan populasi dan persebaran kutudaun, musirn t a m dan penggunaan insektisida. Persebaran verti kal parasitoid dan hiperparasi toid berbeda-beda di pengaruh oleh keberadaan tumbuhan inang dan kutudaun serta ketinggian tempat di atas p e m b laut. Penelitian laboratoriurn menunjukkan bahwa D. r a p e dapat menekan populasi
M.
persicae m p a i 62,7% dan Aphelinus sp. menekan populasi Mpersicae sampai 59,4%. Aphelinus sp. mematikan kutudaun melalui parrtsitisasi dan
host feeding. Secam umum parasitoid berpotensi dij adi kan agens hay ati pengendd i kutudaun.
Untuk
penerapmy a masih di perlukan bany ak penelitian khususny a rnengenai bioekologi, pembiakan masal dan teknik aplikasinya.ABSTRACT
CHANDRA IXSAN. Host Plants, Pamitoids and Hyperparasitoids of Green Peach Aphid, Myzus persicae (Sulzer) (Hornoptem: Aphididae) in the neighburhood of Bogor and Cimjur (West Java). Under the direction of UTOMO
KARTOSU
WONDO as the chairman of the advisory committee, and SYAFRIDA MANUWOTO,SOEMARTONO SOSROMARSONO,
NINA MARYANA and HERMANU TRIWIDODO as a the committee members.The green peach aphid Wtls persicae (Sulzer) is truly a cosmopolitan species, very poIyphagous, reproduces parthenogenetically in the tropics, and has a short life cycle. It is a common pest of many cultivated piants and act as vectors of many plant viruses. The predators, parasitoids and pathogens are the important natural enemies of green peach aphid and other aphids. In general this r e m h investigates the natural host plants, parasitoids, and hyperparasitoids of green peach aphid in the neighbourhood of Bogor and Cianjur (West Java). The special purpose of this resarch is to study the diversity of the host plants, the parasitoids, and hyperparasitoids of the aphid and their vertical distributions in the areas. Besides, the influence of the diversity of host crop plants and the application of insecticide on the aphid parasitoids and hrperparasitoids were also studied. Labot~tory experiments were conducted to investigate the potential of parasitoids as agent of biological control. The results showed that the aphids' host plant diversity is high, and that plant species which are taxonomically related are potential hosts of the aphld. Dominant host plants belong to families Solanaceae and Brassicaceae. There were six species of parasitoids attacking the aphid, i,e. : Aphidius rosae Halida y, Aphidius delicat ers Baker, Aphiditrs sp.,
Diaeretiella r a p e (M' intosh) (Aphidiidae), Aphelinus asychis (Walker), and
Apkelinus sp. (Aphelinidae). Four species of hyperpamitoids found attacking those parasitoids, i-e. : Aphtdencyrtus sp., Coagerus sp., Ooemyrtus sp. (Emyrtidae), and Charips sp. (Chanpidae). Population abundance and d~stribution of the parasitoids of the aphid were influenced by the crops plant species, population density and distribution of the aphid, season, and application of insecticide. The species vertical distribution of the aphids' parasitoids and h yperparasi toids differ and apparently are influenced by the occurrence of the aphids' host plant and elevation above sea level. Laboratory experiment showed that the parasitoids D. r a p e , could suppressed the aphids' population to 62,7%, and the other pamitoid, Aphelinm sp. to 59,4%.
The
last mentioned parasitoids killed its host by host feeding, besides parasitization. The use of those parasitoids in applied biological control programs is recommended, however more researches are still needed, especially in aspectsof
field biaecology, mass rearing and release technologies.SURAT
PERNYATAAN
Saya menyatakan dengan sebenar-benamya bahwa segala pernyataan &lam & s e m i
saya yang be j udul :
TUMBUHAN INANG, PARASITOID DAN HIPERPARASITOID KUTUDAUN
MYZUS PERSICAE (SULZER) (HOMOPTERA: APHIDIDAE) DI SEKITAR
BOGOR DAN CIANJUR, JAWA BARAT
merupakan gagasan atas hasil penelitian saya sendiri dengan bimbingan komisi pembim bing, kecual i yang dengan jelas dinyatakan ruj ukannya.
Disertasi ini belum pemah diajukan untuk memperoleh gelar pada program sejenis di perguruan tinggi manapun.
Semua data clan informasi yang digunakan telah dinyatakan secara jelas dan dapat diperi ksa kebenarannya.
Bogor, Nopember 2004
Nama : Chandra Irsan
TUMBUHAN
INANG,
PARASITOID DAN
HIPERPARASITOID
KUTUDAUN
MYZUS PERSICAE (SULZER)
(HOMOPTERA:
APHIDIDAE) DI SEKITAR
80GOR
DAN
CIANJUR,
JAWA BARAT
Oleh
CHANDRA IRSAN
Disertasi
Sebagai salah satu syarat untuk memperoIeh gelar Doktor Pada Sekolah Pascasarjana, Institut Pertanian Bogor
SEKOLAH
PASCASARJANA
INSTITUT
PERTANIAN BOGOR
Judul Disertasi : Tumbuhan Inang, Paraltoid dm Hiperparasitoid Kutudaun Myzus persicae ( Sdzer) (Hornoptera: Aphididae) di Sekitar Bogor dan Cirtnjur, Jawa Bamt
Nomor Pokok : 995069
Program Studi : Entomologi-Fitopatologi
1. Komisi Pembimbing
Prof.
Dr.
Ir. Utomo Kartosuwondo, M.S.Ketua
n
2. Ketua Program Studi Sekolah Pascasarjana
Entornolog-Fitoptologi
&&1&
Dr.
Ir. Sri Hendrastuti Hidayat, M..
Syafrida Manuwoto, M. Sc.Penuiis dilahirkan di Lubuk Linggau, Sumatera Selatan pada tanggal 19 Februari 1965 sebagai anak ke-enarn dari delapan bersaudara dari Ibu Hj. Zainiar dan Ayah
H.
Widji Amusis (Alm.). Penulis menyelesaikan pendidikan SD pada tahun1976 di
SD
M h m a d i y a h Lubuk Linggau, S M F tahun 1980 di SMP Muhammadiyah Lubuk Linggau dan SMA tahun 1983 di SMA Negeri I Lubuk Linggau.Pada tahun 1988 penulis memperoleh gelar Sajana Pertanian pa& Fakuttas Pertman, Universitas Sriwijaya, Palemhang. Pada tahun 1997, penulis memperoleh gelar Magister Sains Program Studi Entomologi-Fitopatologi, Sekolah Pascasarjana, Insti tut Pertanian Bogor. Pa& tahun 1 999 penul is mendapat kesempatan melanj utkan pendidikan Program Doktor di program studi dan perguruan tinggi yang sama dengan bessiswa yang sama, yaitu dari Departemen Pendidikan Nasional melalui BPPS (Beasiswa Program Pascasa jam).
Sejak tahun 1989 sampai s e k m g penulis kkerja sebagai staf pengajar di Jurusan Hama dan Penyaki t
Turn
buhan, Fakul tas Pertanian, Universi tas Sriwij aya.Alhamdulillah saya panjatkan ke hadirat Allah SWT atas &at, taufik dan hidayah-Nya sehingga saya dapat meny elesaikan peneli tian dm penulisan disertasi. Disertasi ini adalah lapran hasil penelitian dengan judul "Tumbuhan hang, Parasitoid clan Hiperparasitoid Kutudaun W z t r s persicae (Sulzer) (Hornoptera: Aphididae) di Sekitar Bogor clan Cianjur" yang merupakan salah satu syarat untuk memperoleh gelar Doktor di Sekolah Pascasa rj ana, Insti tut Pertanian Bogor.
Ucapan terirna kasih disampaikan kepada Prof. Dr. Ir. Utomo Kartosuwondo,
M.S.
sebagai ketua komisi pembimbing, Prof. Dr. Ir. Syafrida Manuwoto, M.Sc., Prof. Dr. Ir. Soemartono Sosromarsono, M.Sc., Dr. Ir. Nina Maryana, M.Si. dan Dr. Ir. Hermanu Triwidodo, M.Sc, masing-masing sebagai anggota komisi pembimbing yang telah banyak memberikan pengarahan, bimbingan, saran, motivasi, serta bantuandengan penuh keikhlasan selarna pelaksanaan penelitian dan penulisan diserhsi. Kepada kedua orang hta tercinta, Ayahanda H. Widji Anarsis ( A h ) dan bunda
Hj.
Zainiar Arief, bapak clan ibu mertua yang terhormat, Bapak Drs. H. M.Y. Effendy Hasibuan, SH. dan lbu Rr Hj. Sri Hartini, istriku tersayang Eveltine Yusnilanniari S.P., dan anak-anakku tercinta beserta semua saudara-saudaraku yang kuci ntai di sampaikan terima kasih yang &lam karena berkat doa dan dukungan merekalah saya dapat menyelesai kan pendidi kan di Sekolah Pascasarjana lnsti tut Pertanian Bogor. Semoga AfIah S WT memberi balasan amal baik mereka dengan kebaikan yang tak terhingga.Ucapan terirna kasih saya sarnpaikan kepada Kakanda Mahmud
HM
dan Fauzia Anggraini, Bapak Ir.H.
Budiman Anef, Bapak Ir. H. Ibnu Amin, M.Si. serta semua pihak yang telah membantu kelancaran studi saya, mdai dari pendanaan sarnpai kepada pelaksanaan penelitian dan selesainya studi doktor saya ini. Kepada BapakMuhammad Tris, terirna kasih atas semua bantuan dan fasilitas pelaksanaan penelitian di lapangan dan kepada Ibu Aisyah juga saya ucapkan terima kasih atas bantuannya &lam pelaksanaan penelitian di laboratorium. Semoga Allah SWT memberi balasan amal kepada mereka semua dengan balasan yang tak terhingga. Akhirnya semoga penelitian ini bermanfaat bagi yang memerlukannya.
Hasil dan Pernbahasan ... 38
Simpulan ... 53
BAB IV KUTUDAUN Myzw persicae
(SULZER)
(HOMOPTERA: APHDIDAE). PARASITOID DAN HIPERPARASITODNY A PADA TANAMAN BROKOLI. CABAIDAN
KENTANG DI...
CILOTO. KECAMATAN PACET. KABUPATEN CIANJUR 58 ...
Pendahuluan 58
...
Bahan dan Metode 61
...
Hasil dan Pembahasan 63
Simpulan ... 84
...
Daftar Pustaka 84
BAB V POPULASI Myzars persicae
(SULZER)
(HOMOPTERA: APKDIDAE). PARASITOIDDAN HIPERPARASITOID
Dl PERTANAMAN BROKOLI. CABAl DAN KENTANG DENGAN...
DAN TANPA INSEKTISIDA 87
Pendahuluan ... 87
...
Bahan dm Metode 90
...
Hasii dm Pembahasan 94
...
Simpulan 109
BAB
VI
PARASITIS AS1 Diaeretiella rape (M'NOSH)DAN
Aphelinus sp.
PADA KUTUDAUN Myzm persicae (SULZER)...
EIalaman
...
Hasil dan Pembahasan 121
...
Simpulan 134
DaRarPustaka
...
135BABVII
PEMBAHASANUMUM
... 138...
Daftar Pustaka 144
BABVIII SIMPULANDANSARAN ... 145 ...
Simpulan 145
...
DAFTAR
TABEL
Nomor EaIaman
3.1 Keanekaragaman spesies turnbuhan inang Myzus persicae yang sudah
...
di laporkan dan yang ditemukan dalarn penelitian 3 8
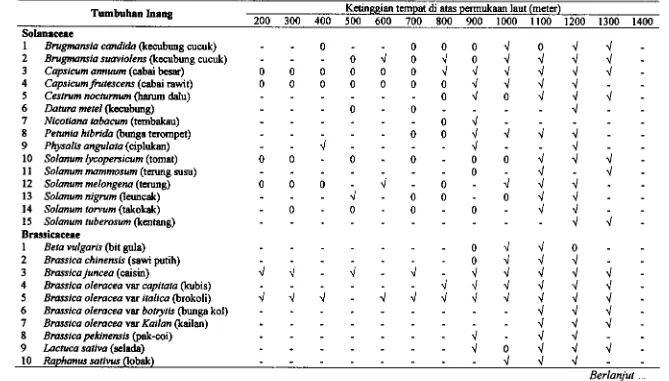
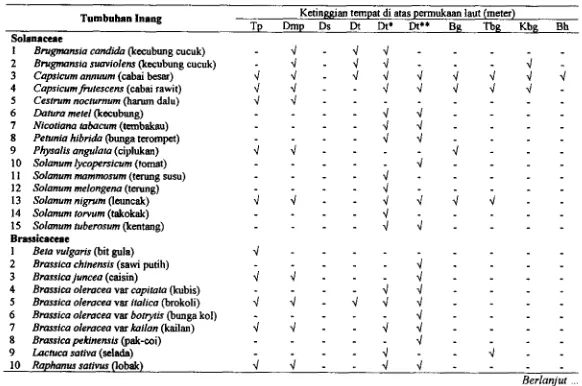
3.2 Tumbuhan inang A@=us persicae clan persebarannya pada ket inggian
...
tempat di atas permukaan laut 40
...
3.3 Letak koloni m p e r s i c a e di tumbuhan inang 43 3.4 Keanekaragaman spesies parasi toid Myzus persicae yang sudah
...
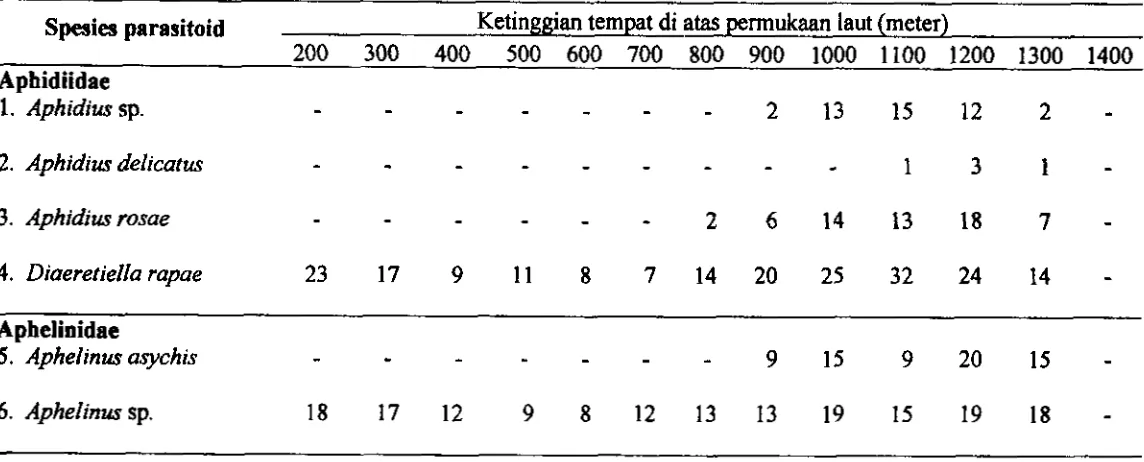
dilaporkan dan yang di temukan &lam peneli tian 45 3.5 Persebaran parasi toid kutudaun A@w persicae berdasarkan ketinggian
...
tempat di atas perrnukaan laut 47
3.6 Keanekaragaman spesies hi perparasitoid pada parasitoid &us persicae yang sudah dilaporkan dan yang ditemukan dalam penelitian ... 49
3.7 Persebaran hiperparasitoid berdasarkan ketinggian tempat di atas ...
pemukaan laut 5 1
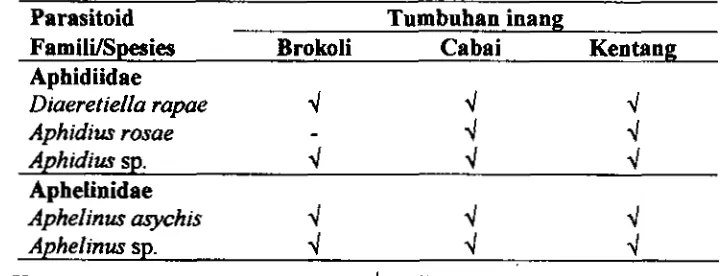
4.1 Asosiasi Myzus persicae dengan tanman brokoli, cabai dm kentang
...
pada rnusim hujan dan musim kernarau 64
4.2 Rerata jurnlah ksdatangan dm koloni A&rus persicae di pertanaman brokoli, cabai dm kentang pada rnusim hujan dan musim kemarau ... 67
4.3 Parasitoid yang memarasit kutudaun A4y.z~~ persicae di tanaman brokoli, cabai dan kentang pada musim hujan dan musim kemarau ... 71 4.4 Rerata populasi mumi aphidiid
dan
aphelinid per tanaman brokoli, cabaidm kentang yang dikolonisasi h&m persicae pada musim hujan
d m
...
musim kemarau 72
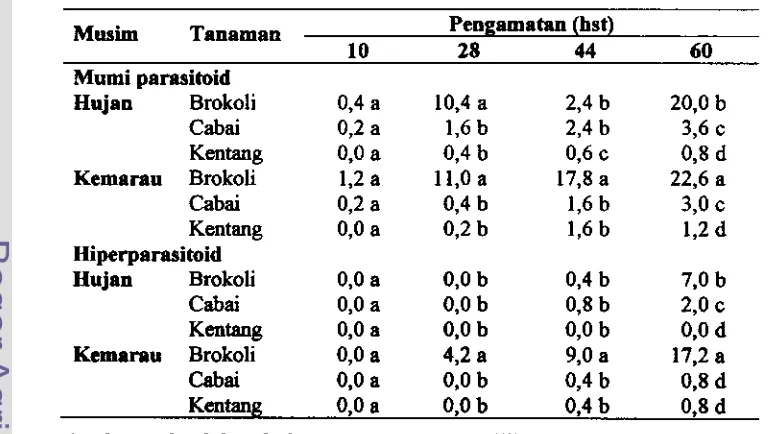
4.5 Rerata murni parasitoid dan hiperparasitoid per tanaman brokoli, cabai clan kentang yang dikolonisasi M y z u persicae pada musim hujan dan musim kemarau
...
78Nomor Halama 5.2 Keane karagaman spesies parasi toid di pertanaman bro koli, cabai dan
kentang dengan dan tanpa insektisida pada musim hujan dm musim kemarau ... 96 5.3 Keane karagaman spesies hiperparasitoid di pertanaman brokoli,
cabai dan kentang dengan dan tanpa insektisida pada musim hujan dm musim kemarau
...
98 5.4 Rerata kelimpahan populasi dan persentase persebaran &us persicaedi pertanaman brokoli, cabai dan kentang dengan dan tanpa insektisida pa& musim hujan dan musim kemarau ... 99
5.5 Rerata populasi imago prasi toid dan hiperparasitoid serta parasi tisasi hiperparasitoid terhadap aphidiid dan aphelinid di pertanaman brokoli, cabai dan kentang dengan dan tanpa insektisida pa& rnusim hujan dan musim kemarau ... 102
6.1 Rerata tiap fase perkembangan kutudaun Wzus persicae yang diparasit oleh Diaeretiella r a p e dan Aphelinus sp.. ... 122 6.2 Kematian serangga inang oleh tusukan ovipositor Diaereriella r a p e
dan host feeding oleh ApheIinus sp. ... 1 23
6.3 Rerata kemat ian Myzus persicae oleh Diaeretiella r a p e dan Aphelinus
... sp.pada masing-masing fase perkembangan inang 124 6.4 Periode waktu pembentukan mumi kutudaun yang diparasit oleh
Diaerel~ella rapae dm ApheIinus sp. ... 126 6.5 Nisbah kelamin imago parasitoid Diaereriella r a p e
dan
Aphelinus sp.pada setiap fase perkembangan inang ... 127 6.6 Penganrh pemkrian pakan terhdap lama hidup imago dan jumlah
DAFTAIR GAMBAR
Nornor Halaman
1.1 Skema hubungan keanekaragaman hunbuhan inang dengan
keanekaragamm parasitoid
c
h
hi perparasitoid y ang berasosiasi dengan kutudaun h@zus persicae ... 5 1.2 Skema penganh musim, ketinggian tempat dan praktik budihya terhadapkeanekaragaman tumbuhan inang persicae dan parasi toid serta .
.
hi perparas~to~dnya ... 7 2.1 Faktor yang mempengaruhi kehidupan Wuspersicae pada turnbuhan
inang ... 2 5
3.1 Perbedaan warna d m bentuk Myzus persicae sehat dan diparasit a) sehat, b) diparasit aphidiid, c) diparasit aphelinid (perksaran 10 kal I ) ... 46
3.2 Lubang keluar imago parasitoid dm hiperparasitoid pa& mumi a) lubang keluar imago parasitoid (perbesaran 10 kali), b) lubang keluar imago ...
hiperparasitoid (perbesaran 7 kal i) 52
4.1 Letak-letak koloni I.tj.zus persicae a) di tanaman brokoli, b) di tanaman cabai, koloninya bercampur dengan Aphis gossypri, c) di tanaman cabai,
...
koloninya mengandung mumi aphidiid, d) di tanaman kentang 66 4.2 Rerata ppulasi M ~ u s persicae per tanaman brokoli, cabai dan kentang
pa& musim hujan dan musim kemarau ... 6 8 4.3 Rerata persebaran kutudaun Myzus persicae di pertanaman brokoli , cabai
...
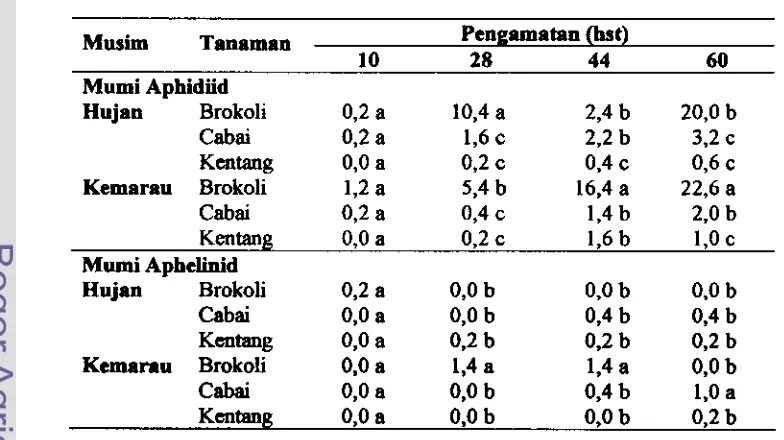
dan kentang pada musim hujan dan musim kemarau 69 4.4 Mumi imago Myztcs persicae yang diparasit aphidiid dengan beberapanimfa di sekitarnya yang diletakkan sebelum menjadi mumi ... 70 4.5 Rerata persebaran aphidiid clan aphelinid di koloni h&us persicae di
pertanaman brokoli, cabai dan kentang pada musim hujan dan musirn kemarau ... 73
4.6 Populasi parasitoid di pertanaman brokoli, cabai dm kentang pada musim hujan ... 75
4.8 Fluktuasi populasi kutudaun, parasitoid dan hiperpamitoid di pertanaman brokoli pada musim kemarau ... 77
4.9 Parasitisasi hiperparasitoid terhadap mumi aphidiid di tanaman brokoli,
...
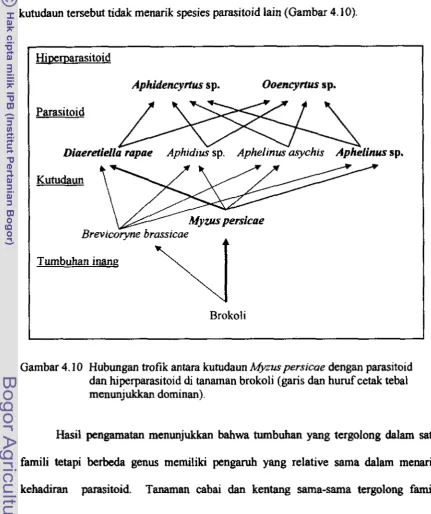
cabai dan kentang pada musim hujan dan musirn kemarau 80 4.1 0 Hubungan tro fi k antara kutudaun Mpus persicae dengan parasitoid dan
hiperparasitoid di tanaman brokoli (garis dan huruf tebal rnenunjukkan ...
dominan) 8 1
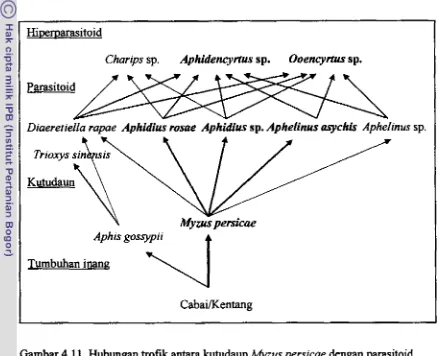
4.1 1 Hubungan trofi k antara kutudaun W u s persicae dengan parasi toid dan hiperpamitoid di tanaman cabal dan kentang (garis dan huruf tebal
...
menunjukkan dominan) 82
5.1 Persebaran parasi toid dl pertanaman brokol i, cabai dan kentang dengan
...
dan tanpa insektisida pada musim hujan dan musim kemarau 104
5.2 Persebaran hipe rparasi toid
di
p e m r n a n brokoli, cabai dm kentang...
dengan dan tanpa insektisida pada musim hujan dan musim kemarau 106
5.3 Mumi aphidiid (a) dan rnumi aphelinid (b) pada koloni h - f m ~ ~ persicae
...
yang menunjukkan populasi kutudaun krkurang 108
6.1 Kernatian kutudaun Mpus persicae yang disebabkan oleh Diaereriella 125 r a p e dan Aphelinus sp. pada fase nimfa dan imago ... 6.2 Preferensi parasi toid Diaeretiella r a p e dan Aphelinus sp. terhadap fase
perkembang inang Myzus persicae, yang ditunjukkan oleh jumlah mumi
...
yang terbentuk dari setiap fase i w g 13 1
6.3 Imago Diaeretiella r a p e yang akan meletakkan telur ke tubuh kutudaun
...
&us persicae 1 3 2
6.4 Imago Aphelinus sp, yang sedang membelakang inang h&zus persicae
...
BAB I
PENDAHULUAN
Latar BelakaogMengetahui keanekaragaman musuh alarni khususnya parasitoid clan kemudian mendapatkan parasitoid yang tepat untuk dijadikan agens hayati pengendali kutudaun
adalah sangat penting. Parasitoid merupakan agens hayati yang pat ing banyak krhasi 1
dibandingkan dengan predator maupun ptogen (Hawkins er a]. 19971, maka sangat beralasan mengapa memilih parasitoid sebagai agens pengendali hayati. Kineja parasitoid dalam menekan populasi kutudaun dipengaruhi antara lain oleh tumbuhan
inang (Kalule & Wright 2002) dan hiperpamitoid (Sullivan & Volkl 1999). Oleh karena ha1 itu kerapkali terjadi, maka penelitian yang berhubungan dengan kedua ha1 tersebut di atas menjadi bagian penting dalam upaya memanfaatkan parasitoid untuk
pengendalian serangga hama yang plifag.
Pemerintah Republik Indonesia meldui Inpres
RI
No.3 Th. 1986,UU
RI No.
12 th. 1992 dm PP RI No.6 Th. 1995 menetapkan pengendalian hamst secara terpadu (PHT) sebagai landasan dalam mengendalikan organisme penganggu tanaman (OPT). Pengendal ian hayati merupakan salah satu komponen utama PHT. Kebij akan pemerintah tersebut telah mendorong peningkatan penelitian y ~ q g berhubungan dengan musuh alarni OPT dan pemanfaatannya (Sosmmarsono 2000). Namun penelitian- penelitian itu urnumnya masi h terbatas dm terpisah-pisah pa& pengunatan, eksplorasi
Penempan pengendalim hayati dilakukan melalui pengelolaan habitat serta perbanyakan dm pelepasan rnusuh alarni secara massal (DeBach 1 979). Pengelolaan habitat bertujuan memberi kesempatan kepada m u s h alarni bertahan hidup dan berkembngbiak pa& suatu habitat, sehingga proses pengendalian &pat bejalan dengan sendirinya (Brewer & Elliot 2004). Perbanyakan dan pelepasan m u s h alami krtujuan menekan populasi hama pda saa? dibutuhkan, karena itu aplikasinya di
lapangan memerlu kan informasi faktor biot i k dan abioti k habitat (Berryman 1 999). Kutudaun merupakan salah satu harna penting karena &pat menjadi hama dan vektor penyakit virus tanaman penting pada tanaman di lapangan maupun di rumah kaca (Hill 1995). Kerugian oleh kutudaun sebagai hama berkisar antara 6-25% dan sebagai vektor dapat mencapai lebih dari 80% (Miles 1987). Besar kecilnya angka kerugian yang disebabkan oleh kutudaun itu erat kaitannya dengan umur clan varietas tanaman serta spesies kutudaun (Kranz er nl. 1978). Kutudaun krsifat monofag,
oligofag dan polifag, dan beberapa spesies di antaranya ada yang sangat poiifag sekaligus menj adi vektor penyakit virus penting tanaman, karena itu sangat merugi kan (Blackman & Eastop 2000).
@zw persicae (Sulzer) (Hornoptera: Aphididae) merupakan salah satu spesies kutudaun penting di daerah subtropis maupun tropis dan bersifat sangat polifag (Hill
1995). Lebih
dari
400 spesies tumbuhan dalam 40 h i l i dapat menjadi inang M persicae (Kranz ef al. 1978, Blackman & &top 2000). Di samping ituM
persiccediketahui dapat menjadi vektor lebih
dari
150 jenis virus persisten maupun non-persisten @astop 1977).
pengeloiaan habitat. Dixon clan Kindmann ( 1998) menyatakan Wwa keberadaan spesies tumbuhan inang mempengaruhi fluktuasi populasi kutudaun. Di samping itu
tumbuhan inang j uga mempengaruhi interaksi antara parasitoid dengan kutudaun serta interaksi antara hiperparasitoid dengan parasitod (Godfray & Muller 1998). Informasi keanekaragaman tumbuhan inang tersebut sangat berguna dalam mengantisi p s i sum ber i nvasi clan tempat berl indung kutudaun rnaupun musuh-musuh alaminya (Unruh & Messing 1997).
Di laporkan ada tiga spesies parasitoid yang memarasit M persicae, yaitu Aphidtus sp., Diaeretiella sp. (Hymenoptera: Aphidiidae) dan Aphelinus sp.
(Hymenoptera: Aphelinidae), di daerah Bogor (Irsan & Sosromarsono 2000). Parasttoid pa& kutudaun di negeri dengan empat musim teiah dimanf~tkan untuk mengendalikan kutudaun clan menunjukkan hasil yang baik (Hagen & van den Bosch 1968, Stary 1988, Carver 1989). Di negeri itu sudah tersedia sscara komersial parasitoid &lam kernasan yang siap diaplikasikan untuk rnengendal i kan kutudaun (Koppert 1999). Orr dan Suh (2000) mengemukakan bahwa dl Amerika Utara pada tahun 1997 terdapat 142 perusahaan komersial yang menjual 130 spesies agens hayati, 46 spesies di antaranya adalah parasitoid. Indonesia pernah mengimpr p s i t o i d
Lyslpklebus testacepes f Cresson) (Hymenoptera: Aphidi idae) untuk mengendalikan kutudaun Aphis craccivora K w h (Hornoptera: Aphididae) pa& kacang-kacangan,
tetapi tidak a& laporan mengenai keberhasilan introduksi m u s h alami tersebut (Soemartono Sosrommono, komunikasi pribd). Pengembangan parasitoid berdasarkm hasil penelitian perlu dilakukan.
parasitoid yang memottasit kutudaun yang tergolong dalam famili Encyrhdae. Kedua spesies hiperparasitoid itu memarasit Trioxys sp. (Hymenoptera: Aphidiidae) yang memarasit kutudaun Aphis gossypti (Glover) (Hornoptera: Aphididae) di Bogor. Di Ci panas encyrtid bperpamsitoid tersebut juga diternukan memarasit Diaeretiella sp. dan Aphelinus sp. yang merupakan parasitoid pada kutudaun
M.
persicae yang berkoloni di tanaman brokoli (Irsan 200 1 a).S e m urnum infomasi tertulis mengenai parasitoid dan hiperpamitoid yang bemsiasi dengan M. persicae di Indonesia masih sangat lmgka. Untuk memanfaatkan parasitoid sebagai agens hayati sangat di perlukan in formasi keanekaragaman spesies parasitoid tersebut clan j uga keanekaragaman spesies hiperparasitoidnya di suatu habitat (Altieri ef a/. 1997).
Untuk mengaplikasikan parasitoid &lam pengendalian M. persicae d~ perlukan informasi hubungan trofik. Hal itu penting mengingat spesies turnbuhan inang kutudaun dapat mempengaruhi kehidupan pamitoid dm hi perparasitoidnya (Panda &
Khush 1995). Barbsa dm Benrey (1 998) menyatakan bahwa spesies atau varietas
tanaman mempengmhi spesies parasitoid yang berasosiasi dengan herbivor yang hidup di tanaman itu. Penelitian hubungian trofi k antara kutudaun
M.
persicae dengan tumbuhan inang, pamitoid dm hiziperparasitoid di Indonesia belurn pernah dilakukan. Inforrnasi penelitian tersebut merupakan landasan penting dalam pelakwnaan pengendalian hama terpadu berbasis pengendalian hayati yang berkelanjutm danramah lxngkungan serta ekonomis (LaSalle 1997).
jenis tanaman tersebut selain mewakili famili juga didasarb pada faktor lainnya yaitu budidaya tanaman ymg luas di masyarakat, nilai ekonomi, nilai gizi clan kebutuhan masyarakat terhadap ketiga jenis tanaman serta penggunaan pestisida y ang intensi f pada ketiga jenis tanaman tersebut. Tanaman yang dipilih diharapkan &pat memberikan gambamn umum &lam perenaman pengembangan pengendalian kutudaun secara hayati
.
Mengingat pentingnya suatu penelitian yang bermuara pada pemanfaatan hasil
penelitian, maka dilakukan penel i tian yang saling menunjang yaitu eksplorasi untuk
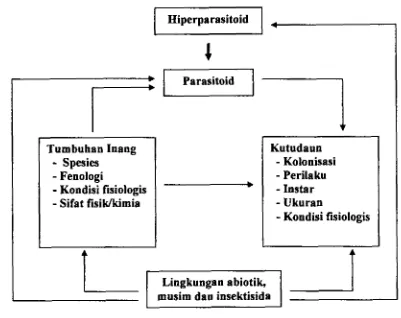
mendapatkan infomasi keanekaragaman tumbuhan inang M. persicae, parasitoid yang berasosiasi dengan kutudaun I tu serta keanekaragaman hi perparasitoid y ang memarasit parasitoid pada l utudaun M persicae. Keanekaragaman parasi toid dan hi perparasitoid tersebut diamati keterkaitanny a dengan keanehgaman twnbuhan inang seperti dipaparkan dengan skema pada Garnbar 1.1.
g Hiperparasitoid
Keanekaragaman
Keanekaragaman
-
= Hubungan trofik,- - - -
+
= Hubungan non-trofikUntuk lebih memahami hubungan keanekaragaman itu secara keseluruhan maka dilakukan penelitian pengaruh habitat berupa pertanaman budidaya yang mendapat perlakuan insektisida dan tanpa insektisida serta pengaruh s p i e s tumbuhan
hang terhadap kelimpahan ppulasi
M.
persicae dan keanekaragaman spesies parasitoid dan hiperparasitoidnya pada m u s h hujan dan rnusim kemarau. Di samping i tu j uga dilakukan penel i tian terhadap parasitisasi parasitoid yang dorninan memarasi tM
persicae unt uk mengetahui potensi parasitoid sebagai agens hayati pengendal ikutudaun.
Tujuan Penelitian
Secara umum penelitian bertujuan untuk mendapatkan informasi tentang keanekaragaman spesies tumbuhan yang menjacfi inang
M.
persicae serta parasitoiddan hi perparasttoid yang berasosiasi dengan kutudaun tersebut. Penelitian juga bertujuan memahami hubungan antara M persicae dengan t u m b h n inang dm parasitoid serta hiperparasitoidnya dalam lingkungan berbeda, khususnya musim dan perlakuan insektisida. Untuk men@ tujuan penelitian tersebut dilakukan serangkaian penelitian sebagai berikut :
a. Spesies tumbuhan yang menj adi inang Ad p e r s u e
b. Spesies prasitoid yang memarasit
M
persicaec. Spesies hiperparasitoid yang memarasit parasitoid kutudaun
M
persicaed. Peranan spesies tumbuhan inang pads kutudaun Ad persicae dan keanekaragaman spesies parasitoid dm hi perparasitoidnya
f. Pmitisasi Diaeretiella r a p e (M'intosh) dan Aphelintrs sp. pada kutudauu
M
persicae
Peri ha1 peranan spesies tumbthn terhadap
M
persicae, parasitoid dan hiperparasitoid yang menj adi tujuan penelitian ini secara skematis disajikan di Gambar1.2. Skema tersebut menggambarkan pengaruh ketinggian tempat (mdpl), prakti k
budidaya dan musim pada hubungan trofik antara tumbuhan inang dengan kutudaun,
parasi toid clan hiperparasitoidnya yang rnempenganh keanekaragaman spesies tumbuhan inang, parasi toid dan hiperparasi toid.
Keanekaragarnan spesia turn buhao inang, parasitoid
dan biperparasitoid
Musim +
...
Hiperparasitoid
t
.,
... Parasitoid
t
Myzus
persicaet
- ... ... *
Tumbuban inang
Praktik Budidaya
+ Ketinggian
tempslt
Garnbar 1 -2 Skema pengaruh musim, ketinggian tempat dan prakti k budidaya terhadap kemekamgaman tumbuhan inang Myzur persime dm parasitoid serta
Manfaat Penetitian
Penelitiam ini di harapkan dapat memberikan informasi baru tentang keanekaragaman spesies parasitoid kutudaun Ad persicae dan keanekmgaman spesies hi perparasitoid yang memarasit parasitoid kutudaun
M
persicae. Tumbuhan i m g yang berpengaruh pada keberadaan dan kelimpahan populasiM
persicae diharapkan dapat di manfaatkan dalam melakukan intervensi pada waktu y ang tepat. Keanekaragaman spesies dan kelirnpahan populasi parasitoid diharapksn dapat mem berikan solusi dalarn memili h parasitoid yang tepat untuk tanaman tertentusebagai agens hayati pengendali kutudaun. Musim hujan
dan
musim kemarau yang berpengaruh pa& kelim pahan populasi dan keanekaragaman spesies paras; toid maupun hi perparasi toid diharapkan &pat memberi kan gambaran tentang fluktuasi populasi parasitoid di lapangan. Habitat yang j uga krpengaruh pa& kelimpahan populasi dan keane karagaman spesies parasi toid maupun hi perparasitoid di harapkan &pat memberi kan pemahaman tentang pentingnya mengelola habitat atau lingkungan untuk konservasi parasi toid.Selain manfaat tersebut di atas, pendekatan metodolog yang digunakan &lam penel i tian ini di harapkan dapat dijadi kan model dalam mempelajari hubungan tumbuhan inang dengar, kutudaun dan parasitoid serta hi perparasitoidnya.
Barbosa P, B m y
B.
1998. The influence of plant on insect parasitoid: implications for consewation biological control. Di Mam: Barbosa P, editor. ConservationBiological Control. San Diego: Academic Press. hlm 5 5-83.
Berryman AA. 1999. The theorical foundations of biological control. Di dalam: Hawkins B A, Comell W , editor. Theoretical Approaches lo Biological
Confrol. Cambridge: Cambridge University Press. hlm 3-2 1.
Blackman RL, Eastop VF. 2000. Aphids on she World's crops: an Identrjicarion
Guide. Second edition. Chichester: Wiley .
Brewer MJ, Elliot NC. 2004. Biologcal control of cereal aphids in North America and mdating effects of host plant and habitat manipulation. Annu R w
En fomol49:2 19-242.
Carver M. 1989. Biological control of aphids. Di dalarn: Minks
AK,
Harrewijn P, editor. Aphids: Their Biulogv, Noturul Enemies and Con~rol. Vol 2C. Amsterdam: Elsevier, hlm 14 1-
165.DeBach P. 1979. Biological Control by Natural Enemies. London: Cambridge Univ Press.
Dixon AFG, Kindlmann P. 1998. Population dynamics of aphids. Di dalam: Dempster JF, Mdean IFG, Dordrecht, editor. Insect Populations in Theory and
in Practice. London: Kluwer Academic. hlm 207-230.
Eastop VF. 1977. World wide importance of aphids as virus vectors. Di dalam:
Harris KF, M m o r o s c h K, editor. Aphids as Virus Vectors. New York:
Academic Press. hlm 1-4 1.
Godfray HCJ, Muller CB. 1998. Host-parasitoid dynamics. Di dalam: Dempster
P,
Mclean IFG, Dordrecht, editor. Insect Population in Theory and in Practice.London: Kluwer Academic. hlm 1 35- 165.
Hagen kS, van den Bosch R. 1968. Impact of pathogens, parasites, and predators on aphids. Annu Rev Entomol13: 3 25 -3 84.
Hawkins BA, Comell HV, Hochberg ME. 1997. Predators, parasitoids, and pathogens
as mortality agents in phytophagous insect population. Ecol Soc Amer
78:2 145-2152.
Hill D. 1995. Agricultural Entomology. Portland: Timber Press.
Irsan C. 200 1 a. Parasitisasi parasitoid sekunder pada parasitoid primer kutudaun (Hornoptera: Aphididae). Di Dalarn: Prosiding Seminar Nasional Pertanian berkelanju~m, Bandar Larnpung, 26-27 Juni 200 1. hlm 699-705.
Irsan C, Sosromarsono S. 2000. Identifi kasi Hymenoptera parasi t pada kutudaun (Hornoptera: Aphididae) di tanaman budidaya di berbagai ketinggian tempat di sekitar Bogor. Di dal am: Prosiding Simposium Keanekuragamun Hayatr Arlhropoda pado System Produksi Pertaninn, Cipayung, 1 6- 1 8 Okt. 2000. hlm
149-1 54.
Kalule T, Wright DJ. 2002. Effect of cabbage cultivars with varying levels of resistence to aphids on the performance of the parasitoid, Aphidius colemani
(Hymenoptera: Braconidae). Bull Enromol Res. 92:53-59.
[Koppert]. 1 999. Koppert Product with Direction for Use. Berkel en Rodenrij s. Koppert Biologcal System.
Kranz J, Schmutterer H, Koch W. 1978. Diseases, Pests and Weeds in Tropical Crop.$. Chichester: Wiley.
LaSalle
J.
1997. Parasitic Hymenoptera, biological control and biodiversity. Di dalam: LaSalle 3, Gadd ID, editor. Hymenoptera and Biodiversity.Wallingford: CAB International. hlrn 197-2 15.
Miles PW. 1987. Feeding process of Aphidoidea in relation to effects on their food plants. Di dalm: Minks
AK,
Harrewijn P, editor. Aphids: Their Biology, IC'atural Enemies and Control. Vol2A. Amsterdam: Elsevier. hlrn 32 1-340. Orr DB, Suh CP-C. 2000. Parasitoids and pedators. Di dalam: Rechcigl JE, RechciglNA , editor. Biological and Biotechnological Con/ rol of Insect Pests. Boca Raton: Luwis Publ. hlm 1-34.
Panda N, Khush GS. 1995. Host Plants Resistance to Insects. Manila: CAB International and International Rice Research Institute.
Sosromarsono S. 2000. Sej arah pengendalian hrtyati serangga hama dengan parasitoid di Indonesia. Di dalam: Workshop on Development and Utilization of parasitoid (Exploration, Identification, Mass Production and Field Release). J mHama dan Penyakit Tumbuhan. Bogor: Institut Pertanian Bogor.
Stary P. 1988. Aphidiidae and Aphelinidae. Di dalam: Minks AK, Harrewijn P,
editor. Aphids: Their Biology, Natural Enemies and Control. Vol 2B. Amsterdam: Elsevier. hlm 171-188.
Sullivan DJ, Vol kl W. 1 999. Hyperparasitism: multitropic, ecology and behavior.
BAB
I1
TINJAUAN PUSTAKA
Kehidupan serangga fi to fag di tentukan terutama oleh hunbuhan inang, sedangkan kehidupan serangga entomofag ditentukan oleh serangga inang yang
di parasi t atau dimangsanya (Price 1 997). Panda dm Khush ( 1 995) menyatakan bahwa turn buhan tempat hidup serangga inang mem pengaruhi secara langsung kehidupan
serangga fitofag. Karenanya varietas atau spesies tanaman dapat rnempengaruhi secara langsung kehidupan serangga fitofag dan selanjutnya akan berpengaruh pada kehidupan serangga entomofag (Schoonhoven et ol . 1 998).
Perkembangan popuIasi htudaun dipenganhi oleh spesies tumbuhan inang dan musuh alarni seperti predator, parasitoid dm patogen serta tuperparasitoid (Memmott & Godfray 1997). Satu spesies kutudaun yang sama, namun hidup pada tumbuhan
inang yang berbeda akan menunj ukkan respons pertumbuhan populasi berbeda (Tsai 1998). Secara umum spesies atau varietas tanaman dapat mempenganhi kinerja parasitoid (Barbosa & Benrey 1998). Tetapi tidak demikian halnya dengan hiperparasitoid, karena hiperparasitoid umumnya tidak menunj ukkan kespesifi kan inang (Sullivan & Volkl 1999). lrsan (2001a) melaporkan bahwa spesies hi perparasitoid y ang sama dapat di temukan memarasit parasitoid kutudaun spesies berbeda pstda tumbuhan inang yang berbeda. Namun pernyataan tersebut memerlukan peneIitian lebih lanj ut karena informasi yang a& masi h sangat terbeztas. Secara umum
Kutudaun Myacsprsicw
Di wilayrth atau negeri dengan empat musirn, M. persicae terdiri dari individu jantan dan betina di m u s h gugur, dan betim partenogenetik di m u s h yang lairmya. Pada musim gugur dan awal musim semi kutudaun hidup di tumbuhan inang primer dm pada akhir musim semi dan rnusim panas hidup di tumbuhan inang sekunder (Dixon 1978, 1987; Blackman & Eastop 1994, 2000). Di daerah tropis, M. persicae terus menems berkembangbiak secara partenogenetik (Kranz er a/. 1978). Kalshoven (1 98 1 ) menyatakan bahwa di dataran tinggi di daerah tropis dapat ditemukan kutudaun jantan dan betina, tetapi pernyataan ini belum a& konfirmasi baru.
M. persicae merupakan salah satu harna penting di daerah subtropis maupun tropis dan bersifat sangat polifag. Lebih dari 400 spesies turnbuhan yang tergolong dalarn 40 famili dapat menjadi tumbuhan inang
M.
persicue (Kranz el a/. 1978; Bl ackman & Eastop 2000).M
persicae juga diketahui dapat menjadi vektor Iebih dari1 50 jenis virus atau strain virus, baik itu virus persisten maupun non-persisten pada tanaman (Hill 1995; Harris & Mararnorosch 1977). Kutudaun itu temasuk salah satu hama dan vektor pen yakit virus penting pada tanaman kacang-kacangan, bit gula, tebu,
kentang, tembakau, kubis-kubisan dan jeruk (van Emden et al. 1969), dan merupakan hama clan vektor penyakit virus yang penting pa& tanaman yang tergolong dalam famiIi Solanaceae (Blackman & Eastop 2000).
Turn buhan Inang dan Kutudaun
sehubungan dengan keberadaafi sumber invasi dan kerusakan yang disebabkan oleh
kutudaun. Marlinda (1 999) melaporkan bahwa kerusakan tanaman cabai Capsicum
annuurn L. yang disebabkan oleh kutudaun A. gossypii di rumah kaca dapat mencapai 100%, artinya a& tanaman cabai yang tidak &pat menghasilkan apa-apa untuk dipanen karena serangan kutudaun tersebut. Hidayat ( 1997) menyatakan bahwa pada tanarnan kentang dapat ditemukan banyak spesies kutudaun yang inang sesungguhnya
bukan kentang. Namun kehadiran kutudaun itu di tanarnan kentang sudah dapat
merugi kan karena kutudaun tersebut berpotensi menularkan virus apabi la di tanaman kentang itu ia sempat menusuk-nusukkan stiletnya mendeteksi kesesuaian inang
(probing), dan sebelum menusuk-nusukkan stilet di tanaman kentang tersebut, kutudaun itu telah mengisap cairan tumbuhan atau menusuk-nusukkan stiletnya pada tumbuhan sakit yang terinfeksi virus.
Kenyataan di lapangan menunjukkan bahwa satu spesies kutudaun &pat diternukan di berbagai spesies hunbuhan inang (Irsan ef a / . 1998). Warna tubuh satu spesies kutudaun dapat berbeda-be& karena hidup di tumbuhan inang yang berbeda. Perbedaan warna itu dipengaruhi oleh tumbuhan inang dan suhu (Takada 1979). Menurut Setokuchi (1 98 I ) , pigmen yang tetkandung di dalam jaringan tumbuhan inang mempengaruhi warm tubuh kutudaun. Di samping itu turnbuhan inang dan
lingkungan juga mempengaruhi ukuran tubuh kutudaun. Ukuran tubuh satu spesies kutudaun dapat berbeda-beda karena kualitas cairan floem yang dihisap dan suhu yang
berbeda (Mayo & Starks 1974,
Dixon
& KindImam 1994). Kcragaman ukuran tubuhIamina daun rnaka ukuran tubuh kutudaun menjadi kecil (Dixon ec a/ 1995, Douglas
1993). Walaupun demikian u k m tubuh kutudaun itu dibatasi oleh ukuran
maksimumny a (maximum body size) (Kindlmann & Dixon 1 992). Kutudaun yang diberi makanan berl impah tidak akan meny ebabkan ukuran tubuhnya besar melebihi ukuran rnaksimumnya.
Tumbuhan hang, Kutudaun drn Parasitoid
Kutudaun memerlukan tumbuhan inang sebagai sumber karbohidrat, lemak, protein, mineral dan vitamin untuk mempertahankan hldup dm menemkan ketunmannya. Setiap spesies atau varietas tumbuhan memiliki kandungm senyawa metablit primer dan sekunder dalam kualitas dan kuantitas yang berbeda (Panda & Khush 1995). Metablit sekunder itu ada yang berakumulasi di tubuh kutudaun dan
selanj utnya dapat mempengaruhi parasitoid yang memarasit kutudaun tersebut (Schoonhoven er al. 1998). Di samping itu struktur permukaan jaringan tumbuhan inang j uga berpengaruh pada aktivitas parasi toid mencari atau menemukan kutudaun untuk dijadikan serangga inang (Vol kl & Stechmann 1998).
Kunjungan prasitoid pada suatu koloni kutudaun erat kaitannya dengan metaborit sekunder yang dihasilkan oleh tumbuhan inang (Panda & Khush 1995).
Besar kecilnya ukuran tubuh kutudaun dari spesies yang sama
akan
berpengaruh pada jenis kelamin pamitoid yang muncul. Parasitoid jantan banyakmuncul dari kutudaun yang tubuhnya bendcuran kecil, sedangkm pamitoid ktina banyak muncul
dari
kutudaun yang tubuhnya berukuran relatif besar. Kemunculan parasitoid berkelamin jantan dari serangga inang yang berukuran kecil clan betina dari serangga inang ymg berukuran ksar umum te jadi pada parasitoid, karena imago parasitoid memilih meledclan telur bakal jantan atau telur bakal betina berdasarkan ukuran tubuh hang yang akan diletaki telur (Godfray 1994, Quicke 1997).Semut dapat berpengaruh negatif terhadap kinerja prasitoid. Menurut Quicke (1997), semut akan memilih kutudaun ymg terparasit untuk dimangsa atau dibawa ke sarangnya. Akibatnya jumlah inhvidu parasitoid yang berhasil bertahan hidup menjadi berkurang karena larva dan atau pupanya dimangsa sernut. Keberadaan semut dapat menghalangi atau mengganggu kunjungan imago parasitoid yang akan meletakkan telur di tubuh kutudaun (Sudd 1987). Di samping itu semut juga dapat meningkatkan laj u parasitisasi parasi toid maupun hiperparasitoid. Peningkatan i tu terj a& karena semut yang mengkonsurnsi embun madu menyebabkan pemukaan daun tidak licin dm tidak menyebabkan imago parasitoid atau hi perparasitoid tergeiincir (Volkl & Stechmann 1998). Kondisi permukaan daun tersebut memudahkan imago parasitoid atau hiperparasitoid bergerak menemukan serangga inangnya.
mengendalikan
S.
graminurn yang hidup pads wietas sorgum dan barley yang resisten. Demikian juga hainya dengan kinerja Aphidiur mhicariae HaI iday &lam menekanM.
persicae pda varietas Chrysan~hernum, y aitu pada varietas resisten lebih baik danpada pada varietas rentan (Wyatt 1971). Dengan demikian dapat dipahamib&wa penggunaan pasitoid benama-ma dengan varietas resisten &pat dijadi kan kombinasi dalarn rnenekan populasi kutudaun.
Nisbah kelamin parasitoid dipengaruhi oleh varietas tanaman inang. Huey- Ling (1986) menyatakan bahwa nisbah kelamin parasitoid yang ditunjukkan dengan perbandingan j antan dan betina yang te jadi pada parasi toid yang memarasi t kutudaun pada varietas resisten lebi h rendah yai tu 1 : 1,4 daripada yang memarasi t kutudaun yang berkoloni pada varietas rentan yaitu 1 :2,4. Perbandi ngan nisbah kelamin tersebut
mempenganhi perkembangan populasi parasitoid pada generasi beri kutnya.
Varietas resisten mempengaruhi asam amino yang diperoleh kutudaun. Asam amino itu berpengamh pada kualitas gizi embun madu yang dihasilkan kutudaun. imago parasitoid yang mengonsumsi embun madu yang kandungan asam aminonya rendah akan menurun kinerj anya dalam memarasit kutudaun (Huey-Ling 1986; Hagen 1986). Parasitoid menuju ke habitat dipandu oleh bau kutudaun dm embun madu yang diekskresikan oleh kutudaun itu (Powell 1989). Penelitian Bouchard &n Cioutier ( 1984) menunj ukkan bahwa aktivitas mencari serangga inang oleh Aphidius nigripes
Parasitoid LCutudaun
Studi pustaka menunj ukkan ads empat famili ordo Hy menoptera y ang memarasit kutudaun. Keempat fmi li itu adalah Aphidiidae (Ichneumonoidea), Aphelinidae, Encyrtidae dan Mymmidae (Chalcidoidea) ( Hagen & van den Bosch
1968). Spesies-spesies aphidiid dan aphelinid mwupakan hymenopteran parasit yang ban yak dilaporkan memarasit kutudaun (Stmy 1 988). Seam urnum keanekaragaman
spesies aphidiid parasitoid yang memarasit kutudaun lebih tinggi daripada aphelinid (van Emden et al. 1969). Dua famili lainnya yaitu Encyrtidae dan Mymaridae belum banyak diteliti (Hagen & van den Bosch 1968). Parasitisasi aphidiid clan aphelinid pa& kutudaun M persrcae membentuk murni dengan morfologi y ang khas, sehingga melalui morfologi murni kutudaun yang terbentuk sudah dapat ditentukan famili parasitoid yang memarasit (Stary 1988).
Apbidiid Parasitoid
Parasitoid bertubuh kecil dengan ukuran satu sampai beberapa millimeter. Imago krwarna kuning kewlclatan atau coklat gelap dengan pola kekuningan, jingga
atau kuning kecoklatan, dan bersifat e n d o m i t soliter (Hagen & van den Bosch 1968). Stay (1988) menyatakan bahwa sedikitnya ada 60 genus dan lebih dari 400 spesies aphidiid yang te1ah diketahui mernarasit kutudztun. Di Asia Timur diternukan
parasitoid dap& memarasit lebih dan satu spesies kutudaun, dan satu spesies b & u n dapat diparasit oleh lebih dari satu spesies parasitoid.
Van Emden
et al. (1969) menyatakan bahwa, ada 40 spesies aphidiid yang memarasitM
persicae, lrsan clanSosromarsono (2000) melaporkan bahwa di sekitar Bogor ada dua spesies aphidiid memarasit
M,
persicae. Kedua spesies tersebut adalah Aphiditrs sp. dan Diaeretiellasp. Lawton (1986) mengungkapkan bahwa beberapa spesies parasitoid kutudaun dari
fami li Aphidi idae mernpunyai inang yang luas, di antaranya Ephedrtrs spp, memarasit antara 50 sampai 100 spesies kutudaun. Sifat poiifag tersebut erat kaitannya dengan kernampuan parasitoid mendetoksifikasi senyawa yang berasaI dari tumbuhan inang yang di hisap oleh kutudaun yang menjadi i nangnya.
T i p reproduksi aphidiid adalah arrhenotoki, yaitu telur yang tidak dibuahi (haploid) menjadi jantan dan yang dibuahi (diploid) menjadi betina (Stay 1988). Stary
(1 970) menyatakan bahwa beberapa spesies ap hi diid ada yang hanya menghasilkan
keturunan betina saja dan t i p ini disebut theliotoki serta ada yang rnenghasilkan keturunan jantan &n betina tetapi secara biologi yang jantan tidak berfungsi dan tipe ini disebut deuterotoki. Nisbah kelamin parasitoid secara umurn adalah 1 : I . Stary (1970) menyatakan bahwa, nisbah kelamin aphidiid 1:2. Tetapi nisbah kelamin itu dapat berubah tergantung Iingkungan terutama suhu.
Siklus perkembangan parasitoid dari telur sampai imago berkisar antara 10-3 0 hari. Perkdam waktu dipengaruhi oleh suhu dan kelembaban. Makin tinggi suhu atau
makin rendah kelembaban menyebabkan siklus hidup parasitoid makin singkat. Pada
embun madu merupakan makanan penting bagi imago parasitoid. Selain embun madu imago parasitoid juga memerlukan nektar, air dan protein melalui host feeding
(Hagen & van den Bosch 1968).
Aphelinid Parasitoid
Aphelinidae merupakan salah satu farnili dari ordo Hymenoptera yang
anggotanya banyak menjadi parasitoid pada kutudaun. Secara umum aphelinid
khususnya genus Aphelintls banyak memarasit kutudaun yang mempunyai sifunkuli berukuran pendek (Hagen & van den Bosch 1968). Van Emden et al. (1969) melaporkan bahwa ada enam spesies aphelinid yang memarasit
M
persicae yang terrnasuk kutudaun yang merniliki sifunku t i panj ang. Irsan dm Sosromarsono (2000) melaporkan ada satu spesies parasitoid tergolong dalam farnili Aphelinidae memarasitM
persicae yaitu Aphelinus sp.Penggunaan spesies Aphelinid sebagai agens hayati pengendali kutudaun sudah banyak dilaporkan (DeBach 1979). Di antaranya Aphelinus mali (Haldeman) yang digunakan untuk mengendalikan kutudaun pada tanaman ape1 di Amerika Serikat dm A. asychis yang digunakan untuk mengendal ikan kutudaun Therioaphis trqolii pada pohon oak di Tasmania (Hagen & van den Bosch 1968).
Kutudaun dapat menghhdari serangan aphelinid dengan menggerakkan antena dm tungkainya sebelum parasi toid hinggap di tubuhnya, atau menggoyang antena, tungkai dan tubuhnya pada saat imago parasitoid mencoba meletakkan telur ke dalam tubuhnya (Gross 1993). Secara fisiologi pada beberapa spesies kutudaun dapat terjadi pengkapsulan terhadap telur dan larva parasitoid di dalam tubuh inang itu (Hagen &
h a g o Aphelimrs mengonsumsi embun madu, oektar serta melakukan host fieding, namun ada yang hanya melakukan host feeding saja. Di Iaboratoriurn A.
agehis hanya hidup tiga hari tanpa makan, tetapi dapat hidup sampai 37 hari jika diberi pakan larutan rnadu (Hagen & van den Bosch 1968).
Parasitisasi aphelinid pada kutudaun menyebabkan perubahan warna kutudaun
yang diparasit
.
Umurnnya warna tubuh kutudaun yang terparasit aphelinid b e m a hitam. Embelan yang ada di bagian kepala dari mumi yang berwarna hitam itu ti& hitam, melainkan transparan. Imago muncul dari murni melalui lubang di bagiandorsal rnumi (Hagen & van den Bosch 1968).
Reproduksi aphel inid secara urnurn bertipe arrhenotoki dan beberapa spesies bersifat deuterotoki (Stary 1 988). Serangga hang dan suhu mernpengaruhi nisbah kelamin. A. my chis yang bertipe arrhenotoki pada M persicae han ya m e n g h a s i h sedikit jantan. Pada suhu tinggi akan banyak jantan dihasilkan, sebaliknya betina akan banyak dihasilkan pada suhu rendah. Banyahya jantan yang dihasilkan pada suhu
tinggi itu disebabkan oleh sedikitnya betina yang kopulasi, sehingga keturunan yang d i h a s i h lebih banyak yang jantan (Hagen & van den Bosch 1 968).
Perilaku menarik dari imago aphelinid ialah tidak &if terbang pada saat
sampai di habitat serangga inang. Biasanya imago parasitoid itu hays beqalan dan atau terbang pada jarak yang pendek di tumbuhan tempat hidup inangnya. Di samping
Semua imago Aphelinus spp. melakukan host feeding, kegiatan itu dimulai
dengan menusuk integumen serangga inang dengan ovipsitornya. Setelah itu imago meletakkan telur dahulu baru melakukan host feeding, tetapi
ada
juga Aphelinus spp. yang melakukan host feeding tanpa meleakan telur. Imago Aphelinus spp. setelah meletakkan telur, akan memutarkan badan dan mulai melakukan host feeding,menghisap inang sampai kering clan mati, yang berakibat pada telur yang diletakkan ikut mati juga. Pada A. mall perilaku tersebut dapat menyebabkan kematian 3-5 kutudaun per hari (Hagen & van den Bosch 1968).
Encyrtid Parasitoid
Secara umurn spesies parasitoid yang tergolong dalam famili Encyrtidae sedikit j umlahnya, dan sebagian besar mereka temasuk dalam genus Aphidenqrroides. Barn
satu spesies yang telah dilaporkan memarasit kutudaun yaitu A. ~horacuphis yang memarasit Nipponaphis sp. clan Metanlpponaphts sp. di Jepang (Hagen & van den Bosch 1968).
Mymarid Parasitoid
Masi h sangat sedikit penelitian rnengenai mymarid yang memarasit kutudaun. Ada dua spesies yang dilaporkan memarasit kutudaun Pertama adalah Polynema striaticome yang dilaporkan memarasit hreartophis bakert (Cowen). Kedua addah
Poiynema schmitzi yang memarasit Pemphigtrs sp. (Hagen & van den Bosch 1968).
Hiperparasitoid prda Parasitoid Kutudaun
1999). Hippamitoid menentukan kine j a parasitoid (LaSalle & Gauld 1997). Kematian parasibid oleh hiperpamitoid berkisar antara 50-90% (Sullivan & Vot
kl
1999).
Irsan
(200 1, 200 1 a) melaporkan bahwa, pamsitisasi hiperpami toid pada parasitoid yang memarasit kutudaun rnencapai lebih dari 90%. Hiperparasitoid juga dapat menurunkan keanehagman dan kelim phan parasitoid ( Waage 1 99 1 ). Parasitoid yang sudah mapan tin@ di suatu habitat kutudaun dapat terganggu oleh kehadiran hiperparasi toid (Altieri et ul. 1997).Hiperparasitoid yang memarasit parasitoid pada kutudaun antara lain tergolong ke dalam genus AIIoq~ta (=Char@), Lyfovsta, Phaenoglyphi (Cynipoidea: Cyni pidae); Dendrocerus (=Lygocem) (Ceraphronoidea: Megaspl idae); Asaphes,
Packneuron, Caruna, Euneura (Chalcidoidea: C halcididae), Aphidencyrt us
(Chalcidoidea: Encyrtidae) clan Tetrasfichus (Chalcidoidea: Eulophidae). Hiperpamitoid itu ada yang bersifat endoparasit dan ektoparasit (Hagen & van den Bosch 1968, SuHivan 1988, Sullivan & Volkl 1999). Sullivan (1988) menyatakan bafiwa, imago hipepmitoid tersebut ada yang meletakkan telur sebelum kutudaun rnenjadi mumi dan ada yang meletakkan telur setelah kutudaun menjadt mumi.
Hiperparasitoid juga bermanfaat dari sudut pengendalian hayati, apabila hiperparasitoid itu (parasitoid tersier) memarasit hi perparasitoid (parasitoid sekunder) yang lain (Sullivan 1 988). Menurut Rosenheim (1 998), hiperparasitoid j uga dapat meningkatkan aktivitas parasitoid mencari koloni kutudaun. Peningkatan a h fitas itu d a p t terjadi karena bila ada kehadim hiperpamitoid, imago parasitoid a k . segem pergi meninggalkan koloni kutudaun pertama dan m e n d koloni kutudaun yang lain sebelum memarasit
mn$
kutudaun di koloni pertama. Perilaku imago parasitoid diTumbuhan Inang, Kutudnun, Parasitoid dan Hiperparasitoid
Kolonisasi kutudaun di tumbuhan hang erat kaitannya dengan spesies, fenologi, kondisi fisiologis dan sifat fisik rnaupun kimia tumbuhan hang itu (Awmack & Leather 2002). Kesesuaian tumbuhan inang dengan kutudaun dan kondisi
lingkungan yang mendukung serta tekanan pamitoid merupakan faktor yang menentukan kolonisasi kutudaun. Di samping itu kelim- popufasi kutudaun juga di pengaruhi oleh faktor biotik be- parasitisasi parasitoid dan hi perparasitoid (Godfray & Muller 19981, serh faktor abiotik yang terdiri dari musim (Dixon & KindImann 1998), dan penggunaan insektisida (Lowery & Isman 1994). Kinerja
parasitoid dalam mengendalikan popuiasi kutudawl dipenganrhl, antara lain oleh pmasitisasi hiperpamitoid (Sullivan 1 988). Pamsitisasi hi perparasi toid merupakan faktor penting yang menurunkan kelimpahan populasi ~ i t o i d yang memarasit kutudaun (lrsan 200 1 ).
Interaksi antara kutudaun dengan tumbuhan inang serta kebe- parasitaid dan hiperparasitoid serta lingkungan yang terdiri dari musim tanam dan penggunaan
insektisida dapat dili hat pad Gambar 2.1. Mengetahui dan memahami interaksi
kutudaun dengan tumbuhan inang &n parasitoid serta hiperprtrasitoidnya pada suatu
Hiperparasitoid
I
Parasitoid
m
Tumbuhan Iuang
-
Spesies1
-
Fenologi-
Kondisi fisiologis1-
-
Sifat fisiklkimiaKutudaun
-
Kolonisasi-
Perilaku1
-lostar-
Ukuran-
Kondisi fisiologis [image:40.620.111.512.103.421.2]musirn dan insektisida
Gambar 2.1 Faktor yang mempengaruhi kehidupan Myzus persicae pada tumbuhan Inang.
Iklim dnn Kutudauu
IHim merupakan faktor penting yang mempengaruhi kelimpahan populasi serangga khususnya kutudaun (Harington et al. 1995; WeIlington el a/. 1999). Rlim
wilayah tropis terdiri dari musim hujan dan musim kemarau, keduanya sangat berpengaruh pada populasi kutudaun (Kranz et al. 1978). C d hujan tinggi &pat menyebabkan populasi kutudaun rendah bahkan hilang h e m kutudaun itu tejatuh oleh deraan air hujan dan hanyut terbawa aliran air hujan pixon 1985). Pada rnusim
Parasitoid (Dixon & Kindlmann 1998; Godfray & MuIIer 1998). Oleh karena itu infmasi popdasi kutudaun dan pamitoidnya yang berhut>unp dengm musim hujan dan musim kemarau menjadi penting ddam m e ~ n Program b
pengendalian kutudaun secara hayati di wilayah tropis.
Altieri MA, Cure JR, Garcia MA. 1997. The role and enhancement of parasitic Hymenoptera biodiversity in agroecosysterns. Di &lam: LaSdle J, Gauld JD, editor. Hymenoptera and Biodiversity. Wallingford: CAB hternasiod
.
hlm 257-275.Awrnack CS, Leather SR. 2002.
Host
plant quality and fecundity in herbivorous insects. Annu Rev Entomol47: 8 1 7-844.Barbosa
P,
Benrey B. 1998. The influence of plant on insect pamitoid: implications for conservation biological control. Di dalam: Barbosa P, editor. Conservation Biological Control. San Diego: Academic Press. hlm 55-83.Bergman JM, Tingey
MW.
1979. Aspects of interaction &ween plant genotypes and biological control. Esa Bull 25(4): 275-279.Blackman RL, Eastop VF. 1994. Aphidr on the World's Trees: an Iderrtflcation and Information Guide. Wallingford: CAJ3 International.
Blackman RL, Eastop VF. 2000. Aphids on the World S Crops: an Ident@cafion and Information Guide. Second edition. Chichester: Wiley.
Bouchard Y, Cloutier C. 1984. Honeydew as a source of host-searching kairomones for the aphid parasitoid Aphidiw nigripes (Hymenoptera: Aphidiidae) Can J
Zoo 62: 254-255.
Brewer
UI,
Elliott NC. 2004. Biological control of cereal aphids in North America and mediating effects of host plant and habitat manipulation. Annu RevEntomol49:Z 19-242.
DeBach P. 1979. Biological Co