Peranan Kadar C3 dan C4 pada Penderita Diabetes Mellitus
Abdul Gani
Bagian/SMF Ilmu Penyakit Dalam Fakultas Kedokteran UNSYIAH RSUD. dr. Zainoel Abidin, Banda Aceh
Abstrak: Sistem komplemen merupakan salah satu mediator utama reaksi radang, dan penting perananya dalam pertahanan tubuh menghadapi infeksi. Aktivitas biologiknya adalah sebagai anafilatoksin, ikut dalam netralisasi virus, khemotaksis lekosit dan opsonisasi, merangsang granulositosis dan pembentukan antibody serta limfokin, mempertinggi sitotoksis sel K(killer cell), inaktifasi endotoksin, dan pada akhirnya menyebabkan lisisnya mikroorganisme. Pada penderita dengan defisiensi komponen komplemen dilaporkan lebih mudah terkena infeksi atau penyakit “kolagen vaskuler”. Demikian pula pada defisiensi komponen komplemen ketiga (C3) dan komponen ke empat (C4). Diduga terjadinya penyakit “kolagen vaskuler” juga didahului dengan infeksi.
Kata kunci: C3 and C4, Diabetes Mellitus
Abstract: The complement system in one of the principal mediators of the inflammatory respon and thereby, serves as an essential function in host defense against infection. The biologic activites of the complement are virus neutralization, anaphylatoxin, chemotaxis of leucocytes, opsonization, enhacing induction of antibody formation, stimulating lymphokine production, enhancing killer cell citotoxicity, inducing granulocytosis, endotoxin inactivation and lysis of microorganisms. Patients with complement deficiencies such as C3 and C4 were reported to have had a high incidence of infections or “collagen vascular disease”. The reason of the occurrence of the “collagen vascular disease” was not known, but it might have been originated by infections. Keywords: C3 and C4, Diabetes Mellitus
PENDAHULUAN
Terdapat bukti bahwa komplemen (C) ikut berperan dalam pertahanan tubuh menghadapi infeksi. Defisiensi komponen komplemen ketiga (C3) dilaporkan sebagai yang bertanggung jawab atas bertambahnya kepekaan terhadap infeksi. Diduga defisiensi komponen komplemen (C4) juga memberi pengaruh yang serupa. Adanya gangguan fungsi yang berkaitan dengan aktivitas komplemen seperti fagositosis (opsonisasi), khemotaksis dan pembentukan anafilatoksin diduga yang menjadi sebab turunnya pertahanan tubuh. Diabetes Mellitus (DM) yang terawat jelek (poorly controlled) akan menurunkan efisiensi pertahanan tubuh.
Dengan perawata DM yang baik (well controlled) ketahanan penderita terhadap
infeksi dapat ditingkatkan.(1,2)
Adapun keseluruhan sebab yang menjelaskan tinggi insidens dan beratnya infeksi pada penderita DM masih banyak belum terungkap. Terdapat laporan bahwa fagositosis dan khemotaksis lekosit dan aktivitas opsonisasi serum
penderita DM menurun.(3)
Peristilahan dalam Sistem Komplemen
Dari komponen-komponen sistem komplemen, sebanyak 9 buah disingkat dengan huruf C besar dan di belakangnya diberi angka sesuai dengan urutan ditemukannya, yakni C1,C2,C3,C4 dan seterusnya sampai C9. Komponen pertama (C1) terdiri dari 3 subkomponen, C1q,C1r,C1s. Garis datar di atas angka merupakan tanda komponen yang aktif, ditandai dengan huruf kecil a, b, c atau d. Huruf a menandai pecahan kecil yang dibebaskan kecairan sekitarnya, sedangkan huruf b menandai pecahan besar yang berikatan dengan komponen aktif yang memecahnya atau dengan suatu komplek imun, sebagai contoh C3a dan C3b. Dalam hal ini C2 merupakan perkecualian, C2a
adalah pecahan yang besar(1,2,4)
. Sistem komplemen akan menjadi aktif lewat 2 jalur, yakni jalur klasik dan jalur alternatif (jalur properdin).
Dikenal 3 komponen lain yang terlibat dalam aktifasi jalur alternatif, yakni faltor B, D dan P (properdin), bila aktif ditulis B, D dan P.
Selain itu terdapat 2 komponen yang berfungsi sebagai inaktifator, yakni C1 Ina dan C3 Ina (1,2,3).
Mekanisme Kerja Sistem Komplemen
Prinsip-prinsip berikut ini merupakan dasar untuk memahami fungsi komplemen (1,2,3).
1. Sistem komplemen termasuk salah satu
mediator reaksi radang. Sistem tersebut merupakan interaksi dari komponen-komponen yang berwujud protein.
2. Komponen-komponen yang berinteraksi
secar runtut (cascade), aktifasi tiap komponen berlangsung setelah aktifasi komponen sebelumnya.
3. Interaksi pada jalur klasik berlangsung
dengan urutan “antigen antibodi – C 142356789”. Sedangkan untuk jalur alternatif urutannya ialah “aktifator – (antibodi) – sistem properdin – C356789”; dalam hal ini peranan antibodi masih dipertanyakan. Kedua jalur teresbut saling berinteraksi; di samping itu juga berinteraksi dengan sistem pembekuan darah.
4. Interaksi 5 komponen pertama (C14235)
bersifat enzimatis, aktifasi berarti merubah
komponen menjadi enzim. Pada mulanya sebagai hasil interaksi dengan antibodi, C1 menjadi aktif. Selanjutnya C4, C2, C3, C5 dan begitu pula dengan B pada jalur alternatif aktifasinya secara berurutan dilakukan oleh komponen sebelumnya yang telah aktif. Adapun antara C5b, C6, C7, C8 dan C9 berinteraksi dengan saling berikatan satu sama lain dalam bentuk ikatan nonkovalen atau ikatan hidrofobi; dengan demikian sifat molekul yang terbentuk sebelumnya berubah.
Aktifasi Jalur Klasik dan Jalur Alternatif Jalur klasik
SKEMA 1. menguraikan tahapan komponen komplemen berinteraksi. Sebagai standard in vitro untuk mempelajari interaksi pada jalur klasik adalah eritrosit dan antibodinya (4,5)
. Aktifasi jalur klasik dimulai
dengan fiksasi C1q pada bagian Fc antibodi (1)
. CRP (C-reactive protein) yang terikat mikroorganisme juga dapat mengikat C1q
untuk memulai rangkaian reaksi aktifasi (4)
. Reaksi terakhir ini sangat penting sebagai pencetus reaksi radang pada saat antibody dari antigen tersebut belum terbentuk. Aktifasi C4 dan C2 menghasilkan fragmen yang sifatnya menyerupai kinin, menaikkan permeabilitas
venula post-kapiler, menyebabkan edema(5)
. Fragmen C4b difiksasi membentuk kompleks EAC14b yang berkemampuan untuk menempel ke berbagai sel, diantaranya ke neutrofil, monosit dan eritrosit; fenomena ini
disebut “immune adherence” (2,3)
. Pemecahan C3 dilakukan oleh kompleks EAC142 (“C3 convertase” jalur klasik). Fragmen C3b yang terfiksasi pada kompleks molekul tersebut kemudian menempel pada sel yang mempunyai reseptor C3b, seperti limfosit B, eritrosit dan fagosit (neutrofil, monosit, makrofag). Dengan cara seperti ini fagositosis menjadi efesien. Oleh C3bIna fragmen C3b diinaktifkan menjadi C3d yang tetap terikat pada kompleks malekul, dan dilepaskan C3c
(3)
. Fragmen C3a dan C5a bersifat sebagai anafilatoksin, mempengaruhi sel mast dan basofil mengeluarkan mediator vasoaktif. Ada pula fragmen C3 dan C5 yang merupakan khemotaksis fagosit, dan masing-masing sifat
fisikokimiawinya menyerupai C3a dan C5a (2,3).
A=antibodi, C-CRP= Kompleks C- reactive protein.
Pengrasakan membran sel (membrane attack) sebagai permulaan sitolisis dimulai setelah C5b terikat pada C423. Selanjutnya C6 tanpa terpecah berikatan dengan C5b, berfungsi menstabilkan C5b. Pada tahap berikutnya kompleks C5b6 terlepas dari C423 dan bereaksi dengan C7. Agar kemampuan lisisnya tidak hilang, kompleks C5b67 harus segera berikatan dengan membran sel; kompleks C5b67 yang bebas bersifat khemotaksis. Kemudian dengan terikatnya C8 dan C9 pada kompleks tersebut terjadilah sitolisis(2,3)
. Suatu mekanisme kontrol mencegah aktifasi komplemen yang berlebih-lebihan. C1 Ina menghambat kerja esterolisis
C1s--Æ memecah C4 dan C2. Dalam 55 detik
sebanyak 90% C1 telah terikat pada C1 Ina (6)
.
Pada suhu 37 0
C waktu paruh C2 hanya sekitar 8 menit, hal ini menjadi pembatas waktu efektif C42 dan C423. Aktivitas C3a dan C5a akan terhenti setelah dipecah anafilaktoksin inaktifator yang beredar di dalam serum. C3b Ina memecah C3b, C4b, dan mungkin juga C5b menjadi fragmen inaktif (1,2,3,6)
.
Jalur alternatif (properdin)
Permulaan aktifasi berupa interaksi antigen (polisakharida atau liposakharida) atau
agregasi imunoglobulin dengan IF (initiating
faktor) yang belum diketahui(7)
; mungkin pula
berupa interaksi antigen antibodi(8)
. Diduga dalam aktifasi ini C1 atau C1s terlibat, namun
tanpa mengikut sertakan C4 dan C2 (6,7)
. Sebagai hasil interaksi tersebut P dan D menjadi P dan D. Selanjutnya D memecah B menjadi Bb dan Ba; pemecahan ini membutuhkan Mg ++ dan C3b atau CoVF. Kompleks C3bBb atau CoVFBb yang kemudian terbentuk merupakan “C3 convertase”. Dilaporkan bahwa enzim proteolisis (plasmin, tripsin, pronase) juga
berfungsi seperti D (5,6)
. Dengan terikatnya properdin (P) pada kompleks C3bBb, kerja C3b Ina dihambat dan kompleks tersebut
menjadi lebih stabil (7)
. Pada aktifasi sistem properdin ini, C3b dapat diperoleh dari hasil aktifasi jalur klasik, atau dari pemecahan C3 oleh trombin dan plasmin pada koagulasi darah, protease lekosit, dan tripsin. Seperti terlihat pada skema.2, C3b lewat alur amplifikasi merangsang terbentuknya C3b lebih banyak; pengaturan amplifikasi ini
dilakukan oleh C3b Ina (6)
.
Skema 2. Aktifasi komplemen pada jalur alternatif (Johnston dkk. 1989).
Ab = antibodi; Ig = imunoglobulin; Ag = antigen; IF = initiating faktor; CoVF = cobra venom factor.
Sistem komplemen
Klasik Alternatif
Amplifikasi C3b
C3a Anafilatoksin
Fagositosis Immune adherence Terikat sel B EAC1423b
C5
Polisakharida
Fagositosis Immune adherence Terikat sel B Ag(Ab)C36Bb (P)
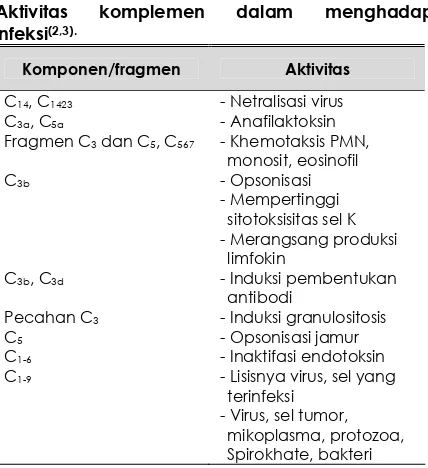
Aktivitas komplemen menghadapi infeksi Tabel 1 di bawah ini merupakan ringkasan aktivitas komplemen menghadapi infeksi (2,4)
. Terdapat bukti-bukti bahwa aktivitas C1 dan C4 menetralkan infektifitas virus (5,6,7)
. Dalam netralisasi tersebut diperlukan fiksasi oleh C5, C6 dan terutama C3b. Fiksasi ini sangat bermanfaat selagi kadar anti bodi masih rendah, yaitu pada permulaan infeksi (7,8,9)
Tabel 1.
Aktivitas komplemen dalam menghadapi infeksi(2,3).
Komponen/fragmen Aktivitas
C14, C1423 - Netralisasi virus
C3a, C5a - Anafilaktoksin
Fragmen C3 dan C5, C567 - Khemotaksis PMN,
monosit, eosinofil
C3b - Opsonisasi
- Mempertinggi sitotoksisitas sel K - Merangsang produksi
limfokin
C3b, C3d - Induksi pembentukan
antibodi
Pecahan C3 - Induksi granulositosis
C5 - Opsonisasi jamur
C1-6 - Inaktifasi endotoksin
C1-9 - Lisisnya virus, sel yang
terinfeksi - Virus, sel tumor,
mikoplasma, protozoa, Spirokhate, bakteri
Sel mast dan basofil yang berikatan dengan C3a dan C5a akan mengeluarkan mediator vasoaktif dan terjadilah vasodilatasi dan keradangan. Fragmen yang berasal dari C3 dan C5 juga memudahkan influks netrofil,
monosit dan eosinofil(3)
. Diduga C3b
menggiatkan fagositosis (opsonisasi) dengan mempengaruhi ikatan antara antibodi dengan
reseptor Fc fagosit (6)
, kemudian opsonisasi menjadi inaktif setelah C3b terpecah menjadi C3d(7)
. C3b juga diperlukan untuk mempertinggi daya sitolisis sel K (killer cell), bekerja pada ikatan antara”antibody-coated
target cell” dengan reseptor Fc sel K (6)
. Dalam kegiatannya, limfosit B dan T ditunjang sistem komplemen C3 menginduksi pembentukan
antibodi (8)
; C3b merangsang produksi limfokin (6)
. C4 berperan pada fase pengenalan benda asing, terbukti dengan penambahan antibodi terhadap C4 replikasi limfosit setelah
dicampur sel asing (6,7,8)
. Fragmen C3 dilaporkan dapat menginduksi granulositosis. Aktivitas C3 terutama yang lewat jalur alternatif, menghasilkan faktor-faktor yang dapat meninggikan kapasitas fagositosis
makrofag (5)
. C5 diperlukan dalam proses fagositosis fungi dan fagositosis lanjutan dari
“C3-coated bacteria” (6)
. Aktifasi komplemen sampai C6 menetralkan efek letal endotoksin
(6,7)
. Sebagai hasil akhir aktifasi komplemen ialah lisisnya virus, sel yang terinfeksi virus, sel tumor, mikoplasma, protozoa, spirokheta dan beberapa strain bakteri(1,2,3,6)
.
Kelainan Sistem Komplemen
1. Jalur klasik
Kelainan sistem komplemen dibagi
menjadi defisiensi komponen komplemen
kengenital dan defisiensi dapatan (6)
. Defisiensi kongenital dari semua komponen telah
dilaporkan, terkecuali defisiensi C9 (2,3,6)
vaskuler”. Defisiensi komplemen kongenital yang dihubungkan dengan infeksi ialah defisiensi C3 (9)
, C5 (6)
, C6 (8)
, C8 (9)
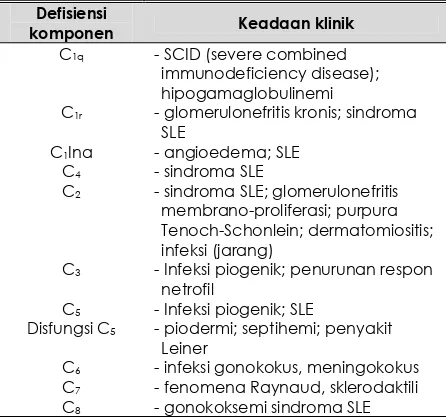
. Beberapa kali septikhemi dialami 2 penderita defisiensi C2, namun sebagian besar dari penderita ini tanpa problem infeksi (2). Keseluruhan keadaan klinik defisiensi komplemen kongenital ini dapat dilihat pada Tabel 2 (2,3)
.
Tabel 2.
Defisiensi komplemen kongenital: kelainan pada jalur klasik (2,3)
Defisiensi
komponen Keadaan klinik
C1q - SCID (severe combined
immunodeficiency disease); hipogamaglobulinemi
C1r - glomerulonefritis kronis; sindroma
SLE
C1Ina - angioedema; SLE
C4 - sindroma SLE
C2 - sindroma SLE; glomerulonefritis
membrano-proliferasi; purpura Tenoch-Schonlein; dermatomiositis; infeksi (jarang)
C3 - Infeksi piogenik; penurunan respon
netrofil
C5 - Infeksi piogenik; SLE
Disfungsi C5 - piodermi; septihemi; penyakit
Leiner
C6 - infeksi gonokokus, meningokokus
C7 - fenomena Raynaud, sklerodaktili
C8 - gonokoksemi sindroma SLE
Karena aktifasi C3 dapat lewat jalur klasik ataupun jalur alternatif, maka bila salah satu jalur terganggu dapat dikompensasi jalur yang lain. Sebagai akibat defisiensi C3, fragmen khemotaksis dari C3 dan C5 tidak terbentuk dan dengan demikian fagositosis
menjadi inefisien (10)
. Hal ini menjelaskan terjadinya infeksi piogenik pada penderita defisiensi C3. Dua dari tiga penderita
defisiensi C3 yang dilaporkan(6)
, respon netrofilnya menurun.
Keterangan tentang hubungan defesiensi komplemen dengan tingginya insidens penyakit “kolagen vaskuler” masih belum jelas. Diduga infeksi sebagai proses pertama dan karena penghalauan kompleks imun terganggu
maka timbullah penyakit tersebut(2)
.
2. Jalur alternatif.
Penderita infeksi piogenik berat yang mulai pertama dilaporkan sebagai penderita defisiensi C3 (6,10)
, ternyata kelainan dasarnya adalah hiperkatabolisme C3 akibat defisiensi C3bIna (2,3,6)
. Pemberian infus plasma atau
C3bIna memulihkan kadar C3 (2,3,4,11)
. Bentuk lain defisiensi C3 karena hiperkatabolisme ditemukan pada penderita lipodistrofi partial dengan nefritis membranoproliferasi (ginjal),
dan penderita dengan infeksi piogenik(6,11)
. Diduga C3 dipecah oleh “C3 nefritic factor”/C3NeF(ginjal). Gangguan aktifasi jalur alternatif yang disertai infeksi juga ditemukan pada penderita “sickle cell disease”(6)
dan panca splenektomi (9)
Sistem Komplemen pada Diabetes Mellitus Pada penelitian yang telah dilakukan, mendapatkan bahwa aktivitas hemolitik total dari komplemen serum penderita diabetes mellitus pada berbagai tingkat regulasi tidak berkurang (8,9,12)
. Hal ini tidak berarti bahwa kadar tiap-tiap komponen komplemen tidak berkurang(12)
. Pada penderita diabetes mellitus tipe 1 mendapatkan bahwa kadar C3 untuk yang mengindap diabetes mellitus kurang dari satu tahun cenderung menurun dan meningkat pada penderita yang telah sakit 1–3 tahun dan lebih 5 tahun. Kadar C4 menurun pada penderita yang mengidap kurang dari 1 tahun dan cenderung turun pada penderita yang telah mengidap antara 1–3 tahun. Pola tersebut dihubungkan dengan ICA (islet cell antibody) yang bersifat mengikat komplemen. Berbeda dengan diabetes mellitus tipe 11, tipe
1 ini diduga sebagai penyakit otoimun(3,11,12)
. Dalam hal ini “complement-fixing ICA”) dianggap yang bertanggung jawab terhadap kerusakan sel beta(3,9,11,12)
. Triolo G dkk (1991) melaporkan bahwa tingginya prevalensi ICA pada diabetes mellitus tipe 1 ada hubungannya dengan lama mengidap sakit; 60% untuk yang mengidap kurang dari 1 tahun; 20% untuk yang 2 – 5 tahun dan hanya 5% untuk yang lebih dari 5 tahun.
KESIMPULAN
secara keseluruhan, apa yang menjadi sebab lebih rentannya penderita diabetes mellitus pada infeksi masih belum jelas. Adapun untuk mendapatkan daya tahan tubuh yang optimal, regulasi gula darah sangat penting.
DAFTAR PUSTAKA
1. Ahmed AEE, Peter JB. Clinical utility of
complement assessment. Clin Diagn Lab Immun 1995;2:509-17
2. Coupes BM, Ken SP, Brenchly PEC,
Short CD, Mallick NP. The temporal relationship between urinary C5b-9 and C3dg ang clinical parameters in human membranous nephropathy. Nephrol Dial Transplant 1993;8:397-401.
3. Triolo G, Giardina E, Casiglia D,
Scarantino G, Bompiani GD. Detection of the terminal fluid-phase complement complex SC5b-9, in the plasma of patients with insulin-dependent (type.1) diabetes mellitus: Relation to increased urinary excretion and plasma von Wilebrand factor, Clin Exp Immunol 1991;84:53-8.
4. Gabrielsson BG, Johansson JM, Lonn M,
Jernas M, Olbers T, Peltonen M, Larsson I, Lonn L, Sjostrom L, Carlsson B, Carlsson LM. High expression of complement components in omental adipose tissue in obese men. Obes Res 2003;11:699-708.
5. Muscari A, Bozzoli C, Puddu GM,
Sangiorgi Z, Dormi A, Rovinetti C, Descovich GC, Puddu P. Association of serum C3 levels with the risk of myocardial infarction. Am J Med 1995;98:357-364.
6. Johnston RB, Newman SL, Struth AG.
An abnormality of the alternate pathway of complement activation in sickle cell disease Am, J Med 1989;288:303-308
7. Peake PW, Kriketos AD, Camphell LV,
Charlesworth JA. Response of the alternative complement pathway to an oral fat load in first degree relatives of subjects with type ll diabetes. Int J Obes Relat Metab Disord Apr.27,.2004[Epub ahead of print].
8. Muscari A, Massarelli G, Bastagli L,
Poggiopollini G, Tomassetti V, Drago G, Martignani C, Pacilli P, Boni P, Puddu P. Relationship of serum C3 to fasting insulin, risk factors and previous ischaemic events in middle aged men. Eur Heart J 2000;231:1081-1090.
9. Weyer C, Tataranni PA, Pratley RE.
Insulin action and insulinemia are closely related to the fasting complement C3, but not acylation stimulating protein concentration. Diabetes Care 2000; 23:779-785.
10. Van Harmelen V, Reynisdottir S,
Cianflone K, Degerman E, Hoffstedt J, Nilsell K, Sniderman A, Arner P. Mechanisms involved in the regulation of free fatty acid release from isolated human fat cells by acylation-stimulating protein and insulin. J Biol Chem 1999;274: 18243-18251.
11. Ford ES. Leukocyte count, erythrocyte
sedimentation rate, and diabetes incidence in a national sample of US adults. Am J Epidemiol 2002; 155:57-64.
12. Pickup JC, Crook MA. Is type ll diabetes