i
SELEKSI PADI HIBRIDA TERHADAP KEKERINGAN
UNTUK PENGEMBANGAN DI LAHAN
SAWAH TADAH HUJAN
LA ODE AFA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
iii Dengan ini saya menyatakan bahwa disertasi yang berjudul “Seleksi Padi Hibrida terhadap Kekeringan untuk Pengembangan di Lahan Sawah Tadah Hujan” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Januari 2013
v Lowland. Under supervision of: BAMBANG S. PURWOKO as chairman, AHMAD JUNAEDI, OTENG HARIDJAJA and ISWARI S DEWI as members of the advisory committee.
The objective of the research was to determine method and character for early selection of rice to drought and to obtain drought tolerant hybrid rice genotypes to be used further in rainfed lowland. Experiments were conducted from April 2011 until July 2012 at Seed Science and Technology Laboratory, IPB and BB Biogen greenhouse Cimanggu, Bogor, at Babakan Experiment Station, IPB, Bogor, and at irrigated lowland, Cikembar, Sukabumi, and rainfed lowland, Gantar, Indramayu. The study consisted of seven experiments. The results showed that: (1) Polyethylene glycol (PEG) 6000 could detect and differentiate the response of hybrid rice genotypes to drought stress in both germination and seedling stage. The use of 25% PEG 6000 in the seedling stage could detect early hybrid rice genotypes tolerant to drought and have a compatibility with grouping of genotypes tolerant to drought based on yield criteria in the field. Variables of root dry weight, shoot dry weight and leaf dryness score were to detect hybrid rice genotypes tolerant to drought at seedling stage. (2) Genotypes BI485A/BP12, BI485A/BP15 and BI559A/BP15 under drought stress conditions decreased root length less (17.6 - 26.9 percent) than the IR64, the sensitive check variety (33.6 percent). Their grain weight, harvest index and grain yield (2.82, 2.58 and 2.72 ton ha-1) were higher than other genotypes. Moreover they were earlier to harvest, shorter in grain filling period and had sensitivity index to drought ≤ 0.50. Hybrid genotype tolerant to drought i.e BI599/BP15 under drought stress conditions had less stomatal density, maintained higher leaf relative water content, and contained non-structural carbohydrates in stem, sheath and leaf less than other genotypes. Genotypes BI485A/BP12, BI485A/BP15 and BI559A/BP15 were tolerant to drought and potentially could be developed for rainfed lowland; (3) The yield of these genotypes under irrigated lowland were not significantly different from those of Ciherang and Hipa 7 (i.e. 5.63, 6.87 and 6.30 ton grain ha-1, respectively). BI599A/BP15 genotypes under severe drought at rainfed lowland yielded 0.90 ton grain ha-1, whereas Hipa 7 and Limboto reached 0.34 and 0.29 ton grain ha-1, respectively.
vii di Lahan Sawah Tadah Hujan. Dibimbing oleh BAMBANG S. PURWOKO sebagai ketua, AHMAD JUNAEDI, OTENG HARIDJAJA dan ISWARI S. DEWI sebagai anggota komisi pembimbing.
Salah satu masalah yang dihadapi dalam peningkatan produksi beras nasional adalah meningkatnya alih fungsi lahan subur dan produktif. Peningkatan produksi dapat diarahkan ke lahan sawah tadah hujan. Di lahan sawah tadah hujan, kekeringan terjadi hampir setiap tahun akibat jumlah curah hujan yang rendah dan pendeknya musim hujan. Padi hibrida diharapkan tetap menunjukkan hasil yang lebih baik pada kondisi kekeringan dibanding varietas unggul inbrida. Salah satu strategi yang dapat dilakukan adalah pengembangan genotipe padi hibrida toleran cekaman kekeringan dengan potensi hasil tinggi dan berumur genjah. Genotipe tersebut diperoleh melalui serangkaian tahapan seleksi. Seleksi terhadap bahan genetik dalam jumlah besar, membutuhkan banyak biaya, tenaga dan waktu, oleh karena itu, perlu didukung metode seleksi yang efektif dan efisien.
viii
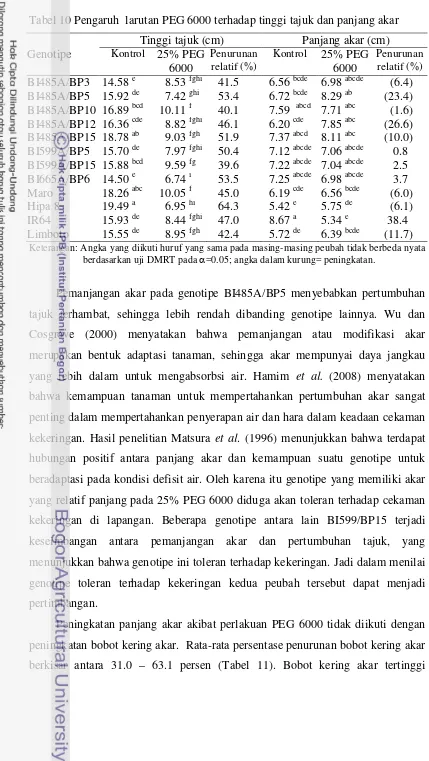
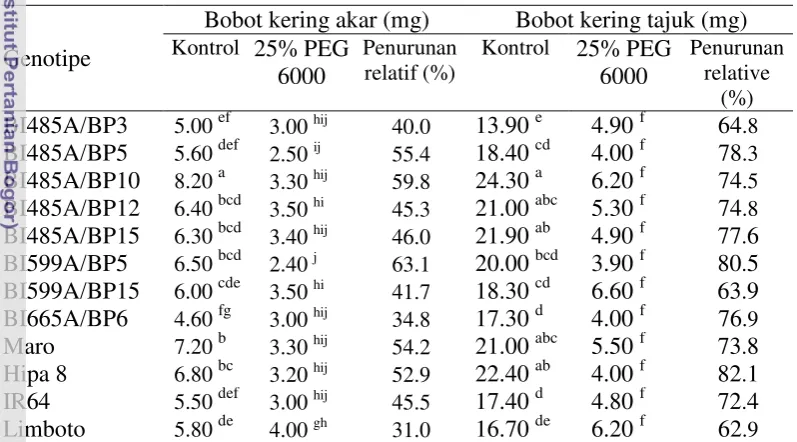
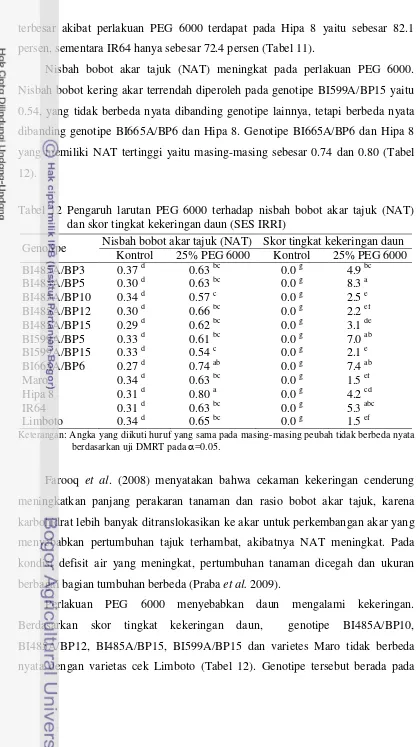
kekeringan berdasarkan karakter bobot kering akar, bobot kering tajuk dan skor tingkat kekeringan daun. Percobaan ke empat yaitu pengujian dengan perlakuan cekaman kekeringan di pot. Percobaan menggunakan rancangan split plot, 3 ulangan. Petak utama adalah perlakuan cekaman kekeringan yang terdiri atas tanpa cekaman kekeringan dan cekaman kekeringan 60% kapasitas lapangan. Anak petak adalah 12 genotipe/varietas (sama dengan percobaan ke dua dan ke tiga). Hasil penelitian menunjukkan bahwa berdasarkan kriteria hasil di pot, genotipe BI485A/BP5, BI485A/BP10, BI485A/BP12, BI485A/BP15 dan BI559A/BP15 dikelompokkan toleran kekeringan.
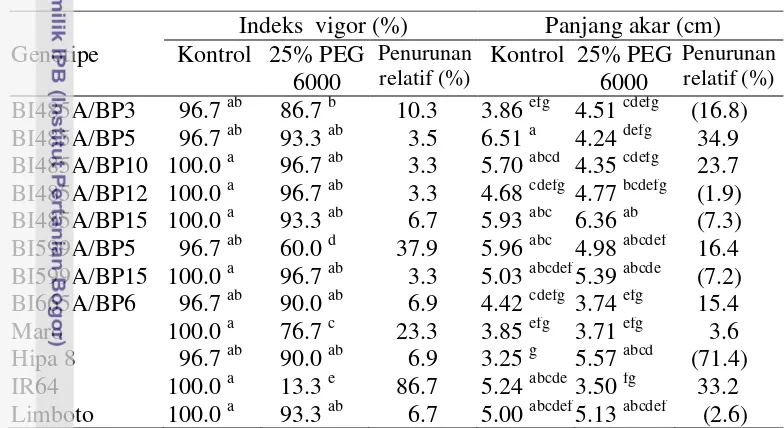
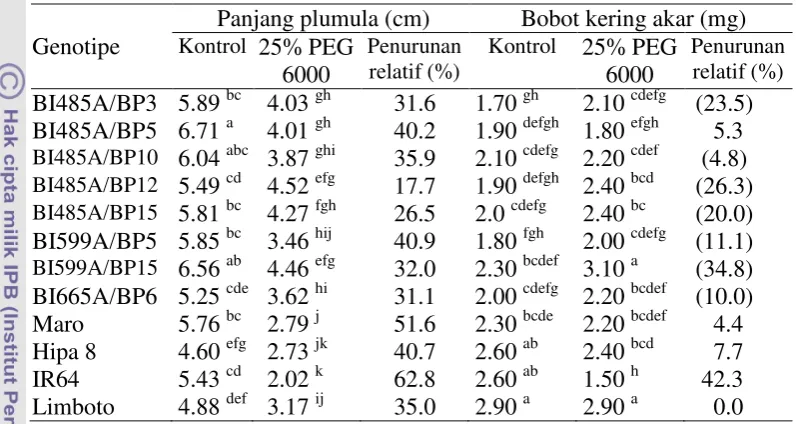
Penelitian ke lima bertujuan mengetahui respon agronomi, morfologi dan fisiologi genotipe padi hibrida terhadap simulasi cekaman kekeringan di lahan sawah dan mendapatkan genotipe hibrida yang berpotensi dikembangkan di lahan sawah tadah hujan telah dilaksanakan di Laboratorium Lapangan Riset Padi Babakan, University Farm IPB, Bogor, pada bulan Desember 2011 sampai Maret 2012. Penelitian menggunakan rancangan split plot, 3 ulangan. Petak utama adalah perlakuan cekaman kekeringan yang terdiri atas tanpa cekaman kekeringan dan cekaman kekeringan pada akhir fase vegetatif hingga dua minggu setelah antesis. Anak petak adalah genotipe/varietas terdiri atas genotipe BI485A/BP3, BI485A/BP5, BI485A/BP10, BI485A/BP12, BI485A/BP15, BI599A/BP5, BI599A/BP15, BI665A/BP6, varietas Maro, Hipa 6, Hipa 7, Hipa 8, IR-64 dan Limboto. Hasil penelitian menunjukkan bahwa genotipe BI485A/BP12, BI485A/BP15 dan BI559A/BP15 pada kondisi cekaman kekeringan mengalami penurunan panjang akar relatif kecil (17.6 – 26.9 persen) dibanding dengan varietas cek peka IR64 (33.6 persen), menghasilkan bobot gabah per rumpun, indeks panen dan daya hasil gabah per hektar (masing-masing sebesar 2.82, 2.58 dan 2.72 ton ha-1
Penelitian ke enam dan ke tujuh bertujuan mengetahui daya hasil pendahuluan genotipe padi hibrida toleran kekeringan di lahan sawah irigasi dan saat mengalami kekeringan di lahan sawah tadah hujan. Penelitian ini dilakukan: (i) di lahan sawah irigasi Desa Bojong, Kecamatan Cikembar, Sukabumi, pada bulan November 2011 sampai Februari 2012 dan (ii) di lahan sawah tadah hujan Desa Sanca, Kecamatan Gantar, Indramayu, pada bulan April sampai Juli 2012. Penelitian menggunakan rancangan acak kelompok, 3 ulangan.Percobaan di lahan sawah irigasi menggunakan 8 genotipe padi hibrida (BI485A/BP3, BI485A/BP5, BI485A/BP10,BI485A/BP12, BI485A/BP15, BI599A/BP5, BI599A/BP15 dan BI665A/BP6) dan 2 varietas cek (Hipa 7 dan Ciherang). Hasil penelitian menunjukkan bahwa genotipe BI485A/BP12, BI485A/BP15 dan BI559A/BP15
ix BI485A/BP15 dan BI599A/BP15) dan 3 varietas cek (Hipa 7, IR64 dan Limboto). Genotipe BI599A/BP15 pada tingkat kekeringan parah mampu menghasilkan gabah 0.90 ton ha-1, sedangkan Hipa 7 (varietas hibrida yang sudah dilepas untuk lahan sawah tadah hujan) dan Limboto (varietas cek toleran kekeringan) hanya menghasilkan masing-masing 0.34 dan 0.29 ton ha-1
Berdasarkan hasil 7 set percobaan maka dapat ditarik kesimpulan bahwa PEG 6 000 konsentrasi 25% pada fase bibit dapat digunakan untuk mendeteksi secara dini padi hibrida/inbrida toleran kekeringan. Peubah bobot kering akar, bobot kering tajuk dan tingkat kekeringan daun merupakan karakter untuk pendugaan padi hibrida/inbrida toleran kekeringan pada fase bibit. Genotipe BI485A/BP12, BI485A/BP15 dan BI559A/BP15 pada kondisi cekaman kekeringan mengalami penurunan panjang akar relatif kecil, daya hasil gabah per hektar yang lebih tinggi serta indeks kepekaan terhadap kekeringan ≤0.50. Genotipe toleran kekeringan (BI599/BP15) pada kondisi cekaman kekeringan memiliki kerapatan stomata yang lebih rendah, tetap mempertahankan kadar air relatif daun yang tinggi serta kandungan karbohidrat non struktural yang lebih rendah pada batang, pelepah dan daun dibanding genotipe/varietas yang peka. Genotipe ini toleran kekeringan dan potensial dikembangkan di lahan sawah tadah hujan. Genotipe BI599A/BP15 yang diuji di lahan sawah tadah hujan pada tingkat kekeringan parah mampu menghasilkan gabah 0.90 ton ha
.
-1
, sedangkan Hipa 7 dan Limboto hanya menghasilkan masing-masing 0.34 dan 0.29 ton ha-1.
xi
©Hak Cipta milik Institut Pertanian Bogor tahun 2013
Hak Cipta dilindungi Undang-Undang
xiii
SAWAH TADAH HUJAN
LA ODE AFA
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor
pada Mayor Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
xiv
Penguji pada Ujian Tertutup : Dr. Ir. Iskandar Lubis, M.S.
(Staf pengajar Departemen Agronomi dan Hortikultura, Fakultas Pertanian IPB)
Dr. Desta Wirnas, SP., M.Si.
(Staf pengajar Departemen Agronomi dan
Hortikultura, Fakultas Pertanian IPB)
Penguji pada Ujian Terbuka : Prof. Dr. Ir. Munif Ghulamahdi, M.S. (Staf pengajar Departemen Agronomi dan Hortikultura, Fakultas Pertanian IPB)
Dr. Suwarno
xv Nama
NIM
: :
La Ode Afa A262090011
Disetujui Komisi Pembimbing
Ketua
Prof. Dr. Ir. Bambang S. Purwoko, M.Sc.
Anggota
Dr. Ir. Ahmad Junaedi, M.Si.
Anggota
Dr. Ir. Oteng Haridjaja, M.Sc.
Anggota Dr. Ir. Iswari S. Dewi
Mengetahui
Ketua Mayor Agronomi dan Hortikultura
Prof. Dr. Ir. Munif Ghulamahdi, M.S.
Dekan Sekolah Pascasarjana
Dr. Ir. Dahrul Syah, M.Sc.Agr.
xvii segala Rahmat dan Karunia-Nya, sehingga penulis dapat menyelesaikan penelitian dan penulisan disertasi yang berjudul ‘Seleksi Padi Hibrida terhadap Kekeringan untuk Pengembangan di Lahan Sawah Tadah Hujan’.
Disertasi ini membahas serangkaian penelitian yang terdiri atas beberapa topik yaitu: (1) Penentuan konsentrasi larutan PEG 6000 yang memberikan cekaman kekeringan, (2) Pengujian dengan 25% PEG 6000 pada fase perkecambahan, (3) Pengujian dengan 25% PEG 6000 dalam kultur hara pada
fase bibit, (4) Pengujian dengan perlakuan cekaman kekeringan di pot, (5) Respon agronomi, morfologi dan fisiologi padi hibrida terhadap cekaman
kekeringan di lahan sawah, (6) Uji daya hasil pendahuluan padi hibrida toleran cekaman kekeringan di lahan sawah irigasi dan (7) Uji daya hasil pendahuluan padi hibrida toleran cekaman kekeringan di lahan sawah tadah hujan. Satu topik penelitian telah dipublikasikan pada Jurnal Agrivigor vol. 11(2):293-300, Mei-Agustus 2012 berjudul “Pendugaan Toleransi Padi Hibrida terhadap Kekeringan dengan Polyetilen Glikol (PEG) 6000”, dan sebagian paper sudah dipresentasikan
pada: (1) Kongres dan Simposium ISSAAS, pada tanggal 7-10 November 2011 di IPB ICC dan Kampus IPB, Bogor, Indonesia, berjudul ”Selection of Drought Tolerance of Hybrid Rice Using Polyethylene Glycol (PEG 6000)”, (2) Seminar
Nasional PERIPI pada Tanggal 6-7 November 2012 di IPB ICC, Bogor, Indonesia, berjudul “Uji Daya Hasil Pendahuluan Genotipe Padi Hibrida Toleran Kekeringan” dan (3) Kongres dan Simposium ISSAAS, pada tanggal 13-16
November 2012 di Bicol University, Legazpi City, the Phillippines, berjudul ”Drought Simulation Effect on Physiology and Agronomy of New Rice Hybrids”.
xviii
tepat waktu; (2) Dr. Ir. Ahmad Junaedi, M.Si selaku anggota Komisi Pembimbing. Dari beliau penulis mendapatkan ide-ide pembanding dan konstruktif terkait penelitian dan penyusunan disertasi ini; (3) Dr. Ir. Oteng Haridjaja, M.Sc selaku anggota Komisi Pembimbing. Dari beliau penulis mendapatkan tentang pemahaman logika berpikir yang benar, ketegasan dan disiplin; dan (4) Dr. Ir. Iswari S. Dewi juga selaku anggota Komisi Pembimbing, atas saran dan ide dalam penelitian disertasi ini. Dari beliau penulis mendapatkan bagaimana memaknai sebuah ide sehingga menjadi lebih bernilai, cara berpikir yang sistematis dan terstruktur. Dengan kecerdasan, keikhlasan, kesabaran dan keluasan wawasan ilmu pengetahuan mereka telah membuka cakrawala berpikir penulis dan memberi sumbangan pemikiran yang amat berharga pada penelitian ini. Semoga sumbangsih yang besar ini menjadi ilmu yang bermanfaat dengan amal jariah yang tak terhingga.
Terima kasih kepada Dr. Ir. Iskandar Lubis, M.S dan Dr. Desta Wirnas, SP., M.Si selaku penguji luar komisi pada ujian tertutup, Prof. Dr. Ir. Munif Ghulamahdi, M.S selaku Ketua Mayor Agronomi dan Hortikultura dan sebagai penguji luar komisi pada ujian terbuka, dan Prof. Dr. Dadang, M.Sc selaku Wakil Dekan Fakultas Pertanian atas semua saran dan koreksi yang konstruktif. Terima kasih pula kepada Dr. Suwarno selaku penguji luar komisi pada ujian terbuka, Dr. Ir. Maya Melati, M.Sc selaku sekretaris Mayor Agronomi dan Hortikultura dan Dr. Ir. Dahrul Syah, M.Sc. Agr. selaku Dekan Pasca Sarjana IPB atas semua saran dan koreksi yang konstruktif.
xix Ilmu dan Teknologi Benih Departemen AGH, IPB atas izin dan bantuannya sehingga penelitian topik 1 dan 2 terlaksana. Kepada kepala Balai BB Biogen, Cimanggu dan staf rumah kaca, disampaikan terima kasih atas izin dan bantuannya sehingga penelitian topik 3 dan 4 terlaksana. Terima kasih penulis sampaikan kepada kepala KP Laboratorium Lapangan Riset Padi Babakan, University Farm, IPB yang telah memberikan izin dan membantu pada penelitian topik 5. Percobaan uji daya hasil pendahuluan (UDHP) di lapangan sangat terbantu oleh Bapak Nurjani di Cikembar, Sukabumi dan Bapak Warlim di Gantar, Indramayu. Penulis menyampaikan apresiasi atas terselesaikannya percobaan 6 dan 7. Penghargaan tinggi kepada Bapak Ir. H. Sutono M.Si atas peminjaman alat pengukur kadar air tanah (TRIME-TDR) dan kepada Bapak Iman dan saudara Joni atas bantuannya selama penelitian. Kepada rekan-rekan mahasiswa pascasarjana AGH terutama AGH 2009 juga disampaikan apresiasi atas kebersamaan dan motivasinya selama studi. Tak lupa diucapkan apresiasi pula pada Forum Mahasiswa Sultra di Bogor atas kebersamaan dan motivasi selama studi.
xx
pengetahuan khususnya di bidang pertanian.
Bogor, Januari 2013
xxi merupakan putra ke dua dari tiga bersaudara, dari Ayahanda L. Waho (Almarhum) dan Ibunda Wa Ode Kaiya (almarhumah). Penulis telah menikah dengan Megawati Asrul Tawulo, S.Sos, M.Si pada tanggal 10 November 2000. Sampai saat ini penulis baru dikaruniai satu orang anak, seorang putri bernama Wa Ode Kaiya Reifana Insyirah (Refa).
Penulis menyelesaikan pendidikan sarjana pada Program Studi Agronomi, Jurusan Budidaya Pertanian, Fakultas Pertanian Universitas Haluoleo, lulus pada tahun 1994. Jenjang strata dua (S2) di Program Studi Agronomi Program Pascasarjana IPB lulus tahun 1998. Selanjutnya, penulis tahun 2009 mengikuti jenjang strata tiga (S3) pada Mayor Agronomi dan Hortikultura, Sekolah Pascasarjana IPB dengan beasiswa BPPS dari Direktorat Jenderal Pendidikan Tinggi, Kementerian Pendidikan dan Kebudayaan Republik Indonesia.
Penulis bekerja sebagai dosen pada Jurusan Agroteknologi Fakultas Pertanian, Universitas Haluoleo, Kendari, Sulawesi Tenggara sejak tahun 2000.
xxiii
Halaman
DAFTAR TABEL……… xvii
DAFTAR GAMBAR……….. xx
DAFTAR LAMPIRAN……….. xxii
PENDAHULUAN ………. 1
Latar Belakang……….. 1
Tujuan Umum ……….. 5
Tujuan Khusus……….. 5
Hipotesis……… 5
TINJAUAN PUSTAKA………. 7
Kendala Budidaya Padi Sawah Tadah Hujan……… 7 Adaptasi Tanaman terhadap Cekaman Kekeringan... 11 Respon Morfologi... 15 Respon Fisiologi... 17 Pemanfaatan Efek Heterosis untuk Toleransi Kekeringan... 21 Metode dan Karakter Seleksi Toleransi Kekeringan... 25 PENDUGAAN DINI TOLERANSI PADI HIBRIDA TERHADAP
KEKERINGAN………... 33
Abstrak………... 33
Abstract………... 33
Pendahuluan………... 34
Bahan dan Metode………... 35 A. Pengujian dengan Larutan Polietilen Glikol (PEG)
6000... 35 A.1. Penentuan Konsentrasi Larutan PEG 6000
yang Memberikan Cekaman Kekeringan... 35 Waktu dan Tempat... 35 Metode Penelitian... 35 Pelaksanaan Percobaan... 36 A.2. Pengujian dengan 25% Larutan PEG 6000
pada Fase Perkecambahan... 36 Waktu dan Tempat... 36 Metode Penelitian... 37 Pelaksanaan Percobaan... 37 A.3. Pengujian dengan 25% PEG 6000 Dalam
xxiv
B. Pengujian dengan Perlakuan Cekaman Kekeringan
di Pot... 39 Waktu dan Tempat... 39 Metode Penelitian... 39 Pelaksanaan Percobaan... 40
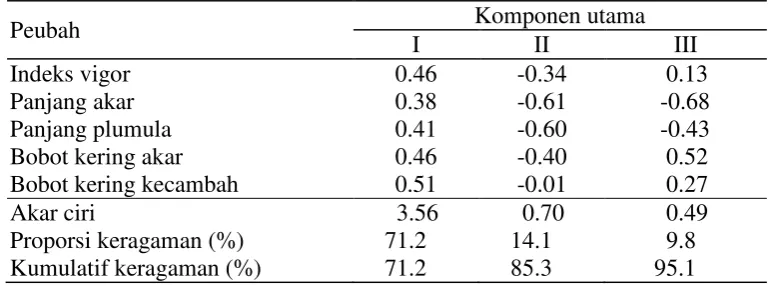
Hasil dan Pembahasan………….………. 43
A. Pengujian dengan Larutan Polietilen Glikol (PEG)
6000... 43 A.1. Penentuan Konsentrasi Larutan PEG 6000
yang Memberikan Cekaman Kekeringan... 43 A.2. Pengujian dengan 25% Larutan PEG 6000
pada Fase Perkecambahan... 47 Peubah Penduga Toleransi Genotipe Padi
Hibrida terhadap Kekeringan pada Fase
Perkecambahan... 51 A.3. Pengujian dengan 25% PEG 6000 Dalam
Kultur Hara pada Fase Bibit... 52 Peubah Penduga Toleransi Genotipe Padi
Hibrida terhadap Kekeringan pada Fase
Bibit... 57 B. Pengujian dengan Perlakuan Cekaman Kekeringan
di Pot... 60 Pengaruh Cekaman Kekeringan terhadap
Pertumbuhan... 60 Kadar Air Relatif Daun... 62
Klorofil……….. 62
Pengaruh Cekaman Kekeringan terhadap
Komponen Hasil dan Hasil... 63
Simpulan………... 68
RESPON AGRONOMI, MORFOLOGI DAN FISIOLOGI PADI HIBRIDA TERHADAP CEKAMAN KEKERINGAN DI LAHAN
SAWAH... 69
Abstrak………. 69
Abstract………. 69
Pendahuluan …….……….……….. 70
xxv A.1. Pertumbuhan Genotipe Padi Hibrida pada
Kondisi Cekaman Kekeringan... 75 A.2. Komponen Hasil dan Hasil Padi Hibrida pada
Kondisi Cekaman Kekeringan... 83 B. Karakter Fisiologi Genotipe Hibrida pada Kondisi
Cekaman Kekeringan……….... 87 B.1. Kerapatan Stomata dan Kadar Air Relatif Daun 87
B.2. Klorofil……… 89
B.3. Karbohidrat Total... 90
Simpulan ……….. 93
UJI DAYA HASIL PENDAHULUAN (UDHP) GENOTIPE PADI
HIBRIDA……… 95
Abstrak……….. 95
Abstract………. 95
Pendahuluan ………. 96
Bahan dan Metode………... 97 A. Uji Daya Hasil Pendahuluan (UDHP) di Lahan
Sawah Irigasi………... 97
Waktu dan Tempat... 97 Metode Penelitian... 97 Pelaksanaan Percobaan... 97 B. UDHP di Lahan Sawah Tadah Hujan... 98 Waktu dan Tempat... 98 Metode Penelitian... 98 Pelaksanaan Percobaan... 99
Hasil dan Pembahasan……… 100
xxvi
B. UDHP di lahan Sawah Tadah Hujan…...……... 108 B.1. Curah Hujan... 108 B.2. Suhu... 110 B.3. Komponen Pertumbuhan... 110 B.3.1. Tinggi Tanaman………. 110 B.3.2. Jumlah Anakan Produktif... 111 B.3.3. Panjang Daun Bendera... 112 B.3.4. Biomasa Total... 112 B.3.5. Umur Berbunga………... 113 B.4. Komponen Hasil dan Hasil... 113 B.4.1. Panjang Malai... 114 B.4.2. Jumlah Gabah Total... 114 B.4.3. Persentase Gabah Isi... 114 B.4.4. Bobot Gabah per Rumpun... 115
Simpulan………. 116
PEMBAHASAN UMUM………... 117
SIMPULAN UMUM DAN SARAN……….. 129
Simpulan Umum....……….. 129
Saran………. 130
DAFTAR PUSTAKA……….. 131
xxvii
Halaman
1 Galur mandul jantan untuk perakitan padi hibrida... 22 2 Galur pemulih kesuburan (restorer) yang telah diseleksi BB padi
untuk perakitan padi hibrida... 23 3 Indeks vigor Situ Bagendit, Inpari 10, Limboto dan IR64 pada
berbagai konsentrasi larutan PEG 6000………... 44 4 Bobot kering kecambah Situ Bagendit, Inpari 10, Limboto dan IR64
pada berbagai konsentrasi larutan PEG 6000....………... 45 5 Indeks vigor Maro, Limboto dan IR64 pada berbagai
konsentrasi larutan PEG 6000... 46 6 Pengaruh larutan PEG 6000 terhadap indeks vigor dan panjang
akar... 48 7 Pengaruh larutan PEG 6000 terhadap panjang plumula dan bobot
kering akar... 49 8 Pengaruh larutan PEG 6000 terhadap bobot kering kecambah... 50 9 Hasil analisis komponen utama beberapa peubah fase
perkecambahan pada perlakuan PEG 6000... 51 10 Pengaruh larutan PEG 6000 terhadap tinggi tajuk dan panjang akar.. 54 11 Pengaruh larutan PEG 6000 terhadap bobot kering akar dan tajuk... 55 12 Pengaruh larutan PEG 6000 terhadap nisbah bobot akar tajuk (NAT)
dan skor tingkat kekeringan daun (SES IRRI) ... 56 13 Hasil analisis komponen utama beberapa peubah fase bibit pada
perlakuan PEG 6000... 58 14 Pengaruh cekaman kekeringan di pot dan genotipe terhadap bobot
kering akar dan bobot kering tajuk... 61 15 Pengaruh cekaman kekeringan di pot dan genotipe terhadap jumlah
gabah isi per malai……...…………... 64 16 Pengaruh cekaman kekeringan di pot terhadap persentase gabah
xxviii
18 Pengaruh cekaman kekeringan di lahan sawah terhadap tinggi
tanaman, jumlah anakan produktif dan luas daun... 76 19 Pengaruh cekaman kekeringan di lahan sawah dan genotipe terhadap
panjang akar saat panen... ……….. 78 20 Pengaruh cekaman kekeringan di lahan sawah dan genotipe terhadap
bobot kering tajuk saat panen………... 80 21 Pengaruh cekaman kekeringan di lahan sawah dan genotipe terhadap
bobot kering akar saat panen………...………. 81 22 Pengaruh cekaman kekeringan di lahan sawah dan genotipe terhadap
umur panen dan periode pengisian biji………... 82 23 Pengaruh cekaman kekeringan di lahan sawah terhadap panjang
malai, jumlah gabah isi dan bobot 1 000 butir... 83 24 Pengaruh cekaman kekeringan di lahan sawah dan genotipe terhadap
persentase gabah hampa dan bobot gabah per rumpun ………... 85 25 Pengaruh cekaman kekeringan di lahan sawah dan genotipe terhadap
hasil gabah per hektar, indeks panen, indeks toleransi kekeringan
dan indeks kepekaan kekeringan ………...…………... 86 26 Distribusi dan frekuensi hujan selama penanaman padi hibrida di
lahan sawah, Desa Bojong, Kecamatan Cikembar, Sukabumi…….... 100 27 Rata-rata curah hujan selama pertumbuhan padi hibrida di lahan
sawah irigasi, Desa Bojong, Kecamatan Cikembar, Sukabumi …..… 101 28 Keadaan suhu selama penanaman padi hibrida di lahan sawah, Desa
Bojong, Kecamatan Cikembar, Sukabumi ……….. 101 29 Komponen pertumbuhan genotipe padi hibrida dan varietas cek di
lahan sawah irigasi………... 102 30 Komponen hasil dan hasil genotipe padi hibrida dan varietas cek di
lahan sawah irigasi………... 105 31 Distribusi dan frekuensi hujan selama penanaman padi hibrida di
lahan sawah tadah hujan, Desa Sanca, Kecamatan Gantar,
xxix 33 Keadaan suhu selama penanaman padi hibrida di lahan sawah tadah
hujan, Desa Sanca, Kecamatan Gantar, Indramayu………….……… 110 34 Komponen pertumbuhan genotipe padi hibrida dan varietas cek di
lahan sawah tadah hujan……….. 111 35 Komponen hasil dan hasil genotipe padi hibrida dan varietas cek di
lahan sawah tadah hujan………... 115 36 Genotipe toleran cekaman kekeringan berdasarkan beberapa metode
pengujian... 118 37 Korelasi indeks vigor, panjang akar, panjang plumul, bobot kering
akar, bobot kering kecambah pada perlakuan PEG 6000 dan bobot gabah per rumpun perlakuan cekaman kekeringan di pot dan
simulasi kekeringan di lahan sawah... 119 38 Korelasi tinggi tajuk, panjang akar, bobot kering akar, bobot kering
tajuk, nisbah bobot akar tajuk, skor tingkat kekeringan daun fase bibit pada perlakuan PEG 6000 dan bobot gabah per rumpun perlakuan cekaman kekeringan di pot dan simulasi kekeringan di
xxxi
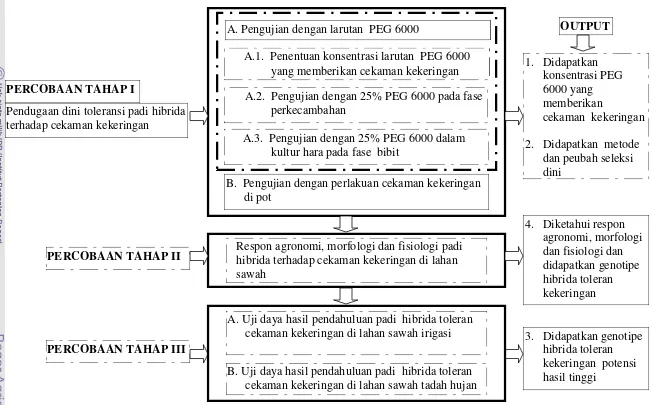
Halaman 1 Bagan alur kegiatan penelitian ………... 6
2 Pola respon peubah panjang plumula (PP) dan panjang akar (PA) varietas Limboto (A), Inpari 10 (B), Situ Bagendit (C) dan IR64 (D)
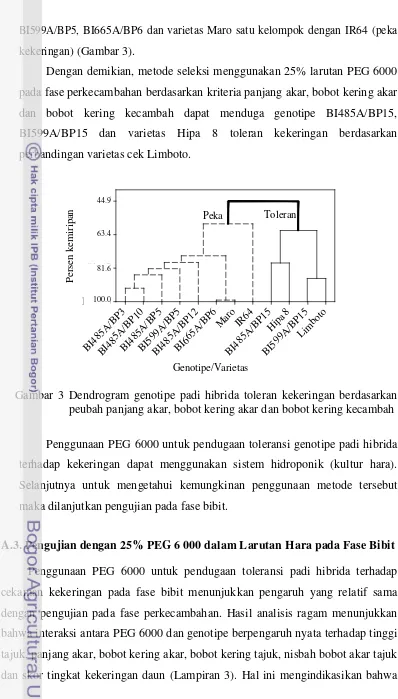
pada berbagai konsentrasi larutan PEG 6000... 44 3 Dendrogram genotipe padi hibrida toleran kekeringan berdasarkan
peubah panjang akar, bobot kering akar dan bobot kering kecambah... 52 4 Penampilan daun beberapa genotipe hibrida dan varietas cek pada
konsentrasi 25% larutan PEG 6000... 57 5 Dendrogram genotipe padi hibrida toleran kekeringan berdasarkan
peubah bobot kering akar, bobot kering tajuk dan skor tingkat
kekeringan daun... 59 6 Pengaruh cekaman kekeringan di pot terhadap kadar air relatif
daun……….. 62
7 Pengaruh cekaman kekeringan di pot terhadap kandungan klorofil
skor SPAD 502……… 63
8 Perubahan kadar air tanah selama periode pengeringan pada
kedalaman ±16 cm………... 77
9 Pengaruh cekaman kekeringan di lahan sawah terhadap kerapatan
stomata... 88 10 Pengaruh cekaman kekeringan di lahan sawah terhadap kadar air
relatif daun……… 89
11 Pengaruh cekaman kekeringan di lahan sawah terhadap kandungan
klorofil... 90 12 Pengaruh cekaman kekeringan di lahan sawah terhadap karbohidrat
non struktural... 91 13 Kadar air tanah selama percobaan di lahan sawah tadah hujan……… 109 14 Penampilan genotipe BI599A/BP15 (H7) dan varietas Limboto
(H12) pada fase reproduktif di lahan sawah tadah hujan pada kondisi
xxxiii
Halaman 1 Hasil analisis tanah sawah Babakan Dramaga (SBD), University
Farm Institut Pertanian Bogor………...…….. 149 2 Rekapituasi hasil analisis ragam pengaruh berbagai konsentrasi
larutan polietilen glikol (PEG) 6000……….... 150 3 Rekapituasi hasil analisis ragam pengujian dengan 25% larutan PEG
6000... 151 4 Rekapituasi hasil analisis ragam pengujian dengan perlakuan
cekaman kekeringan di pot……….. 151 5 Perhitungan kandungan klorofil dan karbohidrat total………. 152 6 Rekapituasi hasil analisis ragam respon agronomi, morfologi dan
fisiologi padi hibrida terhadap cekaman kekeringan di lahan sawah... 152 7 Hasil analisis tanah sawah tadah hujan (STH) Desa Sanca,
Kecamatan Gantar, Kabupaten Indramayu……….. 153 8 Data curah hujan (mm) selama berlangsungnya penelitian di lahan
sawah Desa Bojong, Kecamatan Cikembar, Kabupaten Sukabumi... 154 9 Rekapituasi hasil analisis ragam uji daya hasil pendahuluan (UDHP)
padi hibrida………... 155
10 Data curah hujan (mm) selama berlangsungnya penelitian di lahan sawah tadah hujan Desa Sanca, Kecamatan Gantar, Kabupaten
Indramayu………. 156
11 Hasil pengamatan kadar air tanah selama penelitian di lahan sawah tadah hujan Desa Sanca, Kecamatan Gantar, Kabupaten
Indramayu………. 157
12 Data suhu udara (0C) selama berlangsungnya penelitian di lahan sawah tadah hujan Desa Sanca, Kecamatan Gantar, Kabupaten
Indramayu………. 158
13 Distribusi hujan selama percobaan simulasi cekaman kekeringan di
lahan sawah……….. 159
14 Frekuensi dan jumlah curah hujan bulanan selama percobaan
PENDAHULUAN
Latar Belakang
Beras merupakan salah satu bahan makanan pokok penduduk Indonesia. Jumlah penduduk Indonesia yang semakin meningkat dengan rata-rata laju pertumbuhan per tahun 1.36% (BPS 2009), harus diikuti dengan ketersediaan beras. Ketersediaan beras dapat dicapai melalui peningkatan produktivitas padi. Salah satu upaya peningkatan produktivitas padi dilakukan dengan mengembangkan varietas padi hibrida. Padi hibrida telah terbukti menghasilkan produksi yang sangat tinggi. Rata-rata potensi hasil padi hibrida dapat mencapai 11.4 t ha-1 - 12.6 t ha-1
Meningkatnya kelangkaan air di bidang pertanian akibat perubahan iklim global menyebabkan sawah beririgasi dan tadah hujan sering rawan kekeringan. Cekaman kekeringan merupakan salah satu masalah yang menghambat produktivitas padi, karena pada umumnya pada keadaan kekeringan produksi lebih rendah dan umur panen lebih lama (Boyer 1992; Richards 1996; Nguyen et al. 1997; Blum 2004) dibandingkan dengan kondisi normal. Pengembangan
produktivitas (intensifikasi) merupakan alternatif terbaik untuk dilakukan pada saat perluasan areal sudah semakin sulit seiring dengan berkurangnya lahan produktif di Jawa. Lahan sawah tadah hujan di Indonesia dengan luasan ± 2.1 juta ha diharapkan dapat menjadi lumbung padi ke dua nasional setelah lahan sawah irigasi. Jika produksi padi dapat ditingkatkan sebesar ± 1.0 ton ha
gabah kering atau 15-20% lebih tinggi dibanding varietas unggul inbrida (IR64) (Badan Litbang 2007; Villa et al. 2011).
-1
Permasalahan yang dihadapi di lahan sawah tadah hujan adalah kekeringan hampir terjadi setiap tahun akibat jumlah curah hujan yang rendah dan pendeknya musim hujan (Serraj et al. 2008). Cekaman kekeringan merupakan pembatas utama produksi dan stabilitas produksi padi pada ekosistem tadah hujan (Srinivasan et al. 2008). Lingkungan tersebut memerlukan genotipe toleran yang mampu meminimalkan kehilangan hasil akibat kekeringan dan mempunyai indeks kepekaan yang kecil terhadap kekeringan.
Kekeringan dapat menurunkan laju pertumbuhan akar, tajuk tanaman dan indeks luas daun (Perez et al. 1996; Olsson et al. 1997; Farooq et al. 2008). Menurunnya pertumbuhan akar ini akan menurunkan penyerapan hara dan air sehingga mempengaruhi proses fisiologi antara lain menurunnya fotosintesis. Hal ini dapat menurunkan pertumbuhan dan meningkatkan kehampaan gabah. Pada akhirnya kekeringan menurunkan hasil bahkan sampai menggagalkan panen (Takagi 1976; Van Dat 1986; Samaullah et al. 1996; IRRI 2002). Cekaman kekeringan pada padi dapat terjadi pada pertumbuhan awal, saat berbunga atau pengisian biji. Saat berbunga dan pengisian biji merupakan fase pertumbuhan yang peka bagi tanaman padi terhadap kekeringan. Kekeringan pada fase tersebut dapat menyebabkan tanaman tidak berkembang atau tidak menghasilkan gabah (Lubis et al. 1993; Hijmans dan Serraj 2008), kehilangan produksi dapat mencapai lebih dari 75% (Venuprasad et al. 2008; Serraj et al. 2009).
Salah satu cara untuk menanggulangi masalah tersebut adalah mengembangkan varietas padi yang relatif toleran kekeringan dengan umur genjah, mempunyai peluang yang besar untuk ditanam pada daerah beriklim kering dengan periode hujan singkat. Pengembangan genotipe tanaman yang mengkonsumsi air sedikit merupakan pendekatan yang dapat memberikan harapan untuk keberlanjutan produktivitas tanaman pada daerah yang memiliki kelangkaan air (Serraj et al. 2009; Farooq et al. 2010). Padi hibrida mampu menunjukkan sifat superior (efek heterosis), terutama toleran kekeringan dengan potensi hasil tinggi dan berumur genjah. Perakitan padi hibrida sistem 3 galur yang menggunakan galur mandul jantan (GMJ) tipe Wild abortive (WA) dan Kalinga (KA) dengan galur pemulih kesuburan (R) telah menghasilkan beberapa genotipe padi hibrida yang memiliki nilai standar heterosis tinggi berdasarkan perbandingan terhadap varietas hibrida yang telah dilepas dan varietas cek (Purwoko et al. 2010). Padi hibrida mempunyai kemampuan yang tinggi untuk mengakumulasi bahan kering dan mentranslokasikan cadangan makanan dari batang dan helaian daun ke bagian spikelet (Villa et al. 2011).
mengidentifikasi galur/varietas toleran terhadap kekeringan. Sifat fisik berupa perakaran yang panjang, padat, dan diameter akar yang besar menjadi tolok ukur galur/varietas padi toleran kekeringan (Suardi 1988; Mackill et al. 1996). Perakaran yang padat dan dalam akan meningkatkan serapan air dari tanah. Perbedaan kedalaman perakaran dan penyesuaian ketersediaan air dengan fenologi tanaman merupakan faktor yang menentukan efisiensi penggunaan air (Golluscio dan Oesterheld 2007).
Untuk mengetahui toleransi kekeringan dan memecahkan persoalan banyaknya galur yang akan diuji di lapangan serta kemungkinan pemanfaatan padi hibrida yang berpeluang ditanam pada sawah tadah hujan, maka perlu dicari metode pengujian yang dapat mengidentifikasi secara dini genotipe padi hibrida toleran kekeringan. Metode pengujian yang paling memberikan kesesuaian hasil dengan uji lapangan adalah metode yang tepat untuk mendeteksi secara dini toleransi genotipe tertentu terhadap kekeringan sehingga dapat mempercepat siklus seleksi dalam program perbaikan atau perakitan varietas.
Penggunaan polietilen glikol (PEG) dalam pengujian toleransi benih terhadap kekeringan dengan memperhitungkan indeks kekeringan telah banyak digunakan (Bouslama dan Schapaugh 1984; Nemoto et al. 1995). Penggunaan PEG untuk uji toleransi terhadap kekeringan pada benih padi dilakukan pada tingkat tegangan air -2 dan -12 bar dilakukan di IRRI dan banyak peneliti menggunakan konsentrasi 15-20% dengan PEG 6000. PEG dengan bobot molekul ≥ 6000 telah banyak digunakan dalam melakukan penelitian pengaruh cekaman
air terhadap pertumbuhan tanaman termasuk padi (Balch et al. 1996; Verslues et al. 2006). Selain itu, metode pengujian toleransi terhadap kekeringan yang dianggap dapat menggambarkan kondisi lapangan yang sesungguhnya adalah dengan perlakuan cekaman kekeringan di pot (metode pot). Metode pot dapat memberikan informasi kehilangan air melalui evapotranspirasi selama pertumbuhan dan perkembangan tanaman.
uji lapangan maka pengujian genotipe toleran kekeringan dapat dilakukan dengan cara menanam langsung pada sawah tadah hujan (in situ) atau pada simulasi kekeringan di lahan sawah irigasi.
Penilaian toleransi genotipe terhadap cekaman kekeringan sebaiknya dinyatakan dalam tingkat produktivitas tanaman (Passioura 1983). Beberapa peneliti menyarankan agar kuantifikasi toleransi genotipe terhadap cekaman kekeringan diukur dengan suatu ukuran yang membandingkan secara relatif antara hasil suatu genotipe pada kondisi tercekam dengan kondisi tidak tercekam kekeringan (Fischer dan Maurer 1978; Lin dan Binns 1988; Langer et al. 1999).
Toleransi tanaman terhadap cekaman kekeringan berkaitan dengan mekanisme adaptasi yang dapat dicirikan dengan perubahan-perubahan karakter baik karakter agronomi dan morfologi maupun karakter fisiologi tanaman. Perubahan karakter tersebut merupakan adaptasi yang umum dijumpai pada tanaman yang tumbuh pada cekaman kekeringan, sehingga dapat dijadikan indikator toleransi tanaman terhadap kekeringan. Tanaman yang mengalami cekaman kekeringan akan mempertahankan diri melalui beberapa mekanisme fisiologi yaitu: (i) penghindaran (avoidance): menjaga agar status air di dalam tanaman tidak berkurang, (ii) toleransi (tolerance): mempertahankan fungsi organ walaupun kekurangan air dan (iii) pemulihan (recovery): memperbaiki kondisi tanaman yang telah mengalami kondisi cekaman kekeringan (Blum 2004) serta (iv) melepaskan diri (escape): mempercepat fase reproduktif sebelum terjadi cekaman kekeringan (Fukai dan Cooper 1995).
Tujuan Umum
Tujuan umum penelitian ini adalah untuk mendapatkan metode pengujian dan genotipe padi hibrida toleran kekeringan yang berpotensi dikembangkan di lahan sawah tadah hujan.
Tujuan Khusus
Tujuan khusus penelitian ini adalah:
1. Mengevaluasi metode dan peubah dalam seleksi dini untuk mendapatkan genotipe padi hibrida toleran kekeringan.
2. Mengetahui respon agronomi, morfologi dan fisiologi padi hibrida terhadap cekaman simulasi kekeringan di lahan sawah dan mendapatkan genotipe hibrida yang potensial dikembangkan di lahan sawah tadah hujan.
3. Mengetahui daya hasil padi hibrida toleran cekaman kekeringan di lahan sawah tadah hujan.
Hipótesis
Hipotesis penelitian ini adalah:
1. Metode seleksi toleran kekeringan di laboratorium dan rumah kaca berkorelasi positif dengan toleran kekeringan di lapangan.
2. Toleransi padi hibrida terhadap cekaman kekeringan dapat ditandai dengan karakter fisiologi dan morfologi.
Pendugaan dini toleransi padi hibrida terhadap cekaman kekeringan
PERCOBAAN TAHAP I
B. Pengujian dengan perlakuan cekaman kekeringan di pot
A.3. Pengujian dengan 25% PEG 6000 dalam kultur hara pada fase bibit
A.1. Penentuan konsentrasi larutan PEG 6000 yang memberikan cekaman kekeringan A.2. Pengujian dengan 25% PEG 6000 pada fase
perkecambahan
1. Didapatkan konsentrasi PEG 6000 yang memberikan
cekaman kekeringan 2. Didapatkan metode
dan peubah seleksi dini
Respon agronomi, morfologi dan fisiologi padi hibrida terhadap cekaman kekeringan di lahan sawah
PERCOBAAN TAHAP II
A. Uji daya hasil pendahuluan padi hibrida toleran cekaman kekeringan di lahan sawah irigasi
PERCOBAAN TAHAP III
B. Uji daya hasil pendahuluan padi hibrida toleran cekaman kekeringan di lahan sawah tadah hujan
3. Didapatkan genotipe hibrida toleran kekeringan potensi hasil tinggi
4. Diketahui respon agronomi, morfologi dan fisiologi dan didapatkan genotipe hibrida toleran kekeringan
OUTPUT
Gambar 1 Bagan alur kegiatan penelitian
TINJAUAN PUSTAKA
Kendala Budidaya Padi Sawah Tadah Hujan
Padi dapat dibudidayakan pada berbagai kondisi agroekologi seperti sawah tadah hujan, air-dalam, pasang surut, ladang atau gogo dan sawah irigasi, hal ini tergantung pada jenis/varietas padi. Salah satu zona agroekologi yang sering ditanami padi adalah di lahan sawah tadah hujan (FAO 2004). Luas lahan sawah tadah hujan di Indonesia 2.08 juta ha (Badan Litbang 2005). Sistem produksi padi yang diterapkan adalah teknologi budidaya padi gogo rancah (gora), terutama untuk wilayah yang mempunyai 4-5 bulan basah dalam setahun. Studi yang dilakukan pada tahun 1994 tampak rata-rata hasil padi di lahan sawah tadah hujan pada musim hujan (MH) antara 2.5-4.0 ton ha-1 gabah kering panen (GKP), sedangkan pada musim kemarau (MK) antara 2.0-3.0 ton ha-1
Cekaman kekeringan dan pengaruhnya terhadap produktivitas padi sering tergantung pada banyaknya dan distribusi curah hujan dari total curah hujan musiman. Suatu kasus yang terjadi pada percobaan di IRRI (Los Banos, Philippina) selama musim hujan 2006, dimana curah hujan musiman lebih dari 1 200 mm, tetapi dalam sehari terjadi hujan hanya 320 mm yang bertepatan dengan fase pembungaan menyebabkan penurunan produksi biji dan indeks panen secara tajam dibandingkan dengan kontrol irigasi (Hijmans dan Serraj 2008). Hal ini menunjukkan bahwa distribusi curah hujan dan jumlah hari hujan selama fase pertumbuhan tanaman adalah penting untuk mendukung pertumbuhan dan produksi padi sawah tadah hujan.
GKP (Fagi 1995). Rendahnya produksi padi pada musim kemarau disebabkan oleh rendahnya tingkat ketersediaan air yang tergantung curah hujan. O’Toole (2004) menyatakan padi merupakan jenis tanaman yang dikembangkan pada lingkungan semiaquatik dan secara khusus sangat peka terhadap cekaman kekeringan.
dan fertilisasi); dan (iii) fase pematangan yang dimulai dari pembungaan sampai matang penuh meliputi: fase masak susu, pengerasan, dan pematangan biji (De Datta 1981). Pada pertumbuhan awal yaitu fase vegetatif, kekurangan air dapat menyebabkan pertumbuhan terhambat. Saat berbunga dan pengisian biji merupakan fase pertumbuhan yang peka bagi tanaman padi terhadap kekeringan. Kekeringan pada fase tersebut dapat menyebabkan tanaman tidak berkembang atau tidak menghasilkan gabah (Lubis et al. 1993). Penurunan hasil akibat defisit air setelah fase berbunga terjadi karena penurunan jumlah gabah isi dan meningkatnya sterilitas (kehampaan) (Sikuku et al. 2010). Padi peka terhadap defisit air pada fase reproduksi (Fukai dan Lilley 1994; Pirdashti et al. 2004). Fase bunting dan antesis adalah fase yang paling sensitif (Sikuku et al. 2010). Kekeringan menyebabkan pengurangan terbesar terhadap hasil gabah ketika bertepatan dengan fase reproduksi (Cruz dan O'Toole 1984), sehingga berpengaruh terhadap stabilitas hasil (Babu dan Pathan 1996)
Lahan sawah tadah hujan dengan iklim kering dimana terjadi periode hujan singkat, jumlah dan intensitas curah hujan yang rendah, distribusi curah hujan yang tidak merata sepanjang musim, dan adanya kondisi biofisik-kimia tanah yang beragam menjadi kendala utama budidaya padi di lahan tadah hujan (Quisenberry 1982; Serraj et al. 2008). Setiap kultivar mempunyai kemampuan yang berbeda untuk menghasilkan dalam kondisi cekaman kekeringan. Kehilangan hasil karena cekaman kekeringan dapat berguna dalam menilai toleransi tanaman terhadap kekeringan (Pirdashti et al. 2004). Penanaman varietas toleran kekeringan merupakan salah satu cara untuk mengatasi masalah tersebut (Suardi et al. 2004).
.
Faktor pembatas pertumbuhan tanaman padi di lahan sawah tadah hujan lebih kompleks dibandingkan dengan lahan sawah irigasi. Kendala produksi padi di lahan sawah tadah hujan dihadapkan pada permasalahan kekeringan karena pasokan air hujan yang sulit diprediksi. Mackill et al. (1996) dan Boling et al. (2004) mengemukakan kekeringan di persemaian, kekeringan pada fase vegetatif, dan kekeringan pada fase pembungaan padi yang disebabkan oleh faktor iklim/cuaca yang tidak menentu lebih mempersulit usaha penanggulangan penurunan produksi.
kekeringan dan potensi hasil tinggi untuk meningkatkan dan menstabilkan hasil panen untuk lahan tadah hujan perlu dilakukan (Sikuku et al. 2010).
Perubahan iklim global yang dapat menyebabkan intensitas kejadian iklim ekstrim atau perubahan pola curah hujan yang dapat berimplikasi pada intensitas kejadian kekeringan (efek El-Nino), merupakan tantangan dalam meningkatkan produktivitas tanaman. Data Direktur Jenderal Pengelolaan Lahan dan Air (Ditjen PLA) Departemen Pertanian menunjukkan bencana kekeringan yang terjadi di Indonesia pada periode tahun 1998, 2006 dan 2007 adalah sebagai berikut: (1) pada tahun 1998 seluas 180 701 hektar yang terkena kekeringan, 32 557 hektar mengalami puso (gagal panen) dan kerugian yang ditanggung sebanyak 310 929 ton gabah kering panen, (2) pada tahun 2006 seluas 267 088 hektar yang terkena kekeringan, 63 034 hektar mengalami puso (gagal panen) dan kerugian yang ditanggung sebanyak 527 224 ton gabah kering panen, dan (3) pada tahun 2007 seluas 295 552 hektar yang terkena kekeringan, 17 348 hektar mengalami puso (gagal panen) dan kerugian yang ditanggung sebanyak 365 944 ton gabah kering panen. Fenomena El-nino menjadi ancaman serius pengembangan padi terutama padi sawah tadah hujan.
Kekeringan adalah fenomena yang sering berulang dan faktor pembatas penting pada produksi padi tadah hujan di Asia, terutama curah hujan pendek yang sering terjadi ketika waktu produksi padi. Tercatat 23 juta hektar areal padi (20% dari luas areal padi) di Asia mengalami kekeringan dengan intensitas yang berbeda. Kekeringan merupakan faktor utama yang menyebabkan rendah dan tidak stabilnya produksi padi di wilayah ini (Pandey dan Bhandari 2008). Tujuh puluh persen (70%) petani padi sawah tadah hujan menghadapi masalah kekeringan pada fase reproduktif (Hijmans dan Serraj 2008) dengan penurunan produksi yang sangat parah dibandingkan dengan terjadinya kekeringan pada fase vegetatif. Kekeringan mengurangi produktivitas karena berpengaruh langsung terhadap produksi biomasa dan pembentukan biji (Atlin et al. 2008).
secara berurutan minimal 5 bulan (dengan pelumpuran). Bulan basah adalah bulan dengan curah hujan di atas 200 mm bulan-1 dan bulan kering adalah bulan dengan curah hujan kurang dari 100 mm bulan-1. Fagi dan Las (1998) menyatakan waktu tanam optimum padi sawah bila curah hujan > 200 mm/bulan.
Perubahan iklim dan degradasi sumber daya air dan lahan akibat kerusakan lingkungan hutan dan tata air menyebabkan frekuensi gelombang panas dan periode kekeringan diprediksi akan lebih sering terjadi (Luterbacher et al. 2004; Schär et al. 2004). Hal tersebut
Jumlah kebutuhan air tanaman padi sawah untuk pertumbuhan bibit, pengolahan tanah dan untuk pertumbuhan dari transplanting sampai panen ditentukan oleh banyak faktor, yaitu tipe tanah, topografi, tingkat kekeringan, kedalaman muka air tanah, pemeliharaan tanggul, pemupukan, lamanya tanaman di lapangan, metode pengolahan tanah, dan permintaan evaporasi selama musim pertumbuhan. De Datta (1981) menyatakan bahwa pengolahan lahan semai seluas akan mengacaukan pola dan distribusi curah hujan dan merupakan ancaman serius terhadap kelangsungan pertanian khususnya lahan sawah tadah hujan di dunia termasuk Indonesia karena kekeringan. Ketepatan prediksi awal dan normalitas musim hujan sangat menentukan keberhasilan produksi padi di lahan sawah tadah hujan. Samaullah dan Darajat (2001) menyatakan estimasi faktor tersebut di lapangan sangat sulit ditentukan secara tepat, sebab awal musim hujan dan distribusi curah hujan pada tiap musim di suatu daerah selalu berubah-ubah. Salah satu alternatif yang mungkin dapat dikembangkan untuk mengatasi kendala tersebut adalah menanam varietas toleran kekeringan.
667 m2
Kekeringan dapat didefinisikan menurut perspektif meteorologi, hidrologi dan pertanian. Menurut perspektif meteorologi kekeringan didefinisikan sebagai kurangnya curah hujan dari rata-rata curah hujan normal pada suatu periode waktu tertentu. Kekeringan menurut perspektif hidrologi, merupakan keadaan ketika jumlah air yang tersedia lebih sedikit dibandingkan jumlah air yang dibutuhkan. Di bidang pertanian, definisi kekeringan adalah jumlah air yang tidak mencukupi untuk pertumbuhan optimal tanaman pertanian (Wilhite dan Glantz 1985; Jodo 1995); atau kekeringan terjadi ketika kadar air tanah tidak cukup untuk memenuhi kebutuhan air tanaman (evapotranspirasi) menyebabkan kehilangan produksi. Hal (untuk kebutuhan tanam seluas 1 hektar) membutuhkan air 150-200 mm dan untuk mengairi pertumbuhan tanaman semai selama 21 hari membutuhkan 250-400 mm. Keperluan air dari transplanting sampai panen sebanyak 800-1200 mm, dengan penggunaan air 6-10 mm per hari. Hal ini menunjukkan bahwa tanaman padi sawah sebagai tanaman semiaquatik membutuhkan air yang banyak untuk pertumbuhan dan produksi optimal dan efisiensi penggunaan air sangat ditentukan oleh banyak faktor. Sebanyak 5 000 liter air digunakan untuk menghasilkan 1 kg gabah pada padi sawah irigasi (Al-Saeedi et al. 1999).
Adaptasi Tanaman terhadap Cekaman Kekeringan
ini tergantung pada ketepatan waktu, lama dan keparahan yang dapat mendatangkan bencana kronis atau cekaman kekeringan inherent yang akan membutuhkan perbedaan mekanisme untuk mengatasinya, strategi adaptasi dan tujuan pemuliaan tanaman (Serraj et al. 2008).
Kekeringan merupakan kendala lingkungan yang besar pada tanaman yang dibudidayakan karena mempengaruhi pertumbuhan dan produksi tanaman (Breda et al. 2006; Galle dan Feller 2007).
Diakhir fase I atau awal fase II selama kekeringan, semua proses utama berkontribusi terhadap penghambatan produksi tanaman, termasuk perluasan daun, laju fotosintesis dan pertumbuhan (Serraj et al. 1999). Pada akhir fase II, proses fisiologis dalam mendukung pertumbuhan efektivitasnya mencapai nol dan tidak terjadi pertumbuhan lebih lanjut pada tanaman. Pada fase III proses fisiologis umumnya difokuskan untuk bertahan, yang secara umum melibatkan mekanisme pengaturan osmotik (Serraj dan Sinclair 2002). Peningkatan produksi
Pada kondisi kekeringan, tanaman harus mampu mempertahankan diri melalui beberapa mekanisme adaptasi dan toleransi serta kemampuan untuk pulih setelah cekaman kekeringan terjadi.
tanaman dan produktivitas air pada kondisi kekeringan membutuhkan optimalisasi proses fisiologi yang terlibat pada fase-fase kritis (utamanya fase II) (Serraj et al. 2008).
Tanaman dalam menghadapi kondisi cekaman kekeringan mengembangkan strategi yang berbeda sehingga dikenal kelompok tanaman toleran dan tanaman peka terhadap cekaman lingkungan kekeringan. Tanaman yang memiliki daya adaptasi yang baik (toleran) akan mampu tumbuh dan berproduksi pada lingkungan ekstrim atau tercekam kekeringan meskipun mengalami penurunan produksi (Sikuku et al. 2010). Tanaman yang tidak memiliki daya adaptasi yang baik (peka) akan mengalami hambatan pertumbuhan atau gagal panen atau kematian, yang sangat tergantung pada kapan terjadinya cekaman kekeringan selama periode tumbuh tanaman tersebut (Gunes et al. 2008). Kekeringan dalam waktu singkat yang bertepatan dengan fase pembungaan tanaman menyebabkan penurunan produksi gabah dan indeks panen secara drastis dibanding kontrol (Hijmans dan Serraj 2008). Cekaman kekeringan pada fase sebelum dan setelah antesis mengurangi bobot kering tanaman dan respon tanaman pada awal dan akhir cekaman kekeringan nyata bervariasi. Pengurangan produksi bahan kering kultivar chickpea pada cekaman kekeringan lebih parah berpengaruh pada fase setelah antesis, kemudian fase sebelum antesis (Gunes et al. 2008). Kesuburan serbuk sari (pollen) pada gandum dan eksersi malai serta pecahnya anter pada padi sangat dipengaruhi kekeringan ketika cekaman kekeringan terjadi pada fase reproduksi. Namun ketika cekaman kekeringan dilakukan pada fase vegetatif, gandum tidak terlalu terpengaruh kekeringan dibandingkan padi (Praba et al. 2009). Tingkat cekaman kekeringan sangat tergantung pada waktu dan lamanya cekaman kekeringan (Serraj et al. 2008). Kekeringan menyebabkan kehilangan produksi tanaman dapat melebihi kehilangan produksi dari semua penyebab lainnya, karena tingkat keparahan dan lama cekaman merupakan hal yang kritis (Farooq
Mitra (2001) menyatakan mekanisme yang digunakan tanaman untuk mengatasi cekaman kekeringan dapat dikelompokkan menjadi tiga kategori yaitu melepaskan diri dari kekeringan (drought escape), menghindari kekeringan (drought avoidance) dan mentoleransi kekeringan (drought tolerance). Namun,
tanaman budidaya dapat menggunakan lebih dari satu mekanisme pada waktu mengatasi kekeringan.
Drought escape didefinisikan sebagai kemampuan tanaman untuk
melengkapi siklus hidupnya sebelum tanah dan tanaman mengalami defisit air yang parah. Mekanisme ini melibatkan perkembangan fenologi yang cepat (pembungaan lebih awal dan pemasakan lebih awal) dan remobilisasi asimilat pra-antesis. Drought avoidance adalah kemampuan tanaman untuk mempertahankan potensial air jaringan relatif tinggi meskipun kekurangan kadar air tanah. Tanaman mengembangkan strategi untuk memelihara turgor dengan meningkatkan kedalaman akar atau mengembangkan sistem akar yang efisien untuk memaksimalkan penyerapan air, dan mengurangi kehilangan air melalui pengurangan konduktansi epidermis (stomata dan lenticular), mengurangi radiasi yang diabsorbsi dengan menggulung atau melipat daun dan mengurangi evapotransporasi permukaan daun (luas daun). Mekanisme drought tolerance adalah kemampuan sel-sel jaringan tanaman untuk tetap hidup dan berfungsi meskipun kandungan air dalam jaringan berkurang atau dalam keadaan kekeringan, dimana tanaman mempertahankan turgor dengan pengaturan osmotik untuk menginduksi akumulasi solut dalam sel sehingga elastisitas sel meningkat (Pugnaire et al. 1999; Mitra 2001; De Datta 2002).
adaptasi dan mekanisme respon yang berbeda baik secara morfologi maupun fisiologi.
Respon Morfologi
Mekanisme penghindaran dari kekeringan (avoidance) dengan mengurangi kehilangan air banyak berhubungan dengan respon secara morfologi misalnya perubahan struktural stomata dan mengurangi luas daun, meningkatkan ketebalan daun, pertumbuhan akar yang lebih dalam dan perubahan ketebalan lilin daun (Galle dan Feller 2007; Agbicodo et al. 2009).
Cekaman kekeringan cenderung meningkatkan panjang perakaran tanaman dan rasio akar tajuk (Farooq et al. 2008). Hal tersebut berkaitan dengan upaya tanaman dalam mencapai lapisan tanah yang lebih dalam, karena pada umumnya lapisan yang lebih dalam memiliki kelembaban yang lebih besar dibandingkan dengan tanah yang ada di lapisan atas (Breseghello et al. 2008). Kemampuan tanaman untuk mempertahankan pertumbuhan akar sangat penting dalam mempertahankan penyerapan air dan hara dalam keadaan cekaman kekeringan (Hamim et al. 2008). Perubahan arsitektur akar karena kekeringan tergantung genotipe. Bentuk dan kedalaman akar sangat bervariasi antar kultivar (Price et al. 1997; Venuprasad et al. 2002) tetapi beberapa kultivar padi gogo sangat sensitif terhadap kondisi tanah kering pada fase pembungaan (Atlin et al. 2008). Beberapa kultivar menunjukkan densitas akar yang sama pada perlakuan cekaman dan tanpa cekaman, sedangkan perubahan arsitektur akar yang lain memproduksi akar yang sangat dalam, ketika cekaman kekeringan terjadi (Breseghello et al. 2008).
Sistem akar memainkan peranan penting untuk tanaman dalam memperoleh air dan merupakan komponen penting dalam toleransi terhadap cekaman defisit air. Pertumbuhan pemanjangan akar berhubungan dengan pengaturan potensial osmotik (Sharp et al. 2004). Pada padi, beberapa kelompok telah menunjukkan pentingnya komponen perakaran tertentu yang memberikan kontribusi terhadap hasil yang lebih tinggi dalam lingkungan kekurangan air dan mengidentifikasi QTL yang sesuai untuk sifat toleransi terhadap kekeringan (Bruce et al. 2002).
Fungsi penting lilin daun adalah untuk meningkatkan efisiensi stomata mengontrol pengurangan kehilangan air setelah stomata menutup (Clarke dan Richards 1988). Sebagai respon terhadap cekaman kekeringan, deposisi lilin epikutikular dimulai beberapa hari sebelum cekaman kekeringan parah terjadi dan tanaman toleran sering memiliki lilin daun lebih tebal dibandingkan tanaman peka (Shepherd dan Griffiths 2006).
1994). Penggulungan daun adalah sifat sekunder yang dimiliki oleh tanaman untuk karakter toleransi terhadap cekaman kekeringan (Bänziger et al. 2000).
Tingkat toleransi cekaman kekeringan berbeda antar kultivar. Pada kondisi defisit air yang meningkat pertumbuhan tanaman dicegah sehingga ukuran berbagai bagian tumbuhan berkurang. Kultivar gandum yang toleran juga memelihara kesuburan serbuk sari yang lebih tinggi sedangkan padi toleran memiliki kesuburan spikelet tinggi. Sensitivitas terbesar hasil gandum terhadap cekaman kekeringan terjadi pada tahap awal pengembangan spikelet, sekitar 5 hari setelah penyerbukan. Pada padi, sensitivitas terbesar terjadi tak lama setelah heading (Praba et al. 2009). Defisit air menurunkan hasil padi karena menurunkan
jumlah anakan, panjang malai dan persentase gabah isi. Selain itu, defisit air mempengaruhi umur panen dan hasil gabah karena terjadi penurunan jumlah anakan, panjang malai dan persentase gabah isi (Sikuku et al. 2010).
Respon Fisiologi
Cekaman kekeringan akan menyebabkan adanya perubahan pada proses fisiologi seperti pada pembelahan sel, sintesis protein dan fotosintesis (Mitra 2001; Altman 2003; Zulkarnain et al. 2009). Defisit air menurunkan konduktansi stomata dan kadar air relatif. Sementara aktivitas peroksidase dan akumulasi prolin meningkat pada padi dengan perlakuan cekaman kekeringan (Zulkarnain et al. 2009).
Kadar air relatif daun merupakan ukuran dari adaptasi terhadap cekaman kekeringan dan perkiraan osmotic adjustment, yang dianggap sebagai salah satu mekanisme paling penting untuk adaptasi terhadap lingkungan tanaman dengan air yang terbatas (Jain dan Chattopadhyay 2010). Selama perlakuan cekaman kekeringan pada kacang tunggak cultivar PUSABGD72 dan ICCV2 menunjukkan adanya peningkatan kadar air relatif daun setelah 3 hari perlakuan cekaman kekeringan. Hal ini terjadi akibat peningkatan transportasi air dari kompartemen
lain dari tanaman ke daun untuk menjaga turgor, atau mungkin hasil dari penyesuaian osmotik akibat peningkatan sintesis osmolit. Setelah peningkatan awal ini, kadar air relatif daun pada kedua kultivar tersebut menunjukkan penurunan yang tajam sampai dengan akhir perlakuan (12 hari). Namun, pada akhir perlakuan kadar air relatif daun PUSABGD72 10% lebih tinggi dari ICCV2 (Jain dan Chattopadhyay 2010).
Penutupan stomata tidak selalu berhubungan dengan setiap perubahan kadar air relatif. Kacang tunggak kultivar EPACE-1 menunjukkan respon awal stomata terhadap penipisan air substrat tidak dipicu oleh perubahan kadar air daun. Oleh karena itu, hanya dengan perubahan kadar air relatif saja, tidak dapat digunakan sebagai suatu indikator kekeringan untuk sebagian spesies tanaman. Komunikasi akar – daun, tidak tergantung status air daun yang menginformasikan pucuk tentang perubahan di zona akar tetapi juga ditentukan oleh faktor lainnya (Agbicodo et al. 2009). Gunes et al. (2008) menyatakan bahwa kultivar toleran kekeringan memiliki kadar air relatif daun, asam askorbat dan konsentrasi prolin tinggi, tetapi permeabilitas membran rendah dibandingkan dengan kultivar peka kekeringan.
Prolin adalah salah satu indikator respon adaptif terhadap cekaman kekeringan (Yoshiba et al. 1997; Jain dan Chattopadhyay 2010). Prolin dan kadar ABA pada kacang tunggak kultivar PUSABGD72 dan ICCV2 meningkat setelah 3 hari perlakuan cekaman kekeringan dan akumulasi prolin sangat tinggi pada kultivar PUSABGD72 dibanding ICCV2, 12 hari setelah perlakuan cekaman kekeringan (Gunes et al. 2008; Jain dan Chattopadhyay 2010). Toleransi tanaman pada cekaman kekeringan dapat dilihat dari daya pemulihan (recovery), yaitu terjadinya laju pertumbuhan yang cepat segera setelah cekaman kekeringan terhenti (Chang et al. 1972). Genotipe dengan daya pemulihan baik, memiliki rangsangan pertumbuhan lebih cepat dibanding genotipe kontrol, karena terdapat zat prolin sebagai sumber energi untuk pertumbuhan setelah cekaman kekeringan dihentikan.
Kultivar gandum Excalibur dan RAC875 toleran kekeringan mengadopsi perbedaan sifat secara morfologi dan mekanisme fisiologi untuk mengurangi cekaman kekeringan. Excalibur paling responsif terhadap tingkat ketersediaan air dan menunjukkan tingkat pengaturan osmotik tinggi, konduktansi stomata tinggi, kadar ABA terrendah, dan pemulihan lebih cepat pada perlakuan cekaman kekeringan. RAC875 lebih konservatif dan terkendali, dengan pengaturan osmotik moderat, lilin (waxiness) daun tinggi, kandungan klorofil tinggi, dan pemulihan lebih lambat akibat cekaman kekeringan. Pengaturan osmotik, di samping untuk menjaga turgor dan mempertahankan fungsi selular untuk waktu yang lama pada kondisi kekeringan, memungkinkan tanaman untuk pulih lebih cepat akibat cekaman kekeringan (Izanloo et al. 2008).
ketika potensial air daun berkurang (Morgan 1980). Pinter dan Kalman (1978) menyatakan pengaturan osmotik disebabkan oleh peningkatan akumulasi bahan larut misalnya asam amino prolin dan asparagin sehingga mampu menjaga turgor tanaman. Morgan dan Condon (1986) menemukan hubungan positif antara pembentukan benih (seed set) dan pemeliharaan turgor dan menyimpulkan bahwa genotipe dengan pemeliharaan turgor rendah menghasilkan sedikit benih.
Cekaman kekeringan menyebabkan tanaman mengalami penurunan laju fotosintesis akibat penurunan konduktansi stomata (Morison dan Lawlor 1999; Hamim 2005). Selain hal tersebut, kekeringan yang berat dapat mengakibatkan kerusakan langsung dari perangkat-perangkat fotosintesis seperti fotosistem I dan II (PSI dan PSII) (Genty et al. 1987; Colom dan Vazzana 2003; Subrahmanyam et al. 2006). Salah satu penyebab kerusakan tanaman pada kondisi kekeringan ialah
terjadinya cekaman oksidatif yang disebabkan terakumulasinya senyawa Reactive Oxygen Species (ROS) seperti singlet oksigen (O2), hidrogen peroksida (H2O2),
superoksida (O2¯) dan hidroksil radikal (OH) (Prochazkova et al. 2001). Radikal
tersebut dapat menyebabkan kerusakan melalui beberapa cara yaitu memutus ikatan rantai protein, merusak membran lemak dan bereaksi dengan DNA sehingga menyebabkan mutasi sel (Sgherri dan Nazari-Izzo 1995).
Bosch dan Alegre (2002) menunjukkan bahwa selain terjadi peningkatan aktivitas enzim antioksidan, tanaman juga mengakumulasi metabolit seperti askorbat dan α-tokoferol yang berfungsi untuk mengatur tingkat keaktifan oksigen pada jaringan tanaman. Beberapa mekanisme lain yang sudah umum diketahui ketika terjadi cekaman kekeringan pada tanaman antara lain ialah tanaman akan melakukan pengaturan osmotik dengan mengakumulasi senyawa organik diantaranya glisin betain atau sukrosa dan juga mengakumulasi ion anorganik dari tanah berupa K+, Cl- atau Na+ di dalam sel (Taiz dan Zeiger 2002). Lin dan Wang (2002) juga menyatakan bahwa tiap genotipe tanaman memiliki keseimbangan yang berbeda antara ROS dan antioksidan.
Kekeringan memicu produksi hormon asam absisat (ABA), yang menyebabkan penutupan stomata dan menginduksi ekspresi gen yang terkait dengan cekaman (Shinozaki dan Yamaguchi-Shinozaki 2007). Sel penjaga (guard sel) mengintegrasikan sinyal tingkat ketersediaan air dan CO2, hormon,
cahaya dan kondisi lingkungan lainnya, untuk mengatur ukuran apertur stomata. Hormon ABA adalah antitranspirant endogen yang mengurangi laju kehilangan air melalui pori-pori stomata pada epidermis daun. Peningkatan biosintesis ABA terjadi sebagai respon terhadap kekurangan air, yang menyebabkan redistribusi intraseluler dan akumulasi ABA dalam sel penjaga. Peningkatan kadar ABA menyebabkan efluks ion, hilangnya turgor sel penjaga, dan penutupan pori stomata (Bray 1997; Schroeder et al. 2001).
Penggunaan dasar-dasar genetika akan sangat membantu dalam peningkatan toleransi kekeringan pada tanaman padi (Babu dan Pathan 1996). Padi hibrida cenderung lebih unggul dalam mentolerir kekeringan (Tian et al. 1980). Keragaan
Kelebihan energi eksitasi dari konversi energi cahaya menjadi energi kimia adalah sumber generasi ROS dalam sel penjaga. Oleh karena itu kloroplas tidak hanya memenuhi kebutuhan energi, tetapi juga merupakan sensor informasi lingkungan, dan sinyal redoks kloroplas memungkinkan tanaman untuk membiasakan diri dengan cekaman lingkungan (Pfannschmidt 2003).
tanaman hibrida F1
No.
tergantung pada pemilihan tetua yang akan memberikan hibrida heterotik.
Heterosis (hybrid vigor) pada padi menunjukkan superioritas hibrida (F1) dibandingkan tetua pembentuknya dalam hal kejaguran pertumbuhan (growth vigor), vitalitas, kapasitas reproduktif, ketahanan terhadap cekaman biotik dan ketoleranan terhadap cekaman abiotik, adaptabilitas, hasil biji serta karakteristik lain (Yuan et al. 2003; Virmani dan Kumar 2004). Secara substansial terdapat variasi genetik di dalam Oryza sativa untuk sifat toleransi terhadap kekeringan. Beberapa varietas sangat toleran pada cekaman jangka pendek sekitar pembungaan, sebaliknya pada varietas lain dengan jelas mengurangi pembentukan benih dan indeks panen (Atlin et al. 2008).
Pengembangan padi hibrida yang sesuai dengan kondisi Indonesia adalah sistem tiga galur yang memerlukan mandul jantan sitoplasmik (CMS/GMJ/galur A), galur pelestari (galur B) dan pemulih kesuburan (galur R) (Virmani et al. 1997). Populasi padi hibrida F1 sebagai materi genetik untuk pengujian seleksi toleransi kekeringan dari persilangan dua jenis Galur mandul jantan steril sitoplasmik (GMJ) dengan 15 galur pemulih kesuburan (restorer/R) (Purwoko et al. 2010). GMJ tersebut adalah mandul jantan tipe Wild abortive (WA) dan Kalinga
(KA) beserta galur pelestari pasangan masing-masing yang telah mempunyai sterilitas 100%, karakter agronomis dan morfologi bunga yang mendukung penyerbukan silang, serta tahan terhadap hawar daun bakteri (Tabel 1). Galur mandul jantan merupakan kunci utama dalam perakitan padi hibrida sistem 3 galur.
Tabel 1 Galur mandul jantan untuk perakitan padi hibrida
Tipe GMJ Karakter GMJ
1. Wild abortive (WA) (BI485 dan BI 599)
100 % steril, umur genjah, tahan HDB strain III dan IV
2. Kalinga (KA) (BI665)
100 % steril, sangat tahan HDB strain III, tahan HDB strain IV
Sumber: Purwoko et al. (2010) dan Rumanti (2012)
tahan terhadap hama wereng batang coklat, atau penyakit hawar daun bakteri (Tabel 2).
Tabel 2 Galur pemulih kesuburan (restorer) yang telah diseleksi BB padi untuk perakitan padi hibrida
No. Galur Restorer Karakter restorer
1. IR53942 Jumlah pollen dalam antera banyak, daya gabung baik
2. S4124F Jumlah pollen dalam antera banyak, daya gabung baik, pulen, tahan HDB
3. BP51-1 Jumlah pollen dalam antera banyak, daya gabung baik, kualitas gabah baik
4. BP1028F Jumlah pollen dalam antera banyak, daya gabung baik, agak toleran kering
5. BH25B-MR-2-2-B Genjah, Jumlah pollen dalam antera banyak, daya gabung baik
6. R42 Jumlah pollen dalam antera banyak, daya gabung baik, kualitas gabah baik
7. BP2274-3E-4-1 Jumlah pollen dalam antera banyak, daya gabung baik, tahan WBC
8. CRS 39 (BP2510E-5-3-1) Jumlah pollen dalam antera banyak, daya gabung baik, tahan WBC
9. CRS 8 (BP2276-5E-28-2) Jumlah pollen dalam antera banyak, daya gabung baik, tahan WBC
10. CRS 9 (BP2278-2E-17-1) Jumlah pollen dalam antera banyak, daya gabung baik, tahan WBC
11. SMD9C-7 Toleran kekeringan, umur panen 90 HST 12. SMD10C-6 Toleran kekeringan, umur panen 86 HST 13. SMD11C-12 Toleran kekeringan, umur panen 88 HST 14. SMD12C-9 Toleran kekeringan, umur panen 89 HST 15. SMD13C-6 Toleran kekeringan, umur panen 90 HST Sumber: Purwoko et al. (2010)
perakaran lebih kuat, anakan lebih banyak, jumlah gabah per malai lebih banyak, dan bobot 1 000 butir yang lebih tinggi. Dari aspek fisiologi, padi hibrida memiliki aktivitas perakaran yang lebih luas, area fotosintesis yang lebih luas, intensitas respirasi yang lebih rendah dan translokasi asimilat yang lebih tinggi. Padi hibrida dalam mewujudkan potensinya umumnya memerlukan areal penanaman dengan syarat tumbuh tertentu. Teknologi pemuliaan seperti penggunaan padi hibrida untuk lahan kering akan makin penting di masa depan, dan apabila pemulia padi mampu meningkatkan potensi hasil sekitar 1 ton ha-1 di atas padi gogo lokal, maka akan diperoleh tambahan produksi beras secara siginifikan (Sutaryo et al. 2004).
Padi hibrida yang ideal toleran terhadap kekeringan adalah padi yang memberikan hasil panen lebih tinggi dibandingkan padi lain apabila terjadi cekaman kekeringan dan tetap berpenampilan baik bila tidak ada cekaman kekeringan (Fukai dan Cooper 1995). Sifat yang terdapat pada padi toleran cekaman kekeringan dapat berupa sifat konstitutif dan indusibel (Blum 2002; 2005). Sifat konstitutif tidak dipengaruhi oleh ada atau tidaknya cekaman. Sifat indusibel muncul pada saat tanaman terkena cekaman kekeringan dan merupakan proses adaptasi tanaman.
Metode dan Karakter Seleksi Toleransi Kekeringan
Seleksi untuk toleransi kekeringan pada berbagai jenis tanaman sudah banyak dilakukan dengan menggunakan berbagai metode seleksi. Metode tersebut berupa seleksi langsung dan tidak langsung. Seleksi langsung dilaksanakan pada lahan bercekaman seperti pada lahan sawah tadah hujan (Breseghello et al. 2008; Sie et al. 2008). Seleksi tidak langsung dengan menirukan kondisi cekaman yang terjadi di lapangan dapat dilaksanakan di laboratorium atau di rumah kaca. Seleksi di laboratorium biasanya menggunakan PEG pada kultur in vitro (Lestari 2007; Satoh et al. 2008) dan pada fase kecambah (Ogawa dan Yamauchi 2006; Ai et al. 2008; Satoh et al. 2008; Efendi 2009; Imanparasat dan Hasanpanah 2009; Jajarmi 2009; Gholami et al. 2010; Herawati 2010), pada fase vegetatif dengan menggunakan media pasir (Chazen dan Newman 1994; Efendi 2009). Disamping itu seleksi untuk toleransi kekeringan di rumah kaca dapat dilakukan dengan menggunakan metode lain misalnya dengan menumbuhkan padi pada lapisan lilin (campuran parafin dan vaselin) (Suardi 2002), media campuran tanah dan sabut kelapa (Satoh et al. 2008), media campuran perlit dan tanah (Praba et al. 2009) dalam pot plastik hitam, media tanah dalam pot silinder/pipa PVC (Breseghello et al. 2008; Courtois 2008; Sikuku et al. 2010) dan media hidroponik (Xu et al. 2004; Yue et al. 2006; Courtois 2008) serta dengan menggunakan penanda molekuler (Xiong 2008).
Metode-metode seleksi tersebut, masing-masing memiliki kelebihan dan kekurangan dan pemilihan metode uji sangat tergantung pada ketersediaan bahan/alat pengujian (kemampuan laboratorium), banyaknya bahan yang akan diseleksi, dan kemudahan dalam pelaksanaannya. Samac dan Tasfaye (2003) menyatakan metode yang dipilih harus murah, pelaksanaannya cepat dan handal untuk menyeleksi genotipe dalam jumlah banyak sekaligus dan dapat memisahkan genotipe toleran dan peka (Lewis dan Christiansen 1981; Fukai et al. 2008; Sie et al. 2008).
curah hujan musiman yang tidak menentu serta jenis dan tipe tanah yang berbeda pada suatu wilayah akan memberikan keragaman respon, sehingga menyulitkan dalam menilai genotipe toleran dan peka, bila dilakukan seleksi langsung di lapangan. Kehilangan hasil di lapangan baik sawah tadah hujan maupun sawah irigasi tidak hanya disebabkan oleh faktor kekeringan tetapi juga faktor-faktor lingkungan lainnya yang saling berinteraksi. Spehar dan Sauza (2006) menyatakan pengujian di lapangan dapat menghadapi lebih dari satu cekaman dan faktor-faktor lain yang tidak dapat dikendalikan yang akan mempengaruhi pertumbuhan dan perkembangan tanaman. Adanya berbagai interaksi yang terjadi di lapangan akan menyulitkan untuk memilah antara pengaruh lingkungan kekeringan dan genotipe. Hal tersebut dapat diperbaiki dengan melakukan seleksi pada lingkungan terkontrol seperti di rumah kaca.
Seleksi dengan menggunakan media tanah atau campuran media tanah dengan bahan lain di rumah kaca akan memberikan tekanan seleksi yang hampir mendekati keadaan kekeringan di lapangan. Media tanam yang digunakan dapat diambil dari lingkungan daerah target produksi dan memiliki hampir semua cekaman yang dijumpai di lapangan misalnya aspek jenis dan tekstur tanah. Metode seleksi akan baik bila terdapat konsistensi hasil antara percobaan di rumah kaca (terkontrol) dan di lapangan terhadap cekaman kekeringan pada padi. Samaullah dan Darajat (2001) menyatakan genotipe 2-2 dan S3842-5g-2-3 adalah genotipe toleran kekeringan, baik di rumah kaca maupun di lapangan dalam kondisi tercekam. Kedua galur secara konsisten menampilkan persentase kehampaan gabah yang relatif rendah dan daya hasil relatif tinggi dibanding dengan genotipe lainnya serta nilai indeks kepekaan terhadap kekeringan yang relatif stabil.