ISNA RAHMADINI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
© Hak Cipta milik IPB, tahun 2012 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tidak merugikan kepentingan yang wajar IPB.
Dengan ini saya menyatakan bahwa Tesis Pemurnian dan Karakterisasi Enzim Selulase dari Bakteri yang Diisolasi dari Limbah Rumput Laut adalah karya saya sendiri yang merupakan bagian dari penelitian kelompok peneliti Bioteknologi BBP4BKP tahun anggaran 2010/2011 dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau kutipan dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Mei 2012
ISNA RAHMADINI. Purification and Characterization of Cellulase Enzyme from Bacteria Isolated from Seaweed Waste. Under direction of NISA RACHMANIA MUBARIK and EKOWATI CHASANAH.
Seaweedwasteis a sourceof bacteria thatcanproduce cellulaseenzyme. PMP0126yisolateis one collection isolateof the Research and Development Center for Marine and Fisheries Product Proecessing and Biotechnology Agency for Marine and Fisheries Research and Development Ministry of Marine Affairs and Fisheries obtainedfromseaweed wastefrom Pameungpeukarea, Garut, West Java. The aims this research were to purify, characterize the cellulase enzyme, and identify the bacteria producing the enzyme using 16S-rRNA. PMP 0126yisolatewasa Gram-negativeshortrodshape bacteria. Based on thesequencingof the 16S-rRNA genfrom 1282base pairPMP0126y isolate had96%similaritywith Chryseobacteriumindologenesstrain McR-1. Theisolate had 1.9 cellulolytic index onCarboxymethyl Cellulose(CMC) agar medium. The highestcellulaseactivityobtained onthe thirdday offermentation timewith acellulaseactivityof0.108U/mLandspecificactivityof0.120U/mg.
Initialpurificationof cellulasebyultrafiltrationproducedactivityof0.112U/mL. Purificationthe enzyme byanionexchange chromatographyproducedthe highestpeak of proteininthefraction no. 48withcellulaseactivityof0.154U/mLat 37.3mMNaCl.Optimumactivity ofthe cellulaseenzymeafterultrafiltrationwaspH 5and300C,while optimumactivity ofthe cellulaseenzymebyanionexchange chromatographywaspH 5and400C. At 300C, the enzymeremainedstableup to4hourincubation.Theactivity ofthe cellulasewasincreasedbyaddition ofCaCl2ions
by 53%anddecreased bythe additionof ZnCl2 ions by 78%. Thecellulase showed
the highest activity i.e. 0.149U/mL using treated seaweed wasteGlacilariasp. as substrate. Using SDS-PAGE and zimogram analysis, the molecular weight of the cellulase was estimated to be 39 kDa, 30 kDa, and 14 kDa.
yang Diisolasi dari Limbah Rumput Laut. Dibimbing oleh Nisa Rachmania Mubarik dan Ekowati Chasanah.
Limbah pengolahan rumput laut merupakan salah satu sumber bakteri yang dapat menghasilkan enzim selulase. Industri pengolahan agar-agar dari rumput laut Glacilaria sp. di daerah Pemeungpeuk Garut, Jawa Barat merupakan sumber isolat PMP 0126y yang mampu menghasilkan enzim selulase. Isolat ini merupakan koleksi BBP4BKP yang dapat tumbuh baik pada suhu 37 0C. Hasil pewarnaan Gram isolat PMP 0126y bersifat Gram negatif dengan bentuk batang pendek. Berdasarkan hasil sekuensing gen penyandi16S-rRNA dari 1282 pasang basa, isolat PMP 0126y memiliki kemiripan sebesar 96% dengan bakteri Chryseobacterium indologenes galur McR-1.
Uji kualitatif dilakukan dengan mengukur indeks selulolitik yang dihasilkan oleh bakteri pada media agar-agar yang mengandung Carboxymethyl Cellulose (CMC). Indeks selulolitik yang dihasilkan oleh isolat PMP 0126y pada media agar-agar CMC 1% sebesar 1,9 pada hari kelima dengan suhu inkubasi 37 0C. Uji kuantitatif yang dilakukan terhadap selulase yang dihasilkan oleh isolat PMP 0126y menghasilkan aktivitas selulase tertinggi pada hari ketiga produksi dengan aktivitas selulase sebesar 0,108 U/ml dan aktivitas spesifik sebesar 0,120 U/ml.
Enzim selulase dipekatkan dengan melakukan pengendapan amonium sulfat dan ultrafiltrasi. Persentase amonium sulfat yang terbaik dihasilkan pada 50% amonium sulfat dengan aktivitas selulase yang diperoleh sebesar 0,072 U/ml dan aktvitas spesifik 0,128 U/mg pada endapan. Pemekatan dengan ultrafiltasi menghasilkan aktivitas selulase sebesar 0,112 U/ml dan aktivitas spesifik 0,136 U/mg. Pemurnian selanjutnya dilakukan dengan kromatografi penukar anion (KPA) yang menghasilkan puncak tertinggi pada fraksi ke-48 dengan aktivitas selulase sebesar 0,154 U/ml ketika dielusi dengan NaCl sebesar 37,3 mM. Pra pemurnian enzim selulase dengan ultrafiltrasi menghasilkan rendemen sebesar 17,5% dengan tingkat kemurnian 15,82 kali. Enzim hasil pemurnian dengan KPA menghasilkan rendemen sebesar 19,6% dengan tingkat kemurnian sebesar 15,08 kali. Hasil SDS-PAGE dan zimogram menunjukkan ada tiga protein enzim selulase dari isolat PMP 0126y pada berat molekul yaitu 39 kDa, 30 kDa, dan 14 kDa.
Aktivitas optimum enzim selulase PMP 0126y hasil ultrafiltrasi tertinggi pada bufer sitrat fosfat pH 5 dan suhu 30 0C. Enzim tetap stabil selama 4 jam inkubasi pada suhu 30 0C. Aktivitas relatif tertinggi enzim selulase meningkat dengan penambahan logam CaCl2 sebesar 53% dan menurun pada penambahan
logam ZnCl2 sebesar 78%. Aktivitas enzim selulase tertinggi pada substrat limbah
ISNA RAHMADINI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program StudiBioteknologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Nama : Isna Rahmadini
NIM : P051090191
Disetujui Komisi Pembimbing
Dr. Nisa Rachmania Mubarik, M.Si. Ketua
Dr. Ekowati Chasanah, M.Sc. Anggota
Diketahui
Ketua Program Studi Bioteknologi
Prof. Dr. Ir. Suharsono, DEA.
Dekan Sekolah Pascasarjana
Dr. Ir. Dahrul Syah, M.Sc.
Penulis dilahirkan di Lahat pada tanggal 19 April 1988 dari Ayah H. Hardi Bustanuddin dan Ibu Hj. Muchlisa. Penulis merupakan anak kelima dari lima bersaudara.
Tahun 2005 penulis lulus dari SMA Negeri 2 Lahat dan masuk seleksi PBUD di Universitas Riau pada Program Studi Teknologi Hasil Perikanan (THP) dan berhasil menyelesaikan kuliah pada tahun 2009. Pada tahun yang sama, penulis melanjutkan sekolah dan masuk ke dalam Mayor Multidisiplin, Program Studi Bioteknologi, IPB.
Puji syukur Alhamdulillah penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Februari 2011 ini ialah enzim selulase, dengan judul Pemurnian dan Karakterisasi Enzim Selulase dari Bakteri yang Diisolasi dari Limbah Rumput Laut.
Penulis mengucapkan terima kasih dan penghargaan yang tinggi kepada Ibu Dr. Nisa Rachmania Mubarik, M.Si. selaku ketua komisi pembimbing yang telah memberikan bimbingan dan perhatian penuh dalam penulisan tesis. Ucapan terima kasih dan penghargaan yang tinggi juga kepada Ibu Dr. Ekowati Chasanah, M.Sc. selaku anggota komisi pembimbing yang telah memberikan kesempatan kepada penulis untuk melakukan penelitian dan bimbingan selama penelitian, serta kepada Ibu Ir. Yusro Nuri Fawzya, M.Si. yang telah banyak memberikan saran dan bimbingan selama penelitian. Tidak lupa penulis mengucapkan terima kasih banyak kepada Ibu Prof. Dr. Anja Meryandini, M.S. sebagai penguji ujian tesis dan Bapak Prof. Dr. Ir. Suharsono, DEA. sebagai ketua Program Studi Bioteknologi yang telah memberikan saran dan masukan terhadap penulisan demi kesempurnaan tesis ini. Di samping itu, penulis menyampaikan terima kasih kepada Balai Besar Penelitian Pengembangan Pengolahan Produk dan Bioteknologi Kelautan dan Perikanan (BBP4BKP) yang telah membiayai dan memberikan segala fasilitas kepada penulis untuk melakukan penelitian di Laboratorium Bioteknologi dan Mikrobiologi BBP4BKP Petamburan, Jakarta Pusat.
Fakultas Perikanan dan Ilmu Kelautan Universitas Riau yang sedang sekolah di IPB atas persahabatan, dorongan, semangat, dan bantuan dalam penyelesaian tesis ini. Semoga Allah SWT membalas semua kebaikan yang telah diberikan dengan balasan yang sempurna. Semoga karya ilmiah ini bermanfaat.
Bogor, Mei 2012
Halaman
DAFTAR TABEL ... xv
DAFTAR GAMBAR ... xvi
DAFTAR LAMPIRAN ... xvii
PENDAHULUAN Latar belakang. ... 1
Tujuan Penelitian ... 3
Manfaat Penelitian ... 3
TINJAUAN PUSTAKA Selulosa ... 5
Rumput Laut ... 6
Enzim Selulase... 7
Mikroorganisme Penghasil Enzim Selulase ... 12
Pemekatan Enzim ... 13
Kromatografi Kolom... 15
Elektroforesis ... 19
Identifikasi Mikroorganisme dengan 16S-rRNA ... 20
BAHAN DAN METODE Waktu dan Tempat ... 23
Bahan dan Alat Penelitian... 23
Peremajaan Isolat PMP 0126y ... 24
Pengamatan Morfologi Isolat PMP 0126y... 24
Identifikasi Bakteri secara Molekuler ... 24
Uji Kualitatif Enzim Selulase ... 26
Penentuan Waktu Optimum Produksi Enzim Selulase ... 27
Produksi Enzim Kasar Selulase ... 28
Pemurnian Enzim Selulase ... 29
Analisis Elektroforesis SDS-PAGE dan Zimogram ... 30
Pengukuran Kadar Protein ... 32
Karakterisasi Enzim Selulase ... 32
HASIL Identifikasi Isolat PMP 0126y ... 35
Pertumbuhan dan Produksi Enzim Selulase ... 37
Pemurnian Enzim Selulase ... 40
Analisis Berat Molekul Enzim Selulase Menggunakan SDS- PAGE dan Zimogram ... 42
Pertumbuhan dan Produksi Enzim Selulase ... 50
Pemurnian dan Karakterisasi Enzim Selulase ... 51
SIMPULAN ... 59
DAFTAR PUSTAKA ... 61
Halaman
1 Komposisi kimia rumput laut ... 7
2 Hidrolisis berbagai substrat oleh enzim selulase ... 9
3 Substrat selulosa berdasarkan kelarutan air dan jenis enzim selulase ... 10
4 Metode kromatografi untuk fraksinasi protein ... 15
5 Teknik kromatografi yang digunakan pada pemurnian selulase ... 16
6 Komposisi gel pemisah dan gel penahan untuk sepasang gel ... 30
7 Aktivitas selulase hasil ultrafiltrasi ... 41
8 Hasil uji aktivitas selulase PMP 0126y pada beberapa tahap pemurnian ... 42
Halaman
1 Struktur serat selulosa ... 5
2 Struktur selulosa teratur (kristalin) dan kurang teratur (amorphous) ... 6
3 Pemecahan selulosa menjadi glukosa oleh enzim selulase ... 8
4 Klasifikasi enzim selulase... 9
5 Mekanisme degradasi selulosa ... 11
6 Pemurnian enzim dengan kromatografi penukar ion ... 17
7 Isolat PMP 0126y ... 35
8 Pewarnaan Gram isolat PMP 0126y dengan perbesaran 1000 x ... 35
9 Hasil amplifikasi dari gen penyandi 16S-rRNA isolat PMP 0126y . 36 10 Sebagian sekuen DNA penyandi 16S-rRNA isolat PMP 0126y dari(arah 5’-3’)... 36
11 Pohon filogenetik isolat PMP 0126y ... 37
12 Zona bening isolat PMP 0126y ... 38
13 Kurva pertumbuhan isolat PMP 0126y... 38
14 Kurva aktivitas selulase, aktivitas spesifik, dan jumlah sel bakteri PMP 0126y ... 39
15 Kurva aktivitas selulase, aktivitas spesifik, dan jumlah sel bakteri PMP 0126y pada media glukosa 0,1% ... 39
16 Aktivitas spesifik dari pengendapan amonium selulase dengan amonium sulfat ... 40
17 Profil elusi enzim selulase pada kromatografi DEAE penukar ionmenggunakan matriks Sepharose ... 41
19 Hasil zimogram PMP 0126y pada gel akrilamida yang mengandung CMC 0,1% dan ilustrasi pita yang terbentuk dalam
zimogram ... 44
20 Pengaruh pH terhadap aktivitas selulase PMP 0126y hasil
ultrafiltrasi... 45
21 Pengaruh pH terhadap aktivitas selulase PMP 0126y hasil
kromatografi penukar anion ... 45
22 Suhu optimum aktivitas selulase PMP 0126y hasil ultrafiltrasi
dan kromatografi penukar anion ... 46
23 Pengaruh suhu dan waktu inkubasi terhadap aktivitas
selulase PMP 0126y ... 46
24 Substrat spesifik enzim selulase PMP 0126y hasil ultrafiltrasi ... 47
25 Aktivitas relatif selulase PMP 0126y hasil ultrafiltrasi pada
Halaman 1 Prosedur pembuatan media dan reagen yang digunakan dalam
penelitian ... 73
2 Kurva standar glukosa ... 77
3 Kurva standar bovin serum albumin (BSA) ... 78
4 Kurva hubungan log sel dan kerapatan optis dan jumlah sel isolat
PMP 0126y selama 27 jam pengamatan ... 79
5 Hasil uji aktivitas selulase isolat PMP 0126y ... 80
6 Prosedur delignifikasi limbah rumput laut dengan NaOH dan H2SO4
oleh BBP4BKP ... 82
7 Gambar metafile hasil sekuensing isolat PMP 0126y (primer f) ... 83
PENDAHULUAN
Latar Belakang
Perkembangan industri berbasis hayati termasuk hayati laut dengan memanfaatkan senyawa biologi seperti enzim yang berasal dari mikroorganisme seperti bakteri dan kapang saat ini terus ditingkatkan di berbagai negara. Telah banyak peneliti yang mengisolasi bakteri baru dan memanfaatkan senyawa metabolit bakteri tersebut. Salah satu sumber yang dapat dimanfaatkan pada sektor Kelautan dan Perikanan yaitu limbah hasil pengolahan rumput laut. Mengingat bahwa 75% wilayah Indonesia terdiri atas perairan laut, maka berbagai jenis rumput laut telah banyak dimanfaatkan untuk produk pangan seperti agar-agar maupun karagenan. Pengolahan agar-agar-agar-agar memanfaatkan rumput laut jenis Glacilaria sp., sedangkan karagenan menggunakan rumput laut jenis Eucheuma sp. Berbagai industri rumput laut akan menghasilkan limbah sekitar 65-70% dari bahan baku segar yang masuk dan diolah (Kim et al. 2008).
Peningkatan pengolahan rumput laut Glacilaria sp. untuk diolah menjadi agar-agar tentu saja akan meningkatkan jumlah limbah rumput laut sehingga akan menjadi masalah pencemaran karena limbah tersebut mengandung selulosa yang sulit larut dalam air. Limbah rumput laut Glacilaria sp. mengandung selulosa sebanyak 15-25% (Kim et al. 2008). Salah satu alternatif pemanfaatan yang dapat dilakukan ialah dengan memanfaatkan bakteri asal limbah rumput laut tersebut. Bakteri yang hidup pada limbah ini diduga dapat menghasilkan enzim yang dapat menguraikan limbah selulosa menjadi sumber nutrisi untuk pertumbuhannya. Enzim yang dihasilkan oleh bakteri tersebut dapat menghidrolisis limbah selulosa menjadi glukosa, yang selanjutnya dapat digunakan sebagai bahan untuk fermentasi dalam memproduksi bioetanol. Pemanfaatan limbah selulosa dan bakteri penghasil enzim penghidrolisis selulosa dapat memberikan peluang pada pengembangan bioenergi dari bahan hayati laut.
Crueger 1984). Ketiga enzim ini bekerja secara sinergis mendegradasi selulosa dan melepaskan gula reduksi (selobiosa dan glukosa) sebagai produk akhirnya (Deng & Tabatabai 1994). Enzim selulase akan memutuskan ikatan glikosidik β -1,4 di dalam selulosa yang memiliki ikatan β-1,4-glikosidik pada polimer glukosanya (Jeong et al. 2004) sehingga menjadi gula sederhana turunannya.
Proses hidrolisis selulosa dapat dilakukan dengan menggunakan asam dan suhu tinggi. Proses ini relatif mahal karena kebutuhan energi yang besar serta dapat mengakibatkan degradasi produk monosakarida yang dihasilkan sehingga produk yang akan dihasilkan rendah. Riyanti (2008) juga melaporkan efisiensi proses hidrolisis dengan asam masih rendah karena proses yang dilakukan cukup panjang dan membutuhkan banyak tahap. Kekurangan lain dari proses ini antara lain penanganan limbah asam yang tidak mudah. Baru pada tahun 1980-an, mulai dikembangkan hidrolisis selulosa dengan menggunakan enzim selulase (Coral et al. 2002). Hidrolisis secara enzimatik akan berjalan spesifik dan efisien sehingga produk yang akan dihasilkan lebih tinggi dan menghasilkan produk monosakarida dengan biaya produksi rendah.
NRRL-B-14023 (Ruanglek et al. 2006), Z. mobilis ATCC 10988 (Tanaka et al. 1999).
Selain dalam bidang industri, pemanfaatan enzim selulase dari bakteri dapat memberikan solusi dalam masalah pencemaran yakni mengurangi jumlah limbah selulosa, salah satunya dari industri pengolahan agar-agar dan karagenan, dan mendapatkan produk bernilai tambah dari pemanfaatan limbah rumput laut tersebut. Balai Besar Penelitian Pengembangan Pengolahan Produk dan Bioteknologi Kelautan Perikanan (BBP4BKP) telah melakukan eksplorasi mikrob dari rumput laut termasuk limbah pengolahan rumput laut. Beberapa isolat bakteri yang memiliki aktivitas selulase ekstraseluler yaitu isolat PMP 0126y berhasil diisolasi dari limbah pengolahan agar-agar rumput laut Glacilaria sp. dari daerah Pameungpeuk, Garut Jawa Barat (Munifah et al. 2011).
Tujuan Penelitian
Penelitian ini bertujuan untuk memproduksi, melakukan pemurnian parsial, dan mengkarakterisasi enzim selulase yang dihasilkan oleh isolat PMP 0126y penghasil enzim selulase dari limbah pengolahan rumput laut Glacilaria sp., serta melakukan identifikasi secara molekuler bakteri tersebut.
Manfaat Penelitian
TINJAUAN PUSTAKA
Selulosa
Selulosa merupakan polimer karbohidrat terbanyak yang terdapat di alam (Han & Chen 2007). Selulosa merupakan komponen utama penyusun dinding sel tumbuhan bersama-sama dengan hemiselulosa dan pektin. Komposisi selulosa dalam tumbuhan dapat mencapai 40-50% dari massa tumbuhan sehingga selulosa merupakan biopolimer terbarukan yang paling berlimpah di alam (Milala et al. 2005). Classen (1999) menambahkan bahwa diperkirakan 50% dari biomassa tumbuhan berupa selulosa dan jumlahnya sekitar 50 milyar ton. Selulosa merupakan polimer glukosa yang dihubungkan dengan ikatan β-1,4-D-glukosidik (Gambar 1).
Gambar 1 Struktur serat selulosa (Beguin & Aubert 1994).
Polimer glukosa tersusun secara paralel dan berikatan silang membentuk struktur kristalin yang disebut mikrofibril. Panjang mikrofibril ini bervariasi dari 2.000-15.000 unit glukosa, tergantung organismenya. Bentuk mikrofibril selulosa ditentukan oleh kompleks geometri sintase dan lingkungan lokal. Pada tumbuhan, unit mikrofibril mempunyai jumlah sekitar 3-4 unit dan terdiri atas sekitar 36 rantai selulosa dan seringkali dikemas dalam bentuk lebih besar (Doblin et al. 2002).
kurang teratur (amorphous). Struktur amorphous terjadi karena proses kristalisasi yang berlangsung secara tidak sempurna pada mikrofibril yang terbentuk (Gambar 2). Dimensi serat selulosa dan proporsi dari bagian kristalin dan amorf sangat tergantung pada keadaan alaminya (Linder & Teeri 1997). Setiap serat selulosa tersusun oleh kira-kira 3.000 molekul glukosa dan berat molekulnya diperkirakan mencapai 500.000 (Hardjo et al. 1984).
Gambar 2 Struktur selulosa teratur (kristalin) dan kurang teratur (amorphous) (Beguin & Aubert 1994).
Secara alamiah molekul selulosa tersusun dalam fibril yang terdiri atas beberapa molekul glukosa yang dihubungkan dengan ikatan hidrogen yang kuat mengakibatkan dapat tahan terhadap tarikan tinggi. Fibril-fibril ini membentuk struktur kristal yang dibungkus oleh lignin, oleh karena itu sumber selulosa dari tumbuh-tumbuhan sulit sekali dihidrolisis secara langsung oleh katalis asam. Molekul selulosa berbentuk lurus dan tidak pernah bercabang, serta gugus hidroksilnya bebas membentuk ikatan hidrogen dengan gugus hidroksil molekul selulosa lainnya yang terletak sejajar (paralel) dengannya (Beguin & Aubert 1994).
Rumput Laut
didapatkan presentase masing-masing komponen kadar air sebesar 11.28%, kadar abu 36,05%, kadar lemak 0,42%, kadar protein 1,86%, kadar serat kasar 8,96% dan karbohidrat 41,43% (Harvey 2009).
Jenis rumput laut yang telah banyak dimanfaatkan berasal dari marga Euchema, Gelidium, Gracilaria, Hypnea, dan Sargassum. Selain itu, terdapat jenis lainnya seperti Caulerpa dan Dictosphaeria masih dimanfaatkan dalam skala kecil untuk konsumsi lokal (Atmadja et al. 1996). Beberapa jenis rumput laut memiliki komposisi kandungan selulosa maupun kandungan senyawa kimia lainnya yang berbeda. Berikut ini komposisi kimia dari beberapa jenis rumput laut (Tabel 1).
Tabel 1 Komposisi kimia rumput laut (Kim et al. 2008)
Jenis alga Selulosa
Rumput laut Glacilaria sp. banyak dimanfaatkan dalam industri pengolahan agar-agar. Limbah industri agar-agar yang dihasilkan mengandung selulosa sebesar 15-25% (Kim et al. 2008). Selain itu, limbah agar-agar Glacilaria sp. merupakan salah satu sumber bakteri yang berpotensi menghasilkan enzim selulase. Pemanfaatan limbah agar-agar dan enzim selulase dari bakteri tersebut memegang peranaan yang sangat penting dalam pengembangan bioenergi.
Enzim Selulase
Enzim selulase atau enzim yang dikenal dengan nama sistematik β-1,4 glukan-4-glukano hidrolase adalah enzim yang dapat menghidrolisis selulosa
pemecahan selulosa menjadi glukosa terdiri atas tiga jenis enzim selulase yaitu endo-β-1,4-glukanase, ekso-β-1,4-glukanase, dan β-glukosidase. Endo-β -1,4-glukanase menyerang bagian tengah rantai secara random, ekso-β-1,4-glukanase (selobiohidrolase) memecah unit-unit disakarida (selobiosa) dari ujung rantai, dan
β-glukosidase memecah selobiosa menjadi glukosa (Da silva et al. 2005) (Gambar 3).
Gambar 3 Pemecahan selulosa menjadi glukosa oleh enzim selulase.
Menurut Enari (1983) (Tabel 2) demikian pula Prescott dan Dunns (1981) (Gambar 4) mengelompokkan enzim utama selulase berdasarkan kespesifikan substrat masing-masing enzim yaitu :
1. Endo-β-1,4-glukanase (β-1,4-D-glukan-4-glukanohidrolase, EC 3.2.1.4)
menghidrolisis ikatan glikosidik β-1,4 secara acak. Enzim ini dapat bereaksi dengan selulosa kristal tetapi kurang aktif. Enzim ini secara umum dikenal sebagai CMC-ase atau selulase Cx.
3. β-1,4-D-glukan glukohidrolase (EC.3.2.1.74) menyerang ujung rantai selulosa non pereduksi dan membebaskan glukosa. Enzim ini menghidrolisis selulosa yang telah dilunakkan dengan asam fosfat, selo-oligosakarida dan CMC.
4. β-1,4-glikosidase (β-1,4-D-glukosida glukohidrolase, EC 3.2.1.21) menghidrolisis selobiosa dan rantai pendek selo-oligosakarida yang menghasilkan glukosa. Enzim ini tidak dapat memecah selulosa dan selodekstrin.
Gambar 4 Klasifikasi enzim selulase (Prescott & Dunns 1981).
Tabel 2 Hidrolisis berbagai substrat oleh enzim selulase (Enari 1983)
Jenis Enzim
Berdasarkan kelarutannya, selulosa dapat dibagi menjadi dua katagori yaitu substrat yang larut dalam air dan substrat yang tidak dapat larut dalam air beserta enzim selulase yang menghidrolisis substrat tersebut (Tabel 3).
Tabel 3 Substrat selulosa berdasarkan kelarutan air dan jenis enzim selulase (Zhang et al. 2006)
Substrat Selulosa Enzim Selulase
Larut dalam air
-Rantai pendek (derajat polimerisasi rendah) Silodekstrin
Radio-labeled selodekstrin -Turunan silodekstrin
β-methyllumberlliferil oligosakarida p-nitrofenol oligosakarida
-Turunan selulosa dengan rantai panjang Carboxymethylecellulose (CMC) Dye CMC
-Kromogenik dan turunan fluoreforik Trinitrofenil-karboksimetilselulase
Endo ; endoglukanase, Ekso ; eksoglukanase, BG ; glukosidase, Total ; ketiga tipe enzim selulase.
Perbedaan antara masing-masing enzim selulase terletak pada kespesifikan struktur di sekeliling substrat. Perbedaan kespesifikan dari enzim endoglukanase dan selobiohidrolase bersifat tidak mutlak karena kedua enzim tersebut dapat menghidrolisis ikatan β-1,4 glukosida dari selulosa amorf. Penentuan aktivitas enzim selulase akan sulit apabila filtrat yang akan diukur aktivitas enzimnya merupakan campuran dari berbagai enzim selulase. Enzim-enzim ini tidak hanya dapat menghidrolisis substrat yang sama tetapi juga dapat bekerja secara sinergis memecah substrat yang sama, sehingga menyebabkan aktivitas yang diukur dipengaruhi oleh proporsi dari masing-masing enzim yang ada (Enari 1983).
eksoglukanase seringkali diuji dengan substrat avisel sehingga enzim eksoglukanase disebut dengan aviselase (Zhang et al. 2006).
Tahapan hidrolisis selulosa tergantung kepada struktur selulosa, interaksi antara enzim selulase dengan serat selulosa, mekanisme hidrolisis enzim tersebut di alam dan inhibitor yang terbentuk. Fase adsorbsi dan pembentukan kompleks enzim substrat adalah fase kritis di dalam hidrolisis selulosa. Glukosa dan selobiosa adalah inhibitor enzim dalam menghidrolisis selulosa. Selobiosa menghambat enzim selobiohidrolase dan glukosa menghambat enzim
penghidrolisis selobiosa yaitu β-glukosidase pada kompleks enzim selulase. Selobiosa mempunyai potensi lebih kuat menjadi inhibitor dibandingkan dengan glukosa (Coughlan 1985). Laju hidrolisis enzim selulase ditentukan oleh struktur substrat (Mandels 1985). Struktur kristal lebih sulit dihidrolisis dibandingkan dengan struktur amorf maka hidrolisis dilakukan oleh enzim endoselulase atau endoglukanase (Coughlan 1985) (Gambar 5).
Gambar 5 Mekanisme degradasi selulosa (Beguin & Aubert 1994).
bertambah sampai suhu optimum, kecepatan reaksi enzim naik karena energi kinetik bertambah. Bertambahnya energi kinetik enzim akan mempercepat gerak vibrasi, translasi, dan rotasi baik enzim maupun substrat. Hal ini akan memperbesar peluang enzim dan substrat bereaksi. Ketika suhu lebih tinggi dari suhu optimum, protein enzim berubah konformasi sehingga gugus reaktif terhambat. Perubahan konformasi ini dapat menyebabkan enzim terdenaturasi. Substrat juga dapat berubah konformasinya pada suhu yang tidak sesuai, sehingga substrat tidak dapat masuk ke dalam sisi aktif enzim (Ottaway 1984).
Selain pH dan suhu, faktor lain yang mempengaruhi aktivitas selulase yaitu adanya senyawa penghambat berupa ion logam. Penghambatan tersebut dapat dinetralkan dengan menambahkan sistein sehingga aktivitas enzim dapat berlangsung kembali (Kulp 1975). Beberapa senyawa logam dan senyawa lainnya yang dapat menghambat aktivitas selulase ialah Hg2+, Ag2+, dan Cu2+ (Deng & Tabatai 1994; Oikawa et al. 1994), glukanolakton (Kulp 1975), surfaktan, senyawa pengkelat khususnya Sodium Dodecyl Sulphate (SDS), Ethylene Diamine Tetraacetyc Acid (EDTA) (Oikawa et al. 1994), laktat dalam konsentrasi agak rendah (Chesson 1987), dan etanol serta alkohol lainnya (Ooshima et al. 1985). Senyawa penghambat tersebut dapat menekan seluruh kecepatan hidrolisis dengan menghambat adsorbsi eksoglukanase dan endoglukanase pada selulosa, dan menghambat aksi sinergis eksoglukanase dan endoglukanase yang bekerja pada permukaan selulosa.
Mikroorganisme Penghasil Enzim Selulase
Beberapa contoh genus bakteri yang diketahui mempunyai aktivitas selulolitik ialah Acetobacter, Bacillus, Clostridium, Cellulomonas, Pseudomonas, Cytophaga, Sarcina, dan Vibrio, sedangkan contoh genus cendawan yang mempunyai aktivitas selulolitik ialah Bulgaria, Chaetomium, Helotium, Coriolus, Phanerochaete, Poria, Schizophyllum, Serpula, Aspergillus, Cladosporium, Fusarium, Geotrichum, Myrothecium, Paecilomyces, Penicillium, dan Trichoderma (Rao 1994). Beberapa jenis organisme juga dapat menghasilkan enzim selulase seperti rayap (Watanabe & Tokuda 2001), remis (Xu et al. 2000), dan arabidopsis.
Di alam, degradasi selulosa kebanyakan dilakukan oleh mikroorganisme aerobik. Mikroorganisme aerobik menghasilkan enzim selulase nonkompleks yang terdiri atas endoglukanase, eksoglukanase, dan glukosidase yang bekerja secara sinergis untuk menghidrolisis selulosa. Mikroorganisme anaerobik menghasilkan enzim selulase kompleks yang disebut selulosom (Doi et al. 2003; Bayer et al. 2004). Meskipun mikroorganisme anaerobik hanya menyumbang sekitar 5-10% dari biodegradasi total selulosa di alam, namun peranannya sangat penting karena bertanggung jawab terhadap degradasi daerah anoksik pada danau, laut, dan saluran pencernaan hewan pemamah biak maupun rayap, yang tidak dapat dilakukan oleh mikroorganisme aerobik (Zhang et al. 2006).
Pemekatan Enzim
Pada tahap awal pemurnian enzim biasanya dilakukan klarifikasi dan pengendapan protein enzim. Klarifikasi berfungsi memisahkan larutan enzim dari partikel-partikel yang tidak larut, misalnya debris sel dan partikel substrat. Klarifikasi dapat dilakukan dengan penyaringan atau sentrifugasi. Pemekatan protein enzim merupakan tahap awal dari prosedur pemurnian enzim sebelum tahap pemurnian berikutnya atau dapat pula digunakan untuk keperluan analisis enzim. Pemekatan protein enzim berfungsi untuk meningkatkan konsentrasi protein enzim, mereduksi volume larutan enzim, dan memisahkan protein enzim dengan protein pengotor yang lain (Harris 1989).
etanol), dan imunopresipitasi yang dapat menyebabkan denaturasi protein. Pemekatan protein dengan metode preparatif tetap mempertahankan aktivitas protein misalnya dengan menggunakan pengendapan garam, pengendapan dengan pelarut organik, pengendapan dengan polimer organik, ultrafiltrasi, liofilisasi, dan dialisis (Harris 1989).
Metode pengendapan protein yang biasa dilakukan dalam pengendapan selulase ialah dengan menggunakan amonium sulfat (Jung et al. 2008) dan ultrafiltrasi (Arifin 2006). Amonium sulfat merupakan garam yang paling sering digunakan untuk mengendapkan protein karena memiliki daya larut tinggi di dalam air, relatif tidak mahal, dan kestabilan protein di dalam larutan amonium sulfat (2M- 3M) tahan bertahun-tahun (Scopes 1987).
Prinsip pengendapan dengan garam berdasarkan pada kelarutan protein yang berinteraksi polar dengan molekul air, interaksi ionik protein dengan garam, dan daya tolak menolak protein yang bermuatan sama. Kelarutan protein (pada pH dan suhu tertentu) meningkat pada kenaikan konsentrasi garam (salting in). Kenaikan kelarutan protein akan meningkatkan kekuatan ion larutan. Pada penambahan garam dengan konsentrasi tertentu menyebabkan kelarutan protein menurun (salting out). Molekul air yang berikatan dengan ion-ion garam semakin banyak yang menyebabkan penarikan selubung air yang mengelilingi permukaan protein. Peristiwa ini mengakibatkan protein saling berinteraksi, beragregasi, dan kemudian mengendap (Harris 1989; Scopes 1987).
pori yang berukuran kecil, misalnya Sephadex G-25 buatan Phamacia. Kekurangan metode ini adalah terjadi pengenceran sampel protein (Harris 1989). Ultrafiltrasi merupakan suatu metode untuk mengkonsentrasikan protein dengan menekan cairan larutan protein enzim supaya tertahan di dalam membran. Ukuran cairan yang akan ditahan (retentat) dan yang dikeluarkan (permeat) sesuai dengan ukuran membran yang digunakan. Prinsip pemisahan dengan ultrafiltrasi adalah pemisahan komponen berdasarkan berat molekul (Bollag & Edelstein 1991). Pemisahan komponen ini terjadi karena adanya membran ultrafiltrasi. Membran ultrafiltrasi berfungsi sebagai penghalang (barrier) tipis yang sangat selektif di antara dua fasa, hanya dapat melewatkan komponen tertentu dan menahan komponen lain dari suatu aliran fluida yang dilewatkan melalui membran (Mulder 1996). Proses membran ultrafiltrasi merupakan upaya pemisahan dengan membran yang menggunakan gaya dorong beda tekanan yang dipengaruhi oleh ukuran dan distribusi pori membran (Malleviale 1996).
Kromatografi Kolom
Kromatografi kolom pada prinsipnya yaitu pengaliran suatu cairan melalui kolom yang mengandung bahan pengisi dan substanta yang ingin dipisahkan menjadi beberapa komponen dengan adanya perbedaan terhadap daya ikat bahan pengisi (Tabel 4).
Tabel 4 Metode kromatografi untuk fraksinasi protein (Ersson et al. 1998)
Sifat Protein Jenis Kromatografi
Ukuran dan bentuk Filtrasi gel
Muatan neto dan distribusi grup bermuatan
Penukar ion
Titik isoelektris Kromatofokusing
Hidrofobisitas Interaksi hidrofobik dan fase balik Pengikatan logam Afinitas ion logam terimobilisasi Kandungan tiol yang terbuka Kovalen
Afinitas biospesifik terhadap ligan, inhibitor, reseptor, antibodi, dsb
Afinitas
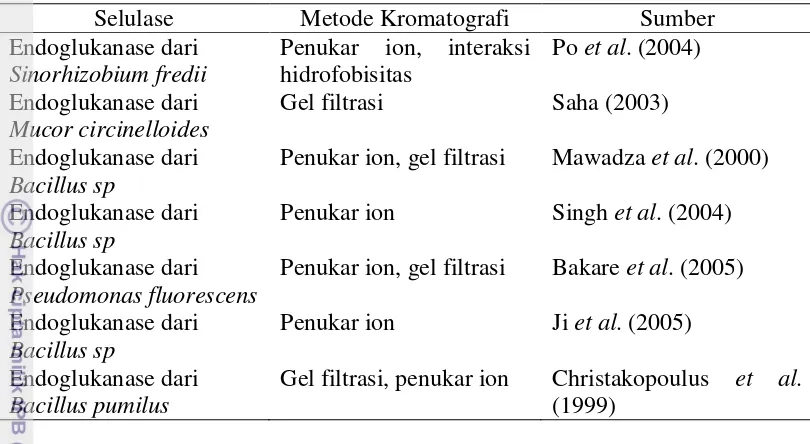
Tabel 5 Teknik kromatografi yang digunakan pada pemurnian selulase
Selulase Metode Kromatografi Sumber
Endoglukanase dari Sinorhizobium fredii
Penukar ion, interaksi hidrofobisitas
Po et al. (2004)
Endoglukanase dari Mucor circinelloides
Gel filtrasi Saha (2003)
Endoglukanase dari Bacillus sp
Penukar ion, gel filtrasi Mawadza et al. (2000)
Endoglukanase dari Bacillus sp
Penukar ion Singh et al. (2004)
Endoglukanase dari Pseudomonas fluorescens
Penukar ion, gel filtrasi Bakare et al. (2005)
Endoglukanase dari Bacillus sp
Penukar ion Ji et al. (2005)
Endoglukanase dari Bacillus pumilus
Gel filtrasi, penukar ion Christakopoulus et al. (1999)
Kolom untuk kromatografi penukar ion biasanya tidak panjang dan memiliki diameter lebih besar dari pada kolom untuk filtrasi gel. Banyaknya sampel yang dimasukkan umumnya sekitar 10-20% dari kapasitas kolom. Pembilasan dengan gradien konsentrasi NaCl yang linier baik digunakan untuk memisahkan molekul-molekul yang memiliki perbedaan muatan bersih yang tidak terlalu besar sedangkan gradien NaCl bertahap baik digunakan untuk memisahkan molekul-molekul yang memiliki perbedaan muatan bersih yang besar.
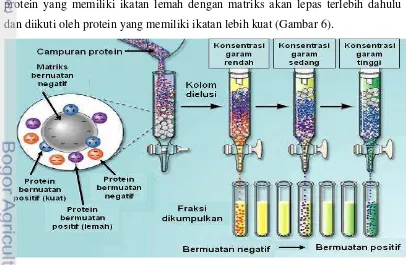
Pada dasarnya prinsip kromatografi penukar ion adalah ion bermuatan bebas dipertukarkan dengan ion yang memiliki tipe muatan yang sama. Protein yang bermuatan negatif dapat ditukar dengan ion klorida. Awalnya gugus fungsional matriks yang bermuatan negatif mengikat ion dari bufer (misalnya Na+). Pada saat sampel dimasukkan ke dalam kolom, maka protein yang bermuatan positif akan menggantikan ion Na+ sedangkan protein yang bermuatan negatif atau netral tidak akan terikat. Protein yang tidak terikat dibilas dengan menggunakan bufer (biasanya dengan konsentrasi 10-50 mM). Selanjutnya ikatan protein yang terikat gugus fungsional matriks akan terlepas setelah dibilas dengan bufer yang mengandung NaCl atau KCl secara linier atau bertahap sehingga protein yang memiliki ikatan lemah dengan matriks akan lepas terlebih dahulu dan diikuti oleh protein yang memiliki ikatan lebih kuat (Gambar 6).
Gambar 6 Pemurnian enzim dengan kromatografi pertukar ion
Pemilihan penukar ion tergantung pada muatan protein target. Muatan bersih protein tergantung pada pH yaitu protein akan bermuatan positif dengan menurunkan pH dan bermuatan negatif dengan menaikkan pH. Pada saat menentukan pH untuk kromatografi, kestabilan protein target pada pH yang dipilih perlu dijaga. Apabila protein stabil pada pH di atas titik isoelektriknya (pI) maka digunakan penukar anion (positif), tetapi bila protein stabil pada pH di bawah pI nya maka digunakan penukar kation (negatif). Jika protein stabil pada rentang 1 unit di atas dan di bawah pI maka kedua penukar ion dapat digunakan. Matriks yang mengikat gugus fungsional menentukan sifat aliran, ion yang dapat diikat, kestabilan mekanik dan kimia. Ada 3 kelompok matriks yang biasanya digunakan, yaitu: 1) polistiren, poliakrilik atau polifenol; 2) selulosa; dan 3) dekstran (Sephadex) atau agarosa (Sepharose). Matriks polistiren dan polifenolik lebih sering digunakan untuk memisahkan molekul-molekul kecil seperti asam-asam amino, peptida kecil, nukleotida, nukleotida siklik, asam-asam-asam-asam organik. Matriks selulosa biasanya digunakan untuk memisahkan protein (termasuk enzim), polisakarida dan asam nukleat. Matriks DEAE-selulosa, CM-selulosa dan fosfoselulosa paling sering digunakan. Matriks polidekstran dan agarosa (misalnya DEAE-Sephadex, CM-Sephadex) digunakan untuk memisahkan protein, hormon, tRNA dan polisakarida (Scopes 1987).
Elektroforesis
Elektroforesis adalah suatu proses perpindahan partikel-partikel bermuatan atau suatu cara untuk memisahkan fraksi-fraksi suatu campuran berdasarkan atas pergerakan partikel koloid yang bermuatan di bawah pengaruh medan listrik (Suhartono 1989). Elektoforesis dengan menggunakan gel polakrilamida sodium dodesil sulfat (SDS-PAGE) merupakan teknik elektroforesis gel yang menggunakan poliakrilamida untuk memisahkan protein yang bermuatan berdasarkan berat molekulnya. Penentuan berat molekul yang menyusun enzim selulase dianalisis dengan menggunakan metode SDS-PAGE (Sodium dodecyl sulphate-polyacrylamide gel electrophoresis). Pada metode ini digunakan 2 gel yaitu gel penahan (stacking gel) dan gel pemisah (separating gel). Gel akrilamida diperoleh dengan cara polimerisasi akrilamida dengan sejumlah crosslinking agent metilen bis akrilamida dan amonium persulfat (APS) sebagai katalisator. Radikal bebas yang terbentuk dari pelarutan amonium persulfat dalam air akan bereaksi dengan akrilamida membentuk akrilamida aktif yang dapat bereaksi satu dengan yang lain membentuk polimer (Janson & Ryden 1998).
Ada beberapa jenis elektroforesis, yaitu elektroforesis kertas, elektroforesis selulosa asetat/nitrat dan elektroforesis gel. Elektroforesis gel berguna untuk pemisahan protein, sedangkan dua jenis lainnya berguna untuk memisahkan molekul yang lebih kecil. Matriks gel dapat berupa pati, agarosa atau poliakrilamida. Saat ini gel poliakrilamida lebih sering digunakan. Matriks ini disusun oleh akrilamida dan N,N’-metilen-bis-akrilamida yang berpolimerisasi
dengan bantuan katalisator amonium persulfat dan N,N,N’,N’tetrametilen diamin
Elektroforesis protein dapat dilakukan dengan proses denaturasi (SDS-PAGE) dan nondenaturasi (Native-(SDS-PAGE). Mekanisme pada SDS-PAGE dijelaskan bahwa protein akan bereaksi dengan SDS yang merupakan detergen anionik membentuk kompleks yang bermuatan negatif. Protein akan terdenaturasi dan terlarut membentuk kompleks berikatan dengan SDS yang berbentuk elips atau batang yang ukurannya sebanding dengan berat molekul protein. Protein dalam bentuk kompleks yang bermuatan negatif ini akan dapat terpisahkan berdasarkan muatan dan ukurannya secara elektroforesis di dalam matriks gel poliakrilamida (Smith 1984).
Berbeda dengan SDS-PAGE, pada gel pemisah disisipi substrat yang akan dihidrolisis oleh enzim selama masa inkubasi yang disebut sebagai zimogram. Elektroforesis zimogram memisahkan protein terlarut yang tidak mengendap atau beragregasi selama elektroforesis. Pada elektroforesis gel yang terdenaturasi, seperti pada SDS-PAGE, molekul-molekul protein yang telah terpisah dengan elektroforesis dapat kehilangan aktivitas biologi dan biokimianya, tetapi pada elektroforesis zimogram aktivitas tersebut masih bertahan (Dunn 1989). Enzim dipisahkan dalam gel denaturasi (SDS), namun dalam kondisi tidak tereduksi. SDS dilepaskan dengan penambahan larutan renaturasi (misalnya detergen Triton X-100) dan kembali terjadi pelipatan protein. Kemudian gel diwarnai dengan pewarna yang sesuai dengan enzim yang diujikan. Metode zimogram bersifat mudah, sensitif, dan kuantitatif dalam menganalisis aktivitas enzim (Kleiner & Stetler-Stevenson 1994; Leber & Balkwil 1997).
Berat molekul protein dapat ditetapkan dengan menggunakan protein standar yang telah diketahui berat molekulnya dan memperbandingkan nilai Rf (mobilitas relatif) yang diperoleh. Pita pada gel dapat divisualisasi dengan pewarnaan, misalnya menggunakan pewarna coomasie blue atau pewarna perak nitrat (Suhartono 1989).
Identifikasi Mikroorganisme dengan 16S-rRNA
digunakan untuk menduga hubungan kekerabatan secara alami antara spesies yang mempunyai kekerabatan dekat sehingga sangat menguntungkan untuk analisis filogenetik bakteri di tingkat famili, genus, spesies, maupun subspesies. (Chen et al. 2000). Woese (1987) menambahkan bahwa molekul 16S-rRNA paling banyak digunakan sebagai target asam nukleat untuk mendeteksi dan mengidentifikasi bakteri yang belum pernah terdeteksi sebelumnya. Sekuen variabel berevolusi pada laju yang berbeda sehingga memberikan cukup informasi untuk menentukan kedekatan atau jauhnya hubungan filogenetik suatu organisme (Woese 1987).
Madigan et al. (2000) menyatakan sekuen gen penyandi 16S-rRNA digunakan untuk menentukan pohon filogenetik dari keragaman makhluk hidup di bumi. Kekerabatan evolusi antar spesies dalam keseluruhan sistem biologi diperlukan parameter yang memenuhi persyaratan sebagai berikut : 1) terdapat pada semua makhluk hidup, 2) fungsinya identik, 3) dapat dibandingkan secara obyektif, dan 4) parameter tersebut berubah sesuai dengan jarak evolusinya sehingga dapat dijadikan sebagai kronometer evolusi yang handal.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini dilakukan pada bulan Februari 2011 sampai Februari 2012 di Laboratorium Bioteknologi, Balai Besar Penelitian Pengembangan Pengolahan Produk dan Bioteknologi Kelautan Perikanan (BBP4BKP), Jakarta.
Bahan dan Alat Penelitian
Bahan yang digunakan pada penelitian ini antara lain ialah isolat PMP 0126y koleksi dari BBP4BKP hasil isolasi dari limbah pengolahan rumput laut Glacilaria sp. dari daerah Pameungpeuk Jawa Barat. Beberapa bahan kimia yang digunakan dalam penelitian ini yaitu agar-agar nutrien (NA), kaldu nutrien (NB), Carboxymethyl Cellulose (CMC), MgSO4.7H2O, K2HPO4, FeSO4.7H2O,
CaCl2.2H2O, ekstrak khamir, NH4NO3, KH2PO4, glukosa. Bahan kimia lain yang
digunakan antara lain yaitu bovin serum albumin (BSA) standar, sodium tartarat, asam dinitrosalisilat (DNS), bufer sitrat-fosfat, bufer asetat, bufer tris-HCl, NaCl, etanol, merah kongo, sodium dodesil sulfat (SDS), Triton X-100, glysin, dan membran ultrafiltrasi yaitu polyetersulfon (Model UFP-10-E-4MA, dengan area permukaan 420 cm2 dan tipe membran sebesar 10.000 NMWC (Nominal Molecular Weigth Cutoff)) (GE Healthcare Bio-Sciences Corp), matriks DEAE SepharoseTM Fast Flow (Amersham Bioscience, Upsalla Sweden).
Applied Biosystem dan BIOMETRA Tprofesional Thermoclyne), Microspin (FV-2400), piranti elektroforesis SDS-PAGE (Amersham Bioscience, Swedia), piranti elektroforesis DNA (Portsmouth NH, USA), batang pengaduk, Akta Purifier (Amersham Biosciences UPC-900, Upsalla Sweden), Blok panas (Biometra), Ultrafiltrasi (Watson Marlow).
Peremajaan Isolat PMP 0126y
Peremajaan isolat PMP 0126y dilakukan dengan menumbuhkan isolat bakteri pada media agar-agar nutrien (NA). Bakteri tersebut diinkubasi di dalam inkubator selama 1 x 24 jam pada suhu 37 0C (Munifah et al. 2011). Kemudian, dilanjutkan dengan pengamatan secara morfologi bakteri yaitu pewarnaan Gram.
Pengamatan Morfologi Isolat PMP 0126y
Morfologi isolat PMP 0126y diamati dengan melakukan pewarnaan Gram yang dilihat dengan menggunakan mikroskop. Pewarnaan Gram dilakukan dengan cara memfiksasi bakteri pada kaca objek gelas dengan menggunakan larutan KH2PO4 (Lampiran 1) sebanyak 3 tetes di atas api bunsen. Preparat olesan bakteri
yang telah difiksasi panas digenangi pewarna ungu kristal violet selama 1 menit, dibilas dengan air, dan ditiriskan. Olesan digenangi iodium Gram selama 1 menit dan dicuci dengan 95% etanol (decoloration solution) selama 30 detik sampai pewarna ungu kristal pada preparat tidak terbilas lagi dan dicuci dengan akuades sampai warna olesan menjadi bening. Olesan digenangi kembali dengan larutan safranin selama 1 menit, dibilas dengan akuades, dan ditiriskan sampai kering. Bakteri yang telah diwarnai diamati dengan mikroskop medan terang pada perbesaran 1000-2000 x (Cappucino & Sherman 1983). Hasil pewarnaan Gram isolat PMP 0126y difoto menggunakan kamera mikroskop (Olympus DP12) yang dikerjakan di laboratorium Mikrobiologi, BBP4BKP.
Identifikasi Bakteri secara Molekuler
analisis isolasi bakteri secara molekuler meliputi a) isolasi DNA total, b) amplifikasi gen penyandi 16S-rRNA dengan PCR, c) verifikasi dengan elektroforesis gel agarosa, d) ekstraksi DNA dari agarosa, e) cycle sequencing, f) purifikasi hasil PCR, dan g) sequencing hasil PCR.
Isolasi DNA Total (Maniatis et al. 1989). Isolasi DNA total dilakukan dengan menggunakan kit Genomic DNA Purification (Fermentas Life Biosciences, EU). Isolat bakteri dikulturkan pada media kaldu nutrien selama 12-14 jam. Sebanyak 1,5 mL kultur dimasukkan ke dalam tabung mikro dan disentrifugasi selama 10 menit dengan kecepatan 9000 x g. Supernatan dibuang dan ditambahkan kultur lagi berulang-ulang sampai diperoleh pelet dalam jumlah yang cukup. Ke dalam pelet ditambahkan 200 µL bufer TE dan 50 µL lisozim (10 mg dalam 167 ml), dibolak-balik dan diinkubasi selama semalam pada suhu 37 0C. Selanjutnya ke dalam tabung mikro ditambahkan 200 µL bufer lisis, diinkubasi pada suhu 65 0C selama 10 menit (setiap 3 menit dilakukan inversi/tabung dibolak-balik). Kemudian ditambahkan 600 µL kloroform, diinversi perlahan sampai terbentuk dua fase yaitu fase atas dan fase bawah. Selanjutnya disentrifugasi pada suhu 4 0C dengan kecepatan 13.000 x g selama 10 menit. Saat sedang dilakukan sentrifugasi, disiapkan larutan pengendapan dengan mencampurkan 80 µL larutan pengendapan dengan 720 µL air distilasi. Setelah sentrifugasi selesai dilanjutkan dengan mengambil fase atas/fase cair (aqueous phase) perlahan-lahan dan dimasukkan ke dalam larutan pengendapan. Pada saat dimasukkan ke dalam larutan pengendapan akan terlihat benang-benang DNA dan didiamkan selama 2 menit pada suhu ruang.
DNA yang mengendap dikeringkan sebelum diresuspensi dengan bufer TE untuk penyimpanan di dalam lemari es suhu 4 0C.
Amplifikasi Gen Penyandi 16S-rRNA dengan PCR (Suwanto et al. 2000). DNA template diamplifikasi dengan PCR menggunakan dua primer universal spesifik untuk bakteri yaitu 63f (5’-CAGGCCTAACACAGGCAAGTC)
dan 1387r (5’-GGGCGGWGTGTACAAGGC) (Marchesi et al. 1998). Ke dalam tabung mikro steril dimasukkan 18 µL ddH2O; 1,0 µL primer 63f; 1,0 µL primer
1387r; dan 25 µL Taq polymerase, kemudian dimasukkan ke dalam PCR. Kondisi PCR terdiri atas tahap: pre-PCR (95 0C, 5 menit), denaturasi (95 0C, 1 menit), annealing atau pelekatan primer (56 0C, 1 menit 15 detik), elongasi atau pemanjangan primer (72 0C, 1 menit 30 detik), post-PCR (72 0C, 7 menit), dan penyimpanan/pendinginan (4 0C). Proses PCR tersebut dilakukan sebanyak 30 siklus. Hasil PCR kemudian divisualisasi dengan elektroforesis 1% gel agarosa.
Proses selanjutnya yaitu ekstraksi DNA dari agarosa, analisis sekuen parsial gen penyandi 16S-rRNA, dan sequencing hasil PCR dilakukan oleh 1st base, Singapura. Data sekuen DNA yang telah diperoleh dibandingkan dengan data sekuen di Gene Bank untuk menentukan pohon filogenetiknya. Analisis klaster dilakukan dengan menggunakan program dari National Center Biotechnology Information (NCBI) (Van de Peer & De Watcher 1993), sedangkan pembuatan pohon filogenetik menggunakan program Clustal X2 dan NJ-plot.
Uji Kualitatif Enzim Selulase
aktivitas selulase (IAS) diperoleh dengan menggunakan rumus sebagai berikut (Kader & Omar 1998):
Indeks selulolitik =
Penentuan Waktu Optimum Produksi Enzim Selulase
Penentuan waktu optimum produksi enzim selulase diawali dengan penentuan waktu penuangan inokulum. Hal ini dilakukan agar dapat diketahui waktu pertumbuhan eksponensial bakteri pada inokulum yang akan digunakan. Penentuan waktu inokulum dilakukan dengan mengkultur 2 lup isolat di dalam 10 mL kaldu nutrien dan diinkubasi selama 12-14 jam, kemudian dituang ke dalam 50 mL media cair CMC. Kultur diinkubasi pada suhu 30 0C di dalam penangas goyang dengan kecepatan agitasi 150 rpm. Pengambilan sampel dilakukan selama 27 jam inkubasi dengan rentang waktu sampling 3 jam untuk diukur nilai Optical Density (OD) pada panjang gelombang 600 nm. Setelah itu, dibuat kurva pertumbuhan bakteri untuk menentukan waktu yang terbaik pada penuangan inokulum pada media produksi. Selanjutnya, dilakukan penghitungan jumlah koloni total pada cawan (TPC) untuk memperkirakan jumlah sel bakteri pada setiap nilai OD yang dihasilkan.
Setelah waktu penuangan inokulum ke dalam media produksi diketahui, dilanjutkan dengan penentuan waktu optimum aktivitas enzim selulase. Sebanyak 5 mL kaldu nutrien yang telah mengandung biakan sel diinokulasikan ke dalam 25 mL media inokulum yang mengandung glukosa 0,1%. Inokulum tersebut dituang ke dalam 250 mL media produksi tanpa glukosa (sebanyak 10% dari media produksi). Waktu penuangan inokulum dilihat dari waktu pertumbuhan eksponensial bakteri (fase pertumbuhan logaritmik) yang telah diketahui dari kurva pertumbuhan bakteri. Pengambilan sampel dilakukan setiap hari selama 6 hari waktu inkubasi dilakukan.
fosfat pH 5, kemudian ditambah dengan 0,2 mL enzim selulase, dikocok kuat dengan vortex, selanjutnya diinkubasi selama 30 menit pada suhu 30 0C, dan reaksi enzim dihentikan dengan pendidihan pada suhu 100 0C selama 15 menit. Setelah itu, diambil sebanyak 1 mL dari campuran reaksi dan ditambah dengan 1 mL DNS, dididihkan pada suhu 100 0C selama 15 menit. Setelah larutan dingin absorbansi diukur pada λ 575 nm. Perlakuan kontrol dan blanko dilakukan secara bersamaan dengan metode dan tahapan yang sama. Pada kontrol, enzim yang akan direaksikan dengan substrat telah diinaktivasi terlebih dahulu dengan memanaskan enzim selama 15 menit dalam air mendidih. Pada blanko, larutan enzim diganti dengan akuades untuk direaksikan dengan substrat. Aktivitas enzim diukur pada setiap pengambilan sampel yang dilakukan sehingga dapat diketahui waktu optimum produksi enzim selulase.
Aktivitas selulase dinyatakan dalam satuan internasional yaitu U/mL. Satu unit merupakan jumlah enzim yang dibutuhkan untuk memecah 1 µmol selulosa menjadi gula pereduksi per menit pada kondisi pengujian. Kadar glukosa yang dihasilkan dari hidrolisis selulosa dengan enzim selulase berdasarkan nilai
absorbansi pada λ 575 nm.
Absorbansi = ((As - Ab) - (Ak - Ab))
Nilai absorbansi yang diperoleh kemudian dimasukkan ke dalam persamaan yang diperoleh dari kurva standar glukosa (Lampiran 2). Kemudian, aktivitas selulase dihitung berdasarkan rumus sebagai berikut (Irawan et al. 2008) yang dimodifikasi.
Aktivitas selulase (U/mL) =
Keterangan : As = Absorbansi sampel
Ab = Absorbansi blanko
Ak = Absorbansi kontrol
V = volume enzim (0,2 mL) t = waktu inkubasi (30 menit)
BM = Berat molekul glukosa (180 Dalton)
Produksi Enzim Kasar Selulase
selulase yang dihasilkan. Media pertumbuhan produksi diinkubasi pada suhu 30 0C di dalam penangas goyang dengan kecepatan agitasi 150 rpm, kemudian enzim selulase dipanen selama waktu produksi tertinggi yang telah didapatkan sebelumnya.
Kultur sel pada media produksi yang mengandung enzim selulase ekstraseluler disentrifugasi pada kecepatan 10.000 x g selama 15 menit untuk memisahkan larutan enzim dengan pelet bakteri. Supernatan hasil sentrifugasi kemudian disimpan pada suhu 10 0C sebagai enzim ekstrak kasar.
Pemurnian Enzim Selulase
Pemurnian awal enzim dilakukan dengan melakukan pemekatan enzim menggunakan ultrafiltrasi dan pengendapan amonium sulfat. Pemekatan enzim ekstrak kasar dengan ultrafiltrasi pada penelitian ini dilakukan dengan menggunakan alat ultrafiltrasi dan membran filtrasi. Enzim ekstrak kasar dimasukkan ke dalam tabung dan kecepatan pompa ultrafiltrasi sebesar 200-250 rpm. Pemekatan enzim dilakukan sampai 10 kali pemekatan, sehingga pada akhirnya akan menghasilkan enzim hasil ultrafiltrasi dan filtrat yang keluar dari membran filtrasi.
dipilih salah satu metode pemekatan berdasarkan hasil uji aktivitas selulase tertinggi.
Enzim hasil pemekatan dimurnikan dengan menggunakan Akta Purifier. Proses purifikasi dengan kromatografi yang dilakukan tergolong ke dalam kromatografi penukar anion (KPA) dengan menggunakan kolom (40 cm, diameter 50 mm). Matriks DEAE SepharoseTM Fast Flow sebagai fase diamnya, dan bufer Tris-HCl 0,05 M pH 8 dengan gradien konsentrasi 1 M NaCl dalam Tris-HCl 0,05 M pH 8 sebagai fase geraknya. Matriks sepharose merupakan cross-linked agarosa 6% berbentuk bola berukuran 45-165 µm, dapat bekerja pada suhu 4-40 0C dan stabil pada pH 2-14 (GE Healthcare). Kecepatan alir eluen 1 mL/menit. Volume selulase yang dimurnikan sebanyak 4 mL. Volume fraksi
yang ditampung masing-masing sebanyak 5 mL. Serapan setiap fraksi yang ditampung diukur oleh alat spektrofotometer (mAu) yang terdapat pada alat Akta Purifier. Hasil pemurnian dengan Akta Purifier selanjutnya diuji aktivitas enzim selulasenya.
Analisis Elektroforesis SDS-PAGE dan Zimogram
Elektroforesis protein dilakukan dengan dua metode yaitu elektroforesis SDS-PAGE dan Zimogram. Elektroforesis dilakukan dengan menggunakan 10% poliakrilamida sebagai gel pemisah dan 4% poliakrilamida sebagai gel pengumpul atau penahan (Tabel 6).
Tabel 6 Komposisi gel pemisah dan gel penahan untuk sepasang gel
Komposisi 10% Gel Pemisah 4% Gel
Penahan (mL) SDS (mL) Zimogram (mL)
Akuades 3,4 2,4 3,05
Substrat CMC - 1 -
1,5 M Bufer Tris-HCl pH 8,8 2,5 2,5 -
0,5 M bufer Tris-HCl pH 6,8 - - 1,25
10% SDS 0,1 0,1 0,05
30% akrilamida/bis 4 4 0,65
10% Amonium Persulfat 0,05 0,05 0,05
TEMED (N,N,N’,N’
-tetrametilen-etilendiamin 0,025 0,025 0,025
dalam tabung mikro. Sampel protein yang telah dicampur dengan bufer sampel dipanaskan di dalam blok panas selama 5-7 menit, kecuali pada sampel untuk zimogram tidak dipanaskan. Kemudian sebanyak 20 µL campuran tersebut dimasukkan ke dalam sumur pada gel penahan menggunakan mikropipet 10 µL. Setelah gel dipasang dalam piranti elektroforesis, sebayak 300-400 mL 1X bufer elektroforesis (Lampiran 1) dituangkan pada tempatnya.
Proses elektroforesis berlangsung selama 2 jam pada tegangan 100 volt dan 50 mA di dalam piranti elektroforesis (Amersham Bioscience, Swedia). Setelah selesai, gel dilepas dan jarak migrasi diukur dari batas atas gel pemisah. Gel SDS-PAGE kemudian direndam dalam larutan pewarna perak nitrat berdasarkan protokol kit Fermentas dengan berbagai tahapan perendaman dengan berbagai larutan yaitu larutan peluntur gel 1 dan 2, larutan sensitizer, larutan pewarna, larutan pencuci gel, dan larutan akhir (Lampiran 1). Setelah itu, pita protein hasil elektroforesis terlihat dan difoto.
Pada gel elektroforesis untuk zimogram, gel kemudian direnaturasi dengan merendam gel di dalam 2,5% Triton X-100 selama satu jam sambil digoyang konstan. Gel ditiriskan dan direndam dalam 0,05 M bufer sitrat fosfat pH 5 selama 1,5-2 jam sambil digoyang perlahan dalam inkubator goyang pada suhu 30 0C. Kemudian gel diwarnai dengan 0,1% kongo merah selama 30 menit, selanjutnya direndam dengan 1 M NaCl selama 15 menit (perendaman dilakukan sebanyak tiga kali). Zona bening di sekitar pita yang terbentuk dibandingkan dengan penanda berat molekul sehingga dapat diketahui berat molekul enzim selulase yang dapat menghidrolisis substrat CMC pada gel akrilamida.
Perkiraan berat molekul relatif ditentukan dengan membandingkan migrasi pita protein dengan pita standar penanda massa molekul relatif berberat molekul rendah (14,4-97 kDa, GE) dan molekul tinggi (53-220 kDa, GE). Standar protein berat molekul rendah terdiri atas Fosforilase b (otot kelinci) 97 kDa, albumin (serum bovin) 66 kDa, ovalbumin (putih telur) 45 kDa, karbonat anhidrase
(eritrosit bovin) 30 kDa, tripsin inhibitor (kedelai) 20,1 kDa, dan α-laktalbumin (susu bovin) 14,4 kDa. Standar berat molekul tinggi terdiri atas miosin (otot
(E. Coli) 116 kDa, transferin (manusia) 76 kDa, dan glutamat dehidrogenase (hati bovin) 53 kDa.
Pengukuran Kadar Protein
Pengukuran kadar protein bertujuan untuk mengukur kandungan protein yang terdapat dalam enzim selulase yang dihasilkan menggunakan metode Bradford (1976). Sebanyak 20 µL enzim direaksikan dengan 1,0 mL Coomassie Brilliant Blue G-250 kemudian dikocok kuat dengan vortex. Absorbansi dibaca
pada λ 595 nm. Blanko menggunakan 20 µL air distilasi yang direaksikan dengan 1,0 mL Coomassie Brilliant Blue G-250. Standar protein menggunakan bovine serum albumin (BSA) pada kisaran 0,1-1,0 mg protein/mL dari 2 mg/mL larutan stok BSA. Pengujian mikro dalam mengukur kadar protein menggunakan BSA pada kisaran 0,01-0,1 mg protein/mL.
Karakterisasi Enzim Selulase
pH Optimum. Pengaruh pH terhadap aktivitas enzim diuji dengan menambahkan 0,2 mL enzim yang direaksikan dengan 1,8 mL substrat. Substrat dibuat dengan mencampurkan 1,8 g CMC ke dalam bufer dengan berbagai tingkatan pH 3-9, antara lain yaitu 0,05 M bufer asetat (3, 4, 5), 0,05 M bufer sitrat fosfat (5, 6, 7), dan 0,05 M bufer tris-HCl (7, 8, 9). Masing-masing enzim diinkubasi pada suhu 30 0C selama 30 menit. Aktivitas enzim selulase diukur sesuai dengan prosedur pengujian sebelumnya.
Suhu Optimum. Pengaruh suhu terhadap aktivitas enzim dilakukan dengan mereaksikan 0,2 mL enzim dengan 1,8 mL substrat di mana substrat dibuat dengan mencampurkan 1,8 g CMC dalam bufer pH optimum. Enzim yang telah dicampurkan dengan substrat kemudian diinkubasi pada tingkatan suhu antara 30 0C sampai dengan 90 0C dengan selang 10 0C selama 30 menit waktu inkubasi. Aktivitas enzim selulase diukur sesuai dengan prosedur pengujian sebelumnya.
pada suhu dan pH optimumnya dikondisi pengujian sebelumnya (Jung et al. 2008).
Substrat Spesifik. Pengujian aktivitas selulase pada berbagai substrat dilakukan dengan CMC teknis, CMC murni, avisel, kertas Whatman filter paper No. 1, limbah rumput laut pengolahan agar-agar PT. Agarindo yang didelignifikasi dengan NaOH 6%, Limbah rumput laut pengolahan agar-agar Pemeungpeuk yang didelignifikasi dengan 4 dan 6% NaOH serta 1% H2SO4,
limbah pengolahan alginat dari rumput laut Sargassum sp. yang dilarutkan dalam bufer pH optimum dan diinkubasi pada suhu optimum selama 30 menit.
Kestabilan Enzim pada Ion Logam dan Bahan Aditif. Kestabilan enzim pada bahan aditif yang diberikan antara lain yaitu ion logam KCl, NaCl (monovalen), CaCl2.2H2O, MgCl2. 6 H2O, ZnCl2 (divalen), FeCl3 (trivalen),
senyawa pengkelat logam EDTA yang ditambahkan sebanyak 5 mM dan 10 mM (Jung et al. 2008). Campuran enzim dengan ion logam diinkubasi pada pH dan suhu optimum enzim.
HASIL
Identifikasi Isolat PMP 0126y
Isolat PMP 0126y merupakan isolat koleksi BBP4BKP yang diisolasi dari limbah hasil pengolahan rumput laut Glacilaria sp. menjadi agar-agar di daerah Pemeungpeuk, Jawa Barat. Isolat PMP 0126y dapat tumbuh baik pada media agar-agar nutrien dan tergolong pada bakteri mesofilik karena tumbuh pada suhu 37 0C.
Isolat PMP 0126y diidentifikasi secara langsung dengan melihat morfologi koloni bakteri (Gambar 7). Ciri morfologi yang dimiliki oleh isolat PMP 0126y yaitu warna koloni kuning jingga, bundar, mengkilat. Berdasarkan hasil pewarnaan Gram menggunakan mikroskop (Olympus DP12) dengan perbesaran 1000 x, isolat PMP 0126y tergolong dalam bakteri Gram negatif berbentuk batang pendek (Gambar 8).
Gambar 7 Isolat PMP 0126y. Gambar 8 Pewarnaan Gram isolat PMP 0126y dengan
perbesaran 1000 x.
Gambar 9 Hasil amplifikasi dari gen penyandi 16S-rRNA isolat PMP 0126y.
1 GAGAGCGGCG TACGGGTGCG GAACACGTGT GCAACCTGCC TTTATCTGGG
51 GGATAGCCTT TCGAAAGGAA GATTAATACC CCATAATATA TTGAATGGCA
101 TCATTTGATA TTGAAAACTC CGGTGGATAG AGATGGGCAC GCGCAAGATT
151 AGATAGTTGG TGAGGTAACG GCTCACCAAG TCAGCGATCT TTAGGGGGCC
201 TGAGAGGGTG ATCCCCCACA YTGGTAMTTG AGACAMGGRC CCAGAMTYCT
251 TACGGGAGGG CAGCCAGTGA AGGAATATTT GGACAATGGG GTGAGAGCCT
301 TGATCCCAGC CATCCCGGCG TGAAAGGACG ACGGCCCTTA TGGGTTGTAA
351 ACTTYTTTTT GTATAGGGGA TAAACCTACC CTCGTGAGGG TAGCTGAAGG
401 TACTATACGA ATAAGCACCG GCTAACTCCG TGCCAGCAGC CGCGGTAATA
451 CGGAGGGTGC AAGCGTTATC CGGATTTATT GGGTTTAAAG GGTCCGTAGG
501 CTGATTTGTA AGTCAGTGGT GAAATCTCAC AGCTTAACTG TGAAACTGCC
551 ATTGATACTG CAAGTCTTGA GTGTTGTTGA AGTAGCTGGA ATAAGTAGTG
601 TAGCGGTGAA ATGCATAGAT ATTACTTAGA ACACCAATTG CGAAGGCAGG
651 TTACTAAGCA ACAACTGACG CTGATGGACG AAAGCGTGGG GAGCGAACAG
701 GATTAGATAC CCTGGTAGTC CACGCCGTAA ACGATGCTAA CTCGTTTTTG
751 GGCTTTTGGG TTCAGAGACT AAGCGAAAGT GATAAGTTAG CCACCTGGGG
801 AGTACGAACG CAAGTTTGAA ACTCAAAGGA ATTGACGGGG GCCCGCACAA
851 GCGGTGGATT ATGTGGTTTA ATTCGATGAT ACGCGAGGAA CCTTACCAAG
901 GCTTAAATGG GGAAATGACA GGCTTAGAAA ATAGGCTTTT CTTCGGACAT
951 TTTTCAAGGT GCTGCATGGT TGTCGTCAGC TCSTGCCCGT GAGGTGTTAA
1001 GGTTAAGTCC TTGCAACGAA GCGCAACCCC TTGTCACTAR TTTGCCATCA
1051 TTTAAKTTGG GGGACTCTAG TKARAACTGC CTACSCCAAG TARARARGAA
1101 AAGKTGGGGA TRAMGTCAAA TCATCACGGC CCTTACGCCT TGGGCCACAC
1151 ACGTAATACA ATGGCCGGTA CAGAGGGCAG CTACACTGCG AAGTGATGCA
1201 AATCTCGAAA GCCGGTCTCA GTTCGGATTG GAGTCTGCAA CTCGACTCTA
1251 TGAAGCTGGA ATCGCTAGTA ATCGCGCATC AG
Gambar 10 Sebagian sekuen DNA penyandi 16S-rRNA isolat PMP 0126y dari
(arah 5’-3’).
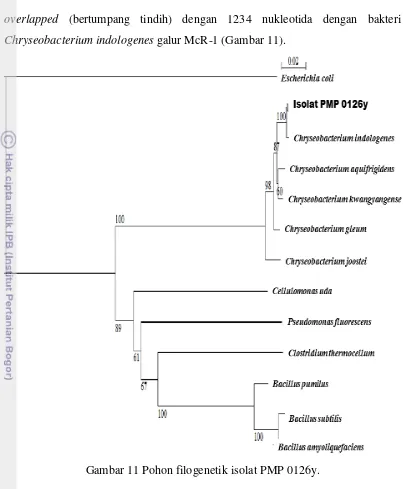
Sekuen komplemen DNA penyandi 16S-rRNA isolat PMP 0126y dianalisis dengan program FASTA dari koleksi Genebank National Center Biotechnology Information (NCBI). Berdasarkan analisis sekuen DNA tersebut, isolat PMP 0126y memiliki kemiripan sebesar 96% dari 1282 nukleotida yang
M + - PMP 0126Y
overlapped (bertumpang tindih) dengan 1234 nukleotida dengan bakteri Chryseobacterium indologenes galur McR-1 (Gambar 11).
Gambar 11 Pohon filogenetik isolat PMP 0126y.
Pertumbuhan dan Produksi Enzim Selulase
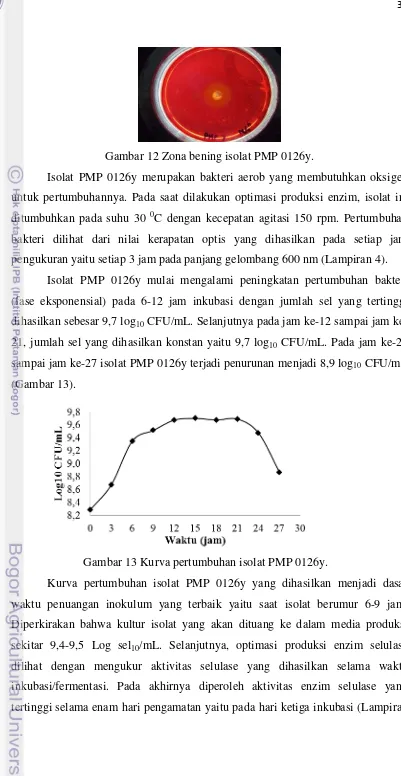
Gambar 12 Zona bening isolat PMP 0126y.
Isolat PMP 0126y merupakan bakteri aerob yang membutuhkan oksigen untuk pertumbuhannya. Pada saat dilakukan optimasi produksi enzim, isolat ini ditumbuhkan pada suhu 30 0C dengan kecepatan agitasi 150 rpm. Pertumbuhan bakteri dilihat dari nilai kerapatan optis yang dihasilkan pada setiap jam pengukuran yaitu setiap 3 jam pada panjang gelombang 600 nm (Lampiran 4).
Isolat PMP 0126y mulai mengalami peningkatan pertumbuhan bakteri (fase eksponensial) pada 6-12 jam inkubasi dengan jumlah sel yang tertinggi dihasilkan sebesar 9,7 log10 CFU/mL. Selanjutnya pada jam 12 sampai jam
ke-21, jumlah sel yang dihasilkan konstan yaitu 9,7 log10 CFU/mL. Pada jam ke-24
sampai jam ke-27 isolat PMP 0126y terjadi penurunan menjadi 8,9 log10 CFU/mL
(Gambar 13).
Gambar 13 Kurva pertumbuhan isolat PMP 0126y.
Kurva pertumbuhan isolat PMP 0126y yang dihasilkan menjadi dasar waktu penuangan inokulum yang terbaik yaitu saat isolat berumur 6-9 jam. Diperkirakan bahwa kultur isolat yang akan dituang ke dalam media produksi sekitar 9,4-9,5 Log sel10/mL. Selanjutnya, optimasi produksi enzim selulase
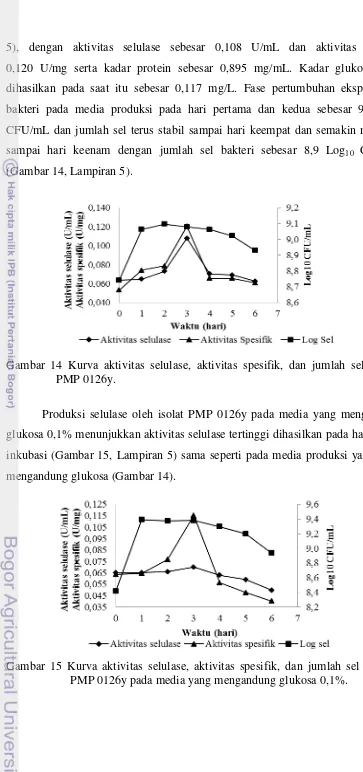
5), dengan aktivitas selulase sebesar 0,108 U/mL dan aktivitas spesifik 0,120 U/mg serta kadar protein sebesar 0,895 mg/mL. Kadar glukosa yang dihasilkan pada saat itu sebesar 0,117 mg/L. Fase pertumbuhan eksponensial bakteri pada media produksi pada hari pertama dan kedua sebesar 9,1 log10
CFU/mL dan jumlah sel terus stabil sampai hari keempat dan semakin menurun sampai hari keenam dengan jumlah sel bakteri sebesar 8,9 Log10 CFU/mL
(Gambar 14, Lampiran 5).
Gambar 14 Kurva aktivitas selulase, aktivitas spesifik, dan jumlah sel bakteri PMP 0126y.
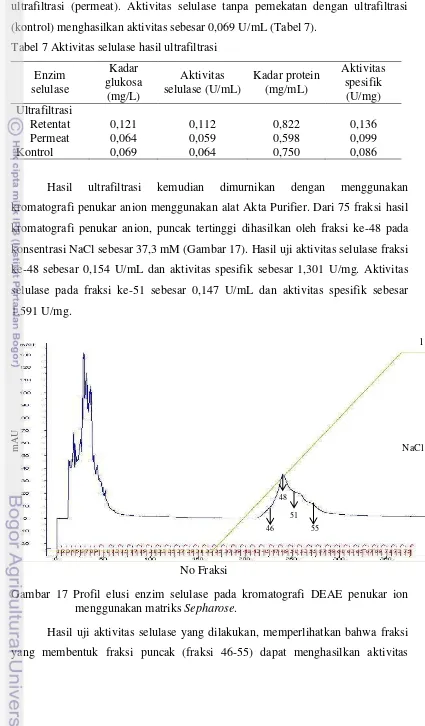
Produksi selulase oleh isolat PMP 0126y pada media yang mengandung glukosa 0,1% menunjukkan aktivitas selulase tertinggi dihasilkan pada hari ketiga inkubasi (Gambar 15, Lampiran 5) sama seperti pada media produksi yang tidak mengandung glukosa (Gambar 14).
Penambahan glukosa sebanyak 0,1% pada media produksi menghasilkan jumlah sel sebesar 9,4 log10 CFU/mL, sedangkan jumlah sel tertinggi pada media
produksi tanpa glukosa hanya 9,1 log10 CFU/mL. Aktivitas selulase yang tertinggi
pada hari ketiga sebesar 0,070 U/mL dan aktivitas spesifik 0,116 U/mg dengan kadar protein sebesar 0,606 mg/mL pada media produksi yang mengandung glukosa 0,1%.
Pemurnian Enzim Selulase
Enzim selulase diproduksi selama 3 hari yang menunjukkan waktu produksi tertinggi, kemudian dimurnikan dengan melakukan pemekatan enzim melalui dua cara pemekatan yaitu pengendapan amonium sulfat dan ultrafiltrasi. Persen kadar amonium sulfat yang menghasilkan aktivitas selulase tertinggi diperoleh pada 50% kejenuhan amonium sulfat. Aktivitas selulase yang dihasilkan sebesar 0,072 U/mL dan aktivitas spesifik sebesar 0,128 U/mg pada endapan, sedangkan pada supernatan dihasilkan aktivitas selulase sebesar 0,068 U/mL dan aktivitas spesifik sebesar 0,105 U/mg. Selulase tanpa penambahan amonium sulfat (kontrol) memiliki aktivitas sebesar 0,064 U/mL dan aktivitas spesifik 0,075 U/mg (Gambar 16, Lampiran 5).
Gambar 16 Aktivitas spesifik dari pengendapan selulase dengan amonium sulfat.
ultrafiltrasi (permeat). Aktivitas selulase tanpa pemekatan dengan ultrafiltrasi (kontrol) menghasilkan aktivitas sebesar 0,069 U/mL (Tabel 7).
Tabel 7 Aktivitas selulase hasil ultrafiltrasi
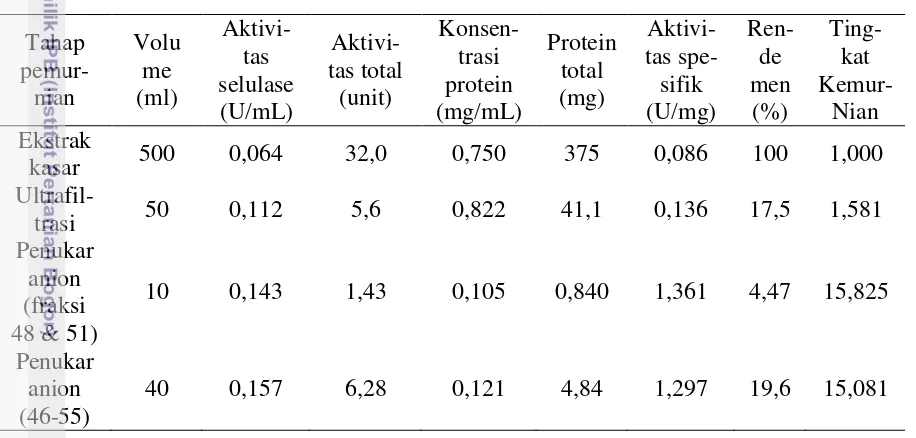
Enzim
Hasil ultrafiltrasi kemudian dimurnikan dengan menggunakan kromatografi penukar anion menggunakan alat Akta Purifier. Dari 75 fraksi hasil kromatografi penukar anion, puncak tertinggi dihasilkan oleh fraksi ke-48 pada konsentrasi NaCl sebesar 37,3 mM (Gambar 17). Hasil uji aktivitas selulase fraksi ke-48 sebesar 0,154 U/mL dan aktivitas spesifik sebesar 1,301 U/mg. Aktivitas
Hasil uji aktivitas selulase yang dilakukan, memperlihatkan bahwa fraksi yang membentuk fraksi puncak (fraksi 46-55) dapat menghasilkan aktivitas
selulase (Lampiran 5). Selulase dari fraksi ke-48 dan 51 digabung dan diukur aktivitasnya sebesar 0,143 U/mL dengan aktivitas spesifik sebesar 1,361 U/mg. Gabungan fraksi ke-46 sampai fraksi ke-55 (tanpa fraksi 48 & 51) menghasilkan aktivitas selulase sebesar 0,157 U/ml dengan aktivitas spesifik sebesar 1,297 U/mg.
Hasil pengukuran tingkat kemurnian enzim selulase yang dihasilkan oleh ultrafiltrasi sebesar 1,58 kali dibandingkan ekstrak kasar, dan tingkat kemurnian enzim hasil penukar anion (fraksi 48 & 51, fraksi 46-55) secara berturut-turut sebesar 15,82 dan 15,08 kali (Tabel 8).
Tabel 8 Hasil uji aktivitas selulase PMP 0126y pada beberapa tahap pemurnian
Tahap
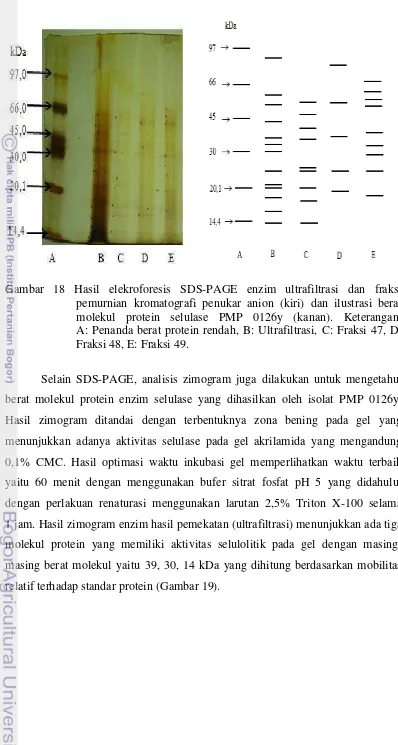
Analisis Berat Molekul Enzim Selulase Menggunakan SDS-PAGE dan Zimogram
Gambar 18 Hasil elekroforesis SDS-PAGE enzim ultrafiltrasi dan fraksi pemurnian kromatografi penukar anion (kiri) dan ilustrasi berat
molekul protein selulase PMP 0126y (kanan). Keterangan: A: Penanda berat protein rendah, B: Ultrafiltrasi, C: Fraksi 47, D:
Fraksi 48, E: Fraksi 49.