PENANDA RAPD (Random Amplified Polymorphic
DNA)
SKRIPSI
OLEH
YANTIKA ROMAULI SIMATUPANG
100805072
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS SUMATERA UTARA
ANALISIS KEANEKARAGAMAN GENETIK MARKISA
(Passiflora
spp.
) DI SUMATERA UTARA BERDASARKAN
PENANDA RAPD (Random Amplified Polymorphic
DNA)
SKRIPSI
Diajukan untuk melengkapi tugas dan memenuhi syarat mencapai gelar Sarjana Sains
OLEH
YANTIKA ROMAULI SIMATUPANG
100805072
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS SUMATERA UTARA
i
Judul : Analisis Keanekaragaman Genetik Markisa (Passifloraspp.) Di Sumatera Utara Berdasarkan Penanda RAPD (Random Amplified PolymorphicDNA)
Kategori : Skripsi
Nama : Yantika Romauli Simatupang Program Studi : Sarjana (S1) Biologi
Nomor Induk Mahasiswa : 100805072
Fakultas : Matematika dan ilmu Pengetahuan Alam Universitas Sumatera Utara
Disetujui di Medan, Mei 2015
Komisi Pembimbing:
Pembimbing 2, Pembimbing 1,
Dr. Suci Rahayu, M.Si. Dr. Saleha Hannum, M.Si.
NIP: 196506291992032002 NIP:197108312000122001
Disetujui Oleh
Departemen Biologi FMIPA USU Ketua
ii
PERNYATAAN
ANALISIS KEANEKARAGAMAN GENETIK MARKISA (Passiflora
spp.
) DI SUMATERA UTARA BERDASARKAN PENANDA RAPD
(Random Amplified Polymorphic
DNA)
SKRIPSI
Saya mengakui bahwa skripsi ini adalah hasil karya sendiri. Kecuali beberapa kutipan dan ringkasan yang masing-masing disebutkan sumbernya.
Medan, Mei 2015
iii
Puji dan syukur penulis ucapkan ke hadirat Tuhan Yang Maha Esa atas segala berkat dan penyertaanNya sehingga penulis dapat menyelesaikan penulisan skripsi ini yang berjudul “Analisis Keanekaragaman Genetik Markisa (Passiflora
spp.) Di Sumatera Utara Berdasarkan Penanda RAPD (Random Amplified
Polymorphic DNA)” sebagai syarat untuk mencapai gelar sarjana sains di
Departemen Biologi FMIPA USU Medan.
Ucapan terimakasih terbesar penulis sampaikan kepada kedua orang tua penulis, ayahanda dan Alm. ibunda tercinta, Syarifuddin Simatupang dan Alm. Remida Silalahi, S.Pd yang selalu memberikan do’a, dukungan, semangat,
pengorbanan serta kasih sayangnya yang besar kepada penulis. Kepada seluruh keluarga besarku atas segala bantuan yang diberikan kepada penulis baik moril maupun materil.
Terimakasih penulis sampaikan kepada ibu Dr. Saleha Hannum, M.Si. selaku pembimbing 1 dan ibu Dr. Suci Rahayu, M.Si. selaku pembimbing 2 yang telah memberikan bimbingan, saran, motivasi dan waktu selama penyusunan skripsi ini. Ucapan terimakasih juga penulis sampaikan kepada ibu Dra. Elimasni, M.Si. dan bapak Dr. Salomo Hutahaean, M.Si. selaku dosen penguji yang telah memberikan banyak saran demi kesempurnaan penulisan skripsi ini. Tidak lupa juga penulis ucapkan terimakasih kepada ibu Dr. Nursahara Pasaribu M.Sc. selaku ketua Departemen Biologi serta semua staf yang bekerja di Departemen Biologi.
Dalam kesempatan ini penulis juga menyampaikan ucapan terimakasih kepada rekan mahasiswa/i Biologi. Kepada ‘ma gizibe’ AnitaDoris, Santa Lusia, Chrestina Sidabutar yang selalu menemani dan menerima kegilaanku. Kepada sahabatku Norton, Edwardman, Doni, Yuli, Nurhayati, Anisa Rilla, Nurfithri apriani, Nova, Maria, Bobby Hutabarat, Ilham buat waktu dan tamparan kalimat yang membuat penulis sadar dan termotivasi. Terimakasih kepada motivator dan guruku bang Imam Aulia dan kak ai atas waktu dan ilmu praktek serta teori yang diberikan. Tak terlupakan teman-teman stambuk 2010 ‘BioRev’ yang namanya
tak bisa disebutkan satu persatu, terimakasih sudah membuatku dewasa dan berjiwa sosial. Kepada rekan asisten genetika Vahnoni, Sandi, bang Tombak, Jordan, Nasir, Wilda, Nolo dan lainnya. Semuanya yang tidak dapat penulis tuliskan satu persatu, terimakasih atas kerjasamanya selama di bangku perkuliahan.
Akhirnya dengan segala kerendahan hati, penulis menyadari bahwa skripsi ini masih jauh dari kesempurna. Oleh karena itu, penulis mengharapkan kritik dan saran yang bersifat membangun demi kesempurnaan penulisan skripsi ini. Sebelum dan sesudahnya penulis mengucapkan terima kasih dan semoga Tuhan YME membalas semua kebaikan dengan balasan yang setimpal.
Medan, Mei 2015
iv
ANALISIS KEANEKARAGAMAN GENETIK MARKISA (Passifloraspp.)
DI SUMATERA UTARA BERDASARKAN PENANDA RAPD
(Random Amplified PolymorphicDNA)
ABSTRAK
Penelitian keanekaragaman genetik markisa (Passiflora spp.) di Sumatera Utara berdasarkan penanda RAPD telah dilakukan di Laboratorium Genetika dan Laboratorium Terpadu, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara. Tujuan dari penelitian ini adalah untuk mengkarakteristik keanekaragaman genetik markisa (Passiflora sp.) berdasarkan penanda RAPD. Analisis RAPD dilakukan terhadap 31 aksesi markisa yang dikoleksi dari 4 kabupaten yaitu kabupaten HUMBAHAS, Tapanuli Utara, Simalungun, dan Karo yang mewakili 4 spesies (P. edulis, P. ligularis, P. quadrangularis, P. foetida) dari genus Passiflora dengan menggunakan 4 primer acak (diseleksi dari 7 primer acak). Hasil elektroforesis menunjukkan 73 pita polimorfik (98%) yang berukuran dari 250 bp sampai 2500 bp. Koefisien kemiripan genetik berkisar antara 0,80 sampai 0,97 berdasarkan analisis data biner dari pita RAPD dengan metode UPGMA (Unweight Pair- Group Method Arithmatic) melalui program NTSYS (Numerical Taxonomy and Multivariate System) versi 2.11a. Hasil ini mengungkapkan bahwa tingkat variasi dalam dan antara spesies tinggi. Analisis cluster menunjukkan bahwa pengelompokan markisa terjadi berdasarkan wilayah pengkoleksian sampelnya Kedekatan jarak geografis wilayah pengkoleksian dan kesamaan jenis markisa berdasarkan warna buah tidak menjamin kedekatan hubungan genetik markisa berdasarkan penanda RAPD.
v
ABSTRACT
The research about Genetic diversity analysis of the passion fruit (Passifloraspp.) in north Sumatera based on RAPD markers has been done in Genetic Laboratory and Integrated Laboratory, University of North Sumatera. The objective of this study was to characterize the genetic diversity of Passiflora sp. collection by RAPD marker. RAPD analysis was done to 31 passion fruit (Passiflora sp.) accessions that collected from 4 regional is region HUMBAHAS, North Tapanuli, Simalungun, and Karo that represented 4 species (P. edulis, P. ligularis, P. quadrangularis, P. foetida) from genus of Passiflora using 4 random primers (selected from 7 random primers). The result of electrophoresis showed 73 reproducible polymorphic bands (98%) are ranging in size from 250 bp to 2500 bp. The genetic similarity showed coefficients ranged from 0,8 to 0,97 based on Unweight Pair- Group Method Aritchmatic and Numerical Taxonomy (UPGMA) and Multivariate System (NTSYS) version 2.11a computer program,. The result revealed high levels of variation within and among species. Cluster analysis showed that grouping of passion fruit occurs by area collecting of the sample. Geographical proximity and similarity of the area where passion fruit had been collecting and based on the color of the fruit do not guarantee the genetic relationship of passion fruit by RAPD.
vi 2.2. Morfologi dan Syarat Tumbuh Markisa 6 2.2.1. Morfologi Markisa 6
3.3.3. Uji Kualitas dan Kuantitas DNA 13 3.3.4. Amplifikasi DNA 13
3.4. Analisis Data 15
vii
4.2. Analisis Profil Pita RAPD 17 4.3. Analisis Keanekaragaman Genetik Tanaman Markisa 21
4.3.1. Analisis Hubungan Genetik Markisa Ungu (P. edulis)
21
4.3.2. Analisis Hubungan Genetik Markisa (Passifloraspp.)
23
BAB 5. Kesimpulan dan Saran
5.1. Kesimpulan 27
5.2. Saran 27
Daftar Pustaka 28
viii
DAFTAR TABEL
No. Tabel Judul Halaman
ix
No. Gambar Judul Halaman
3.1. Pola Terjemahan Pita DNA 15 4.1. Hasil elektoforesis DNA genom dari 9 sampel
markisa menggunakan gel agarose 0,8%
16
4.2. Profil Pita RAPD pada primer Akansha 19 4.3. Dendogram kemiripan genetik markisa ungu
(Passiflora edulis) berdasarkan 4 primer RAPD
22
4.4. Dendogram kemiripan genetik markisa (Passiflora
spp.) berdasarkan 4 primer RAPD
x
DAFTAR LAMPIRAN
No. Lampiran Judul Halaman
Lampiran 1.ole Koleksi 31 Aksesi Tanaman Markisa di Sumatera Utara 32 Lampiran 2. Hasil Pengukuran Kuantitas DNA dengan
Menggunakan Nanophotometer
33
Lampiran 3 Gambar Hasil Amplifikasi DNA Markisa berdasarkan Teknik RAPD
34
iv
ABSTRAK
Penelitian keanekaragaman genetik markisa (Passiflora spp.) di Sumatera Utara berdasarkan penanda RAPD telah dilakukan di Laboratorium Genetika dan Laboratorium Terpadu, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara. Tujuan dari penelitian ini adalah untuk mengkarakteristik keanekaragaman genetik markisa (Passiflora sp.) berdasarkan penanda RAPD. Analisis RAPD dilakukan terhadap 31 aksesi markisa yang dikoleksi dari 4 kabupaten yaitu kabupaten HUMBAHAS, Tapanuli Utara, Simalungun, dan Karo yang mewakili 4 spesies (P. edulis, P. ligularis, P. quadrangularis, P. foetida) dari genus Passiflora dengan menggunakan 4 primer acak (diseleksi dari 7 primer acak). Hasil elektroforesis menunjukkan 73 pita polimorfik (98%) yang berukuran dari 250 bp sampai 2500 bp. Koefisien kemiripan genetik berkisar antara 0,80 sampai 0,97 berdasarkan analisis data biner dari pita RAPD dengan metode UPGMA (Unweight Pair- Group Method Arithmatic) melalui program NTSYS (Numerical Taxonomy and Multivariate System) versi 2.11a. Hasil ini mengungkapkan bahwa tingkat variasi dalam dan antara spesies tinggi. Analisis cluster menunjukkan bahwa pengelompokan markisa terjadi berdasarkan wilayah pengkoleksian sampelnya Kedekatan jarak geografis wilayah pengkoleksian dan kesamaan jenis markisa berdasarkan warna buah tidak menjamin kedekatan hubungan genetik markisa berdasarkan penanda RAPD.
v
GENETIC DIVERSITY ANALYSIS OF THE PASSION FRUIT (Passifloraspp.)IN NORTH SUMATERA BASED ON RAPD MARKERS
(Random Amplified PolymorphicDNA)
ABSTRACT
The research about Genetic diversity analysis of the passion fruit (Passifloraspp.) in north Sumatera based on RAPD markers has been done in Genetic Laboratory and Integrated Laboratory, University of North Sumatera. The objective of this study was to characterize the genetic diversity of Passiflora sp. collection by RAPD marker. RAPD analysis was done to 31 passion fruit (Passiflora sp.) accessions that collected from 4 regional is region HUMBAHAS, North Tapanuli, Simalungun, and Karo that represented 4 species (P. edulis, P. ligularis, P. quadrangularis, P. foetida) from genus of Passiflora using 4 random primers (selected from 7 random primers). The result of electrophoresis showed 73 reproducible polymorphic bands (98%) are ranging in size from 250 bp to 2500 bp. The genetic similarity showed coefficients ranged from 0,8 to 0,97 based on Unweight Pair- Group Method Aritchmatic and Numerical Taxonomy (UPGMA) and Multivariate System (NTSYS) version 2.11a computer program,. The result revealed high levels of variation within and among species. Cluster analysis showed that grouping of passion fruit occurs by area collecting of the sample. Geographical proximity and similarity of the area where passion fruit had been collecting and based on the color of the fruit do not guarantee the genetic relationship of passion fruit by RAPD.
BAB 1
PENDAHULUAN
1.1. Latar Belakang
Markisa (Passiflora sp.) merupakan salah satu jenis buah hortikultura yang
berpotensi besar dalam perdagangan buah di pasar dunia. Buah dari genus
Passiflora ini memiliki hal-hal yang menarik seperti memiliki bentuk dan warna
yang eksotis dan memiliki aroma yang khas (Ashari, 1995), selain itu memiliki
penarik di bidang farmatik seperti sebagai obat penenang, antipasmodik dan
antibakterial serta insektisida (Fajardoet al., 1998). Markisa merupakan tanaman
penting baik secara ekonomi maupun sosial, dan telah banyak dilakukan
penelitian untuk mengembangkan varietas yang adaptif terhadap sistem tanam dan
kondisi iklim yang berbeda (Nasutionet al.,2011).
Tanaman markisa bukanlah tanaman asli Indonesia, tetapi merupakan
tanaman yang berasal dari Amerika Selatan yaitu Brazil, yang menyebar sampai
ke Indonesia. Di negara asalnya markisa tumbuh liar di hutan-hutan basah yang
mempunyai ratusan spesies. Di Indonesia, markisa mulai dibudidayakan sejak
tahun 2003, karena sebelum tahun 2003 keberadaan dan manfaatnya belum bagitu
disadari masyarakat Indonesia. Ada beberapa provinsi di Indonesia yang
membudidayakan markisa yaitu Sumatera Utara, Sumatera Barat, Lampung dan
Sulawesi Selatan, namun yang menjadi sentra terbesar produksi markisa adalah
Sumatera Utara dan Sulawesi Selatan (BPPHP, 2004).
Jenis markisa yang dikembangkan di Sumatera Utara pada umumnya
adalah markisa ungu (P. edulis) yang akan diolah menjadi minuman berupa jus
ataupun sirup, namun ada juga markisa yang dibudidayakan untuk konsumsi
langsung yang sering juga disebut buah meja yaitu markisa konyal (P. ligularis)
(Karmila, 2013). Di Indonesia ada 3 jenis markisa yang telah dibudidayakan,
meliputi markisa asam dengan kulit buah berwarna ungu (Passiflora edulis f.
edulis Sims.), markisa asam dengan kulit buah kuning (P. edulis f. flavicarva
2
ada 4 varietas unggul markisa yang sudah dilepas yaitu markisa asam varietas
Malino dari Sulawesi Selatan tahun 2000, markisa asam varietas Berastagi tahun
2000, markisa manis varietas Super Solinda dan Gumanti tahun 2001 yang
merupakan hasil seleksi indigenos, sedangkan kegiatan pemuliaan tanaman
markisa untuk mendapatkan varietas unggul belum banyak dilakukan
(Karsinahet al.,2007).
Ketersediaan varietas unggul merupakan syarat yang harus dipenuhi pada
era industrialis pertaniaan dan liberalis perdagangan. Varietas unggul dapat dirakit
jika tersedia plasma nutfah atau sumberdaya genetik yang mempunyai karakter
sesuai dengan yang dikehendaki (Karsinah et al.,2007). Salah satu kendala dalam
perbaikan mutu tanaman adalah terbatasnya koleksi plasma nutfah dan informasi
genetik tanaman (Langga et al., 2012), oleh karena itu diperlukan
langkah-langkah dalam upaya penyediaan materi genetik dalam memperbaiki tanaman
dengan cara eksplorasi, konservasi dan evaluasi (Karsinahet al.,2007). Salah satu
usaha yang dapat dilakukan untuk memperbaiki mutu markisa adalah dengan
menganalisis sumberdaya genetik markisa secara molekuler (Langgaet al.,2012).
Informasi berdasarkan analisis molekuler diharapkan mampu menjawab
permasalahan dalam karakterisasi tanaman markisa yang ada di Indonesia.
Berdasarkan hal tersebut maka penelitian genetik markisa dengan menggunakan
penanda molekuler perlu dilakukan. Penanda molekuler yang dapat digunakan
adalah penanda DNA yang berupa fragmen/pita-pita DNA. Penelitian penanda
DNA lebih akurat karena tidak dipengaruhi lingkungan (Yasminingsih, 2009).
Penanda Random amplified polymorphic DNA (RAPD) merupakan salah
satu penanda DNA berbasis PCR yang banyak digunakan dalam mengidentifikasi
keanekaragaman pada tingkat intraspesies maupun antarspesies
(Fajardo et al., 1998). Teknik ini mendeteksi polimorfisme ruas nukleotida pada
DNA dengan menggunakan sebuah primer tunggal yang memiliki rangkaian
nukleotida acak. Marka RAPD bersifat lebih sederhana dibandingkan marka
lainnya seperti mikrosatelit atau simple sequence repeat (SSR), restriction length
polymorphism (RLFP), ataupun amplified length polymorphism (AFLP)
(Bardakchi, 2001). Hal ini dikarenakan teknik RAPD tidak memerlukan informasi
memerlukan probe DNA yang spesifik.
Studi keanekaragaman genetik dari genusPassifloradengan menggunakan
penanda RAPD telah dilakukan (Fajardo et al., 1998; Crochemore et al., 2003;
Viana et al., 2003).Di Indonesia, studi keanekaragaman genetik Passiflora telah
dilakukan dengan menggunakan penanda ISSR (Nasution et al., 2011) yang
meneliti 13 aksesi markisa yang berasal dari kebun petani markisa yang ada di
Sulawesi Selatan. Studi keanekaragaman genetik Passiflora yang terdapat di
Sumatera Utara dengan menggunakan penanda RAPD sampai saat ini belum ada
informasi, oleh karena itu penelitian ini perlu dilakukan untuk mengetahui
keanekaragaman genetik markisa yang terdapat di Sumatera Utara.
1.2. Permasalahan
Rumusan masalah dalam penelitian ini adalah bagaimana keanekaragaman
genetik markisa (Passiflora sp.) yang terdapat di Sumatera Utara berdasarkan
penanda molekular RAPD?
1.3. Tujuan Penelitian
Tujuan yang akan dicapai dalam penelitian ini adalah untuk mengetahui
keanekaragaman genetik markisa (Passiflorasp.) yang terdapat di Sumatera Utara
dengan menggunakan analisis RAPD dan mengelompokkannya berdasarkan pola
pita yang tampak pada analisis RAPD.
1.4. Hipotesis
Hipotesis penelitian ini adalah tingginya keanekaragaman genetik markisa
yang terdapat dari Sumatera Utara berdasarkan analisis RAPD.
1.5. Manfaat Penelitian
Penelitian ini diharapkan dapat menghasilkan data molekuler markisa yang
terdapat dari Sumatera Utara dengan menggunakan analisis RAPD yang
4
BAB 2
TINJAUAN PUSTAKA
2.1. Klasifikasi Markisa
Markisa mula-mula disebut passion fruit. Menurut sejarah, tanaman markisa
berasal dari daerah tropis Amerika Selatan, tepatnya di daerah Brazil, Venezuela,
Kolumbia, dan Peru. Di Indonesia, markisa banyak ditanam di dataran tinggi di
Goa, Malino, Sulawesi Selatan dan Sumatera Utara (Sunarjono, 1997). Buah
markisa yang pertama kali dikenal ditempat asalnya adalah markisa kuning dan
markisa ungu (Rukmana, 2003).
Klasifikasi botani tanaman markisa adalah sebagai berikut (Sharma, 1993):
Kingdom :Plantae
Divisio :Spermatophyte
Sub divisi :Angiospermae
Kelas :Dicotiledoneae
Ordo :Paretales
Famili :Passifloraceae
Genus :Passiflora
Spesies :Passiflorasp.
Di Indonesia terdapat empat jenis markisa yang dibudidayakan, yaitu
markisa ungu (Passiflora edulis),markisa kuning (Passiflora edulis f. flavicarpa),
markisa erbis (Passiflora quadrangularis), dan markisa konyal (Passiflora
ligularis).
2.1.1. Markisa Asam (Siuh)
Markisa asam (Passiflora edulis Sims.) mempunyai nama umum granadillaatau.
Passion fruit yang termasuk dalam famili Passifloraceae. Diperkirakan ada 500
spesies Passiflora dalam famili ini, diantara ciri-ciri tersebut P. edulis Sims.
memiliki ciri-ciri spesifik markisa. Menurut LITBANG (2010), dalam spesies ini
a. Forma edulis atau forma ungu dikenal dengan markisa ungu. Yang termasuk
dalam forma ini adalah markisa asam dengan kulit buah berwarna ungu (purple),
merah (red), dan hitam (black granadilla) disebut juga siuh atau purple passion
fruit (P. edulis f. edulis Sims.). Markisa ini merupakan bahan baku utama industri
pengolahan sari buah markisa dan sirup konsentrat.
b. Forma flavicarpa atau forma kuning dikenal dengan markisa kuning, yaitu
markisa asam dengan kulit buah berwarna kuning disebut juga rola atau yellow
passion fruit (P. edulisSimsf. flavicarpaDeg.).
Markisa ungu berasal dari Brazil bagian Selatan yaitu Paraguay hingga
Argentina bagian Utara, sedangkan asal markisa kuning tidak diketahui mungkin
berasal dari Amazon wilayah Brazil, hibrid antara P. edulis dan P. ligularis,
namun studi secara sitologi tidak menunjukkan teori hibrid. Pendapat lain
menyatakan bahwa markisa kuning berasal dari Australia. Pada tahun 1930-1950
markisa ungu dan kuning telah menyebar ke berbagai negara di dunia. Di sebagian
besar negara-negara penghasil markisa, kultivar-kultivar markisa kuning yang
umumnya dibudidayakan. Di Indonesia markisa asam yang telah dibudidayakan
secara komersial adalah markisa ungu. (LITBANG, 2010).
2.1.2. Markisa Manis (Konyal)
Selain markisa asam, di Indonesia juga terdapat markisa manis yang sering
disebut pula dengan markisa konyal (Passiflora ligularis). Markisa konyal
merupakan spesies umum yang berasal dari Brazil dan menyebar ke daerah tropis
dan subtropis, termasuk Indonesia. Markisa konyal banyak dijualbelikan sebagai
buah segar di tempat-tempat tertentu karena rasanya manis walaupun aromanya
relatif tidak ada. Sentra produksi markisa ini adalah di Kabupaten Solok,
Sumatera Barat dengan lahan 2.710 ha dan produksi sebanyak l2.710 ton pada
tahun 1997 (Karmila, 2013).
2.1.3. MarkisaGiant(Erbis)
Di Indonesia terdapat markisa dengan ukuran besar yang disebut juga
dengan erbis (Passiflora quadrangularis L.) atau markisa sayur. Buah erbis ini
6
dan buahnya hanya untuk sari buah segar (Sunarjono, 1997).
2.2. Morfologi dan Syarat Tumbuh Markisa
2.2.1. Morfologi Markisa
Markisa merupakan tumbuhan semak atau pohon yang hidup menahun (perennial)
dan bersifat merambat atau menjalar hingga sepanjang 20 meter atau lebih.
Batang tanaman berkayu tipis, bersulur dan memiliki banyak percabangan yang
kadang-kadang tumbuh timpang tindih, Pada stadium muda, cabang tanaman
berwama hijau dan setelah tua berubah menjadi hijau kecoklatan. Daur, tanaman
sangat rimbun, tumbuh secara bergantian pada batang atau cabang. Tiap helai
daun bercaping tiga dan bergerigi, berwarna hijau mengkilap (Rukmana, 2003).
Semua jenis markisa termasuk penyerbuk silang melalui lebah madu. Meskipun
demikian, penyerbukan sendiri masih dapat berlangsung baik (Sunarjono, 1997).
Tanaman markisa yang berasal dari buah mulai berbuah setelah umur 9-10
bulan, sedangkan yang berasal dari stek, mulai berbuah dari awal, yaitu sekitar 7
bulan. Buah markisa digolongkan kedalam buah klimaterik karena pola respirasi
markisa meningkat seiring dengan perubahan pigmen warna dan gas volatile
tertentu. Respirasi dan produk etilen akan menurun saat buah mencapai tingkat
kematangan penuh dan mulai mengalami pembusukan (Karmila, 2013).
2.2.2. Syarat Tumbuh Markisa
Menurut Danarto et al. (2012), untuk dapat tumbuh dan berproduktivitas baik,
tanaman markisa mempunyai kekhususan persyaratan tumbuh. Faktor yang
terpenting dalam menentukan pertumbuhan markisa adalah sebagai berikut:
a. Lokasi
Tanaman markisa merupakan tanaman subtropis, sehingga jika ditanam di
Indonesia harus di daerah-daerah yang mempunyai ketinggian antara 800-1.500 m
dpl dengan curah hujan minimal 1.200 mm/tahun, kelembaban nisbi antara
80-90%, suhu lingkungan antara 20-30 °C, dan tidak banyak angin.
b. Tanah
Tanaman markisa dapat tumbuh diberbagai jenis tanah, terutama pada yang
berdrainase baik dengan ketinggian tanah antara 600-1.500 m dpl. Jika markisa
ditanam pada ketinggian di bawah 600 dpl maka produktifitas tanaman markisa
akan mengalami penurunan. Hal ini dikarenakan daerah dengan ketinggian
tersebut lama penyinaran matahari lebih rendah, serta suhu lingkungan juga
kurang mendukung sehingga menghambat pembungaan.
2.3. Marka Molekuler
Variasi teknik molekuler sangat beragam tergantung cara pelaksanaan untuk
mendapatkan data maupun tingkatan target data yang diinginkan. Salah satu cara
untuk mendapatkan data yang sering digunakan pada saat ini adalah dengan
menggunakan marka/penanda. Dari sejarah perkembangannya, penanda/marka ini
dapat dikelompokkan sebagai penanda morfologi, penanda sitologi, dan penanda
molekular. Penanda molekular dapat dibedakan menjadi penanda isozim dan
penanda DNA (Nuraida, 2012). Penggunaan penanda molekuler berupa DNA
digunakan seiring dengan perkembangan ilmu pengetahuan tentang biologi
molekuler. Penanda ini telah digunakan sejak tahun 1980-an, merupakan
pendekatan untuk lebih meningkatkan informasi genetik yang belum dapat
diperoleh dengan penanda protein (Langga et al., 2012). Marka molekuler yang
berbasis DNA jenisnya sangat banyak dan mempunyai variabilitas yang besar,
sedangkan marka yang berbasis protein memiliki keragaman yang lebih rendah
(Warta Biogen, 2008). Marka molekuler yang pertama kali dikenal adalah marka
protein yang secara genetik dikenal sebagai marka isozim. (Yasminingsih, 2009).
Menurut Nuraida (2012), suatu penanda (marker) adalah suatu karakter
atau sifat yang dapat diturunkan atau diwariskan pada keturunannya dan dapat
digunakan untuk mengkarakteristik/mendeteksi genotip tertentu. Penanda
molekular merupakan teknik yang efektif dalam analisis genetik dan telah
diaplikasi secara luas dalam program pemuliaan tanaman. Pemanfaatan marka
DNA sebagai alat bantu seleksi Marker Assisted Selection (MAS) lebih
menguntungkan dibandingkan dengan seleksi secara fenotipik. Seleksi dengan
bantuan marka molekular didasarkan pada sifat genetik tanaman saja, tidak
dipengaruhi oleh faktor lingkungan (Azrai, 2005). Pemilihan jenis marka
8
kesesuaiannya dengan fasilitas dan materi yang dimiliki untuk melakukan seleksi.
Penyiapan atau purifikasi gen target juga sangat menentukan keberhasilan dari
seleksi yang dilakukan (Syafaruddin & Santoso, 2011).
Jenis marka molekular pada tanaman ada dua yaitu penanda mendasarkan
teknik PCR dan yang tidak mendasarkan teknik PCR. Penanda molekular yang
mendasarkan teknik PCR antara lain RAPD (Random Amplified Polymorphic
DNA), AFLP (Amplified Fragment Length Polymorphism) dan SSR (Simple
Sequence Repeats) yang lebih mendasarkan pada sequencing DNA. Sedangkan
penanda molekuler yang tidak mendasarkan teknik PCR hanya ada satu jenis yaitu
RLFP (Restriction Fragment Length Polymorphism) (Azrai, 2005). Setiap
penanda molekuler memiliki teknik yang berbeda-beda baik dalam hal jumlah
DNA yang dibutuhkan, dana, waktu, prosedur pelaksanaan, tingkatan
polimorfisme dan pengujian secara statistik. Penanda tersebut masing-masing
mempunyai kelebihan dan kelemahan, oleh karena itu kombinasi beberapa teknik
akan memberikan data yang lebih komprehensif dan akurat. Penentuan teknik
yang digunakan sangat penting untuk mendapatkan hasil sesuai dengan yang
diinginkan. Umumnya strategi pemilihan teknik berdasarkan pada tujuan studi,
ketersediaan dana dan fasilitas serta kemampuan sumber daya manusia
(Afifah, 2012).
2.4. Penanda Molekuler RAPD
Teknik RAPD (Random Amplified Polymorphic DNA) merupakan salah satu
metode yang dapat digunakan dalam analisis DNA tanaman dengan menggunakan
alat Polymerase Chain Reaction (PCR). Alat ini berguna mengamplifikasi DNA
hasil ekstraksi dalam jumlah kecil dan waktu singkat. Penanda molekular RAPD
dihasilkan melalui proses amplifikasi DNA secara in vitro dengan PCR
(Williamet al.,1990).
Dasar polimorfisme DNA berdasarkan Polymerase Chain Reaction (PCR)
menggunakan primer tunggal dari urutan nukleotida acak dan terdeteksi sebagai
produk amplifikasi segmen DNA acak. Polimorfisme, disebut juga dengan
random amplified polymorphic DNA (RAPD marker) yang membedakan antara
berdasarkan untaian primer (Javornik & Kump, 1993). Menurut Syafaruddin &
Santoso (2011), RAPD adalah penanda berbasis PCR dengan menggunakan 10
basa primer acak. Teknik RAPD tidak memerlukan informasi awal tentang urutan
basa suatu spesies. Yang diperlukan adalah DNA yang relatif murni dan dalam
jumlah yang relatif kecil dibandingkan dengan RFLP. Oleh karenanya RAPD
dapat diterapkan pada hampir semua jenis tanaman. DNA sebagai pembawa
informasi genetik terdapat dalam sel khususnya di dalam inti sel dan untuk
mendapatkannya dapat dilakukan dengan proses ekstraksi, sehingga, memudahkan
untuk mengidentifikasi DNA yang disebut isolasi DNA (Langgaet al.,2012).
Metode RAPD merupakan pengembangan teknik PCR untuk mendeteksi
keanekaragaman genetik atau mengidentifikasi potongan DNA spesifik yang
berkomplementer dengan DNA cetakan. RAPD bertujuan untuk menghasilkan
banyak copy dari DNA eetakan. Potongan-potongan acak yang umumnya
berukuran antara 250-2000 pasangan basa (bp) diamplifikasi menggunakan PCR
dengan primer tunggal, yang pada umumnya berukuran 10 pasang basa. Reaksi
RAPD umumnya menghasilkan 3-10 potongan DNA. Produk amplifikasi
biasanya dianalisis dengan elektroforesis pada gel agarosa yang dilanjutkan
pengecatan denganethidium bromide(Yasminingsih, 2009).
PCR adalah suatu metode untuk melipat gandakan suatu pita DNA secara
in vitro. Metode PCR dikembangkan oleh Kary Mullis pada tahun 1986
(Irawan, 2008). Proses terjadinya reaksi amplifikasi melalui tiga tahapan yaitu, (a)
denaturasi, merupakan proses awal yang dilakukan dengan pemanasan hingga
96°C selama 30-60 detik. Pada suhu ini DNA utas ganda akan memisah menjadi
utas tunggal, (b) annealing, setelah DNA menjadi utas tunggal, suhu diturunkan
ke kisaran 40-60°C selama 20-40 detik untuk memberikan kesempatan bagi
primer untuk menempel pada DNA template di tempat yang komplemen dengan
sekuen primer, (c) ekstension/elongasi, merupakan proses akhir dimana dilakukan
dengan menaikkan suhu ke kisaran suhu kerja optimum enzim DNA polymerase,
biasanya 70-72°C. Pada tahap ini DNA polymerase akan memasangkan dNTP
yang sesuai pada pasangannya. Jika basa pada template adalah A, maka akan
dipasang dNTP T, begitu seterusnya (pasangan A adalah T, dan C dengan G).
10
ekstensi bergantung pada panjangnya daerah yang akan diamplifikasi, secara
umumnya adalah 1 menit untuk setiap 1000 bp (Erlich, 1989). Tiga langkah
tersebut merupakan satu siklus PCR. Proses ini disebut chain reactionatau reaksi
berantai sebab hasil dari langkah ke tiga, yaitu dua pita ganda, masing-masing
akan menjalani siklus PCR, mengalami denaturasi, annealing, dan pemanjangan
lagi dan demikian seterusnya. Karena setiap hasil daur selalu melakukan reaksi
ulang yang sama inilah maka disebut reaksi berantai (Irawan, 2008).
Prosedur penggunaan RAPD ini mempunyai keutungan seperti sederhana,
mudah dalam hal preparasi, dapat dilakukan secara maksimal untuk sampel dalam
jumlah banyak, jumlah DNA yang diperlukan relatif sedikit, dan pengerjaannya
tidak menggunakan senyawa radioaktif (Syafaruddin & Santoro, 2011). Menurut
Anggraeni (2008), penanda RAPD memiliki beberapa kelebihan, diantaranya
tidak membutuhkan latar belakang pengetahuan genom yang akan dianalisis,
primer universal dapat digunakan untuk organisme prokariotik maupun eukariotik,
mampu menghasilkan karakter yang relatif lebih murah, mudah dalam preparasi,
dan memberikan hasil lebih cepat dibandingkan dengan analisis keragaman
molekulernya. Kelebihan lain yang lebih spesifik adalah teknik RAPD lebih
sederhana, yaitu: (1) DNA tidak perlu dipotong dengan enzim restrikasi, (2)
sampel DNA yang diperlukan relatif sedikit, (3) tidak perlu memindahkan DNA
ke membran nilon, (4) tidak memerlukan hibridisasi DNA dan (5) tidak
memerlukan prosedurlabeling.
Penanda RAPD telah berhasil digunakan untuk tujuan dalam bidang
pemuliaan tanaman antara lain: (1) penyusunan peta genetik, (2) analisis struktur
genetik populasi, (3) sidik jari individu, (4) pemetaan sifat-sifat, dan (5) penanda
khas pada bagian genom (Yasminingsih, 2009). Penanda RAPD telah banyak
digunakan untuk mempelajari keanekaragaman genetik seperti, jeruk (Karsinahet
al., 2002), jarak pagar (Yasminingsih, 2009), mentimun (Julisaniah et al., 2008)
dan kelapa (Roslim, 2003).
BAB 3
METODOLOGI PENELITIAN
3.1. Waktu dan Lokasi
Penelitian ini dilakukan pada bulan Juli 2014 hingga Februari 2015 di
Laboratorium Terpadu dan Laboratorium Genetika, Fakultas Matematika dan
Ilmu Pengetahuan Alam, Universitas Sumatera Utara.
3.2. Alat dan Bahan
Alat yang digunakan dalam penelitian ini adalah water bath, shaker, spin down,
nanophotometer, UV transluminator, PCR (Polymerase Chain Reaction),
elektroforesis, sentrifugasi, freezer, pipet mikro, tip, mortar dan porselin, hot
plate, autoklaf, neraca analitik, tabung ependorf, erlenmeyer, alumunium foil,
gunting, kertas label.
Bahan tanaman yang digunakan dalam penelitian ini adalah pucuk daun
markisa. Bahan kimia yang digunakan adalah buffer ekstraksi CTAB (100 mM
Tris HCL, 4M Nacl, 20 mM EDTA, 3% CTAB, 6% PVP dan 0,2% β
-merkaptoetanol), Kloroform, Isoamilalkohol, Isopropanol, Alkohol 70%, 5 M
NaCl, sodium asetat, Fenol, Buffer TE, TAE 1X, Agarose, Promega GoTaq®
Green Master Mix, ddH20 steril, primer RAPD (Tabel 3.1.),Ethium bromide1%,
Loading dye6X, RNA-se, Proteinase-K.
3.3. Prosedur Penelitian
3.3.1. Pengkoleksian Sampel
Sampel tanaman markisa dikoleksi dari 4 kabupaten di Sumatera Utara
yaitu kabupaten Karo, kabupaten Simalungun, kabupaten HUMBAHAS
(Humbang Hasundutan) dan kabupaten Tapanuli Utara yang merupakan daerah
pembudidaya tanaman markisa secara komersial. Jenis markisa yang dikoleksi
12
(Passiflora edulis f. flavicarpa Deg.), markisa asam merah (Passiflora edulis f.
edulis Sims.), markisa konyal (Passiflora ligularis L.), markisa erbis (Passiflora
quadrangularis), F1 (markisa ungu besar hasil persilangan antara markisa ungu
dengan markisa merah) dan markisa liar (Passiflora foetida) (Lampiran 1.).
3.3.2. Isolasi DNA
DNA diisolasi dari bagian daun yang masih muda (bagian pucuk) dengan
menggunakan metode CTAB (Doyle & Doyle, 1990) yang dimodifikasi oleh Lade
et al. (2014), Sebanyak 0,5 g daun muda digerus dalam cawan porselin dengan
menambahkan 1 ml buffer CTAB (100 mM Tris HCL, 4M Nacl, 20 mM EDTA,
3% CTAB, 6% PVP dan 0,2% β-merkaptoetanol) tanpa menggunakan nitrogen
cair. Hasil gerusan dipindahkan kedalam tabung eppendorf 2 ml. Ditambahkan
RNase 1 µl dari 10 mg/ml kemudian inverted dan diinkubasi pada suhu 37 oC
selama 15 menit. Kemudian ditambahkan 10 µl Proteinase-K (1 mg/ml) dan
diinkubasi dalam waterbath pada suhu 65 oC selama 1 jam (inverted 15 menit
sekali), setelah itu didiamkan di suhu ruang.
Proses pemurnian diawali dengan penambahan klorofom: isoamilalkohol
(24:1) hingga mencapai 1 kali volume sampel, kemudian dihomogenkan dengan
dibolak-balik (inverted) kemudian disentrifuse (eppendorf centrifuge 5430R)
dengan kecepatan 10.000 rpm selama 15 menit pada suhu 4 °C. Supernatan
diambil dengan pipet mikro dan dipindahkan ke eppendorf baru yang steril.
Ditambahkan 5 M NaCl sebanyak setengah volume supernatan dan diinkubasi di
es selama 15 menit. Sodium asetat ditambahkan sebanyak 1/10 volume supernatan
dan isopropanol dingin sampai tube penuh (inverted secara perlahan, dilihat
adanya benang-benang halus dari DNA).Selanjutnya larutan tersebut diendapkan
dengan cara diinkubasi pada suhu -20 °C selama 1 jam. Larutan disentrifuse
(eppendorf centrifuge 5430R)dengan kecepatan 10.000 rpm selama 15 menit pada
suhu 4 °C, dan cairan dibuang hingga terlihat endapan DNA berupa pelet pada
ujung eppendorf. Endapan yang merupakan DNA dicuci dengan alkohol/EtOH
70%, kemudian disentrifuse (eppendorf centrifuge 5430R) kembali dengan
kecepatan 10.000 rpm selama 10 menit pada suhu 4°C. Alkohol/EtOH dibuang
telah kering dilarutkan dengan ddH20 steril.
3.3.3. Uji Kualitas dan Kuantitas DNA
Uji kualitas dan kuantitas DNA dilakukan untuk melihat konsentrasi dan
kemurniannya dengan menggunakan nanophotometer (IMPLEN P-360
NanoPhotometer P-Class) dan elektroforesis (C.B.S. Scientific EPS-300 X)
dengan menggunakan gel agarose 0,8%. Dilakukan dengan menimbang 0,32 g
bubuk agarose dan menambahkan 40 ml TAE 1X kemudian dipanaskan dengan
hot plate sampai mendidih/ warna larutan menjadi bening. Larutan agarose
didinginkan sampai suhu ± 60 °C, kemudian dituangkan kedalam cetakan
elektroforesis yang telah disiapkan dan dibiarkan sampai memadat. Selanjutnya,
larutan TAE 1X dimasukkan kedalam bak elektroforesis sampai gel agarose
terendam. Sampel DNA yang akan diuji diambil sebanyak 5 µl dan ditambahkan
loading dye 6X sebanyak 1 µl, selanjutnya sampel yang telah disiapkan
dimasukkan dalam sumur gel dan dielektroforesis selama 30 menit pada voltase
70 V dan 100 mA. Hasil elektroforesis tersebut diwarnai dengan melakukan
perendaman pada larutan ethidium bromide (EtBR 1%) selama 15 menit
kemudian direndam dalam aquades selama 10 menit. Kualitas DNA hasil
elektroforesis diamati di bawah UV transluminator (G.BOX SYNGENE) dan
didokumentasikan dengan alatKodak gel logicdengansoftware.
Uji kuantitas DNA dilakukan dengan pengukuran Kemurnian dan
konsentrasi DNA dapat dihitung melalui perbandingan A260 nm dan A280 nm
menggunakan nanophotometer (IMPLEN P-360 NanoPhotometer P-Class).
3.3.4. Amplifikasi DNA dengan Teknik RAPD
Amplifikasi DNA genom markisa dilakukan jika sampel DNA sudah
memenuhi kualitas dan kuantitas yang baik. Amplifikasi dilakukan dalam mesin
PCR (Polymerase Chain Reaction) menggunakan 7 primer acak (Tabel 3.1.).
14
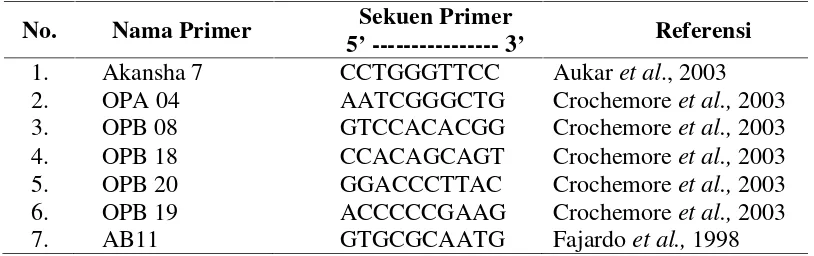
Tabel 3.1. Primer acak yang digunakan untuk analisis RAPD
No. Nama Primer Sekuen Primer
5’---3’ Referensi
1. Akansha 7 CCTGGGTTCC Aukaret al., 2003 2. OPA 04 AATCGGGCTG Crochemoreet al.,2003 3. OPB 08 GTCCACACGG Crochemoreet al.,2003 4. OPB 18 CCACAGCAGT Crochemoreet al.,2003 5. OPB 20 GGACCCTTAC Crochemoreet al.,2003 6. OPB 19 ACCCCCGAAG Crochemoreet al.,2003 7. AB11 GTGCGCAATG Fajardoet al.,1998
Reaksi amplifikasi selanjutnya dilakukan dengan memasukkan tabung yang telah
berisi bahan untuk reaksi PCR ke dalam blok mesin PCR(ependorf vapo protect)
dengan waktu yang digunakan adalah: inisiasi denaturasi 95 °C selama 30 detik,
selanjutnya denaturasi 95 °C selama 1 menit, annealing (suhu optimum primer)
34oC selama 30 detik, dan ekstension pada suhu 72 °C selama 1 menit, diikuti
ekstension akhir pada suhu 72 °C selama 5 menit, pendinginan setelah siklus
selesai pada suhu 4 oC. Reaksi PCR dilakukan sebanyak 40 siklus. Hasil reaksi
PCR dielektroforesis pada 1,2% gel agarose. Elektroforesis dilakukan selama 90
menit pada tegangan 70 V, 500 mA pada suhu ruang. Pewarnaan hasil
elektroforesis dilakukan dengan merendam agarose dalam larutan ethidium
bromide (EtBR 1%) selama 10 menit. Gel yang telah diwarnai kembali direndam
dalam aquades steril selama 3 menit dan dilanjutkan pengamatan pita hasil
amplifikasi dengan menggunakan uv transiluminator (G.BOX SYNGENE).
Tabel 3.2. Bahan untuk satu kali reaksi PCR
No. Komponen PCR Volume
1. GoTaq® Green Master Mix, 2X 10 µl
2. DNA template (<250 ng) 2 µl
3. Primer (10 pmol/µl) 1 µl
4. Nuclease free water 7 µl
3.4. Analisis Data
3.4.1. Pita Polimorfik DNA
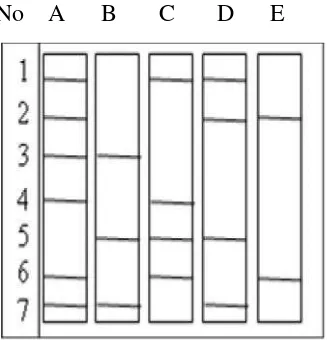
DNA hasil PCR diterjemahkan kedalam data biner berdasarkan ada
tidaknya pita, dengan ketentuan nilai 0 (nol) untuk tidak ada pita, dan nilai 1
(satu) untuk adanya pita pada suatu posisi yang sama dari setiap individu yang
dibandingkan. Cara pemberian nilai dapat dilihat pada Gambar 3.1.
No A B C D E No A B C D E
Gambar 3.1. Pola Terjemahan Pita DNA
3.4.2. Analisis Similaritas
Data biner pita DNA diproses dengan bantuan program Numerical Taxonomy and Multivariate Analysis System(NTSYS) ver. 2.11a.Hasil analisis ini
disajikan dalam bentuk pohon dendogram.
16
BAB 4
HASIL DAN PEMBAHASAN
4.1. Analisis Kualitatif dan Kuantitatif Pita Hasil Isolasi DNA Tanaman Markisa
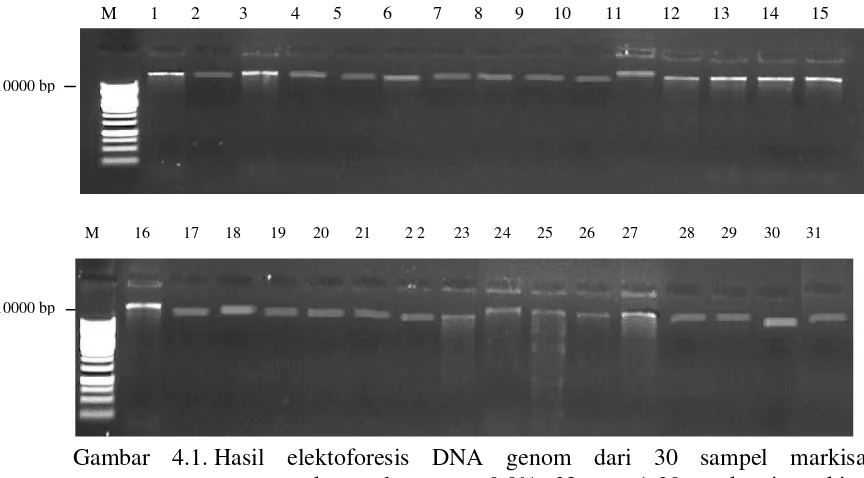
Hasil isolasi DNA genom dari 31 sampel markisa dielektroforesis pada gel
agarose 0,8%. Uji ini dilakukan untuk mengetahui kualitas pita DNA yang
diperoleh sehingga dapat digunakan untuk proses PCR. Hasil elektroforesis DNA
genom menunjukkan adanya satu pita utuh pada setiap sumur gel.
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
M 16 17 18 19 20 21 2 2 23 24 25 26 27 28 29 30 31
Gambar 4.1. Hasil elektoforesis DNA genom dari 30 sampel markisa menggunakan gel agarose 0,8%. Nomor 1-30 = aksesi markisa (Lampiran 1.)
Pita utuh DNA genom ini ditandai dengan tidak adanyasmear(DNA tidak
terdegradasi dan terkontaminasi). Ukuran pita DNA genom yang didapat adalah
diatas 10000 bp. Menurut Aulia (2014), ciri DNA yang utuh pada elektroforesis
adalah DNA dapat bermigrasi pada pori-pori agarose dalam buffer pada arus
tertentu dan tidak tercacah (smear). Menurut Syafaruddin & Santoso (2011), DNA
yang utuh ditandai dengan tidak adanyasmearDNA yang dielektroforesis. Hal ini
10000 bp
menjadi penting karena pada proses PCR, DNA yang masih utuh akan lebih
memberikan hasil yang relatif lebih akurat.
Hasil pengukuran kuantitas dan kemurnian dari 31 DNA genom markisa
dengan nanofotometer (IMPLEN P-360 NanoPhotometer P-Class) menunjukkan
kemurnian DNA yang baik dan kuantitasnya yang cukup (Lampiran 2).
Kemurnian yang diperoleh berkisar antara 1,368-2,112. Menurut Sambrook &
Russel (1989), batas kemurnian yang biasa digunakan dalam analisis molekuler
pada rasio A260 /A280adalah 1,8-2,0. Kemurnian yang diperoleh tidak semuanya
memenuhi batas kriteria untuk dapat digunakan dalam analisis molekuler (<1,8),
namun pada penelitian yang telah dilakukan oleh Minarsihet al., (2011), terhadap
keragaman genetik Ganoderma spp., nilai kemurnian 1,069 merupakan nilai
terendah tetapi sudah dapat digunakan untuk proses PCR-RAPD. Selain itu pada
penelitian yang telah dilakukan oleh Bayzura (2014) & Aulia (2014) terhadap
tanaman kelapa sawit, nilai kemurnian 1,5 - 1,7 sudah dapat digunakan untuk
melakukan proses PCR-RAPD. Rendahnya nilai rasio A260/A280 (<1,8), dapat
disebabkan oleh kontaminasi protein dan bahan organik lainnya sebaliknya
kontaminasi fenol ditandai dengan tingginya nilai rasio tersebut (>2,0) (Linacero
et al.,1998).
Konsentrasi DNA genom markisa yang diperoleh berkisar antara 42,5
sampai 2552 ng/µl. Konsentarsi DNA yang biasanya digunakan dalam proses
PCR adalah 0,5-50 ng/µl, maka konsentrasi yang didapatkan pada penelitian ini
sudah cukup memenuhi syarat dalam melakukan RAPD. Menurut Haris et al.
(2003), konsentrasi DNA akan berdampak pada kualitas fragmen hasil
amplifikasi. Konsentrasi DNA yang rendah akan menghasilkan fragmen yang
tipis, sebaliknya konsentrasi DNA yang tinggi akan menyebabkan fragmen
terlihat tebal sehingga sulit membedakan antar fragmen.
4.2. Analisis Profil Pita RAPD Tanaman Markisa
Amplifikasi dengan menggunakan primer RAPD yang dilakukan terhadap DNA
genom markisa menghasilkan produk PCR yang dapat dibaca dan diskor,
18
dapat mengamplifikasi DNA genom dan menghasilkan pita DNA polimorfik,
sedangkan primer lainnya tidak menghasilkan pita.
Primer yang dapat mengamplifikasi DNA genom pada 31 aksesi yaitu
primer Akansha 7, OPA 04, OPB 08, dan OPB 18 sedangkan primer OPB 19,
OPB 20 dan AB 11 tidak dapat mengamplifikasi pita DNA genom. Menurut
Kumar et al. (2011), ketidakcocokan antara primer dengan template DNA dapat
mengakibatkan ketiadaan total produk PCR. Keempat primer yang menghasilkan
pita DNA polimorfik selanjutnya digunakan untuk mengamplifikasi DNA genom
dari 31 aksesi markisa (Lampiran 3.).
Primer yang digunakan dalam analisis RAPD adalah sintetis
oligonukleotida pendek (10 pasang basa) dengan sekuens acak untuk
mengamplifikasi kuantitas nanogram dari total genomik DNA
(Kumar et al., 2011). Menurut Hannum (2001), RAPD merupakan hasil
berpasangnya nukleotida primer dengan nukleotida genom tanaman, semakin
banyak primer yang digunakan akan semakin terwakili bagian-bagian genom,
sehingga semakin tergambar keadaan genom tanaman yang sesungguhnya.
Perbedaan jumlah dan polimorfisme pita DNA yang dihasilkan dari setiap primer
menggambarkan kekompleksan genom tanaman yang diamati
(Grattapagliaet al, 1992).
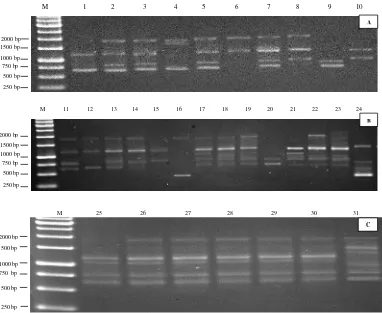
Profil pita DNA pada primer Akansha 7 terhadap 31 aksesi markisa
(Gambar 4.2.) menunjukkan pola pita yang berbeda antar aksesi dengan total pola
pita yang dihasilkan sebanyak 25 pola dengan ukuran475–2279bp.
Hasil amplifikasi menyatakan bahwa dengan penggunaan primer Akansha
7 pada 31 aksesi markisa menghasilkan profil pita yang baik dengan keadaan pita
jelas. Hal ini dipengaruhi oleh kondisi DNA hasil ekstraksi yang menunjukkan
adanya kesamaan sekuens antara DNA cetakan dengan primer. Menurut Langga
(2012), semakin banyak sekuens yang dapat dikenali oleh primer pada sebuah
M 1 2 3 4 5 6 7 8 9 10
M 11 12 13 14 15 16 17 18 19 20 21 22 23 24
M 25 26 27 28 29 30 31
Gambar 4.2. Profil Pita RAPD tanaman markisa dengan primer Akansha. M= Marker 1kb, nomor 1-31 = aksesi markisa (Lampiran 1.)
Jumlah dan intensitas pita DNA yang dihasilkan setelah amplifikasi
DNA dengan PCR sangat tergantung bagaimana primer mengenal urutan DNA
komplementernya pada cetakan DNA. Hasil amplifikasi DNA tidak selalu
memperoleh pita dan intensitas yang sama. Intensitas pita DNA produk PCR pada
setiap primer sangat dipengaruhi oleh kemurnian dan konsentrasi cetakan DNA.
Cetakan DNA yang mengandung senyawa-senyawa seperti polisakarida dan
senyawa fenolik, serta konsentrasi DNA yang terlalu kecil sering menghasilkan
pita DNA amplifikasi yang redup atau tidak jelas (Weeden et al., 1992 dalam
Harahap, 2014). Menurut Hannum (2001), jumlah pita DNA yang dihasilkan oleh
setiap primer bergantung pada sebaran situs yang homolog dengan sekuen primer
pada genom.
Keempat primer yang digunakan memperlihatkan bahwa jumlah pita yang
dihasilkan pada setiap primer berbeda. Dari keempat primer tersebut
20
yang dihasilkan bervariasi yaitu 229-2279 bp. Persentase pita polimorfik sebesar
100% (akansha 7, OPA 04, OPB 08) dan 94% pada primer OPB 18.
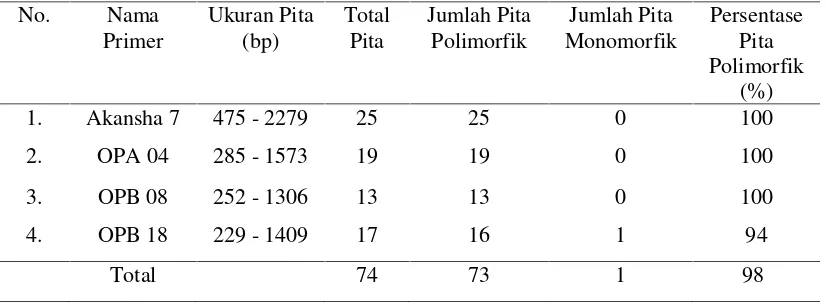
Tabel 4.1. Persentase Pita Polimorfik pada 4 primer
No. Nama
Jumlah dan ukuran pita tertinggi terdapat pada primer Akansha 7 yang berjumlah
25 pita pada ukuran sekitar lebih dari 2279 bp, sedangkan jumlah dan ukuran pita
terendah terdapat pada primer OPB 18 dengan jumlah 13 pita pada ukuran 229 bp
Primer Akansha 7 menghasilkan 25 pita DNA dengan persentase
polimorfik sebesar 100%, pada penelitian Lade et al. (2014), dari 10 primer Akansha yang dioptimasi untuk mengamplifikasi DNA markisa liar (P. foetida),
primer Akansha 7 merupakan primer dengan angka amplifikasi tertinggi yaitu 9
pita fragmen.
Primer OPA 04, OPB 08, dan OPB 18 menghasilkan pita berturut-turut
adalah 19, 13 dan 17 pita. Dari ketiga primer tersebut dapat dibandingkan bahwa
primer OPA 04 merupakan primer yang dapat menghasilkan pita terbanyak. Hal
ini juga ditunjukkan dalam penelitian yang telah dilakukan oleh Crochemoreet al.
(2003), tentang keanekaragaman genetik passion fruit (Passiflora spp.) dengan
penggunaan primer yang sama yaitu primer OPA 04 memperlihatkan angka
amplifikasi tertinggi sebanyak 33 pita, primer OPB 18 mengamplifikasi sebanyak
4.3. Analisis Keanekaragaman Genetik Tanaman Markisa
4.3.1. Analisis Hubungan Genetik Markisa Ungu (Passiflora edulis)
Hubungan genetik setiap populasi tanaman markisa ungu dalam penelitian ini
dapat ditentukan berdasarkan kemiripan genetik antar individu tanaman dalam
populasi yang sama dan antar populasi yang berbeda dengan cara membandingkan
pita RAPD dari setiap individu tanaman.
Hasil analisis pengelompokan aksesi markisa ungu yang dikoleksi dari 3
kabupaten berdasarkan penanda RAPD dengan 4 primer yang terseleksi, dibuat
matriks kemiripan genetik berdasarkan Coefficient Dice antar individu markisa
ungu (Lampiran 4). Koefisien kemiripan genetik pada aksesi markisa ungu
berkisar antara 0,97-0,58. Kemiripan genetik tertinggi terdapat pada kabupaten
HUMBAHAS antara aksesi MUH1 dengan MUH2 sebesar 0,97 (97%). Hal ini
menunjukkan bahwa aksesi MUH1 dengan MUH2 berkerabat dekat.
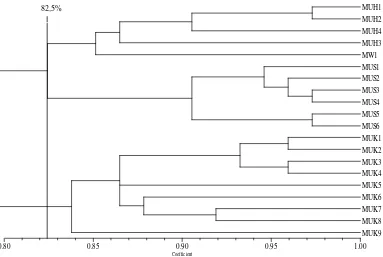
Berdasarkan kemiripan genetik secara grafik yang diperlihatkan dalam
bentuk dendogram (Gambar 4.3.) dapat dilihat bahwa aksesi markisa ungu
mengelompok berdasarkan wilayah pengkoleksiannya. Hal ini mengindikasikan
bahwa markisa ungu pada wilayah yang sama memiliki latar belakang genetik
yang cukup dekat.
Aksesi markisa ungu dari kabupaten Simalungun mengelompok pada
kemiripan genetik yang lebih tinggi dibandingkan dua kabupaten lainnya yaitu
90,5%, hal ini mencermikan bahwa hubungan genetik antar individu pada
kabupaten tersebut lebih dekat. Pada kemiripan genetik 82,5% aksesi markisa
ungu dari kabupaten HUMBAHAS dan Simalungun mengelompok, dan pada
akhirnya bergabung dengan markisa ungu dari kabupaten Karo pada kemiripan
genetik 80%. Hal ini menunjukkan bahwa markisa ungu dari kabupaten
HUMBAHAS mempunyai hubungan genetik yang lebih dekat dengan markisa
22
Gambar 4.3. Dendogram kemiripan genetik markisa ungu (Passiflora edulis) berdasarkan 4 primer RAPD. MU = markisa ungu; MW= markisa liar. H= HUMBAHAS; S= Simalungun; K= Karo.
Analisis pengelompokkan pada aksesi markisa ungu ini memperlihatkan
bahwa setiap individu mengelompok berdasarkan wilayah pengkoleksiannya. Hal
ini mungkin disebabkan karena adanya perbedaan lingkungan tempat tumbuh
pada setiap wilayah pengkoleksian, sehingga dapat mempengaruhi genetik dari
markisa tersebut. Menurut Viana et al. (2006), dalam penelitiannya tentang
keanekaragaman genetik markisa kuning yang dikoleksi dari tiga lokasi yang
berbeda menunjukkan bahwa markisa kuning mengelompok pada setiap
populasinya. Individu pada populasi yang sama, bekerja dan
bersaing/berkompetisi, dan menyesuaikan diri dengan lingkungan tempatnya
tumbuh, sehingga menunjukkan kesamaan genetik antar individu. Dari hasil
penelitian yang dicapai, markisa kuning pada satu populasi mempunyai
heritabilitas sekitar 92% untuk jumlah sifat dari buah pada populasi yang sama.
Menurut Falconer (1987)dalam Vianaet al.(2006), satu genotip yang dievaluasi
dari lingkungan yang berbeda akan membentuk kelompok yang berbeda sesuai
lingkungannya. Perbedaan lingkungan menunjukkan adanya perbedaan respon
markisa pada penelitian ini mengelompok pada koefisien kemiripan genetik 80%
kemudian terpisah berdasarkan masing-masing kabupaten.
Pengelompokkan individu-individu tanaman markisa berdasarkan
kemiripan genetik, tidak bisa hanya dari kesamaan warna buahmya saja. Seperti
yang telah dilakukan dalam penelitian Hannum (2001), terhadap hubungan
genetik kelapa genjah yang berwarna buah hijau (populasi kelapa genjah GHJ dan
GHN) memperlihatkan bahwa masing-masing individu populasi kelapa
mengelompok pada populasi masing-masing pada kemiripan genetik 83%, kedua
populasi itu hanya mempunyai kemiripan genetik 69%.
Keanekaragaman genetik pada satu spesies dari suatu populasi dipengaruhi
oleh sejumlah faktor evolusi, termasuk sistem persilangan, penyebaran biji, aliran
gen, seleksi alam, dan jangkauan geografis (Hamrick & Godt, 1989), sehingga
keanekaragaman genetik Passiflora cukup luas karena adanya penyebaran
geografis dari setiap genusnya. Menurut Lopes (1991), hal ini disebabkan karena
dalam kebanyakan kasus, produsen markisa memproduksi sendiri bibit baik dari
biji yang dikumpulkan dilingkungan mereka atau dari buah-buahan segar yang
dibeli dipasar yang tidak diketahui asalnya. Akibatnya hubungan genetik yang
dekat dapat diamati diantara aksesi markisa yang plasma nutfahnya dikoleksi dari
lokasi yang sama atau origin (pedigree) yang sama tetapi dari lahan yang berbeda.
4.3.2. Analisis Hubungan Genetik Markisa (Passiflora spp.) dari Sumatera Utara
Hasil analisis pengelompokan 31 markisa (markisa asam ungu (Passiflora edulis),
markisa asam kuning (Passiflora edulis f. flavicarpa Deg.),markisa asam merah
(Passiflora edulis f. edulis Sims.), markisa konyal (Passiflora ligularis),markisa
erbis (Passiflora quadrangularis), F1 (markisa ungu besar hasil persilangan antara
markisa merah dengan markisa ungu) dan markisa liar (Passiflora foetida L.))
yang dikoleksi dari 4 kabupaten berdasarkan penanda RAPD dengan 4 primer
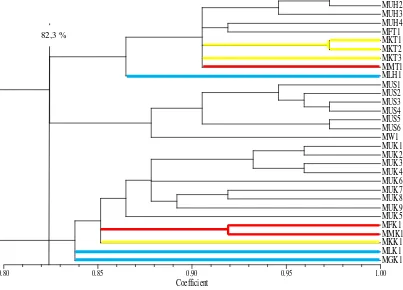
acak ditampilkan dalam bentuk dendogram (Gambar 4.4.).
Keseluruhan markisa pada penelitian ini mengelompok pada koefisien
kemiripan genetik 80% kemudian terpisah berdasarkan masing-masing kabupaten.
Markisa asal kabupaten Karo terpisah dari 2 kabupaten lainnya dan mengelompok
24
Tapanuli Utara membentuk satu kelompok pada kemiripan genetik 82,3%
kemudian memisah pada kemiripan 86,8%.
Gambar 4.4. Dendogram kemiripan genetik markisa (Passiflora sp.) berdasarkan 4 primer RAPD. MU = markisa ungu; MK= markisa kuning, MM= markisa merah; ML= markisa konyal; MG= markisa giant/erbis; MW= markisa liar; dan MF= markisa F1. H= HUMBAHAS; T= Tapanuli Utara; S= Simalungun; K= Karo. = markisa kuning;
= markisa merah; = markisa beda spesies (markisa giant/erbis & markisa konyal); = markisa ungu
Markisa asal kabupaten Simalungun mempunyai hubungan genetik yang
lebih dekat dengan markisa asal kabupaten Humbahas dan Tapanuli Utara,
dibandingkan dengan markisa asal kabupaten Karo. Populasi markisa yang jarak
geografisnya lebih dekat (markisa dari kabupaten Simalungun dengan kabupaten
Karo) ternyata mempunyai hubungan genetik yang lebih jauh dibandingkan
markisa yang jarak geografisnya lebih jauh (kabupaten Simalungun dengan
kabupaten HUMBAHAS dan Tapanuli Utara). Menururt Hannum (2001), dalam
penelitian mengenai hubungan genetik kelapa genjah, menyatakan bahwa jauh
dekatnya daerah asal tidak menjamin jauh dekatnya hubungan genetik kelapa
genjah. Hal ini juga didukung dari hasil penelitian Pandin (2000) dalam Hannum
(2001), yang diamati pada kelapa Dalam Mapanget (DMT) dan Dalam Tenga
(DTA) yang jarak geografis daerah asalnya lebih dekat dari pada daerah asal
kelapa Dalam Palu (DPU), ternyata mempunyai kemiripan genetik yang lebih
rendah dibandingkan dengan kemiripan genetik antar DMT dan DPU, dan antar
DTA dan DPU berdasarkan primer acak RAPD.
Pada dendogram dapat dilihat bahwa pada kabupaten HUMBAHAS dan
Tapanuli Utara, markisa konyal (MLH1) dan markisa asam (MUH1, MUH2,
MUH3, MUH4, MFT1, MKT1, MKT2, MKT3, dan MMT1) memiliki kemiripan
genetik 86,5%. Keseluruhan markisa asam pada kabupaten ini mengelompok dan
memiliki kemiripan genetik 90,5%, tetapi markisa ungu (MUH4) dan markisa
persilangan (MFT1) membentuk satu kelompok dan memisah dari markisa asam
lainnya pada kemiripan genetik 91,9%. Kemiripan genetik markisa tertinggi pada
kabupaten ini adalah 97% yaitu antara markisa asam ungu (MUH1 dan MUH2)
dan markisa asam kuning (MUKT1 dan MUKT2).
Markisa liar (Passiflora foetida L), aksesi MW1, mengelompok bersama
markisa dari kabupaten Simalungun pada kemiripan genetik 88%, tetapi markisa
liar ini memisah dengan aksesi lain yang mengelompok pada kemiripan genetik
90,5% yang mana merupakan jenis dari markisa ungu (Passiflora edulis Sims.).
Hubungan genetik markisa dari kabupaten Simalungun mempunyai kekerabatan
yang cukup tinggi antar individunya.
Pada kabupaten Karo dapat dilihat bahwa pada koefisien kemiripan
genetik 83,9%, markisa (Passiflora sp.) membentuk 3 kelompok yaitu kelompok
markisa asam (MUK, MKK, MMK, MFK), kelompok markisa konyal (MLK1)
dan kelompok markisa giant/erbis (MGK1). Kelompok markisa asam ini memisah
dari 2 kelompok lainnya dan berkelompok pada kemiripan genetik 85,1% dengan
membentuk kembali 3 kelompok yang berbeda yaitu kelompok markisa ungu
(MUK) yang akan membentuk satu kelompok pada koefisien kemiripan 86,4%,
kelompok markisa kuning (MKK), dan kelompok markisa merah (MMK) yang
akan bergabung dengan markisa persilangan (MFK) pada kemiripan genetik
91,9%.
Markisa konyal (Passiflora ligularis) dan markisa giant/erbis (Passiflora
26
markisa asam (Passiflora edulis), sehingga pada dendogram dapat dilihat adanya
pemisahan koefisien kemiripan genetik diantara ketiganya. Sedangkan markisa
asam dengan spesies yang sama tetapi forma berbeda yaitu markisa merah
(Passiflora edulisf.edulisSIMS), markisa kuning (Passiflora edulisf.flavicarpa)
dan markisa F1 (hasil persilangan antara markisa merah (Passiflora edulif. edulis
Sims.) dengan markisa ungu (Passiflora edulis Sims.) mempunyai kemiripan
genetik sebesar 85,1% - 97% (pada ketiga kabupaten).
Menurut Bellon et al. (2007), pada penelitiannya, aksesi markisa asam
(Passiflora edulis Sims.) ini menunjukkan variabilitas genetik yang tinggi.
Variabilitas yang tinggi ini menunjukkan meskipun taksonomi berasal dari spesies
yang sama (markisa ungu dengan markisa kuning) namun secara genetik berbeda.
Passiflora edulis f. flavicarpa merupakan populasi dari Passiflora edulis yang
diperoleh dari hasil mutasi Passiflora edulis dan bukan dari hasil persilangan
antaraP. edulis dengan spesies lainnya, sehingga pada sistem klasifikasi tanaman
ini didasarkan pada kemiripan morfologi dan genetik (Pope & Degener dalam
Martin & Naksone, 1970).
Dalam penelitian yang telah dilakukan oleh Halimas (2014) tentang studi
morfologi dan anatomi markisa, ketiga markisa asam ini memperlihatkan
beberapa ciri kesamaan. Dari analisis kemiripan morfologi yang dilakukan antara
markisa asam merah dan asam kuning didapatkan tingkat kemiripan morfologi
sebesar 69%, sedangkan kemiripan morfologi antara kedua markisa ini dengan
markisa konyal memiliki tingkat kemiripan morfologi sebesar 31 %. Suranto
(2002) menjelaskan bahwa jarak genetik atau hubungan kekerabatan diantara
varieras dapat menggambarkan perbedaan genetik antar varietas. Dalam penelitian
ini juga diketahui bahwa markisa F1 yang merupakan hasil persilangan antara
markisa ungu (P.edulis) dengan markisa merah (P.edulis f. edulis) memiki
BAB 5
KESIMPULAN DAN SARAN
5.1 Kesimpulan
Analisis RAPD dengan menggunakan 4 primer acak terseleksi pada 31
aksesi markisa dari Sumatera Utara menghasilkan polimorfisme yang cukup tinggi
yaitu 73 pita polimorfik (98%). Markisa (Passiflora spp.) mengelompok pada
masing-masing wilayah pengkoleksiannya. Koefisien kemiripan genetik markisa
berkisar antara 0,80 sampai 0,97. Markisa dengan spesies yang sama tetapi forma
berbeda (P. edulis f.) membentuk satu kelompok yang berbeda dan terpisah
dengan spesies lainnya (P. lingularis dan P. quadrangularis). Hubungan genetik
markisa antara kabupaten HUMBAHAS dan kabupaten Simalungun relatif lebih
dekat dibandingkan kabupaten Karo. Kedekatan jarak geografis wilayah
pengkoleksian dan kesamaan jenis markisa (warna buah) tidak menjamin
kedekatan hubungan genetik antar wilayah markisa ungu berdasarkan penanda
RAPD. Hal ini mengindikasikan bahwa setiap spesies dalam genus yang sama
memiliki genetik yang berbeda. Dengan demikian adanya variasi genetik yang
tinggi pada setiap tanaman markisa yang diuji memiliki potensi untuk
dilakukannya program pemuliaan markisa demi mendapatkan markisa yang lebih
berkualitas.
5.2 Saran
Perlu dilakukan penelitian lebih lanjut untuk mengetahui keanekaragaman
genetik tanaman markisa dengan menggunakan primer yang berbeda atau dengan
28
DAFTAR PUSTAKA
Afifah, E,N. 2012. Penggunaan Penanda Molekuler Untuk Mempercepat Dan Mempermudah Perbaikan Kualitas Tanaman Teh(Camellia sinensis (L.)
O. Kuntze). Makalah Seminar Budidaya Pertanian. Yogyakarta: Universitas Gadjah Mada.
Anggereini, E. 2008. Random Amplified Polymorphic DNA (RAPD), Suatu Metode Analisis DNA Dalam Menjelaskan Berbagai Fenomena Biologi.
Biospecies.1(2): 73-76.
Ashari, S. 1995. Hortikultur Aspek Budidaya. UI-press. Jakarta.
Aulia, I. 2014. Pengaruh Pemberian 2,4-D dan Frekuensi Subkultur Terhadap Perubahan Genetik Kalus dari Bunga Betina Kelapa Sawit (Elaeis guineensisJacq.). [Skripsi]. Medan: Universitas Sumatera Utara.
Azrai, M. 2005. Ulasan Pemanfaatan Marka Molekuler dalam Proses Seleksi Pemuliaan Tanaman.Jurnal Agro Biogen. 1(1): 26-37.
Bardakci, F. 2001. Random Amplified Polymorphic DNA (RAPD) Markers.
Turk J Biol.25: 185-196.
Bellon G, Faleiro F.G, Junqueira K.P, Junqueira N.T.V. 2007. Genetic Variability of Wild and Commercial Passion Fruit (Passiflora edulis
Sims.) Accessions Using RAPD Markers.Rev. Bras. Frutic Jaboticabal. 29: 124-127.
Anonim. 2004. [BPPHP] Buletin Teknopro Hortikultura Direktorat Pengolahan dan Pemasaran Hasil Hortikultura. Edisi 70.
Crochemore, M.L., Molinari, H. B. C., and Vieira, L. G. E. 2003. Genetic Diversity in Passion Fruit (Passiflora spp.) Evaluated by RAPD Markers.An International Journal.46:521-524.
Danarto, S., Affianto, A., Bantara, J., Adi, N.J., Sanyoto, R. 2012. Produksi Agroforesty. Indonesia Foresty and Governance Institute. pp 21-28.
Doyle, J.J., and Doyle J.L. 1990. Isolation of Plant DNA from Fresh Tissue.
Focus12:13-15.
Erlich, H.A. 1989. Polymerase Chain Reaction. Journal of Clinical Imunology.
9(6):34-47.
Fajardo, D., Angel. F., Grum, M., Tohme, J., and Lobo, M. 1998. Genetic Variation Analysis of The Genus Passiflora L. Using RAPD Markers.
Grattapaglia, D., J. Chaparro, P. Wilcox., S. McCord, D. Werner., H. Amerson.,
S. McKeand, F. Bridgwater., R. Whetten., D.O’Malley, and R. Sederoff.
1992. Mapping in Woody Plants with RAPD markers; Application to Breeding in Forestry and Horticulture. In, Application of RAPD Technology to Plant Breeding. Joint Plant breeding Symposia Society of America, Madison, WI.
Halimas, A.W. 2014. Studi Morfologi dan Anatomi Beberapa Aksesi Markisa Koleksi Balai Penelitian Tanaman Buah Kebun Percobaan Berastagi Sumatera Utara. [Skripsi]. Medan; Universitas Sumatera Utara.
Hamrick JL, Godt MJW. 1989. Allozyme Diversity in Plant Species. In: Brown AHD, Clegg MT, Kahler AL, Weir BS .(eds). Plant Population Genetics, Breeding, and Germplasm. Resources. Sinauer Associates, Sunderland. pp 43–63.
Hannum, S. 2001. Hubungan Genetik Empat Populasi Kelapa Genjah Berdasarkan Penanda RAPD (Random Amplified Polymorphic DNA). [Tesis]. Bogor: Institut Pertanian Bogor.
Harahap, A.S. 2014. Keragaman Genetik Aren Asal Sulawesi Tenggara Brdasarkan Penanda RAPD (Random Amplified Polymorphic DNA). [Tesis]. Medan: Universitas Sumatera Utara.
Haris, T.N. 1994. Development and Germination Studies of The Sugar Palm (Arenga pinnata Merr) Seed. [Disertasi]. Malaysia: Universitas Putra Malaysia.
Irawan, B. 2008. Genetika Molekuler. Air Langga University Press. Surabaya.
Javornik, B., and Kump, B. 1993. Random Amplified Polymorphic DNA (RAPD) Markers in Buckwheat.Fagopyrum.13:35-39.
Julisaniah, N. I., Sulistyowati, L., and Sugiharto, A.N. 2008. Analisis Kekerabatan Mentimun (Cucumis sativus L.) Menggunakan Metode RAPD-PCR dan Isozim.Biodiversitas.9(2):99-102.
Karmila. 2013. Analisis Kelayakan Financial Usaha Tani Markisa Konyal
(Passiflora ligularis) di Desa Arosuka Kecamatan Gunung Talang Kabupaten Solok Provinsi Sumatera Barat. [Skripsi]. Bengkulu: Universitas Bengkulu.
Karsinah, Silalahi. F.H., and Mashur, A. 2007. Eksplorasi dan Karakterisasi Plasma Nutfah Tanaman Markisa.J. Hort. 17(4)-.297-306.
Kumar, N.S., and Gurusubramanian G., 2011. Random Amplified Polymorphic DNA (RAPD) Markers and Its Application.Journal Sci Vis.11 (3): 116-124.