PENINGKATAN KERAGAMAN GENETIK LADA
(Piper nigrum L.) VARIETAS CIINTEN MELALUI IRADIASI
SINAR GAMMA DAN SELEKSI TERHADAP PENYAKIT
BUSUK PANGKAL BATANG (BPB)
NUR LAELA WAHYUNI MEILAWATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis yang berjudul Peningkatan Keragaman Genetik Lada (Piper nigrum L.) Varietas Ciinten melalui Iradiasi Sinar Gamma dan Seleksi terhadap Penyakit Busuk Pangkal Batang (BPB) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam datar pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Oktober 2016
RINGKASAN
NUR LAELA WAHYUNI MEILAWATI. Peningkatan Keragaman Genetik Lada (Piper nigrum L.) Varietas Ciinten melalui Iradiasi Sinar Gamma dan Seleksi terhadap Penyakit Busuk Pangkal Batang (BPB). Dibimbing oleh AGUS PURWITO dan NURLIANI BERMAWIE.
Lada merupakan tanaman introduksi dan selalu diperbanyak secara vegetatif, sehingga keragaman genetiknya sempit. Salah satu kendala dalam budidaya lada adalah penyakit busuk pangkal batang (BPB) yang disebabkan oleh Phytophthora capcisi. Keragaman genetik yang tinggi penting untuk menghasilkan varietas baru. Salah satu cara untuk meningkatkan keragaman genetik adalah melalui iradiasi sinar gamma. Jarak genetik dan hubungan kekerabatan antar genotipe/ aksesi plasma nutfah lada perlu diketahui untuk membantu pemulia tanaman dalam usaha merakit varietas unggul. Penelitian ini bertujuan untuk (1) mengetahui pengaruh iradiasi sinar gamma terhadap radiosensitivitas dan respon lada varietas Ciinten pada fase benih dan fase benih dengan radikula, (2) mengidentifikasi keragaman genetik mutan putatif lada varietas Ciinten berdasarkan penanda morfologi dan SSR, dan (3) Seleksi tanaman lada varietas ciinten hasil iradiasi sinar gamma terhadap penyakit busuk pangkal batang (BPB). Penelitian terdiri dari tiga percobaan/kegiatan. Penelitian dilaksanakan di rumah kaca Balittro, Laboratorium Molekuler Pemuliaan Tanaman, Balittro dan BB Biogen, serta Laboratorium Penyakit, Balittro pada bulan September 2015 sampai dengan April 2016.
Bahan tanaman yang digunakan pada percobaan I yaitu biji lada varietas Ciinten pada fase benih dan fase benih dengan radikula. Rancangan percobaan menggunakan Rancangan Split plot dengan petak utama adalah fase benih (benih dan benih dengan radikula) dan anak petak adalah dosis iradiasi dengan tujuh taraf yaitu (0, 25, 50, 75, 100, 125, 150) Gy. Masing-masing perlakuan terdiri dari tiga ulangan, setiap ulangan terdiri dari 60 benih. Bahan pada percobaan II dan III menggunakan 27 individu hasil iradiasi sinar gamma dan kontrol. Percobaan II yaitu isolasi DNA dari daun lada menggunakan metode CTAB dilanjutkan dengan uji kemurnian dan kuantitas DNA. Amplifikasi DNA dengan menggunakan PCR menggunakan 9 primer. Data yang diamati dianalisis menggunakan program NTSys dan minitab untuk mendapatkan dendogram. Hubungan kekerabatan antar mutan putatif berdasarkan karakter morfologi dan molekuler (SSR). Percobaan III yaitu seleksi ketahanan terhadap penyakit busuk pangkal batang dengan cara menginokulasi daun dengan inokulum P. capsici. Percobaan menggunakan Rancangan Acak Lengkap (RAL), apabila berbeda nyata dengan kontrol akan diuji lanjut dengan uji Dunnet pada taraf 5%. Pengamatan dilakukan 72 jam setelah inokulasi dengan mengukur luas bercak.
mengakibatkan tinggi tanaman, panjang daun semakin terhambat pertumbuhannya sehingga jumlah daun dan jumlah ruas semakin sedikit. Dosis iradiasi 25 dan 50 Gy pada fase benih dan 25 Gy pada fase benih dengan radikula nyata meningkatkan keragaman genetik berdasarkan karakter morfologi kuantitatif, morfologi kualitatif dan anatomi.
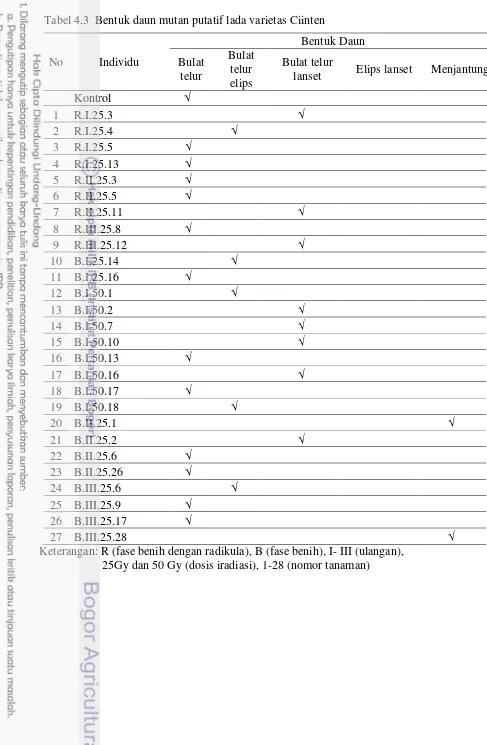
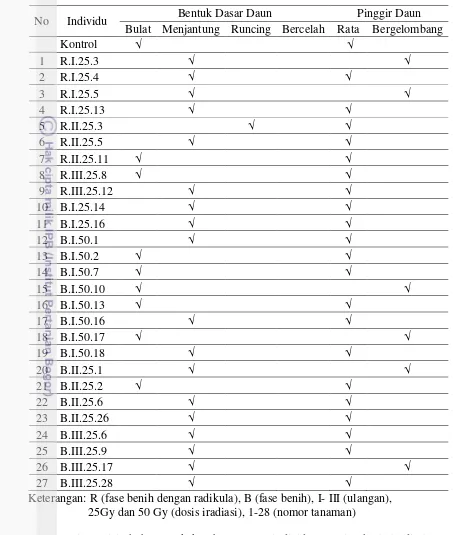
Pada percobaan II, karakter yang digunakan untuk penanda morfologi adalah tinggi tanaman, panjang daun, lebar daun, jumlah daun, jumlah ruas dan jumlah cabang. Hasil pengelompokkan berdasarkan karakter morfologi menunjukkan kesamaan antar mutan putatif sebesar 18.15%. Terdapat keragaman atau perubahan beberapa karakter daun mutan terhadap tetua. Perubahan tersebut terdapat pada bentuk daun, bentuk pangkal daun dan tepi daun. Hasil analisis SSR didapatkan lima primer yang menghasilkan pita polimorfis yaitu primer Psol10, Psol15, Psol16, Psol17, Psol18. Hasil analisis keragaman mutan putatif berdasarkan penanda SSR, memiliki tingkat kesamaan 63%.
Hasil penelitian pada percobaan III menunjukkan bahwa keragaman genetik yang tinggi penting untuk menghasilkan varietas baru, khususnya untuk pemuliaan ketahanan terhadap infeksi Phytophthora capsici. Terdapat 14 mutan yang memiliki luas bercak < 1 mm2, tetapi hanya 10 mutan yang memiliki nilai luas bercak daun yang berbeda nyata dengan kontrol. Sembilan mutan memiliki nilai luas bercak daun lebih rendah dibandingkan kontrol pada kisaran 0.17-0.60 mm2 yaitu MP11, MP16, MP17, MP18, MP19, MP20, MP23, MP25, MP26 sehingga termasuk kategori sangat tahan, sedangkan satu mutan putatif memiliki ukuran bercak daun lebih tinggi dibandingkan kontrol yaitu MP 2 (6.34 mm2) termasuk kategori sangat peka. Berdasarkan klasifikasi terdapat 14 mutan putatif bersifat sangat tahan, enam mutan putatif bersifat tahan, lima mutan putatif yang bersifat moderat tahan seperti kontrol, serta dua individu lainnya bersifat sangat peka terhadap P. capsici.
SUMMARY
NUR LAELA WAHYUNI MEILAWATI. Genetic Diversity Improvement of Black Pepper (Piper nigrum L.) Ciinten Variety through Gamma Irradiation and Selection against Phytophthora Foot Rot Diseases. Supervised by AGUS PURWITO and NURLIANI BERMAWIE.
Black pepper is an introduced species and has always been propagated vegetatively, therefore has narrow genetic diversity. One of the obstacles in the cultivation of pepper was foot rot disease caused by Phytophthora capcisi. High genetic diversity is important to produce new varieties. One way to increase the genetic diversity is through gamma ray irradiation. Genetic distance and phylogenetic relationship among accessions in black pepper germplasm will help plant breeders assemble superior varieties. This study aimed to (1) understand the effect of gamma ray irradiation on radiosensitivity and response of black pepper Ciinten variety at seed phase and seed with radicle, (2) identify genetic diversity of putative black pepper Ciinten varieties based on morphology and SSR markers, (3) select the plant of Ciinten variety resulted from gamma ray irradiation against foot rot disease.
The plant material used in experiment I were pepper seeds Ciinten variety at seed phase and seed with the radicle. The experiment used Split plot design with two factors. The main factor was treatment of seeds (seeds and seeds with radicle) and the sub plot was dosage irradiation with seven levels i.e. (0, 25, 50, 75, 100, 125, 150) Gy. Each treatment consisted of three replications, each replication consisted of 60 seeds. The research was conducted in the greenhouse of Indonesian Spice and Medicinal Crops Research Institut (ISMCRI), Molecular Plant Breeding laboratory of ISMCRI and ICABIOGRAD, Disease Laboratory of ISMCRI from September 2015 to April 2016. The plant material used in the experiments II and III were 27 individuals of black pepper plant resulted from gamma irradiation and control. Observations were made on morphological characters and SSR pattern. Isolation of DNA used CTAB methods followed by identification of purity and quantity of DNA. PCR aamplification using 9 primers. The experiment II used NTSYS program and Minitab to get dendogram. The experiment III was selection of resistance to foot rot disease by inoculated the leaves with P. capsici inoculum. The experiment used completely randomized design (CRD). Observations were made 72 hours after inoculation, by measuring the size of lesion. Data were analyzed statistically, when where was significant different from the controls would be tested further by Dunnet test at 5% level.
variation based on quantitative and qualitative morphological characters and anatomy.
The results of experiment II, the character used for morphological markers were plant height, leaf length, leaf width, leaf number, number of segments, and number of branches. Dendogram showed 18.15% similarity level. There were variations or changes in mutant leaves character compare to the origin. The changes appear in leaf shape, shape of leaf base and leaf margin. Molekular study showed that from PAGE visualization with five primers produce polymorphic bands. Those primers were Psol10, Psol15, Psol16, Psol17, Psol18. Results of analyses based on SSR markers showed 63% similarity level.
Results of the experiment III showed that high genetic diversity necessary to generate new varieties, especially for breeding resistance to Phytophthora capsici disease. There were 14 mutants that have lesion size <1 mm2, but only 10 mutants that have broad leaf lesion significantly different from control. Nine mutants have broad leaf lesion lower than the control between 0.17-0.60 mm2 that MP11, MP16, MP17, MP18, MP19, MP20, MP23, MP25, MP26 thus categorized as highly resistant, while the putative mutant has leaf lesion size higher than the control, namely MP2 (6.34 mm2) was highly sensitive category. Based on the classification there were 14 putative mutants highly resistant, six putative mutants resistant, 5 putative mutants that were moderately resistant as control, as well as two other individuals were very sensitive to P. capsici.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan IPB.
PENINGKATAN KERAGAMAN GENETIK LADA (Piper
nigrum L.) VARIETAS CIINTEN MELALUI IRADIASI SINAR
GAMMA DAN SELEKSI TERHADAP PENYAKIT BUSUK
PANGKAL BATANG (BPB)
NUR LAELA WAHYUNI MEILAWATI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Peningkatan Keragaman Genetik Lada (Piper nigrum L.) Varietas Ciinten melalui Iradiasi Sinar Gamma dan Seleksi terhadap Penyakit Busuk Pangkal Batang (BPB)
Nama : Nur Laela Wahyuni Meilawati
NIM : A253130291
Disetujui oleh Komisi Pembimbing
Dr Ir Agus Purwito, MSc Agr Dr Ir Nurliani Bermawie
Ketua Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu EK, MS Dr Ir Dahrul Syah, MSc Agr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah Subhanahu Wa Ta’ala atas
segala karunia-Nya sehingga tesis yang berjudul “Peningkatan Keragaman Genetik Lada (Piper nigrum L.) Varietas Ciinten melalui Iradiasi Sinar Gamma dan Seleksi terhadap Penyakit Busuk Pangkal Batang (BPB)” ini berhasil diselesaikan. Penulis mengucapkan terimakasih yang sebesar-besarnya kepada:
1. Dr Ir Agus Purwito, MScAgr dan Dr Nurliani Bermawie atas ide, gagasan dan ilmu yang sangat berharga yang beliau sampaikan kepada penulis sejak awal penyusunan proposal, bimbingan selama penelitian dan penulisan tesis.
2. Dr Ir Yudiwanti Wahyu EK, MS selaku ketua program studi Pemuliaan dan Bioteknologi Tanaman, Dr Dewi Sukma, SP MSi sebagai penguji sidang dan seluruh dosen, karyawan, teknisi di IPB atas ilmu dan bantuannya.
3. Dr Syarifah Iis Aisyah, MSc Agr selaku penguji luar komisi atas semua saran dan masukkannya.
4. Kepala Balai Penelitian Tanaman Rempah dan Obat yang telah memberikan izin untuk menempuh pendidikan magister di Institut Pertanian Bogor.
5. Kepala Badan Penelitian dan Pengembangan Pertanian, Kementrian Pertanian atas kesempatan belajar dan beasiswa pendidikan magister yang telah diberikan kepada penulis.
6. Dra Natalini Nova selaku ketua RPTP lada di Balittro yang telah membantu penulis untuk melaksanakan penelitian dan pendanaan melalui anggaran APBN tahun 2015.
7. Rekan-rekan di Balittro atas perhatian, doa dan dukungannya.
8. Bapak (Sutopo MM), Ibu (Rian Herini S.), Bapak mertua (M. Khusaeri), Ibu mertua (Suparni) atas semua doa dan kasih sayangnya.
9. Suami (Mochammad Rizal, SSi Apt), anak pertama (Naufa Nazihah Rufaidah), anak kedua (Muhammad Qowiyyun Alghifari) atas keikhlasannya mendoakan umi.
10.Rekan-rekan pascasarjana PBT angkatan 2013 dan 2014 atas kebersamaan dan perjuangannya.
Penulis sadari masih ada kekurangan yang perlu diperbaiki dalam tesis ini. Semoga tesis ini dapat bermanfaat untuk penelitian selanjutnya.
Bogor, Oktober 2016
DAFTAR ISI
3 RESPON TANAMAN LADA (Piper nigrum L.) VARIETAS CIINTEN TERHADAP IRADIASI SINAR GAMMA
L.) VARIETAS CIINTEN BERDASARKAN PENANDA
7 SIMPULAN UMUM 50
DAFTAR PUSTAKA 51
LAMPIRAN 60
DAFTAR TABEL
Tabel 3.1 Persentase hidup tanaman lada hasil iradiasi sinar gamma fase benih dan fase benih dengan radikula
20 Tabel 3.2 Pertumbuhan lada varietas Ciinten karakter tinggi
tanaman, panjang daun, lebar daun, tebal daun, tebal batang, jumlah daun dan jumlah ruas terhadap perlakuan dosis iradiasi sinar gamma umur 8 BST
23
Tabel 3.3 Pertumbuhan lada varietas Ciinten pada karakter jumlah cabang terhadap perlakuan dosis iradiasi sinar gamma umur 8 BST
23
Tabel 3.4 Persentase bentuk daun, bentuk dasar daun, pinggir daun lada hasil iradiasi sinar gamma
25 Tabel 3.5 Pengaruh iradiasi tanaman terhadap jumlah, kerapatan,
indeks stomata pada lada varietas Ciinten umur 8 BST
26 Tabel 4.1 Individu mutan putatif lada Ciinten yang digunakan untuk
penanda morfologi dan SSR
31 Tabel 4. 2 Primer yang digunakan untuk analisa SSR pada tanaman
lada
32 Tabel 4.3 Bentuk daun mutan putatif lada varietas Ciinten 34 Tabel 4.4 Bentuk dasar daun dan pinggir daun mutan putatif lada
varietas Ciinten
35 Tabel 4.5 Kelompok hasil dendogram berdasarkan penanda
morfologi
36 Tabel 5.1 Populasi mutan putatif lada varietas Ciinten hasil iradiasi
sinar gamma
42 Tabel 5.2 Klasifikasi luas bercak daun lada berdasarkan Xu et al.
(2014) dengan modifikasi
43 Tabel 5.3 Persentase bercak pada 27 mutan putatif Ciinten hasil
iradiasi sinar gamma dan satu tetua lada varietas Ciinten
DAFTAR GAMBAR
Gambar 1.1 Bagan alir penelitian 5
Gambar 3.1 Bentuk daun lada 1. Bulat telur, 2. Bulat telur-elips, 3. Bulat telur lonjong, 4. Elips panjang, 5. Bentuk jantung
19 Gambar 3.2 Tepi daun lada 1.lurus dan 2. Bergelombang 19 Gambar 3.3 Bentuk pangkal daun lada 1. Membulat, 2. Bentuk
jantung, 3. Runcing, 4. Tidak simetris-Bercelah
munculnya radikula terhadap beberapa dosis iradiasi sinar gamma sinar gamma dan tetua berdasarkan penanda morfologi dengan karakter tinggi tanaman, panjang daun, lebar daun, jumlah daun, jumlah ruas, jumlah cabang
36 sinar gamma dan tetua berdasarkan karakter molekuler marka SSR dengan lima primer
38
Gambar 5.1 Respon daun lada Ciinten hasil iradiasi sinar gamma (a) kontrol, (b) 25Gy, (c) 50Gy, (d) 75Gy, (e) 100Gy, (f) 25Gy terhadap infeksi Phytophthora capsici pada (b)(c)(d)(e) fase benih dan (f) fase benih dengan radikula inkubasi 72 jam
44
DAFTAR LAMPIRAN
Lampiran 1 Karakteristik Lada Varietas Ciinten 59 Lampiran 2 Hasil Scoring Penanda SSR dengan lima primer 61 Lampiran 3 Data morfologi yang digunakan untuk analisis
pengelompokkan
1 PENDAHULUAN
1.1 Latar Belakang
Lada (Piper nigrum L.) merupakan tanaman yang dimanfaatkan sebagai bumbu masakan, obat herbal, anti bakteri dan anti oksidan. Kebutuhan lada dunia mencapai 350 ribu ton/tahun. Kontribusi Indonesia sebagai pengekspor lada mencapai 29% dari kebutuhan dunia, terbesar kedua setelah Vietnam. Produksi lada nasional tahun 2014 mencapai 91.941 ton (Direktorat Jenderal Perkebunan, 2014). Salah satu kendala dalam budidaya lada adalah penyakit busuk pangkal batang (BPB) yang disebabkan oleh Phytophthora capcisi (Muller 1937). Penyakit ini dapat mematikan tanaman dan dapat menyerang seluruh bagian tanaman lada. Serangan pada daun akan menimbulkan bercak dan meluas keseluruh permukaan daun, sedangkan serangan pada pangkal batang dan akar dapat menyebabkan tanaman mati (Manohara et al. 2005). Tingkat serangan cendawan P. capsici pada tanaman lada sangat dipengaruhi oleh tingkat ketahanan tanaman, virulensi cendawan tersebut dan faktor lingkungan. Kehilangan hasil lada akibat penyakit BPB pada triwulan ketiga tahun 2010 sebesar 16 milyar rupiah (Direktorat Perlindungan Perkebunan 2011).
Upaya mengatasi penyakit BPB telah dilakukan antara lain dengan perbaikan kultur teknis dengan pemberian nutrisi sehingga meningkatkan ketahanan tanaman (Manohara et al. 2005). Pengendalian secara kimia atau agensia hayati maupun pengendalian terpadu (Wahyuno et al. 2007) juga telah dilakukan, namun upaya tersebut belum sesuai harapan. Pengendalian penyakit secara kimiawi berdampak negatif terhadap lingkungan. Salah satu alternatif untuk mengatasi masalah penyakit pada tanaman lada adalah dengan menggunakan varietas lada yang tahan. Pembentukan varietas tahan merupakan pendekatan yang efektif, ekonomis dan ramah lingkungan.
Ciinten adalah salah satu varietas unggul lada yang baru dilepas Balittro (Bermawie et al. 2015b), mempunyai malai panjang dan ukuran biji lebih besar (Setiyono & Udarno 2011), hasil per pohon, malai per tanaman, jumlah biji per malai, bobot dan panjang tangkai malai lebih tinggi dibandingkan dengan lada varietas unggul Petaling-1 (Bermawie et al. 2013). Hasil pengujian secara in vitro varietas Ciinten ini termasuk moderat tahan terhadap penyakit BPB sehingga diperlukan perbaikan varietas untuk meningkatkan karakter tersebut. Keberhasilan pembentukan varietas tahan ditentukan oleh tersedianya plasma nutfah dengan keragaman genetik yang tinggi. Salah satu cara untuk meningkatkan keragaman adalah dengan induksi mutasi (Suwarno & Silitonga 2006). Pemuliaan mutasi telah berhasil meningkatkan keragaman genetik pada tanaman yang diperbanyak dengan biji maupun vegetatif seperti kalus nilam (Kadir et al. 2007), kalus tebu (Suhesti 2015), rimpang jahe (Bermawie et al. 2015a).
membutuhkan waktu 13 hari untuk muncul radikula/ calon akar. Benih yang telah muncul radikula diduga lebih sensitif terhadap iradiasi sinar gamma, sehingga dalam penelitian ini digunakan dua fase perlakuan yaitu fase benih dan benih dengan radikula. Ini bertujuan untuk mengetahui bagian tanaman mana yang paling respon optimal dalam menangkap efek iradiasi sinar gamma.
Zanzibar & Sudrajat (2009) menjelaskan ketika radiasi ionisasi diserap ke dalam material biologis, radiasi tersebut akan beraksi secara langsung terhadap target sel kritis atau secara tidak langsung melalui pembangkitan metabolit yang dapat memodifikasi komponen-komponen sel penting. Penggunaan irasiasi sinar gamma dengan berbagai dosis dalam hubungannya dengan perkecambahan benih telah dicoba pada berbagai tanaman seperti pada Capsicum annuum L. yang telah diberi dosis rendah yaitu 2, 4, 8, dan 16 Gy merangsang pertumbuhan dan resistensi terhadap cekaman (Kim et al. 2005). Benih Triticum aestivum L. dengan pemberian dosis iradiasi 10, 20, 30 dan 40 kR menunjukkan lebih superior dibandingkan kontrol untuk beberapa karakter (Singh & Balyan 2009), dapat meningkatkan daya dan kecepatan berkecambah pada Triticum durum pada dosis
10 dan 20 Gy (Melki & Marouani 2009), dapat merangsang parameter pertumbuhan (perkecambahan, panjang akar dan hipokotil) Lactuca sativa pada dosis 5 dan 30 Gy (Marcu et al. 2012), dapat meningkatkan daya berkecambahn, indeks vigor, laju rata-rata pertumbuhan Terminalia arjuna pada dosis 25 Gy (Akshatha et al. 2013).
1.2 Perumusan Masalah
Lada merupakan tanaman introduksi dan selalu diperbanyak secara vegetatif sehingga keragaman genetiknya sempit (Purseglove et al. 1981). Bunga lada merupakan bunga hermaprodit dan bunga jantan yang terletak dalam satu pohon (andromonoecious) (Daryanto & Satifah 1984). Hasil pengamatan pada enam varietas lada menunjukkan lebih dari 94% memiliki bunga hermaprodit dan sekitar (2.6 – 5.3) % memiliki bunga hanya berkelamin jantan (Setiyono 2008). Hal ini menyebabkan terjadinya penyerbukan sendiri sehingga benih yang dihasilkan bersifat homozigot. Hal ini juga menyebabkan keragaman genetik lada rendah.
Salah satu cara peningkatan keragaman genetik suatu tanaman dapat dengan induksi mutasi (Suwarno & Silitonga 2006). Induksi mutasi menggunakan bantuan mutagen seperti mutagen fisik (radiasi) maupun mutagen kimia. Penggunaan mutagen fisik diantaranya dengan sinar gamma, sinar x, electron beam dan ion beam. Induksi mutasi menggunakan iradiasi dapat menimbulkan mutasi gen dan aberasi kromosom yang bisa menghasilkan keragaman genetik yang luas sebagai materi dasar untuk penciptaan varietas baru (Chen 2002). Respon tanaman terhadap efek radiasi sinar gamma dipengaruhi oleh laju dosis radiasi yang digunakan. Dosis yang umum digunakan adalah (40–400) Gy (Ismachin 1988).
baru, telah diterapkan pada benih dan stek berakar lada Karimunda, Paniyur 1, dan lainnya dengan dosis 1-4 krad. Dosis radiasi yang semakin tinggi berpengaruh pada penampilan tanaman, pertumbuhan tanaman yang abnormal, perubahan klorofil dan penampilan daun yang roset (Peter et al. 2007). Hadipoentyanti (2007) telah melakukan mutasi radiasi dengan sinar gamma pada benih lada varietas Natar 1 dengan dosis dosis 1, 2, 3, 4 dan 5 krad dan hasilnya menunjukkan setelah tanam di lapang, tanaman mulai berbuah pertama pada umur 4-5 tahun. Dari beberapa variasi genetik yang diperoleh di lapang terdapat satu mutan yang mempunyai karakteristik yang berbeda dibanding dengan varietas asalnya (Natar 1) perbedaan terlihat pada bentuk daun lebih sempit (jorong-lanset), rata-rata panjang bulir 13 cm, memiliki fruit set (buah jadi) lebih banyak 80% dan buah masak serentak dalam satu bulir serta buah tidak mudah rontok dari bulir walaupun telah masak (warna merah). Karakter tersebut lebih superior dibanding yang dimiliki varietas asal yaitu Natar 1.
Induksi mutasi dapat menyebabkan perubahan pada sifat morfologi, selain itu dapat merubah sifat ketahanan terhadap penyakit. Ketahanan suatu tanaman berkaitan dengan kemampuan tanaman untuk mencegah, menghambat atau memperlambat perkembangan penyakit. Salah satu mekanisme ketahanan tanaman yaitu adanya sifat fisik tanaman yang berfungsi sebagai penghalang penyakit seperti lapisan lilin, stomata, bulu daun (trichoma), lentisel yang dapat mencegah patogen untuk masuk atau menginfeksi bagian tanaman (Brown 1988). Pada penelitian ini, induksi mutasi dengan sinar gamma diharapkan akan menimbulkan perubahan genetik yang mengarah kepada ketahanan terhadap penyakit, khususnya penyakit busuk pangkal batang.
1.3 Tujuan Penelitian
1.3.1 Tujuan Umum
Meningkatkan keragaman genetik lada varietas Ciinten untuk sifat ketahanan terhadap penyakit busuk pangkal batang.
1.3.2 Tujuan Khusus
1. Mengetahui pengaruh iradiasi sinar gamma terhadap radiosensitivitas dan respon lada varietas Ciinten pada fase benih dan fase benih dengan radikula 2. Mengidentifikasi keragaman genetik lada varietas Ciinten hasil iradiasi sinar
gamma berdasarkan penanda morfologi dan SSR
3. Seleksi tanaman lada (Piper nigrum l.) varietas Ciinten hasil iradiasi sinar gamma terhadap penyakit busuk pangkal batang (BPB)
1.4 Hipotesis Penelitian
1. Perlakuan pada fase benih akan menghasilkan radiosensitivitas yang lebih rendah tetapi menghasilkan keragaman yang lebih tinggi dibandingkan dengan perlakuan pada fase benih dengan radikula
3. Diperoleh mutan putatif yang tahan terhadap P. capsici
1.5 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai dosis iradiasi lada varietas Ciinten pada fase benih dan fase benih dengan radikula yang mampu menimbulkan keragaman. LD20 dan LD50 merupakan dosis yang dapat menimbulkan mutasi pada sifat yang diinginkan sehingga dapat dimanfaatkan untuk program pemuliaan tanaman lada. Munculnya keragaman genetik pada sifat ketahanan akibat iradiasi diharapkan dapat dimanfaatkan untuk merakit varietas lada unggul tahan penyakit busuk pangkal batang (BPB).
1.6 Kerangka Penelitian
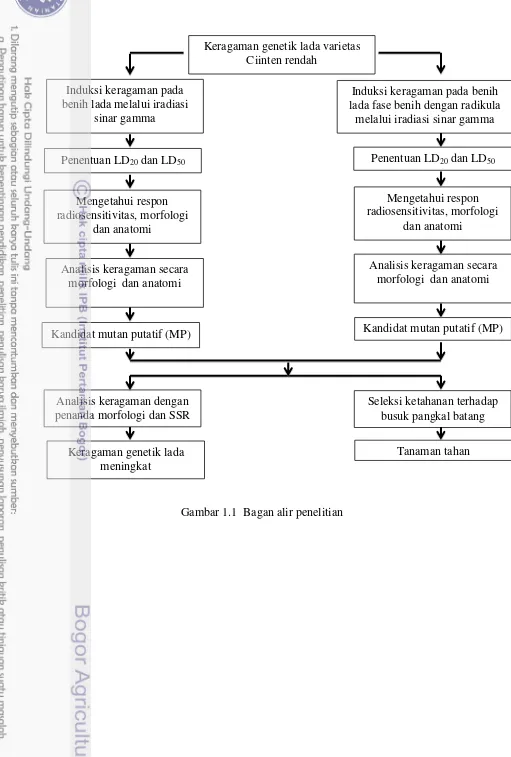
Kerangka penelitian yang dilakukan meliputi iradiasi pada fase benih dan fase benih dengan radikula, analisis respon iradiasi dengan mengamati keragaman melalui karakter morfologi, anatomi, molekuler dan uji ketahanan daun terhadap infeksi P. capsici yang menyebabkan penyakit busuk pangkal batang. Penelitian terdiri atas tiga percobaan untuk mencapai tujuan dan menjawab hipotesis dengan alur penelitian seperti Gambar 1. Lada varietas Ciinten diiradiasi dengan dosis (0, 25, 50, 75, 100, 125 dan 150) Gy. Iradiasi dilakukan terhadap dua fase yaitu fase benih dan fase benih dengan radikula. Percobaan 1 dilakukan untuk mengetahui LD20 dan LD50, mengetahui respon morfologi kuantitatif dan kualitatif dan respon karakter anatomi (stomata). Diharapkan terjadi peningkatan keragaman lada di antara LD20 dan LD50.
Gambar 1.1 Bagan alir penelitian
Keragaman genetik lada varietas Ciinten rendah
Induksi keragaman pada benih lada melalui iradiasi
sinar gamma
Penentuan LD20 dan LD50
Mengetahui respon radiosensitivitas, morfologi
dan anatomi
Analisis keragaman secara morfologi dan anatomi
Kandidat mutan putatif (MP)
Analisis keragaman dengan penanda morfologi dan SSR
Keragaman genetik lada meningkat
Induksi keragaman pada benih lada fase benih dengan radikula
melalui iradiasi sinar gamma
Penentuan LD20 dan LD50
Mengetahui respon radiosensitivitas, morfologi
dan anatomi
Kandidat mutan putatif (MP)
Seleksi ketahanan terhadap busuk pangkal batang
Tanaman tahan Analisis keragaman secara
2 TINJAUAN PUSTAKA
2.1 Sistematika Tumbuhan
Klasifikasi tanaman lada menurut taksonomi secara rinci adalah sebagai berikut:
Divisi : Spermatophyta Subdivisi : Angiospermae Class : Dicotyledoneae Ordo : Piperales Familia : Piperaceae Genus : Piper
Spesies : Piper nigrum L.
Genus Piper ditemukan oleh Linnaeus dan memiliki banyak spesies. Sekitar 600–2.000 spesies di antaranya tersebar di daerah tropis. Dari jumlah tersebut, terdapat beberapa spesies yang telah dibudidayakan (Rukmana 2003) dan memiliki nilai ekonomi tinggi, antara lain lada (Piper nigrum L). Beberapa varietas lada yang dikembangkan di Indonesia antara lain Petaling 1, Petaling 2, Natar 1, Natar 2, Chunuk, Lampung Daun Kecil (LDK) (Nuryani et al. 1993), Bengkayang, Malonan1 (Rostiana et al. 2015) dan Ciinten (Bermawie et al. 2015b).
2.2 Syarat Tumbuh
Lada merupakan jenis tanaman tropis sehingga hanya dapat dikembangkan di daerah tropis. Faktor yang berpengaruh pada pertumbuhan dan perkembangan lada harus diketahui supaya berhasil dalam pengembangannya. Persyaratan tumbuh yang cocok untuk tanaman lada adalah sebagai berikut :
2.2.1 Iklim
Tanaman lada dapat tumbuh dan berproduksi dengan baik pada daerah yang memiliki tipe iklim A, B dan C. Menurut Schmidt & Ferguson, tipe A merupakan iklim amat basah (0–1.5 bulan kering), tipe B merupakan iklim basah (1.5–3 bulan kering) dan tipe C iklim agak basah (3-4.5 bulan kering). Dengan Curah hujan 2.000-3.000 mm/tahun. Suhu udara 20oC (minimum) hingga 34oC (maksimum) dengan kisaran suhu terbaik antara 20-27oC pada pagi hari, 26-32oC pada siang hari dan 24-30oC pada sore hari, dan kelembaban udara 50% - 100% (Syakir 2002).
2.2.2 Lahan
pertumbuhan dan produktivitas lada dipengaruhi oleh kedalaman air tanah (Rismunandar & Riski 2007).
2.3 Deskripsi Tanaman Lada
Lada secara morfologi tergolong tanaman dimorfik yang memiliki dua macam sulur yaitu sulur panjat (Orthotropic climbing shoot) dan sulur buah (Axillary plagiotropic fruiting branches), sedangkan pada lada perdu hanya memiliki sulur buah saja. Berdasarkan morfologinya perbedaan yang jelas antara sulur panjat dan sulur buah yaitu sulur panjat memiliki akar lekat (hold fast), sedangkan sulur buah tidak memilikinya. Sulur panjat secara fisiologi memiliki sifat negatif fototrof, sedangkan sulur buah bersifat fototrof (Syakir 2002).
2.3.1 Akar
Lada termasuk anggota tanaman dikotil. Benihnya akan tumbuh membentuk akar lembaga dan berkembang menjadi akar tunggang. Akar lada akan terbentuk pada buku-buku di ruas batang pokok dan cabang. Berdasarkan peranannya, akar lada dibagi menjadi dua jenis walaupun pada dasarnya hanya satu jenis. Kedua akar tersebut ialah akar yang tumbuh dari buku didalam tanah dan di atas tanah. Akar yang tumbuh dari buku didalam tanah akan membentuk akar lateral dan berfungsi sebagai pengisap zat makanan (feeding roots). Akar yang tumbuh dari buku di atas tanah berfungsi sebagai pelekat untuk menopang batang pokok dan menjalar pada tiang atau pohon penunjang. Akar lateral dengan akar serabut yang tebalnya sekitar 30 cm berada di dalam lapisan tanah bagian atas (top soil) dan dapat masuk kedalam tanah 1–2 meter. Jumlah akar lateral rata-rata 10–20 buah dengan panjang 3–4 meter, tergantung kesuburan tanah. Perakaran lada sangat sensitif terhadap genangan air yang berkepanjangan (Wahid 1996). Menurut Bermawie et al. (2015b) akar lada pada varietas Ciinten memiliki jumlah akar banyak dan daya lekat tinggi.
2.3.2 Batang
Tanaman lada memiliki satu batang pokok dengan dua macam cabang (dimorphicy). Cabang tersebut ialah cabang orthotropis (vertikal) dan cabang plagiotropis (horisontal). Cabang orthotropis tumbuh membentuk kerangka dasar pohon lada hingga berdiameter 4 - 6 cm, mengayu dan beruas dengan panjang rata-rata 5 - 12 cm. Cabang plagiotropis dengan akar pelekat terbentuk dari buku antar ruas yang pertumbuhannya agak membengkak. Dari buku tersebut tumbuh sehelai daun dan kuntum yang selanjutnya tumbuh menjadi cabang. Kedua jenis cabang tersebut akan membentuk percabangan (Wahid 1996). Bermawie et al. (2015b) menyatakan bahwa lada Ciinten memiliki batang tua berwarna coklat, warna batang muda berwarna hijau, panjang ruas (7.63 1.32) cm, percabangan polimorfik, diameter ruas (10.00 3.56) mm, memiliki sulur gantung serta sulur cacing yang sedikit.
2.3.3 Daun
dan bagian bawah pucuk dengan titik-titik kelenjar. Berdasarkan letak tumbuhnya bentuk daun lada beraneka ragam, daun pada batang bagian atas berbeda dengan daun pada batang bagian bawah (Wahid 1996). Warna daun tua pada lada varietas Ciinten yaitu Hijau tua YGG 147 A, warna daun muda yaitu Hijau muda YGG 145 A, dan warna seludang yaitu hijau kemerahan (Bermawie et al. 2015b).
2.3.4 Bunga
Bunga (organum reproductivum) berbentuk malai, agak menggelantung, panjang malai bervariasi dari 3-25 cm, tergantung varietas. Panjang malai varietas Ciinten sebesar (11.441.11) cm, bobot malai masak (10.912.01) gram. Malai tidak bercabang, berporos tunggal, dan terdapat sekitar 150 bunga kecil. Malai tumbuh berhadapan dengan daun dari cabang atau ranting plagiotropis. Bunga lada dapat berupa uniseksual, yaitu monoecious (berumah satu) dan dioecious (berumah dua). Monoecious yaitu pada satu tanaman terbentuk bunga betina dan bunga jantan secara terpisah. Bila bunga jantan dan bunga betina berada dalam satu bunga sehingga disebut tanaman hermaphrodit. Tanaman dioecious yaitu masing-masing bunga jantan dan bunga betina berada terpisah pada pohon yang berlainan. Bunga lada tumbuh dalam ketiak, kelopak berdaging, tidak bermahkota, benang sari sebanyak 2 - 4 helai, berukuran panjang 1 mm, dan terletak di kanan dan kiri bakal buah (Wahid 1996). Bunga lada varietas Ciinten memiliki warna malai krem kehijauan YGG 149 A dan arah malai yang menggantung (Bermawie et al. 2015b).
2.3.5 Buah
Buah lada tidak bertangkai, berbenih tunggal, berbentuk bulat, berdiameter 4–6 mm, dan berdaging. Buah lada berukuran rata-rata 3-4 mm. Embrionya sangat kecil. Berat 100 benih lada sekitar 3-8 gram dengan rata-rata berat normal buah 4.5 gram (Wahid 1996). Jumlah buah per malai bervariasi, pada varietas lada Ciinten 79.2314.47, ukuran buahnya besar dan aromanya kuat. Berat 1000 butir buah segar (g) sebesar 155.2 9.66 gram. Kulit buah lada berwarna hijau saat masih muda dan akan berubah warna menjadi kuning orange atau merah setelah masak. Buah yang berkulit hijau akan menjadi kehitaman setelah dijemur dibawah terik sinar matahari, dan disebut lada hitam. Buah yang dipanen setelah masak, direndam kemudian dikupas kulitnya lalu dikeringkan di bawah sinar matahari, dan disebut lada putih (Bermawie et al. 2015b).
2.4 Pemuliaan Mutasi
misalnya pada tunas, biji, dan bagian tanaman yang lain. Secara molekuler mutasi terjadi karena adanya perubahan urutan (sequence) nukleotida DNA kromosom yang menyebabkan terjadinya perubahan pada protein yang dihasilkan (Poespodarsono 1988).
Mutasi dapat terjadi secara alami (spontan) dan secara buatan/induksi. Mutasi alami berasal dari radioaktif dan sinar kosmik serta pengaruh temperatur. Peluang terjadi mutasi alami di alam sangat kecil yaitu sekitar 10-7-10-6 (IAEA 1997). Mutasi induksi merupakan mutasi buatan yang terjadi melalui campur tangan manusia. Mutasi ini dilakukan untuk meningkatkan peluang terjadinya mutasi sehingga menghasilkan perubahan sifat yang diinginkan. Agen atau bahan yang menyebabkan terjadinya mutasi induksi disebut dengan mutagen. Mekanisme mutasi induksi yaitu mutagen merusak DNA inti kemudian selama proses perbaikan, perubahan terjadi secara acak dan dapat diwariskan pada generasi berikutnya. Mutagen terdiri dari mutagen fisik dan mutagen kimia (Simmonds 1979). Poespodarsono (1988) mengelompokkan mutagen dalam tiga golongan yaitu mutagen kimia, mutagen fisik iradiasi dan mutagen non radiasi.
2.4.1 Mutagen Kimia
Mutagen kimia merupakan salah satu penyebab mutasi pada makhluk hidup yang berasal dari senyawa kimia seperti etil metan sulfonat (EMS), dietil sulfat (DES), metil metan sulfonat (MMS), natrium azida (Talebi et al. 2012). Mutagen ini umumnya menghasilkan mutasi induksi yang menyebabkan substitusi pasangan basa yang mengakibatkan perubahan asam amino. Perubahan ini mengubah fungsi protein tetapi tidak menghapus fungsinya.
Bidabadi et al. (2012) menyatakan bahwa perlakukan berbagai dosis (150, 200 dan 250 mM) dan periode waktu (30 dan 60 menit) dari EMS pada tunas pucuk dapat meningkatkan variasi pada pisang (Musa spp.). Hasil analisis menunjukkan bahwa rata-rata jumlah tunas per eksplan, kelangsungan hidup (%), dan bobot segar tunas pucuk dipengaruhi oleh dosis EMS dan memberikan perbedaan yang signifikan pada jangka waktu perlakuan. Perlakuan 60 menit/250 mM dan 30 menit/200 mM menunjukkan peningkatan maksimal dalam produksi eksplan aktif. Persentase variasi fenotipe yang berasal menunjukkan peningkatan sebesar (10-14)% setelah tiga bulan kultur, dengan perlakuan 60 menit/200 mM dan 30 menit/250 mM perlakuan EMS dibandingkan dengan kontrol. Talebi et al. (2012) menyatakan nilai LD25 dan LD50 diamati berdasarkan pada pengurangan pertumbuhan benih setelah perlakuan konsentrasi EMS 0.25% dan 0.50% untuk variasi padi MR219.
antara lain tetrasomik, triploid, tetraploid, pentaploid, heksaploid, dan oktaploid (Ariyanto et al. 2011).
2.4.2 Mutagen Fisik
Peran teknologi nuklir sangat besar dalam pemuliaan tanaman karena dapat menghasilkan varietas unggul baru. Teknologi nuklir memiliki kemampuan dalam menginduksi mutasi pada materi genetik. Teknik ini diduga memiliki energi cukup tinggi untuk menimbulkan perubahan pada struktur atau komposisi materi genetik (Soeranto 2003). Iradiasi gamma pada umumnya digunakan untuk induksi mutasi pada benih dan bahan perbanyakan vegetatif tanaman. Pemuliaan mutasi pada tanaman yang diperbanyak secara vegetatif lebih efektif karena dapat mengubah satu atau beberapa karakter tanpa mengubah karakteristik kultivar asalnya (Nagatomi 1996).
Iradiasi pada tanaman dapat menimbulkan abnormalitas karena telah terjadi perubahan pada tingkat genom, kromosom, dan DNA sehingga menghasilkan variasi genetik. Keragaman genetik dapat terjadi apabila suatu tanaman diberikan perlakuan dosis pada tingkat tertentu. Tingkat keberhasilan iradiasi dalam meningkatkan keragaman sangat ditentukan oleh radiosensitivitas tanaman yang diiradiasi, karena tingkat radiosensitivitas tanaman bervariasi. Radiosensitivitas dapat diukur berdasarkan nilai LD50 (lethal dose 50) yaitu tingkat dosis yang menyebabkan kematian 50% dari populasi tanaman yang diiradiasi (Shu 2009) atau penghambatan tumbuh 50 %.
Kematian sel dapat terjadi secara langsung karena kerusakan DNA serta tidak langsung karena adanya pengaruh toksik dan radikal bebas ion H2O2 dan OH– yang dihasilkan dari radiolisis air. Material yang paling banyak mengalami iradiasi adalah air yang kemudian terurai menjadi H2O+. Reaksi selanjutnya akan membentuk radikal bebas yang kemudian bergabung dengan peroksida, apabila keduanya bereaksi pada molekul lain maka akan membentuk senyawa yang akan mempengaruhi sistem biologi tanaman (Borzouei 2013).
Oksigen merupakan faktor yang paling penting dalam menyebabkan kerusakan pada jaringan tanaman yang diradiasi sinar gamma. Keberadaannya bersama sama dengan air dalam bahan tanaman akan menghasilkan kerusakan biologis dan genetik pada sel tanaman. Broetjes & Harten (1988); Alpen (1994) menyatakan bahwa semakin banyaknya oksigen dan molekul air (H2O) berada dalam materi yang diradiasi, maka semakin banyak radikal bebas yang terbentuk. Pengaruh radiokimia yang disebabkan oleh radiasi ionisasi adalah terbentuknya ion radikal positif dan elektron bebas sebagai berikut:
H2O H2O+ (ion) +e -H2O H+ + OHo e e-aq
e-aq Ho + OH
Ho + O2 HO2o
Jika terdapat oksigen, maka Ho atau e-aq akan membentuk radikal HO2o. Hydrogen peroksidase (H2O2) dan OH- merupakan agen pengoksida yang pada reaksi kimia berikutnya dengan molekul-molekul lebih besar akan merusak fungsi sel. Selain itu juga terbentuk radikal bebas sepertiHo yaitu ion yang labil sehingga banyak menghasilkan benturan ke berbagai arah, yang akibatnya akan membuat perubahan atau mutasi di tingkat DNA, tingkat sel, maupun jaringan, bahkan sampai kematian pada tanaman (Ahnstroem 1977; Datta 2001).
Iradiasi sinar gamma telah banyak digunakan pada berbagai tanaman dalam rangka peningkatan keragaman genetik untuk toleransi cekaman abiotik dan biotik serta peningkatan kuantitas dan kualitas hasil, diantaranya: tomat (Ishfaq et al. 2012), wortel (Nagananda et al. 2013), kentang (Ahmad et al. 2010), kedelai (Alify et al. 2013), padi (Shanthi et al. 2010), sorghum (Soeranto & Sihono 2010), dan gandum (Singh & Balyan 2009; Borzouei et al. 2010; Plamenov et al. 2013).
2.5 Radiosensitivitas
Radiosensitivitas adalah tingkat sensitivitas tanaman terhadap iradiasi (Harten 1998; Datta 2001). Keberhasilan radiasi untuk meningkatkan keragaman populasi sangat ditentukan oleh radiosensitivitas tanaman (Banerji & Datta 1992). Beberapa hal yang mempengaruhi radiosensitivitas, di dalam IAEA (1977) disebutkan adanya dua faktor utama yang mempengaruhi radiosensitivitas yaitu faktor lingkungan dan faktor biologi. Faktor lain yang juga mempengaruhi radiosensitivitas seperti genotipe, bagian tanaman yang digunakan dengan memberikan perlakuan terhadap organ reproduksi tanaman seperti benih, stek batang, serbuk sari, akar rhizome, kultur jaringan dan sebagainya, stadia perkembangan sel tanaman, jumlah kromosom, umur jaringan, oksigen, temperatur, penyimpanan pasca iradiasi dan dosis iradiasi. Nilai radiosensitivitas tanaman juga berkaitan erat dengan kandungan air di dalam sel tanaman (Herison et al. 2008).
Tingkat sensitivitas secara visual ini dapat diamati dari respon yang diberikan tanaman secara morfologi tanaman, sterilitas maupun dosis lethal 50 (LD50). LD50 adalah dosis yang menyebabkan kematian 50% dari populasi yag diiradiasi. Dalam menentukan LD50 atau LD20 dapat menggunakan program best-fitting curve. Harten (1998) menyatakan bahwa pemberian dosis iradiasi disekitar dosis letal (LD20-LD50) dapat meningkatkan keragaman genetik. Dosis yang digunakan untuk menginduksi keragaman sangat menentukan keberhasilan terbentuknya tanaman mutan. Dosis iradiasi yang digunakan dipengaruhi oleh jenis tanaman yang digunakan, fase tumbuh saat tanaman diiradiasi, ukuran bahan tanam dan tingkat ketebalan bahan yang akan diiradiasi (Shu et al. 2012).
2.6 Marka Genetik
penanda jenis ini adalah pengamatannya mudah dan langsung dapat dilihat dengan mata, tetapi memiliki kelemahan karena dipengaruhi oleh tahap perkembangan tanaman dan lingkungan. Jumlah marka morfologi sangat terbatas sehingga kadang sulit membedakan antar genotipe yang diamati, secara morfologi kelihatan sama tetapi sebenarnya berbeda akibat adanya interaksi intra dan inter gen.
Marka biokimia, seperti isozim merupakan marka yang sangat efektif, pewarisannya bersifat kodominan, sehingga dapat membedakan individu yang homosigot dan heterosigot (McDonald & McDermont 1993) dan dapat dapat diaplikasikan dengan mudah dan murah (Bermawie & Pool 1991; Mondini et al. 2009). Marka isozim dapat digunakan untuk menganalisis keragaman genetik populasi maupun mengidentifikasi perbedaan genetik antar aksesi (Crawford 1990), tetapi marka isozim jumlahnya terbatas, ekspresinya dipengaruhi oleh lingkungan dan tahap perkembangan tanaman (McDonald & McDermont 1993; Mangolin et al. 1997; Garkava et al. 2000) serta tingkat polimorfisme yang relatif rendah.
Marka molekuler sering kali dikenal sebagai sidik jari DNA karena mengacu pada pita polimorfisme berupa fargmen DNA. Keunggulan utama penanda molekuler adalah (i) keakuratan yang tinggi dan tidak dipengaruhi oleh lingkungan yang mempengaruhi ekspresi gen, (ii) dapat diuji pada semua tingkat perkembangan tanaman, (iii) pada pengujian ketahanan hama dan penyakit tidak tergantung pada organisme pengganggu tersebut, (iv) seleksi pada tingkat genotipe ini dapat mempercepat proses seleksi dan hemat pada pengujian selanjutnya di lapangan (Kasim et al. 2002).
Marka molekuler (DNA) yang ideal memiliki kriteria sebagai berikut: a) memiliki tingkat polimorfisme yang sedang sampai tinggi, b) terdistribusi merata diseluruh genom, c) memberikan resolusi perbedaan genetik yang cukup, d) pewarisan bersifat kodominan (dapat membedakan kondisi homozigot dan heterozigot dalam organisme diploid), e) berprilaku netral, f) secara teknik sederhana, cepat dan murah, g) butuh sedikit jaringan dan DNA sampel, h) berkaitan erat dengan fenotipe, i) tidak memerlukan informasi tentang genom organisme, dan j) data mudah dipertukarkan antar laboratorium (Mondini et al. 2009; Agarwal et al. 2008; Weising et al. 2005).
Marka molekuler DNA tersebut dapat dikelompokkan menjadi dua yaitu, penanda DNA tanpa PCR (non-PCR based techniques) seperti RFLP, dan penanda DNA berdasarkan PCR yang meliputi RAPD, AFLP, SSR, CAPS, SCAR, SSCP dan DNA Barkoding (Zulfahmi 2013). Pemulia menggunakan marka DNA untuk identifikasi keragaman plasma nutfah, identifikasi genotipe, galur, kultivar dan varietas untuk melihat kemurnian benih, memecahkan ketidakpastian tetua, penelusuran tetua dan juga untuk melindungi varietas tanaman yang dikembangkan melalui identifikasi individu.
2.6.1 Marka SSR
(Hamada et al. 1982). Teknik PCR pada SSR hanya menggunakan DNA dalam jumlah kecil dengan daerah amplifikasi yang kecil, sekitar 100-300 bp (basepair) dari genom. SSR memiliki kelebihan yang dapat diandalkan, dapat diulang dan biaya yang kompetitif apabila dibandingkan dengan marka yang lain (Singh et al. 2007). Marka SSR dapat diamplikasikan tanpa merusak bahan tanaman karena hanya sedikit saja dalam ekstraksi DNA atau dapat menggunakan bagian tanaman lain, seperti biji dan serbuk sari. Pertimbangan lain adalah marka SSR terdistribusi secara melimpah dan merata dalam genom, variabilitasnya sangat tinggi (banyak alel dalam lokus) dan sifatnya kodominan dengan lokasi genom yang telah diketahui (Zulfahmi 2013).
Keunggulan lain dari SSR ini adalah produk PCR dapat langsung difraksinasi menggunakan elektroforesis, baik pada gel akrilamida maupun agarose. Tetapi gel akrilamid memiliki keunggulan karena memiliki ukuran pori yang kecil, sehingga mampu mendetekasi alel yang memiliki tingkat polimorfis rendah dan alel per lokus sekalipun susunan basanya berbeda 2 bp, sedangkan gel agarose membutuhkan kuantitas DNA lebih banyak, dan membutuhkan susunan yang basa lebih besar. Sekuen SSR pendek maka secara efisien dapat diamplifikasi menggunakan PCR dengan sekuen pengapitnya sebagai primer. Panjang primer yang digunakan biasanya berkisar antara 18-25 bp. Tingkat polimorfismenya biasanya tergantung pada variasi jumlah pengulangan unik spesifik dalam lokus mikrosatelit yang berevolusi lebih cepat dibandingkan dengan DNA disekitarnya, sehingga menjadikannya sangat polimorfik (Zulfahmi 2013). Variasi jumlah ulangan mikrosatelit dapat dideteksi menggunakan elektroforesis hasil amplifikasi produk DNA pada suatu gel dengan standar sekuen yang memisahkan fragmen dengan perbedaan setara dengan satu nukleotida. Perkembangan yang cepat untuk sejumlah penanda molekuler genetik yang didukung oleh praktek-praktek dalam pemuliaan tanaman menjadikan penanda molekuler lebih efektif dibandingkan dengan fenotipiknya (Singh et al. 2007).
Kemudahan SSR dalam mengamplifikasi dan mendeteksi fragmen-fragmen DNA serta tingginya tingkat polimorfisme yang dihasilkan menyebabkan metode ini ideal untuk dipakai dalam studi genetik. Salah satu contoh SSR dapat diaplikasikan untuk mempelajari keragaman genetik, identifikasi plasma nutfah dan studi evolusi serta identifikasi kultivar, pengujian progeny serta gene tagging. Powel et al. (1996) mengemukakan bahwa SSR telah dikarakterisasi pada banyak spesies tanaman meliputi jagung, padi, kedelai, tomat, barley dan brassica. Tingkat polimorfis yang tinggi terlihat pada kelapa sawit, dengan amplifikasi berdasarkan PCR.
2.7 Hama dan Penyakit pada Lada
2.7.1 Hama
sehingga menyebabkan gejala layu dan mati sebagian. Serangga dewasa menyerang bagian tanaman seperti pucuk, bunga dan buah sehingga dapat menurunkan kualitas dan kuantitas produksi. Serangan pada tingkat serangan berat dapat menyebabkan kematian tanaman. Berbagai stadium penggerek batang selalu ditemukan pada saat yang sama berupa telur, larva, pupa atau imago. Pada awal musim hujan biasanya ditemukan telur dan larva muda. Pertengahan musim hujan ditemukan pupa dan imago, sedangkan pada akhir musim hujan ditemukan telur dan larva dan pada musim kemarau, semua stadium jumlahnya sangat rendah (Deciyanto & Suprapto 1996).
Hama penghisap bunga (Diconocoris hewetti) atau nyamuk lada/ enduk-enduk/ kapal terbang/ fui kichong (Bangka). Stadia nimfa maupun dewasa merusak bunga dan tandan bunga. Serangan ringan menyebabkan tandan rusak, salah bentuk dan buah yang terbentuk hanya sedikit. Serangan berat dapat menyebabkan seluruh bunga rusak, tangkai bunga menjadi hitam dan gugur sebelum waktunya. Hama ini juga memakan buah muda. Perkembangan D. hewetti
dipengaruhi oleh varietas lada. Secara umum D. hewetti lebih berpotensi sebagai
hama pada lada varietas Lampung Daun Lebar (LDL) dibandingkan dengan Chunuk Hal ini ditunjukkan oleh masa perkembangan pradewasa yang lebih singkat (13 hari), keperidian yang lebih banyak (24.5 butir), serta laju pertambahan intrinsik yang lebih tinggi (0.0827) pada varietas LDL dibandingkan dengan varietas Chunuk (Laba 2005).
Hama penghisap buah (Dasynus piperis)/ kepik/ kepinding/ walang sangit/ semunyung (Bangka)/ bilahu (Belitung, Kalimantan). Stadia nimfa maupun serangga dewasa menghisap cairan buah, menyebabkan buah menjadi hampa/ kosong, kering/ busuk kemudian gugur. Serangan pada buah muda (umur 4-5 bulan) menyebabkan butiran buah gugur sebelum tua, sehingga tandan banyak yang ksosng. Serangan pada buah tua (umur 6-9 bulan) menyebabkan buah kering Serangga dewasa kurang lebih bisa hidup 3 bulan di lapang (IPC 2011). Setiap stadium D. piperis dapat dijumpai secara bersamaan di lapangan dan menyebar pada tajuk tanaman. Hal tersebut menunjukkan bahwa serangga selalu dijumpai sepanjang tahun pada tanaman lada. Namun demikian populasinya bergantung pada musim buah (Deciyanto 1991).
2.7.2 Penyakit
Penyakit busuk pangka batang (BPB) yang disebabkan Phytophthora capsici, merupakan penyakit utama pada lada. Di Indonesia, gejala BPB pertama kali dilaporkan pada tahun 1885, dan diidentifikasi disebabkan oleh P. palmivora var. piperis (Muller 1937). Phytophthora mudah terbawa air, tanah atau bagian tanaman yang terserang sehingga jamur patogen tersebut kemungkinan terdapat pada daerah pengembangan lada. Phytophthora telah ditemukan hampir di semua pertanaman lada di Indonesia. Populasi Phytophthora memiliki virulensi yang bervariasi terhadap tanaman lada budi daya maupun lada liar (Wahyuno et al. 2007b; 2010) sehingga perlu dipertimbangkan dalam mendapatkan varietas lada tahan BPB.
tanaman menunjukkan serangan lebih lanjut. Pangkal batang yang terserang menjadi berwarna hitam, pada keadaan lembab akan mengeluarkan lender berwarna biru muda. Serangan pada akar, menyebabkan tanaman layu dan daun menguning (IPC 2011).
Penyakit utama lainnya adalah seperti penyakit kuning, dan kerdil/ keriting. Penyakit kuning mematikan pertanaman lada terutama di Bangka dan Kalimantan. Penyakit ini disebabkan adanya serangan nematoda (Radhopholus similis dan Meloidogyne incognita), jamur (Fusarium oxysporum), kesuburan tanah yang rendah, serta rendahnya kelembaban tanah atau kadar air tanah. Luka-luka akibat serangan nematoda akan memudahkan terjadinya infeksi jamur F. oxysporum. Nematoda menyerang akar lada dengan menusuk dan menghisap cairan sehingga terjadi pelukaan akar. Luka tersebut dimasuki jamur dan menyebabkan matinya jaringan pembuluh akar sehingga peredaran air dan unsur hara terganggu (Direktorat Jenderal Bina Produksi Perkebunan 2002).
3
RESPON TANAMAN LADA (Piper nigrum L.) VARIETASCIINTEN TERHADAP IRADIASI SINAR GAMMA
Abstrak
Lada merupakan tanaman introduksi dan selalu diperbanyak secara vegetatif, sehingga keragaman genetiknya sempit. Keragaman genetik yang tinggi penting untuk menghasilkan varietas baru. Salah satu cara untuk meningkatkan keragaman genetik adalah melalui iradiasi sinar gamma. Penelitian ini bertujuan untuk mengetahui respon lada varietas Ciinten pada fase benih dan fase benih dengan radikula terhadap iradiasi sinar gamma. Rancangan percobaan menggunakan Rancangan Split plot dengan petak utama fase benih (benih dan benih dengan radikula) dan anak petak dosis iradiasi dengan tujuh taraf yaitu (0, 25, 50, 75, 100, 125, 150) Gy. Masing-masing perlakuan terdiri dari tiga ulangan, setiap ulangan terdiri dari 60 benih. Kedua fase menghasilkan keragaan pada karakter tinggi tanaman, panjang daun, lebar daun, tebal daun, tebal batang, jumlah daun dan jumlah ruas yang menunjukkan perbedaan secara signifikan antar dosis. Radiosensitivitas lada pada fase benih dengan radikula lebih tinggi dibandingkan dengan fase benih ditunjukkan oleh nilai LD50 (Lethal Dose 50). LD50 pada lada fase benih yaitu 68.15 Gy, sedangkan LD50 fase benih dengan radikula yaitu 30 Gy. Tingginya dosis iradiasi yang diberikan pada kedua fase perlakuan, mengakibatkan tinggi tanaman, panjang daun, lebar daun, tebal daun, tebal batang semakin terhambat pertumbuhannya sehingga jumlah daun dan jumlah ruas semakin sedikit. Dosis iradiasi 25 dan 50 Gy pada fase benih dan 25 Gy pada fase benih dengan radikula nyata meningkatkan keragaman genetik berdasarkan karakter morfologi kuantitatif, morfologi kualitatif dan anatomi. Kata kunci: radiosensitivitas, iradiasi, radikula, anatomi, morfologi
Abstract
Black pepper is an introduced species and has always been propagated vegetatively, so it has narrow genetic variability. High genetic variation is necessary to produce new varieties. Increasing genetic variation can be done through gamma ray irradiation. This research aims to evaluate response of black pepper Ciinten variety at seed and radicle emergence phases to gamma irradiation. The experimental design used was split plot with the main factor was seed phases (seed and seed with radicle) and the sub plot was dose of irradiation with seven levels (0, 25, 50, 75, 100, 125, 150) Gy. Each treatment consisted of three replications, each replication consisted of 60 seeds. Both phases showed significant differences in perfomances among dose in plant height, leaf length, leaf width, leaf thick, stem thick, number of leave, number of internode. Radiosensitivity of pepper on radicle emergence phase was higher than the seed phase indicated by LD50 (Lethal Dose 50). LD50 at seed phase was 68.15 Gy,
whereas LD50 of the radicle emergence phase was 30 Gy. The higher irradiation
significantly increasde genetic variability based on quantitative and qualitative morphological characters and anatomy.
Keywords: radiosensitivity, irradiation, radicle, anatomy, morphology
3.1 Pendahuluan
Lada (Piper nigrum L.) merupakan tanaman yang buahnya berfungsi sebagai bumbu masakan, obat herbal, anti bakteri dan anti oksidan. Kebutuhan lada dunia mencapai 350 ribu ton/tahun. Kontribusi Indonesia sebagai pengekspor lada mencapai 29% dari kebutuhan dunia, terbesar kedua setelah Vietnam (IPC 2013). Produksi lada nasional tahun 2014 mencapai 91.941 ton (Direktorat Jenderal Perkebunan 2014). Lada pada umumnya diperbanyak dengan cara vegetatif sehingga memiliki keragaman genetik yang sempit. Keragaman genetik yang tinggi diperlukan untuk menghasilkan varietas baru. Salah satu cara untuk meningkatkan keragaman genetik adalah dengan induksi mutasi (Suwarno & Silitonga 2006). Induksi mutasi dapat dilakukan menggunakan mutagen fisik maupun kimiawi (Chopra 2005). Mutagen fisik dengan iradiasi sinar gamma lebih banyak digunakan karena memiliki energi dan daya tembus tinggi, memiliki frekuensi dan spektrum iradiasi dan tergantung pada dosis dan laju dosis yang digunakan.
Radiosensitivitas merupakan tingkat sensitivitas tanaman terhadap radiasi (Harten 1998). Uji radiosensitivitas dilakukan untuk mendapatkan dosis iradiasi yang efektif menghasilkan mutan dan mengetahui frekuensi serta spektrum mutasi (Abdullah et al. 2009). Sensitivitas pada radiasi dapat diukur berdasarkan nilai LD (Lethal dose) yaitu dosis yang dapat menyebabkan kematian dari populasi tanaman yang diiradiasi. Variabilitas mutan tertinggi terdapat pada mutan hasil iradiasi sinar gamma antara LD20 dan LD50 (Soeranto 2003). Nilai LD didapatkan dengan menggunakan program analisis statistik untuk mencari persamaan model terbaik. Bagian tanaman mana yang paling respon terhadap dosis iradiasi dapat diketahui melalui fase pertumbuhan yang optimal dalam menangkap efek iradiasi, sehingga penelitian ini menggunakan dua fase pertumbuhan yaitu fase benih dan fase benih dengan radikula.
radikula), oleh sebab itu digunakan dua perlakuan fase benih (benih dan benih dengan radikula). Penelitian ini bertujuan untuk menentukan radiosensitivitas dan mengetahui respon lada varietas Ciinten pada fase benih dan fase munculnya radikula terhadap iradiasi sinar gamma.
3.2 Bahan dan Metode
3.2.1 Waktu dan Tempat
Penelitian ini dilaksanakan dari bulan Juni 2014 sampai bulan April 2015. di Pusat Aplikasi Isotop dan Radiasi (PAIR) BATAN, Jakarta Selatan. Pengamatan morfologi dilakukan di rumah kaca Balittro serta pengamatan stomata dilakukan di Laboratorium Mikroteknik IPB, Darmaga.
3.2.2 Bahan dan Alat
Bahan tanaman yang digunakan adalah biji lada varietas Ciinten pada fase benih dan fase benih dengan radikula. Alat yang digunakan yaitu alat saprodi pertanian, tanah, pupuk, polibag.
3.2.3 Metodologi Penelitian
Fase benih yaitu biji lada yang berwarna merah, direndam dalam air selama dua jam, kemudian dipisahkan antara kulit dan bijinya. Fase benih dengan radikula yaitu biji lada yang sudah terpisah dari kulitnya, lalu diletakkan pada cawan petri yang telah dialasi dengan kertas saring lembab. Fase benih dengan radikula yang terseleksi yaitu benih yang memiliki radikula dengan ukuran sekitar 0.1-1cm pada hari ke-12 hingga hari ke-15. Kedua fase diberi perlakuan dosis iradiasi sinar gamma. Penelitian menggunakan Split Plot dengan dua faktor. Petak utama yaitu fase benih (benih dan benih dengan radikula) dan anak petak yaitu dosis iradiasi dengan tujuh taraf yaitu (0, 25, 50, 75, 100, 125, 150) Gy. Masing-masing perlakuan terdiri dari tiga ulangan, setiap ulangan terdiri dari 60 benih. Benih hasil iradiasi sinar gamma pada kedua fase masing-masing ditanam dengan media pasir di dalam bak persemaian, setelah benih tumbuh dan memiliki 3-4 daun, tanaman lada dipindah ke dalam polibag dengan media tanah dan pupuk kandang (2:1) yang bertujuan untuk merangsang pertumbuhan tanaman.
3.2.3.1 Respon Radiosensitivitasterhadap Iradiasi Sinar Gamma
Radiosensitivitas yaitu sensitivitas suatu materi genetik terhadap radiasi. Ini dapat diukur berdasarkan nilai LD (Lethal Dose) yaitu dosis yang menyebabkan kematian dari populasi tanaman yang di iradiasi. Dosis optimum yang dapat menghasilkan mutan terbanyak umumnya diperoleh di sekitar dosis lethal (Datta 2001). Harten (1998) menyatakan bahwa pemberian dosis iradiasi disekitar dosis letal (LD20-LD50) dapat meningkatkan keragaman genetik. Radiosensitivitas dihitung berdasarkan persentase tanaman yang hidup 80% (LD20) dan persentase tanaman hidup 50% (LD50) dengan menggunakan program curve fit analysis (Soeranto 2003).
3.2.3.2 Respon Karakter Morfologi terhadap Iradiasi Sinar Gamma
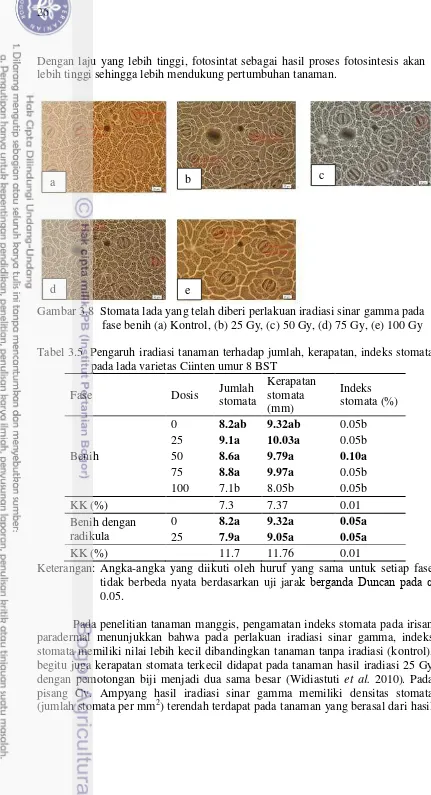
panjang daun (cm), lebar daun (cm), tebal daun, tebal batang, jumlah daun dan jumlah ruas saat tanaman umur 8 BST (Bulan Setelah Tanam). Pengamatan dilakukan pada seluruh tanaman lada yang hidup setelah diberi perlakuan iradiasi pada masing-masing dosis. Data dianalisis uji F pada taraf nyata 5% dengan menggunakan program SAS, bila hasilnya berbeda nyata maka akan diuji lanjut dengan uji DMRT (Duncan’s Multiple Range Test-DMRT). Karakter kualitatif yang diamati yaitu bentuk daun, bentuk pangkal daun, bentuk tepi daun yang diamati berdasarkan IPGRI (1995) (Gambar 3.1- 3.3), sedangkan warna daun yang diamati dengan menggunakan Colour Chart Royal Horticultural Society (RHS 2007).
Gambar 3.1 Bentuk daun lada 1. Bulat telur, 2. Bulat telur-elips, 3. Bulat telur lonjong, 4. Elips panjang, 5. Bentuk jantung
Gambar 3.2 Tepi daun lada 1. Lurus dan 2. Bergelombang
Gambar 3.3 Bentuk pangkal daun lada 1. Membulat, 2. Bentuk jantung, 3. Runcing, 4. Tidak simetris-Bercelah
3.2.3.3 Respon Karakter Anatomi (Stomata) terhadap Iradiasi Sinar Gamma
Stomata diamati pada masing-masing dosis dengan 10 sampel individu setiap dosis pada daun ketiga dan keempat. Sampel stomata menggunakan metode preparat awetan. Permukaan bawah daun diolesi dengan cat kuku, setelah mengering lekatkan pada selotip bening lalu dikelupas dan diletakkan di atas gelas objek. Pengamatan dengan mikroskop pada bidang pandang perbesaran 40x. Pengamatan meliputi banyaknya stomata, panjang dan lebar stomata, kerapatan stomata serta indeks stomata. Setiap sampel preparat diamati tiga bidang pandang. Kerapatan Stomata = Jumlah stomata
Indeks Stomata = Jumlah stomata x 100% Jumlah stomata + Jumlah sel epidermis
Respon karakter anatomi terhadap dosis iradiasi dianalisis ragam dengan uji F pada taraf nyata 5% dengan menggunakan program SAS, bila hasilnya berbeda nyata maka akan diuji lanjut dengan uji DMRT (Duncan’s Multiple Range Test -DMRT).
3.3 Hasil dan Pembahasan
3.3.1 Radiosensitivitas
Perlakuan dosis iradiasi menunjukkan bahwa semakin tinggi dosis yang diberikan mengakibatkan persentase hidup tanaman lada semakin rendah, baik pada fase benih maupun fase radikula. Persentase hidup tanaman lada yang diberi perlakuan dosis 25 Gy pada fase benih sebesar 85 %, menurun pada dosis 50 Gy menjadi 70 % dan menurun tajam pada dosis 100 Gy menjadi 1.7 %. Persentase hidup fase radikula pada dosis 25 Gy sebesar 80.2%, dan menurun tajam hingga 3.1% pada dosis 50 dan 75 Gy, hingga tidak ada yang tumbuh pada dosis 100 Gy (Tabel 3.1). Hal ini kemungkinan bahwa dosis > 100 Gy mengakibatkan terjadinya kerusakan DNA pada tanaman lada yang menyebabkan tanaman mati.
Tabel 3.1 Persentase hidup tanaman lada hasil iradiasi sinar gamma pada fase benih dan fase benih dengan radikula
Dosis (Gy)
Persentase tumbuh (%)
Fase Benih Fase benih dengan radikula
0 100 100
25 85.0 80.2
50 70.0 3.1
75 31.7 3.1
100 27.3 -
125 1.7 -
150 1.7 -
Keterangan: - tanaman tidak tumbuh
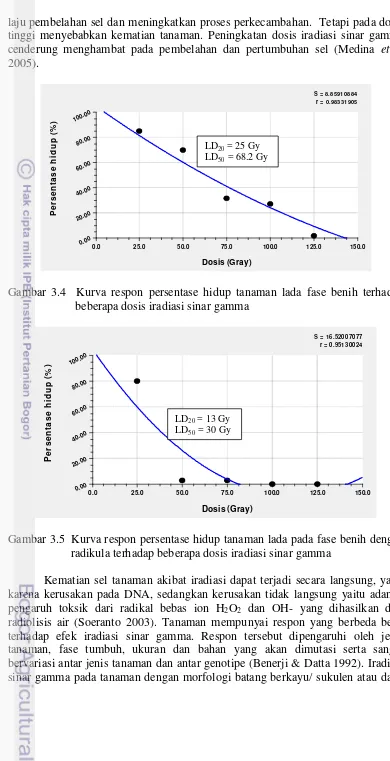
laju pembelahan sel dan meningkatkan proses perkecambahan. Tetapi pada dosis tinggi menyebabkan kematian tanaman. Peningkatan dosis iradiasi sinar gamma cenderung menghambat pada pembelahan dan pertumbuhan sel (Medina et al 2005).
Gambar 3.4 Kurva respon persentase hidup tanaman lada fase benih terhadap beberapa dosis iradiasi sinar gamma
Gambar 3.5 Kurva respon persentase hidup tanaman lada pada fase benih dengan radikula terhadap beberapa dosis iradiasi sinar gamma
Kematian sel tanaman akibat iradiasi dapat terjadi secara langsung, yaitu karena kerusakan pada DNA, sedangkan kerusakan tidak langsung yaitu adanya pengaruh toksik dari radikal bebas ion H2O2 dan OH- yang dihasilkan dari radiolisis air (Soeranto 2003). Tanaman mempunyai respon yang berbeda beda terhadap efek iradiasi sinar gamma. Respon tersebut dipengaruhi oleh jenis tanaman, fase tumbuh, ukuran dan bahan yang akan dimutasi serta sangat bervariasi antar jenis tanaman dan antar genotipe (Benerji & Datta 1992). Iradiasi sinar gamma pada tanaman dengan morfologi batang berkayu/ sukulen atau daun
sukulen/ tidak sukulen dapat menghasilkan respon yang berbeda. Respon tanaman terhadap iradiasi sinar gamma juga berhubungan dengan faktor biologis lainnya seperti faktor genetika, dan juga faktor lingkungan seperti oksigen, kadar air, penyimpanan pasca iradiasi dan suhu (Ahnstroem 1977).
Radiosensitivitas yaitu sensitivitas suatu materi genetik terhadap iradiasi, diukur berdasarkan nilai LD (Lethal Dose) yaitu dosis yang menyebabkan kematian dari populasi tanaman yang diiradiasi. Dosis yang menghasilkan persentase hidup tanaman 80% (LD20) dan 50% (LD50) dihitung dengan menggunakan rumus best fitting curve, danmenghasilkan rumus persamaan kurva Quadratic Fit yaitu y = a+bx+cx2 (y = 59.346 + 0.136x – 0.00040x2) untuk fase benih memiliki nilai LD20 = 25 Gy dan LD50 = 68.2 Gy, sedangkan untuk fase benih dengan radikula menghasilkan persamaan Quadratic Fit y = a+bx+cx2 (y = 59.462 + 0.924x – 0.0034x2) memiliki kisaran LD20 = 13 Gy dan LD50 = 30 Gy. Huruf y merupakan persentase hidup (%), sedangkan huruf x merupakan dosis iradiasi sinar gamma yang diberikan.
Radiosensitivitas pada kedua fase yang berbeda disebabkan pada fase benih dengan radikula memiliki kadar air lebih tinggi. Ini dikarenakan terjadi imbibisi hingga munculnya radikula saat diletakkan dalam media kertas saring pada cawan petri, dibandingkan dengan kadar air pada fase benih. Sejalan dengan penelitian Chan (2009) pada benih pepaya yang diimbibisi jauh lebih sensitif terhadap iradiasi (LD50 = 50-87 Gy) dibandingkan benih pepaya kering (LD50 > 300 Gy).
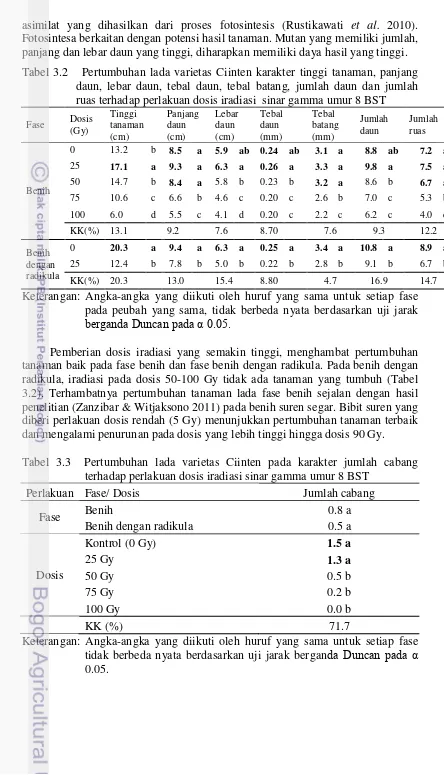
3.3.2 Karakter Morfologi Kuantitatif
Hasil analisis ragam dengan rancangan split plot menunjukkan adanya interaksi pada karakter tinggi tanaman, panjang daun, lebar daun, tebal daun, tebal batang, jumlah daun dan jumlah ruas. Pada karakter jumlah cabang tidak ada interaksi antara perlakuan benih dan dosis. Dosis 25 Gy pada fase benih dapat meningkatkan tinggi tanaman, panjang daun, lebar daun, tebal daun, tebal batang, jumlah daun dan jumlah ruas, sedangkan dosis 50 Gy memiliki panjang daun, tebal batang dan jumlah ruas tidak berbeda nyata dengan kontrol dan dosis 25 Gy, tetapi berbeda nyata dengan perlakuan dosis (75-100) Gy (Tabel 3.2).
Purnamaningsih et al. (2011) menyatakan bahwa tanaman Artemisia yang telah diberi perlakuan iradiasi sinar gamma menghasilkan galur-galur mutan yang beragam pada karakter tinggi tanaman, bentuk daun dan umur berbunga. Demikian juga penelitian Suhesti (2015) pada kalus tebu menunjukkan semakin tinggi dosis iradiasi yang diberikan berdampak pada penurunan kemampuan tumbuh/ viabilitas tanaman tebu, tinggi tanaman dan jumlah daun. Penelitian Taheri et al. (2014) pada Curcuma alismatifolia menunjukkan pemberian iradiasi dengan dosis 20 Gy menurunkan secara signifikan jumlah daun semua varietas dibandingkan kontrol.
asimilat yang dihasilkan dari proses fotosintesis (Rustikawati et al. 2010). Fotosintesa berkaitan dengan potensi hasil tanaman. Mutan yang memiliki jumlah, panjang dan lebar daun yang tinggi, diharapkan memiliki daya hasil yang tinggi. Tabel 3.2 Pertumbuhan lada varietas Ciinten karakter tinggi tanaman, panjang
daun, lebar daun, tebal daun, tebal batang, jumlah daun dan jumlah ruas terhadap perlakuan dosis iradiasi sinar gamma umur 8 BST
Fase Dosis
Pemberian dosis iradiasi yang semakin tinggi, menghambat pertumbuhan tanaman baik pada fase benih dan fase benih dengan radikula. Pada benih dengan radikula, iradiasi pada dosis 50-100 Gy tidak ada tanaman yang tumbuh (Tabel 3.2). Terhambatnya pertumbuhan tanaman lada fase benih sejalan dengan hasil penelitian (Zanzibar & Witjaksono 2011) pada benih suren segar. Bibit suren yang diberi perlakuan dosis rendah (5 Gy) menunjukkan pertumbuhan tanaman terbaik dan mengalami penurunan pada dosis yang lebih tinggi hingga dosis 90 Gy. Tabel 3.3 Pertumbuhan lada varietas Ciinten pada karakter jumlah cabang
terhadap perlakuan dosis iradiasi sinar gamma umur 8 BST
Perlakuan Fase/ Dosis Jumlah cabang