PENAMBAHAN
ASCORBYL PHOSPHATE
MAGNESIUM
SEBAGAI SUMBER VITAMIN C DAN
IMPLANTASI DENGAN ESTRADIOL-17
β
HENGKY JULIUS SINJAL
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi dengan judul “ Kajian
Penampilan Reproduksi Induk Ikan Lele (
Clarias gariepinus
) Betina melalui
Penambahan
Ascorbyl Phosphate Magnesium
sebagai Sumber Vitamin C pada Pakan
dan Implantasi Estradiol-17
β
”
adalah karya saya sendiri dengan arahan komisi
pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi di
mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan
maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan
dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Agustus 2007
Hengky Julius Sinjal
gariepinus
) betina melalui Penambahan
Ascorbyl Phosphate Magnesium
Sebagai
Sumber Vitamin C pada Pakan dan Implantasi Estradiol-17
β
. Komisi
Pembimbing: M. Zairin Jr, R. Affandi, B. Purwantara, dan W. Manalu.
Penelitian ini dilakukan untuk menentukan pengaruh kombinasi
ascorbyl
phosphate magnesium
sebagai sumber vitamin C dan implantasi estradiol-17
β
pada
kecepatan pematangan gonad, kualitas telur, dan larva ikan lele (
Clarias gariepinus
).
Dalam percobaan ini ikan diberikan beberapa kombinasi perlakuan yang terdiri atas 2
faktor, yaitu penambahan
ascorbyl phosphate magnesium
pada pakan dengan dosis
0, 600, 1200, dan 1800 mg/kg pakan dan implantasi estradiol 17-
β
dengan dosis 00,
250, dan 500
μ
g/kg induk ikan, sehingga jumlah kombinasi perlakuan adalah 12.
Delapan belas ekor induk dengan berat 250–328 gr ditebar ke dalam jaring apung
(3X2X2m) untuk satu perlakuan sehingga terdapat 12 buah jaring apung dengan total
induk ikan berjumlah 216 ekor. Pakan diberikan dua kali sehari secara
at satiation
.
Pemeriksaan kematangan gonad dilakukan dua minggu sekali dengan
menggunakan metode kanulasi, bersamaan waktunya dilakukan pengambilan sampel
darah dengan menggunakan spuit 2.5 ml yang berheparin untuk keperluan analisis
kandungan estradiol plasma darah. Induk yang telah matang gonad diinjeksi dengan
ovaprim sebanyak 2 kali untuk memacu ovulasi. Telur-telur yang dihasilkan oleh
induk betina ditetaskan di akuarium, kemudian dihitung daya tetas telurnya, dan
persentase larva abnormal yang dihasilkan induk tersebut. Ketahanan larva diuji
dengan memelihara larva sebanyak 100 ekor tanpa diberi makan. Larva diamati setiap
hari dan mortalitasnya dicatat. Jika larva yang hidup tinggal 20 ekor (20%) maka
pengamatan larva diakhiri. Kandungan vitamin C, protein, fosfolipid, lemak, prolin,
dan hidroksiprolin dianalisis pada ovarium, telur, larva 0 hari, dan larva 2 hari.
Berdasarkan evaluasi kombinasi penambahan
ascorbyl phosphate magnesium
dan implantasi estradiol pada induk ikan lele memperlihatkan respons nyata pada
kinerja penampilan reproduksi yang meliputi kecepatan pematangan gonad, indeks
gonad somatik, fekunditas relatif, daya tetas telur, ketahanan hidup larva, dan larva
abnormal yang dihasilkan. Kombinasi yang terbaik untuk mempercepat pematangan
gonad dan memperbaiki kualitas telur dan larva adalah kombinasi penambahan
ascorbyl phosphate magnesium
1200 mg/kg pada pakan dan implantasi estradiol-17
β
dengan dosis 250
μ
g/kg. Dengan semakin meningkatnya penambahan
ascorbyl
phosphate magnesium
pada pakan menghasilkan peningkatan kandungan vitamin C
pada telur sehingga menghasilkan ketahanan hidup larva yang tinggi dan mengurangi
larva abnormal.
Catfish (
Clarias gariepinus)
with Supplemeted by
Ascorbyl Phosphate
Magnesium
as a Source of Vitamin C in Diets and Implanted with
Estradiol-17
β
. Advisors: M. Zairin Jr, R. Affandi, B. Purwantara, dan W. Manalu.
The experiment was conducted to determine the effect of dietary
ascorbyl
phospahate magnesium
as a source of vitamin C and implanted with estradiol-17
β
on the gonad maturation, egg and larva quality of African Catfish
Clarias
gariepinus
. Fish were treated by various combinations of dietary dosage of
ascorbyl phosphate magnesium (0, 600, 1200, and 1800 mg/kg of feed) and
estradiol-17
β
(00, 250, and 500
μ
g/kg). Two hundreds sixteen and eighteen pairs of
broodstock fish were used for this experiment. Fish were fed with the experimental
diets two times a day
at satiation
. The gonad somatic index, egg diameter, fecundity,
hatching rate of the eggs, survival rate, and percentage of abnormal larvae were
determined. Results of the experiment indicated that supplementation of ascorbyl
phosphate magnesium and estradiol-17
β
stimulated gonad development and increased
hatching rate, fecundity and survival rate, and reduced percentage of abnormal larvae.
Combination of ascorbyl phosphate magnesium 1200 mg/kg feed and estradiol-17
β
250
μ
g/kg gave the best reproductive performance.
Key word :
African Catfish
Clarias gariepinus, Ascorbyl phosphate magnesium,
estradiol-17
β
, reproduction performance
Hak cipta milik Institut Pertanian Bogor tahun 2007
Hak cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencamtumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian,
penulisan karya ilmiah, penyusunan laporan, penulisan kritik
atau tinjauan suatu masalah
PENAMBAHAN
ASCORBYL PHOSPHATE
MAGNESIUM
SEBAGAI SUMBER VITAMIN C DAN
IMPLANTASI DENGAN ESTRADIOL-17
β
HENGKY JULIUS SINJAL
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Departemen Budidaya Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
implantasi dengan estradiol-17
β
Nama
: Hengky Julius Sinjal
Nomor Pokok
: C061020041
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Muhammad Zairin Jr., M.Sc. Dr. Ir. Ridwan Affandi, DEA
Ketua
Anggota
Dr. drh. Bambang Purwantara, M.Sc Prof. Dr. Ir. Wasmen Manalu
Anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Ilmu Perairan
Prof. Dr. Ir. H. Enang Harris, MS. Prof. Dr. Ir. Khairil Anwar Notodiputro, MS.
karena hanya dengan berkat anugerah-Nya penulis dapat menyelesaikan penulisan
disertasi ini. Peningkatan produksi perikanan dan peningkatan kualitas dan kuantitas
benih ikan merupakan
issue
yang mendasari penelitian ini, dengan judul ” Kajian
Penampilan Reproduksi Induk Ikan Lele (
Clarias gariepinus
) Betina Melalui
Penambahan
Ascorbyl Phosphate Magnesium
Sebagai Sumber Vitamin C pada Pakan
dan Implantasi Estradiol-17
β
”
Pada kesempatan ini penulis menyampaikan ucapan terima kasih kepada Bapak
Prof. Dr. M. Zairin Jr., M.Sc sebagai ketua komisi pembimbing, Bapak Dr. Ridwan
Affandi, DEA dan Bapak Dr. Drh. Bambang Purwantara, M.Sc, Bapak Prof. Dr. Ir.
Wasmen Manalu sebagai anggota komisi pembimbing yang telah memberikan
nasihat, petunjuk dan bimbingan yang sangat berharga selama proses penelitian
sampai dengan penyelesaian studi. Terima kasih kepada Prof. Dr. Enang Harris, MS.
Ketua Program Studi Ilmu Perairan dan DR. Chairul Muluk mantan Ketua Program
Studi Ilmu Perairan yang terus memberikan motivasi dan nasehat agar penulis cepat
menyelesaikan studi.
Terima kasih dan penghargaan disampaikan kepada pimpinan Universitas Sam
Ratulangi, dan Fakultas Perikanan dan Ilmu Kelautan, Manado yang telah
memberikan kesempatan kepada penulis untuk melanjutkan pendidikan. Terima kasih
dan penghargaan juga disampaikan kepada pimpinan Institut Pertanian Bogor dan
Sekolah Pascasarjana, yang telah menerima penulis untuk belajar dan menyelesaikan
studi di lembaga ini.
Pada kesempatan ini saya sampaikan ucapan terima kasih kepada Departemen
Pendidikan Nasional Republik Indonesia, pemerintaha daerah Sulawesi Utara, dan
yayasan mandiri atas bantuan dana penelitian dan penulisan disertasi.
Terima kasih juga disampaikan kepada Kepala Balai Riset Perikanan Budidaya
Air Tawar, Departemen Kelautan dan Perikanan yang telah mengizinkan penulis
bantuan tenaga teknis lapangan selama saya melakukan penelitian.
Banyak pihak yang telah membantu dan mendukung penulis selama proses
perkuliahan, penelitian sampai penulisan disertasi; oleh karena itu ucapan terima
kasih disampaikan kepada: staf administrasi dan perpustakaan Departemen Budidaya
Perairan: Yuli, Hani, Asep, dan
Dessy
; teknisi laboratorium: Ibu Lina, Bapak Wasjan,
Bapak Ranta. Teman-teman di Program Studi Ilmu Perairan: Suradi, Gunarto
Lantama, Surya Darwisito, Bambang utomo, Yulfiperius, dan Adharto Utiah;
Rekan-rekan di Bogor: Anderson Kumenaung, Orbanus Naharia, Nurdin Jusuf, Ridwan
Lasabuda, Agung Windarto, Alfret Luasunaung, Adnan Wantasen, Edwin Ngangi,
Donata Pandin, Tommy Lolowang, Hasnawaty, Joice Rimper, Jack Mamangkey,
Deny Karwur, Erly Kaligis, Hengky Manoppo, Teo Lasut, Ari Mirah, dan Yosep
Karamoy.
Terima kasih kepada Ayahanda Hendrik Petrus Sinjal (Alm) dan Ibunda
Eleonora Lumintang, adik-adik, kakak-kakak, adik-adik ipar dan kakak-kakak ipar,
atas bantuan doa dan dana dalam penyelesaian studi penulis.
Secara khusus dengan segala ketulusan hati, saya sampaikan terima kasih
kepada istriku dan anakku yang tercinta Joshua Nirai Okinawa Sinjal dan Hideyuki
William Michael Sinjal yang merupakan sumber inspirasi saya dan dengan penuh
pengertian, kesabaran, dan rela ditinggal lama tanpa didampingi secara fisik oleh
penulis sebagai seorang ayah.
Penulis menyadari bahwa apa yang penulis lakukan masih belum memadai
karena keterbatasan waktu dan dana. Oleh karena itu kritik dan saran penyempurnaan
selanjutnya penulis sangat dihargai. Diharapkan apa yang penulis lakukan dapat
bermanfaat dalam memacu perkembangan teknologi pembenihan ikan di Indonesia.
Bogor, Agustus 2007
Penulis dilahirkan di Manado pada tanggal 19 Juli 1958 sebagai anak dari
pasangan Bapak Hendrik Petrus Sinjal (Alm) dan Ibu Eleonora Lumintang. Penulis
menyelesaikan pendidikan sekolah dasar di Manado pada tahun 1970, sedangkan
pendidikan menengah tingkat pertama dan menengah tingkat atas jurusan IPA
diselesaikan masing-masing pada tahun 1973 dan 1976.
Pendidikan sarjana ditempuh pada Program Studi Budidaya Perairan, Fakultas
Perikanan Universitas Sam Ratulangi, Manado masuk pada tahun 1977 dan selesai
pada tahun 1985. Pada tahun 1991 penulis melanjutkan studi ke program Master
Science pada Program Studi Marine Biology, University of the Ryukyus Okinawa
Jepang dan selesai pada tahun 1993. Sejak tahun 1986 penulis telah bekerja sebagai
staf pengajar pada Fakultas Perikanan dan Ilmu Kelautan, Universitas Sam Ratulangi,
Manado. Pada tahun 1995 penulis menikah dengan Ir. Jeane Rimber Indy MSc dan
dikaruniai dua orang anak Joshua Okinawa Nirai Sinjal dan Hideyuki Michael
William Sinjal.
Pada tahun 2002 penulis diberi kesempatan mengikuti program doktor pada
Program Studi Ilmu Perairan, Sekolah Pascasarjana Institut Pertanian Bogor, dengan
bantuan dana beasiswa pendidikan pascasarjana yang diperoleh dari Departeman
DAFTAR GAMBAR...
DAFTAR LAMPIRAN...
xi
xii
I
PENDAHULUAN...
1
Latar Belakang... ...
Perumusan Masalah...
Tujuan dan Manfaat...
Hipotesis...
1
4
6
6
II
TINJAUAN PUSTAKA... 7
Kematangan Gonad Ikan...
Vitelogenesis...
Peranan Vitamin C pada Reproduksi Ikan...
Peranan Estradiol-17
β
pada Reproduksi Ikan...
Kualitas Telur Ikan...
Kerangka Teoretis...
7
10
12
15
17
19
III
METODE PENELITIAN... 22
Waktu dan Tempat Penelitian...
Bahan dan Alat...
Metode Penelitian...
Pelaksanaan Penelitian...
Analisis Data...
22
22
24
26
30
IV
HASIL DAN PEMBAHASAN... 31
Hasil...
Kecepatan Pematangan Gonad, Indeks Gonad Somatik, Diameter
Telur...
Fekunditas, Daya Tetas Telur, Ketahanan Hidup Larva, dan
Keabnormalan Larva...
Kadar Estradiol-17
β
dalam Plasma Darah...
Kandungan Vitamin C Ovarium, Telur, dan Larva...
Kandungan Protein Telur, dan Larva...
Kandungan Lemak, Telur dan Larva...
Rasio Hidroksiprolin/Prolin Ovarium dan Larva...
Pembahasan...
31
31
37
41
43
46
47
48
50
V
KESIMPULAN DAN SARAN...
69
Tabel Halaman
1
Komposisi pakan dengan penambahan
ascorbyl phosphate magnesium
0,
600, 1200, dan 1800 mg/kg pakan...
23
2 Komposisi proksimat pakan dengan penambahan
ascorbyl phosphate
magnesium
0, 600, 1200, dan 1800 mg/kg pakan...
24
3
Perlakuan berbagai kombinasi penambahan
ascorbyl phosphate magnesium
dan implantasi estradiol-17
β
...
24
4
Kualitas air selama percobaan...
27
5
Nilai rataan indeks kematangan gonad, diameter telur, fekunditas, derajat
tetas telur, ketahanan hidup larva dan larva abnormal pada ikan lele yang
diberikan berbagai kombinasi
ascorbyl phosphate magnesium
dan
1
Proses vitelogenesis pada ikan (Aida et al., 1991)...
16
2 Wadah penelitian yang terbuat dari bambu dengan 12 jaring apung dengan
ukuran 3 X 2 X 2 m yang di atasnya ditutupi dengan jaring...
23
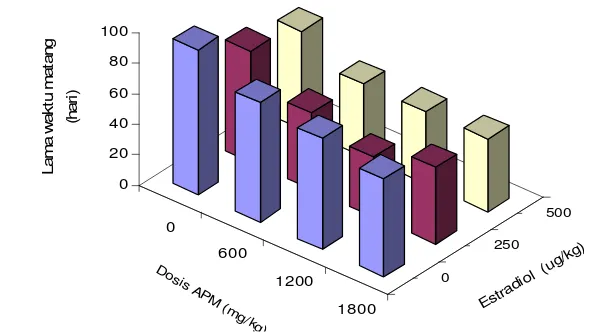
3 Nilai rataan lama waktu matang ikan lele yang diberi berbagai kombinasi
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
31
4 Nilai rataan indeks gonad somatik ikan lele yang diberi berbagai
kombinasi
ascorbyl phosphate magnesium
(APM) dan hormon
estradiol-17
β
...
34
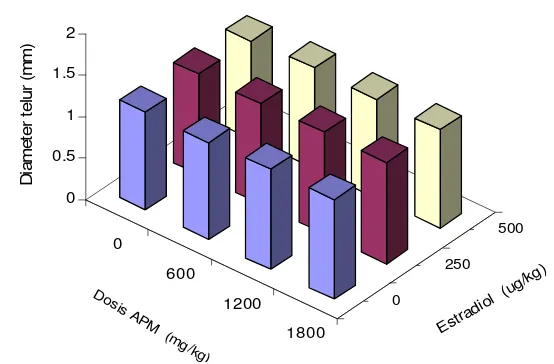
5 Nilai rataan diameter telur ikan lele yang diberi berbagai kombinasi
ascorbyl phosphate magnesium
(APM) dan hormon estradiol-17
β
...
35
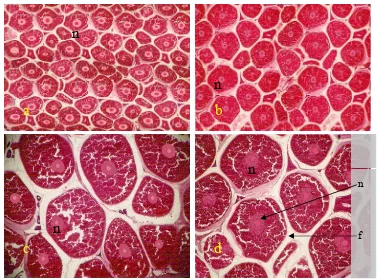
6 Struktur histologi gonad ikan lele pada kombinasi dosis
ascorbyl
phosphate magnesium
00 mg/kg dan hormon estradiol-17
β
00
μ
g/ml...
36
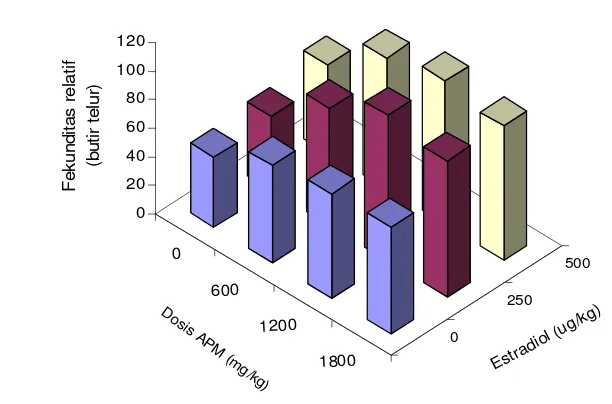
7 Nilai
rataan
fekunditas relatif ikan lele yang diberi berbagai
kombinasi
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
37
8 Nilai rataan daya tetas telur ikan lele yang diberi berbagai kombinasi
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...;...
38
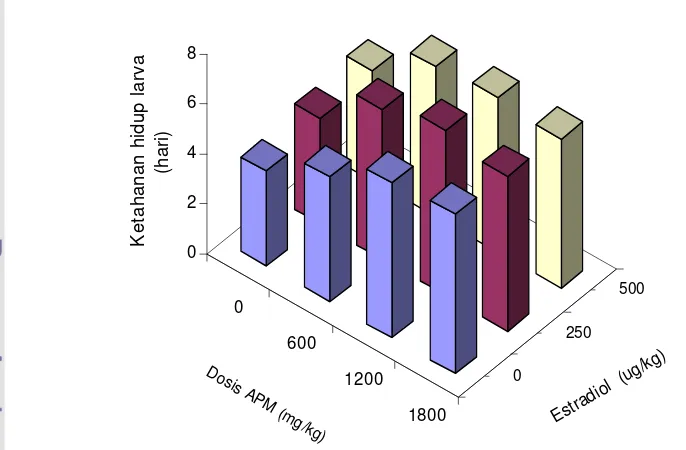
9 Nilai rataan ketahanan hidup larva ikan lele yang diberi berbagai
kombinasi
ascorbyl phosphate magnesium
(APM) dan
estradiol-17
β
...
39
10 Nilai rataan larva abnormal ikan lele yang diberi berbagai kombinasi
ascorbyl phosphate magnesium
(APM) dan estradiol 17
β
...
40
11 Gambaran morfologis larva: normal (a) abnormal (b, c, dan, d) dari hasil
penetasan induk ikan lele (
Clarias gariepinus
)...
41
12 Kadar
estradiol-17
β
plasma darah ikan lele pada berbagai kombinasi
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
selama
percobaan...
42
13 Nilai kandungan vitamin C pada ovarium, telur, larva 0 hari, dan larva 2
hari ikan lele yang diberi berbagai kombinasi
ascorbyl phosphate
magnesium
(APM) dan estradiol-17
β
...
43
14 Nilai kandungan protein pada telur, larva 0 hari, dan larva 2 hari ikan lele
yang diberi berbagai kombinasi
ascorbyl phosphate magnesium
(APM)
dan estradiol-17
β
...
45
15 Nilai kandungan vitamin C ovarium ikan lele pada hari ke 0, 42, dan 98
yang diberi berbagai kombinasi
ascorbyl phosphate magnesium
(APM)
17 Nilai rasio hidroksiprolin/prolin (HP/P) pada telur, larva 0 hari, dan larva
2 hari ikan lele yang diberi berbagai kombinasi
ascorbyl phosphate
Lampiran
Halaman
1 Komposisi
vitamin mix
dan
mineral mix
………... 79
2
Cara pembuatan pelet hormon estradiol-17
β
...…... 80
3
Prosedur penyiapan preparat histologi gonad... 81
4 Prosedur
radioimmunoassai……….. 84
5 Prosedur
analisis
lipida nonpolar (Takeuchi, 1988)…..………..……
85
6
Prosedur analisis vitamin C dengan alat HPLC (Schuep et al. 1994).
86
7
Nilai lama waktu matang ikan lele yang diberi berbagai kombinasi
dosis
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
87
8
Nilai indeks gonad somatik ikan lele yang diberi berbagai kombinasi
dosis
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
87
9
Nilai diameter telur ikan lele yang diberi berbagai kombinasi dosis
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
88
10 Nilai fekunditas ikan lele yang diberi berbagai kombinasi dosis
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
88
11
Nilai derajat tetas telur ikan lele yang diberi berbagai kombinasi dosis
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
89
12
Nilai ketahanan hidup larva ikan lele yang diberi berbagai kombinasi
dosis
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
89
13 Nilai larva abnormal ikan lele yang diberi berbagai kombinasi dosis
ascorbyl phosphate magnesium
(APM) dan hormon estradiol-17
β
...
90
14 Kandungan vitamin C ovarium ikan lele pada hari ke 0, 42, dan 98
yang diberi berbagai kombinasi dosis
ascorbyl phosphate magnesium
(APM) dan estradiol-17
β
...
90
15
Nilai rataan kadar estradiol (ng/ml) plasma darah ikan lele yang diberi
berbagai kombinasi
ascorbyl phosphate magnesium
(APM) dan
estradiol yang diambil setiap dua minggu selama 98 hari...
91
16 Kandungan vitamin C pada ovarium, telur, larva 0, dan larva 2 hari
ikan lele yang diberi berbagai kombinasi
ascorbyl phosphate
magnesium
(AMP) dan implantasi estradiol-17
β
...
92
17
Kandungan protein pada telur, larva 0, dan larva 2 hari ikan lele yang
diberi berbagai kombinasi
ascorbyl phosphate magnesium
(AMP) dan
implantasi estradiol-17
β
...
92
20
Analisis ragam lama waktu matang ikan lele...
94
21
Analisis ragam indeks kematangan gonad ikan lele...
94
22 Analisis
ragam
diameter telur ikan lele...
95
23 Analisis
ragam
fekunditas telur ikan lele...
95
24
Analisis ragam daya tetas telur ikan lele...
96
25
Analisis ragam ketahanan hidup larva ikan lele...
96
26
Analisis ragam larva abnormal ikan lele...
97
27
Analisis ragam kandungan estradiol plasma darah ikan lele
berdasarkan waktu pengamatan...
98
28 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
00 mg/kg dan estradiol 00
μ
g/kg………....
99
29 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
00 mg/kg dan estradiol 250
μ
g/kg…………..…………...
100
30 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
00 mg/kg dan estradiol 500
μ
g/kg………...……….
101
31 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
600 mg/kg dan estradiol 00
μ
g/kg………...
102
32 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
600 mg/kg dan estradiol 250
μ
g/kg……….
103
33 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
600 mg/kg dan estradiol 500
μ
g/kg……….
104
34 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
1200 mg/kg dan estradiol 00
μ
g/kg………..……...
105
35 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
1200 mg/kg dan estradiol 250
μ
g/kg……….…………...
106
36 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
1200 mg/kg dan estradiol 500
μ
g/kg………...
107
37 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
1800 mg/kg dan estradiol 00
μ
g/kg………..……...
108
38 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
1800 mg/kg dan estradiol 250
μ
g/kg……..……….
109
39 Sebaran diameter telur ikan lele yang diberi
ascorbyl phosphate
magnesium
1800 mg/kg dan estradiol 500
μ
g/kg……….………..
110
Penguji Luar komisi ujian tertutup: Prof. Dr. Ir. Ing Mokoginta, MS
I.
PENDAHULUAN
Latar Belakang
Salah satu usaha yang mutlak dibutuhkan untuk mengembangkan budi daya ikan adalah penyediaan benih yang bermutu dalam jumlah yang memadai dan waktu yang tepat. Selama ini usaha ke arah tersebut telah dilakukan, namun belum berhasil dengan
baik. Kekurangan persediaan benih yang bermutu dalam jumlah dan waktu yang tepat disebabkan oleh belum optimalnya penanganan induk dan larva yang dihasilkan. Perbaikan kualitas dan kuantitas telur melalui perbaikan kualitas pakan induk merupakan alternatif dalam upaya mengatasi masalah tersebut. Kandungan nutrisi pakan ikan adalah salah satu faktor penentu dalam perkembangan oosit, terutama pada
awal perkembangan telur.
Informasi kebutuhan nutrisi untuk ikan-ikan budi daya yang tersedia umumnya hanya sebatas kebutuhan nutrien makro, seperti lemak dan protein, sedangkan informasi kebutuhan mikro nutrien, seperti vitamin dan mineral, masih sangat terbatas.
Selain itu, penggunaan mikro nutrien ini dalam ransum pakan induk hanya mengacu kepada kebutuhan ransum secara umum untuk pertumbuhan. Dalam kondisi seperti ini, sulit mengembangkan teknologi produksi benih berkualitas secara massal. Berbagai penelitian membuktikan bahwa kualitas pakan termasuk nutrien mikro yang merupakan faktor penting yang berhubungan erat dengan kematangan gonad, jumlah
telur yang diproduksi, dan kualitas telur dan larva (Watanabe 1988).
Kualitas telur merupakan refleksi keadaan kimia nutrisi kuning telur yang sangat dipengaruhi oleh kesehatan dan gizi pakan yang diterima oleh induk Blaxter (1969). Beberapa nutrien mikro maupun asam-asam lemak esensial yang terkandung dalam ransum pakan ikan akan diakumulasikan dalam telur untuk digunakan sebagai energi
maupun senyawa pembentuk jaringan tubuh selama proses perkembangan embrio (embriogenesis) maupun larva. Saat telur menetas, sumber energi untuk perkembangan larva ikan sangat bergantung pada material bawaan telur yang telah disiapkan oleh induk dan fase ini merupakan fase yang paling kritis. Material telur yang mengalami defisiensi gizi akan menimbulkan gangguan dalam perkembangan larva dan akhirnya
Keberadaan nutrien dalam telur ini merupakan akumulasi nutrien pada fase pematangan gonad. Secara alamiah proses vitelogenesis memerlukan interaksi faktor eksternal dan internal. Faktor eksternal yang mempengaruhi vitelogenesis antara lain
temperatur, naik turunnya permukaan air, curah hujan, debit air, feromon, dan pakan. Pakan induk yang dapat mempengaruhi vitelogenesis adalah pakan yang berkualitas, yaitu pakan yang mengandung protein, lemak, vitamin E, vitamin C, dan mineral yang sesuai dengan kebutuhan ikan sebagai bahan pembentuk vitelogenin. Faktor internal adalah ketersediaan hormon-hormon steroid, gonad terutama estradiol-17β dalam tingkat yang dapat merangsang vitelogenesis.
Dalam upaya untuk lebih meningkatkan kualitas telur dan larva ikan lele, perlu diadakan perbaikan pengelolaan reproduksi dengan cara mempercepat kematangan gonad melalui penggunaan hormon eksogen dan perbaikan nutrisi induk terutama kebutuhan akan vitamin C. Informasi kebutuhan vitamin C saat siklus reproduksi serta
pengaruhnya pada perkembangan ovarium dan perkembangan larva ikan lele belum ada, padahal informasi ini sangat bermanfaat dalam penyusunan ransum yang tepat untuk induk ikan lele sehingga dapat digunakan untuk memperbaiki penampilan reproduksi ikan tersebut.
Vitamin C merupakan salah satu nutrien mikro yang dibutuhkan oleh induk ikan
dalam proses reproduksi. Hal ini didasarkan pada adanya fluktuasi kandungan vitamin C ovarium selama berlangsungnya siklus reproduksi pada beberapa spesies ikan dan udang yang ditangkap di alam seperti pada udang Palaemon serratus (Guary et al. 1975), ikan Carassius carassius (Saeymour 1981), Godus morhua (Agrawal dan
Mahajan 1980; Sandnes dan Braekkan 1981), Oreochromis sp (Azwar 1997). Kandungan vitamin C dalam ovarium akan meningkat pada awal perkembangannya dan kemudian menurun pada fase akhir sebelum ovulasi. Ikan tidak mampu mensintensis vitamin C (Faster dalam Sandnes 1991) sehingga untuk mempertahankan metabolisme sel, vitamin C mutlak harus diperoleh dari luar tubuh karena tidak terdapat
enzim L-gulonolakton oksidase yang dibutuhkan untuk biosintesis vitamin C (Dabrowski 2002).
karena kekurangan pakan mungkin dapat menyebabkan kadar gonadotropin rendah yang dihasilkan oleh kelenjar adenohipofisis, respons ovari yang kurang atau mungkin kegagalan ovari untuk menghasilkan jumlah estrogen yang cukup (Toelihere 1981).
Seperti yang telah dijelaskan di atas, di samping ketersediaan materi baik kualitas maupun kuantitas untuk mendukung proses reproduksi, kerja hormon juga diperlukan untuk mempercepat dan meningkatkan proses sintesis vitelogenin dan penyerapannya oleh telur.
Estradiol-17β merupakan hormon perangsang biosintesis vitelogenin di hati. Di samping itu estradiol-17β di dalam darah memberikan rangsangan balik terhadap hipofisis dan hipotalamus ikan. Rangsangan yang diberikan oleh estradiol-17β kepada hipofisis ikan adalah rangsangan dalam proses pembentukan gonadotropin yang juga berperan dalam membantu proses penyerapan vitelogenin oleh telur. Vitelogenin yang disintesis di hati dengan bantuan hormon estradiol-17β disekresikan ke dalam aliran darah menuju gonad. Oleh karena adanya peranan estradiol-17β pada biosintesis vitelogenin maka penambahan estradiol-17β melalui implan pada induk ikan diharapkan dapat meningkatkan ketersediaan estradiol-17β yang optimun untuk merangsang hati mensintesis dan mensekresi vitelogenin ke dalam darah sehingga konsentrasi vitelogenin dalam darah akan meningkat. Dengan demikian, penyerapan
vitelogenin oleh oosit akan berjalan dengan lancar.
Dengan melakukan pendekatan-pendekatan di atas maka diharapkan telur yang dihasilkan akan mempunyai derajat pembuahan dan derajat tetas yang tinggi sehingga larva yang dihasilkan berkualitas baik dengan ketahanan hidup yang prima. Namun,
usaha-usaha pendekatan tersebut sering dilakukan secara parsial dan tidak bersifat menyeluruh. Pendekatan yang baik adalah mengkombinasikan antara lingkungan, pakan, dan hormon. Faktor lingkungan sangat kompleks dan sukar ditiru sehingga kombinasi antara pakan dan hormonlah yang sangat dimungkinkan, tetapi dengan memperhatikan faktor lingkungan optimal yang mendukung proses reproduksi.
demikian, perlu dilakukan penelitian untuk mengevaluasi pengaruh penambahan
ascorbyl phosphate magnesium sebagai sumber vitamin C pada pakan dan hormon estradiol-17β pada pematangan gonad, kualitas telur, dan ketahanan hidup larva ikan lele dumbo (Clarias gariepinus). Dengan kombinasi tersebut diharapkan akan lebih memaksimalkan proses reproduksi sehingga diharapkan materi yang masuk dapat dengan optimal diserap telur dan pada akhirnya dapat meningkatkan kualitas telur dan larva.
Perumusan Masalah
Masalah yang dihadapi dalam budi daya ikan lele adalah kurangnya informasi tentang nutrisi induk. Ketersediaan nutrien induk untuk proses vitelogenesis sangat bergantung pada kualitas pakan yang diberikan. Material telur yang mengalami definsiensi gizi akan menimbulkan gangguan pada perkembangan larva dan akhirnya akan mengalami kematian. Salah satu unsur mikro nutrien yang penting dalam proses
vitelogenesis dan embriogenesis adalah vitamin C. Pada proses vitelogenesis, vitamin C sebagai donor elektron dalam reaksi hidroksilasi biosintesis hormon steroid yang diperlukan bagi berlangsungnya proses tersebut. Selain itu, vitamin C juga berfungsi sebagai anti oksidan yang akan melindungi kolesterol dari kerusakan akibat terjadinya proses oksidasi sehingga kebutuhan kolesterol untuk proses biosintesis hormon
estrogen dapat terpenuhi. Pada proses embriogenesis, vitamin C berperan dalam metabolisme lemak, yaitu dalam reaksi biosintesis karnitin, yang berfungsi mentransfer asam lemak rantai panjang dari sitosol ke mitokondria untuk dikonversi menjadi energi melalui proses β-oksidasi. Dengan demikian, kebutuhan energi selama proses tersebut berlangsung dapat dipasok dalam jumlah yang cukup sesuai kebutuhan. Sumber energi
dan nutrien esensial bagi perkembangan larva ikan ketika telur menetas bergantung pada materi bawaan yang telah dipersiapkan oleh induk.
Vitamin C mempunyai fungsi sebagai kofaktor enzim prolil dan lisin hidroksilase yang mengkatalis hidroksilasi prolin dan lisin, yang esensial untuk biosintesis jaringan
vitamin C telur cepat menurun (Sato et al. 1987) karena pada saat itu terjadi pembentukan tulang dan jaringan ikat.
Umumnya, spesies ikan tidak mempunyai kemampuan untuk melakukan
biosintesis vitamin C. Ketidakmampuan ikan mensintesis vitamin C disebabkan karena tidak adanya enzim gulunolakton oksidase yang berperanan dalam mengkonversi L-gulunolakton ke bentuk 2-keto-L-gunulolakton, sebagai tahap akhir dalam sintesis vitamin C. Untuk itu kebutuhan vitamin C ikan harus dipasok dari luar.
Selain itu, pemberian pakan yang tidak optimal menyebabkan kurangnya energi
untuk mendukung proses reproduksi, terutama dalam mensintesis hormon-hormon yang terlibat dalam proses perkembangan telur (vitelogenesis) seperti estradiol-17β. Estradiol-17β adalah hormon steroid yang disintesis pada lapisan granulosa yang kemudian bekerja merangsang biosintesis vitelogenin di hati. Sintesis vitelogenin dirangsang oleh estradiol-17β yang memasuki sistem peredaran darah kemudian merangsang hati mensintesis dan mensekresi vitelogenin. Konsentrasi estradiol-17β di dalam plasma darah yang meningkat selama periode pertumbuhan oosit dapat digunakan sebagai indikator vitelogenesis (Fostier et al. 1978, King dan Pankhurst 2004). Dengan kata lain, estradiol-17β bertanggung jawab dalam sintesis vitelogenin.
Dengan adanya peranan vitamin C dan estradiol-17β seperti tersebut di atas, perlu diujicobakan peranan tersebut pada ikan uji, agar diperoleh informasi pengaruh kombinasi ascorbyl phosphate magnesium sebagai sumber vitamin C dan estradiol-17β pada pematangan gonad dan kualitas telur pada ikan lele dumbo. Informasi ini sangat penting dalam menyusun suatu ransum yang tepat bagi pemenuhan gizi dan dosis
Tujuan dan Manfaat
Penelitian ini bertujuan untuk:
1. Untuk mendapatkan dosis kombinasi ascorbyl phosphate magnesium dan
estradiol-17β yang optimum dalam pematangan gonad, kualitas telur, dan larva ikan lele sebagai ikan uji.
2. Untuk melihat pengaruh kombinasi ascorbyl phosphate magnesium dan estradiol-17β pada penampilan reproduksi induk ikan lele serta keterkaitannya dengan komposisi vitamin C, protein, lemak, fosfolipid, dan ratio hidroksiprolin/prolin
ovarium, telur, dan larva.
Hasil penelitian ini diharapkan dapat digunakan sebagai dasar dalam manajemen induk, dengan pemberian pakan dan perlakuan hormonal yang tepat pada induk ikan sehingga dapat mempercepat pematangan gonad, meningkatkan kuantitas, kualitas telur, dan ketahanan hidup larva ikan ikan lele.
Hipotesis
Dengan pemberian ascorbyl phosphate magnesium pada pakan dan implantasi
hormon estradiol-17β yang optimal dapat meningkatkan kualitas vitelogenin yang
II. TINJAUAN PUSTAKA
Kematangan Gonad Ikan
Kematangan gonad adalah tahapan tertentu perkembangan gonad sebelum dan
sesudah memijah. Selama proses reproduksi, sebagian energi dipakai untuk perkembangan gonad. Bobot gonad ikan akan mencapai maksimum sesaat ikan akan memijah kemudian akan menurun dengan cepat selama proses pemijahan berlangsung sampai selesai. Menurut Effendie (1997), umumnya pertambahan bobot gonad ikan betina pada saat stadium matang gonad dapat mencapai 10-25 persen dari bobot tubuh
dan pada ikan jantan 5-10 persen. Lebih lanjut dikemukakan bahwa semakin rneningkat tingkat kematangan gonad, diameter telur yang ada dalam gonad akan menjadi semakin besar. Pendapat ini diperkuat oleh Kuo et al. (1974) bahwa kematangan seksual pada ikan dicirikan oleh perkembangan diameter rata-rata telur dan
melalui distribusi penyebaran ukuran telurnya.
Ikan lele (Clarias batrachus) pertama kali matang kelamin pada umur satu tahun (Chinabut et al. 1991) dengan ukuran panjang tubuh sekitar 20 cm dan ukuran berat tubuh 100 sampai 200 gram (Mollah dan Tan 1983; Suyanto 1986). Di Thailand, ikan lele yang hidup di alam memijah pada musim penghujan dari bulan Mei sampai
Oktober (Chinabut et al. 1991).
Perkembangan sel telur (oosit) diawali dari germ cell yang terdapat dalam lamela dan membentuk oogonia. Oogonia yang tersebar dalam ovarium menjalankan suksesi pembelahan mitosis dan ditahan pada "diploten" dari profase meiosis pertama. Pada stadia, ini oogonia dinyatakan sebagai oosit primer
(Harder 1975). Oosit primer kemudian menjalankan masa tumbuh yang meliputi dua fase. Pertama adalah fase previtelogenesis, ketika ukuran oosit membesar akibat pertambahan volume sitoplasma (endogenous vitelogenesis), namun belum terjadi akumulasi kuning telur. Kedua adalah fase vitelogenesis, ketika terjadi akumulasi material kuning telur yang disintesis oleh hati, kemudian
saat perkembangan oosit terjadi perubahan morfologis yang mencirikan stadianya. Menurut Nagahama (1983) stadium oosit dapat dicirikan berdasarkan volume sitoplasma, penampilan nukleus dan nukleolus, serta keberadaan butiran kuning
telur. Berdasarkan kriteria ini, oosit dapat diklasifikasikan ke dalam beberapa kelas. Yamamoto dalam Nagahama (1983) membaginya ke dalam 8 kelas, yaitu stadia kromatin-nukleolus, perinukleolus (yang terdiri atas awal dan akhir nukleolus), stadium oil drop stadium yolk primer, sekunder, tertier, dan stadium matang. Sedangkan Chinabut et al. (1991) membagi oosit dalam 6 kelas untuk
Clarias sp, dimana stadia nukleolus dan perinukleolus dikategorikan sebagai stadium pertama, dan setiap stadium dicirikan sebagai berikut:
-stadium 1 : Oogonia dikelilingi satu lapis set epitel dengan pewarnaan
hematoksilin-eosin plasma berwarna merah jambu, dengan inti yang besar di tengah.
-stadium 2 : Oosit berkembang ukurannya, sitoplasma bertambah besar, inti biru terang dengan pewarnaan, dan terletak masih di tengah sel. Oosit dilapisi oleh satu lapis epitel.
-stadium 3 : Pada stadium ini berkembang sel folikel dan oosit membesar dan provitilin nukleoli mengelilingi inti.
-stadium 4 : Euvitilin inti telah berkembang dan berada disekitar selaput inti Stadium ini merupakan awal vitelogenesis yang ditandai dengan
adanya butiran kuning telur pada sitoplasma. Pada stadium ini,
oosit dikelilingi oleh dua lapis sel dan lapisan zona radiata tampak jelas pada epitel folikular.
-stadium 5 : Stadia peningkatan ukuran oosit karena diisi oleh kuning telur. Butiran kuning telur bertambah besar dan memenuhi sitoplasma
dan zona radiata terlihat jelas.
-stadium 6 : Inti mengecil dan selaput inti tidak terlihat, inti terletak di tepi. Zona radiata, sel folikel, dan sel teka terlihat jelas.
telur yang terkandung di dalamnya semakin membesar sebagai hasil dari akumulasi kuning telur, hidrasi, dan pembentukan butir-butir minyak yang berjalan secara bertahap.
Secara garis besar, perkembangan gonad ikan dapat dibagi menjadi dua tahap, yaitu tahap pertumbuhan gonad ikan sampai ikan menjadi dewasa kelamin dan selanjutnya adalah pematangan gamet. Tahap pertama berlangsung mulai dari ikan menetas hingga mencapai dewasa kelamin dan tahap kedua dimulai setelah ikan mencapai dewasa, dan terus berkembang selama fungsi reproduksi masih tetap berjalan
normal (Lagler et al. 1977).
Tam et al. (1986) menyatakan bahwa pada saat menjelang ovulasi akan terjadi peningkatan diameter oosit karena diisi oleh massa kuning telur yang homogen akibat adanya peningkatan kadar estrogen dan vitelogenin. Sementara itu, menurut Bagenal (1969), ukuran telur juga berperan dalam kelangsungan hidup ikan. Benih ikan brown
trout yang berasal dari telur yang berukuran besar mempunyai daya hidup yang lebih tinggi daripada benih ikan yang berasal dari telur yang berukuran kecil. Hal ini terjadi karena kandungan kuning telur yang berukuran besar lebih banyak sehingga larva yang dihasilkan mempunyai persediaan makanan yang cukup untuk membuat daya tahan tubuh yang lebih tinggi dibanding dengan telur-telur yang berukuran kecil.
Woynarovich dan Horvath (1980) menyatakan bahwa induk yang pantas dipijahkan adalah induk yang telah melewati fase pembentukan kuning telur (fase vitellogenesis) dan masuk ke fase dorman. Fase pembentukan kuning telur dimulai sejak terjadinya penumpukan bahan-bahan kuning telur da!am sel telur dan berakhir
setelah sel telur mencapai ukuran tertentu atau nukleolus tertarik ke tengah nukleus. Setelah fase pembentukan kuning telur berakhir, sel telur tidak mengalami perubahan bentuk selama beberapa saat, tahap ini disebut fase istirahat (dorman). Menurut Lam (1985), apabila rangsangan diberikan pada saat ini, maka akan menyebabkan terjadinya migrasi inti ke perifer, kemudian inti pecah atau melebur pada saat
Ukuran sel telur ada hubungannya dengan fekunditas. Makin banyak telur yang
dipijahkan ukuran telurnya makin kecil, misalnya ikan cod (diameternya 1-1,7mm) produksinya 10 juta telur. Salmon Atlantik yang memiliki diameter telur 5-6 mm, produksi telurnya 2.000-3.000 butir (Blaxter 1969), sedangkan untuk ikan belut dengan diameter telur 1–1,5 mm produksinya 2.200–5.400 telur (Sidthimunka 1972).
Vitelogenesis
Sintesis vitelogenin (prekursor kuning telur) di dalam hati disebut vitelogenesis. Vitelogenin diangkut dalam darah menuju oosit, lalu diserap secara selektif dan disimpan sebagai kuning telur. Vitelogenin ini berupa glikofosfoprotein yang
mengandung kira-kira 20% lemak, terutama fosfolipid, trigliserida, lipoprotein, dan kolesterol. Berat molekul vitelogenin untuk beberapa jenis ikan diketahui antara 140- 220 kDa (Tyler 1991; Komatsu dan Hayashi 1997).
Proses oogenesis pada teleost terdiri atas dua fase, yaitu pertumbuhan oosit (vitelogenesis) dan pematangan oosit. Vitelogenesis merupakan aspek penting dalam
pertumbuhan oosit yang meliputi rangkaian proses (1) adanya sirkulasi estrogen
(estradiol-17β) dalam darah menggertak hati untuk mensintesis dan mensekresikan
vitelogenin yang merupakan prekursor protein kuning telur; (2) vitelogenin diedarkan menuju lapisan permukaan oosit yang sedang tumbuh; (3) secara selektif, vitelogenin akan ditangkap oleh reseptor dalam endositosis, dan (4) terjadi translokasi sitoplasma
membentuk badan kuning telur bersamaan dengan pembelahan proteolitik dari vitelogenin menjadi subunit lipoprotein kuning telur, lipovitelin, dan fosvitin. Adanya vitelogenin menunjukkan terjadinya akumulasi lipoprotein kuning telur di dalam oosit. Pada beberapa jenis ikan selama pertumbuhan oosit terjadi peningkatan Indeks Somatik Gonad (IGS) 1 sampai 20% atau lebih.
Pada ikan betina, ovari berespons terhadap peningkatan konsentrasi gonadotropin
dengan meningkatkan secara tidak langsung produksi estrogen, yakni estradiol-17β
(E2). Estradiol-17β beredar menuju hati, memasuki jaringan dengan cara difusi dan
Pembesaran oosit disebabkan terutama oleh penimbunan kuning telur. Seperti pada kebanyakan ikan, kuning telur merupakan komponen penting oosit ikan Teleostei. Ada tiga tipe material kuning telur pada ikan Teleostei: butiran kecil minyak,
gelembung kuning telur (yolk vesicle) dan butiran kuning telur (yolk globule). Secara umum, butiran kecil minyak yang kita kenal dengan lipid yang berantai panjang (asam lemak tidak jenuh) pertama kali muncul di daerah perinuklear dan kemudian berpindah ke periferi (tepi sel) pada tahap selanjutnya. Urutan kemunculan material kuning telur bervariasi antarspesies. Pada rainbow trout, butiran kecil muncul segera setelah
dimulainya pembentukan gelembung kuning telur (Yamamoto et al. 1965 dalam Nagahama 1983).
Fenomena penimbunan material kuning telur oleh oosit ikan dibagi menjadi dua fase, yakni sintesis kuning telur di dalam oosit atau vitelogenesis endogen dan penimbunan prekursor (bahan pembentuk) kuning telur yang disintesis di luar oosit
atau vitelogenesis eksogen (Matty 1985). Gelembung kuning telur positif-PAS (mukopolisakarida atau glikoprotein) umumnya merupakan struktur yang pertama muncul dalam sitoplasma oosit selama pertumbuhan sekunder oosit, dan pertama kali muncul di zona terluar dan zona midkortikal pada oosit.
Ketika vitelogenesis berlangsung, sebagian besar sitoplasma telur matang
ditempati oleh banyak gelembung kuning telur yang padat dengan asam lemak dan dikelilingi oleh selapis membran pembatas. Selama tahap akhir vitelogenesis, globula kuning telur beberapa ikan Teleostei bergabung satu sama lain membentuk masa tunggal kuning telur.
Perkembangan gonad ikan betina terdiri atas beberapa tingkat yang dapat didasarkan atas pengamatan secara mikroskopis dan makroskopis. Secara mikroskopis perkembangan telur diamati untuk menilai perkembangan ovarium antara lain tebal dinding indung telur, keadaan pembuluh darah, inti butiran minyak, dan kuning telur. Secara makroskopis perkembangan ovarium ditentukan dengan mengamati warna
indung telur, ukuran butiran telur, dan volume rongga perut ikan.
akan bervariasi. Pada tahap perkembangan awal, oogonia terlihat masih sangat kecil, berbentuk bulat dengan inti sel yang sangat besar dibandingkan dengan sitoplasmanya. Oogonia terlihat berkelompok tapi kadang-kadang ada juga yang berbentuk tunggal.
Sementara itu oogonia terus membelah diri dengan cara mitosis. Pada ikan yang mempunyai siklus reproduksi tahunan atau tengah tahunan akan terlihat adanya puncak-puncak pembelahan oogonia. Pada ikan yang memijah sepanjang tahun, perbanyakan oogonia akan terus menerus sepanjang tahun.
Transformasi oogonia menjadi oosit primer banyak terjadi pada tahap
pertumbuhan yang ditandai dengan munculnya kromosom. Segera setelah itu, folikel berubah bentuk, dari semula yang berbentuk skuamosa menjadi berbentuk kapsul oosit. Inti sel terletak pada bagian sentral dibungkus oleh lapisan sitoplasma yang tipis. Pada perkembangan selanjutnya, oosit membentuk lapisan korion, membran, granulosa, membran, dan teka. Juga butir-butir lemak mulai terlihat ditumpuk pada sitoplasma
dan bersamaan dengan itu muncul cortical alveoli. Pada saat ini, ketersediaan vitamin C mutlak diperlukan karena dengan peningkatan kadar asam lemak, kebutuhan vitamin C semakin meningkat pula. Vitamin C dapat mencegah terjadinya oksidasi pada unit-unit asam lemak, terutama asam lemak tidak jenuh (Machlin 1990). Butir-butir lemak ini selanjutnya akan bertambah besar pada vitelogenesis yang diawali dengan
pembentukan vakuola-vakuola yang kemudian diikuti dengan munculnya globula kuning telur, bersamaan dengan itu oosit membengkak secara menyolok. Kuning telur pada ikan terdiri atas fosfoprotein dan lipoprotein yang dihasilkan oleh hati kemudian disalurkan ke dalam peredaran darah.
Peranan Vitamin C pada Reproduksi Ikan
Variasi kadar vitamin C ovarium pada saat siklus reproduksi dari berbagai spesies ikan telah dicatat oleh beberapa peneliti sehingga menimbulkan spekulasi kemungkinan pentingnya senyawa ini saat ovarium berkembang. Kadar vitamin C ikan karper Krusian (Carassius carassius) saat siklus reproduksi berkisar dari 92 sampai 203 ug/g (Saeymour 1981), ikan cod Atlantik (Gadus morrhua) berkisar dari 80 sampai 203 ug/g (Sandnes dan Braekkan 1981), dan karper India dari 225 sampai 286 ug/g) (Agrawal dan Mahajan 1980). Cho et al. (1979) mendapatkan bahwa kadar vitamin C ovarium ikan trout (Oncorhynchus mykiss) mencapai maksimum pada 451 ug/g bobot basah pada saat akan ovulasi. Dengan memperhatikan indeks gonad somatik, Sandnes dan Braekkan (1981) mencatat bahwa akumulasi vitamin C tertinggi menjelang GSI mencapai maksimum, kemudian menurun saat terjadi ovulasi. Pengamatan pada ikan kod Atlantik memperlihatkan bahwa kandungan vitamin C pada stadia awal pertumbuhan ovarium adalah 150 ug/g dan tertinggi mencapai 500 ug/g (Sandnes 1984). Menurut Ishibashi et al. (1994), perubahan vitamin C ovarium selama periode pematangan berkaitan dengan peningkatan ukuran oosit karena akumulasi material kuning telur. Agrawal dan Mahajan (1980) mencatat bahwa kandungan vitamin C darah ikan karper India yang ditangkap di alam mencapai titik terendah saat musim pemijahan, yaitu 17,95-19,65 ug/ml, dan saat pertumbuhan ovarium kadar vitamin C mencapai kisaran 20,39-25,95 ug/ml. Disimpulkan pula bahwa ada mobilisasi vitamin C yang diperoleh dari pakan alami ke ovarium saat siklus reproduksi.
Penelitian Alava et al. (1993) memperlihatkan bahwa pemberian askorbil-2-fosfat magnesium, suatu bentuk turunan vitamin C, dalam ransum dapat menstimulasi perkembangan gonad induk udang Penaeus japonicus betina. Percobaannya dengan menggunakan pakan yang disuplementasi askorbil-monofosfat magnesium masing-masing 500, 1000, dan 1500 mg/kg. Setelah pemeliharaan 170 hari, nilai IGS induk betina mencapai 2.40, 2.51, dan 1.81%, sedangkan nilai IGS induk jantan adalah 0.76, 0.87, dan 0.91%. Untuk kontrol tidak diperoleh data karena induk mati sebelum berakhimya percobaan. Penelitian Ishibasi et al. (1994) terhadap ikan Japanese parrot
(Oplegnathus fasciatus) memperlihatkan peningkatan indeks gonad somatik dengan peningkatan dosis vitamin C yang diberikan. Ikan yang menerima pakan dengan suplementasi vitamin C 0, 300, 1000, dan 3000 mg/kg memperlihatkan nilai IGS masing-masing 0.5, 0.9, 1.4, dan 2.2 % untuk induk betina, dan 0.4, 0.6, 0.5, dan 0.8 untuk induk jantan. Pengamatan secara mikrokospis terhadap ovarium juga memperlihatkan persentase induk yang mencapai aktivitas vitelogenesis meningkat dengan peningkatan dosis vitamin C. Induk yang menerima pakan tanpa suplementasi vitamin C tidak menunjukkan adanya oosit pada fase vitelogenesis, sedangkan dengan perlakuan suplementasi vitamin C 300, 1000, dan 3000 mg/kg pakan, jumlah induk yang ovarinya mencapai stadium vitelogenesis hingga matang adalah 20, 40, dan 80%. Soliman et al. (1986) mengamati pengaruh asam askorbat pada penampilan reproduksi ikan Oreochromis mossambicus melaporkan bahwa ikan yang menerima pakan dengan suplementasi vitamin C biasa 1250 mg/kg memperlihatkan gejala siap mijah lebih cepat dua minggu dibandingkan induk yang menerima pakan tanpa suplementasi vitamin C.
Percobaan Priyono et al. (1996) mencatat bahwa ikan bandeng (Chanos chanos
vitamin C dalam ransum yang diterima oleh induk dapat ditransfer ke telur, dan disiapkan untuk perkembangan embrio. Pengamatannya pada telur ikan Oreochromis mossambicus dimana induk yang menerima pakan dengan suplementasi vitamin C 1250 mg/kg pakan mengandung vitamin C 201.83 ug/g dan daya tetas telur mencapai 89.33%, sedangkan kandungan vitamin C telur dari induk yang menerima pakan tanpa vitamin C tidak terdeteksi dan mempunyai daya tetas 56.90%, dan 85% pascalarva yang dihasilkan mengalami gangguan pertumbuhan tulang belakang. Percobaan Akiyama et al. (1990) pada ikan sardin (Sardinops sagaxmelanosticia) menunjukkan bahwa tidak ditemui telur yang menetas dari induk yang menerima pakan dengan suplementasi vitamin C 80 mg/kg pakan, sedangkan induk yang menerima pakan dengan suplementasi vitamin C lebih tinggi, yaitu 3200 mg/kg pakan menghasilkan daya tetas telur lebih baik. Vitamin C yang ditransfer dari induk ke material telur berperan dalam mendukung perkembangan embrio (Sandnes 1991). Menurut Sandnes
et al. (1984) kandungan vitamin C telur 20 ug/g merupakan batas terendah untuk perkembangan normal embrio ikan trout.
Peranan Estradiol-17β pada Reproduksi Ikan
Proses vitelogenesis pada ikan melibatkan beberapa hormon, dan pada ikan ada dua macam hormon gonadotropin yang dihasilkan oleh adenohipofisis yang berperan
sebagai follicle stimulating hormone (FSH) dan luteinizing hormone (LH). Hormon tersebut adalah FSH (GTH I), yang bekerja merangsang perkembangan folikel melalui sekresi estradiol-17β pada ovari dan LH (GTH II) yang dibutuhkan untuk proses pematangan akhir oosit (Nagahama 1983). Gonadotropin yang dihasilkan akan bekerja pada sel teka sebagai tempat sintesis testosteron. Testosteron yang dihasilkan oleh
lapisan sel teka akan masuk ke dalam lapisan granulosa. Di dalam lapisan granulosa testosteron diubah menjadi estradiol-17β dengan bantuan enzim aromatase.
Estradiol-17β merupakan perangsang dalam biosintesis vitelogenin di hati. Di
samping itu, estradiol-17β yang terdapat dalam darah memberikan rangsangan balik
terhadap hipofisis dan hipotalamus ikan. Rangsangan yang diberikan oleh estradiol-17β
terhadap hipofisis ikan adalah rangsangan dalam proses pembentukan gonadotropin.
nantinya berperan dalam proses biosintesis estradiol-17β pada lapisan granulosa. Siklus
hormonal terus berjalan di dalam tubuh ikan selama terjadinya proses vitelogenesis (Nagahama 1983 dan Yaron 1995). Menurut Aida et al. (1991) proses vitellogenesis
pada ikan terjadi seperti pada Gambar 1.
Gambar 1. Proses vitelogenesis pada ikan (Aida et al. 1991)
Beberapa penelitian menunjukkan bahwa peningkatan konsentrasi
estradiol-17β akan meningkatkan konsentrasi vitelogenin darah dan konsentrasi
estradiol-17β tinggi pada saat vitelogenesis pada European sea bass (Dicebtrachus labrax)
(Hassin et al. 1991); salmon (Salmo gairdneri) (Van Bohemen et al. 1981); mas koki (Pankhurst et al. 1986); jambal siam (Pangasius hypophthalmus) (Indriastuti 2000 dan Monijung 2002). Penelitian untuk melihat hubungan tersebut telah dilakukan pada ikan trout, Salmo trutta dan rainbouw trout Salmo gairdneri
(Hjartarson et al. 1991), Striped bass Morone sexatilis (Sullivan et al. 1991), dan
Clarias macrocepalus (Tan-Fermin et al. 1997). Sintesis vitelogenin di hati sangat
dipengaruhi oleh estradiol-17β yang merupakan stimulator dalam biosintesis
vitelogenin. Selain itu, sintesis tersebut dipengaruhi juga oleh androgen yang ada
Umpan balik
dalam tubuh ikan (testosteron) dan melalui perubahan androgen menjadi estrogen oleh enzim aromatase hati (Yaron 1995). Dengan demikian, peningkatan GtH dapat meningkatkan estradiol, dan pola kandungan estradiol seiring dengan
perkembangan telur (Yaron 1995; Tan-Fermin et al. 1997).
Kualitas Telur Ikan
Kualitas telur adalah kemampuan telur untuk menghasilkan larva yang berdaya
hidup. Kemampuan telur untuk menghasilkan larva yang berdaya hidup dapat ditentukan dengan beberapa faktor antara lain faktor fisik, kimia, genetik, dan fisiologi selama terjadi proses awal pada telur. Jika salah satu faktor essensial ini tidak ada, perkembangan telur akan gagal dalam beberapa stadia. Telur merupakan hasil akhir dari proses gametogenesis setelah oosit mengalami fase pertumbuhan yang panjang
yang sangat bergantung pada gonadotropin dari pituitari. Perkembangan diameter telur pada oosit teleostei umumnya karena akumulasi kuning telur.
Ada tiga macam material kuning telur yang berbeda, yaitu 1) oil droplet (butir minyak), 2) yolk vesicle (vesikula kuning telur), dan 3) yolk globule (bola kecil kuning telur). Dalam vitelogenesis yang sedang berkembang, sitoplasma telur yang matang
ruangannya diisi oleh bola-bola kecil kuning telur saling bersatu dengan yang lainnya membentuk menjadi massa kuning telur.
Pembuahan atau fertilisasi adalah proses bergabungnya inti sperma dengan inti sel telur dalam sitoplasma sehingga membentuk zigot, dimana asosiasi ini merupakan
mata rantai awal dan sangat penting pada proses fertilisasi. Laju pembuahan sering digunakan sebagai parameter untuk mendeteksi kualitas telur.
Fertilisasi dan proses aktivasi pada telur ikan menjadi penting untuk beberapa perkembangan embrio. Selama fertilisasi dan aktivasi, pada telur-telur ikan teleost terjadi reaksi kortikal. Alveoli kortikal melebur dan melepaskan kandungannya
korion telur selama proses aktivasi akibat dari reaksi enzim. Telur yang kualitasnya bagus memiliki korion yang keras.
Selain hal-hal di atas parameter lain yang dapat juga menjadi patokan kualitas
telur adalah transparansi telur dan distribusi butiran lemak (Mc Evoy 1984). Kualitas telur yang baik umumnya transparan dan jelas kelihatan serta pembelahan awal yang simetris.
Selama oogenesis, kuning telur mengakumulasi sejumlah besar granula kuning telur dan lipid yang terisi pada bagian tengah. Kisaran diameter granula telah diukur
antara 6-24 µm (Linhart et al. 1995). Jumlah dan distribusi lemak (butir lemak) sangat bervariasi. Ukuran diameternya antara 1-1.5µm, dan diketahui bahwa butir lemak berfungsi sebagai cadangan energi dan fungsi hidrostatik. Distribusi butir-butir lemak ini juga menjadi parameter kualitas telur.
Ukuran telur berkorelasi dengan ukuran larva. Larva yang besar lebih tahan tanpa
pakan dibandingkan dengan larva yang kecil yang ditetaskan dari telur yang kecil. Hubungan positif antara ukuran larva dan ukuran telur telah dilaporkan untuk Salmon salar, Onchorhynchus mykiss (Kamler 1992), dan turbot (Scopththalmus maximus L) (Kjorsvik et al. 2003).
Beberapa peneliti menunjukkan bahwa telur yang berukuran besar menghasilkan
kelangsungan hidup yang lebih tinggi. Kamler (1992) mengajukan sebuah persamaan kelangsungan hidup untuk ikan pelagis laut, laju mortalitas telur dan larva berbanding terbalik dengan ukuran telur. Bila tidak ada makanan eksternal, larva yang lebih besar yang berasal dari telur yang besar dapat bertahan hidup lebih lama dibanding larva yang
berasal dari telur yang kecil.
Pada kondisi yang baik seperti di pembenihan, ukuran telur tidak memberikan dampak secara langsung pada kelangsungan hidup dan pertumbuhan pada ikan Rainbow trout (Salmo gairdneri: Pitman 1979; Springate dan Bromage 1985), Salmo salar (Thorpe et al. 1984), catfish Clarias macrocepalus. (Reagen dan Conley 1977),
dan carp (Kirpchnikov 1966 dalam Kjorsvik et al. 1990).
pertumbuhan. Komponen yang diketahui esensial untuk kehidupan organisme (terutama untuk organisme yang tidak dapat mensintesis nutrien), seperti vitamin C harus ada dalam pakan dalam jumlah tertentu untuk kebutuhan biologi organisme
tersebut. Oleh karena itu, parameter biokimia kualitas telur dapat digunakan untuk mengevaluasi kandungan biokimia telur bahkan sebelum fertilisasi.
Materi yang diperlukan selama perkembangan dapat dibagi dua, yaitu 1) diperlukan secara langsung untuk sintesis jaringan embrionik dan 2) digunakan untuk energi metabolisme. Jumlah nutrien yang diperlukan jelas bervariasi bergantung pada
beberapa faktor, antara lain waktu inkubasi, ukuran ikan pada waktu menetas, dan lamanya anak-anak ikan memerlukan persediaan makanan endogen sebelum menemukan semua keperluan dari sumber lain. Kandungan protein lipid dan karbohidrat berkorelasi positif dengan kelangsungan hidup larva (Kamler 1992)
Kerangka Teoretis
Secara alami proses vitellogenesis memerlukan interaksi antara faktor eksternal dan faktor internal. Faktor eksternal yang mempengaruhi vitelogenesis antara lain temperatur, curah hujan, debit air, feromon, dan pakan. Pakan merupakan komponen
penting dalam proses pematangan gonad karena proses vitelogenesis pada dasarnya adalah proses akumulasi nutrien dalam sel telur sehingga ketersediaan nutrien pada sel telur akan menentukan kualitas telur dan pada akhirnya juga pada perkembangan larva. Di pihak lain, faktor internal yang terpenting adalah tersedianya hormon-hormon steroid gonad terutama estradiol-17β pada tingkat yang dapat merangsang vitelogenesis.
Proses perkembangan gonad ikan dimulai dengan adanya respons dari hipotalamus terhadap sinyal lingkungan, kemudian merangsang pituitari untuk menghasilkan gonadotropin yang nantinya akan mempengaruhi sintesis testosteron
yang diaromatasi menjadi estradiol-17β. Estradiol-17β merupakan perangsang biosintesis vitelogenin di hati, dan dapat memberikan umpan balik terhadap hipofisis serta hipotalamus ikan untuk menghasilkan gonadotropin. Sintesis vitelogenin ini distimulir oleh estradiol-17β yang masuk ke dalam sistem vaskuler dan merangsang hati untuk mensintesis dan mensekresi vitelogenin. Selanjutnya vitelogenin ini dilepas
ditimbun sebagai komponen kuning telur. Peningkatan konsentrasi estradiol-17β akan meningkatkan konsentrasi vitelogenin darah. Hal ini menunjukkan bahwa konsentrasi estradiol-17β dalam tubuh ikan sejalan dengan perubahan konsentrasi vitelogenin darah.
Vitamin C mempunyai peranan dalam proses vitelogenesis dan embriogenesis. Hal ini terjadi karena vitamin C mempunyai peranan penting dalam reaksi hidroksilasi biosintesis hormon steroid, metabolisme lemak, dan sintesis kolagen (Masumoto et al. 1991; Linder 1992: Piliang 1995).
Vitamin C memainkan peranan penting dalam reaksi hidroksilasi biosintesis hormon steroid yang diperlukan bagi berlangsungnya proses tersebut. Selain itu, vitamin C juga berfungsi sebagai antioksidan. Pada proses embriogenesis, vitamin C berperan dalam metabolisme lemak, yaitu dalam reaksi biosintesis karnitin, yang berfungsi mentransfer asam lemak rantai panjang dari sitosol ke mitokondria untuk
dikonversi menjadi energi melalui proses β-oksidasi. Dengan demikian, kebutuhan energi selama proses tersebut berlangsung dapat dipasok dalam jumlah yang cukup sesuai kebutuhan. Sumber energi dan nutrien esensial bagi perkembangan larva ikan ketika telur menetas bergantung pada materi bawaan yang telah dipersiapkan oleh induk.
Vitamin C mempunyai fungsi sebagai kofaktor enzim prolil dan lisil hidroksilase yang mengkatalis hidroksilasi prolin dan lisin, dan esensial untuk perkembangan normal jaringan kolagen yang banyak terdapat pada ovarium. Kolagen merupakan penyusun utama dinding dalam kantong ovarium. Hal inilah yang menyebabkan
adanya fluktuasi kandungan vitamin C ovarium selama berlangsungnya siklus reproduksi. Selain itu, kolagen dibutuhkan untuk perkembangan embrio dan larva, karena kolagen merupakan komponen utama pada kulit dan jaringan ikat serta zat-zat pembentuk tulang dan gigi.
Defisiensi vitamin C pada ikan dapat disebabkan kurang tersedianya
senyawa ini dalam pakan yang diberikan karena mudah larut dalam air dan hilang selama proses pembuatan pakan. Di samping itu, ikan tidak mampu mensintesis vitamin C, walaupun sel-selnya membutuhkan vitamin C. Dari hasil percobaan Azwar (1977) diketahui bahwa ascorbyl phosphate magnesium
III. BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan di Instalasi Riset Perikanan Budidaya Air Tawar
(BRPBAT), Depok, Jawa Barat. Penelitian ini dimulai sejak Juni sampai Desember
2005.
Bahan dan Alat
Pakan uji
Selama penelitian, jenis pakan yang digunakan adalah pakan dalam bentuk
pelet dengan komposisi berdasarkan komposisi pakan buatan untuk induk ikan
lele (Khoironi, 2002), kemudian ditambahkan vitamin C yang berupa
ascorbyl
phosphate magnesium
(Showa Denko Jepang) dengan bobot molekul 379,61
serta kandungan asam askorbat 46%. Komposisi pakan uji dan proksimat pakan
dapat dilihat pada Tabel 2 dan 3.
Hewan Uji
Dalam percobaan ini hewan uji yang akan digunakan adalah induk ikan lele
dumbo (
C. gariepinus
) dari hasil pembesaran petani ikan yang ada di Dermaga Bogor.
Induk ikan adalah hasil pemijahan dari satu induk dan berumur 6 bulan. Jumlah ikan
yang digunakan adalah 216 ekor induk ikan lele betina pada tingkat kematangan
gonad II dan 216 ekor ikan jantan. Masing-masing perlakuan mempunyai 18 ekor
ikan. Ukuran panjang dan bobot ikan uji masing-masing 32–38 cm dan 250–328 gram.
Hormon implantasi
Hormon yang digunakan adalah estradiol-17
β
(Sigma Chemical Company) dan
bubuk kolesterol (5-cholesten-3
β
-ol), cocoa butter, alkohol 98%, 2-phenoxyethanol dan
betadine (prosedur selengkapnya pada Lampiran 03).
Wadah
Ikan dipelihara dalam 12 buah jaring terapung yang diletakkan di dalam
kolam dengan ukuran panjang, lebar, dan kedalaman masing-masing 3.0 meter,
untuk mencegah agar ikan uji tidak melompat ke luar wadah (Gambar 2). Untuk
pemijahan digunakan akuarium sebanyak 40 buah dengan ukuran 60 X 50 X 50 cm
yang dilengkapi dengan pipa-pipa aerasi. Akuarium tersebut juga akan digunakan
[image:40.612.127.507.162.410.2]untuk proses inkubasi, penetasan telur, dan pemeliharaan larva.
Gambar 2. Wadah penelitian yang terbuat dari bambu dengan 12 jaring apung
dengan ukuran 3 X 2 X 2 m yang di atasnya ditutupi dengan jaring
Tabel 1. Komposisi bahan pakan dengan penambahan
ascorbyl phosphate
magnesium
(APM) 0, 600, 1200, dan 1800 mg/kg pakan
Persentase
Ascorbyl Phosphate Magnesium
(APM)
Bahan Pakan (%)
0
600
1200
1800
Tepung ikan
23.28
23.28
23.28
23.28
Tepung kedelai
28.08
28.08
28.08
28.08
Pollard
36.29
36.29
36.29
36.29
Minyak ikan
1.37
1.37
1.37
1.37
Minyak jagung
1.37
1.37
1.37
1.37
Cholin Chlorida
0.50
0.50
0.50
0.50
Mineralª
5.00
5.00
5.00
5.00
Vitamin mix
(tanpa vitamin C)
0.61
0.61
0.61
0.61
CMC
2.00
2.00
2.00
2.00
Selulosa
1.50
1.44
1.38
1.32
[image:40.612.103.521.496.695.2]Tabel 2. Komposisi proksimat pakan dengan perlakuan penambahan
ascorbyl
phosphate magnesium
(APM) 0, 600, 1200, dan 1800 mg/kg pakan.
Persentase
Ascorbyl Phosphate Magnesium
(APM)
Komposisi
Proksimat
0
600
1200
1800
Protein
36.42
36.12
35.98
36.67
Lemak
7.47
7.89
7.24
7.95
Karbohidrat
36.56
36.47
36.12
36.98
Abu
10.34
10.65
9.87
10.80
Serat kasar
8.56
8.90
8.24
8.95
DE (kkal/g pakan)
308.65
308.90
309.46
309.96
Vit. C (mg/kg pakan) 9.56
204.76
386.90
564.82
Metode Penelitian
Perlakuan
Metode yang digunakan pada penelitian ini adalah model eksperimental dengan
menggunakan Rancangan Faktorial 4 X 3 sehingga terdapat 12 interaksi perlakuan. Faktor
pertama adalah 4 dosis
ascorbyl phosphate magnesium
yang berbeda dalam pakan ikan,
yaitu 0, 600, 1200, dan 1800 mg/kg pakan. Faktor kedua adalah dosis hormon
estradiol-17
β
yang berbeda, yaitu 0, 250, dan 500
μ
g/kg bobot tubuh. Secara rinci rancangan
perlakuan tersebut dapat dilihat pada Tabel 3.
Tabel 3. Perlakuan berbagai kombinasi dosis
ascorbyl phosphate magnesium
(APM)
dan implantasi estradiol-17
β
(E2)
Dosis APM
(mg/kg pakan)
Dosis estradiol-17
β
(
μ
g/kg bobot tubuh)
0 250
500
0
0.00 : 0.00 (A)
0.00 : 250 (B)
0.00 : 500 (C)
600
600 : 0.00 (D)
600 : 250 (E)
600 : 500 (F)
1200
1200 : 0.00 (G)
1200 : 250 (H)
1200 : 500 (I)
Parameter yang dievaluasi
Adapun peubah yang diamati adalah sebagai berikut
1.
Diameter telur, yang diukur sebanyak 100 butir dengan menggunakan
mikroskop yang dilengkapi dengan mikrometer okuler.
2.
Pengamatan perkembangan gonad, yang dilakukan dengan membuat preparat
histologis ovarium dan diambil di awal, tengah, dan akhir penelitian.
3.
Fekunditas relatif
Fr =
Wt
Ft
dengan Fr : Fekunditas relatif
Ft : Fekunditas total (Butir telur)
Wt : Bobot tubuh ikan (Gram)
4.
Daya tetas telur
DT =
x
100
F
a
dengan DT : Daya tetas telur
a : Jumlah telur yang menetas
F : jumlah total telur yang ditetaskan
5. Indeks Gonad Somatik (IGS)
IGS (%) =
Bg
Bo
x 100
dengan IGS : Indeks Gonad Somatik (%)
Bo : Bobot ovarium
Bg : Bobot gonad
6. Perhitungan ketahanan hidup larva dilakukan dengan memelihara 100 ekor larva
yang baru menetas di akuarium dengan tidak diberi makanan. Larva diamati setiap
hari, mortalitasnya dicatat. Perhitungan lama waktu (hari) dihentikan jika larva
7. Analisis proksimat pakan, telur, dan larva, yang dilakukan mengikuti protokol
yang dikemukakan Takeuchi (1988).
8. Analisis fospolipida pada telur menggunakan kromatografi gas (Takaeuchi 1988)
9. Analisis kandungan vitamin C pada ovarium, telur dan larva mengikuti prosedur
yang dikemukakan oleh Schuep
et al
. (1994).
10. Analisis hidroksiprolin dan prolin pada ovarium dan larva menggunakan
spektrofotometer (Apriyantono
et al
. 1989).
11. Analisis estradiol-17
β
pada plasma darah ikan dengan teknik radioimunoassay
Pelaksanaan Penelitian
Pematangan gonad
Sebelum induk ikan dimasukkan ke dalam jaring, dilakukan pemeriksaan jaring
apung dari kebocoran. Penempatan jaring apung berdasarkan perlakuan dilakukan secara
acak. Jaring apung ditempatkan di kolam dengan air yang mengalir dengan pergantian air
sebanyak 100 sampai 200% dalam sehari. Pembersihan sisa-sisa makanan dan kotoran di
dasar jaring apung dilakukan setiap hari. Sebelum dilakukan percobaan, ikan uji
diaklimatisasi selama 1 minggu. Selama periode aklimatisasi, ikan diberi pakan dari
perlakuan kontrol sebanyak 2 % bobot tubuh perhari.
Ikan uji yang digunakan dalam penelitian ini ditandai secara individu dengan
menggunakan tasi (tagging) yang berwarna dan bernomor yang diselipkan pada sirip
punggung. Setiap perlakuan menggunakan satu wadah dan tiap wadah diisi 18 ekor induk
ikan betina. Selama penelitian, ikan yang dipelihara diberi pakan buatan 2 kali sehari
at
satiation
pada pagi dan sore hari. Bobot pakan yang diberikan pada ikan setiap perlakuan
dicatat. Pengukuran parameter kualitas air untuk suhu dilakukan setiap hari pagi dan sore
hari. Kandungan oksigen dan pH diukur sekali seminggu, sedangkan kandungan
karbondioksisida dan amonia diukur tiga kali, yaitu pada awal, tengah, dan akhir penelitian.
Tabel 4. Parameter fisika kimia air selama percobaan
Parameter Kisaran
Suhu
27.5 – 31
0C