TRANSFORMASI GENETIK Nicotiana benthamiana DENGAN
GEN PEMBUNGAAN Hd3a DARI PADI
LAILA NUR SYAFITRI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
LAILA NUR SYAFITRI. Transformasi Genetik Nicotiana benthamiana dengan Gen Pembungaan Hd3a dari Padi. Dibimbing oleh SUHARSONO dan SRI KOERNIATI.
Pembungaan adalah salah satu peristiwa penting dalam siklus hidup tanaman tingkat tinggi dan karakter yang sangat penting untuk menentukan kemampuan spesies beradaptasi pada berbagai kondisi lingkungan. Gen Hd3a adalah salah satu gen yang berhubungan dengan waktu pembungaan di padi. Ekspresi berlebih Hd3a menyebabkan pembungaan lebih awal pada padi. Penelitian ini bertujuan untuk melakukan transformasi genetik Nicotiana benthamiana dengan gen Hd3a dari padi. Transformasi genetik dilakukan melalui bantuan Agrobacterium tumefaciens LBA4404 dengan metode kokultivasi. Seleksi tunas transgenik putatif dilakukan dengan 30 mg/l higromisin. Hasil penelitian menunjukkan bahwa proses transformasi N. benthamiana menghasilkan tunas transgenik putatif dengan nilai efisiensi transformasi cukup tinggi yaitu 86%. Analisis molekuler dengan PCR terhadap tanaman transgenik putatif menunjukkan bahwa tanaman tersebut mengandung gen hpt yang bertanggung jawab terhadap resistensi tanaman terhadap higromisin. Keberadaan bagian gen hpt dalam genom Nicotiana benthamiana transgenik ini juga menunjukkan keberadaan gen pembungaan Hd3a. Tunas Nicotiana benthamiana transgenik yang mengandung gen Hd3a berbunga lebih awal, sehingga gen Hd3a mempunyai peranan dalam memicu pembungaan.
ABSTRACT
LAILA NUR SYAFITRI. Genetic Transformation of Nicotiana benthamiana with Hd3a flowering gene from rice. Supervised by SUHARSONO and SRI KOERNIATI.
TRANSFORMASI GENETIK Nicotiana benthamiana DENGAN
GEN PEMBUNGAAN Hd3a DARI PADI
LAILA NUR SYAFITRI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul : Transformasi Genetik Nicotiana benthamiana dengan Gen Pembungaan Hd3a dari Padi
Nama : Laila Nur Syafitri NIM : G34060962
Disetujui :
Pembimbing I,
Pembimbing II,
Prof. Dr. Ir. Suharsono, DEA. Dr. Ir. Sri Koerniati, M.Sc. NIP 19610428 198703 1 003 NIP 19610915 198603 2 001
Diketahui :
Ketua Departemen Biologi,
Dr. Ir. Ence Darmo Jaya Supena, M.Si. NIP 19641002 198903 1 002
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga penulis dapat menyelesaikan karya ilmiah ini. Judul yang dipilih dalam penelitian yang dilaksanakan pada bulan Agustus 2010 hingga Mei 2011 ini ialah Transformasi Genetik Nicotiana benthamiana dengan Gen Pembungaan Hd3a dari Padi.
Terima kasih penulis ucapkan kepada Bapak Prof. Dr. Ir. Suharsono, DEA dan Ibu Dr. Ir. Sri Koerniati, M.Sc selaku pembimbing atas segala bimbingan, dukungan, pengarahan, nasihat, kesabaran dan saran yang telah diberikan selama penelitian dan penulisan karya ilmiah ini. Terima kasih juga penulis ucapkan kepada Bapak Dr. Ir. Ahcmad Farajallah, M.Si selaku dosen penguji yang telah bersedia menguji dan memberikan saran serta masukan dalam penulisan karya ilmiah ini.
Penulis mengucapkan terima kasih kepada proyek penelitian KKP3T yang berjudul : “Perbaikan genetik Jatropha curcas untuk produksi biji dan toleransinya terhadap pH rendah dan aluminium melalui pendekatan biologi molekuler”dengan SPK no 642/LB.620/I.1/2/2009 tanggal 20 Februari 2009 atas nama Dr. Ir. Suharsono, DEA yang telah membiayai penelitian ini.
Terima kasih kepada Kepala Pusat Penelitian Sumberdaya Hayati dan Bioteknologi beserta seluruh staf dan karyawan atas sarana, prasarana, dan bantuannya selama penulis melakukan penelitian di Laboratorium Biorin dan Biologi Molekuler Seluler Tanaman. Penulis juga menyampaikan terima kasih kepada Pak Mulya, Mbak Pepi, Mbak Nia, Mbak Sarah, Pak Asep, dan Pak Iri atas bantuan, dan kerjasamanya. Terima kasih kepada teman-teman biologi angkatan 43. Terima kasih kepada rekan-rekan peneliti di Laboratorium Biorin dan Biologi Molekuler Seluler Tanaman yaitu Pak Muzuni, Bu Hanum, Bu Yohana, Pak Ulung, Bu Ratna, Pak Radit, Kak Nurul, Kak Anita, Kak Ophie, Fajri, Yulita, Indah, Iin, Rian, serta semua pihak yang tidak dapat disebutkan satu-persatu atas segala kerjasamanya, bantuan, nasihat, diskusi yang diberikan, saling menguatkan semangat, persahabatan, kekeluargaan dan keceriaan. Ungkapan terima kasih juga disampaikan kepada Bapak, Ibu, kakak, dan adik penulis, serta seluruh keluarga, atas segala doa, pengertian, dukungan, kesabaran, dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2012
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 5 Desember 1987 dari ayah Syafril MZ dan ibu Nursyamsi. Penulis merupakan anak ketiga dari empat bersaudara.
Tahun 2006 penulis lulus dari SMA Negeri 103 Jakarta dan pada tahun yang sama lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis terpilih masuk Program Studi Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... viii
DAFTAR LAMPIRAN ... viii
PENDAHULUAN Latar Belakang... 1
Tujuan Penelitian ... 1
BAHANDAN METODE Waktu dan Tempat Penelitian ... 1
Bahan ... 1
Metode ... 1
Persiapan Tanaman In Vitro ... 1
Penumbuhan Agrobacterium tumefaciens ... 2
Transformasi Genetik N. benthamiana ... 2
Seleksi, Regenerasi, dan Perbanyakan Tunas ... 2
Analisis Tanaman Transgenik ... 2
Analisis Molekuler ... 2
Analisis Fenotipe ... 3
HASIL Transformasi Genetik Nicotiana benthamiana ... 3
Analisis Tanaman Transgenik ... 4
PEMBAHASAN ... 5
SIMPULAN ... 6
SARAN ... 6
DAFTAR PUSTAKA ... 6
DAFTAR TABEL
Halaman
1 Rata-rata Efisiensi transformasi N. benthamiana ... 4
2 Rata-rata jumlah tunas transgenik putatif tiap eksplan ... 4
DAFTAR GAMBAR
Halaman 1 Peta fisik daerah T-DNA di dalam pCambia1300-Hd3a ... 12 Eksplan pada media seleksi yang mengandung higromisin pada umur 6 minggu setelah tanam ... 3
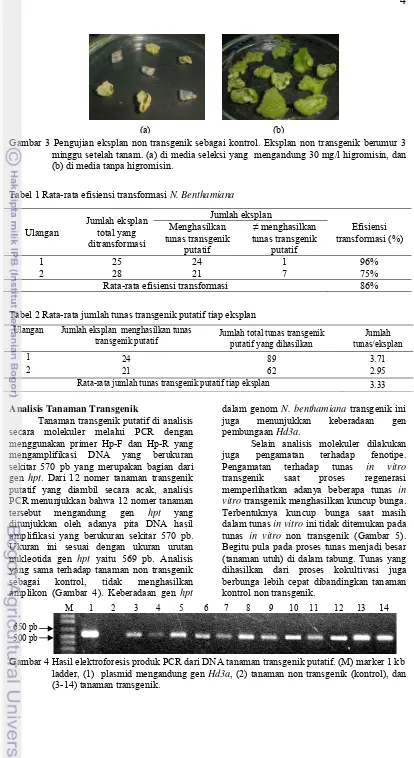
3 Pengujian eksplan non transgenik sebagai kontrol ... 4
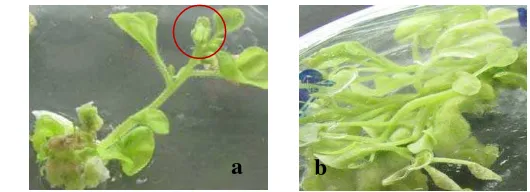
4 Hasil elektroforesis produk PCR dari DNA tanaman transgenik putatif ... 4
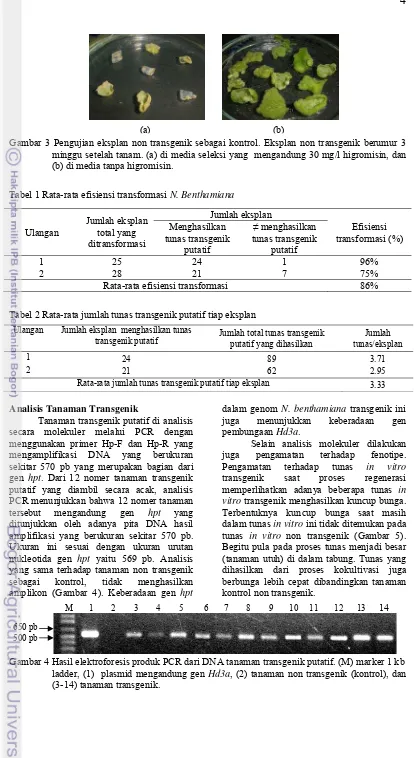
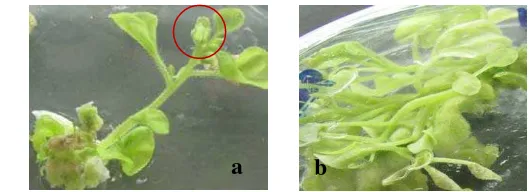
5 Pertumbuhan tunas in vitro pada umur 6 minggu setelah tanam ... 5
DAFTAR LAMPIRAN
Halaman 1 Komposisi media MS (Murashige & Skoog 1962) ... 8PENDAHULUAN
Latar Belakang
Pembungaan merupakan transisi dari fase vegetatif ke fase generatif/reproduksi. Pembungaan adalah salah satu peristiwa penting dalam siklus hidup tanaman tingkat tinggi dan karakter yang sangat penting untuk menentukan kemampuan spesies beradaptasi pada berbagai kondisi lingkungan. Adapun faktor lingkungan yang dapat mempengaruhi pembungaan meliputi panjang hari (fotoperiod), suhu, dan ketersediaan air.
Gen Hd3a adalah salah satu gen yang berhubungan dengan waktu pembungaan. Gen Hd3a yang berasal dari padi telah diisolasi oleh Tamaki et al. (2007). Gen ini menyandi aktivator utama pembungaan pada padi dalam kondisi hari pendek (Kojima et al. 2002; Tamaki et al. 2007). Selain gen Hd3a, pada Arabidopsis thaliana dikenal gen flowering locus T (FT) yang berperan memacu pembungaan pada tanaman hari panjang. Berdasarkan percobaan, ekspresi berlebih (overexpression) Hd3a dan FT menyebabkan pembungaan lebih awal pada padi dan juga pembungaan lebih awal pada A. thaliana. Kemiripan fungsi dan sekuen menunjukkan bahwa Hd3a merupakan ortolog dari FT (Kojima et al. 2002).
Transformasi genetik merupakan perubahan genetik karena adanya DNA asing yang masuk dan terintegrasi di dalam kromosom sel (hidup) inang. Transformasi genetik dilakukan untuk mengintegrasikan gen ke dalam sel tanaman untuk menghasilkan tanaman baru yang mampu mengekspresikan gen tersebut. Salah satu teknik transformasi genetik yang digunakan adalah melalui bantuan Agrobacterium tumefaciens. Teknik transformasi tersebut merupakan teknik transformasi secara tidak langsung yang paling sering digunakan. Teknik ini mempunyai beberapa keunggulan seperti efisiensi transformasi dengan salinan gen tunggal lebih tinggi dan dapat dilakukan dengan peralatan laboratorium yang sederhana.
Tanaman Nicotiana benthamiana merupakan tanaman model yang biasa digunakan untuk menguji peranan suatu gen.
Tujuan Penelitian
Penelitian ini bertujuan mendapatkan tanaman Nicotiana benthamiana transgenik yang mengandung Hd3a.
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan pada bulan Agustus 2010 − Mei 2011 di Laboratorium BIORIN (Biotechnology Research Indonesia-The Netherlands) dan Biologi Molekular Seluler Tanaman, Pusat Penelitan Sumberdaya Hayati dan Bioteknologi IPB (PPSHB-IPB), Institut Pertanian Bogor.
Bahan
Nicotiana benthamiana yang ditumbuhkan secara in vitro digunakan sebagai bahan tanaman yang di transformasi secara genetik. Agrobacterium tumefaciens galur LBA4404 yang mengandung plasmid pCambia 1300-Hd3a yang membawa gen Hd3a di bawah kontrol promoter 35S CaMV (Sulistyaningsih 2012) digunakan untuk melakukan transformasi genetik. Primer Hp-F (5’AAGGAATCGGTCAATACACTAC3’) dan Hp-R (5’ACTATCGGCGAGTACTTC TACA3’) digunakan untuk menganalisis keberadaan gen hpt di dalam tanaman transgenik putatif. Primer Hp-F didesain berdasarkan urutan nukleotida ke- 416-437 dan primer Hp-R didesain berdasarkan urutan nukleotida ke- 963-984 dari gen hpt (Hannum 2012). Peta fisik daerah T-DNA disajikan pada Gambar 1.
Metode
Persiapan Tanaman In Vitro
Biji N. benthamiana disterilisasi dalam laminar air flow dengan cara merendam biji dalam alkohol 70% selama 2 menit. Selanjutnya biji dicuci menggunakan aquades steril sebanyak satu kali, lalu direndam dalam 20% larutan bayclean (atau 1,05% NaClO) selama 5-10 menit. Kemudian biji dicuci kembali menggunakan aquades steril sebanyak 3-4 kali. Biji yang sudah disterilisasi ditanam dalam media dasar
Gambar 1 Peta fisik daerah T-DNA di dalam
pCambia 1300-Hd3a
. LB: left border, RB: right border, 35S pro: promoter 35S dari Cauliflower mosaic virus (CaMV), Hd3a: gen Hd3a daripadi, hpt: gen resistensi terhadaphigromisin(Sulistyaningsih 2012). L B hpt 35S Pro R B
2
Murashige dan Skoog (MS) (Lampiran 1), yang mengandung 3 g/l phytagel, dan diletakkan di dalam ruang gelap selama 3 hari, kemudian dipindahkan ke dalam ruangan bercahaya, suhu 26-27oC. Setelah biji berkecambah, tunas disubkultur ke botol kultur dan dipelihara dalam ruang kultur selama satu bulan.
Penumbuhan Agrobacterium tumefaciens Agrobacterium tumefaciens galur LBA4404 yang membawa gen Hd3a di dalam plasmid pC1300-Hd3a diperbanyak dengan menumbuhkan satu koloni tunggal di dalam 10 ml media LB (1% tripton, 0,5% yeast extract, 1% NaCl) cair yang mengandung 50 mg/l kanamisin dan 25 mg/l rifampisin. Biakan digoyang menggunakan inkubator bergoyang pada suhu ruang selama semalam (kondisi gelap) hingga OD600
mencapai 0,5-0,8.
Transformasi genetik N. benthamiana
Sebelum melakukan transformasi, biakan A. tumefaciens diendapkan menggunakan alat sentrifuse pada kecepatan 5000 rpm selama 10 menit dan endapan diresuspensi dengan penambahan 10 ml media Murashige dan Skoog (MS) cair yang mengandung 0,5 mg/l BAP dan 100 mg/l asetosiringon hingga OD600 mencapai 0,5-0,8.
Transformasi dilakukan dengan metode kokultivasi menggunakan A. tumefaciens menurut prosedur Horsch et al. (1985) yang dimodifikasi. Daun-daun N. benthamiana yang diperoleh dari kultur in vitro yang berumur 4 minggu dipotong menjadi berukuran ± 1 cm2 dan direndam dalam suspensi A. tumefaciens OD600 0,5-0,8,
digoyang menggunakan inkubator bergoyang pada suhu ruang selama 10-15 menit. Selanjutnya eksplan dikeringkan dengan tissu steril dan ditanam pada media MS padat yang mengandung 0,5 mg/l BAP, dan 100 mg/l asetosiringon selama 3 hari di dalam ruang gelap.
Seleksi, Regenerasi, dan Perbanyakan Tunas
Setelah 3 hari, eksplan dicuci dengan aquades steril sebanyak tiga kali dan dengan larutan 200 mg/l cefotaxime. Eksplan dikeringkan dengan tissu steril dan dipindahkan ke media seleksi yaitu media MS yang mengandung 0,5 mg/l BAP, 30 mg/l higromisin, dan 200 mg/l cefotaxime. Selanjutnya, eksplan di simpan dalam ruang kultur pada suhu 26-27oC. Tunas yang
terbentuk dipindahkan ke dalam media MS yang mengandung 30 mg/l higromisin, dan 200 mg/l cefotaxime hingga tunas menjadi besar. Eksplan yang menghasilkan tunas transgenik putatif dihitung untuk mengetahui efisiensi transformasi. Rumus efisiensi transformasi disajikan pada Lampiran 2.
Eksplan non transgenik (sebagai kontrol) yang tidak di kokultivasi dengan A. tumefaciens diperlakukan dengan metode dan kondisi yang sama dengan eksplan yang di kokultivasi dengan A. tumefaciens yang di tanam pada dua media perlakuan yaitu mengandung higromisin dan tanpa higromisin.
Analisis Tanaman Transgenik Analisis Molekuler
ditambah dengan RNAse (10 µg/µl) sebanyak 0,1 x volume dan diinkubasikan pada suhu 37oC selama semalam.
Analisis molekuler untuk mengetahui keberadaan gen hpt (hygromycin phosphotransferase) dilakukan dengan PCR (Polymerase Chain Reaction). PCR dilakukan dengan total volume 10 µl yang mengandung 100 ng DNA total (1 µl), 10 pmol primer Hp-F (0,5 µl), 10 pmol primer Hp-R (0,5 µl), 2 mM dNTP (1 µl), 1 unit taq polimerase (0,2 µl), 1x buffer (1 µl) dan air bebas ion (ddH2O) hingga mencapai volume
10 µl. Kondisi PCR adalah pra-PCR 95oC selama 5 menit, denaturasi 94oC selama 30 detik, annealing (penempelan) 56oC selama 30 detik, extension (pemanjangan) 72oC selama 1 menit, pasca-PCR 72oC selama 5 menit. PCR dilakukan sebanyak 30 siklus. Hasil PCR kemudian dicek dengan elektroforesis di gel agarosa 1% dalam larutan buffer penyangga 1xTAE (40 mM Tris-acetate, 1 mM EDTA) pada tegangan 100 volt selama 27 menit. Pita DNA pada agarosa divisualisasi dengan cahaya UV setelah gel direndam di larutan ethidium bromida (0,5 µg/ml).
Analisis Fenotipe
Analisis fenotipe dilakukan dengan pengamatan terhadap munculnya kuncup bunga pada tunas in vitro.
HASIL
Transformasi Genetik Nicotiana
benthamiana
Eksplan N. benthamiana yang telah dikokultivasi dengan A. tumefaciens yang mengandung pC1300-Hd3a yang mampu
bertahan hidup di dalam media seleksi yang
mengandung 30 mg/l higromisin membentuk
tonjolan kalus mulai minggu ke-2. Pada minggu ke-4 dan ke-5, eksplan mulai menghasilkan tunas dari tonjolan kalus
tersebut. Sedangkan eksplan yang tidak
dikokultivasi dengan A. tumefaciens yang di tanam di dalam media seleksi yang sama mengalami kematian dan tidak menghasilkan tunas (Gambar 2). Eksplan yang mati menunjukkan tidak mempunyai gen resistensi terhadap higromisin.
Untuk mengetahui efisiensi media seleksi, eksplan non transgenik (sebagai kontrol) yang tidak dikokultivasi dengan A. tumefaciens yang mengandung pC1300-Hd3a ditanam di dua media yang berbeda yaitu media yang mengandung higromisin dan media tanpa higromisin. Eksplan yang ditanam pada media yang mengandung higromisin mengalami kematian dan tidak mampu menghasilkan tunas, sedangkan eksplan yang ditanam pada media tanpa higromisin tumbuh dengan baik dan mampu menghasilkan tunas (Gambar 3).
Tunas yang tumbuh dari eksplan di media seleksi adalah tunas transgenik putatif. Berdasarkan jumlah eksplan yang menghasilkan tunas transgenik putatif, rata-rata efisiensi transformasi pada N. benthamiana adalah 86% (Tabel 1). Kemampuan eksplan menghasilkan tunas dalam media yang mengandung higromisin dapat menunjukkan eksplan tersebut mengandung gen hpt yang difusikan dengan gen Hd3a.
Setiap eksplan transforman menghasilkan jumlah tunas berbeda. Rata-rata jumlah tunas transgenik tiap eksplan berkisar 3 – 4 tunas (Tabel 2).
(b) (a)
4
Gambar 3 Pengujian eksplan non transgenik sebagai kontrol. Eksplan non transgenik berumur 3 minggu setelah tanam. (a) di media seleksi yang mengandung 30 mg/l higromisin, dan (b) di media tanpa higromisin.
Tabel 1 Rata-rata efisiensi transformasi N. Benthamiana
Tabel 2 Rata-rata jumlah tunas transgenik putatif tiap eksplan
Analisis Tanaman Transgenik
Tanaman transgenik putatif di analisis secara molekuler melalui PCR dengan menggunakan primer Hp-F dan Hp-R yang mengamplifikasi DNA yang berukuran sekitar 570 pb yang merupakan bagian dari gen hpt. Dari 12 nomer tanaman transgenik putatif yang diambil secara acak, analisis PCR menunjukkan bahwa 12 nomer tanaman tersebut mengandung gen hpt yang ditunjukkan oleh adanya pita DNA hasil amplifikasi yang berukuran sekitar 570 pb. Ukuran ini sesuai dengan ukuran urutan nukleotida gen hpt yaitu 569 pb. Analisis yang sama terhadap tanaman non transgenik sebagai kontrol, tidak menghasilkan amplikon (Gambar 4). Keberadaan gen hpt
dalam genom N. benthamiana transgenik ini juga menunjukkan keberadaan gen pembungaan Hd3a.
Selain analisis molekuler dilakukan juga pengamatan terhadap fenotipe. Pengamatan terhadap tunas in vitro transgenik saat proses regenerasi memperlihatkan adanya beberapa tunas in vitro transgenik menghasilkan kuncup bunga. Terbentuknya kuncup bunga saat masih dalam tunas in vitro ini tidak ditemukan pada tunas in vitro non transgenik (Gambar 5). Begitu pula pada proses tunas menjadi besar (tanaman utuh) di dalam tabung. Tunas yang dihasilkan dari proses kokultivasi juga berbunga lebih cepat dibandingkan tanaman kontrol non transgenik.
Gambar 4 Hasil elektroforesis produk PCR dari DNA tanaman transgenik putatif. (M) marker 1 kb ladder, (1) plasmid mengandung gen Hd3a, (2) tanaman non transgenik (kontrol), dan (3-14) tanaman transgenik.
Ulangan
Jumlah eksplan total yang ditransformasi
Jumlah eksplan
Efisiensi transformasi (%) Menghasilkan
tunas transgenik putatif
≠ menghasilkan tunas transgenik
putatif
1 25 24 1 96%
2 28 21 7 75%
Rata-rata efisiensi transformasi 86%
Ulangan Jumlah eksplan menghasilkan tunas
transgenik putatif Jumlah total tunas transgenik putatif yang dihasilkan tunas/eksplan Jumlah
1 24 89 3.71
2 21 62 2.95
Rata-rata jumlah tunas transgenik putatif tiap eksplan 3.33
(b) (a)
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Gambar 5 Pertumbuhan tunas in vitro pada umur 6 minggu setelah tanam. (a) tunas transgenik yang berbunga, dan (b) tunas non transgenik.
PEMBAHASAN
Nicotiana banyak digunakan sebagai tanaman model untuk transformasi genetik karena tanaman ini mudah diregenerasikan, mudah tumbuh dan berumur relatif pendek. Chaidamsari et al. (2006) telah menggunakan N. tabacum untuk menguji ekspresi gen APETALA1 dari tanaman coklat, sedangkan penelitian ini menggunakan N. benthamiana untuk menguji ekspresi gen pembungaan Hd3a dari tanaman padi.
Regenerasi tanaman pada penelitian ini dilakukan melalui organogenesis yaitu pembentukan organ atau tunas langsung dari eksplan tanpa melalui pembentukan kalus terlebih dahulu. Eksplan pada penelitian ini adalah daun. Untuk melakukan proses transformasi, daun N. benthamiana terlebih dahulu dilukai dengan dipotong menjadi beberapa bagian. Pelukaan pada tanaman akan menyebabkan sel pada bagian tanaman terluka mengeluarkan senyawa kelompok fenol yang dikenal dengan nama asetosiringon. Asetosiringon berperan menginduksi gen-gen vir yang berfungsi dalam mentransfer T-DNA ke dalam sel tanaman dan penambahan asetosiringon meningkatkan efisiensi infeksi Agrobacterium (Orlikowska et al. 1995), sehingga dapat meningkatkan jumlah sel yang transforman. Untuk meningkatkan efisiensi transformasi, pada penelitian ini asetosiringon juga ditambahkan dalam media kokultivasi. Menurut Ozawa (2009), penggunaan asetosiringon merupakan salah satu kondisi terbaik yang perlu diciptakan agar transformasi lebih efektif.
Transformasi genetik pada penelitian ini menggunakan gen Hd3a di bawah kendali promoter 35S CaMV yang difusikan dengan gen resistensi terhadap higromisin yang merupakan gen penanda seleksi. Gen penanda seleksi sangat penting dalam kegiatan transformasi genetik karena berguna
untuk menyeleksi sel, jaringan, organ atau tanaman yang sudah mengalami transformasi (transgenik). Penelitian ini menggunakan 30 mg/l higromisin untuk seleksi karena pada konsentrasi ini eksplan non transgenik tidak mampu melakukan regenerasi untuk membentuk tunas, dan kemudian eksplan mengalami kematian (Gambar 2 dan 3). Berdasarkan jumlah eksplan yang beregenerasi di media seleksi, rata-rata efisiensi transformasi pada penelitian ini yaitu 86%. Efisiensi transformasi ini adalah tinggi jika dibandingkan dengan transformasi pada Jatropha curcas L. (Sulistyaningsih 2012).
Analisis PCR dilakukan dengan menggunakan primer untuk gen penyeleksi hpt. Posisi gen hpt pada daerah T-DNA dalam pC1300-Hd3a berdampingan dengan LB (batas kiri) dan Hd3a berdampingan dengan RB (batas kanan) (Gambar 1). Keberadaan gen hpt dapat merupakan indikasi keberadaan gen lain dalam satu T-DNA yang sama di dalam genom tanaman transgenik. Hal ini disebabkan oleh proses integrasi daerah T-DNA di dalam genom tanaman yang dimulai dari RB (Sheng&Citovsky 1996). Dengan demikian jika hasil PCR menunjukkan keberadaan gen hpt dalam genom tanaman transgenik maka gen target sisipan Hd3a juga telah terintegrasi dalam genom tanaman tersebut. Hasil PCR terhadap 12 tunas yang diduga transgenik yang dapat hidup di media seleksi yang diambil secara acak menunjukkan bahwa kedua belas tanaman tersebut mengandung gen hpt yang bertanggungjawab terhadap resistensi tanaman terhadap higromisin yang ditunjukkan oleh adanya pita DNA hasil amplifikasi yang berukuran sekitar 570 pb.
Pengamatan fenotipe terhadap tanaman transgenik menunjukkan bahwa N. benthamiana transgenik menghasilkan kuncup bunga pada saat proses regenerasi tunas dan berbunga pada saat tunas di dalam
8
tabung (Gambar 5), sedangkan tunas non transgenik tidak dapat menghasilkan kuncup bunga pada proses tersebut. Hal ini menunjukkan bahwa gen Hd3a dapat memicu pembungaan pada N. benthamiana. Hasil ini memperkuat penelitian sebelumnya pada padi. Padi transgenik yang mengekspresikan gen Hd3a berbunga lebih cepat dari pada padi tipe liarnya pada kondisi hari pendek (Kojima et al. 2002). Ekspresi gen Hd3a di Jatropha curcas juga menghasilkan tunas yang berbunga sangat dini pada saat tunas masih di dalam tabung. Pembungaan dini ini disebabkan oleh ekspresi gen Hd3a di bawah kendali promoter konstitutif p35S CaMV (Sulistyaningsih 2012) .
SIMPULAN
Proses transformasi Nicotiana benthamiana melalui Agrobacterium tumefaciens telah berhasil dilakukan dengan nilai efisiensi transformasi cukup tinggi. Analisis molekuler dengan PCR terhadap tanaman transgenik putatif menunjukkan bahwa tunas yang tumbuh di media seleksi mengandung gen hpt. Karena gen hpt difusikan dengan gen Hd3a, maka tanaman transgenik ini mengandung gen Hd3a. Tunas transgenik yang mengandung gen Hd3a berbunga awal, sehingga gen Hd3a mempunyai peranan dalam memicu pembungaan.
SARAN
Tanaman transgenik perlu diaklimatisasi hingga menghasilkan biji generasi T1 untuk analisis segregasi dan kestabilan transgen, serta ekspresi gen.
DAFTAR PUSTAKA
Chaidamsari T, Samanhudi, Budiani A, Poerwanto R, Santoso D. 2006. Ekspresi fenotipe gen APETALAI kakao (TcAPI) pada eksplan tembakau. Menara Perkebunan 74(1): 1-9.
Hannum S. 2012. Isolasi, pengklonan, dan analisis ekspresi gen penyandi copper-zinc superoxide dismutase (CuZn-SOD) dari Melastoma malabathricum
L. [Disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor. Horsch RB, Fry JE, Hoffman NL, Eichholtz
D, Rogers SG, Fraley RT. 1985. A simple and general method for transferring genes into plants. Science 227 : 1229-1231.
Kojima S, Takahashi Y, Kobayashi Y, Monna L, Sasaki T, Araki T, Yano M. 2002. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant CellPhysiol. 43: 1096-1105.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassay with tobacco tissue cultures. Plant Physiol 15:473-497.
Orlikowska TK, Cranston HJ, Dyer WE. 1995. Factor influencing Agrobacterium tumefaciens–mediated transformation and regeneration of the sunflower cultivar contennial. Plant Cell Tiss. & Org. Cult. 40: 85-91.
Ozawa K. 2009. Establishment of a high efficiency Agrobacterium mediated transformation system of rice (Oryza sativa L.). Plant Science 176: 522-527.
Sheng J, Citovsky V. 1996. Agrobacterium -plant cell DNA transport: have virulence protein, will travel. Plant cell 8: 1699-1710.
Suharsono. 2002. Konstruksi pustaka genom kedelai kultivar Slamet. Hayati 9(3):67-70.
Sulistyaningsih YC. 2012. Rekayasa ekspresi gen pembungaan Hd3a pada tanaman Jarak Pagar (Jatropha curcas L.) [Disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor. Tamaki S, Matsuo S, Wong HL, Yokoi S,
8
Lampiran 1 Komposisi media MS (Murashige & Skoog 1962)
Bahan Konsentrasi
(mg/l)
Hara makro NH4NO3 1650
CaCl2.2H2O 440
MgSO4.7H2O 370
KNO3 1900
KH2PO4 170
Hara mikro H3BO3 6,2
Na2MoO4.2H2O 0,25
CoCl2.6H2O 0,025
KI 0,83
ZnSO4.7H2O 8,6
MnSO4.4H2O 22,3
CuSO4.5H2O 0,025
Na2EDTA 37,2
FeSO4.7H20 27,8
Vitamin Myo-Inositol 100
Pyridoxin-HCl 0,5
Nicotinic acid 0,5
Thiamin-HCl 0,1
Glycine 2
Sukrosa 30 g/l
9
Lampiran 2 Perhitungan efisiensi transformasi dan jumlah tunas transgenik putatif tiap eksplan
% efisiensi transformasi = jumlah eksplan menghasilkan tunas transgenik putatif
jumlah eksplan total yang ditransformasi × 100%
jumlah tunas transgenik putatif tiap eksplan = jumlah total tunas transgenik putatif
ABSTRAK
LAILA NUR SYAFITRI. Transformasi Genetik Nicotiana benthamiana dengan Gen Pembungaan Hd3a dari Padi. Dibimbing oleh SUHARSONO dan SRI KOERNIATI.
Pembungaan adalah salah satu peristiwa penting dalam siklus hidup tanaman tingkat tinggi dan karakter yang sangat penting untuk menentukan kemampuan spesies beradaptasi pada berbagai kondisi lingkungan. Gen Hd3a adalah salah satu gen yang berhubungan dengan waktu pembungaan di padi. Ekspresi berlebih Hd3a menyebabkan pembungaan lebih awal pada padi. Penelitian ini bertujuan untuk melakukan transformasi genetik Nicotiana benthamiana dengan gen Hd3a dari padi. Transformasi genetik dilakukan melalui bantuan Agrobacterium tumefaciens LBA4404 dengan metode kokultivasi. Seleksi tunas transgenik putatif dilakukan dengan 30 mg/l higromisin. Hasil penelitian menunjukkan bahwa proses transformasi N. benthamiana menghasilkan tunas transgenik putatif dengan nilai efisiensi transformasi cukup tinggi yaitu 86%. Analisis molekuler dengan PCR terhadap tanaman transgenik putatif menunjukkan bahwa tanaman tersebut mengandung gen hpt yang bertanggung jawab terhadap resistensi tanaman terhadap higromisin. Keberadaan bagian gen hpt dalam genom Nicotiana benthamiana transgenik ini juga menunjukkan keberadaan gen pembungaan Hd3a. Tunas Nicotiana benthamiana transgenik yang mengandung gen Hd3a berbunga lebih awal, sehingga gen Hd3a mempunyai peranan dalam memicu pembungaan.
ABSTRACT
LAILA NUR SYAFITRI. Genetic Transformation of Nicotiana benthamiana with Hd3a flowering gene from rice. Supervised by SUHARSONO and SRI KOERNIATI.
PENDAHULUAN
Latar Belakang
Pembungaan merupakan transisi dari fase vegetatif ke fase generatif/reproduksi. Pembungaan adalah salah satu peristiwa penting dalam siklus hidup tanaman tingkat tinggi dan karakter yang sangat penting untuk menentukan kemampuan spesies beradaptasi pada berbagai kondisi lingkungan. Adapun faktor lingkungan yang dapat mempengaruhi pembungaan meliputi panjang hari (fotoperiod), suhu, dan ketersediaan air.
Gen Hd3a adalah salah satu gen yang berhubungan dengan waktu pembungaan. Gen Hd3a yang berasal dari padi telah diisolasi oleh Tamaki et al. (2007). Gen ini menyandi aktivator utama pembungaan pada padi dalam kondisi hari pendek (Kojima et al. 2002; Tamaki et al. 2007). Selain gen Hd3a, pada Arabidopsis thaliana dikenal gen flowering locus T (FT) yang berperan memacu pembungaan pada tanaman hari panjang. Berdasarkan percobaan, ekspresi berlebih (overexpression) Hd3a dan FT menyebabkan pembungaan lebih awal pada padi dan juga pembungaan lebih awal pada A. thaliana. Kemiripan fungsi dan sekuen menunjukkan bahwa Hd3a merupakan ortolog dari FT (Kojima et al. 2002).
Transformasi genetik merupakan perubahan genetik karena adanya DNA asing yang masuk dan terintegrasi di dalam kromosom sel (hidup) inang. Transformasi genetik dilakukan untuk mengintegrasikan gen ke dalam sel tanaman untuk menghasilkan tanaman baru yang mampu mengekspresikan gen tersebut. Salah satu teknik transformasi genetik yang digunakan adalah melalui bantuan Agrobacterium tumefaciens. Teknik transformasi tersebut merupakan teknik transformasi secara tidak langsung yang paling sering digunakan. Teknik ini mempunyai beberapa keunggulan seperti efisiensi transformasi dengan salinan gen tunggal lebih tinggi dan dapat dilakukan dengan peralatan laboratorium yang sederhana.
Tanaman Nicotiana benthamiana merupakan tanaman model yang biasa digunakan untuk menguji peranan suatu gen.
Tujuan Penelitian
Penelitian ini bertujuan mendapatkan tanaman Nicotiana benthamiana transgenik yang mengandung Hd3a.
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan pada bulan Agustus 2010 − Mei 2011 di Laboratorium BIORIN (Biotechnology Research Indonesia-The Netherlands) dan Biologi Molekular Seluler Tanaman, Pusat Penelitan Sumberdaya Hayati dan Bioteknologi IPB (PPSHB-IPB), Institut Pertanian Bogor.
Bahan
Nicotiana benthamiana yang ditumbuhkan secara in vitro digunakan sebagai bahan tanaman yang di transformasi secara genetik. Agrobacterium tumefaciens galur LBA4404 yang mengandung plasmid pCambia 1300-Hd3a yang membawa gen Hd3a di bawah kontrol promoter 35S CaMV (Sulistyaningsih 2012) digunakan untuk melakukan transformasi genetik. Primer Hp-F (5’AAGGAATCGGTCAATACACTAC3’) dan Hp-R (5’ACTATCGGCGAGTACTTC TACA3’) digunakan untuk menganalisis keberadaan gen hpt di dalam tanaman transgenik putatif. Primer Hp-F didesain berdasarkan urutan nukleotida ke- 416-437 dan primer Hp-R didesain berdasarkan urutan nukleotida ke- 963-984 dari gen hpt (Hannum 2012). Peta fisik daerah T-DNA disajikan pada Gambar 1.
Metode
Persiapan Tanaman In Vitro
Biji N. benthamiana disterilisasi dalam laminar air flow dengan cara merendam biji dalam alkohol 70% selama 2 menit. Selanjutnya biji dicuci menggunakan aquades steril sebanyak satu kali, lalu direndam dalam 20% larutan bayclean (atau 1,05% NaClO) selama 5-10 menit. Kemudian biji dicuci kembali menggunakan aquades steril sebanyak 3-4 kali. Biji yang sudah disterilisasi ditanam dalam media dasar
Gambar 1 Peta fisik daerah T-DNA di dalam
pCambia 1300-Hd3a
. LB: left border, RB: right border, 35S pro: promoter 35S dari Cauliflower mosaic virus (CaMV), Hd3a: gen Hd3a daripadi, hpt: gen resistensi terhadaphigromisin(Sulistyaningsih 2012). L B hpt 35S Pro R B
2
Murashige dan Skoog (MS) (Lampiran 1), yang mengandung 3 g/l phytagel, dan diletakkan di dalam ruang gelap selama 3 hari, kemudian dipindahkan ke dalam ruangan bercahaya, suhu 26-27oC. Setelah biji berkecambah, tunas disubkultur ke botol kultur dan dipelihara dalam ruang kultur selama satu bulan.
Penumbuhan Agrobacterium tumefaciens Agrobacterium tumefaciens galur LBA4404 yang membawa gen Hd3a di dalam plasmid pC1300-Hd3a diperbanyak dengan menumbuhkan satu koloni tunggal di dalam 10 ml media LB (1% tripton, 0,5% yeast extract, 1% NaCl) cair yang mengandung 50 mg/l kanamisin dan 25 mg/l rifampisin. Biakan digoyang menggunakan inkubator bergoyang pada suhu ruang selama semalam (kondisi gelap) hingga OD600
mencapai 0,5-0,8.
Transformasi genetik N. benthamiana
Sebelum melakukan transformasi, biakan A. tumefaciens diendapkan menggunakan alat sentrifuse pada kecepatan 5000 rpm selama 10 menit dan endapan diresuspensi dengan penambahan 10 ml media Murashige dan Skoog (MS) cair yang mengandung 0,5 mg/l BAP dan 100 mg/l asetosiringon hingga OD600 mencapai 0,5-0,8.
Transformasi dilakukan dengan metode kokultivasi menggunakan A. tumefaciens menurut prosedur Horsch et al. (1985) yang dimodifikasi. Daun-daun N. benthamiana yang diperoleh dari kultur in vitro yang berumur 4 minggu dipotong menjadi berukuran ± 1 cm2 dan direndam dalam suspensi A. tumefaciens OD600 0,5-0,8,
digoyang menggunakan inkubator bergoyang pada suhu ruang selama 10-15 menit. Selanjutnya eksplan dikeringkan dengan tissu steril dan ditanam pada media MS padat yang mengandung 0,5 mg/l BAP, dan 100 mg/l asetosiringon selama 3 hari di dalam ruang gelap.
Seleksi, Regenerasi, dan Perbanyakan Tunas
Setelah 3 hari, eksplan dicuci dengan aquades steril sebanyak tiga kali dan dengan larutan 200 mg/l cefotaxime. Eksplan dikeringkan dengan tissu steril dan dipindahkan ke media seleksi yaitu media MS yang mengandung 0,5 mg/l BAP, 30 mg/l higromisin, dan 200 mg/l cefotaxime. Selanjutnya, eksplan di simpan dalam ruang kultur pada suhu 26-27oC. Tunas yang
terbentuk dipindahkan ke dalam media MS yang mengandung 30 mg/l higromisin, dan 200 mg/l cefotaxime hingga tunas menjadi besar. Eksplan yang menghasilkan tunas transgenik putatif dihitung untuk mengetahui efisiensi transformasi. Rumus efisiensi transformasi disajikan pada Lampiran 2.
Eksplan non transgenik (sebagai kontrol) yang tidak di kokultivasi dengan A. tumefaciens diperlakukan dengan metode dan kondisi yang sama dengan eksplan yang di kokultivasi dengan A. tumefaciens yang di tanam pada dua media perlakuan yaitu mengandung higromisin dan tanpa higromisin.
Analisis Tanaman Transgenik Analisis Molekuler
ditambah dengan RNAse (10 µg/µl) sebanyak 0,1 x volume dan diinkubasikan pada suhu 37oC selama semalam.
Analisis molekuler untuk mengetahui keberadaan gen hpt (hygromycin phosphotransferase) dilakukan dengan PCR (Polymerase Chain Reaction). PCR dilakukan dengan total volume 10 µl yang mengandung 100 ng DNA total (1 µl), 10 pmol primer Hp-F (0,5 µl), 10 pmol primer Hp-R (0,5 µl), 2 mM dNTP (1 µl), 1 unit taq polimerase (0,2 µl), 1x buffer (1 µl) dan air bebas ion (ddH2O) hingga mencapai volume
10 µl. Kondisi PCR adalah pra-PCR 95oC selama 5 menit, denaturasi 94oC selama 30 detik, annealing (penempelan) 56oC selama 30 detik, extension (pemanjangan) 72oC selama 1 menit, pasca-PCR 72oC selama 5 menit. PCR dilakukan sebanyak 30 siklus. Hasil PCR kemudian dicek dengan elektroforesis di gel agarosa 1% dalam larutan buffer penyangga 1xTAE (40 mM Tris-acetate, 1 mM EDTA) pada tegangan 100 volt selama 27 menit. Pita DNA pada agarosa divisualisasi dengan cahaya UV setelah gel direndam di larutan ethidium bromida (0,5 µg/ml).
Analisis Fenotipe
Analisis fenotipe dilakukan dengan pengamatan terhadap munculnya kuncup bunga pada tunas in vitro.
HASIL
Transformasi Genetik Nicotiana
benthamiana
Eksplan N. benthamiana yang telah dikokultivasi dengan A. tumefaciens yang mengandung pC1300-Hd3a yang mampu
bertahan hidup di dalam media seleksi yang
mengandung 30 mg/l higromisin membentuk
tonjolan kalus mulai minggu ke-2. Pada minggu ke-4 dan ke-5, eksplan mulai menghasilkan tunas dari tonjolan kalus
tersebut. Sedangkan eksplan yang tidak
dikokultivasi dengan A. tumefaciens yang di tanam di dalam media seleksi yang sama mengalami kematian dan tidak menghasilkan tunas (Gambar 2). Eksplan yang mati menunjukkan tidak mempunyai gen resistensi terhadap higromisin.
Untuk mengetahui efisiensi media seleksi, eksplan non transgenik (sebagai kontrol) yang tidak dikokultivasi dengan A. tumefaciens yang mengandung pC1300-Hd3a ditanam di dua media yang berbeda yaitu media yang mengandung higromisin dan media tanpa higromisin. Eksplan yang ditanam pada media yang mengandung higromisin mengalami kematian dan tidak mampu menghasilkan tunas, sedangkan eksplan yang ditanam pada media tanpa higromisin tumbuh dengan baik dan mampu menghasilkan tunas (Gambar 3).
Tunas yang tumbuh dari eksplan di media seleksi adalah tunas transgenik putatif. Berdasarkan jumlah eksplan yang menghasilkan tunas transgenik putatif, rata-rata efisiensi transformasi pada N. benthamiana adalah 86% (Tabel 1). Kemampuan eksplan menghasilkan tunas dalam media yang mengandung higromisin dapat menunjukkan eksplan tersebut mengandung gen hpt yang difusikan dengan gen Hd3a.
Setiap eksplan transforman menghasilkan jumlah tunas berbeda. Rata-rata jumlah tunas transgenik tiap eksplan berkisar 3 – 4 tunas (Tabel 2).
(b) (a)
4
Gambar 3 Pengujian eksplan non transgenik sebagai kontrol. Eksplan non transgenik berumur 3 minggu setelah tanam. (a) di media seleksi yang mengandung 30 mg/l higromisin, dan (b) di media tanpa higromisin.
Tabel 1 Rata-rata efisiensi transformasi N. Benthamiana
Tabel 2 Rata-rata jumlah tunas transgenik putatif tiap eksplan
Analisis Tanaman Transgenik
Tanaman transgenik putatif di analisis secara molekuler melalui PCR dengan menggunakan primer Hp-F dan Hp-R yang mengamplifikasi DNA yang berukuran sekitar 570 pb yang merupakan bagian dari gen hpt. Dari 12 nomer tanaman transgenik putatif yang diambil secara acak, analisis PCR menunjukkan bahwa 12 nomer tanaman tersebut mengandung gen hpt yang ditunjukkan oleh adanya pita DNA hasil amplifikasi yang berukuran sekitar 570 pb. Ukuran ini sesuai dengan ukuran urutan nukleotida gen hpt yaitu 569 pb. Analisis yang sama terhadap tanaman non transgenik sebagai kontrol, tidak menghasilkan amplikon (Gambar 4). Keberadaan gen hpt
dalam genom N. benthamiana transgenik ini juga menunjukkan keberadaan gen pembungaan Hd3a.
Selain analisis molekuler dilakukan juga pengamatan terhadap fenotipe. Pengamatan terhadap tunas in vitro transgenik saat proses regenerasi memperlihatkan adanya beberapa tunas in vitro transgenik menghasilkan kuncup bunga. Terbentuknya kuncup bunga saat masih dalam tunas in vitro ini tidak ditemukan pada tunas in vitro non transgenik (Gambar 5). Begitu pula pada proses tunas menjadi besar (tanaman utuh) di dalam tabung. Tunas yang dihasilkan dari proses kokultivasi juga berbunga lebih cepat dibandingkan tanaman kontrol non transgenik.
Gambar 4 Hasil elektroforesis produk PCR dari DNA tanaman transgenik putatif. (M) marker 1 kb ladder, (1) plasmid mengandung gen Hd3a, (2) tanaman non transgenik (kontrol), dan (3-14) tanaman transgenik.
Ulangan
Jumlah eksplan total yang ditransformasi
Jumlah eksplan
Efisiensi transformasi (%) Menghasilkan
tunas transgenik putatif
≠ menghasilkan tunas transgenik
putatif
1 25 24 1 96%
2 28 21 7 75%
Rata-rata efisiensi transformasi 86%
Ulangan Jumlah eksplan menghasilkan tunas
transgenik putatif Jumlah total tunas transgenik putatif yang dihasilkan tunas/eksplan Jumlah
1 24 89 3.71
2 21 62 2.95
Rata-rata jumlah tunas transgenik putatif tiap eksplan 3.33
(b) (a)
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Gambar 5 Pertumbuhan tunas in vitro pada umur 6 minggu setelah tanam. (a) tunas transgenik yang berbunga, dan (b) tunas non transgenik.
PEMBAHASAN
Nicotiana banyak digunakan sebagai tanaman model untuk transformasi genetik karena tanaman ini mudah diregenerasikan, mudah tumbuh dan berumur relatif pendek. Chaidamsari et al. (2006) telah menggunakan N. tabacum untuk menguji ekspresi gen APETALA1 dari tanaman coklat, sedangkan penelitian ini menggunakan N. benthamiana untuk menguji ekspresi gen pembungaan Hd3a dari tanaman padi.
Regenerasi tanaman pada penelitian ini dilakukan melalui organogenesis yaitu pembentukan organ atau tunas langsung dari eksplan tanpa melalui pembentukan kalus terlebih dahulu. Eksplan pada penelitian ini adalah daun. Untuk melakukan proses transformasi, daun N. benthamiana terlebih dahulu dilukai dengan dipotong menjadi beberapa bagian. Pelukaan pada tanaman akan menyebabkan sel pada bagian tanaman terluka mengeluarkan senyawa kelompok fenol yang dikenal dengan nama asetosiringon. Asetosiringon berperan menginduksi gen-gen vir yang berfungsi dalam mentransfer T-DNA ke dalam sel tanaman dan penambahan asetosiringon meningkatkan efisiensi infeksi Agrobacterium (Orlikowska et al. 1995), sehingga dapat meningkatkan jumlah sel yang transforman. Untuk meningkatkan efisiensi transformasi, pada penelitian ini asetosiringon juga ditambahkan dalam media kokultivasi. Menurut Ozawa (2009), penggunaan asetosiringon merupakan salah satu kondisi terbaik yang perlu diciptakan agar transformasi lebih efektif.
Transformasi genetik pada penelitian ini menggunakan gen Hd3a di bawah kendali promoter 35S CaMV yang difusikan dengan gen resistensi terhadap higromisin yang merupakan gen penanda seleksi. Gen penanda seleksi sangat penting dalam kegiatan transformasi genetik karena berguna
untuk menyeleksi sel, jaringan, organ atau tanaman yang sudah mengalami transformasi (transgenik). Penelitian ini menggunakan 30 mg/l higromisin untuk seleksi karena pada konsentrasi ini eksplan non transgenik tidak mampu melakukan regenerasi untuk membentuk tunas, dan kemudian eksplan mengalami kematian (Gambar 2 dan 3). Berdasarkan jumlah eksplan yang beregenerasi di media seleksi, rata-rata efisiensi transformasi pada penelitian ini yaitu 86%. Efisiensi transformasi ini adalah tinggi jika dibandingkan dengan transformasi pada Jatropha curcas L. (Sulistyaningsih 2012).
Analisis PCR dilakukan dengan menggunakan primer untuk gen penyeleksi hpt. Posisi gen hpt pada daerah T-DNA dalam pC1300-Hd3a berdampingan dengan LB (batas kiri) dan Hd3a berdampingan dengan RB (batas kanan) (Gambar 1). Keberadaan gen hpt dapat merupakan indikasi keberadaan gen lain dalam satu T-DNA yang sama di dalam genom tanaman transgenik. Hal ini disebabkan oleh proses integrasi daerah T-DNA di dalam genom tanaman yang dimulai dari RB (Sheng&Citovsky 1996). Dengan demikian jika hasil PCR menunjukkan keberadaan gen hpt dalam genom tanaman transgenik maka gen target sisipan Hd3a juga telah terintegrasi dalam genom tanaman tersebut. Hasil PCR terhadap 12 tunas yang diduga transgenik yang dapat hidup di media seleksi yang diambil secara acak menunjukkan bahwa kedua belas tanaman tersebut mengandung gen hpt yang bertanggungjawab terhadap resistensi tanaman terhadap higromisin yang ditunjukkan oleh adanya pita DNA hasil amplifikasi yang berukuran sekitar 570 pb.
Pengamatan fenotipe terhadap tanaman transgenik menunjukkan bahwa N. benthamiana transgenik menghasilkan kuncup bunga pada saat proses regenerasi tunas dan berbunga pada saat tunas di dalam
8
tabung (Gambar 5), sedangkan tunas non transgenik tidak dapat menghasilkan kuncup bunga pada proses tersebut. Hal ini menunjukkan bahwa gen Hd3a dapat memicu pembungaan pada N. benthamiana. Hasil ini memperkuat penelitian sebelumnya pada padi. Padi transgenik yang mengekspresikan gen Hd3a berbunga lebih cepat dari pada padi tipe liarnya pada kondisi hari pendek (Kojima et al. 2002). Ekspresi gen Hd3a di Jatropha curcas juga menghasilkan tunas yang berbunga sangat dini pada saat tunas masih di dalam tabung. Pembungaan dini ini disebabkan oleh ekspresi gen Hd3a di bawah kendali promoter konstitutif p35S CaMV (Sulistyaningsih 2012) .
SIMPULAN
Proses transformasi Nicotiana benthamiana melalui Agrobacterium tumefaciens telah berhasil dilakukan dengan nilai efisiensi transformasi cukup tinggi. Analisis molekuler dengan PCR terhadap tanaman transgenik putatif menunjukkan bahwa tunas yang tumbuh di media seleksi mengandung gen hpt. Karena gen hpt difusikan dengan gen Hd3a, maka tanaman transgenik ini mengandung gen Hd3a. Tunas transgenik yang mengandung gen Hd3a berbunga awal, sehingga gen Hd3a mempunyai peranan dalam memicu pembungaan.
SARAN
Tanaman transgenik perlu diaklimatisasi hingga menghasilkan biji generasi T1 untuk analisis segregasi dan kestabilan transgen, serta ekspresi gen.
DAFTAR PUSTAKA
Chaidamsari T, Samanhudi, Budiani A, Poerwanto R, Santoso D. 2006. Ekspresi fenotipe gen APETALAI kakao (TcAPI) pada eksplan tembakau. Menara Perkebunan 74(1): 1-9.
Hannum S. 2012. Isolasi, pengklonan, dan analisis ekspresi gen penyandi copper-zinc superoxide dismutase (CuZn-SOD) dari Melastoma malabathricum
L. [Disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor. Horsch RB, Fry JE, Hoffman NL, Eichholtz
D, Rogers SG, Fraley RT. 1985. A simple and general method for transferring genes into plants. Science 227 : 1229-1231.
Kojima S, Takahashi Y, Kobayashi Y, Monna L, Sasaki T, Araki T, Yano M. 2002. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant CellPhysiol. 43: 1096-1105.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassay with tobacco tissue cultures. Plant Physiol 15:473-497.
Orlikowska TK, Cranston HJ, Dyer WE. 1995. Factor influencing Agrobacterium tumefaciens–mediated transformation and regeneration of the sunflower cultivar contennial. Plant Cell Tiss. & Org. Cult. 40: 85-91.
Ozawa K. 2009. Establishment of a high efficiency Agrobacterium mediated transformation system of rice (Oryza sativa L.). Plant Science 176: 522-527.
Sheng J, Citovsky V. 1996. Agrobacterium -plant cell DNA transport: have virulence protein, will travel. Plant cell 8: 1699-1710.
Suharsono. 2002. Konstruksi pustaka genom kedelai kultivar Slamet. Hayati 9(3):67-70.
Sulistyaningsih YC. 2012. Rekayasa ekspresi gen pembungaan Hd3a pada tanaman Jarak Pagar (Jatropha curcas L.) [Disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor. Tamaki S, Matsuo S, Wong HL, Yokoi S,
TRANSFORMASI GENETIK Nicotiana benthamiana DENGAN
GEN PEMBUNGAAN Hd3a DARI PADI
LAILA NUR SYAFITRI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
8
Lampiran 1 Komposisi media MS (Murashige & Skoog 1962)
Bahan Konsentrasi
(mg/l)
Hara makro NH4NO3 1650
CaCl2.2H2O 440
MgSO4.7H2O 370
KNO3 1900
KH2PO4 170
Hara mikro H3BO3 6,2
Na2MoO4.2H2O 0,25
CoCl2.6H2O 0,025
KI 0,83
ZnSO4.7H2O 8,6
MnSO4.4H2O 22,3
CuSO4.5H2O 0,025
Na2EDTA 37,2
FeSO4.7H20 27,8
Vitamin Myo-Inositol 100
Pyridoxin-HCl 0,5
Nicotinic acid 0,5
Thiamin-HCl 0,1
Glycine 2
Sukrosa 30 g/l
9
Lampiran 2 Perhitungan efisiensi transformasi dan jumlah tunas transgenik putatif tiap eksplan
% efisiensi transformasi = jumlah eksplan menghasilkan tunas transgenik putatif
jumlah eksplan total yang ditransformasi × 100%
jumlah tunas transgenik putatif tiap eksplan = jumlah total tunas transgenik putatif