ANALISIS PERTUMBUHAN KEDELAI (
Glycine max
(L.)
Merr.) DI BAWAH CEKAMAN NAUNGAN
WIDYA MERITA NINGRUM A24070108
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
Widya Merita N.1, Abdul Qadir2
1
Mahasiswa Departemen Agronomi dan Hortikultura, Fakultas Pertanian, IPB, A24070108
2Staf Pengajar Departemen Agronomi dan Hortikultura, Fakultas Pertanian, IPB
Abstract
The purpose of this research was to study the growth analyze of two varieties soybean under shade condition. This research was conducted in controlled condition of green house at IPB garden experiment, Leuwikopo, Dramaga, Bogor on March until June 2011. The experiment was arranged in Split Plot Design by two factors and three replications, with the levels of
second factors nested under the levels of first factors. The first factor is shade consist of N0
=non-shade (under full sunlight) and N50=50% shade. The second factors is two varieties consist of
G1=Godek (shade sensitive) and G2=Ceneng (shade tolerant). The 50% shading was made
artificially using black plastic paranet with 50% light transmitted. Agronomic characters measured were: plant height, total leaves, and time of flowering. Physiological characters measured were chlorophylls content (chl a, chl b, and ratio chl a/b). Growth analyze measured were Leaf Area Index (LAI), Net Assimilation Rate (NAR), Leaf Area Ratio (LAR), and Relative Growth Rate (RGR). The result of research indicated that Ceneng has a higher plant height, low of total leaves, faster time of flowering, higher of chl a, chl b, lower ratio of chl a/b, higer LAI, LAR, and lower NAR, RGR.
RINGKASAN
WIDYA MERITA NINGRUM. Analisis Pertumbuhan Kedelai (Glycine max
(L.) Merr.) di Bawah Cekaman Naungan. (Dibimbing oleh ABDUL QADIR).
Penelitian ini bertujuan untuk mengetahui pengaruh naungan (50%) terhadap pertumbuhan dua varietas kedelai (Glycine max (L.) Merr.) melalui peubah-peubah analisis pertumbuhan tanaman. Penelitian dilaksanakan mulai
bulan Maret sampai Juni 2011 di Kebun Percobaan Ilmu dan Teknologi Benih, Leuwikopo, Dramaga, Bogor. Analisis tanah dilakukan di Laboratorium Analisis Tanah Balai Penelitian Tanaman Obat dan Aromatik, Cimanggu, Bogor dan analisis klorofil dilakukan di Laboratorium Plant Analysis and Chromatography, Departemen Agronomi dan Hortikultura, FAPERTA, IPB.
Penelitian ini menggunakan rancangan petak terbagi menggunakan tiga ulangan dengan anak petak tersarang pada petak utama dengan tiga ulangan. Penelitian ini terdiri dari dua faktor, yang pertama adalah petak utama berupa intensitas cahaya yang diatur menggunakan naungan paranet, yang terdiri dari dua tingkat naungan (N0=0% (tanpa naungan) dan N50=50% (dengan naungan paranet 50%)) dan faktor kedua adalah dua varietas kedelai (G1=genotipe peka naungan (Godek) dan G2=genotipe toleran naungan (Ceneng)) sebagai anak petak, sehingga terdapat 12 satuan percobaan. Untuk mengetahui pengaruh nyata akibat intensitas cahaya rendah dan genotipe kedelai data dianalisa dengan menggunakan uji F. Uji lanjut dilakukan bila pengaruh perlakuan berpengaruh nyata terhadap peubah yang diukur dengan uji DMRT pada taraf kesalahan 5%.
Peubah yang diamati pada penelitian ini meliputi; karakter agronomi (tinggi tanaman, jumlah daun trifoliat dan umur berbunga), pengamatan analisis
pertumbuhan (Indeks Luas Daun (Leaf Area Index), Laju Asimilasi Neto (Net
Assimilation Rate), Nisbah Luas Daun (Leaf Area Ratio), Laju Pertumbuhan
Data iklim mikro di lokasi penelitian meliputi suhu rata-rata selama penelitan sekitar 33.90C, kelembaban relatif sekitar 65%. Daya kecambah benih dari kedua genotipe kurang dari 80% sehingga penyulaman dilakukan pada saat tanaman berumur 2 MST untuk mempertahankan populasi.
Hama yang dijumpai diantaranya adalah belalang, dan kutu putih. Tidak ada penyakit yang dijumpai pada saat penelitian. Pengendalian hama belalang menggunakan insektisida kontak Decis 25EC dengan dosis 0.5 ml/l. Gulma yang dijumpai saat pelaksanaan penelitian diantaranya adalah Mimosa pudica, Boreria
Laevis, Phyllantus niruri, Oxlalis barerieli. Pengendalian gulma dilakukan secara
manual seminggu sekali.
Hasil penelitian menunjukkan bahwa naungan sebesar 50% mempengaruhi pertumbuhan tanaman kedelai untuk kedua genotipe. Naungan 50% meningkatkan
tinggi tanaman, menurunkan jumlah daun trifoliat, mempercepat waktu pembungaan, meningkatkan jumlah klorofil a dan b, menurunkan rasio klorofil a/b, meningkatkan Indeks Luas Daun, menurunkan Laju Asimilasi Bersih, meningkatkan Nisbah Luas Daun, menurunkan Laju Pertumbuhan Relatif, meningkatkan Luas Daun Spesifik, menurunkan berat kering total tanaman dan berat polong.
ANALISIS PERTUMBUHAN KEDELAI (
Glycine max
(L.)
Merr.) DI BAWAH CEKAMAN NAUNGAN
Skripsi sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Fakultas Pertanian Institut Pertanian Bogor
WIDYA MERITA NINGRUM A24070108
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
LEMBAR PENGESAHAN
Judul Skripsi :
ANALISIS PERTUMBUHAN KEDELAI (
Glycine
max
(L.) Merr.) DI BAWAH CEKAMAN
NA-UNGAN
Nama :
WIDYA MERITA NINGRUM
NIM :
A24070108
Menyetujui, Pembimbing
Ir. Abdul Qadir, MS. NIP. 19620927 198703 1 001
Mengetahui, Ketua Departemen
Dr. Ir. Agus Purwito, MSc. Agr NIP. 19611101 198703 1 003
RIWAYAT HIDUP
Penulis dilahirkan di Kediri, pada tanggal 24 Maret 1989. Penulis merupakan anak pertama dari tiga bersaudara dari pasangan Sugian Budi Martono dan Tri Agustin.
Penulis memulai pendidikan pada tahun 1995 di SD Negeri Pare 1, Kediri. Pada tahun 2001 penulis melanjutkan pendidikan ke SMP Negeri 2 Pare dan lulus pada tahun 2004. Penulis melanjutkan pendidikan ke SMA Negeri 2 Pare dan lulus pada tahun 2007.
KATA PENGANTAR
Puji Syukur penulis panjatkan kehadirat Allah SWT yang telah memberikan kekuatan dan hidayah-Nya, sehingga penulis dapat menyelesaikan penelitian dan menuliskannya dalam skripsi yang berjudul Analisis Perumbuhan
Kedelai (Glycine max (L.) Merr) Di Bawah Cekaman Naungan. Selama penelitian dan penyelesaian skripsi ini, penulis telah banyak memperoleh bantuan dan dukungan dari berbagai pihak. Untuk itu penulis mengucapkan terimakasih kepada:
1. Ir. Abdul Qadir, MS. selaku dosen pembimbing skripsi yang telah
membimbing penulis selama menjalankan penelitian dan penyusunan skripsi. 2. Dr. Ir. Suwarto, M.Si dan Ir. Diny Dinarti, M.Si selaku dosen penguji yang
telah memberi masukan dan kritikan untuk perbaikan skripsi ini.
3. Dr. Ir. Muh. Syukur, M.Si selaku pembimbing akademik penulis yang telah membimbing penulis selama menjalani perkuliahan di Institut Pertanian Bogor.
4. Bapak Sugian Budi Martono, Ibu Tri Agustin, adik-adikku, yaitu Septyan dan
Fibri, serta seluruh keluarga di Kediri atas kasih sayang, doa dan dorongannya. 5. Ir. Bregas Budianto, Ass.Dpl, Anto, Sriyo, yang telah membantu instrumen
alat.
6. Pak Nana, Bu Eli, Bibi, Pak Bambang, Pak Haryanto, dan Kak Arif atas bantuan yang luar biasa selama penelitian berlangsung.
7. Elly Zunara atas doa, bantuan, dan perhatiannya.
8. Rikania, Nyoman Ari, Fani, S. Khalimah, Tatied, Nurul dan Beni yang selalu
memberi perhatian dan bantuan.
9. Semua pihak yang telah membantu dalam penyelesaian studi dan skripsi ini,
yang tidak dapat penulis sebutkan satu-persatu.
Akhirnya, penulis berharap semoga hasil penelitian ini berguna bagi yang memerlukan.
Bogor, November 2011
DAFTAR ISI
Halaman
DAFTAR ISI ... i
DAFTAR TABEL ... iii
DAFTAR GAMBAR ... vi
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 3
Hipotesis ... 3
TINJAUAN PUSTAKA ... 4
Botani Kedelai ... 4
Lingkungan Tumbuh ... 5
Sistem Produksi ... 6
Faktor yang Mempengaruhi Fotosintesis ... 7
Adaptasi Terhadap Cahaya dan Naungan ... 8
Analisis Pertumbuhan tanaman ... 9
METODOLOGI ... 13
Tempat dan Waktu ... 13
Bahan dan Alat ... 13
Metode Penelitian ... 13
Pelaksanaan Penelitian ... 14
Pengamatan ... 16
HASIL DAN PEMBAHASAN ... 19
Kondisi Umum ... 19
Karakter Agronomi ... 21
Tinggi Tanaman ... 21
Jumlah Daun Trifoliat ... 23
Waktu Berbunga ... 25
Karakter Fisiologi ... 26
Klorofil a ... 26
Klorofil b... 28
Rasio Klorofil a/b... 29
Analisis Pertumbuhan Tanaman ... 30
Indeks Luas Daun ... 30
Laju Asimilasi Bersih ... 32
Nisbah Luas Daun ... 34
Laju Pertumbuhan Relatif ... 36
KESIMPULAN DAN SARAN ... 46
DAFTAR PUSTAKA ... 47
DAFTAR TABEL
Nomor Halaman
1. Rekapitulasi Hasil Sidik Ragam Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Karakter Agronomi, Analisis Pertumbuhan
Tanaman dan Fisiologi ... 20
2. Pengaruh Naungan Terhadap Tinggi Tanaman Dua Varietas Kedelai 21
3. Pengaruh Naungan Terhadap Jumlah daun Trifoliat Dua Varietas
Kedelai ... 23
4. Pengaruh Naungan terhadap Peubah Waktu Berbunga Dua Varietas
Kedelai ... 25
5. Pengaruh Naungan terhadap Jumlah Klorofil a pada Dua Genotipe
Kedelai ... 26
6. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Jumlah
Klorofil a Tanaman Kedelai pada Fase Berbunga Penuh ... 27
7. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Jumlah
Klorofil a Tanaman Kedelai pada Fase Mulai Berpolong ... 27
8. Pengaruh Naungan terhadap Jumlah Klorofil b pada Dua Genotipe
Kedelai ... 28
9. Pengaruh naungan, Genotipe, dan Interaksi terhadap Jumlah
Klorofil b Tanaman Kedelai Fase Berpolong Penuh ... 29
10. Pengaruh Naungan terhadap Rasio Klorofi a/b pada Dua Genotipe
Kedelai ... 30
11. Pengaruh Naungan terhadap Indeks Luas Daun pada Dua Genotipe
Kedelai ... 31
12. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Indeks Luas
Daun pada Fase Berbunga Penuh ... 32
13. Pengaruh Naungan terhadap Laju asimilais Bersih pada Dua
Genotipe Kedelai ... 33
14. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Laju
15. Pengaruh Naungan terhadap Nisbah Luas Daun pada Dua Genotipe
Kedelai ... 35
16. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Nisbah
Luas Daun Tanaman Kedelai pada Fase Berbunga Penuh ... 35
17. Pengaruh Naungan terhadap Laju Pertumbuhan Relatif pada Dua
Genotipe Kedelai ... 36
18. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Nisbah
Luas Daun Tanaman Kedelai pada Fase Berbunga Penuh ... 36
Lampiran
1. Analisis Ragam Peubah Tinggi Tanaman pada Perlakuan Naungan
dan Genotipe Kedelai ... 54
2. Analisis Ragam Peubah Jumlah Daun Trifoliate pada Perlakuan
Naungan dan Genotipe Kedelai ... 54
3. Analisis Ragam Peubah Waktu Berbunga pada Perlakuan Naungan
dan Genotipe Kedelai ... 55
4. Analisis Ragam Peubah Kolofil a pada Perlakuan Naungan dan
Genotipe Kedelai ... 56
5. Analisis Ragam Peubah Klorofil b pada Perlakuan Naungan dan
Genotipe Kedelai ... 57
6. Analisis Ragam Peubah Rasio Klorofil a/b pada Perlakuan Naungan
dan Genotipe Kedelai ... 58
7. Analisis Ragam Peubah Indeks Luas Daun pada Perlakuan Naungan
dan Genotipe Kedelai ... 59
8. Analisis Ragam Peubah Laju Asimilasi Bersih pada Perlakuan
Naungan dan Genotipe ... 60
9. Analisis Ragam Peubah Nisbah Luas Daun pada Perlakuan Naungan
dan Genotipe Kedelai ... 61
10. Analisis Ragam Laju Pertumbuhan Relatif pada Perlakuan Naungan
dan Genotipe Kedelai ... 62
DAFTAR GAMBAR
Nomor Halaman
Teks
1. Luas Daun Spesifik (LDS) Dua Genotipe Kedelai pada Kondisi Tanpa
Naungan (0%) dan Naungan (50%) Fase Berpolong Penuh ... 39
2. Persentase Pembagian Biomassa Dua Genotipe Kedelai pada Kondisi
Tanpa Naungan (0%) dan Naungan (50%) Fase Berpolong Penuh ... 44
Lampiran
1. Denah Percobaan dengan Dua Naungan (Naungan 50% (N50) dan Tanpa Naungan (N0)) dan Dua Genotipe Kedelai (Genotipe Peka
(G1) dan Genotipe Toleran (G2)) ... 52
2. Alat yang digunakan dalam penelitian; Spektrofotometer (1), Tabung
Reaksi (2), Timbangan Digital (3), Solarimeter (4) ... 52
3. Benih kedua Genotipe: Godek (G1) dan Ceneng (G2) ... 53
4. Pertumbuhan Tinggi Tanaman Kedelai Genotipe: Godek (G1) dan Ceneng (G2) pada Fase Mulai Berbunga dengan Naungan 50% (N50)
PENDAHULUAN
Latar Belakang
Kedelai merupakan tanaman pangan berupa semak yang tumbuh tegak. Kedelai jenis liar Glycine ururiencis, merupakan kedelai yang menurunkan
berbagai kedelai yang kita kenal sekarang (Glycine max (L) Merril). Kedelai
berasal dari daerah Manshukuo (Cina Utara). Di Indonesia, kedelai mulai dibudidayakan sejak abad ke-17 sebagai tanaman makanan dan pupuk hijau. Penyebaran tanaman kedelai ke Indonesia berasal dari daerah Manshukuo
kemudian menyebar ke daerah Mansyuria, Jepang (Asia Timur) dan ke negara-negara lain di Amerika dan Afrika (Prihatman, 2000).
Kedelai mengandung protein, isoflavon, dan serat untuk kesehatan. Kedelai adalah sumber protein diet, termasuk semua asam amino esensial. Kedelai juga merupakan sumber lesitin atau fosfolipid. Isoflavon dan lesitin kedelai telah dipelajari secara ilmiah untuk kesehatan. Seperti isoflavon genistein yang diyakini memiliki efek estrogen di tubuh, sebagai hasilnya kadang-kadang disebut fitoestrogen (Mayo Foundation, 2010).
Kedelai termasuk kedalam famili Leguminosae yang merupakan sumber pangan dan pakan, hal ini terbukti dengan kedudukan famili ini di urutan kedua setelah Graminae (Baharsjah, 1992). Komoditi ini hingga kini produksinya terus menurun. Produksi kedelai di Indonesia pernah mencapai puncaknya pada tahun 1992 yaitu sebanyak 1.87 juta ton. Produksi terus mengalami penurunan hingga hanya 0.672 juta ton pada tahun 2003. Data tersebut menunjukkan selama 11 tahun produksi kedelai merosot mencapai 64 persen (Atman, 2009).
Sehubungan dengan permasalahan diatas setidaknya ada lima strategi penting yang harus dilaksanakan untuk menjamin keberhasilan peningkatan produksi kedelai nasional, yaitu: (1) Perbaikan harga jual; (2) pemanfaatan potensi lahan; (3) intensifikasi pertanaman; (4) perbaikan proses produksi; dan (5) konsistensi program dan kesungguhan aparat. Harga jual yang rendah di tingkat petani sehingga kurang kompetitif dibandingkan komoditas palawija lainnya, merupakan salah satu faktor utama yang menyebabkan petani kurang berminat membudidayakan kedelai. Peningkatan harga jual di tingkat petani merupakan kunci utama dalam mengembalikan minat petani untuk menanam kedelai. Pemanfaatan potensi lahan yang tersedia untuk mendukung peningkatan produksi kedelai antara lain dapat dilakukan dengan penanaman kedelai sebagai tanaman sela, diantaranya penanaman kedelai secara tumpang sari dengan ubi kayu, kelapa
sawit, kelapa, atau tanaman tua lainnya. Perbaikan proses produksi, konsistensi program dan kesungguhan aparat berkaitan dengan kerjasama antara petani dan aparat yang berkaitan untuk menggunakan sistem produksi yang sesuai dan pelaksanaan program yang kontinyu (Subandi dalam Atman, 2009).
Ditinjau dari ketersediaan teknologi dan sumber daya lahan maka pengembangan kedelai di dalam negeri masih dapat diperluas guna meningkatkan produksi menuju swasembada kedelai (Hermanto, 2010). Guna menanggulangi penurunan lahan dalam rangka produksi kedelai diperlukan usaha perluasan lahan melalui optimasi pemanfaatan lahan baik tanaman perkebunan maupun tanaman pohon lainnya. Optimasi pemanfaatan lahan misalnya dapat berupa pemanfaatan lahan gawangan tanaman perkebunan dengan penanaman kedelai sebagai tanaman sela (intercropping plant). Pemanfaatan lahan gawangan untuk produksi kedelai
mengalami beberapa kendala, diantaranya karena penutupan tajuk tanaman perkebunan yang menyebabkan lahan gawangan sebagian besar ternaungi dari cahaya matahari (Atman, 2009).
Kemampuan adaptasi dari tanaman yang toleran intensitas cahaya rendah dengan tanaman yang peka erat kaitannya dengan karakter-karakter fisiologi fotosintetik tanaman tersebut (Soverda et al., 2009).
Analisis pertumbuhan merupakan suatu cara untuk mengikuti dinamika fotosintesis yang diukur oleh produksi bahan kering. Pertumbuhan tanaman dapat diukur tanpa mengganggu tanaman, yaitu dengan pengukuran tinggi tanaman atau jumlah daun, tetapi sering kurang mencerminkan ketelitian kuantitatif. Akumulasi bahan kering sangat disukai sebagai ukuran pertumbuhan. Akumulasi bahan kering mencerminkan kemampuan tanaman dalam mengikat energi cahaya matahari melalui proses fotosintesis, serta interaksinya dengan faktor-faktor lingkungan lainnya (Sumarsono, 2008).
Komponen analisis pertumbuhan diantaranya adalah laju pertumbuhan
relatif (Relatif Growth Rate), laju unit daun (Unit Leaf Rate), nisbah luas daun (Leaf Area Ratio), luas daun spesifik dan rasio berat daun (Specific Leaf Area and
Leaf Weight Ratio), indeks luas daun (Leaf Area Index), laju tumbuh pertanaman
(Crop Growth Rate), lamanya luas daun dan lamanya biomassa (Leaf area
Duration and Biomass Duration) (Beadle, 1993).
Potensi hasil yang akan diperoleh dapat diketahui dengan mempelajari peubah-peubah pertumbuhan pada tanaman kedelai terhadap cekaman naungan atau intensitas cahaya rendah dengan genotipe dan tingkat naungan tertentu.
Tujuan
Tujuan dari penelitian ini adalah untuk mengetahui pengaruh naungan terhadap pertumbuhan dua genotipe kedelai melalui peubah-peubah analisis pertumbuhan tanaman.
Hipotesis
Hipotesis yang diajukan dalam penelitian ini, yaitu:
1. Naungan berpengaruh terhadap pertumbuhan tanaman.
TINJAUAN PUSTAKA
Botani Kedelai
Tanaman kedelai (Glycine max (L.) Merr.) merupakan anggota dari
famili Leguminosae, subfamili Papilionideae, dan termasuk ke dalam genus
Glycine L. (Johnson and Bernard, 1963). Bibit kedelai berkecambah dengan tipe
perkecambahan epigeal dengan kotiledon tebal dan berdaging, berwarna kuning atau hijau. Tanaman ini biasanya tegak dan merupakan herba tahunan yang lebat dengan tinggi mencapai dua meter dan kadang-kadang agak merambat. Sistem
perakaran tunggang bercabang dengan panjang akar mencapai dua meter. Akar lateral menyebar secara horizontal hingga 2.5 meter (Giller dan Dashiell, 2010).
Tanaman kedelai mempunyai bunga sempurna, yaitu dalam satu bunga terdapat alat kelamin jantan (benang sari/serbuk sari) dan alat kelamin betina (putik). Bunga kedelai berwarna ungu atau putih (Fachruddin, 2000). Bunga kedelai biasanya berukuran panjang sekitar enam sampai tujuh milimeter dan secara keseluruhan ukurannya kecil. Struktur bunga kedelai yang sedemikian rupa menjadikan bunga tersebut melakukan suatu pembatasan terhadap penyerbukan, yakni penyerbukan yang mereka kontrol sendiri, yaitu penyerbukan sendiri (
self-pollination). Penyebrukan sendiri, yaitu kepala putik diserbuki oleh tepung sari
dari bunga yang sama (Kartono, 2005).
Kedelai merupakan tanaman hari pendek, yakni tidak akan berbunga bila lama penyinaran (panjang hari) melampaui batas kritis. Apabila lama penyinaran kurang dari batas kritik, maka kedelai akan berbunga. Pada lama penyinaran 12 jam hampir semua varietas kedelai dapat berbunga beragam dari 20-60 hari setelah tanam. Apabila lama penyinaran melebihi periode kritik, tanaman kedelai akan meneruskan pertumbuhan vegetatifnya tanpa pembungaan (Karamoy, 2009). Di Indonesia, tanaman kedelai pada umumnya mulai berbunga pada umur 30-50 hari setelah tanam (Fachruddin, 2000). Faktor lain seperti suhu, nutrisi, intensitas cahaya mungkin mempengaruhi respon kedelai yang sesuai untuk pembungaan
mencapai 75 persen. Kedelai biasanya menghasilkan banyak bunga daripada buah yang terbentuk. (Johnson and Bernard, 1963).
Buah kedelai berbentuk polong dan setiap polong berisi satu sampai empat biji. Biji umunya berbentuk bulat atau bulat pipih sampai bulat lonjong. Ukuran biji berkisar antara 6-30 gram/100 biji. Ukuran biji diklasifikasikan menjadi tiga kelas, yaitu biji kecil (6-10 gram/100 biji), sedang (11-12 gram/100 biji), dan besar (13 atau lebih/100 biji). Warna kulit biji bervariasi antara lain, kuning, hijau, cokelat, dan hitam (Fachruddin, 2000).
Lingkungan Tumbuh
Kedelai sebagai tanaman legum memiliki areal kesesuaian lingkungan dalam hal lintang, ketinggian tempat, suhu, panjang hari, dan kelembaban.
Panjang hari dan intensitas penyinaran (radiasi) surya penting untuk diperhatikan dalam budidaya tanaman kedelai. Faktor iklim terutama radiasi surya perlu diperhatikan (Baharsjah, 1992).
Di Indonesia kedelai dapat tumbuh dan berproduksi dengan baik di dataran rendah samapi ketinggian 900 meter diatas permukaan laut (dpl). Meskipun demikian telah banyak varietas atau genotipe kedelai dalam negeri maupun introduksi yang dapat beradaptasi dengan baik di dataran tinggi (pegunungan) ± 1 200 m dpl. (Rukmana dan Yuniarsih, 1996). Kedelai biasanya ditanam di daerah dengan garis lintang 550 LU atau 550 LS, pada ketinggian dari permukaan laut sampai dengan 2 000 mdpl (Giller dan Dashiell, 2010).
Kisaran suhu untuk pertumbuhan kedelai adalah 10-350C, diatas suhu 350C tanaman dapat tumbuh namun kurang baik, dan diatas suhu 400C produksinya hampir tidak ada. Suhu yang kurang sesuai terhadap tanaman kedelai dapat mengakibatkan berkurangnya inisiasi bunga dan pembentukan polong (Baharsjah, 1992). Kondisi iklim yang cocok untuk penanaman kedelai di Indonesia umumnya adalah daerah dengan suhu antara 25-270C (Rukmana dan Yuniarsih, 1996). Meskipun tanaman tumbuh dengan baik pada temperatur
10-350C, namun suhu optimum untuk pertumbuhan dan perkembangan pada
Pada umumnya kedelai menghendaki tanah yang berstruktur remah dengan keasaman sedang (pH 5-7) (Baharsjah, 1992). Nilai pH ideal bagi pertumbuhan kedelai dan bakteri Rhizobium adalah 6.0-6.8. Apabila pH diatas 7.0
tanaman kedelai mengalami klorosis sehingga tanaman menjadi kerdil dan daunnya menguning. Sementara pada pH di bawah 5.0 kedelai mengalami keracunan Al, Fe, dan Mn, sehingga pertumbuhannya terganggu. Untuk
menaikkan pH, dilakukan pengapuran misalnya dengan Kalsit (CaCO3), Dolomit
(CaMg(CO3)2), atau kapur bakar. Pemberian kapur dilakukan sekitar 2-4 minggu sebelum tanam, bersamaan dengan pengolahan lahan. Tanaman kedelai dapat ditanam pada berbagai jenis tanah dengan drainase dan aerasi yang baik. Jenis tanah yang sangat cocok untuk kedelai adalah Aluvial, Regosol, Grumosol, Latosol, dan Andosol (Fachruddin, 2000).
Kedelai tergolong pada tanaman yang tidak tahan kekeringan dan kelebihan air. Kekeringan akan menurunkan hasil, sedangkan pengairan berlebihan dalam ketersediaan air terbatas disamping menurunkan hasil juga mengurangi luas pertanaman. Teknik irigasi dan teknik konservasi air khusus dikembangkan dalam usaha memanipulasi status air tanah agar sesuai dengan kebutuhan air kedelai (Sumarno dan Harnoto, 1993). Kondisi iklim yang cocok untuk penanaman kedelai di Indonesia umumnya adalah daerah dengan kelembaban udara (RH) rata-rata 65% dan curah hujan paling optimum antara 100-200 mm/bulan (Rukmana dan Yuniarsih, 1996). Kedelai membutuhkan setidaknya 500 mm air selama musim pertumbuhan untuk perkembangan yang baik dengan konsumsi air dalam kondisi optimal adalah 850 mm (Giller dan Dashiell, 2010).
Sistem Produksi
Di sebagian besar negara, kedelai ditanam dalam baris. Seperti kebanyakan tanaman tahunan lainnya, hampir segala sesuatu tentang budidaya kedelai mekanik, dari penyusunan tanah untuk penanaman, budidaya, aplikasi bahan kimia, dan panen. Praktek budidaya seperti persiapan lahan konservasi atau penanaman langsung semua merujuk kepada cara untuk mengurangi atau menghilangkan tahap pengolahan tanah. Budidaya tersebut menguntungkan, yaitu sistem budidaya lebih organik, mengurangi jumlah mesin yang digunakan, dan mengurangi biaya keseluruhan produksi. Pemanenan terjadi sekali per tahun. Kedelai dipanen dan dipotong dari tangkai dan dipisahkan biji dari polong dan sisa tanaman (Aquino, 2002).
Umur panen kedelai ditentukan oleh beberapa faktor, yaitu varietas dan ketinggian tempat penanaman. Di daerah dataran tinggi, umur tanaman kedelai
siap panen lebih lama 10-20 hari dibandingkan dengan di daerah dataran rendah. Ciri–ciri umum tanaman kedelai siap panen diantaranya adalah: polong berwarna kuning kecoklatan secara merata, daun sudah banyak yang kering dan rontok, batang sudah mengering (Fachruddin, 2000).
Faktor yang Mempengaruhi Fotosintesis
Pertanian pada dasarnya merupakan sistem pemanfaatan energi matahari melalui proses fotosintesis. Produksi tanaman budidaya pada dasarnya tergantung pada ukuran dan efisiensi sistem fotosintesis. Tempat utama terjadinya fotosintesis pada legum pangan adalah pada daun. Tidak seperti pada tanaman serealia dimana kegiatan fotosintesis pada malai dapat memberikan andil sampai 50 persen atau lebih dari fotosintat yang dibutuhkan oleh biji-biji yang sedang
mengisi, polong-polong hijau dari legum tidak menunjukkan adanya fiksasi CO2
dari udara (Baharsjah, 1992).
proses yang terjadi pada fotosintesis. Pada tanaman alfalfa (Medicago sativa),
yang diamati selama dua hari di akhir musim panas dengan pengaruh awan menutupi beberapa waktu, menunjukkan bahwa penambatan CO2 paling banyak terjadi sekitar tengah hari ketika tingkat cahaya paling tinggi dan cahaya sering membatasi fotosintesis terlihat dengan menurunnya laju penambatan CO2 ketika tumbuhan terkena bayangan awan sebentar (Salisbury dan Ross, 1991).
Kisaran laju fotosintesis telah diteliti oleh Ogren dan Rinne pada Glycine
max, yaitu sebesar 12-24 CO2dm-2h-1. Laju fotosintesis berubah dengan bertambahnya umur tanaman. Faktor-faktor yang mempengaruhi laju fotosintesis yaitu, intensitas serta lamanya penyinaran, difusi CO2, karboksilasi, translokasi, dan banyaknya klorofil per satuan luas daun (Baharsjah, 1992).
Adaptasi Terhadap Naungan
Pada tumbuhan dikotil, daun yang ternaungi berukuran lebih besar tapi lebih tipis dibandingkan dengan daun dari tanaman yang biasa hidup dibawah jumlah cahaya matahari yang cukup. Daun matahari (daun dari tanaman yang tidak ternaungi) menjadi lebih tebal daripada daun naungan karena membentuk sel palisade yang lebih panjang atau membentuk tambahan lapisan sel palisade. Berdasarkan bobot, daun naungan umumnya juga mempunyai klorofil yang lebih banyak, khususnya klorofil b, terutama karena tiap kloroplas mempunyai lebih banyak grana dibandingkan dengan pada daun matahari. Kloroplas daun yang ternaungai mempunyai protein stroma lebih sedikit, termasuk rubisko, dan mungkin juga protein pengangkut elektron tilakoid lebih sedikit daripada daun matahari. Daun naungan menggunakan lebih banyak energi untuk menghasilkan pigmen pemanen cahaya yang memungkinkannya mampu menggunakan semua cahaya dalam jumlah terbatas yang mengenainya (Salisbury dan Ross, 1991).
Salisbury dan Ross (1991) selanjutnya juga menjelaskan bahwa Alocasia
sebagai tanaman ternaungi mempunyai repon yang khas. Pada tanaman Alocasia,
tingkat cahaya yang sangat rendah mereka biasanya berfotosintesis pada laju yang lebih tinggi dibandingkan spesies yang lainnya dan titik kompensasi cahayanya sangat rendah.
Adaptasi tanaman, terutama adaptasi tanaman kedelai terhadap naungan telah banyak diteliti. Naungan berpengaruh terhadap karakter morfologi, anatomi, fisiologi, dan produksi kedelai dengan meningkatkan tinggi tanaman (Mulyana 2006; Anggraeni 2010), luas daun trifoliat (Mulyana 2006; Kisman 2007), kandungan klorofil a dan b (Jufri dan Mulyana 2006; Muhuria dan Kisman 2007; Anggraeni 2010), menurunkan jumlah daun trifoliat (Mulyana 2006; Anggraeni 2010), tebal daun (Jufri 2006; Anggraeni 2010), kerapatan stomata (Mulyana 2006; Anggraeni 2010), polong isi, polong hampa, polong total, bobot biji per tanaman, dan bobot 100 butir (Jufri dan Mulyana, 2006; Anggraeni 2010).
Analisis Pertumbuhan tanaman Indeks Luas Daun (ILD) atau Leaf Area Index (LAI)
Pengukuran luas daun tanaman merupakan salah satu pengamatan yang cukup penting dalam penelitian fisiologi dan agronomi. Hasil pengukuran tersebut dapat digunakan untuk menentukan indeks luas daun, laju asimilasi netto, efisiensi fotosintesis, dan potensi fotosintesis daun. Cara pengukuran luas daun ini dapat dilakukan dengan atau tanpa memotong daun dari tanaman. Pada penelitian-penelitian tertentu, misalnya transpirasi dan fotosintesis, sering diperlukan pengukuran luas daun tanpa pemotongan daun dari tanamannya (Sutoro, 1986).
LAI (Leaf Area Index) merupakan nisbah luas daun (satu sisi permukaan
saja) tanaman terhadap luas permukaan tanah tempat tanaman tersebut tumbuh (Salisbury dan Ross, 1991). Indeks Luas Daun (ILD) yang dibatasi dengan pengertian perbandingan luas daun total dengan luas tanah yang ditutupi atau luas daun diatas suatu luasan tanah, yang diusulkan untuk disebut Leaf Area Index
(LAI). Luas daun dan luas lahan memiliki satuan yang sama, sehingga peubah analisis ini tidak memiliki satuan atau suatu nisbah yang tidak memiliki dimensi (Watson dalam Sitompul dan Guritno, 1995).
LAI (Leaf Area Index) merupakan total bahan daun dalam suatu
fotosintesis jaringan per unit satuan luas pada permukaan atas tanah. LAI dari tajuk tanaman memainkan peranan penting dalam mengontrol interaksi antara lingkungan darat dan variabel atmosfer. Akurasi dan presisi Indeks Luas Daun yang berasal dari pengindraan jauh atau citra jarak jauh sangat penting untuk studi perubahan iklim, terutama pada skala regional dan lokal untuk meningkatkan proses parameterisasi dalam berbagai kelas model (Gobron, 2008).
Laju Asimilasi Bersih (LAB) atau Net Assimilation Rate (NAR)
Tingkat Asimilasi Bersih (LAB) berdasarkan luas daun, berat kering, protein dan kandungan klorofil telah diukur untuk banyak spesies. Pentingnya luas daun dalam menentukan hasil telah banyak dibahas, tetapi penelitian terbaru sampai pada keterbatasan pengukuran luas daun total (Ohlrogge, 1963).
Laju asimilasi bersih adalah laju penimbunan berat kering per satuan luas daun per satuan waktu. LAB merupakan ukuran rata-rata efisiensi fotosintesis daun dalam suatu komunitas tanaman budidaya. LAB paling tinggi nilainya pada saat tumbuhan masih kecil dan sebagian besar daunnya terkena sinar matahari langsung. Dengan bertumbuhnya tanaman budidaya dan dengan meningkatnya LAI, makin banyak daun terlindung, menyebabkan penurunan LAB sepanjang musim pertumbuhan. Laju asimilasi bersih merupakan ukuran rata-rata efisiensi fotosintesis daun dalam suatu komunitas tanaman budidaya (Gardner et al., 1991).
LAB atau NAR dalam banyak literatur sering disebut juga dengan HSD (Harga Satuan Daun). Hal tersebut kurang tepat untuk menyatakan pertambahan bahan baru tanaman. Hal ini didasarkan atas alasan bahwa bahan baru tanaman yang dibentuk tidak hanya berasal dari produk reduksi CO2 dalam proses fotosintesis, tetapi juga unsur hara yang diambil akar dari dalam tanah. Sementara NAR mempunyai pengertian tingkat asimilasi CO2 bersih, yaitu jumlah total CO2 yang diambil tanaman dikurangi dengan jumlah yang hilang melalui respirasi (Sitompul dan Guritno, 1995). Laju asimilasi bersih dapat menggambarkan produksi bahan kering atau merupakan produksi bahan kering per satuan luas daun dengan asumsi bahan kering tersusun sebagian besar dari CO2 (Kastono et
Nisbah Luas Daun (NLD) atau Leaf Area Ratio (LAR)
Suatu peubah pertumbuhan yang dapat digunakan untuk mencerminkan morfologi tanaman adalah nisbah luas daun (NLD), yaitu hasil bagi dari luas daun dengan berat kering total tanaman. Indeks ini mencangkup proses pembagian dan translokasi asimilat ke tempat sintesa bahan daun dan efisiensi penggunaan substrat dalam pembentukan luasan daun (Sitompul dan Guritno, 1995).
Laju Pertumbuhan Relatif (LPR) atau Relative Growth Rate (RGR)
Merupakan pertambahan berat kering tanaman pada suatu waktu tertentu (Beadle, 1993). Laju Pertumbuhan Relatif (LPR) merupakan peningkatan berat kering tanaman dalam suatu interval waktu, erat hubungannya dengan berat awal tanaman. Asumsi yang digunakan untuk persamaan kuantitatif LPR adalah bahwa
pertambahan biomassa tanaman per satuan waktu tidak konstan tetapi tergantung pada berat awal tanaman. Bahwa keseluruhan tanaman yang dinyatakan dalam biomassa total tanaman dipertimbangkan sebagai suatu kesatuan untuk menghasilkan bahan baru tanaman (Sitompul dan Guritno, 1995).
LPR dapat digunakan untuk mengukur produktivitas (efisiensi) biomassa awal tanaman, yang berfungsi sebagai modal, dalam menghasilkan bahan baru tanaman. Perbedaan LPR dapat terjadi diantara spesies akibat perbedaan dalam laju fotosintesis dan efisiensi biomassa. Dalam aspek biosintesis, tanaman yang mengandung banyak protein per unit biomassa seperti tanaman kacang-kacangan akan membentuk biomassa yang lebih sedikit per satuan substrat (karbohidrat) yang tersedia dari tanaman yang mengandung protein lebih sedikit dari tanaman serealia. Energi yang dibutuhkan akan meningkat dengan peningkatan kandungan protein, sementara energi tersebut diperoleh dari proses perombakan (respirasi aerobik atau fermentasi) dari substrat. Tanaman yang tergolong ke dalam tanaman C4 seperti jagung mempunyai LPR yang tinggi, jauh lebih tinggi dari LPR tanaman golongan C3 seperti Theobroma cacao (Sitompul dan Guritno, 1995).
Luas Daun Spesifik (LDS) atau Specific Leaf Area (SLA)
organela fotosintesis. Kuanta cahaya merupakan faktor yang dominan dari biomassa tanaman dalam memicu aktifitas sifat dalam tanaman (genetik) yang mengendalikan nilai luas daun spesifik. Tanggapan luas daun spesifik kepada perubahan kuanta radiasi dalam jangka pendek cukup sensitif (Sitompul dan Guritno, 1995). Nilai luas daun spesifik yang semakin besar mengindikasikan daun semakin tipis dan nilai luas daun spesifik tidak berpengaruh langsung terhadap bobot biji (Sutoro et al., 2008).
METODOLOGI
Tempat dan Waktu
Penelitian dilaksanakan di Rumah Plastik di Kebun Percobaan Ilmu dan Teknologi Benih IPB, Leuwikopo, Dramaga, Bogor. Waktu pelaksanaan penelitian dimulai dari bulan Maret sampai dengan Juni 2011. Analisis klorofil dilakukan di Laboratorium Plant Analysis and Chromatography, Departemen
Agronomi dan Hortikultura, Fakultas Pertanian IPB. Analisis tanah dilakukan di Laboratorium Analisis Tanah, Balai Penelitian Tanaman Obat dan Aromatik,
Bogor.
Bahan dan Alat
Bahan tanaman yang digunakan adalah dua genotipe kedelai, yang terdiri dari genotipe toleran (Ceneng) dan genotipe peka (Godek) terhadap naungan. Media tanam yang digunakan adalah campuran tanah dan pupuk kandang yang dicampur seluruhnya dengan perbandingan 1:1 yang kemudian dimasukkan kedalam polibag. Bahan kimia yang digunakan meliputi alkohol dan aseton 80% untuk analisis klorofil. Karbofuran 3G untuk mencegah lalat bibit. Pestisida digunakan sesuai dengan kebutuhan. Peralatan yang dibutuhkan adalah alat ukur (meteran), timbangan digital, solarimeter, termohigrometer, label, paranet 50%, gelas ukur, mortar, spektrofotometer, polibag, plastik untuk pembuatan rumah plastik, bambu, kawat, tali, screen dan alat budidaya untuk menanam kedelai
dalam polibag.
Metode Penelitian
Percobaan disusun dengan menggunakan Rancangan Split Plot Design
dengan anak petak tersarang pada petak utama dengan tiga ulangan. Petak utama terdiri dari dua tingkat naungan dan faktor kedua adalah dua genotipe/varietas kedelai sebagai anak petak, sehingga terdapat 12 satuan percobaan. Petak utama
genotipe kedelai, yaitu G1=genotipe peka dan G2=genotipe toleran terhadap cahaya. Model aditif linier yang digunakan adalah:
Yijk=µ + Ni + Uk(Ni) + Gj + (NG)ij + Eijk
keterangan:
Yijk = nilai pengamatan pada kelompok ke-i yang memperoleh taraf dari
faktor naungan ke-j dan faktor genotipe kedelai ke-k
µ = nilai rataan umum
Ni = pengaruh aditif dari taraf naungan ke-i, i = 0, 1
Uk(Ni) = pengaruh ulangan ke-k dalam naungan ke-i , k = 1, 2, 3 Gj = pengaruh aditif dari taraf ke-j faktor genotipe, j = 1, 2
(NG)ij = pengaruh interaksi taraf ke-i faktor naungan dan taraf ke-j faktor genotipe kedelai
Eijk = galat percobaan
Pelaksanaan Penelitian Persiapan
Tahap awal penelitian ini adalah persiapan bangunan tanam yang berupa rumah plastik dan pemasangan paranet dalam rumah plastik sesuai perlakuan. Rumah plastik ini nantinya berfungsi sebagai penahan/pelindung tanaman dari hujan. Persiapan benih meliputi benih pengujian daya berkecambah. Langkah selanjutnya yaitu persiapan media tanam dan polibag. Media tanam dengan campuran tanah dan pupuk kandang (1:1) dicampur dan dimasukkan kedalam polibag. Pengukuran kapasitas lapang dilakukan setelah persiapan media tanam untuk menentukan volume penyiraman pada masing–masing polibag.
Analisis Tanah
Penanaman dan Pemeliharaan
Kegiatan pertama untuk sebelum penanaman adalah pengujian daya berkecambah benih kedelai dari dua genotipe. Pengujian daya berkecambah dilakukan di Laboratorium Benih Leuwikopo, Departemen Agronomi dan Hortikultura, IPB. Pengujian daya berkecambah ini menggunakan metode UKDdp dengan menggunakan media kertas buram. Hitungan pertama untuk menghitung daya berkecambah dilakukan pada hari ketiga dan hitungan kedua pada hari kelima setelah pengujian.
Karakteristik benih kacang-kacangan yang baik diantaranya adalah daya tumbuh tinggi, lebih dari 80%, tidak tercampur dengan varietas lain atau dapat dikatakan tingkat kemurniannya tinggi, yakni antara 98%-100%, memiliki kecepatan tumbuh (vigor) yang baik, biji berwarna mengkilat, tidak keriput,
bernas, dan bebas dari gigitan serangga, dan tidak tercampur dengan kotoran, gulma, atau biji tanaman lain (Fachruddin, 2000).
Penanaman dilakukan jika rumah plastik dan naungan sesuai perlakuan telah selesai dibuat dan siap untuk digunakan. Benih dari masing-masing genotipe untuk setiap ulangan ditanam dalam polibag. Penanaman dilakukan dalam polibag ukuran 35 cm×35 cm dengan tiga benih dalam satu polibag. Benih kedelai yang telah dimasukkan dalam polibag yang berisi media tanam diberi Karbofuran 3G sebanyak ± 3 butir/lubang untuk mencegah serangan serangga lalat bibit.
Pengamatan
Pengamatan agronomi yang dilakukan diantaranya:
1. Tinggi tanaman (cm)
Pengukuran tinggi tanaman dilakukan setiap minggu hingga tanaman mulai berbunga. Tinggi tanaman diukur dari kotiledon sampai titik tumbuh yang terletak diujung batang.
2. Jumlah daun trifoliat (helai)
Jumlah daun dihitung mulai daun trifoliat pertama sampai daun yang sudah terbuka penuh dan dilakukan setiap minggu hingga tanaman mulai berbunga.
3. Umur berbunga (HST)
Perhitungan dilakukan satu kali, yaitu saat tanaman sudah berbunga 75% dari jumlah tanaman contoh setiap perlakuan
Pengamatan analisis pertumbuhan diantaranya:
1. Indeks Luas Daun (ILD) atau Leaf Area Index (LAI)
Adalah luas daun (A) pada tiap satuan luas tanah (P)
ILD= LAI =
Pengukuran ILD ini dengan cara memotong bagian daun dari tanaman, kemudian dilakukan pengukuran terhadap luas masing–masing daun dari masing–masing tanaman contoh.
2. Laju Asimilasi Bersih (LAB) atau Net Assimilation Rate (NAR) g/m2/hari
Adalah jumlah total CO2 yang diambil tanaman dikurangi dengan jumlah yang hilang melalui respirasi. Dihitung dengan laju peningkatan bobot kering tanaman pada saat tertentu (t) tiap satuan luas daun (L) per satuan waktu (t)
NAR= ( ) ×(( ))
Bobot kering tanaman didapat dengan cara mengambil sampel tanaman beserta akarnya, kemudian ditimbang dan dikeringkan dengan menggunakan oven, selanjutnya ditimbang lagi untuk mendapatkan bobot kering.
3. Nisbah Luas Daun (NLD) atau Leaf Area Ratio (LAR) m2/g
Adalah perbandingan luas daun (L) terhadap bobot kering tanaman yang ada (W)
Luas daun didapat dengan cara yang sama seperti pada pengukuran luas daun ILD dan bobot kering juga didapat dengan cara yang sama seperti dalam mendapatkan bobot kering tanaman pada LAB.
4. Laju Pertumbuhan Relatif (LPR) atau Relative Growth Rate (RGR) g/g/hari
Adalah suatu peningkatan bobot kering (W) tiap satuan waktu (T)
LPR=RGR=
Bobot kering didapat dengan cara yang sama seperti dalam mendapatkan bobot kering tanaman pada LAB.
5. Luas Daun Spesifik (LDS) atau Specific Leaf Area (SLA) m2/g
Adalah hasil bagi luas daun (A) dengan berat daun (BD) seperti yang di-tunjukkan persamaan berikut.
LDS=
BD A
Semua pengamatan analisis pertumbuhan tanaman diukur sebanyak lima kali, yaitu pada stadium buku ketiga (V3), stadium mulai berbunga (R1), stadium berbunga penuh (R2), stadium mulai berpolong (R3), dan stadium berpolong penuh (R4).
Pengamatan karakter Fisiologi, yaitu:
Karakter fisiologi yang diamati adalah kandungan klorofil. Kandungan klorofil diukur lima kali, yaitu pada stadium buku ketiga (V3), stadium mulai berbunga (R1), stadium berbunga penuh (R2), stadium mulai berpolong (R3), dan stadium berpolong penuh (R4) pada daun ketiga dari pucuk dengan menggunakan metode Sims dan Gamon (2002).
Analisis klorofil di laboratorium dilakukan dengan cara sebagai berikut: 1. Contoh daun diambil dan digerus dengan mortar, kemudian ditambah aseton
80% secukupnya untuk memudahkan penggerusan.
2. Selanjutnya ekstrak klorofil pada mortar dipindahkan kedalam tabung reaksi 3. Kemudian ekstrak klorofil dalam tabung reaksi diencerkan dengan aseton 80%
hingga 10 ml dan ekstrak klorofil dipindahkan ke wadah spektrofotometer yang kemudian akan diukur oleh alat spektrofotometer
Klorofil a ( mol/100cm2)
= 0.01373 ×A663- 0.000897 ×A537- 0.0030464 ×A647 ×Fp ×Vol
Luas (cm2)×100
Klorofil b ( mol/100cm2)
= 0.02405 ×A647- 0.004305 ×A537- 0.00507 ×A663 ×Fp ×Vol
Luas (cm2)×100
Keterangan:
A = Nilai absorban yang terbaca pada spektrofotometer Fp = Faktor pengencer
Vol = Volume aseton 80%
Pengamatan biomassa tanaman meliputi biomassa akar, batang, daun, serta
HASIL DAN PEMBAHASAN
Kondisi Umum
Penelitian dilaksanakan mulai bulan Maret sampai Juni 2011 dalam kondisi terkontrol di rumah plastik. Penyiraman dilakukan secara manual untuk menggantikan kehilangan air oleh tanaman dengan cara pemberian air pada masing-masing polibag hingga mencapai kadar air kapasitas lapang secara berkala dua hari sekali. Data iklim mikro di lokasi penelitian meliputi suhu rata-rata selama penelitan sekitar 33.90C, kelembaban relatif sekitar 65%. Daya
ber-kecambah benih dari kedua genotipe kurang dari 80% sehingga penyulaman dilakukan pada saat tanaman berumur 2 MST untuk mempertahankan populasi.
Hama dan penyakit yang ditemui saat pelaksanaan penelitian berlangsung cenderung tidak banyak, hal ini disebabkan kondisi lingkungan penelitian yang cukup terkontrol dengan adanya rumah plastik. Beberapa hama yang dijumpai diantaranya adalah belalang dan kutu putih. Tidak ada penyakit yang dijumpai pada saat penelitian.
Pengendalian hama dilakukan pada saat pertama kali ditemukan adanya hama dan kemudian dilakukan secara teratur selama satukali dalam seminggu. Penyemprotan insektisida digunakan untuk menekan perkembangan hama belalang dan melindungi populasi tanaman kedelai. Pengendalian hama belalang menggunakan insektisida kontak Decis 25EC dengan dosis 0.5 ml/l.
Gulma yang dijumpai saat pelaksanaan penelitian diantaranya adalah
Mimosa pudica, Boreria Laevis, Phyllantus niruri, Oxlalis barerieli. Pengendalian
gulma dilakukan secara manual seminggu sekali.
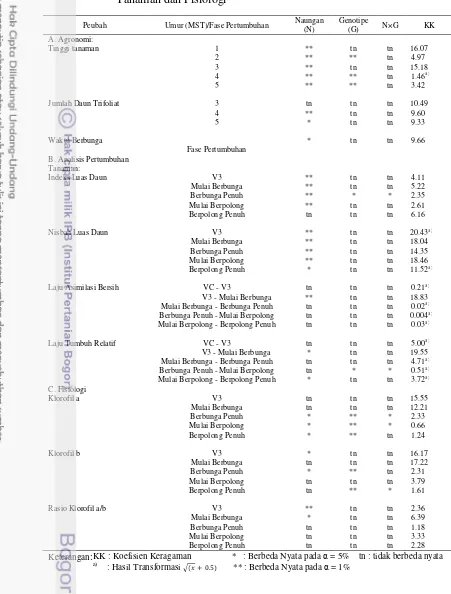
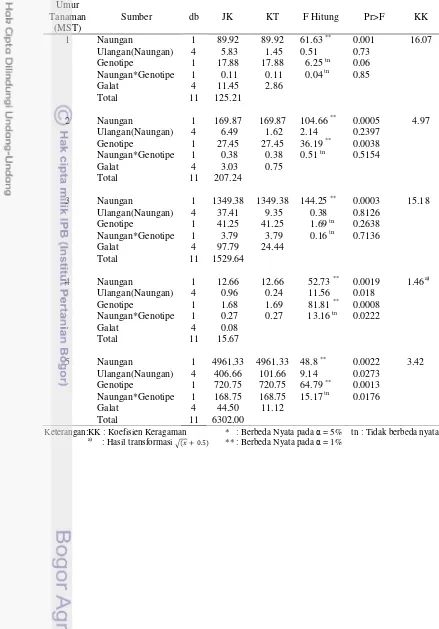
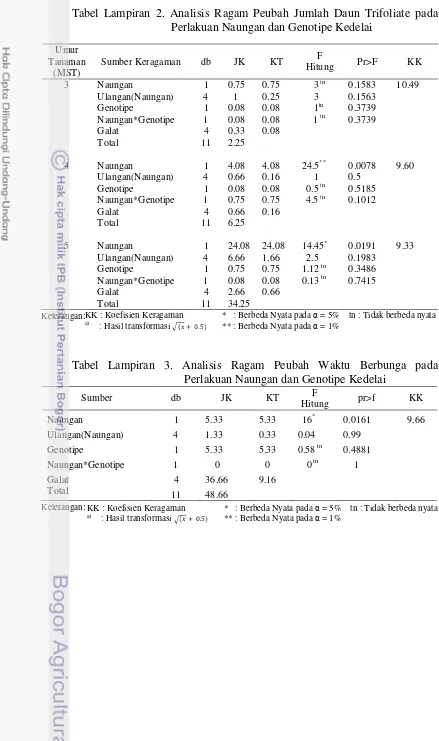
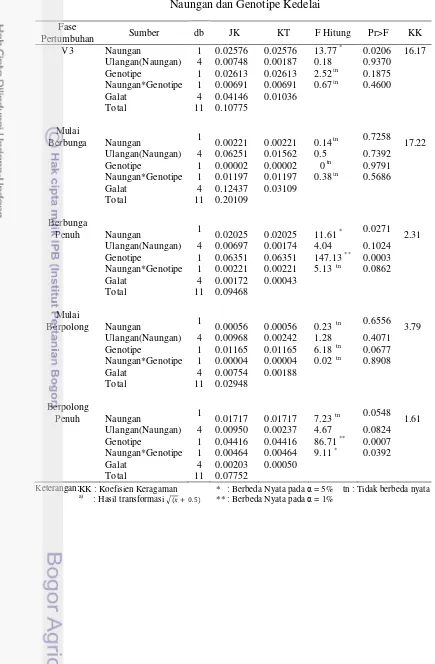
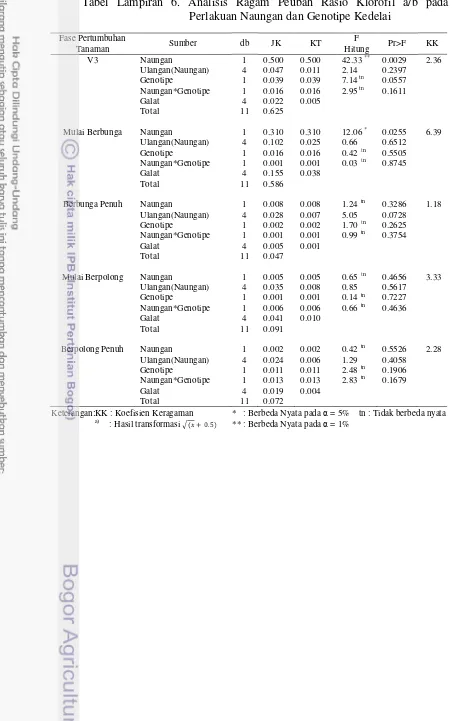
Tabel 1. Rekapitulasi Hasil Sidik Ragam Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Karakter Agronomi, Analisis Pertumbuhan Tanaman dan Fisiologi
Peubah Umur (MST)/Fase Pertumbuhan Naungan (N) Genotipe (G) N×G KK
A. Agronomi:
Tinggi tanaman 1 ** tn tn 16.07
2 ** ** tn 4.97
3 ** tn tn 15.18
4 ** ** tn 1.46a)
5 ** ** tn 3.42
Jumlah Daun Trifoliat 3 tn tn tn 10.49
4 ** tn tn 9.60
5 * tn tn 9.33
Waktu Berbunga * tn tn 9.66
Fase Pertumbuhan B. Analisis Pertumbuhan
Tanaman:
Indeks Luas Daun V3 ** tn tn 4.11
Mulai Berbunga ** tn tn 5.22
Berbunga Penuh ** * * 2.35
Mulai Berpolong ** tn tn 2.61
Berpolong Penuh tn tn tn 6.16
Nisbah Luas Daun V3 ** tn tn 20.43a)
Mulai Berbunga ** tn tn 18.04
Berbunga Penuh ** tn tn 14.35
Mulai Berpolong ** tn tn 18.46
Berpolong Penuh * tn tn 11.52a)
Laju Asimilasi Bersih VC - V3 tn tn tn 0.21a)
V3 - Mulai Berbunga ** tn tn 18.83
Mulai Berbunga - Berbunga Penuh tn tn tn 0.02a)
Berbunga Penuh - Mulai Berpolong tn tn tn 0.004a)
Mulai Berpolong - Berpolong Penuh tn tn tn 0.03a)
Laju Tumbuh Relatif VC - V3 tn tn tn 5.00a)
V3 - Mulai Berbunga * tn tn 19.55
Mulai Berbunga - Berbunga Penuh tn tn tn 4.71a)
Berbunga Penuh - Mulai Berpolong tn * * 0.51a)
Mulai Berpolong - Berpolong Penuh * tn tn 3.72a)
C. Fisiologi
Klorofil a V3 tn tn tn 15.55
Mulai Berbunga tn tn tn 12.21
Berbunga Penuh * ** * 2.33
Mulai Berpolong * ** * 0.66
Berpolong Penuh * ** tn 1.24
Klorofil b V3 * tn tn 16.17
Mulai Berbunga tn tn tn 17.22
Berbunga Penuh * ** tn 2.31
Mulai Berpolong tn tn tn 3.79
Berpolong Penuh tn ** * 1.61
Rasio Klorofil a/b V3 ** tn tn 2.36
Mulai Berbunga * tn tn 6.39
Berbunga Penuh tn tn tn 1.18
Mulai Berpolong tn tn tn 3.33
Berpolong Penuh tn tn tn 2.28
Keterangan: KK : Koefisien Keragaman * : Berbeda Nyata pada = 5% tn : tidak berbeda nyata
Karakter Agronomi
Tinggi Tanaman
Berdasarkan hasil rekapitulasi sidik ragam (Tabel 1) naungan berpengaruh sangat nyata terhadap tinggi tanaman mulai dari 1 MST hingga 5 MST (awal pembungaan), genotipe berpengaruh sangat nyata pada 1 MST dan berpengaruh nyata pada 2 dan 5 MST, sedangkan genotipe tidak berpengaruh terhadap tinggi tanaman pada 3 dan 4 MST, tidak terdapat interaksi antara naungan dengan genotipe. Analisis ragam peubah tinggi tanaman ditampilkan pada Tabel Lampiran 1. Pengaruh naungan terhadap tinggi tanaman dua genotipe kedelai kedelai disajikan pada Tabel 2.
Tabel 2. Pengaruh Naungan Terhadap Tinggi Tanaman Dua Genotipe Kedelai Kedelai
Perlakuan 1 MST 2 MST 3 MST 4 MST 5 MST
Naungan ---cm---
0% 7.79b 13.75b 21.95b 8.80b (42.75b) 77.16b
50% 13.26a 21.27a 43.16a 10.86a (71.58a) 117.83a
Genotipe ---cm---
Godek (G1) 9.30a 16.00b 30.70a 9.45a (57.50a) 89.75b
Ceneng (G2) 11.75a 19.02a 34.41a 10.20a (56.83a) 105.25a
Keterangan: angka dalam tanda kurung ( ) merupakan nilai rata-rata sebelum transformasi. Angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%
Berdasarkan Tabel 2 terlihat bahwa naungan meningkatkan tinggi tanaman. Tanaman kedelai yang di tanam pada kondisi ternaungi dengan naungan buatan sebesar 50% lebih tinggi dibandingkan dengan tanaman kedelai yang ditanam pada kondisi cahaya penuh (tanpa naungan) untuk kedua genotipe. Hal ini sejalan dengan hasil penelitian Mulyana (2006); Soverda et al. (2009);
Anggraeni (2010) menunjukkan bahwa pemberian naungan 50%-55% pada tanaman kedelai memberikan pengaruh berupa pertambahan tinggi tanaman dibandingkan dengan tanaman kedelai yang ditanam pada kondisi cahaya penuh. Kondisi ini terjadi karena tanaman yang berada di bawah naungan mengalami etiolasi. Kaufman, et. al. (1989) menjelaskan bahwa proses pemanjangan batang
melibatkan sel yang membelah dan memanjang secara aktif. Proses tersebut
Weafer dan Clements (1938) menyatakan bahwa batang dari tanaman yang ternaungi biasanya tumbuh lebih panjang daripada daun dari tanaman yang ditanam pada cahaya penuh sebagai akibat dari usaha untuk mendapatkan cahaya. Ciri khas pemanjangan batang pada tanaman yang ternaungi terutama akibat peningkatan panjang dari sel-sel tanaman. Fuller (1955) menyatakan bahwa etiolasi merupakan kondisi dimana tanaman tidak mendapat cukup cahaya kemudian tanaman tersebut gagal membentuk klorofil sehingga daun menjadi berwarna kekuningan dan menunjukkan beberapa struktur khusus seperti mudah rebah, batang yang sukulen, dan daun yang tidak berkembang.
Kedua genotipe (Ceneng dan Godek) yang ditanam dibawah naungan memiliki batang yang kecil, panjang, dan mudah rebah. Karamoy (2009) menjelaskan bahwa tanaman yang ditanam di bawah naungan hingga 40%
mengakibatkan penurunan diameter batang.
Menurut Crawley (1986) tanaman ternaungi dengan jarak daun dalam kanopi menjadi lebih rapat membuat tanaman meningkatkan pertumbuhan batang yang lebih tinggi untuk mendapatkan cahaya yang cukup guna bersaing. Tanaman yang demikian mengalokasikan sumberdaya secara proporsional kearah batang dan biasanya akan semakin bertambah tinggi selama diperlukan.
Hasil penelitian oleh Lakitan (1993) menunjukkan bahwa peningkatan pemanjangan batang sering menguntungkan bagi tumbuhan yang berkompetisi untuk mendapatkan cahaya. Lambers et al. (1998) menjelaskan bahwa tanaman
dengan mekanisme penghindaran naungan (shade-avoiding) yang tumbuh pada
kondisi lingkungan yang ternaungi akan meningkatkan pemanjangan batang dan tangkai, mengurangi jumlah cabang (meningkatkan dominasi apikal). Oosting (1958) menyatakan bahwa cahaya menghalangi produksi auksin maupun beberapa substansi pengontrol pertumbuhan pada tanaman. Tanaman yang tumbuh pada lingkungan yang gelap, menghasilkan auksin maksimum sehingga tumbuh memanjang secara cepat dan sukulen.
daun dari tanaman yang ternaungi berwarna hijau tua jika dibandingkan dengan tanaman yang tidak ternaungi.
Jumlah Daun Trifoliat
Berdasarkan hasil rekapitulasi sidik ragam (Tabel 1) naungan tidak berpengaruh nyata pada 3 MST, berpengaruh pada 4 dan 5 MST. Pengaruh genotipe dan interaksi antara naungan dan genotipe tidak menunjukkan pengaruh nyata mulai 3 MST hingga 5 MST (awal pembungaan). Analisis ragam peubah jumlah daun trifoliate disajikan pada Tabel Lampiran 2. Pengaruh naungan terhadap jumlah daun trifoliat dua varietas kedelai disajikan pada Tabel 3.
Tabel 3. Pengaruh Naungan terhadap Jumlah Daun Trifoliat Dua Varietas Kedelai
Perlakuan 2 MST 3 MST 4 MST 5 MST
Naungan ---helai---
0% 1.00a 3.00a 4.88a 10.16a
50% 1.00a 2.50a 3.66b 7.33b
Genotipe ---helai---
Godek (G1) 1.00a 2.83a 4.33a 8.50a
Ceneng (G2) 1.00a 2.66a 4.16a 9.00a
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%.
Dari Tabel 3 dapat dilihat bahwa jumlah daun trifoliat pada kedua genotipe yang ditanam pada kondisi lingkungan yang ternaungi lebih sedikit daripada genotipe yang ditanam pada kondisi cahaya penuh. Daun pada tanaman yang ternaungi menjadi lebih lebar jika dibandingkan dengan daun pada tanaman yang mendapat cahaya penuh (tanpa naungan). Hal serupa juga dilaporkan oleh
Tamaki dan Naka (1972); Mulyana (2006); Anggraeni (2010) bahwa pada tanaman yang ternaungi akan menurunkan jumlah daun, daun menjadi lebih tipis, dan lenih lebar.
Oosting (1958) menyatakan bahwa tanaman yang tumbuh pada lingkungan yang gelap tumbuh menjadi tinggi, kurus, dengan jarak antar buku yang panjang, dan relatif memiliki jumlah daun yang sedikit.
Dijelaskan lebih lanjut oleh Lambers et al. (1998), bahwa tanaman dengan
mekanisme penghindaran naungan (shade-avoiding) yang tumbuh pada kondisi
lingkungan yang ternaungi akan meningkatkan total luas daun dan mengurangi ketebalan daun.
Kaufman et al. (1989) menyatakan bahwa cahaya merah mendukung
perluasan daun, dengan meningkatkan pembelahan sel dan pembesaran sel. Hormon berperan dalam mengatur proses perluasan daun tersebut, khususnya sitokonin dan auksin. Hal tersebut mungkin disebabkan oleh cahaya merah bertindak untuk meningkatkan sintesis sitokinin dan auksin sehingga menyebabkan pelepasan satu atau kedua hormon tersebut maupun untuk meningkatkan sensitivitas dalam sel.
Daun pada kedua genotipe yang ditanam pada kondisi lingkungan tanpa
naungan memiliki daun yang lebih sempit. Oosting (1958) menjelaskan bahwa tanaman yang tumbuh dalam kondisi cahaya penuh (full sunlight) adalah lebih kecil, tebal, dan lebih keras dibandingkan dengan daun dari tanaman yang tumbuh pada naungan.
Weafer dan Clements (1938) menjelaskan bahwa akibat dari tanaman yang ditanam pada kondisi ternaungi adalah daun yang lebih tipis dengan satu lapis sel-sel palisade. Pada kondisi naungan yang sangat rapat, ketika sedikit sekali intensitas cahaya yang diterima oleh daun, seringkali jaringan palisade tidak dapat terbentuk dan sebagai akibatnya hanya terbentuk jaringan spons yang seragam.
Kaufman et al. (1989) menyatakan bahwa intensitas cahaya
mempengaruhi perluasan daun. Secara umum daun yang berada pada kondisi intensitas cahaya yang rendah akan cenderung memiliki permukaan yang luas, tipis, dan lebih hijau (lebih banyak klorofil per unit luas daun) jika dibandingkan dengan daun pada tanaman yang tumbuh pada kondisi cahaya matahari penuh. Daun yang lebar pada daun tanaman yang hanya mendapat sedikit intensitas cahaya digunakan agar daun tersebut dapat mendapatkan cahaya lebih banyak, hal ini merupakan ekspresi dari adaptasi lingkungan oleh daun.
penuh, lebih tebal karena daun tersebut membentuk sel-sel palisade yang lebih panjang dan terdiri dari beberapa lapisan.
Ehleinger (1988) dalam BjÖrkman dan Adams (1995) menyatakan bahwa
secara umum daun pada tanaman yang berkembang di bawah naungan memiliki orientasi horizontal dan tersusun dalam satu lapisan, sedangkan daun dari tanaman yang mendapat cahaya matahari penuh memiliki sudut daun lebih tajam.
Waktu Berbunga
Berdasarkan hasil rekapitulasi sidik ragam (Tabel 1) naungan mempengaruhi waktu berbunga pada tanaman kedelai. Genotipe dan interaksi tidak menunjukkan pengaruh terhadap waktu berbunga. Analisis ragam peubah waktu berbunga ditampilkan pada Tabel Lampiran 3. Pengaruh naungan terhadap peubah waktu berbunga dua varietas kedelai disajikan pada Tabel 4.
Tabel 4. Pengaruh Naungan terhadap Peubah Waktu Berbunga Dua Varietas Kedelai
Perlakuan Rata-Rata (HST)
Naungan
0% 32.00a
50% 30.66b
Genotipe
Godek (G1) 32.00a
Ceneng (G2) 30.66a
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%.
Berdasarkan Tabel 4 dapat dilihat bahwa naungan mempercepat waktu berbunga pada kedelai yang ditanam pada kondisi ternaungi. Kaufman (1989) menjelaskan bahwa tanaman yang dengan batang yang mengalami pemanjangan,
biasanya diikuti dengan pembungaan yang cepat.
Parker (2004) menjelaskan bahwa tanaman tomat yang ditanam pada
naungan memiliki lebih sedikit bunga, batang yang memanjang, daun menjadi lebih sedikit, lebih sedikit cabang, pada struktur internal daun dijumpai sedikit sel dan selapis sel pelindung yang tipis.
bentuk yang mudah berganti tergantung pada kualitas cahaya. Cahaya dengan panjang gelombang 660 nm dapat mengubah pigmen menjadi bentuk yang mengawali kejadian kearah terbentuknya induksi pembungaan.
Karakter Fisiologi Klorofil a
Berdasarkan rekapitulasi sidik ragam (Tabel 1) naungan tidak berpengaruh nyata pada fase vegetatif ketiga dan fase mulai berbunga, berbeda nyata pada fase berbungapenuh, mulai berpolong, dan berpolong penuh. Genotipe tidak berpengaruh nyata terhadap kandungan klorofil a pada fase vegetatif ketiga dan mulai berbunga, namun berpengaruh sangat nyata pada fase berbunga penuh, mulai berpolong, dan berpolong penuh.
Dari Tabel 5 dapat dilihat bahwa secara umum naungan meningkatkan jumlah klorofil a pada fase berpolong penuh. Interaksi (Tabel 6 dan 7) antara naungan dan genotipe pada fase berbunga penuh dan fase mulai berpolong menunjukkan bahwa naungan menyebabkan peningkatan klorofil a pada genotipe kedua, yaitu genotipe Ceneng sedangkan pada genotipe G1, yaitu Godek, tidak menunjukkan kenaikan. Pengaruh naungan terhadap jumlah klorofil a pada dua genotipe kedelai disajikan pada Tabel 5. Pengaruh naungan, genotipe, dan interaksinya terhadap jumlah klorofil a tanaman kedelai pada fase berbunga penuh disajikan pada Tabel 6 dan pada fase mulai berpolong disajikan pada Tabel 7..
[image:39.595.88.514.26.819.2]
Tabel 5. Pengaruh Naungan terhadap Jumlah Klorofil a pada Dua Genotipe Kedelai
Perlakuan V3 Mulai Berbunga Berpolong Penuh
Naungan --- mol/100cm2---
0% 1.93a 3.27a 4.17b
50% 1.98a 3.03a 4.38a
Genotipe --- mol/100cm2---
Godek (G1) 2.08a 3.19a 4.01b
Ceneng (G2) 1.84a 3.11a 4.47a
Tabel 6. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Jumlah Klorofil a Tanaman Kedelai pada Fase Berbunga Penuh
Genotipe 0% Naungan 50% Rata-Rata
--- mol/100cm2---
Godek (G1) 2.47Bb 2.80Ba 2.64b
Ceneng (G2) 3.02Ab 3.12Aa 3.07a
Rata-Rata 2.75B 2.96A
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%. Kapital: kolom
Tabel 7. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Jumlah Klorofil a Tanaman Kedelai pada Fase Mulai Berpolong
Genotipe 0% Naungan 50% Rata-Rata
--- mol/100cm2---
Godek (G1) 3.39Bb 3.44Ba 3.42b
Ceneng (G2) 3.52Ab 3.65Aa 3.58a
Rata-Rata 3.45B 3.55A
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%. Kapital: kolom
Kaufman et al. (1989) menyatakan bahwa klorofil a terdapat pada panjang gelombang 430 dan 662 nm, sedangkan klorofil b pada panjang gelombang 453 dan 642 nm. Klorofil a biasanya terdapat dalam jumlah dua kali lebih banyak daripada klorofi b. klorofil a dapat ditemukan pada hampir semua organisme fotosintetik. Graham et al. (2006) menyatakan bahwa klorofil a merupakan
klorofil yang paling banyak terdapat pada kloroplas daun. Klorofil a memantulkan cahaya hijau sehingga daun tanaman selalu tampak hijau. Spektrum cahaya yang diserap oleh klorofil a adalah merah dan biru-violet.
Jumlah klorofil a yang tinggi pada tanaman ternaungi dijelaskan oleh Langenheim dan Thimann (1982) yang menyatakan bahwa komposisi klorofil pada daun yang ternaungi lebih tinggi dan perbandingan klorofil dan karotenoid tinggi daripada daun pada tanaman dengan cahaya penuh. Hasil penelitian Muhuria et al. (2006); Mulyana (2006); Kisman et al. (2007); dan Anggraeni
(2010) menunjukkan bahwa genotipe Ceneng yang ditanam pada naungan 50% memiliki lebih banyak klorofil a daripada yang ditanam pada kondisi tanpa naungan.
Muhuria (2006) menyatakan bahwa daun yang ternaungi memiliki lebih banyak grana per volume kloroplas, kloroplas lebih besar, dan rasio klorofil yang lebih besar daripada daun yang berkembang pada kondisi cahaya matahari penuh.
Klorofil b
Berdasarkan hasil rekapitulasi sidk ragam (Tabel 1) naungan berpengaruh terhadap jumlah klorofil b pada fase vegetatif ketiga dan pada fase berbunga penuh. Genotipe berpengaruh sangat nyata pada fase berbunga penuh dan fase berpolong penuh. Interaksi antara naungan dan genotipe berpengaruh nyata pada saat fase berpolong penuh. Analisis ragam peubah klorofil b ditampilkan pada Tabel Lampiran 5. Pengaruh naungan terhadap jumlah klorofil b pada dua genotipe kedelai disajikan pada Tabel 8. Pengaruh naungan, genotipe, dan interaksi terhadap jumlah klorofil b tanaman kedelai fase berpolong penuh disajikan pada Tabel 9.
Tabel 8. Pengaruh Naungan terhadap Jumlah Klorofil b pada Dua Genotipe Kedelai
Perlakuan V3 Mulai
Berbunga
Berbunga Penuh
Mulai Berpolong
Naungan --- mol/100cm2---
0% 0.58b 1.009a 0.85b 1.13a
50% 0.67a 1.037a 0.93a 1.14a
Genotipe --- mol/100cm2---
Godek (G1) 0.67a 1.022a 0.82b 1.11a
Ceneng (G2) 0.58a 1.024a 0.97a 1.17a
Tabel 9. Pengaruh naungan, Genotipe, dan Interaksi terhadap Jumlah Klorofil b Tanaman Kedelai Fase Berpolong Penuh
Genotipe Naungan Rata-Rata
0% 50%
--- mol/100cm2
---Godek (G1) 1.27Ba 1.39Ba 1.33b
Ceneng (G2) 1.43Aa 1.47Aa 1.45a
Rata-Rata 1.35A 1.43A
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%. Kapital: kolom
Berdasarkan Tabel 8 Dapat dilihat bahwa secara umum naungan meningkatkan pembentukan klorofil b. Hal ini sejalan dengan penjelasan Weafer dan Clements (1966) bahwa tanaman ternaungi akan memiliki komponen klorofil yang lebih banyak dibandingkan dengan tanaman yang tumbuh pada kondisi cahaya penuh. Salisbury dan Ross (1991) menyatakan bahwa tanaman ternaungi lebih banyak mengandung klorofil b karena tiap kloroplas mempunyai lebih banyak grana dibandingkan dengan daun pada tanaman tanpa naungan.
Graham et al. (2006) menyatakan bahwa klorofil b dan karotenoid
merupakan pigmen asesoris. Korofil b dan karotenoid menyerap cahaya yang berbeda dengan klorofil a. Klorofil b dan karotenoid meningkatkan penyerapan dari cahaya tampak (visible light) yang berguna untuk fotosintesis. Klorofil b dan
karotenoid mentrasfer energi yang telah diserap ke klorofil a. Baik klorofil a, klorofil b, dan karotenoid menyerap energi dari cahaya tampak untuk fotosintesis.
Rasio Klorofil a/b
Berdasarkan hasil rekapitulasi sidik ragam (Tabel 1) naungan berpengaruh sangat nyata pada fase vegetatif ketiga dan berpengaruh nyata pada fase mulai berbunga. Genotipe dan interaksi antara genotipe dan naungan tidak berpengaruh
nyata terhadap rasio klorofil a/b. Analisis ragam peubah rasio klorofil a/b disajikan pada Tabel Lampiran 6. Pengaruh naungan terhadap rasio klorofi a/b
pada dua genotipe kedelai disajikan pada Tabel 10.
di-sampaikan oleh Jufri (2006); Muhuria (2006); Mulyana (2006); Kisman (2007) bahwa naungan menurunkan rasio klorofil a/b.
Tabel 10. Pengaruh Naungan terhadap Rasio Klorofi a/b pada Dua Genotipe Kedelai
Perlakuan V3 Mulai
Berbunga
Berbunga Penuh
Mulai Berpolong
Berpolong Penuh Naungan
100% 3.34a 3.24a 3.21a 3.04a 3.02a
50% 2.93b 2.92b 3.15a 3.09a 3.05a
Genotipe
Godek (G1) 3.08a 3.12a 3.19a 3.08a 3.01a
Ceneng (G2) 3.20a 3.04a 3.16a 3.05a 3.07a
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%.
Penjelasan mengenai rendahnya rasio antara klorofil a dengan klorofil b dikemukakan oleh Lambers et al. (1998) bahwa rasio antara klorofil a dan klorofil
b rendah pada daun dari tanaman yang ternaungi. Daun-daun tersebut memiliki klorofil lebih banyak untuk berasosiasi dengan Light Harvesting Complex (yang
mengandung lebih sedikit klorofil a daripada klorofil b) dibanding dengan fotosistem. Penurunan rasio klorofil a/b adalah karena refleksi dari pembentukan pada sistem LHC. Proporsi terbanyak dari LHC terdapat di grana terbesar dari grana daun-daun yang ternaungi. Secara umum lebih lanjut dijelaskan bahwa daun yang ternaungi akan lebih banyak mengandung klorofil per kloroplas, klorofil per berat kering, dan rasio klorofil a/b dibandingkan dengan daun tanaman yang tumbuh pada kondisi cahaya penuh.
Analisis Pertumbuhan Tanaman Indeks Luas Daun
Berdasarkan hasil rekapitulasi sidik ragam (Tabel 1) naungan berpengaruh
sangat nyata terhadap indeks luas daun pada fase vegetatif ketiga hingga fase mulai berpolong, tidak nyata pada fase berpolong penuh. Genotipe berpengaruh
daun pada dua genotipe kedelai disajikan pada Tabel 11. Pengaruh naungan, genotipe, dan interaksinya terhadap indeks luas daun pada fase berbunga penuh disajikan pada Tabel 12.
Sitompul dan Guritno (1995) menyatakan bahwa indeks luas daun merupakan perbandingan luas daun total dengan luas tanah yang ditutupi atau luas daun diatas suatu luasan tanah. Harga indeks luas daun >1 menggambarkan adanya saling menaungi diantara daun yang mengakibatkan daun yang ternaungi pada lapisan bawah tajuk mendapat cahaya yang kurang dan karenanya mempunyai laju fotosintesis yang lebih rendah dari daun yang tidak ternaungi.
Secara umum, tanaman kedelai akan meningkat nilai indeks luas daunnya sesuai dengan tahap perkembangan hingga mencapai luas daun maksimum. Bila dilihat pada Tabel 11, peningkatan maksimum indeks luas daun terjadi pada fase
pertumbuhan berpolong penuh. Menurut Gardner et al. (1991) dalam tajuk tanaman dengan nilai indeks luas daun yang tinggi, daun yang muda pada pucuk tanaman menyerap radiasi paling banyak, memiliki laju asimilasi CO2 yang tinggi, dan mentranslokasikan sejumlah besar hasil asimiasi ke bagian tumbuhan yang lain. Sebaliknya, daun-daun yang lebih tua pada dasar tajuk dan terlindung mempunyai laju asimilasi CO2 yang rendah dan memberikan lebih sedikit asimilasi kepada bagian tumbuhan yang lain.
Tabel 11. Pengaruh Naungan terhadap Indeks Luas Daun pada Dua Genotipe Kedelai
Perlakuan V3 Berbunga Mulai Berpolong Mulai Berpolong Penuh
Naungan ---m2/m2
---0% 0.67a 0.96b 4.96b 5.73a
50% 0.48b 1.35a 5.41a 6.21a
Genotipe ---m2/m2
---Godek (G1) 0.55b 1.16a 5.11a 6.09a
Ceneng (G2) 0.60a 1.15a 5.26a 5.85a
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%.
Berdasarkan Tabel 12 terlihat bahwa naungan meningkatkan indeks luas
Tetapi jika dilihat dari perbedaan rata-rata indeks luas daun pada kondisi tanpa naungan dan kondisi ternaungi, genotipe Ceneng juga mengalami kenaikan indeks luas daun.
Tabel 12. Pengaruh Naungan, Genotipe, dan Interaksinya terhadap Indeks Luas Daun pada Fase Berbunga Penuh
Genotipe Naungan Rata-Rata
0% 50%
---m2/m2---
Godek (G1) 1.69Ab 2.05Aa 0.55b
Ceneng (G2) 1.90Aa 2.05Aa 0.60a
Rata-Rata 1.79B 2.05A
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji lanjut DMRT 5%. Kapital: kolom.
Sitompul dan Guritno (1995) menyatakan bahwa indeks luas daun pada kebanyakan tanaman di lapangan adalah nol untuk tanaman yang ditanam dengan biji, dan selama beberapa minggu kemudian dapat berada dibawah 1.0 selanjutnya peningkatan indeks luas daun secara cepat hingga mencapai




![Growth responses of soybean [Glycine max (L ) Merr ] to drought and aluminium stress](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)