Efektivitas Cendawan Duddingtonia flagrans dan Saccharomyces cerevisiae Dalm Pengendalian Cacing Haemonchus contortus Pada Domba
Teks penuh
Gambar


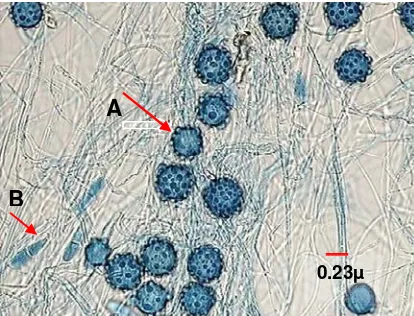

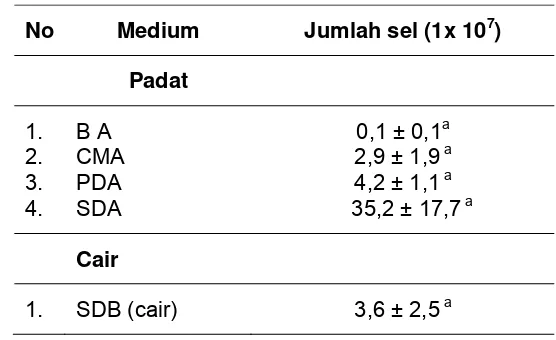
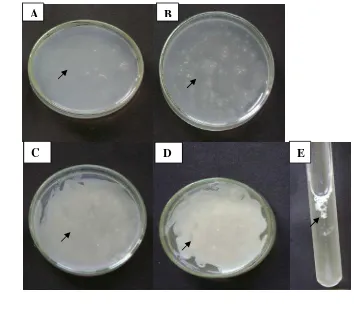

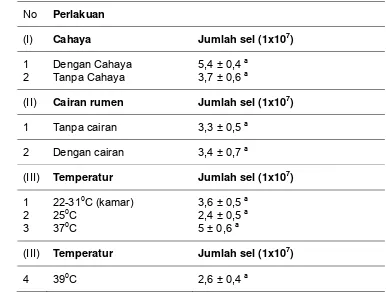

Dokumen terkait
Effe\ctif pada stadium beluru dewasa. Effektif pada stadium
Rataan jumlah telur cacing pada enam kelompok domba percobaan ..... Rataan berat badan pada enam kelompok domba Percobaan (kg)
Pada eksperimen kedua, sebanyak 30 botol biakan yang berisi 5 g tinja dari domba yang terinfeksi cacing Haemonchus contortus dengan jumlah telur cacing dalam tinja kurang
contortus dihitung dengan perkalian antara rata-rata berat tinja dalam satu hari dengan rata-rata jumlah ttgt campuran dibagi dengan rata- rata jumlah cacing betina
dan Haemonchus contortus pada sapi Madura di desa Kraton dan desa Pejagan Kecamatan Bangkalan Kabupaten Bangkalan Provinsi Jawa Timur.. Pengambilan sampel feses
Akan tetapi walaupun jumlah populasi Haemonchus contortus paling rendah pada waktu istirahat 105 hari, namun jika dihubungkan dengan sifat pertumbuhan rumput Brachiaria brizantha maka
Pada pengamatan patologi anatomi belum ditemukan perubahan yang nyata (P > 0,05) terhadap ukuran alat bantu reproduksi betina (vulva flep) dan jantan (gubernakulum dan
Larva masuk tubuh domba melalui mulut, termakan bersama rumput, kemudian larva masuk kedalam abomasum dan hidup di bagian mukosa, berkembang menjadi cacing dewasa, siap kawin
