DETEKSI VIGOR BlOKlMlAWl DAN VIGOR FlSlOLOGl
UNTUK FENOMENA PEMULIHAN VIGOR PADA TINGKAT AWAL
DETERIORASI DAN DEVIGORASI
BENlH KEDELAI
(Glycine
max (L.) Merr.) MELALCll PROSES INVIGORASI'
Oleh
ENY
WlDAJATl
AGR. 93510
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
DAFTAR SINGKATAN ATP D Dl3 DMRT DNA GDH KCT KST MDH MPC PK-r PKs UKDdp VDS v g VKT
v g b i o k
~ g f i r i o l
VP v s s
= Adenosine triphosphate
= Nilai Delta
= Daya Berkecambah
= Duncan Multiple Range Test
= Deoxyribonucleic acid
= Glutamat dehidrogenase
= Kecepatan Turnbuh
= ~ e s e r e m ~ a k a n Tumbuh
= Malat dehidrogenase
= Mesin Pengusang Cepat
= Periode Konservasi sebelurn tanam
= Periode Konservasi sebelum simpan
= Uji Kertas Digulung didirikan dalam plastik
= Vigor Daya Simpan
= Vigor
= Vigor Kekuatan Tumbuh
= Vigor Biokimiawi
= Vigor Fisiologi
= Viabilitas Potensial
RINGKASAN
ENY WIDAJATI. Deteksi Vigor Biokimiawi dan Vigor Fisiologi untuk Fenomena Pemulihan Vigor Pada Tingkat Awal Deteriorasi dan Devigorasi Benih Kedelai
(Glycine max (L.) Merr.) Melalui Proses Invigorasi. (Di bawah bimbingan
SJAMSOE'OED SADJAD sebagai ketua, M.S. SAENI, MAGGY T.
SUHARTONO, SATRlYAS ILYAS dan BAMBANG S. PURWOKO masing- masing sebagai anggota)
Tujuan penelitian adalah mempelajari proses deteriorasi dan devigorasi
yang ditengarai oleh asam fitat, P teresterifikasi oleh mitokondria, aktivitas enzim
fitase, aktivitas enzim peroksidase di dalarn benih kedelai yang memungkinkan
untuk dijadikan indikator vigor biokimiawi maupun DB dan KST sebagai indikator
mutu fisiologi. Tujuan lainnya adalah mempelajari tingkat pulih vigor sebagai
kriteria analisis benih dalam variasi benih yang mengalami deteriorasi dan
devigorasi. Penelitian ini juga bertujuan untuk mempelajari mekanisme invigorasi
pada lot benih yang pulih vigor dengan indikasi fisiologi dan indikasi biokimiawi.
Penelitian ini juga bertujuan untuk mempelajari aplikasi fenomena invigorasi pada
lot-lot benih yang diduga sudah mundur namun masih dalam batasan pulih vigor,
sehingga dapat ditentukan apakah lot benih tersebut dapat digunakan untuk tujuan
blending.
Benih yang digunakan adalah benih kedelai varietas Wilis. Penelitian terdiri
dari dua seri yaitu ; 1) Studi kriteria analisis vigor benih dengan mekanisme pulih
vigor d m 2) Pemanfaatan invigorasi untuk deteksi tingkat pulih vigor. Penelitian
pertarna mempelajari mekanisme pulih vigor, hubungan antara parameter vigor
fisiologi dan biokimiawi sebagai indikasi pulih vigor dan konsumsi energi selarna
perkecambahan. Penelitian kedua mempelajari aplikasi pulih vigor untuk kriteria
analisis vigor b e ~ h dan aplikasi pulih vigor dalam seed blending. Proses invigorasi
dengan meningkatnya tolok ukur DB, KST, aktivitas enzim peroksidase, aktivitas
enzim fitase, jurnlah P teresterifikasi dan menurunnya asam fitat benih. Berdasarkan
tolok ukur tersebut lot benih yang memiliki DB lebih tinggi dari 92 % dan lebih
rendah dari 77 % mengalami proses invigorasi sangat kecil. Proses invigorasi yang
besar terjadi pada lot benih dengan kisaran DB 77-92%, pada kisaran tersebut
perlakuan osmotic priming PEG 6000 dapat meningkatkan DB sampai 14%. Proses
invigorasi juga ditunjukkan oleh meningkatnya ketebalan pita protein pada
elektroforesis benih yang diberi perlakuan osmotic priming PEG 6000 maupun
KH2P04.
Tolok ukur DB, KST, aktivitas enzim peroksidase, asam fitat dan P
teresterifikasi dapat digunakan untuk deteksi ragam viabilitas benih deteriorasi
maupun devigorasi. Aktivitas enzim peroksidase, P teresterifikasi, DB dan KST
menurun sejalan dengan meningkatnya proses deteriorasi dan devigorasi. Pada benih
yang mengalami deteriorasi dan devigorasi lanjut menunjukkan kandungan asam
fitat tetap tinggi.
Diternukan adanya korelasi yang tinggi antara
v , ~ ~ ' ~ '
denganv,b'ok,
kecualipada tolok ukur P teresterifikasi pada beberapa taraf deteriorasi. Aktivitas enzim
peroksidase dan P teresterifikasi memiliki korelasi positif dengan tolok ukur DB dan
KST, sedangkan asam fitat memiliki korelasi negatif dengan kedua tolok ukur
tersebut.
Tingkat pulih vigor dapat digunakan untuk kriteria analisis benih vigor tinggi
dan sedang. Berdasarkan tolok ukur DB, KST, aktivitas enzim peroksidase dan asam
fitat, lot benih vigor tinggi memiliki DB lebih tinggi dari 93 % dan lot benih vigor
berdasarkan tolok ukur KST lot benih yang digunakan pada penelitian ini termasuk
kriteria vigor sedang.
Tingkat pulih vigor dapat digunakan untuk deteksi viabilitas blended seed lot dengan tolok ukur asam fitat dan KST. Blending antara benih vigor tinggi dan vigor sedang dapat &lakukan. Untuk benih v i g o ~ tinggi (DB sekitar 95 %) clan benih vigor
SUMMARY
ENY WIDAJATI. Biochemical and Physiological Vigor Detection for Vigor
Recovery in Early Deterioration and Devigoration of Soybean Seed (Glycine m a x
(L.) Merr) through the Invigoration Mechanism. (Under supervision of
SJAMSOE'OED SADJAD as chairman, M.S. SAENI, MAGGY T.
SUHARTONO, SATRIYAS ILYAS and BAMBANG S. PUWOKO as members
of advisory committe).
The objectives of the experiment were to study the mechanism of
deterioration and devigoration indicated by phosphorous metabolism; to obtain the
analysis criteria of seed viability; and to obtain the optimum ratio of blended seed.
The experiments were conducted at Seed Science and Technology Laboratory, Plant
Molecular and Cellular Biology Laboratory of IUC for Biotechnology, Bogor
Agricultural University from January 1997 to October 1998.
Wilis was the cultivar used throughout the experiment. The experiments
consisted of 1) study of seed vigor criteria based on vigor recovery mechanism and
2) application of invigoration to detect level of vigor recovery. The first step of this
experiment was conducted to study vigor recovery mechanism; and relationship
between the physiological and biochemical parameters as vigor recovery indicators
and the energy consumption during seed germination. The second step of this experiment was conducted to study vigor recovery application for seed vigor criteria
and to detect vigor recovery in relation to seed blending.
The germination capacity, uniformity of seedling growth, activity of
peroxidase and phytic acid content were found as parameters capable of detecting
seed viability of deteriorated or devigorated seed lots. Invigoration was indicated by
increase in germination capacity, uniformity of seedling growth and activity of
peroxidase, and decrease in phytic acid content. Vigor recovery level could be used
to analyze seed viability. Seed lots with germination capacity higher than 92 %
level of 14% germination capacity was obtained by the medium seed lot (77-92 %
germination capacity). Mechanism of seed invigoration was detected by increase in thickness of protein band in SDS-PAGE analysis of protein extracted from cotyledon of 4 day old seedling.
There was high correlation between physiological and biochemical vigor parameters. The activity of peroxidase and esterified P showed positive correlation with gemination capacity and uniformity of seedling growth, while phytic acid content showed negative correlation.
Vigor recovery could be used as criteria for vigor analysis. Based on analysis of germination capacity, uniformity of seedling growth, phytic acid content, activity of peroxidase and esterifed P, high vigor seed lot was determined for those showing germination capacity higher than 93 %, while such level for medium vigor seed lot ranged from 78-92%.
DETEKSI VIGOR BlOKlMlAWl DAN VIGOR FtSlOLOGl UNTUK FENOMENA PEMULIHAN VIGOR PADA TINGKAT AWAL
DETERlORASl DAN DEVlGORASl BENlH KEDELAI (Glycine max (L.)Merr.) MELALUI PROSES INVlGORASl
Oleh : ENY WIDAJATI
AGR 9351 0
Disertasi sebagai salah satu syarat untuk memperoleh gelar Doktor dalam Ilmu-ilmu Pertanian
pada
Program Pascasarjana lnstitut Pertanian Bogor
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR 1999
6
Judul Disertasi : DETEKSI VIGOR BlOKlMlAWl DAN VIGOR FlSlOLOGl UNTUK FENOMENA PEMULIHAN
VIGOR PADA TINGKAT AWAL DETERlORASl DAN DEVlGORASl BENlH KEDELAI (Glycine max
(L.) Merr.) MELALUI PROSES INVIGORASI Nama Mahasiswa : ENY WlDAJATl
Nomor Pokok : 93570
Menyetujui :
1 Komisi Pemb~rnblng
-/
-&7\p
-- Prof. Dr Ir Sjamsoe'oed Sadjad, MA(Ketua) ,
- & e %-> ,
</ Z N
/
,fly/*
Prof. Dr Ir M. Sri Saeni, MS
,
Dr Ir ~ a g g $ ~ h e n a w i d a Suhartono(Anggota) (Andota)
Dr Ir Hj. Satriyas Ilyas, MS
d
Dr Ir Barnbang Sapta Purwoko, MSc
(Anggota) (Anggota)
2. Ketua Program-Studi Agronomi
Dr Ir Sudirman Yahya, MSc
ram Pascasarjana
RIWAYAT HIDUP
PenuIis dilahirkan pada tanggal 6 Januari 1961 di Jakarta dari orang tua
Bapak Sukito dan Ibu Singgang Haryani.
Pada tahun 1979 penulis lulus dari SMA Negeri 28 Jakarta. Penulis
memperoleh gelar Sarjana Pertanian tahun 1983 dan Magister Sain tahun 1992 dari
Institut Pertanian Bogor.
Penulis bekerja sebagai staf pangajar di Jurusan Budi Daya Pertanian
KATA PENGANTAR
Puji dan syukur penulis panjatkan ke hadirat Allah SWT, karena hanya atas
perkenan-Nyalah penulis dapat menyelesaikan penulisan disertasi ini.
Pada kesempatan ini penulis menyampaikan terima kasih sebesar-besamya
kepada :
1 . Rektor Institut Pertanian Bogor dan Direktur Pascasarjana yang telah
memberikan kesempatan kepada penulis untuk mengikuti program S3 di
Institut Pertanian Bogor serta Team Managemen Program Doktor atas
beasiswa yang diberikan.
2. Prof. Dr Ir Sjamsoe'oed Sadjad, MA sebagai ketua komisi pembimbing atas
pengarahan dan bimbingannya mulai dari sejak awal kuliah sampai dengan
selesainya penulisan disertasi ini.
3. Prof. Dr Ir M.Sri Saeni, MS, Dr Ir Bambang S. Purwoko,MSc, Dr Ir Maggy
T. Suhartono dan Dr Ir Hj. Satriyas Ilyas, MS atas bimbingan dan
pengarahannya mulai dari menyusun rencana penelitian sarnpai dengan
penulisan disertasi ini.
4. Kepala laboratorium Ilmu dan Teknologi Benih, Jurusan Budi Daya Pertanian,
Kepala laboratorium Biologi Molekuler dan Seluler Tanaman, Kepala
laboratorium Mikrobilogi dan Biokimia PAU Bioteknologi, IPB beserta staf
yang telah memberikan ban- dan fasilitas selama penulis melaksanakan
penelitian.
5. Dr Ir M. Yusuf dan Dr Ir Sony Suharsono atas saran dan bantuannya selama
pelaksanaan penelitian.
6. Rekan Dr fr Tati Budiarti, MS, Dr Ir Setia Hadi, MS, Rahmat, Bambang K.
serta rekan-rekan di Iaboratorium Ilmu dan Teknologi Benih yang tidak dapat
disebutkan
S3.
7. Dr Ir Udin S. Nugraha, MS dan Dr Ir Yuliana Maria Diah Ratnadewi yang
telah bersedia menjadi penguji luar komisi pada ujian terbuka dan saran-saran
yang telah diberikan untuk penyempurnaan disertasi.
8. Suami dan anak-anak tersayang (Pardomuan, Brarn, Ririn dan Johan) atas
segala doa, bantuan dan kesabarannya selarna penulis menempuh program S3.
9. Kedua orang tua (Bapak Sukito d m Ibu S. Haryani) atas kasih sayang, doa dan
bantuan yang diberikan kepada penulis dengan tulus ikhlas.
10. Keluarga Bapak D.Silaban atas doa dan bantuannya.
Akhirnya penulis berharap semoga tulisan ini dapat bermanfaat untuk
pengembangan Ilmu Benih khususnya bidang Analisis Benih dan bermanfaat bagi
yang memerlukannya.
DAFTAR IS1
Halarnan
DAFTARISI
. . .
DAFTARTABEL. . .
DAFTARGAMBAR. . .
I
.
PENDAHULUANLatarBelakang
. . .
Tujuan Penelitian. . .
Hipotesis. . .
I1
.
TINJAUAN PUSTAKA. . .
Viabilitas Benih
...
Komposisi Cadangan Energi Benih
. . .
Metabolisme Senyawa P selama Perkecambhan
. . .
Invigorasi Benih
Konsurnsi Energi Benih
. . .
Blending
. . .
111
.
BAHAN DAN METODEPenelitian I
.
Studi Kriteria Analisis Vigor Benih dengan. . .
Mekanisme Pulih Vigor
Penelitian I1
.
Pemanfaatan Invigorasi untuk Deteksi. . .
Tingkat Pulih Vigor
IV
.
HASIL DAN PEWAHASANPenelitian I
.
Studi Kriteria Analisis Vigor Benih dengan Mekanisme Pulih Vigor. . .
l a
.
Studi Mekanisme Pulih Vigor...
. . .
.
1 b Hubungan
vgfiso'
danv,biok
1 c
.
Pemantapan Mekanisme Pulih Vigor. . .
Id
.
Konsurnsi Energi selama Perkecambahan...
Penelitian I1
.
Pemanfaatan Invigorasi untuk Deteksi T i g k a t. . .
Pulih Vigor
2a
.
Aplikasi Pulih Vigor untuk Kriteria Analisis. . .
Viabilitas Benih
2b
.
Aplikasi Pulih Vigor dalam Seed Blending. . . .
V.
KESIMPULAN. . .
DAFTAR TABEL
Nomor Halaman
. . .
1 Variasi rasio blending lot benih vigor tinggi dan sedang
2 Rekapitulasi nilai F dari hasil analisis ragam faktor tingkat
viabilitas yang diperoleh dengan deteriorasi maupun
devigorasi dan perlakuan osmotic priming dengan PEG
. . .
6000 terhadap beberapa tolok ukur viabilitas
3 Pengaruh tingkat viabilitas yang diperoleh dengan proses
deteriorasi dengan tolok ukur DB dan KST
. . .
Pengaruh tingkat viabilitas yang diperoleh dengan proses devigorasi dengan tolok ukur DB clan KST. . .
Pengaruh osmotic priming dengan PEG 6000 pada benih yang
mengalami deteriorasi dan devigorasi dengan tolok ukur
D B d a n K s ~
. . .
Pengaruh interaksi tingkat viabilitas yang diperoleh dengan
proses devigorasi dan osmotic priming dengan tolok ukur
aktivitas enzim peroksidase
. . .
Pengaruh interaksi tingkat viabilitas yang diperoleh dengan
proses deteriorasi dan osmotic priming dengan tolok ukur
. . .
kandunganasamfitat
Pengaruh tingkat viabilitas yang diperoleh dengan proses
...
devigorasi dengan tolok ukur kandungan asam fitat
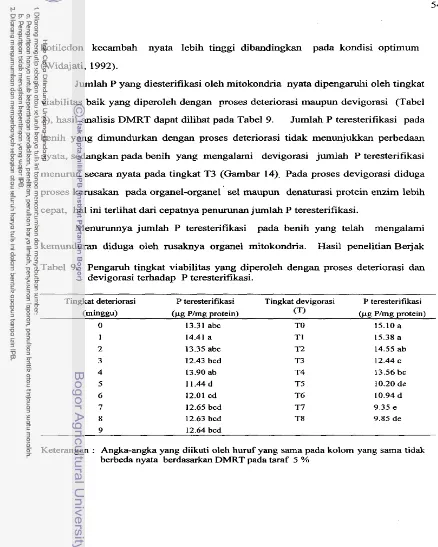
Pengamh tingkat viabilitas yang diperoleh dengan proses
. . .
deteriorasi dan devigorasi terhadap P teresterifikasi
Rekapitulasi nilai F pengaruh osmotic priming dengan PEG 6000
danKH2P01
. . .
Pengaruh osmotic priming dengan tolok ukur DB, &T, asam f3atdan aktivitas enzim fitase
. . .
12 Rata-rata panjang akar, panjang hipokotil, bobot kering kecambah
dan kandungan asarn fitat kecambah kedelai yang
. . .
ditumbuhkan dalam kondisi gravitasi < 1 dan kontrol.
13 Rekapitulasi nilai F analisis ragarn pengaruh variasi rasio
blending ( A ) dan osmotic priming dengan KH2P04 (B)
....
14 Pengaruh osmotic priming dengan tolok ukur DB, &T dan P
. . .
Pengaruh interaksi variasi rasio blending dengan osmoric priming
dengan KH2P04 terhadap tolok ukur kandungan asam fitat
Lampiran
Darnpak perlakuan osmotic priming PEG 6000 terhadap lot benih
dengan variasi tingkat deteriorasi.
. . .
Darnpak perlakuan osmotic priming PEG 6000 terhadap lot benih
dengan variasi tingkat devigorasi.
. . .
. . .
Komposisi larutan untuk ekstraksi rnitokondria
Komposisi larutan untuk elektroforesis.
. . .
Analisis ragam pengaruh tingkat viabilitas yang diperoleh denganproses deteriorasi dan osmotic priming dengan tolok ukur
DB
. . .
Analisis ragam pengaruh tingkat viabilitas yang diperoleh denganproses deteriorasi dan osmotic priming dengan tolok ukur
KST
. . .
Analisis ragam pengaruh tingkat viabilitas yang diperoleh denganproses deteriorasi dan osmotic priming dengan tolok ukur
. . .
alctivitas enzim peroksidase.
Analisis ragam pengaruh tingkat viabilitas yang diperoleh dengan
proses deteriorasi dan osmotic priming dengan tolok ukur
asamfitat
. . .
Analisis ragam pengaruh tingkat viabilitas yang diperoleh denganproses deteriorasi clan osmoticpriming dengan tolok ukur
P
. . .
teresterifikasi.
Analisis ragam pengaruh tingkat viabilitas yang diperoleh dengan
proses devigorasi dan osmotic priming dengan tolok ukur
DB
. . .
Analisis ragam pengaruh tingkat viabilitas yang diperoleh denganproses devigorasi dan osmotic priming dengan tolok ukur
Analisis ragam pengaruh tingkat viabilitas yang diperoleh dengan
proses devigorasi dan osmotic priming dengan tolok ukur
. . .
aktivitas enzim peroksidase
Analisis ragam pengaruh tingkat viabilitas yang diperoleh dengan
proses devigorasi dan osmotic priming dengan tolok ukur
asamfitat
. . .
Analisis ragam pengaruh tingkat viabilitas yang diperoleh denganproses devigorasi dan osmotic priming dengan tolok ukur P
. . .
teresterifikasi.
Analisis ragam pengaruh osmoticpriming dengan tolok ukur DB
Analisis ragam pengaruh osmotic priming dengan tolok ukur
KST
. . .
Analisis ragam pengaruh osmotic priming dengan tolok ukur
asamfitat
. . .
Analisis ragam pengaruh osmotic dengan tolok ukur
. . .
aktivitas enzim fitase
Rata-rata DB, KST, kandungan asam fitat dan P teresterifikasi
. . .
oleh mitokondria pada berbagai rasio blending
Analisis ragam pengaruh variasi rasio lot benih dan osmotic
. . .
priming dengan tolok ukur DB.Analisis ragam pengaruh variasi rasio lot benih dan osmotic
priming dengan tolok ukur K S ~
. . .
Analisis ragam pengaruh variasi rasio lot benih dan osmotic
. . .
priming dengan tolok ukur asam fitatAnalisis ragam pengaruh variasi rasio lot benih dan osmotic
DAFTAR GAMBAR
Nomor -' Halaman
Teks
Garis-garis viabilitas benih dalam konsepsi Steinbauer -
. . .
Sadj ad (Sadj ad, 1994) 7
Diagram sintesis fitin dan mekanisme penimbunan fitin
. . .
(Bewley dan Black, 1994) 13
Skema mekanisme sintesis ATP (Anderson dan Gupta,
1986)
. . .
17 Diagram pergerakan benih dalam alat blender conveyorspiral vertikal (Greg el al., 1970)
. . .
25Teoritis tentang aplikasi osmotic priming untuk
. . .
indikator blending. 25
Skema pelaksanaan penelitian.
. . .
27. . .
Metode osmotic priming. 28
. . .
Alat untuk membuat kondisi gravitasi < 1. 3 1
Teoritis kriteria vigor benih berdasarkan tingkat pulih
vlgor
...
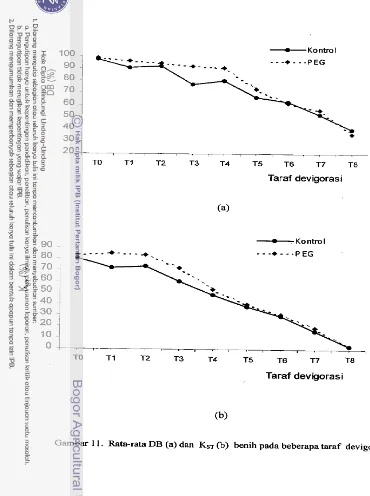
32Rata-rata DB (a) dan KST (b) benih pada beberapa taraf
. . .
deteriorasi 45
Rata-rata DB (a) dan &T @) benih pada beberapa taraf
. . .
devigorasi 47
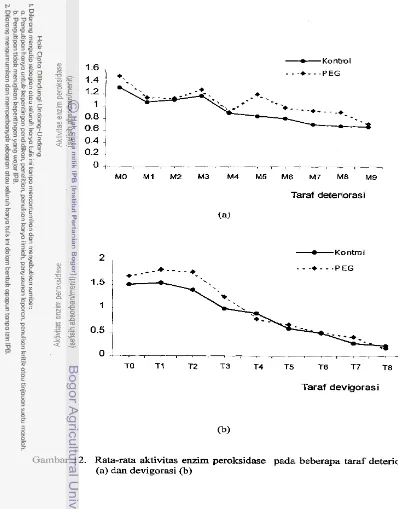
Rata-rata aktivitas enzim peroksidase pada beberapa
taraf deteriorasi (a) dan devigorasi (b)
...
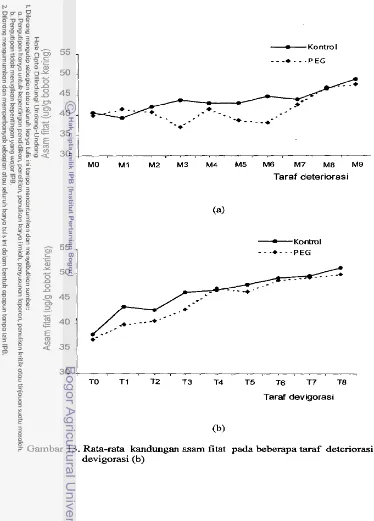
50Rata-rata kandungan asam fitat pada beberapa taraf
deteriorasi (a) dan devigorasi (b)
...
53Rata-rata P teresterifikasi pada beberapa taraf
deteriorasi (a) clan devigorasi (b)
. . .
5 5Hubungan antara aktivitas enzim peroksidase dengan
DB pada beberapa taraf deteriorasi (a) clan
. . .
devigorasi (b) 5 8
Hubungan antara aktivitas enzim peroksidase dengan
KST pada beberapa taraf deteriorasi (a) dan
Hubungan antara asam fitat dengan DB pada beberapa
. . .
taraf deteriorasi (a) dan devigorasi (b).
Hubungan antara asam fitat dengan I& pada beberapa
. . .
taraf deteriorasi (a) dan devigorasi (b)
Hubungan antara P teresterifikasi dengan DB pa&
beberapa taraf deteriorasi (a) dan devigorasi (b)
. .
Hubungan antara P teresterifikasi dengan %T pada
beberapa taraf deteriorasi (a) dan devigorasi (b)
. .
Pemindahan gugus fosfat dari fosfoenolpiruvat ke ADP
(Lehninger, 1990)
. . .
Analisis SDS-PAGE pola pita protein yang diekstrakdari kotiledon kecambah kedelai b e m u r 4 hari
. .
~enampakan kecambah pada kondisi kontrol (a) dan
. . .
gravitasi < 1 (b)
Kurva tingkat pulih vigor pada beberapa taraf
deteriorasi dengan tolok ukur DB.
. . .
Kurva tingkat pulih vigor pada beberapa taraf devigorasi. . .
dengan tolok ukur DB.
Kurva tingkat pulih vigor pada beberapa taraf deteriorasi dengan tolok ukur I(ST.
. . .
Kurva tingkat pulih vigor pada beberapa taraf
devigorasi dengan tolok
ukur
I(ST.. . .
Kuwa tingkat pulih vigor pada beberapa taraf
deteriorasi dengan tolok ukur aktivitas enzim
peroksidase.
. . .
Kurva tingkat pulih vigor pada beberapa taraf
devigorasi dengan tolok ukur aktivitas enzim
peroksidase.
. . .
Kurva tingkat pulih vigor pada beberapa tarafdeteriorasi dengan tolok ukur asam fitat.
. . .
Kurva tingkat pulih vigor pada beberapa taraf devigorasi dengan tolok ukur asarn fitat. . .
Kurva tingkat pulih vigor pada beberapa taraf. . . .
deteriorasi dengan tolok ukur P teresterifikasi Kurva tingkat pulih vigor pada beberapa tarafdevigorasi
. . .
Kurva tingkat pulih vigor pada beberapa variasi rasio blending dengan tolok ukur DB
. . .
Kurva tingkat pulih vigor pada beberapa variasi rasio blending dengan tolok ukur I(ST
. . .
Kurva tingkat pulih vigor pada beberapa variasi rasioblending dengan tolok ukur asam fitat
. . .
Kurva tingkat pulih vigor pada beberapa variasi rasio blending dengan tolok ukur P teresterifikasi. . .
I. PENDAHULUAN
Latar Belakang
Selama periode penyimpanan benih mengalami kemunduran yang dise-
babkan oleh faktor-faktor alami. Proses ini disebut deteriorasi. Kemunduran
benih dapat juga tejadi oleh tindakan non alami, misalnya oleh deraan Mesin
Pengusang Cepat (MPC) dengan menggunakan uap etanol. Untuk membedakan
dari istilah deteriorasi, proses kemunduran non alami oleh Sadjad (1994) disebut
devigorasi. Beberapa peneliti menyebutnya sebagai deteriorasi terkontrol.
Perubahan yang terjadi akibat deraan uap etanol menurut hasil penelitian Pian
(1981) adalah menurunnya potensi tumbuh rnaksimum, daya berkecambah dan vigor
benih yang merupakan perwujudan fisiologi. Perwujudan biokimiawi akibat deraan
uap etanol adalah menurunnya aktivitas enzim, respirasi benih, laju sintesis dan
meningkatnya kebocoran hasil metabolisme. P e m j u d a n sitologik terlihat pada
kerusakan tudung akar embrio benih yaitu terlepasnya plasma sel dari dinding sel
dan hubungan antar sel menjadi renggang.
Perubahan metabolik, sitologik dan genetik terjadi selama proses kemun-
duran viabilitas benih. Pada benih-benih yang mengalami kemunduran te jadi
akumulasi krornosom yang rusak, denaturasi lipoprotein membran sel dan asam
nukleat (Roberts, 1972). Hasil penelitian Pian (198 1) menunjukkan adanya
kesarnaan antara deteriorasi dan devigorasi dilihat secara fisiologi, biokimiawi,
maupun sitologi.
Studi ultrasbuktur pada tudung akar dari beberapa tingkat kemunduran
benih, menunjukkan bahwa semua organel sel terpengaruh oleh proses penuaan.
Namun pada lot benih yang mengalami kemunduran tipe I dengan protoplas masih
tersebut menghilang setelah imbibisi 48 jam. Hal ini menunjukkan adanya
mekanisme perbaikan. Kemunduran tipe I1 dengan protoplas sudah tidak
beraturan dan organel-organel sudah mengalami degenerasi, tidak dapat pulih
kembali. Demikian pula pada tingkat kemunduran yang lebih lanjut pada tipe I11
(Roberts, 1972).
Faktor internal benih dan kondisi lingkungan kadang-kadang menyebabkan
pertumbuhan bibit lambat atau abnormal. Benih-benih yang sudah mengalami
p e n m a n viabilitas biasanya sangat peka terhadap kondisi sub optimum. Salah satu
upaya untuk mengatasi kondisi tersebut diperlukan suatu perlakuan sebelum tanam
sehingga benih dapat mengatasi kondisi sub optimum, rnetabolisrne perkecambahan
yang terjadi lebih awal dan pengaruh pen- benih dapat dikurangi.
Perlakuan tertentu pada benih yang telah mengalami kemunduran dapat
meningkatkan vigor. Proses bertambahnya vigor benih disebut invigorasi (Sadjad,
1994). Murray d m Wilson (1987) mengemukakan bahwa salah satu usaha untuk
menambah vigor benih adalah dengan perlakuan priming. Priming adalah suatu
metode untuk mengatur jurnlah air yang diimbibisi oleh benih, serta mengatur
kecepatan masuknya air ke dalam benih. Ada beberapa metode priming yaitu
hydropriming, osmotic priming dan matriconditioning. Hydropriming yaitu priming
dengan cara merendam benih dalam air beberapa jam lalu diikuti dengan perlakuan
inkubasi pada kondisi kelembaban nisbi 100% (Koch, Hoffinan dan Steiner , 1992 ;
Fujikura e t al., 1993). Osmotic priming yaitu priming dengan cara menempatkan
benih dalam larutan osmotik, metode ini sering juga disebut osmopriming atau
Fenomena invigorasi ditunjukkan oleh indikasi fisiologi maupun biokimiawi.
Perlakuan osmotic priming dengan PEG 6000 pada benih kedelai yang telah
mengalami devigorasi dapat meningkatkan Daya Berkecambah (DB), Kecepatan
Tumbuh (I(CT), Keserempakan Tumbuh (I(ST) dan persentase pemunculan bibit
(Nursandi, 1990; Sagala, 1990 ; Shatters et al., 1994). Hasil penelitian Shatters et
al. (1994) menunjukkan adanya peningkatan intensitas pewarnaan pada analisis
izoenzim glutamat dehidrogenase (GDH) pada benih yang diberi perlakuan osmotic
priming dibanding kontrolnya. Kandungan protein terlarut lebih tinggi pada benih
yang diberi perlakuan osmotic priming dib&ding kontrol (Smith dan Cobb, 1992).
Hasil penelitian Armstrong dan McDonald (1992) menunjukkan bahwa pada
perlakuan osmotic priming terjadi pemantapan integritas membran.
Viabilitas benih merupakan fokus pengembangan ilmu benih (Sadjad, 1992).
Dalam pengembangan ilmu benih tersebut diharapkan dapat ditemukan indikasi-
indikasi viabilitas benih yang dapat diukur dengan tolok ukur yang spesifik. Pada
penelitian ini diharapkan dapat diperoleh indikasi viabilitas yang spesifik dalam
kaitannya dengan invigorasi benih.
Proses invigorasi ditunjukkan oleh tingkat pulih vigor yaitu selisih nilai
viabilitas benih, baik yang dinyatakan dengan tolok ukur fisiologi maupun
biokimiawi antara lot yang diinvigorasi dan kontrolnya (tanpa invigorasi). Hasil
penelitian Widajati, Suwarno dan Murniati (1990) pada benih kacang tanah
menunjukkan bahwa benih vigor tinggi dan vigor rendah tidak menunjukkan
peningkatan vigor dengan perlakuan invigorasi, sedangkan pada benih vigor sedang
terjadi peningkatan vigor secara nyata. Fenomena yang sarna juga terjadi pada
benih kedelai (Sagala, 1990) dan benih jagung (Setiarini, 1995). Hasil tersebut
dengan garis Nilai Delta pada Periode I11 konsepsi Steinbauer-Sadjad. Analog
dengan garis Nilai Delta, maka fenomena pulih vigor tersebut diduga dapat
digunakan untuk paiameter viabilitas suatu lot benih. Proses invigorasi pada
penelitian ini diharapkan dapat digunakan untuk kriteria viabilitas suatu lot benih.
Lot benih pada hakekatnya m e ~ p a k a n campuran individu-individu benih baik,
sedang dan jelek, oleh karena itu informasi viabilitas benih mengikuti kurva
distribusi normal. Dalam prakteknya dua atau lebih lot benih dapat diblending.
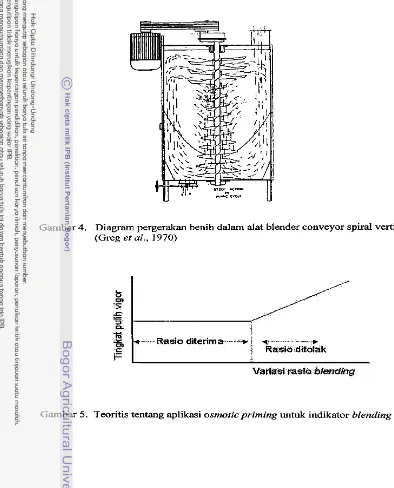
sehingga didapatkan komposisi lot benih yang lebih baik. Greg et al. (1970)
mendefinisikan blending adalah mencampur dua atau lebih lot benih, sehingga
dihasilkan lot benih yang lebih homogen.
Pemanfaatan benih yang sudah menurun viabilitasnya dalam budi daya tanarnan
diduga akan dapat menghasilkan pertanaman yang lebih seragam di lapang jika
benih yang sudah menurun viabilitasnya diblending dengan benih berviabilitas
tinggi. Penentuan dapat tidaknya benih tersebut diblending diduga dapat dilakukan
dengan uji invigorasi. Lot benih yang sudah menurun viabilitasnya dan setelah uji
invigorasi dapat pulih vigor termasuk kelompok benih vigor sedang dan dapat
digunakan untuk blending. Blended seed mempakan campuran benih vigor tinggi
dan sedang dengan berbagai variasi rasio. Batas rasio yang dapat diterima adalah
rasio-rasio yang memiliki delta relatif konstan apabila campuran lot tersebut
diinvigoraqi. Jika ha1 ini dapat dilakukan maka penambahan waktu dan tenaga untuk
penyularnan dapat ditiadakan dan benih yang sudah mengalami kemunduran dapat
dimanfaatkan dan pertanaman yang dihasilkan lebih baik.
Senyawa P di dalam benih sangat penting peranannya dalam metabolisme
perkecambahan benih. Fitin adalah kompleks garam kalsium, magnesium dan
kecil namun merupakan surnber fosfat dan unsur mineral penting bagi benih.
Selain itu fitin merupakan cadangan Fe, Mn dan Cu (Bewley dan Black, 1985).
Benih-benih yang pulih vigor dan diberi perlakuan invigorasi diduga mengalami
peningkatan aktivitas enzim-enzim hidrolisis, terrnasuk enzim fitase sebagai
penghidrolisis fitin. Pemanfaatan cadangan P dalam benih yang mengalami
deteriorasi dan devigorasi akan menurun. Perlakuan invigorasi diduga dapat
meningkatkan pemanfaatan cadangan P selama perkecambahan.
Tujuan Penelitian
Penelitian bertujuan mempelajari proses deteriorasi dan devigorasi yang
ditengarai oleh asam fitat, P teresterifikasi oleh mitokondria, aktivitas enzim fitase,
aktivitas enzim peroksidase di dalam benih kedelai yang memungkinkan untuk
&ja&ikan indikator vigor biokimiawi maupun DB dan ST sebagai indikator mutu
fisiologi. Tujuan lainnya adalah mempelajari tingkat pulih vigor sebagai kriteria
analisis benih dalam variasi benih yang mengalarni deteriorasi dan devigorasi.
Penelitian ini juga bertujuan untuk mempelajari mekanisme invigorasi pada lot
benih yang pulih vigor dengan indikasi fisiologi dan indikasi biokimiawi. Penelitian
ini juga bertujuan untuk mempelajari aplikasi fenomena invigorasi pada lot-lot benih
yang diduga sudah mundur narnun masih dalam batasan pulih vigor, sehingga dapat
ditentukan apakah lot benih tersebut dapat digunakan untuk tujuan blending.
Hipotesis
Hipotesis yang diajukan dalam penelitian ini adalah :
1. Tingkat viabilitas clan kepulihan vigor benih dari lot benih yang mengalami
deteriorasi maupun devigorasi dapat diindikasikan dengan indikasi fisiologi dan
2. Kandungan asarn fitat, aktivitas enzirn fitase, aktivitas enzim peroksidase,
jumlah P teresterifikasi oleh mitokondria, serta pola pita protein dapat
rnenjelaskan mekanisme invigorasi.
3. Indikasi kecambah kuat pada pengujian vigor benih berkorelasi positif dengan
lot benih yang masih mengalami pulih vigor
4. Tingkat invigorasi dapat rnengindikasikan vigor benih.
5 . Pada lot benih yang pulih vigor dapat dilakukan blending, narnun lot benih hasil
11. TINJAUAN PUSTAKA
Viabilitas Benih
Viabilitas benih merupakan daya hidup benih yang ditunjukkan oleh
fenomena pertumbuhan benih atau gejala metabolismenya. Periode viabilitas benih
merupakan suatu perjalanan waktu dari seluruh hidup benih. Pada Konsepsi
Steinbauer-Sadjad (Garnbar 1) viabilitas dimulai dari antesis sampai benih mati.
Periode viabilitas dibagi atas Periode I sebagai Periode Pembangunan Benih,
Periode I1 yaitu Periode Simpan dan Periode 111 yaitu Periode Kritikal (~adjad,
1994).
Keterangan:
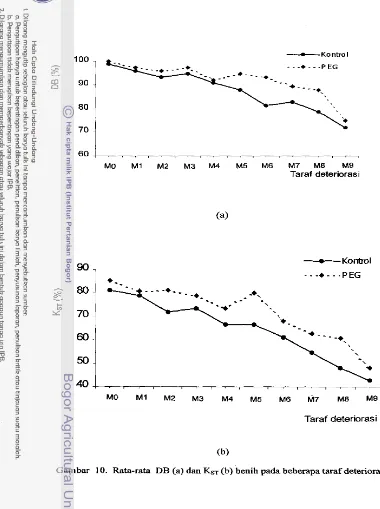
Vp = Viabilitas potensial; Vg = Vigor; PKs = Periode Konservasi sebelum simpan; PK.r = Periode Konservasi sebelum tanam; Vss = Viabilitas sesungguhnya; D = Nilai Delta.
Gambar I . Garis-garis viabilitas benih dalam Konsepsi Steinbauer- Sadjad (Sadjad, 1994)
Periodisasi viabilitas benih tidak sekadar menjabarkan proses viabilitas be-
[image:29.526.39.459.27.592.2](Sadjad, 1994). Dengan demikian viabilitas benih dalarn bentuk garis dapat
menjabarkan status viabilitas benih secara absolut d m simulatif. Lot benih pada
Periode I1 mengindikasikan bahwa lot benih masih memiliki vigor yang tinggi,
sebaliknya lot benih yang sudah berada pa& Pcriode I11 mengindikasikan lot benih
yang tidak memiliki vigor untuk disimpan lagi.
Parameter lot benih mencakup Viabilitas Potensial (Vp) dan Vigor (V, )
(Sadjad, 1994). Apabila lot benih memiliki pertumbuhan normal pada kondisi
optimum, lot benih itu memiliki kemampuan potensial. Masalah yang dihadapi
adalah bahwa kondisi di lapang produksi tidak selalu dalam kondisi optimum,
sehingga apabila lot benih tersebut menghadapi kondisi sub optimum, kemampuan
potensial itu belurn tentu dapat mengatasinya. Lot benih mempunyai kemampuan
lebih dari potensial, apabila mampu menghasilkan pertanaman normal dalam kondi-
si sub optimum. Lot benih demikian mempunyai parameter viabilitas yang disebut
vigor yang tinggi
.
Vigor benih merupakan unsur kualitas benih yang dinyatakan sebagai suatu
potensial untuk berkecambah cepat dan pertumbuhan bibit yang cepat pada kondisi
lapang yang beragam (Heydecker, 1972). Vigor benih berbeda dengan viabilitas
benih atau kemampuan berkecambah. Dalam suatu lot benih persentase viabilitas
atau perkecambahan memberikan informasi tentang perkiraan jurnlah maksimurn
bibit yang kemungkinan dihasilkan dalam kondisi laboratorium yang optimum.
Indeks vigor suatu lot benih mengindikasikan kemungkinan penampakan di lapang,
sehingga lebih mendekati perkiraan pertanaman di lapang, kecepatan tumbuh ta-
naman dan produksi. Ching (1982) mengemukakan bahwa ekspresi vigor benih
dipengaruhi oleh beberapa faktor yaitu faktor genetik, kondisi perkembangan,
Abdul Baki (1980) dalam Ching (1982) mengemukakan beberapa me-
kanisme biokimia yang dapat dikaitkan dengan vigor benih yaitu : (1) Kecepatan
dan kemampuan reorganisasi dan pulihnya kembali kondisi sitoplasma dan kondisi
membran pada awal imbibisi benih ; (2) efisiensi dalam penggunaan substrat se-
derhana misalnya glukosa untuk komponen selular ; (3) kecepatan sintesis ECNA
dan protein ; (4) sintesis dan kompetensi mitokondria khususnya enzim-enzim siklus
Krebs, senyawa untuk transport elektron (sitokrom) dan efisiensi fosforilasi ; (5)
stabilitas protein dan enzim ; (6) integritas plastida
Ching (1982) menyatakan bahwa vigor benih ditinjau dari beberapa aspek : (1)
efisiensi kepulihan dan reaktivasi keseluruhan sistern yang ada pada benih, semakin
kompeten awal pembentukan sistem-sistem dari membran, enzim, protein, asam
nukleat dan organel-organel sel maka semakin tinggi vigor benih tersebut ; (2)
sintesis yang cepat dan cukup bagi enzim-enzim dan organel untuk degradasi
cadangan makanan dalam mensuplai substrat untuk perturnbuhan bibit; (3) kecepatan
penyarnpaian informasi genetik dalam transkripsi dan translasi mRNA untuk enzim-
enzim anabolik dan protein struktural, tRNA untuk sintesis protein dari jaringan dan
organ yang berbeda-beda, rRNA untuk ribosom dan replikasi DNA untuk sel-sel
baru ; (4) adanya lingkungan mikro biosintesis yang optimum khususnya substrat,
energi, koenzim, kofaktor, efektor, aktivator, kekuatan ionik, pH, air, suhu, oksigen
dan sebagainya.
Untuk mendeteksi parameter viabilitas lot benih tertentu digunakan tolok
ukur yang spesifik, misalnya tolok ukur DB untuk parameter Vp dan Keserempakan
Tumbuh (KsT) merupakan tolok ukur Vg (Sadjad, 1994)
.
Vigor Kekuatan Turnbuh(VKT) dapat dicerminkan oleh keserempakan pertumbuhan benih. Pertanaman yang
tinggi. Hasil penelitian Saenong (1986) menunjukkan bahwa benih dengan
keserempakan tumbuh tinggi mernpunyai vigor daya simpan (VDS) yang tinggi.
Oleh karena itu K S ~ dapat dijadikan tolok ukur V D ~ .
Viabilitas benih diindikasikan oleh berbagai tolok ukur, baik tolok ukur yang
secara langsung menilai pertumbuhan benih, maupun secara tidak langsung dengan
menilai gejala metabolismenya atau mengamati kondisi beberapa organel sel
(Sadjad, 1994). Kandungan ATP pada benih yang telah mengalami proses imbibisi
merupakan indikasi viabilitas benih (Ching, 1973). Pada Brassica napus L.
ditunjukkan bahwa kandungan ATP benih yang sudah dilembabkan selarna 4 hari
berkorelasi positif dengan panjang kecarnbah, bobot basah maupun bobot kering
kecarnbah. Perbedaaan viabilitas benih yang disebabkan oleh proses kemunduran
alami maupun buatan dengan suhu tinggi, menunjukkan kandungan ATP lebih
rendah secara nyata dibandingkan benih baik.
Ching er al. (1977) meneliti beberapa indikasi vigor benih pada Hordeum
vulgare L. yaitu bobot benih, kandungan ATP bibit urnur 3 hari, kandungan total
adenosin fosfat bibit umur 3 hari, aktivitas enzim a amilase endosperma bibit umur
5 hari, bobot kering bibit umur 7 hari dan panjang bibit urnur 3 hari dikorelasikan
dengan kecepatan tumbuh di lapang. Semua indikasi yang diteliti menunjukkan
korelasi positif, namun bobot benih, kandungan ATP, kandungan total adenosin
fosfat dan bobot kering bibit urnur 7 hari merupakan indikasi vigor benih yang baik
untuk memprakirakan kecepatan tumbuh di lapang.
Komposisi Cadangan Energi Benih
Cadangan energi di dalam benih sebagian besar tersusun oleh karbohidrat,
kalah penting, walaupun dalam jumlah yang sedikit misalnya, fitin, alkaloid, zat
pengatur tumbuh dan vitamin. Pati merupakan karbohidrat yang paling umum
ditemukan sebagai cadangan makanan benih. Pati tersimpan dalam bentuk
amilosa dan amilopektin (Bewley dan Black, 1985).
Lemak sebagai cadangan makanan benih dalam bentuk trigliserida yaitu ester
dari gliserol dan asam lemak (Copeland, 1976). Asam lemak dibedakan atas asam
lemak jenuh dan tidak jenuh. Asam lemak tidak jenuh memiliki satu atau lebih
ikatan rangkap misalnya oleat, linoleat dan linolenat. Sebaliknya asarn lemak yang
tidak ada ikatan rangkapnya disebut asam lemak jenuh, misalnya palmitat, stearat
dan laurat. Komposisi asam lemak pada benih kedelai berdasarkan persentase bobot
lemak sebagai berikut : palmitat 11 %, stearat 2 O h , oleat 20 %, linoleat 64 %,
linolenat 3 %.
Osborne (1924) dalam Bewley dan Black (1994) mengklasifikasikan protein di
dalam benih menjadi empat kelas berkaitan dengan kelarutannya, albumin yaitu
protein yang larut di dalam air dan buffer pH netral ; glutelin yaitu protein yang larut
di dalam asam atau basa, prolamin yaitu protein yang larut di dalam alkohol 70-90 %
; dan globulin yaitu protein yang larut dl dalam larutan garam. Protein simpan yang
terdapat pada benih kedelai yang rnasak sebagian besar adalah globulin yaitu
glycinin dan P-conglycinin.
Fitin adalah kompleks garam kalsium, magnesium dan kalium dari asam
12
dan kadang-kadang Na (Bewley dan Black, 1994). Strvktur molekul asam fitat
sebagai berikut
Kandungan unsur anorganik dalam fitin bervariasi antar spesies. Pada kedelai
kandungan Mg (0.22%), Ca (0.13 %) dan K (2.18 %) berdasarkan persentase bobot
kering. Pada sel benih fitin berada dalam bentuk globoid di dalamprotein body.
Dari hasil penelitian pada perkembangan endosperma benih jarak dengan
menggunakan mikroskop elektron, hipotesis tentang sintesis fitin dapat dilihat pada
Gambar 2. Ada kemungkinan bahwa aparatus golgi juga berperan dalam
pengemasan dan pembentukan vesikel transpor
.
Jalur biosintesis asam fitat adalah: myo-inositol-1-P disintesis dari Glc-6-P selanjutnya ditambahkan Lima fosfat
sehingga terbentuk asam myoinositol heksafosfat. ATP berperan sebagai donor P
Keterangan :
CER = retikulum endoplasmik sisternal
CW = dinding sel, G = globoid, L = oil body
M
= mitokondria , PB =protein body, pl = plastidTahapan sintesis fitin sebagai berikut : (I) sintesis fitin berasosiasi dengan retikulum
endoplasmik sisternal sebelum dikemas dalam vesikel transpor ; (11) terjadi migrasi
ke membran vakuola ; (111) d m IV) te rjadi fusi-fitin dengan membran vacuola ; (IV)
dan (V) partikel fitin dilepas ke dalam lumen vakuola ; (VI) kondensasi dalam
vakuola dan mernbentuk globoid
Garnbar 2. Diagram sintesis fitin dan mekanisme penimbunan fitin (Bewley d m Black, 1994).
Hasil penelitian Earley dan DeTurk (1944) menunjukkan bahwa selama
perkembangan benih jagung te rjadi akumulasi P dalam bentuk fitin dalarn jumlah
besar. Peran fisiologi asam fitat adalah sebagai cadangan simpan P, terlihat dari
sebagian besar P pada benih gandum yang masak dalam bentuk asam fitat
(Williams, 1970). Senyawa ini &an dihidrolisis menjadi fosfat anorganik, pada
awal-awal perkecarnbahan. Asam fitat juga terlibat dalam sintesis protein (Morton
Akumulasi asam fitat selama perkembangan benih sangat dipengaruhi oleh
kondisi lingkungan tumbuh tanaman induk. Pada kondisi cekaman kecepatan
akumulasi maksimum asam fitat pada aleuron benih gandum terjadi pada hari ke-23,
sedangkan pada kondisi normal terjadi pada hari ke-28 (Williarns,l970).
Kandungan asam fitat dalam benih bervariasi antar kultivar maupun spesies.
Hasil analisis kandungan asam fitat pada benih jagung Arjuna dan Genjah Kuning
sarna, sedangkan pada Hawai SS. kandungannya lebih rendah (Suwarno, 1995).
Metabolisme Senyawa P selama Perkecambahan
Senyawa P di dalam benih sebagian besar berada dalam bentuk organik,
sedangkan dalam bentuk anorganik jumlahnya sangat kecil. Di antara senyawa
organik yang mengandung P di dalam benih adalah asam nukleat, fosfolipid,
nukleotida dan fitin (Mayer dan Mayber, 1982). Fitin merupakan mayoritas
cadangan P di dalam benih, kurang lebih 80 % dari total P dalam benih. Sebagian
besar P ada dalam bentuk terikat, sehingga ortofosfat menjadi faktor pembatas
dalam beberapa reaksi yang berkait dengan P, misalnya sintesis fosfolipid, protein,
asam nukleat dan proses pembentukan energi.
Pada benih kapas yang sedang berkecambah kandungan fitin merosot tajam
dan hampir habis pada hari ke-6 (Ergle d m Guin (1959) dalarn Mayer dan Mayber,
1982), sedangkan jumlah fosfat anorganik meningkat sampai 16 kali. Menurunnya
kandungan fitin secara tajam selama perkecambahan benih juga terlihat pada benih
gandum, oat, lettuce dan benih-benih lain. Berdasarkan fenomena tersebut
diperkirakan bahwa fitin merupakan cadangan P dl dalam benih yang sangat
Pada benih kedelai kandungan asam fitat sebagai produk hidrolisis fitin oleh
fitase menunjukkan peningkatan mulai hari ke-1 sampai hari ke-3 secara tajam, lalu
terjadi penurunan secara tajam pada hari k e 4 (Widajati, 1992). Pada benih yang
ditumbuhkan pada kondisi cekaman tekanan osmotik terlihat jumlah sisa asarn yang
lebih banyak dibanding benih yang ditumbuhkan dalarn kondisi optimum. Hal ini
menunjukkan bahwa benih yang ditanam dalam kondisi sub optimum kurang dapat
memanfaatkan asam fitat di dalam benih, sehingga energi untuk perkecambahan
lebih sedikit. Kurangnya energi untuk perkecambahan terlihat pada bobot kering
kecambah yang lebih rendah.
Enzim fitase merupakan enzim yang mendegradasi fitin pada awal
perkecambahan benih. Enzim tersebut memiliki semua sifat enzim fos-
fomonoesterase non spesifik (Williams, 1970). Jika enzim tersebut mengkatalisis
reaksi pembentukan ATP, maka tipe reaksinya sebagai berikut :
Asam fitat
+
inositol pentafosfat inositol tetrafosfatn
ADP ATP ADP ATP
Fitase menghidrolisis fitin melepaskan P, kation-kation dan myoinositol.
Fosfat yang dilepaskan dimanfaatkan dalam proses respirasi atau pembentukan makromolekul di bagian poros embrio. Myoinositol diduga digunakan untuk pem-
bentukan dinding sel, karena senyawa tersebut adalah prekursor pentosil dan uranosil
yang biasanya berasosiasi dengan pektin dan polisakarida-polisakarida dinding sel
lainnya (Bewley dan Black, 1985).
Perubahan senyawa P dalarn benih yang sedang berkecambah diantaranya
adalah perubahan nukleosida dan nukleotida (Brown (1965) dalam Mayer dan
sedangkan ATP meningkat hampir 10 kali pada jam-jam pertama imbibisi sampai 16
jam, kemudian setelah itu terjadi penurunan.
ATP m e ~ p a k a n senyawa berenergi yang diperlukan untuk setiap jalur bi-
osintesis maupun kerja biologi misalnya pergerakan, transpoe, perbaikan dan
sebagainya. Selama perkecambahan, poros embrio tidak hanya memiliki enzim,
substrat dan kofaktor untuk sintesis ATP, tetapi juga enzim-enzim yang meng-
konversi adenin dan adenosin menjadi ATP (Ching, 1982). ATP selular dibentuk
dari ADP melalui sistem transport elektron Woroplas, mitokondria, membran
retikulum endoplasmik, membran nukleus, melalui jalur glikolisis dan siklus Krebs.
Pada benih yang sedang berimbibisi tampaknya ada beberapa kemungkinan
mekanisme sintesis ATP (Anderson dan Gupta, 1986). Skema mekanisme sintesis
ATP dapat dilihat pada Gambar 3 . Dari beberapa substrat tersebut, adenin dan ade-
nosin nyata meningkatkan sintesis ATP, masing-masing sebesar 206 % dan 175 %.
Substrat tersebut juga terdapat dl dalam benih. Hasil penelitian Anderson (1977)
menunjukkan adenin dan adenosin menurun selama perkecambahan dan dikonversi
menjadi AMP oleh fosforibosilpirofosfat transferase d m adenosin kinase lalu
menjadi ATP.
Sintesis ATP pada poros embrio kedelai dipengaruhi oleh jenis substrat se-
bagai prekursor ATP (Anderson, 1977). Beberapa jenis substrat yang digunakan
untuk larutan imbibisi adalah adenin, adenosin, inosin, hiposantin, guanin,
guanosin, A m , CAMP, D-ribosa, buffer fosfat pH 6 dan glukosa. Dengan meng-
gunakan larutan substrat untuk imbibisi, sintesis ATP menjadi Iebih besar 93 %
dibanding kontrol aquades.
Pada benih kering jumlah ATP sangat kecil, setelah imbibisi jumlah ATP
Tingkat ADP tetap rendah dan tidak banyak berubah selama proses imbibisi
(Anderson dan Gupta, 1 986).
ADENINE ADENOSINE
DE NOVO
\ /
0- AMP-ADP-ATP
Gambar 3. Skema mekanisme sintesis ATP (Anderson dan Gupta, 1986)
Hasil penelitian Abu-Shakra dan Ching (1967) menunjukkan bahwa
terdapat perbedaan aktivitas mitokondria pada benih kedelai yang baru dipanen
beberapa bulan dibandingkan benih yang sudah berumur 3 tahun. Walaupun benih
yang sudah disimpan 3 tahun memiliki DB 80 %, namun kecepatan tumbuhnya
lebih Iambat. Mitokondria dari kedua lot benih tersebut memperlihatkan absorbsi
oksigen yang sarna, namun jurnlah fosfat anorganik yang diesterifikasi oleh
mitokondria dari benih yang baru dipanen dua kali lebih banyak dari benih tua.
Invigorasi Benih
..
Invigorasi adalah suatu proses bertambahnya vigor benih (Sadjad, 1994).
Pada benih-benih yang telah mundur sarnpai tingkat tertentu, pemulihan pada
organel-organel selnya dapat terjadi (Berjak (1968) dalam Roberts, 1972). Pada
kemunduran tipe I, struktur mitokondria yang abnormal dapat pulih kembali
lanjut, ketidak normalan dari struktur organel sel tidak dapat pulih. Invigorasi
dapat dilakukan dengan metode priming (Murray dan Wilson, 1987).
Priming pada prinsipnya adalah mengatur jurnlah air yang diimbibisi oleh
benih, serta mengatur kecepatan masuknya air ke dalam benih (Murray dan Wilson,
1987). Perlakuanpriming sering pula disebut dengan istilah lain yaitu conditioning.
Beberapa metode priming dapat dilakukan yaitu :
1 . Hydropriming yaitu priming dengan cara merendam benih dalam air selarna 5
jam lalu diikuti dengan perlakuan inkubasi pada kondisi kelembaban nisbi
100 % dan suhu kamar selama 3 hari (Fujikura et al., 1993; ~ o c h et al., 1992)
2 . Osmotic priming yaitupriming dengan cara menempatkan benih dalam larutan
osmotik, misalnya larutan PEG, KH2P04, KC1, K3P04, KN03, MgS04, NaC1,
gliserol d m sebagainya (Murray dan Wilson, 1987). Osmolicpriming sering juga
disebut dengan istilah osmopriming (Khan, 1992; Garcia, Jimenez dan Vazquez-
Ramos, 1995) atau osmoconditioning (Khan, 1992 ; Armstrong dan McDonald,
1992).
3 . Matriconditioning yaitu priming dengan cara menempatkan benih pada media
padatan yang ielah dilembabkan, misalnya pada vermikulit (Khan, 1992; Jeng
d m Sung, 1994) dan abu gosok (Yunitasari, 1995)
Menurut Murray dan Wilson (1987) perlakuan osmotic priming pada beberapa
tanaman, misalnya barley, bit, wortel, bawang, kedelai dan jagung dapat mem-
percepat perkecarnbahan pada suhu rendah dan memperbaiki keserempakan tumbuh.
Hasil penelitian Brocklehurst, Dearman dan Drew (1987) menunjukkan bahwa
osmotic priming pada beberapa benih sayuran menunjukkan adanya invigorasi yang
terlihat dari perkecambahan yang cepat dan seragam. Perlakuan osmotic priming
seledri dan 47 % pada bawang dibanding kontrol. Pengeringan kembali setelah
benih diberi perlakuan osmotic priming menunjukkan bobot tanaman yang lebih
tinggi 34 % pada wortel, 142 % pada seledri d m 32 % pada bawang
Hasil penelitian Munthe (1 992) menunjukkan bahwa perlakuan osmotic
priming pada benih kacang tanah dengan larutan KHzP04 menunjukkan invi-
gorasi, terlihat dari tolok ukur bobot kering kecambah total yang lebih tinggi
bila dibandingkan dengan perlakuan PEG 6000. Namun pada tolok ukur DB dan
Ks7, kedua perlakuan tersebut memberikan pengaruh yang sama. Perlakuan osmo-
tik -7.5 bar atau setara dengan 41.54 g KH2P04 I 1 air atau 260.6 g PEG '6000/1
air merupakan perlakuan yang menghasilkan peningkatan DB terbesar yaitu dari
78 % menjadi 87 %.
Perlakuan PEG 6000 dapat meningkatkan vigor benih kedeiai. Proses invi-
gorasi tersebut terlihat dari waktu perkecambahan yang lebih awal, peningkatan
kecepatan berkecambah dan pemunculan bibit pada s d u sub optimum yaitu ~ O C
dan 1 5 O ~ . Konsentrasi optimum untuk osmotic priming adalah 30 g PEG
6000/100 ml air dengan periode waktu 4-8 hari. Perlakuan pengeringan ke
bobot kering awal tidak mempengaruhi efek dari osmotic priming (Knypl dan
Khan, 1981)
Perlakuan osmotic priming dengan PEG pada benih tomat menunjukkan
adanya proses invigorasi. Konsentrasi PEG 6000 29 dengan periode imbibisi
12 hari dapat meningkatkan DB d m panjang akar serta bobot kering bibit. Semakin
meningkatnya konsentrasi PEG dan semakin lamanya periode imbibisi
menyebabkan p e n m a n viabilitas (Saxena dan Singh, 1987). Hasil penelitian
Alvrado, Bradford dan Hewitt (1987) pada benih tomat juga menunjukkan bahwa
menunjukkan adanya proses invigorasi terlihat dari pemunculan bibit yang lebih
awal dan seragam serta merniliki bobot kering tanaman dan indeks luas d a m yang
Iebih tinggi dibanding kontrol.
Perlakuan hydropriming sebelum semai akan menghasilkan pengaruh yang
baik bila perlakuan tersebut dilakukan dalam kondisi aerob (Koch et al., 1992).
Pada benih Triticum aestivum L. perlakuan hidrasi dalam kondisi aerob
menghasilkan
b,
DB, panjang plurnula dan bobot kering akar yang lebih tinggidibanding kondisi anaerob. Pada benih yang baik (DB 95 O/o) tidak terlihat
perbedaan pengaruh antara kontrol dengan perlakuan hidrasi. Perbedaan antar per-
lakuan terlihat pada benih dengan DB 79 % yang diperoleh melalui proses pengu-
sangan cepat, dengan cara benih berkadar air 18 % d i s h p a n pada suhu 45 OCselama
4 hari. Pada benih Avena sativa L. hidrasi benih sebelum tanam dalam kondisi aerob
dapat mempercepat tumbuh benih di lapang maupun produksi yang lebih tinggi
(Hoffman, Koch dan Steiner, 1992).
Matriconditioning dengan vermikulit sebelurn tanarn memberikan pengaruh
positif terhadap benih kacang tanah yang dimundurkan secara buatan dengan
perlakuan suhu 45OC dan kelembaban nisbi 75 % selama 6 hari (Jeng dan Sung,
1994). Perlakuan matricondifioning meningkatkan bobot kering dan panjang bibit
serta aktivitas beberapa enzim. Beberapa enzim yang meningkat aktivitasnya ada-
lah katalase, peroksidase, isositrat liase dan malat sintetase.
Proses invigorasi juga dapat terjadi pada benih kedelai yang mengalami
kemunduran buatan. Benih yang telah diusangkan dengan perlakuan suhu 41°c
dan kelembaban nisbi 100 O h selama 48 jam, memiliki DB 52 %. Lot benih
tersebut dapat mengalami proses invigorasi setelah perlakuan osmotic priming, yang
invigorasi pada lot tersebut juga ditunjukkan oleh tolok ukur &T dan persentase
pemunculan bibit.
Aktivitas malat dehidrogenase hanya sedikit dipengaruhi oleh perlakuan
osmoticpriming pada benih kedelai yang tidak dimundurkan. Pada benih yang di-
mundurkan selama 48 jam terlihat bahwa osmotic priming menurunkan intensitas
pewarnaan pada enzim malat dehidrogenase. Analisis aktivitas isoenzim glutamat
dehidrogenase (GDH) pada benih baik menunjukkan osmotic priming tidak
menyebabkan peningkatan intensitas pewarnaan. Namun osmotic priming dapat
meningkatkan intensitas pewarnaan isoenzim GDH pada benih yang diusangkan
selama 48 jam. Sedangkan isoenzim esterase merupakan isoenzim yang paling
sensitif terhadap perlakuan pengusangan (Shatters et al., 1994).
Hydropriming adalah suatu perlakuan dengan merendam benih dalarn air
selama 5 jam diikuti dengan inkubasi dalam wadah tertutup dengan kelembaban
nisbi 100% pada suhu 2 3 O ~ selama 3 hari. Perlakuan tersebut dapat menginvigorasi
benih yang ditunjukkan oleh tolok ukur kecepatan berkecambah benih caulzj7ower
khususnya pada lingkungan suhu rendah (IoOC). Perlakuan hydropriming hanya
efektif pada benih yang tidak diusangkan. Pada benih yang diusangkan secara fisik
perlakuan osmotic priming dengan PEG 6000 menunjukkan invigorasi dilihat dari
peningkatan persentase bibit normal yang lebih tinggi dibanding perlakuan
hydropriming (Fujikura et al., 1993 )
Pada benih kedelai yang diberi perlakukan hydropriming dengan air
menunjukkan kebocoran membran yang lebih tinggi dibandingkan benih yang diberi
perlakuan dengan PEG 6000. Perlakuan dengan PEG 6000 akan mengatur laju
penyerapan air, sehingga memungkinkan pemantapan integritas membran
Hasil penelitian Smith dan Cobb (1992) pada benih cabai yang diberi per-
lakuan osmotic priming dengan 300 mM NaCl menunjukkan adanya pengaruh
terhadap metabolisme benih. Kandungan protein terlarut meningkat 64 % pada
benih yang diberi perlakuan osmotic priming dibanding kontrol. Demikian pula
aktivitas enzim glukosa-6-fosfat dehidrogenase meningkat 50 O h , sedangkan enzim
6-fosfoglukonat dehidrogenase tidak menunjukkan perbedaan antara benih yang
diberi perlakuan osmotic priming dengan kontrol.
Osmofic priming secara urnum dapat memperbaiki kualitas benih, karena
mempercepat perkecambahan dan memperbaiki keseragaman pertumbuhan bibit.
Pengaruh yang menguntungkan pada osmotic priming berkaitan dengan perubahan
fisiologi yang terjadi pada embrio. Perbaikan penampakan bibit sesudah osmotic
priming dapat diterangkan dengan mekanisme perbaikan DNA secara lengkap
selama periode osmoticpriming (Osborne dalam Lanteri et al., 1994 )
Hasil penelitian Lanteri ef al. (1994) menunjukkan adanya korelasi
positif induksi sintesis DNA dengan menurunnya waktu rata-rata untuk
berkecambah. Pada benih cabai yang telah diberi perlakuan osmotic priming
reinduksi sintesis D N A 12 jam lebih awal dari pada benih yang tidak diberi
perlakuan. Perlakuan osmofic priming dengan -1.1 MPa dengan PEG 6000 baik
pada cabai maupun tomat menunjukkan perlakuan yang paling efektif untuk
meningkatkan sintesis DNA.
Konsumsi Energi Benih
Status energi benih selama pembentukan, penyimpanan dan perkecambahan
sangat penting dalam kaitannya dengan ekspresi vigor benih (Ching, 1982), makin
benih seperti endosperma pada monokotil, kotiledon pada dikotil dan gametopit
pada Girnnospermae, status energinya berubah secara bertahap karena imbibisi,
stratifikasi dan aktivitas biosintesis.
Efisiensi fosforilasi pada mitokondria benih kedelai bervigor tinggi lebih
efisien dibandingkan benih bewigor rendah karena proses deteriorasi (Abu-Shakra
dan Ching, 1967). FosforiIasi yang sangat efisien juga te jadi pada mitokondria yang
diisolasi dari jagung dan gandurn hibrida (Ching, 1982). Fosforilasi yang efisien
merupakan gambaran status energi dan berkorelasi positif dengan pertumbuhan.
Tingkat konsurnsi energi selama perkecambahan dapat ditunjukkan oleh pe-
manfaatan cadangan asam fitat di d d a m benih (Widajati, 1992). Benih kedelai yang
bervigor tinggi mampu memanfaatkan asarn fitat lebih baik dibandingkan dengan
benih bervigor rendah. Kemampuan tersebut diduga berkaitan erat dengan ke-
tersediaan energi untuk sintesis senyawa baru pada embrio, yang tercermin dari
bobot kering kecambah yang lebih tinggi pada benih bewigor tinggi.
Ketersediaan glukosa pada benih yang sedang berkecambah meng-
gambarkan status energi benih tersebut. Hal ini ditunjukkan adanya korelasi positif
antara kandungan glukosa dengan bobot kering kecambah dan tingkat viabilitas pada
benih kedelai (Widajati, 1992). Jumlah glukosa yang tinggi berkaitan dengan
aktivitas enzim amilase yang tinggi selama proses perkecambahan.
Blending
Lot benih pada hakekatnya merupakan campuran individu-individu benih
baik, sedang dan jelek, oleh karena itu informasi viabilitas mengikuti kurva distribusi
normal. Dalam aspek praktisnya kita dapat mencampur antar dua atau lebih lot
(1970) mendefinisikan bahwa blending adalah mencarnpur dua atau lebih lot benih
sehingga dihasilkan lot benih yang lebih homogen.
Lot benih dengan DB yang sudah sedikit menurun di bawah standar
kelulusan dapat diblending dengan lot benih dengan DB tinggi, sehingga dihasilkan
lot yang perkecambahannya berada dalam standar kelulusan. Blending dapat
dilakukan juga pada lot-lot dengan perbedaan campuran varietas lain, konsentrasi biji
gulma rnaupun kotoran fisik, sehingga diperoleh lot campuran yang lebih seragam.
Blending yang benar sulit dilaksanakan dan memerlukan ketelitian pada alatnya
sehingga akan diperoleh lot campuran yang benar-benar seragam. ' Alat blending
dapat dilihat pada Gambar 4.
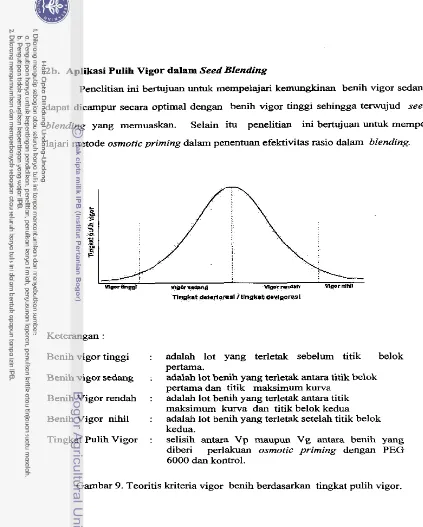
Fenomena pulih vigor diduga dapat dimanfaatkan untuk indikator dalam penen-
tuan suatu lot benih dapat digunakan untuk tujuan blending. Lot benih yang sudah
menurun viabilitasnya dan setelah uji invigorasi termasuk kelompok benih vigor
sedang dapat digunakan untuk blending. Blended seed mempakan campuran benih
vigor tinggi dan sedang dengan berbagai variasi rasio. Batas rasio yang dapat
diterima adalah rasio-rasio yang merniliki delta relatif konstan apabila campuran lot
Gambar 4. Diagram pergerakan benih dalam alat blender conveyor spiral vertikal (Greg ef al., 1970)
111. BAHAN DAN METODE
Penelitian dilaksanakan di Laboratorium Ilmu dan Teknologi Benih, Jurusan
Budi Daya Pertanian, Fakultas Pertanian, Laboratorium Biologi Molekuler dan
Seluler Tanaman, Laboratorium Mikrobiologi clan Biokimia, PAU Bioteknologi
IPB. Pelaksanaan penelitian dimulai bulan Januari 1997 sampai Oktober 1998.
Penelitian ini terdiri atas dua seri penelitian (Gambar 6).
Penelitian I. Studi Kriteria Analisis Vigor Benih dengan Mekanisme Pulih Vigor.
Penelitian ini bertujuan untuk meneliti mekanisme pemulihan vigor untuk
kriteria analisis vigor benih.
1 a. Mekanisrne Pulih Vigor.
Benih kedelai varietas Wilis yang digunakan merupakan has11 panen tanggal
20 Januari 1997 dari Laboratorium Ilmu dan Teknologi Benih Leuwikopo,
Darmaga. Lot-lot benih dengan ragam viabilitas dibuat secara alami dan buatan.
Pembuatan lot secara alami dilakukan dengan menyimpan benih dalam kondisi
terbuka dengan taraf penyimpanan 0, 1, 2
,...,
9 minggu (MO sampai dengan M9).Pembuatan lot dengan cara buatan dilakukan melalui proses devigorasi dengan pe-
ngusangan cepat dalam deraan uap etanol dengan MPC IPB 77-1 pada taraf TO, TI,
T2, T3, T4, T5, T6, T7, T8, dimana T O benih tanpa penderaan, T1 = (10 + 10) menit,
yang artinya dalam hembusan etanol selama 10 menit dan dibiarkan dalam etanol
jenuh selama 10 menit. T2 = 2 x TI,
...
T8 = 8 x T I . Benih dilembabkanselama 6 jam sebelum diberi perlakuan etanol dan dikeringkan ke keadaan semula
Penelitian I
Variasi lot benih oleh deteriorasi
Lot benih dengan pulih vigor Variasi lot benih terlinggi oleh devigorasi
Mekanisrne pulih vigor dengan parameter
biok
Penelitian I 1 Pernanfaatan invigorasi untuk kriteria analisis benih vigor tinggi dan vigor sedang
I
Blended seed lot berbagai variasi rasio
,
lnvigorasif
Oeteksi viabilitas blended seed lot
U = rataan m u m
Ai = pengaruh faktor tingkat viabilitas
Bj = pengaruh faktor invigorasi
(AB)ij = pengaruh interaksi faktor A ke-i dengan faktor B ke-j
Eij k = pengaruh acak
Vigor benih dengan indikasi fisiologi (VgfiSiO')
Parameter Viabilitas Potensial (Vp) diamati dengan tolok ukur DB.
Parameter vigor (Vg) diamati dengan tolok ukur KST.
Vigor benih dengan indikasi biokimiawi
(vgbiok)
Indikasi biokimiawi diamati dengan tolok ukur kandungan asam fitat, jurnlah P
yang diesterifikasi oleh mitokondria dan aktivitas enzim peroksidase.
l b . Hubungan antara V,"~'O' dan
vgbiok
Berdasarkan data penelitian l a dilakukan analisis regresi linier
Y =
a


![STUDI PERLAKUAN HIDRASI-DEHIDRASI PADA DUA LOT BENIH KEDELAI (Glycine max [L.] Merr.) VARIETAS ANJASMORO YANG MENGALAMI DETERIORASI](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)