PENDUGAAN PARAMETER GENETIK
KETAHANAN TANAMAN CABAI
TERHADAP ANTRAKNOSA (Colletotrichum acutatum)
MENGGUNAKAN ANALISIS DIALEL
YULIA IRAWATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Pendugaan Parameter Genetik Ketahanan Tanaman Cabai terhadap Antraknosa (Colletotrichum acutatum) Menggunakan Analisis Dialel adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian tesis ini.
Bogor, Juni 2011
Yulia Irawati
ABSTRACT
YULIA IRAWATI. Genetic Parameters Estimation of Pepper (Capsicum
annuum L.) Resistance to Anthracnose (Colletotrichum acutatum) Using
Diallel Analysis. Supervised by SRIANI SUJIPRIHATI and WIDODO.
Anthracnose (Colletotrichum acutatum) is considered of the major disease that may cause significant yield losses in pepper. Resistance variety is important for controlling disease infection since other methods were not effective. The aim of this research is to identify pepper genotype resistance and estimating genetic parameters of pepper resistance to anthracnose. This experiment were done in three steps: (1) screening of pepper resistance to anthracnose, (2) estimating genetic parameters of pepper resistance to anthracnose using diallel analysis and (3) determining the selection character using path analysis. The first experiment was conducted using 21 genotypes and C. acutatum isolate BKT 04. Mature green fruit were inoculated using microinjection method. The result showed that based on disease incidence, there was no genotype which resistance to anthracnose. However, there were three genotypes (IPB C15 x IPB C10, IPB C15 and IPB C8) which resistance based on lesion diameter. Set population from half diallel crosses which involve 6 parents and 15 crosses with C. acutatum isolate BKT 04 were used in the second experiment. Disease resistance percentage and lesion diameter were used for estimating genetic parameters and combining ability. Disease incidence and lesion diameter were used for estimating heterosis. The results show that there is no non alellic interaction, gene effects for resistance to anthracnose were additive and dominance, dominance effect larger than additive effect, the degree of dominance was overdominance, genes distribution in the parents was assymetrical, anthracnose resistance was controlled by one gene group, broad-sense heritability was high and narrow-sense heritability was low. General combining ability of IPB C15 was high for disease resistance percentage and lesion diameter, specific combining ability of IPB C15 x IPB C9 was high for disease resistance percentage and specific combining ability of IPB C15 x IPB C10 and IPB C4 x IPB C2 were high for lesion diameter. Negative heterosis was expected in disease resistance traits, IPB C15 x IPB C9 had negative heterosis for disease incidence and IPB C10 x IPB C9 had negative heterosis for lesion diameter. The thickness of fruit flesh is recomended for selection criteria for developing anthracnose resistant pepper varieties.
RINGKASAN
YULIA IRAWATI. Pendugaan Parameter Genetik Ketahanan Cabai
terhadap Antraknosa (Colletotrichum acutatum) Menggunakan Analisis
Dialel. Di bawah bimbingan SRIANI SUJIPRIHATI dan WIDODO.
Penyakit antraknosa merupakan salah satu dari penyakit utama pada cabai. Penggunaan kultivar resisten merupakan cara yang terbaik untuk mengendalikan penyakit ini. Perakitan varietas tahan akan lebih mudah jika tersedia informasi kendali genetik karakter yang diinginkan. Perilaku genetik gen-gen karakter yang diinginkan dapat dipelajari melalui studi genetik menggunakan metode analisis dialel. Pendugaan parameter genetik ketahanan cabai terhadap antraknosa terdiri dari tiga percobaan yaitu: (1) pengujian ketahanan cabai terhadap antraknosa, (2) pendugaan parameter genetik ketahanan cabai terhadap antraknosa menggunakan analisis dialel, (3) penentuan karakter seleksi menggunakan analisis lintasan.
Percobaan pertama menggunakan 21 genotipe dan isolat C. acutatum BKT 04. Buah hijau yang sudah matang diinokulasi menggunakan metode suntik dengan mikroinjeksi. Peubah yang diamati adalah kejadian penyakit dan diameter bercak. Hasil pengujian ketahanan cabai terhadap antraknosa menunjukkan tidak terdapat genotipe yang tahan pada peubah kejadian penyakit. Pada peubah diameter bercak terdapat tiga genotipe yang tahan, yaitu IPB C15 x IPB C10, IPB C15 dan IPB C8.
Percobaan kedua menggunakan enam genotipe tetua dan 15 genotipe F1 yang disusun dalam populasi half diallel. Peubah ketahanan (1-kp) dan diameter bercak digunakan untuk menduga nilai parameter genetik dan daya gabung, sementara peubah kejadian penyakit dan diameter bercak digunakan untuk menduga nilai heterosis. Hasil percobaan menunjukkan bahwa ketahanan terhadap penyakit antraknosa yang disebabkan oleh C. acutatum isolat BKT 04 dikendalikan oleh satu kelompok gen, gen pengendali ketahanan adalah dominan, derajat dominansi dikategorikan sebagai overdominan. Aksi gen pengendali ketahanan adalah aditif dan dominan, ragam dominan lebih besar dibandingkan dengan ragam aditif. Nilai heritabilitas arti luas tergolong tinggi dan nilai heritabilitas arti sempit tergolong rendah. Genotipe IPB C15 merupakan genotipe cabai yang memiliki daya gabung umum tinggi untuk ketahanan dan diameter bercak. Sementara untuk pendugaan daya gabung khusus, persilangan IPB C15 x IPB C9 merupakan kombinasi persilangan dengan nilai DGK tinggi pada peubah ketahanan. Persilangan IPB C15 x IPB C10 dan IPB C4 x IPB C2 memiliki nilai DGK terbaik pada peubah diameter bercak. Untuk pendugaan nilai heterosis, genotipe IPB C15 x IPB C9 merupakan genotipe yang memiliki nilai heterosis negatif dan genotipe IPB C10 x IPB C9 merupakan genotipe dengan nilai heterobeltiosis negatif pada peubah kejadian penyakit. Sementara pada peubah diameter bercak, genotipe IPB C10 x IPB C9 merupakan genotipe dengan nilai heterosis negatif dan genotipe IPB C15 x IPB C10 merupakan genotipe dengan nilai heterobeltiosis negatif.
dalam percobaan ini. Hasil analisis lintasan menunjukkan bahwa peubah tebal kulit buah dapat direkomendasikan sebagai karakter kriteria seleksi ketahanan cabai terhadap antraknosa.
© Hak Cipta milik IPB, tahun 2011 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
PENDUGAAN PARAMETER GENETIK
KETAHANAN TANAMAN CABAI
TERHADAP ANTRAKNOSA (Colletotrichum acutatum)
MENGGUNAKAN ANALISIS DIALEL
YULIA IRAWATI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PRAKATA
Puji syukur ke hadirat Allah SWT atas berkat rahmat-Nya akhirnya penulis dapat menyelesaikan penelitian dan penulisan tesis yang berjudul “Pendugaan
Parameter Genetik Ketahanan Tanaman Cabai terhadap Antraknosa
(Colletotrichum acutatum) Menggunakan Analisis Dialel”.
Penelitian dan penulisan tesis dilakukan di bawah bimbingan Prof. Dr. Ir. Sriani Sujiprihati, MS sebagai Ketua Komisi Pembimbing dan Dr. Ir. Widodo, MS sebagai Anggota Komisi Pembimbing. Penelitian ini didanai oleh Hibah Fundamental tahun 2006-2007.
Selama melakukan penelitian dan penulisan tesis, penulis mendapatkan bantuan dan dukungan dari berbagai pihak, karena itu penulis ingin menyampaikan penghargaan yang setinggi-tingginya dan ucapan terima kasih kepada:
1. Kedua orang tua tercinta, Iwan Sarwono dan Sri Hartini, serta adik tersayang, Febryadi Bondan Dwipayana, yang dengan penuh kesabaran senantiasa memberikan kasih sayang, kekuatan dan semangat kepada penulis sehingga dapat menyelesaikan penelitian dan penulisan tesis ini.
2. Prof. Dr. Ir. Sriani Sujiprihati, MS sebagai Ketua Komisi Pembimbing dan Dr. Ir. Widodo, MS sebagai Anggota Komisi Pembimbing atas motivasi, bimbingan dan arahan yang telah diberikan kepada penulis.
3. Dr. Rahmi Yunianti, SP. MSi sebagai penguji luar komisi yang telah memberikan bantuan dan masukan yang sangat berharga baik dalam penyelesaian penelitian maupun dalam penulisan tesis.
4. Dr. Muhamad Syukur, SP. MSi atas bantuan dan masukan selama melakukan penelitian.
5. Ketua Program Studi Pemuliaan dan Bioteknologi Tanaman, Ketua Program Studi Agronomi, beserta seluruh staf pengajar Departemen Agronomi dan Hortikultura.
7. Pak Gusti, Nita dan Arif yang sangat membantu dalam pengolahan data dan penyelesaian penulisan tesis.
8. Kepala Balai, Ketua dan Anggota Kelti Pemuliaan, Kepala Bagian Kepegawaian beserta staf, Kepala Bagian Pelayanan Teknis beserta staf, Kepala Bagian Jasa Penelitian beserta staf dan seluruh pegawai Balai Penelitian Tanaman Buah Tropika atas bantuan yang telah diberikan sehingga penulis dapat menyelesaikan studi di SPS IPB.
9. Kepala Balai, Kepala Bagian Kepegawaian beserta staf, Ketua dan Anggota Kelti Pemuliaan Balai Penelitian Tanaman Hias, terutama Ibu Susi dan Teh Dedeh atas bantuan yang diberikan selama penulis melakukan detasir.
10. Teman-teman Program Studi Agronomi bidang Pemuliaan Tanaman Angkatan 2003: Millah, Mba Ismi, Mba Niken, Uni Reni, Ade, Nila, Imay dan Apri atas bantuan, kebersamaan dan dorongan semangat kepada penulis. 11. Teman-teman Ponytail, Pondok Putri dan Graha Matudilipa, Uni Neni, Uni
Susi, Dian, Nana, Mba Wiwi, Cici dan Uni Ida atas kebersamaan, bantuan dan motivasi yang telah diberikan kepada penulis.
12. Nazly dan Akhyar atas bantuan selama melakukan penelitian dan penulisan tesis serta motivasi yang telah diberikan kepada penulis.
13. Fitri, Riry dan Liza atas kebersamaan selama di Solok serta bantuan dan dorongan semangat sehingga penulis dapat menyelesaikan studi di IPB. 14. Staf pegawai Departemen Agronomi dan Hortikultura dan Sekolah
Pascasarjana IPB atas bantuan dan kerjasamanya. Semoga informasi dari penelitian ini dapat bermanfaat.
Bogor, Juni 2011
RIWAYAT HIDUP
Penulis dilahirkan di Bandung, Jawa Barat pada tanggal 31 Desember 1977. Penulis merupakan anak pertama dari Bapak Iwan Sarwono dan Ibu Sri Hartini.
Penulis lulus dari SDN Kranji I Bekasi pada tahun 1990 dan pada tahun 1993 penulis menyelesaikan pendidikan di SMP Negeri 2 Bandung. Penulis melanjutkan pendidikan di SMA Negeri 5 Bandung dan lulus pada tahun 1996, di tahun yang sama penulis melanjutkan pendidikan di Program Studi Pemuliaan Tanaman Fakultas Pertanian Universitas Padjadjaran. Pada tahun 2003 penulis melanjutkan pendidikan pada Program Studi Agronomi bidang Pemuliaan Tanaman Program Pascasarjana IPB.
DAFTAR ISI
Halaman
DAFTAR TABEL ... xi
DAFTAR GAMBAR ... xii
DAFTAR LAMPIRAN ... xiii
1 PENDAHULUAN Latar Belakang ... 1
Tujuan Penelitian ... 3
Hipotesis ... 3
2 TINJAUAN PUSTAKA Klasifikasi, Botani dan Syarat Tumbuh Cabai ... 4
Penyakit Antraknosa ... 6
Analisis Dialel ... 8
Heterosis ... 10
3 BAHAN DAN METODE Pengujian Ketahanan Cabai Terhadap Antraknosa ... 13
Pendugaan Parameter Genetik Ketahanan Cabai Terhadap Antraknosa Menggunakan Analisis Dialel Pendugaan Parameter Genetik Menggunakan Pendekatan Hayman ... 16
Pendugaan Daya Gabung ... 20
Pendugaan Heterosis ... 21
Penentuan Karakter Seleksi Menggunakan Analisis Lintasan ... 21
4 HASIL DAN PEMBAHASAN Pengujian Ketahanan Cabai Terhadap Antraknosa ... 24
Pendugaan Parameter Genetik Ketahanan Cabai Terhadap Antraknosa Dengan Menggunakan Analisis Dialel Pendugaan Parameter Genetik menggunakan Pendekatan Hayman ... 28
Pendugaan Daya Gabung ... 34
Pendugaan Heterosis ... 37
Penentuan Karakter Seleksi Menggunakan Analisis Lintasan ... 39
5 KESIMPULAN DAN SARAN ... 45
DAFTAR PUSTAKA ... 47
DAFTAR TABEL
Halaman 1 Kelas ketahanan berdasarkan kejadian penyakit ... 15
2 Kelas ketahanan berdasarkan diameter bercak ... 15
3 Populasi half dialel dalam menduga parameter genetik untuk ketahanan cabai terhadap antraknosa ... 16
4 Sidik ragam untuk analisis daya gabung metode 2 ... 20
5 Respon 21 genotipe cabai terhadap antraknosa (kejadian penyakit) .... 24
6 Respon 21 genotipe cabai terhadap antraknosa (diameter bercak) ... 26
7 Kuadrat tengah ketahanan cabai terhadap antraknosa ... 29
8 Pendugaan parameter genetik ketahanan tanaman cabai terhadap antraknosa ... 29
9 Sebaran Vr + Wr ... 31
10 Kuadrat tengah daya gabung ketahanan cabai terhadap antraknosa .... 34
11 Nilai DGU ketahanan cabai terhadap antraknosa ... 35
12 Nilai DGK ketahanan cabai terhadap antraknosa ... 35
13 Nilai duga heterosis dan heterobeltiosis ketahanan cabai terhadap antraknosa ... 38
14 Nilai koefisien korelasi antar karakter ... 43
DAFTAR GAMBAR
Halaman 1 Koloni C. acutatum berwarna oranye hasil inokulasi ... 28
2 Buah cabai hasil inokulasi ... 28
3 Hubungan antara peragam (Wr) dan ragam (Vr) untuk ketahanan (1 - kp) ... 32
4 Hubungan antar peragam (Wr) dan ragam (Vr) untuk diameter bercak 33
DAFTAR LAMPIRAN
Halaman 1 Genotipe-genotipe yang digunakan dalam pengujian ketahanan
terhadap antraknosa ... 52
2 Deskripsi genotipe tetua IPB C2 ... 53
3 Deskripsi genotipe tetua IPB C4 ... 54
4 Deskripsi genotipe tetua IPB C8 ... 55
5 Deskripsi genotipe tetua IPB C9 ... 56
6 Deskripsi genotipe tetua IPB C10 ... 57
7 Deskripsi genotipe tetua IPB C15 ... 58
8 Nilai tengah genotipe-genotipe yang digunakan dalam analisis lintasan 59 9 Sidik ragam analisis dialel metode Hayman peubah ketahanan ... 60
10 Sidik ragam umum analisis dialel metode Griffing peubah ketahanan ... 60
11 Sidik ragam daya gabung peubah ketahanan ... 60
12 Sidik ragam analisis dialel metode Hayman peubah diameter bercak ... 61
13 Sidik ragam umum analisis dialel metode Griffing peubah diameter bercak ... 61
14 Sidik ragam daya gabung peubah diameter bercak ... 61
PENDAHULUAN
Latar Belakang
Cabai (Capsicum annuum L.) adalah komoditas sayuran yang bernilai ekonomi tinggi sehingga banyak diusahakan petani. Buah cabai terutama digunakan sebagai penambah rasa dan warna dalam masakan. Buah cabai segar mengandung provitamin A (caroten), vitamin B, C dan E (Purseglove et al. 1981). Buah cabai juga digunakan dalam industri farmasi karena mengandung senyawa
capsaicin.
Produktivitas cabai merah di Indonesia masih jauh dari potensi produksinya yang mencapai 12 ton per hektar (Duriat 1996). Berdasarkan data BPS (2011) produksi cabai nasional pada tahun 2010 adalah 6.57 ton/ha. Rendahnya produktivitas cabai disebabkan oleh beberapa faktor, diantaranya dikarenakan oleh serangan penyakit. Widodo (2007) dalam studi kasus yang dilakukan pada periode 1994-1997 di Lampung dan Jawa Barat melaporkan bahwa serangan hama dan penyakit menempati urutan pertama sebesar 61.4 % dibandingkan dengan masalah lain seperti penurunan kesuburan tanah, masalah pengairan dan harga pasar.
Penyakit antraknosa merupakan salah satu dari penyakit utama pada cabai. Antraknosa berada pada kisaran urutan kedua dan ketiga diantara 10 hama dan penyakit utama pada cabai pada tahun 2001-2006 (Widodo 2007). Penyakit antraknosa menyerang daun, batang dan buah dengan gejala timbulnya titik berwarna gelap atau luka yang menjorok ke dalam. Pada buah penyakit ini seringkali bersifat laten dan biasanya mengakibatkan buah busuk dan gugur (Agrios 2005). Perkiraan penurunan hasil akibat serangan antraknosa di pulau Jawa berkisar antara 40-60 % (Hartman dan Wang 1992).
mengurangi resiko pencemaran lingkungan serta gangguan kesehatan terhadap manusia akibat pemakaian fungisida.
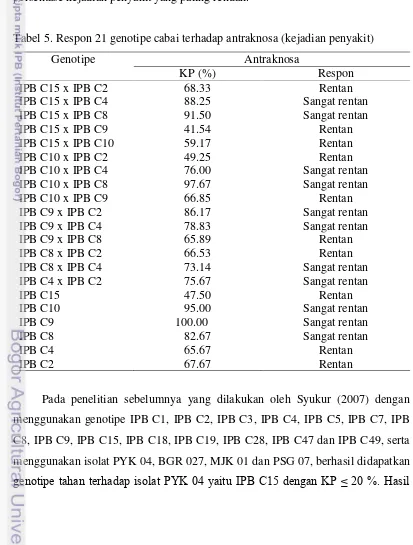
Penelitian untuk mendapatkan varietas cabai tahan antraknosa telah banyak dilakukan. Syukur (2007) telah berhasil mendapatkan genotipe tahan antraknosa yaitu genotipe IPB C15 yang tahan terhadap isolat PYK 04 dan moderat terhadap isolat BGR 027, MJK 01 dan PSG 07. Penelitian tersebut menunjukkan bahwa genotipe IPB C15 memberikan reaksi ketahanan yang berbeda ketika diinokulasi dengan isolat yang berbeda, oleh karena itu dalam penelitian ini digunakan isolat BKT yang berasal dari Bukittinggi yang tidak digunakan dalam penelitian sebelumnya untuk mengetahui reaksi ketahanan dari genotipe-genotipe yang telah diuji sebelumnya.
Perakitan varietas tahan akan lebih mudah jika tersedia informasi kendali genetik karakter yang diinginkan. Perilaku genetik gen-gen karakter yang diinginkan dapat dipelajari melalui studi genetik menggunakan metode analisis silang dialel. Selain untuk mempelajari kendali genetik suatu sifat, analisis silang dialel juga digunakan untuk menduga nilai daya gabung dalam hibrida serta membantu pemulia dalam meningkatkan dan menyeleksi populasi segregan (de Sousa dan Maluf 2003).
Fenomena heterosis terjadi ketika nilai rata-rata F1 lebih baik daripada nilai rata-rata tetuanya. Hibrida F1 dikembangkan berdasarkan adanya fenomena heterosis tersebut, karena itu informasi mengenai heterosis penting dalam pembentukan hibrida varietas tahan antraknosa.
Genotipe-genotipe yang digunakan dalam penelitian ini adalah genotipe-genotipe yang mempunyai karakter agronomi yang baik dan juga genotipe-genotipe yang mempunyai ketahanan terhadap penyakit tertentu yang telah diuji dalam beberapa penelitian sebelumnya. Diharapkan melalui penelitian ini bisa didapatkan genotipe tahan antraknosa yang mempunyai karakter agronomi yang baik.
Tujuan Penelitian
Tujuan dari penelitian ini adalah:
1. Mengidentifikasi tingkat ketahanan cabai terhadap antraknosa.
2. Menduga parameter genetik dan daya gabung dari genotipe-genotipe yang diuji.
3. Memperoleh informasi fenomena heterosis pada karakter ketahanan.
4. Menentukan karakter seleksi untuk perakitan varietas cabai tahan antraknosa.
Hipotesis
1. Terdapat genotipe cabai tahan antraknosa.
2. Terdapat tetua yang memiliki daya gabung umum yang baik dan terdapat pasangan persilangan yang memiliki daya gabung khusus yang baik pada karakter ketahanan cabai terhadap antraknosa.
3. Terdapat fenomena heterosis pada karakter ketahanan.
TINJAUAN PUSTAKA
Klasifikasi, Botani dan Syarat Tumbuh Cabai
Genus Capsicum termasuk dalam kingdom Plantae, divisio Magnoliophyta, kelas Magnoliopsida, ordo Rosales, dan famili Solanaceae (Bosland dan Votava 2000). Para ahli taksonomi modern mengidentifikasi lima spesies utama yang telah dibudidayakan, yaitu C. annuum L., C. frutescens L., C. chinense Jacquin, C. pendulum Wildenow,dan C. pubescens Ruiz & Pavon. Lima spesies utama tersebut dapat dibedakan dari kombinasi karakter bunga dan buah. C. annuum
berbunga putih dengan anter biru sampai ungu, mempunyai calyx yang berlekuk dan berbunga serta berbuah tunggal setiap buku. C. frutescens berbunga agak kehijauan dengan anter biru, calyx yang tidak berlekuk dan pada umumnya berbuah tunggal setiap buku serta beberapa bunga majemuk pada bukunya. C. chinense berbunga putih atau putih kehijauan dengan anter biru, calyx berlekuk dan pada umumnya berbuah 1-3 setiap buku. C. pendulum berbunga putih dengan bintik kuning pada dasarnya, anter kuning, tangkai bunga panjang, dan berbunga tunggal. C. pubescens berbunga besar keunguan, buah oranye, dan biji berwarna gelap (Greenleaf 1986). Tanaman yang berasal dari Amerika Selatan ini semua spesies budidayanya merupakan diploid (2n = 2x = 24) (Purseglove et al. 1981).
Cabai merupakan tanaman menyerbuk sendiri, walaupun demikian penyerbukan silang dapat terjadi pada beberapa genotipe dan pada lingkungan tertentu. Bunga cabai merupakan bunga sempurna dimana alat reproduksi jantan dan betina terdapat dalam satu bunga. Helaian mahkota bunga cabai merah berjumlah lima atau enam. Pada dasar bunga terdapat daun buah berjumlah lima yang kadang-kadang berlekuk. Setiap bunga memiliki satu putik, kepala putik berbentuk bulat. Terdapat lima sampai delapan helai benang sari dengan kepala sari yang berbentuk lonjong berwarna biru keunguan. Tepung sari berbentuk lonjong terdiri dari tiga segmen berwarna kuning mengkilat. Dalam satu kotak sari terdapat sekitar 11.000-18.000 tepung sari (Kusandriani 1996)
Cabai dapat dibudidayakan di dataran rendah maupun dataran tinggi. Agar mampu tumbuh dengan optimal cabai merah memerlukan kisaran suhu antara 18-27 oC. Suhu optimal untuk pertumbuhan dan pembungaan berkisar antara 21-27 oC dan untuk pembuahan antara 15.5-21 oC (Sumarni 1996). Bunga tidak terbuahi pada suhu di bawah 16 oC atau di atas 32 oC karena produksi tepung sari yang tidak viabel (Rubatzky dan Yamaguchi 1999).
Curah hujan yang baik untuk pertumbuhan tanaman cabai adalah sekitar 600-1250 mm per tahun (Sumarni 1996). Tanaman cabai merah dapat tumbuh pada berbagai jenis tanah asal drainase dan aerasi tanah cukup baik karena tanaman sangat peka terhadap genangan. Tanaman yang tergenang cenderung mengalami kerontokan daun dan terserang penyakit akar. Tanah harus mengandung cukup bahan organik, unsur hara, air, bebas dari gulma, dan patogen. Tingkat kemasaman (pH) tanah yang paling sesuai berkisar antara 6.5-7.0 (Rubatzky dan Yamaguchi 1999).
saat pembentukan bunga dan buah dapat menggugurkan bunga dan membusuknya buah. Penggunaan mulsa dapat meningkatkan hasil, baik pada musim kemarau maupun musim hujan. Peningkatan hasil ini terjadi karena mulsa dapat mempertahankan struktur tanah tetap gembur, memelihara kelembaban dan suhu tanah, mengurangi kehilangan unsur hara, dan menekan pertumbuhan gulma (Sumarni 1996).
Penyakit Antraknosa
Nama antraknosa diambil dari kata anthrax yang berarti karbon (Agrios 2005). Nama tersebut digunakan karena gejala yang ditimbulkan oleh penyakit ini menimbulkan bercak berwarna hitam seperti warna karbon.
Penyakit ini menimbulkan gejala pada seluruh bagian tanaman termasuk akar, batang, daun, bunga dan buah (Bailey et al. 1992). Pada umumnya gejala pertama muncul berupa bercak kecil basah berwarna kuning atau kehitaman yang membesar dengan cepat. Bercak kemudian berkembang sampai diameter 3-4 cm berwarna dari merah gelap sampai coklat muda dengan jaringan cendawan berwarna gelap yang terdiri dari kumpulan seta atau aservuli yang menghasilkan massa spora berwarna pucat atau salem yang tersebar atau berbentuk lingkaran konsentrik pada bercak (Hadden dan Black 1989). Gejala pada biji dapat menimbulkan kegagalan berkecambah, pada kecambah dapat menimbulkan rebah kecambah, dan pada tanaman dewasa dapat menimbulkan mati pucuk (Suryaningsih 1996).
terinfeksi tersebut digunakan untuk musim tanam berikutnya maka cendawan dapat menyebar ke area tanam baru tersebut.
Penyakit antraknosa berkembang dengan cepat dalam lingkungan yang hangat dengan kelembaban tinggi. Kelembaban relatif udara 95 % yaitu pada cuaca berkabut dan berembun dapat membantu inisiasi infeksi dan perkembangan penyakit selanjutnya (Suryaningsih 1996).
Spesies Colletotrichum acutatum dalam keadaan tahapan aseksual (anamorf) termasuk dalam kelompok Coelomycetes dan dalam keadaan tahapan seksual (teleomorf) termasuk dalam genus Glomerella dan dimasukan dalam kelompok Ascomycetes (Wharton dan Uribeondo 2004). Beberapa genus yang termasuk dalam kelompok Ascomycetes, seperti Glomerella, jarang menghasilkan spora seksual (askospora) dan lebih sering menghasilkan spora aseksual (konidia), karena itu genus Glomerella lebih dikenal dalam nama tahapan aseksualnya yaitu
Colletotrichum (Agrios 2005).
Koloni spesies C. acutatum pada awalnya berwarna putih kemudian berubah warna menjadi pink atau oranye. Konidia berbentuk elipsoid dan meruncing setidaknya pada salah satu ujungnya (Peres et al. 2005) dan berukuran 8.5-16.5 x 2.5-4 µm (Sutton 1992). Suhu optimum untuk spesies ini adalah 25ºC (Wharton dan Uribeondo 2004).
Tahap awal dari pertumbuhan cendawan dalam proses infeksi dari spesies ini pada dasarnya sama dengan spesies dari genus Colletotrichum lainnya, yaitu persentuhan konidia pada permukaan tanaman, penempelan konidia pada permukaan tanaman, perkecambahan konidia, produksi apresoria, penetrasi ke epidermis tanaman, pertumbuhan dan pembentukan koloni pada jaringan tanaman serta produksi aservuli dan pembentukan spora. Proses terjadinya tiap tahapan tersebut dalam proses infeksi bervariasi bergantung antara lain pada jaringan inang dan isolat cendawan tersebut (Wharton dan Uribeondo 2004).
tidak dapat bersaing atau berkembang pada sisa daun atau pada tanah jika ada organisme lain yang aktif (Peres et al. 2005).
Keberhasilan kolonisasi pada tanaman inang oleh suatu patogen bergantung pada kemampuan patogen tersebut untuk mengatasi sistem pertahanan tanaman tersebut. Ketahanan buah muda pada kolonisasi spesies C. acutatum kemungkinan berhubungan dengan salah satu dari empat mekanisme pertahanan yang dimiliki tanaman, yaitu pembentukan awal senyawa racun yang menghambat pertumbuhan patogen terjadi pada buah muda, buah muda tidak menyediakan substrat yang sesuai untuk memenuhi kebutuhan nutrisi dan energi bagi patogen, enzim patogen tidak sesuai untuk membentuk kolonisasi pada buah muda, dan produksi fitoaleksin pada buah muda (Wharton dan Uribeondo 2004).
Analisis Dialel
Persilangan dialel adalah persilangan dengan menggunakan semua kombinasi persilangan yang mungkin dilakukan diantara sekelompok tetua termasuk selfing. Tujuan dari persilangan dialel adalah untuk mengevaluasi dan memilih tetua berdasarkan turunan terbaik dan evaluasi turunan terbaik (Ganefianti 2010). Genotipe yang digunakan dalam persilangan dialel dapat berupa individu, klon, galur dan lain-lain (Hayman 1954).
Di dalam analisis silang dialel, pendugaan parameter genetik sudah dapat dilakukan pada F1, tanpa harus membentuk populasi F2, BCP1 dan BCP2. Analisis dialel dilakukan berdasarkan asumsi-asumsi segregasi diploid, tidak ada perbedaan diantara persilangan resiprok, tidak ada interaksi gen non alelik, tidak ada multiple allelism, tetua homozigot dan gen-gen menyebar bebas diantara tetua (Hayman 1954).
Ploidi tanaman cabai adalah diploid (Greenleaf 1986), dengan demikian segregasi gen-gen yang terjadi merupakan segregasi diploid. Gen-gen yang mengendalikan suatu karakter harus menyebar diantara tetua-tetua persilangan. Untuk memenuhi asumsi ini maka dipilih tetua yang mewakili tahan, moderat dan rentan.
dan daya gabung khusus yang didapatkan melalui metode Griffing (Singh dan Chaudary 1979). Parameter genetik yang diperoleh dari metode Hayman meliputi keragaman karena pengaruh lingkungan, keragaman karena pengaruh aditif, keragaman karena pengaruh efek dominan, pengaruh dominan, penduga sebaran gen dalam tetua untuk menduga proporsi gen negatif dan positif pada tetua, rata-rata tingkat dominansi, proporsi gen-gen dominan dan resesif pada tetua, jumlah kelompok gen yang mengendalikan sifat dan menimbulkan dominansi, heritabilitas arti luas dan heritabilitas arti sempit.
Daya gabung adalah kemampuan tetua untuk menghasilkan kombinasi persilangan yang unggul dalam satu atau seri persilangan. Evaluasi daya gabung ini bertujuan untuk memilih tetua-tetua atau genotipe yang akan dijadikan tetua dalam pembentukan kultivar hibrida. Ada dua macam daya gabung yaitu Daya Gabung Umum (DGU) dan Daya Gabung Khusus (DGK). DGU adalah rata-rata penampilan tetua dalam satu seri persilangan, sementara DGK adalah kemampuan bergabung dari tetua dalam persilangan tertentu (Chaudhary 1982). Menurut Falconer (1981) efek DGU dan DGK adalah indikator penting dari nilai potensial suatu galur murni dalam kombinasi hibrida. Welsh (1981) menyatakan bahwa kemampuan berkombinasi umum (DGU) merupakan hasil dari aksi gen aditif, sedangkan kemampuan berkombinasi spesifik (DGK) merupakan hasil dari gen dominan, epistasis, dan aditif.
Menurut Griffing (1956) diacu dalam Roy (2000) untuk melakukan analisis daya gabung diperlukan tiga set materi genetik yaitu tetua, F1 hasil silangan serta resiproknya. Griffing menggunakan ketiga set materi genetik tersebut untuk merumuskan empat metode dalam analisis daya gabung, yaitu metode 1 (full diallel) melibatkan tetua, F1 hasil persilangan serta resiproknya dengan analisis [n(n-1)/2]; metode 2 melibatkan tetua dan F1 hasil silangannya tanpa resiproknya dengan analisis [n(n+1)/2]; metode 3 melibatkan F1 hasil silangan serta resiproknya tanpa tetua dengan analisis n(n-1); serta metode 4 yang hanya melibatkan F1 hasil silangan saja dengan analisis (n-1)/2.
ketahanan terhadap CMV dan ChiVMV (Riyanto 2007), ketahanan terhadap Begomovirus (Ganefianti 2010), ketahanan terhadap antraknosa, hawar
Phytophthora dan layu bakteri (Putri 2010), ketahanan terhadap powdery mildew
(Nandadevi et al. 2003) dan ketahanan terhadap thrips dan mite (Jagadeesha et al. 2006).
Heterosis
Pada persilangan tanaman menyerbuk silang seperti jagung, penampilan F1
selalu lebih baik daripada kedua tetuanya. Fenomena ini dinamakan heterosis atau vigor hibrida. Heterosis adalah peningkatan ukuran atau tingkat pertumbuhan suatu keturunan dibandingkan dengan tetuanya (Duvick 1999). Pada persilangan dua galur murni tanaman menyerbuk sendiri seperti serealia, beberapa hasil persilangan memang menampilkan fenomena heterosis tetapi nilainya sangat rendah. Pada persilangan tanaman menyerbuk silang seperti jagung, heterosis bisa mencapai lebih dari 200 %, sedangkan pada tanaman menyerbuk sendiri seperti gandum, nilai heterosis hanya 10 % (Roy 2000).
Pada beberapa kasus fenomena heterosis tidak muncul sama sekali dan pada kasus yang lain penampilan F1 lebih rendah dibandingkan tetuanya. Dari
perbandingan nilai tengah F1 terhadap tetuanya, terdapat dua jenis heterosis, yaitu (1) heterosis, dimana nilai tengah F1 lebih besar bila dibandingkan dengan nilai tengah kedua tetuanya dan (2) heterobeltiosis, dimana nilai tengah F1 lebih besar bila dibandingkan dengan nilai tengah tetua terbaiknya.
Menurut Chaudhary (1984) heterosis dapat dibedakan menjadi dua tipe, yaitu:
(1) Euheterosis
dalam populasi. Euheterosis seimbang adalah heterosis dalam arti sebenarnya, yang muncul dari kombinasi gen yang seimbang antara kemampuan adaptasi dan karakter agronomi yang lebih baik. Heterosis ini yang digunakan dalam progam pemuliaan untuk mengembangkan varietas hibrida.
(2) Pseudo-heterosis
Pseudo-heterosis pada dasarnya adalah suatu fenomena dimana suatu hasil persilangan, secara kebetulan menampilkan ekspresi vigor dan pertumbuhan di atas rata-rata tetapi bersifat sementara dan tidak adaptif.
Menurut Roy (2000) terdapat tiga teori yang berkaitan dengan heterosis yaitu:
(1) Teori dominansi
Teori ini menyatakan bahwa F1 bersifat heterotik karena gen resesif yang
mengekspresikan sifat inferior dalam keadaan homosigot tertutupi oleh gen dominan dan jumlah lokus dimana gen resesif berada dalam keadaan homosigot berkurang pada F1 bila dibandingkan dengan tetuanya, sehingga F1 memperlihatkan penampilan yang lebih superior dari tetua terbaiknya. (2) Teori overdominansi
Teori ini menyatakan bahwa genotipe heterozigot lebih superior dibandingkan dengan genotipe homozigot, dimana genotipe heterosigot memperlihatkan perkembangan homeostatis yang lebih besar daripada genotipe homosigot yang memiliki fenotipe yang lebih rentan terhadap cekaman lingkungan.
(3) Teori epistasis
Teori ini menyatakan bahwa interaksi antar gen berperan dalam ekspresi heterosis, dimana heterosis merupakan ekpresi dari gen yang memperlihatkan pengaruh aditif dengan dominan lengkap.
dari kombinasi aktivitas zat-zat kimia dalam tanaman sebagai hasil dari aksi gen komplementer pada hibrida (Chaudhary 1984).
BAHAN DAN METODE
Penelitian terdiri dari tiga percobaan, yaitu 1) pengujian ketahanan cabai terhadap antraknosa; 2) pendugaan parameter genetik ketahanan cabai terhadap antraknosa dengan menggunakan analisis dialel; dan 3) penentuan karakter seleksi dengan menggunakan analisis lintasan. Penelitian dilaksanakan pada bulan Februari-Juli 2007. Penanaman dilakukan di rumah plastik Kebun Percobaan IPB Tajur Bogor yang terletak pada ketinggian ± 250 m di atas permukaan laut. Perbanyakan dan pemeliharaan C. acutatum dilakukan di Laboratorium Klinik Tanaman Departemen Proteksi Tanaman Fakultas Pertanian IPB, dan pengujian ketahanan dilakukan di Laboratorium Pendidikan Pemuliaan Tanaman Departemen Agronomi dan Hortikultura Fakultas Pertanian IPB.
Pengujian Ketahanan Cabai terhadap Antraknosa
Pengujian ketahanan cabai terhadap antraknosa ini menggunakan bahan tanaman koleksi Bagian Genetika dan Pemuliaan Tanaman AGH IPB. Genotipe-genotipe yang digunakan dapat dilihat pada Lampiran 1 dan deskripsi tetua disajikan pada Lampiran 2-7. Isolat C. acutatum yang digunakan adalah BKT 04 yang merupakan koleksi Dr. Widodo dari Laboratorium Mikologi Tumbuhan Departemen Proteksi Tanaman IPB.
Benih cabai diberi perlakuan perendaman dalam air hangat (50 oC) selama satu malam sebelum disemai. Benih disemai dalam tray yang berisi media semai yang sudah disterilisasi dengan pemanasan pada suhu 150 oC selama tiga jam. Pemeliharaan yang dilakukan pada saat persemaian adalah penyiraman dan pemberian pupuk daun dengan dosis 2 g/l. Setelah empat minggu kemudian dipindahkan ke dalam polibag berdiameter 35 cm berisi 8 kg media tanam steril. Media tanam yang digunakan adalah campuran tanah dan pupuk kandang dengan perbandingan 1:1. Sterilisasi media tanam dilakukan dengan pemanasan media tanam di atas api selama delapan jam. Pemeliharaan meliputi penyiraman, pemupukan dan pengendalian hama penyakit.
sebagai perlakuan. Setiap satuan percobaan terdiri dari 20 buah cabai yang dipanen pada saat buah masih berwarna hijau berumur sekitar seminggu sebelum buah berwarna merah.
Perbanyakan inokulum dilakukan dengan cara mengambil koloni dari biakan murni kemudian dibiakkan pada media PDA dalam cawan petri. Biakan diinkubasi pada suhu 28 oC, diberi lampu selama 12 jam/hari selama 5-7 hari. Pemanenan konidia dilakukan dengan cara mencuci cawan dua kali dengan air steril sebanyak 15 ml kemudian permukaan isolat digosok menggunakan gelas L untuk mengambil konidia. Suspensi kemudian disaring menggunakan dua lapis kain saring untuk memisahkan potongan miselia. Kepadatan inokulum dihitung menggunakan haemocytometer sampai mencapai 5 x 105 konidia/ml.
Inokulasi dilakukan dengan menyuntikkan 2 µl suspensi konidia ke permukaan buah. Untuk buah berukuran kurang dari 4 cm inokulasi dilakukan pada satu titik sementara untuk buah yang berukuran lebih dari 4 cm inokulasi dilakukan pada dua titik dengan jarak 4 cm. Setelah inokulasi buah kemudian diinkubasi dalam suhu ruang dan ditempatkan di atas kawat dalam bak-bak plastik yang pada dasarnya diberi kertas tissue basah kemudian bak tersebut ditutup plastik hitam. Reaksi penyakit diamati lima hari setelah inokulasi.
Pengamatan yang dilakukan meliputi kejadian penyakit (KP) dan diameter bercak. Kejadian penyakit dihitung berdasarkan persentase buah yang terkena serangan dengan melihat ada atau tidak adanya bercak pada lima hari setelah inokulasi, dengan rumus berdasarkan AVRDC (2002):
x 100 %
Persentase dari genotipe yang diuji kemudian ditentukan kelas ketahanannya sesuai dengan Tabel 1.
Tabel 1. Kelas Ketahanan Berdasarkan Kejadian Penyakit
No Persentase Kelas Ketahanan
1 0 ≤ KP ≤10 Sangat Tahan
2 10 < KP ≤ 20 Tahan
3 20 < KP ≤ 40 Moderat
4 40 < KP ≤ 70 Rentan
5 KP > 70 Sangat Rentan
Sumber: Yoon (2003)
Penentuan skor pada buah yang telah diinokulasi dengan mengukur diameter bercak yang ditimbulkan sesuai dengan Tabel 2.
Tabel 2. Kelas Ketahanan Berdasarkan Diameter Bercak
No Diameter Bercak (cm) Kelas Ketahanan
1 0 Imun
2 0.1 – 0.5 Sangat Tahan
3 0.6 – 1 Tahan
4 1.1 – 2 Rentan
5 >2 Sangat Rentan
Sumber: AVRDC (1990)
Pendugaan Parameter Genetik Ketahanan Cabai terhadap Antraknosa
dengan Menggunakan Analisis Dialel
Tabel 3. Populasi half dialel dalam menduga parameter genetik untuk ketahanan cabai terhadap antraknosa
♂
♀ IPB C2 IPB C4 IPB C8 IPB C9 IPB C10 IPB C15 IPB C2 2 x 2
IPB C4 4 x 2 4 x 4
IPB C8 8 x 2 8 x 4 8 x 8
IPB C9 9 x 2 9 x 4 9 x 8 9 x 9
IPB C10 10 x 2 10 x 4 10 x 8 10 x 9 10 x 10
IPB C15 15 x 2 15 x4 15 x 8 15 x 9 15 x 10 15 x 15
Analisis Data:
1. Pendugaan parameter genetik menggunakan pendekatan Hayman
Pendugaan parameter genetik ketahanan cabai terhadap antraknosa dilakukan dengan analisis dialel menggunakan pendekatan Hayman (Singh dan Chaudhary 1979).
a. Analisis ragam
Populasi dialel dianalisis menggunakan rancangan kelompok lengkap teracak dengan tiga ulangan menggunakan model statistik:
Yij= µ + αi+ βj+ εij
Keterangan:
Yij = Nilai pengamatan dari perlakuan genotipe ke-i
µ = Nilai rata-rata
αi = Pengaruh perlakuan genotipe ke-i
βj = Pengaruh kelompok ke-j
εij = Pengaruh galat percobaan pada perlakuan genotipe ke-i
b. Pendugaan ragam dan peragam
∑
xij
Rata-rata tetua (
ML0) =2
1 ∑ X ij
Ragam tetua (V0L0) =
∑ (
Xij)
2-
n - 1
i=j n
n 2
1 n
∑
XijRagam array (Vri) =
∑ (
Xij)
2 - j=ln - 1 j=l n
1 n
Rata-rata ragam array (V1L1) =
∑
Vrin i=l
n 2
1 n
∑
XiRagam rata-rata array (V0L1) =
∑ (
Xi)
2 - i=ln– 1 i=l n
n 2
1 n
∑
XijXi’jPeragam tetua dan keturunan (Wri) =
∑ (
XijXi’j) -
j=l;i=ln – 1 j=l;i=l n
1
Rata-rata peragam tetua dan array (W0L1) =
∑
Wrin
Perbedaan rata-rata tetua dan rata-rata semua keturunan 1 1 n 2
(ML1– ML0)2 =
∑
Xij -∑
Xijn n i=l;j=l i=j
n
c. Uji hipotesis
Kesahihan hipotesis diuji dengan koefisien regresi, menggunakan ragam dan peragam.
b = (Cov (Wr,Vr)/(Var (Vr))
SE (b) = [(Var (Wr) – b * (Cov (Wr, Vr))/ (Var (Vr) * (n-1))]1/2
Uji hipotesis: H0 : b = 1 H1 : b≠ 1
Jika b = 1, maka tidak terdapat interaksi gen non alelik
d. Grafik Wr – Vr
Parabola diperoleh dengan menghubungkan titik-titik dari persamaan:
Wri = (Vri x V0L0)1/2
Regresi diperoleh dengan menghubungkan titik-titik dari persamaan:
Wri = Wr–bVr + bVri
Intersep regresi diperoleh dari:
ɑ = Wr - bVr
semakin dekat letak tetua dengan pangkal persilangan sumbu x-y, kandungan gen dominannya relatif semakin tinggi, sebaliknya semakin jauh letak tetua dengan pangkal persilangan sumbu x-y semakin kecil kandungan gen dominannya.
e. Pendugaan komponen ragam
Pendugaan komponen ragam yang dilakukan adalah:
D = V0L0 – E
F = 2V0L0 – 4W0L0 – 2(n-2) E/n
H1 = V0L0 – 4W0L1 + 4 V1L1 – (3n – 2) E/n H2 = 4V1L1 – 4 V0L1 – 2E
h2 = 4(ML1 – ML0)2 – 4(n-1) E/n S2 = ½ [Var (Wr – Vr)]
SE (D) = [(n5 + n4)/n5] * (S2)
SE(H1) = [(n5 + 41n4– 16n3 + 16n2)/n5] * (S2) SE(H2) = [(36n4)/n5] * (S2)
SE(h2) = [(16n4 + 16n2– 32n + 16)/n5] *(S2) SE(E) = [(n4)/(n5] * (S2)
Keterangan:
D : komponen ragam karena pengaruh aditif
F : nilai tengah Fr untuk semua array, Fr adalah peragam pengaruh aditif dan non aditif pada array ke-r
H1 : komponen ragam karena pengaruh dominan
H2 : perhitungan untuk menduga proporsi gen negatif dan positif pada tetua
h2 :pengaruh dominansi (sebagai jumlah aljabar dari semua persilangan saat heterozigous)
E : komponen ragam karena pengaruh lingkungan
Jika intersep bernilai positif atau D > H1 maka interaksi yang terjadi adalah
dominansi sebagian. Jika bernilai negatif atau D < H1maka interaksi yang
terjadi adalah overdominansi. Jika D = H1 interaksi yang terjadi adalah
dominan lengkap. Jika garis regresi menyentuh batas parabola maka tidak terdapat dominansi.
f. Pendugaan parameter lain
Parameter lain yang diduga adalah: Rata-rata tingkat dominansi = (H1/D)1/2
Proporsi gen-gen dengan pengaruh positif dan negatif dalam tetua = H2/4H1
Proporsi gen-gen dominan dan resesif dalam tetua = [(4DH1)1/2 + F] / [(4DH1)1/2 – F]
Jumlah kelompok gen yang mengendalikan sifat dan menimbulkan dominansi = h2/H2
Heritabilitas arti luas (h2bs)
Heritabilitas arti sempit (h2ns)
= (½ D + ½ H1 – ½ H2 –½ F) / ( ½ D +½ H1 – ½ H2 –½ F +E)
Jika korelasi negatif, nilai Wri + Vri-nya paling rendah, berarti mengandung gen dominan paling banyak.
2. Pendugaan Daya Gabung
Untuk menduga nilai daya gabung umum (DGU) dan daya gabung khusus (DGK) dilakukan analisis dialel menggunakan metode 2 Griffing (Singh dan Chaudhary 1979).
Model statistika yang digunakan adalah :
Yij = m + gi + gj + sij+ 1/bc ∑∑eijkl
Keterangan:
Yij = nilai tengah genotipe i x j m = nilai tengah umum
gi = daya gabung umum (DGU) tetua ke-i gj = daya gabung umum (DGU) tetua ke-j sij = pengaruh daya gabung khusus
1/bc ∑∑eijkl = nilai tengah pengaruh galat.
Komponen ragam untuk daya gabung disajikan pada Tabel 4. Tabel 4. Sidik ragam untuk Analisis Daya Gabung Metode 2
SK Db JK KT KTH
DGU P – 1 JKdgu KTdgu σ2e+ σ2dgk+ (p+2) σ2dgu
DGK P(p-1)/2 JKdgk KTdgk σ2e+ σ2dgk
Galat (r-1)[(p-1)+p(p-2)/2] JKgalat KTgalat σ2e
Pengaruh daya gabung umum (gi) = n (Yi.+Y.j) –1/n2 Y
Keterangan:
gi = nilai daya gabung umum galur ke-i
Yi. = jumlah nilai tengah persilangan genotipe ke-i
Y.j = jumlah nilai tengah selfing genotipe ke-j
Pengaruh daya gabung khusus (sij) = (Yji+Yij)- n(Yi.+Yii+Y.j+Yjj) + 1/n2 Y
Keterangan:
sij = nilai daya gabung khusus persilangan antara galur ke-i dan ke-j Yij = nilai tengah persilangan antara galur ke-i dan ke-j
Yji = nilai tengah persilangan antara galur ke-j dan ke-i Yi. = jumlah nilai tengah persilangan genotipe ke-i Yii = jumlah nilai tengah selfing genotipe ke-i Y.j = jumlah nilai tengah persilangan genotipe ke-j Yjj = jumlah nilai tengah selfing genotipe ke-j
Y.. = total nilai tengah genotipe
3. Pendugaan Heterosis
Nilai heterosis diduga berasarkan nilai tengah kedua tetua (mid parent) dan nilai tengah tetua terbaik (best parent) atau heterobeltiosis.
µF1 - µMP
Heterosis = x 100 % µMP
µF1 - µMP
Heterobeltiosis = x 100 %
µ
BPKeterangan :
µF1 = nilai tengah turunan
µMP= nilai tengah kedua tetua = (P1 + P2) µBP = nilai tengah tetua terbaik
Penentuan Karakter Seleksi Menggunakan Analisis Lintasan
Tajur II Bogor. Percobaan menggunakan metode Rancangan Kelompok Lengkap Teracak dengan tiga ulangan dan genotipe sebagai perlakuan, setiap satuan percobaan terdiri dari 20 tanaman. Data diameter bercak diperoleh dari percobaan pengujian ketahanan cabai terhadap antraknosa (Percobaan 1). Genotipe-genotipe yang digunakan beserta nilai tengahnya disajikan pada Lampiran 8. Peubah yang diambil dari penelitian Pranita (2007) adalah: tinggi tanaman, tinggi dikotomus, diameter kanopi, panjang daun, lebar daun, waktu panen, bobot per buah, panjang buah, tebal daging buah, diameter tengah buah dan jumlah buah total.
Analisis Data
1. Keeratan hubungan antar karakter dianalisis menggunakan analisis korelasi Pearson dengan persamaan sebagai berikut:
[∑ (Xi – X) (Yi – Y)]
r =
√∑(Xi - X)2(Yi – Y)2 dimana:
r = koefisien korelasi
Xi = nilai pengamatan ke-i pada peubah pertama X = rataan karakter pertama
Yi = nilai pengamatan ke-i pada peubah kedua Y = rataan karakter kedua
2. Analisis lintasan dihitung berdasarkan persamaan simultan sebagai berikut (Singh dan Chaudary 1979):
r11r12 . . . r1p C1 r1y
r21r22 . . . r2p C2 r2y
. . . . . . . . . . . . = . . . . . . . . . . . . . . . . . . rp1rp2 . . . rpp Cp rpy
Berdasarkan persamaan di atas, nilai C dapat dihitung menggunakan rumus: C = Rx-1Ry
Keterangan:
Rx = matriks korelasi antar peubah bebas Rx-1 = invers matriks Rx
C = vektor koefisien lintasan yang menunjukkan pengaruh langsung setiap peubah bebas yang telah dibakukan terhadap peubah tak bebas
HASIL DAN PEMBAHASAN
Pengujian Ketahanan Cabai Terhadap Antraknosa
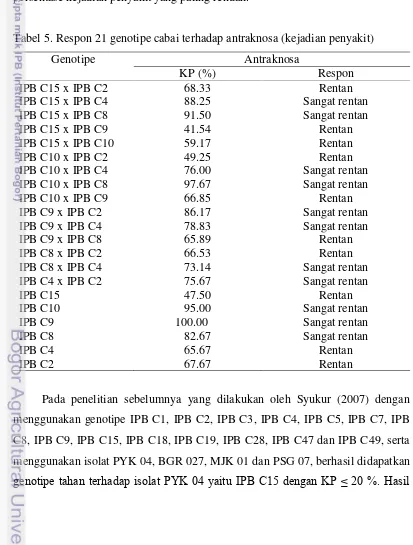
[image:38.595.100.513.252.797.2]Pada pengujian ketahanan terhadap antraknosa yang dilakukan pada buah hijau yang sudah matang, dari pengamatan kejadian penyakit (Tabel 5) tidak ada tetua maupun hibrida yang tahan terhadap antraknosa. Dari 21 genotipe yang diuji hibrida IPB C15 x IPB C9 (41.54 %) dan tetua IPB C15 (47.5 %) menunjukkan persentase kejadian penyakit yang paling rendah.
Tabel 5. Respon 21 genotipe cabai terhadap antraknosa (kejadian penyakit)
Genotipe Antraknosa
KP (%) Respon
IPB C15 x IPB C2 68.33 Rentan
IPB C15 x IPB C4 88.25 Sangat rentan
IPB C15 x IPB C8 91.50 Sangat rentan
IPB C15 x IPB C9 41.54 Rentan
IPB C15 x IPB C10 59.17 Rentan
IPB C10 x IPB C2 49.25 Rentan
IPB C10 x IPB C4 76.00 Sangat rentan
IPB C10 x IPB C8 97.67 Sangat rentan
IPB C10 x IPB C9 66.85 Rentan
IPB C9 x IPB C2 86.17 Sangat rentan
IPB C9 x IPB C4 78.83 Sangat rentan
IPB C9 x IPB C8 65.89 Rentan
IPB C8 x IPB C2 66.53 Rentan
IPB C8 x IPB C4 73.14 Sangat rentan
IPB C4 x IPB C2 75.67 Sangat rentan
IPB C15 47.50 Rentan
IPB C10 95.00 Sangat rentan
IPB C9 100.00 Sangat rentan
IPB C8 82.67 Sangat rentan
IPB C4 65.67 Rentan
IPB C2 67.67 Rentan
yang sama juga didapatkan oleh Putri (2010) yang melakukan penelitian terhadap 25 genotipe cabai dengan menggunakan isolat PYK 04, dimana genotipe IPB C15 termasuk dalam kategori tahan dengan nilai KP 5 %.
Pada penelitian ini genotipe IPB C15 termasuk dalam kategori rentan dengan KP 47.5 %, berdasarkan kelas ketahanan yang digunakan oleh Yoon (2003) (Tabel 1), walaupun bila dibandingkan dengan tetua lainnya yang mempunyai nilai KP lebih dari 60 %, tetua ini mempunyai nilai KP yang paling rendah, selain itu terdapat satu genotipe F1 yaitu IPB C15 x IPB C9 yang memiliki nilai KP yang paling rendah diantara 21 genotipe yang diuji (41.54 %) (Tabel 5). Perbedaan hasil tersebut disebabkan oleh perbedaan isolat yang digunakan, dimana dalam penelitian ini digunakan isolat BKT 04. Isolat BKT 04 berasal dari daerah Bukittinggi yang berada pada ketinggian 909-941 m dpl dengan suhu antara 16.1-24.9 ºC, sementara itu isolat PYK 04 berasal dari daerah Payakumbuh yang berada pada ketinggian 514 m dpl dengan suhu rata-rata 26 ºC. Isolat PYK dan BKT diduga merupakan strain yang berbeda sehingga menimbulkan respon ketahanan yang berbeda ketika diujikan pada genotipe yang sama.
Penelitian Syukur et al. (2009) yang dilakukan terhadap 14 genotipe cabai dan empat isolat C. acutatum menunjukkan bahwa genotipe yang sama memberikan respon yang berbeda terhadap isolat yang berbeda. Dari penelitian tersebut diketahui bahwa tingkat ketahanan cabai terhadap antraknosa sangat dipengaruhi oleh faktor genotipe dan isolat yang digunakan, dimana pengaruh isolat merupakan penyumbang keragaman terbesar. Kombinasi antara sifat struktural dan reaksi biokimia yang digunakan dalam mekanisme pertahanan tanaman terhadap serangan penyakit, berbeda antara setiap sistem kombinasi inang-patogen. Bahkan pada inang dan patogen yang sama, kombinasi tersebut dapat menghasilkan respon tanaman yang berbeda dalam kondisi tertentu, seperti perbedaan umur, jenis organ dan jaringan tanaman yang terserang, keadaan hara tanaman serta kondisi cuaca (Syukur 2007).
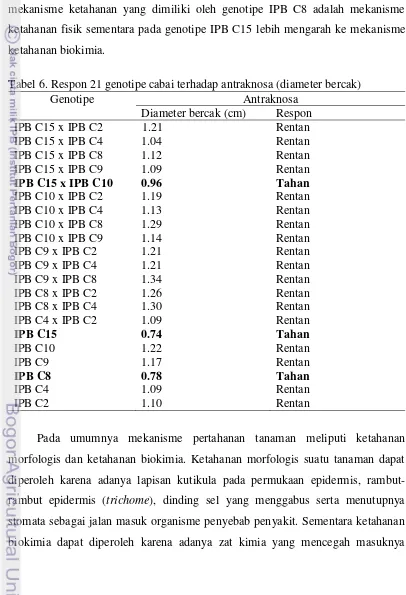
diduga disebabkan oleh adanya mekanisme pertahanan tertentu dari masing-masing genotipe yang dapat mencegah perluasan infeksi antraknosa sehingga diameter bercak yang dihasilkan lebih kecil dibandingkan dengan genotipe lainnya. Syukur et al. (2009) menyatakan bahwa terdapat indikasi bahwa mekanisme ketahanan yang dimiliki oleh genotipe IPB C8 adalah mekanisme ketahanan fisik sementara pada genotipe IPB C15 lebih mengarah ke mekanisme ketahanan biokimia.
Tabel 6. Respon 21 genotipe cabai terhadap antraknosa (diameter bercak)
Genotipe Antraknosa
Diameter bercak (cm) Respon
IPB C15 x IPB C2 1.21 Rentan
IPB C15 x IPB C4 1.04 Rentan
IPB C15 x IPB C8 1.12 Rentan
IPB C15 x IPB C9 1.09 Rentan
IPB C15 x IPB C10 0.96 Tahan
IPB C10 x IPB C2 1.19 Rentan
IPB C10 x IPB C4 1.13 Rentan
IPB C10 x IPB C8 1.29 Rentan
IPB C10 x IPB C9 1.14 Rentan
IPB C9 x IPB C2 1.21 Rentan
IPB C9 x IPB C4 1.21 Rentan
IPB C9 x IPB C8 1.34 Rentan
IPB C8 x IPB C2 1.26 Rentan
IPB C8 x IPB C4 1.30 Rentan
IPB C4 x IPB C2 1.09 Rentan
IPB C15 0.74 Tahan
IPB C10 1.22 Rentan
IPB C9 1.17 Rentan
IPB C8 0.78 Tahan
IPB C4 1.09 Rentan
IPB C2 1.10 Rentan
patogen, adanya substansi racun sitoplasmik dan adanya ion hara yang berperan sebagai antagonis (Crowder 2006).
Mekanisme pertahanan tanaman juga dapat berupa mekanisme pasif dan aktif. Mekanisme pasif melibatkan pertahanan struktural dan kandungan komponen antimikroba pada tanaman yang berfungsi untuk mencegah kolonisasi patogen. Pada mekanisme aktif, tanaman mencegah perluasan serangan patogen dengan bereaksi secara langsung pada saat tanaman tersebut terserang oleh patogen. Reaksi tersebut melibatkan reaksi biokimia, seperti respon hipersensitif nekrotik, dan modifikasi sel serta jaringan. Kim et al. (2004) menyatakan bahwa terdapat hubungan genetik, biokimia dan struktural antara reaksi hipersensitif atau
programmed cell death (PCD) dengan penebalan kutikula yang merefleksikan mekanisme pertahanan tertentu. Pada buah cabai yang diinokulasi oleh C. gloeosporioides, terbentuk formasi jaringan pemisah di sekitar luka akibat inokulasi setelah reaksi awal PCD yang memisahkan jaringan sehat dengan jaringan yang terinfeksi.
Salah satu cara untuk membedakan spesies C. acutatum dengan spesies lainnya adalah dengan melihat perbedaan warna koloni. Koloni C. acutatum
berwarna pink atau oranye, sementara koloni C. gloeosporioides berwarna abu-abu (Wharton dan Uribeondo 2004), walaupun Forster dan Adaskaveg (1999) menemukan isolat C. acutatum yang diambil dari almond mempunyai dua fenotipe, yaitu abu-abu dan pink.
Gambar 1. Koloni C. acutatum berwarna oranye hasil inokulasi
Gambar 2. Buah cabai hasil inokulasi
Keterangan: buah yang menampakkan gejala (kiri) dan buah yang tidak menampakkan gejala (kanan).
Pendugaan Parameter Genetik Ketahanan Cabai Terhadap Antraknosa
dengan Menggunakan Analisis Dialel
Pendugaan Parameter Genetik Menggunakan Pendekatan Hayman
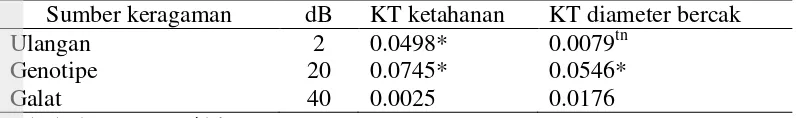
Tabel 7. Kuadrat tengah ketahanan cabai terhadap antraknosa
Sumber keragaman dB KT ketahanan KT diameter bercak
Ulangan 2 0.0498* 0.0079tn
Genotipe 20 0.0745* 0.0546*
Galat 40 0.0025 0.0176
* = berbeda nyata, tn = tidak nyata
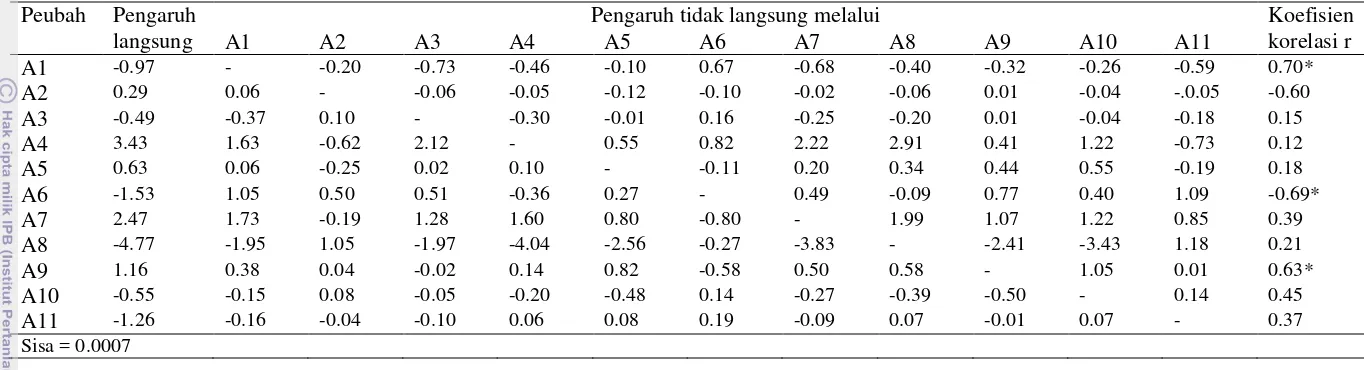
Hasil pendugaan parameter genetik untuk ketahanan tanaman cabai terhadap antraknosa disajikan pada Tabel 8. Hasil uji koefisien regresi b (Wr,Vr) tidak berbeda nyata dengan satu untuk peubah ketahanan dan diameter bercak (Tabel 8). Menurut Roy (2000) jika nilai b (Wr,Vr) tidak berbeda nyata maka tidak terdapat interaksi gen non alelik, dengan demikian tidak ada interaksi antar gen pada kedua peubah tersebut dan hal ini menunjukkan bahwa salah satu asumsi analisis silang dialel dapat terpenuhi.
Tabel 8. Pendugaan parameter genetik ketahanan tanaman cabai terhadap antraknosa
Parameter genetik Ketahanan Diameter bercak
b(Wr, Vr) 0.476tn 1.013tn
D 0.038* 0.036*
F 0.056tn 0.047*
H1 0.121* 0.069*
H2 0.096* 0.044*
h2 0.005tn 0.062*
E 0.001tn 0.006*
(H1/D)1/2 1.785 1.388
H2/4H1 0.197 0.159
Kd/Kr 2.380 2.850
h2/H2 0.047 1.415
r - 0.587 - 0.797
h2bs h2ns
0.956 0.751
0.14 0.277
*= berbeda nyata, tn = tidak berbeda nyata
Nilai D dan nilai H1 menunjukkan pengaruh aditif dan pengaruh dominan.
Nilai D dan H1 yang berbeda nyata pada ketahanan dan diameter bercak
menunjukkan bahwa kedua peubah tersebut dipengaruhi oleh pengaruh aditif dan dominan. Pada pengujian ketahanan terhadap antraknosa ini pengaruh dominan lebih besar dibandingkan dengan pengaruh aditif karena nilai H1 yang lebih besar
daripada nilai D pada ketahanan (D=0.038;H1=0.121) dan diameter bercak
[image:43.595.108.509.378.598.2]Distribusi gen dalam tetua dapat dilihat dari nilai H2. Apabila gen menyebar
secara merata dalam tetua nilai H2 tidak berbeda nyata dan demikian pula
sebaliknya. Pada ketahanan dan diameter bercak nilai H2 berbeda nyata, hal
tersebut menunjukkan bahwa gen dalam tetua menyebar secara tidak merata. Hal ini juga ditunjukkan oleh nilai H2/4H1 yang kurang dari 0.25 (Singh & Chaudary
1979), dimana nilai H2/4H1 pada ketahanan adalah 0.197 dan pada diameter
bercak adalah 0.159 (Tabel 8).
Proporsi gen-gen positif terhadap gen-gen negatif dapat terlihat dari nilai H1
dan H2. Apabila H1 > H2 maka gen-gen yang banyak adalah gen-gen positif
demikian pula sebaliknya. Nilai H1 dan H2 pada Tabel 8 menunjukkan bahwa pada
pengujian ketahanan tanaman cabai terhadap antraknosa ini gen-gen positif lebih banyak daripada gen-gen negatif, dimana nilai H1 lebih besar daripada H2 pada
ketahanan (0.121;0.096) dan pada diameter bercak (0.069;0.044).
Tingkat dominansi pada ketahanan dan diameter bercak adalah over dominan. Hal ini ditunjukkan oleh nilai (H1/D)1/2 lebih dari satu untuk ketahanan
(1.785) dan diameter bercak (1.388) (Tabel 8). Menurut Hayman (1954) nilai (H1/D)1/2 lebih dari satu menunjukkan over dominansi.
Nilai Kd/Kr menunjukkan proporsi gen dominan terhadap gen resesif, jika Kd/Kr > 1 maka gen-gen dominan lebih banyak di dalam tetua, sebaliknya apabila nilai Kd/Kr < 1 maka gen-gen resesif lebih banyak di dalam tetua. Dari hasil analisis diketahui bahwa nilai Kd/Kr karakter ketahanan cabai terhadap antraknosa lebih besar dari satu (Tabel 8) dimana nilai Kd/Kr pada ketahanan adalah (2.380) sedangkan nilai Kd/Kr pada diameter bercak adalah (2.850). Hal ini menunjukkan gen-gen dominan lebih banyak di dalam tetua.
Simpangan rata-rata F1 dari rata-rata tetua yang ditunjukkan oleh nilai h2.
Nilai h2 tidak berbeda nyata untuk ketahanan tetapi berbeda nyata untuk diameter bercak. Nilai h2/H2 menunjukkan jumlah kelompok gen pengendali. Jumlah
kelompok gen pengendali ketahanan terhadap antraknosa adalah satu kelompok gen (Tabel 8).
genetik. Hal tersebut juga ditandai oleh nilai E yang tidak berbeda nyata pada ketahanan. Nilai duga heritabilitas arti sempit dalam percobaan ini tergolong rendah, yaitu 0.14 untuk ketahanan dan 0.27 untuk diameter bercak (Tabel 8). Hal ini menunjukkan aksi gen aditif berperan kecil dalam ketahanan terhadap antraknosa.
Hasil penghitungan r (Wr+Vr<Yr) pada percobaan ini adalah negatif (Tabel 8). Jika nilai r (Wr+Vr<Yr) negatif maka nilai kuantifikasi yang tinggi dominan terhadap yang rendah (Jagau 1993). Hal ini menunjukkan bahwa ketahanan cabai terhadap antraknosa dikendalikan oleh gen resesif. Dari Gambar 3 dan Gambar 4 dapat dilihat bahwa garis lurus Wri memotong sumbu tegak (Wr) di bawah
pangkal sumbu ordinat, hal ini menandakan terdapat pengaruh overdominansi pada ketahanan dan diameter bercak. Unay et al. (2004) menyatakan bila garis lurus Wri memotong sumbu tegak (Wr) di bawah pangkal sumbu ordinat maka
terdapat pengaruh overdominansi pada karakter tersebut. Hal ini sesuai dengan nilai (H1/D)1/2 yang lebih dari satu untuk ketahanan dan diameter bercak (Tabel
8).
Nilai Urutan dominansi tetua berdasarkan (Wr+Vr) pada ketahanan adalah IPB C4 (0.0018), IPB C2 (0.0144), IPB C8 (0.0169), IPB C15 (0.0374), IPB C10 (0.0577), IPB C9 (0.0645) (Tabel 9). Urutan dominansi ketahanan juga dapat terlihat pada Gambar 3. Dimana semakin dekat letak titik nilai tetua pada titik nol maka tetua tersebut semakin banyak mengandung gen dominan serta sebaliknya (Sousa dan Malouf 2003).
Tabel 9. Sebaran Vr + Wr
Genotipe Ketahanan Diameter Bercak
IPB C15 0.0374 0.0394
IPB C10 0.0577 0.0190
IPB C9 0.0645 0.0036
IPB C8 0.0169 0.0793
IPB C4 0.0018 0.0072
Gambar 3. Hubungan antara peragam (Wr) dan ragam (Vr) untuk ketahanan
Urutan dominansi tetua berdasarkan (Wr+Vr) pada diameter bercak adalah IPB C2 (-0.0021), IPB C9 (0.0036), IPB C4 (0.0072), IPB C10 (0.0190), IPB C15 (0.0394) dan IPB C8 (0.0793) (Tabel 9). Pada peubah ketahanan, tetua IPB C4 adalah tetua yang paling banyak mengandung gen dominan karena nilainya paling dekat dengan titik nol, sedangkan tetua IPB C9 adalah tetua yang paling banyak mengandung gen resesif karena paling jauh dari titik nol (Gambar 3). Pada diameter bercak, tetua yang paling banyak mengandung gen dominan adalah IPB C2 karena paling dekat dengan titik nol, sedangkan tetua yang paling banyak mengandung gen resesif adalah IPB C8 karena paling jauh dari titik nol (Gambar 4).
Wri Yr dugaan IPB C4
IPB C2
IPB C8 IPB C15 IPB C10
Gambar 4. Hubungan antara peragam (Wr) dan ragam (Vr)
untuk diameter bercak.
Dari hasil analisis diketahui bahwa ketahanan terhadap antraknosa isolat BKT 04 lebih dipengaruhi oleh aksi gen dominan. Ragam genetik dominan merupakan penyebab utama ketidaksamaan diantara kerabat. Ragam ini merupakan basis utama bagi heterosis dan kemampuan daya gabung (Syukur 2007). Ketahanan terhadap antraknosa dikendalikan oleh gen resesif. Hasil tersebut sesuai dengan hasil penelitian yang dilakukan oleh Syukur (2007) yang melakukan penelitian dengan menggunakan isolat PYK 04, BGR 027, MJK 01 dan PSG 07. Hal ini dapat ditunjukkan dengan nilai r dari peubah ketahanan dan diameter bercak yang bernilai negatif. Hal tersebut juga terlihat pada Gambar 3 dan Gambar 4 dimana tetua IPB C15 yang termasuk tahan pada peubah diameter bercak dan mempunyai persentase kejadian penyakit yang terendah diantara tetua lainnya, berada pada titik yang jauh dari titik sumbu. Hal tersebut menunjukkan bahwa tetua IPB C15 banyak mengandung gen resesif, sementara tetua lainnya membawa gen dominan.
Dari sudut pandang pemuliaan tanaman, ketahanan yang dikendalikan oleh gen dominan lebih menguntungkan daripada ketahanan yang dikendalikan oleh gen resesif, karena akan diturunkan pada hibrida F1, walaupun jika hanya salah satu tetua yang memiliki alel ketahanan. Perakitan varietas tahan dengan gen resesif akan sulit dan memakan banyak waktu, walaupun demikian ketahanan
IPB C2
IPB C4 IPB C10
IPB C15
IPB C8
IPB C9
yang dikendalikan oleh gen resesif lebih dapat bertahan lama dibandingkan dengan ketahanan yang dikendalikan oleh gen dominan (Kim et al. 2007).
Pendugaan Daya Gabung
Pada percobaan pendugan daya gabung digunakan peubah ketahanan dan diameter bercak. Kuadrat tengah daya dabung umum dan daya gabung khusus pada ketahanan dan diameter bercak berbeda nyata (Tabel 10). Hal ini menunjukkan satu atau lebih genotipe cabai penggabung yang baik pada karakter ketahanan terhadap antraknosa. Hal tersebut juga mengindikasikan bahwa karakter ketahanan terhadap antraknosa dipengaruhi aksi gen aditif dan dominan. Pengertian dari daya gabung yang baik adalah kapasitas suatu tetua untuk menghasilkan keturunan yang superior bila digabungkan dengan tetua lain (Borojevic 1990).
Tabel 10. Kuadrat tengah daya gabung ketahanan cabai terhadap antraknosa Sumber keragaman KT diameter bercak KT ketahanan
Daya gabung umum 0.032* 0.024*
Daya gabung khusus 0.020* 0.028*
Galat 0.007 0.001
Pada peubah diameter bercak karena perhitungan DGU dan DGK berdasarkan diameter bercak luka pada buah maka yang diharapkan adalah nilai DGU dan DGK yang negatif karena semakin kecil diameter bercak maka semakin tahan genotipe tersebut terhadap antraknosa. Pada peubah diameter bercak genotipe yang memiliki nilai DGU negatif adalah IPB C15 (-0.123) dan IPB C8 (-0.003) (Tabel 11), walaupun termasuk dalam kategori rentan tetapi tetua IPB C15 dan IPB C8 memiliki daya gabung yang lebih baik bila dibandingkan dengan tetua-tetua lainnya. Tetua dengan nilai daya gabung umum yang baik mempunyai kemungkinan untuk mengembangkan galur yang baik pada generasi selanjutnya. Pada karakter diameter bercak terdapat enam persilangan yang memiliki nilai DGK negatif dimana genotipe IPB C15 x IPB C10 dan IPB C4 x IPB C2, masing-masing memiliki nilai DGK yang terendah, yaitu (-0.08) (Tabel 12).
Tabel 11. Nilai daya gabung umum (DGU) ketahanan cabai terhadap antraknosa Genotipe DGU diameter bercak DGU ketahanan
IPB C15 - 0.123 0.088
IPB C10 0.031 - 0.031
IPB C9 0.055 - 0.031
IPB C8 - 0.003 - 0.057
IPB C4 0.007 - 0.011
IPB C2 0.034 0.041
Pada peubah ketahanan karena perhitungan DGU dan DGK menggunakan nilai 1-kp maka yang diharapkan adalah nilai positif. Genotipe IPB C15 dan IPB C2 memiliki nilai DGU positif untuk peubah ketahanan yaitu 0.088 dan 0.041 (Tabel 11), hal tersebut menunjukkan bahwa kedua tetua tersebut walaupun termasuk dalam kategori rentan namun memiliki daya gabung yang lebih baik dibandingkan dengan tetua-tetua lainnya untuk karakter kejadian penyakit antraknosa. Dari analisis daya gabung khusus diperoleh delapan persilangan yang menghasilkan DGK positif walaupun genotipe-genotipe tersebut termasuk dalam kategori rentan. Sujiprihati (1996) menyatakan bahwa daya gabung khusus (DGK) yang positif untuk karakter produksi menunjukkan bahwa tetua tersebut mempunyai kombinasi hibrida yang tinggi dengan salah satu tetua yang digunakan. Persilangan IPB C15 x IPB C9 memiliki nilai DGK tertinggi, yaitu (0.26) (Tabel 12).
Tabel 12. Nilai daya gabung khusus (DGK) ketahanan cabai terhadap antraknosa Genotipe DGK diameter bercak DGK ketahanan
IPB C15 x IPB C10 - 0.08 0.08
IPB C15 x IPB C9 0.03 0.26
IPB C15 x IPB C8 0.12 - 0.21
IPB C15 x IPB C4 0.03 - 0.23
IPB C15 x IPB C2 0.17 - 0.08
IPB C10 x IPB C9 - 0.07 0.13
IPB C10 x IPB C8 0.13 - 0.15
IPB C10 x IPB C4 - 0.04 0.02
IPB C10 x IPB C2 - 0.01 0.23
IPB C9 x IPB C8 0.20 0.16
IPB C9 x IPB C4 0.02 - 0.01
IPB C9 x IPB C2 - 0.01 - 0.14
IPB C8 x IPB C4 0.17 0.07
IPB C8 x IPB C2 0.11 0.08
[image:49.595.108.510.516.752.2]Tetua dengan nilai DGU yang baik memiliki kemungkinan yang lebih besar untuk menghasilkan keturunan dengan nilai DGK yang baik, bila dibandingkan dengan tetua yang memiliki nilai DGU yang rendah (Borojevic 1990). Hal ini terlihat pada peubah ketahanan dimana genotipe IPB C15 yang memiliki nilai DGU yang paling tinggi bila dibandingkan dengan tetua lainnya, yaitu 0.0883 (Tabel 11), menghasilkan F1 dengan DGK tertinggi dibandingkan dengan F1 lainnya, yaitu IPB C15 x IPB C9 dengan nilai DGK (0.26) (Tabel 12).
Hal yang berbeda terlihat pada peubah diameter bercak, dimana genotipe dengan nilai DGK negatif rendah, yaitu IPB C4 x IPB C2 dengan nilai DGK -0.08 (Tabel 11) diperoleh dari tetua dengan nilai DGU yang positif yaitu IPB C4 (0.007) dan IPB C2 (0.034) (Tabel 11). Sementara genotipe IPB C15 x IPB C10 yang memiliki nilai DGK yang sama dengan genotipe IPB C4 x IPB C2 yaitu -0.08 diperoleh dari tetua dengan nilai DGU negatif yaitu IPB C15 (-0.123) dan tetua dengan DGU positif yaitu IPB C10 (0.031). Hasil yang berbeda antara daya gabung kedua peubah mengindikasikan bahwa nilai DGU belum tentu dapat digunakan untuk menduga nilai DGK suatu varietas. Kemampuan nilai DGU untuk menduga nilai DGK akan meningkat apabila tetua yang digunakan dalam analisis dialel tersebut dalam jumlah yang besar sehingga kombinasi persilangan dalam populasi tersebut juga dalam jumlah yang besar (Borojevic 1990).
Pendugaan Heterosis
Peubah yang digunakan dalam pendugaan nilai heterosis adalah kejadian penyakit dan diameter bercak. Nilai yang diharapkan adalah nilai heterosis negatif karena semakin rendah persentase kejadian penyakit dan semakin kecil diameter bercak maka semakin tahan genotipe tersebut terhadap antraknosa.
Nilai tengah tetua dan F1 adalah spesifik pada tiap parameter genetik oleh karena itu nilai yang dihasi