SULAWESI SELATAN
JUSRI NILAWATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi dengan judul Reproduksi Ikan Telmatherina sarasinorum (Kottelat, 1991) Sebagai Dasar Konservasi di Danau Matano Sulawesi Selatan adalah karya saya dengan arahan dari Komisi Pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Februari 2012
as the Foundation of Conservation in Lake Matano South Sulawesi. Under direction of SULISTIONO, DJADJA S. SJAFEI†, M.F. RAHARDJO, and ISMUDI MUCHSIN.
The research was aimed to analyze characteristics of preferential spawning habitat, spatial and temporal distribution, and reproduction of T. sarasinorum in spawning habitat. Research was carried out in Lake Matano from September 2008 to August 2009 at different sampling sites. Fish were caught by using mini beach seine of 10 m length, 3 m depth, and 3 mm mesh size. Habitat condition was analyzed, and water physical chemical characteristics were measured monthly. Fish standard length, total weight, and gonad weight were measured. Results showed that the fish preferred to spawn in sandy gravel arena and root arena shaded by vegetation or boulders. Mean standard length of male and female were 54,50 mm and 48,60 mm, respectively. Male has larger body and dominant in population. Population was dominated by mature individuals. Male and female’s first standard length at gonadal maturation was 40,28 mm and 37,95 mm, respectively. Mean fecundity was 224. Oocyte and spermatocyte development of T. sarasinorum was asynchronic; the species spawned in batches. Oocyte diameter ranged between 0,50 and 1,75 mm. The spawning peaked at the end of dry season with gradual increased of water level. The fish protected the freshly laying eggs. In addition, the sneakers and single males seemed picking the sperm remnants. Currently, conservation of the fish and its habitat is urgently required cause of increasing anthropogenics surrounding the lake.
JUSRI NILAWATI. Reproduksi Ikan Telmatherina sarasinorum (Kottelat, 1991) Sebagai Dasar Konservasi di Danau Matano Sulawesi Selatan. Dibimbing oleh SULISTIONO, DJADJA S. SJAFEI, M.F. RAHARDJO dan ISMUDI MUCHSIN.
Danau Matano adalah salah satu pusat biodiversitas dunia yang memiliki endemisitas tinggi. Ikan Telmatherina sarasinorum merupakan salah satu ikan endemik Danau Matano. Ikan ini mempunyai bentuk dan warna yang indah sehingga berpotensi untuk dikembangkan sebagai ikan hias, tetapi hingga saat ini belum dapat dipelihara di luar habitat aslinya.
Penelitian reproduksi ikan Telmatherina sarasinorum di Danau Matano bertujuan untuk menganalisis karakteristik habitat, distribusi spasial dan temporal dan reproduksi T. sarasinorum yang berada pada habitat pemijahan di Danau Matano. Penelitian dilakukan di Danau Matano Sulawesi Selatan dari bulan September 2008 sampai dengan Agustus 2009 pada 15 lokasi sampling. Ikan contoh ditangkap dengan pukat pantai mini berukuran panjang 10 m, tinggi 3 m dan mata jaring 3 mm.
Parameter fisik kimiawi perairan dan substrat pemijahan diukur setiap bulan. Kualitas air dan substrat pemijahan dianalisis. Habitat dan tingkah laku pemijahan ikan dianalisis melalui pengamatan bawah air dengan melakukan snorkeling. Suhu, pH dan oksigen terlarut diukur dengan water quality checker Horiba. Padatan tersuspensi total dan padatan terlarut total diukur di laboratorium. Kecerahan diukur dengan papan putih secara horisontal.
Ikan contoh ditentukan jenis kelamin dan warnanya sebelum diawetkan. Panjang baku (PB), berat total, berat gonad, tingkat kematangan gonad, indeks kematangan gonad, faktor kondisi relatif, fekunditas dan diameter telur ditentukan. Jaringan histologi gonad dibuat untuk analisis tingkatan perkembangannya. Tingkah laku pemijahan ikan di habitatnya dicatat langsung di dalam air pada kertas plastik, difoto dan direkam dengan kamera vidio.
yang siap memijah, yang ditandai dengan tingkat kematangan gonad (TKG), indeks kematangan gonad (IKG) dan faktor kondisi yang tinggi. IKG rata-rata ikan jantan lebih rendah daripada IKG rata-rata betina. Sebagian besar tingkat perkembangan oosit dan spermatosit terdapat di dalam gonad yang sama; ikan T. sarasinorum adalah pemijah sebagian. Ketiga kelompok ukuran diameter telur terdapat di dalam gonad, dengan komposisi telur berukuran kecil jumlahnya paling banyak dan telur berukuran besar jumlahnya paling sedikit. Puncak pemijahan terjadi pada akhir musim kemarau dengan kenaikan muka air secara perlahan, dan musim hujan. Ukuran panjang baku ikan jantan petama kali matang kelamin (TKG IV) adalah 40,28 mm sedangkan ikan betina adalah 37,95 mm. Fekunditas rata-rata adalah 224 butir (kisaran 64 – 488 butir).
Tingkah laku pemijahan ditandai dengan ikan berpasangan. Ikan kawin di arena pemijahan pada substrat pasir di antara kerikil dan batu, atau pada alga yang menyelimuti perakaran. Pasangan ikan sering diikuti oleh ikan jantan pengganggu pemijahan dan jantan tunggal lain. Ikan-ikan jantan ini memakan sisa sperma yang tidak membuahi telur. Jantan utama, dan kadang-kadang betina, menyembunyikan telur di antara alga atau di dalam pasir. Pemijahan paling banyak terjadi pada siang hari.
© Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
SULAWESI SELATAN
JUSRI NILAWATI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Ilmu Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup: 1. Dr. Ir. Didik Wahju Hendro Tjahjo, M.Si. 2. Dr. Ir. Ridwan Affandi
Penguji Luar Komisi pada Ujian Terbuka: 1. Dr. Ir. Gadis Sri Haryani
Puji dan syukur penulis panjatkan ke hadirat Tuhan Yang Maha Pengasih dan Maha Penyayang, karena atas segala rahmat dan hidayah-Nya penulis dapat menyelesaikan penulisan disertasi berjudul “Reproduksi Ikan Telmatherina sarasinorum (Kottelat, 1991) sebagai Dasar Konservasi di Danau Matano Sulawesi Selatan”.
Penulis menyampaikan terima kasih yang sebesar-besarnya kepada:
1. Dr. Ir. Sulistiono, M.Sc. sebagai ketua komisi pembimbing, Prof. Dr. Ir. M.F. Rahardjo dan Prof. Dr. Ir. Ismudi Muchsin sebagai anggota komisi pembimbing yang telah memberikan arahan dan bimbingan selama penyusunan disertasi ini.
2. Penghargaan yang setinggi-tingginya kepada Dr. Ir. Djadja S. Sjafei (Alm.) atas bimbingan dan arahan kepada penulis, dan semoga amal ibadah beliau diterima Allah SWT.
3. Penguji luar komisi serta Pimpinan IPB, Pimpinan Departemen dan Pimpinan Program Studi Ilmu Perairan yang telah berkenan menyumbangkan buah pikiran untuk memperkaya tulisan ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Februari 2012
Pada kesempatan ini penulis menyampaikan ucapan terima kasih yang sebesar-besarnya kepada:
1. Kementerian Pendidikan Nasional, Direktorat Jenderal Pendidikan Tinggi atas bantuan beasiswa BPPS 2005 dan Hibah Doktor.
2. Pt Inco Tbk. Indonesia atas bantuan akomodasi di lapangan.
3. The Stability of Rainforest Margins in Indonesia (Storma) atas bantuan dana penelitian.
4. Rektor Universitas Tadulako Palu atas bantuan dan kesempatan yang telah diberikan kepada penulis untuk melanjutkan pendidikan S3.
5. Ayahanda E. Soeparman dan Ibunda Sunarmi atas curahan kasih sayang, semangat, dorongan serta doa bagi penulis dalam menuntut ilmu.
6. Papa Yasin Tantu (Alm.) yang kasih sayangnya dapat penulis rasakan melalui kehangatan keluarga dan Mama Quraisyin Abdulwali (Almh.) atas kasih sayang, perhatian dan doa di saat hidupnya.
7. Aki Sukanta Astraperwata (Alm.) yang melalui penanya senantiasa mendorong penulis untuk senantiasa berjuang menuntut ilmu.
8. H. Hasan Athori (Alm.) dan Uwa dr. Sunarli (Alm.) atas nasihat dan dorongan untuk menyelesaikan studi.
9. Adinda Rizki Abdussalam, SH, Afiati Nurrohmah, SH, Arif Rachman Saleh, Amd., Senti Elisah, S.Ei., Dody Djatmika Sutisna, dan seluruh keluarga besar Sukanta Astraperwata dan Sumaatmadja atas dukungan semangat dan doa yang telah diberikan. Kakanda Isma Y. Tantu, Muh. Rum Y. Tantu, Usman Y. Tantu, M.Kes, S.Sos., Rukyani Y. Tantu, Maryam Y. Tantu, Adinda Isra Y. Tantu, Ir. Rizal Y. Tantu, M.Si., dan Irfan Y. Tantu, dan seluruh keluarga besar Tantu-Abdulwali yang telah memberikan dukungan semangat dan doa kepada penulis.
12. Teman-teman Program Studi Ilmu Perairan (Bu Maja, Bu Sriati, Pak Fadly, Pak Eman, Pak Heru, Pak Erly, Bu Niken dan Bu Ina) atas dukungan semangat dan kerja sama yang telah diberikan selama masa studi.
Penulis dilahirkan di Bandung pada tanggal 13 Juni 1967 dari orang tua Ayah E. Soeparman S.A. dan Ibu Sunarmi. Penulis adalah anak pertama dari empat bersaudara. Pendidikan sarjana ditempuh di Jurusan Manajemen Sumber Daya Perairan Fakultas Perikanan Universitas Sam Ratulangi Manado dan lulus tahun 1990. Pada tahun 1991 penulis mulai bekerja sebagai staf pengajar pada Fakultas Perikanan Universitas Sam Ratulangi Manado. Pada tahun 1993 penulis melanjutkan studi ke Department of Biology, Faculty of Science University of Waterloo Canada dan meraih gelar Master of Science pada tahun 1995.
Pada tahun 1998 penulis menikah dengan Ir. Fadly Y. Tantu, M.Si. dan dikaruniai empat orang putera puteri yaitu Fauzia Noorchaliza (11 tahun), Fatimah Noorasysyifa, Fadhilah Noor Nabiilah (7 tahun) dan Fathan Noor Ilmi Fadly Tantu (2,5 tahun).
Pada tahun 2003 penulis pindah tugas sebagai staf pengajar pada Program Studi Perikanan Fakultas Pertanian Universitas Tadulako Palu. Kemudian pada tahun 2005 penulis melanjutkan studi pada Program Studi Ilmu Perairan Sekolah Pascasarjana Institut Pertanian Bogor.
Karya ilmiah yang sudah dipublikasikan adalah:
DAFTAR TABEL ... xxiii
DAFTAR GAMBAR ... xxv
DAFTAR LAMPIRAN ... xxvii
1. PENDAHULUAN Latar Belakang... 1
Tujuan dan Manfaat Penelitian ... 9
Kebaruan ... 9
Ruang Lingkup Penelitian ... 9
2. HABITAT PEMIJAHAN IKAN T. sarasinorum DI DANAU MATANO Pendahuluan ... 11
Bahan dan Metode ... 12
Hasil dan Pembahasan ... 15
Kesimpulan... 33
3. SEBARAN SPASIAL-TEMPORAL IKAN T. sarasinorum DI DANAU MATANO Pendahuluan ... 35
Bahan dan Metode ... 36
Hasil dan Pembahasan ... 37
Kesimpulan... 47
4. REPRODUKSI DAN TINGKAH LAKU PEMIJAHAN IKAN T. sarasinorum DI DANAU MATANO Pendahuluan ... 49
Bahan dan Metode ... 51
Hasil dan Pembahasan ... 54
Kesimpulan... 86
5. PEMBAHASAN UMUM ... 87
6. KESIMPULAN DAN SARAN ... 95
1. Klasifikasi ukuran substrat ... 13 2. Lokasi pengamatan, jumlah arena pemijahan dan deskripsi
karakter substrat pemijahan ... 19 3. Kisaran parameter fisik kimiawi perairan pada setiap lokasi
sampling di Danau Matano ... 24 4. Kisaran parameter fisik kimiawi perairan pada setiap bulan
di Danau Matano ... 28 5. Nilai indeks kematangan gonad ikan T. sarasinorum jantan dan
1. Sketsa lokasi penelitian di Danau Matano ... 14 2. Kontur dasar di kedua tipe habitat pemijahan ... 15 3. Arena pemijahan T. sarasinorum: arena batu berpasir (atas)
dan arena perakaran (bawah) ... 16 4. Curah hujan rata-rata harian wilayah dan tinggi muka air di
Danau Matano selama penelitian ... 21 5. Fluktuasi parameter fisik kimiawi perairan secara spasial di
Danau Matano ... 25 6. Fluktuasi parameter fisik kimiawi perairan secara spasial di
Danau Matano ... 29 7. Pengelompokan habitat pemijahan ikan T. sarasinorum di
Danau Matano ... 31 8. Jumlah ikan rata-rata yang tertangkap pada masing-masing lokasi
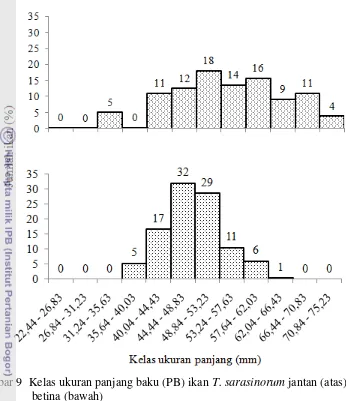
sampling selama periode penelitian ... 37 9. Kelas ukuran panjang baku (PB) ikan T. sarasinorum jantan (atas)
dan betina (bawah) ... 40 10. Sebaran jumlah rata-rata ikan secara temporal ... 42 11. Jumlah ikan T. sarasinorum jantan menurut warna secara
spasial ... 44 12. Jumlah ikan T. sarasinorum jantan menurut warna secara
temporal ... 46 13. Jumlah ikan T. sarasinorum jantan menurut warna per kelas
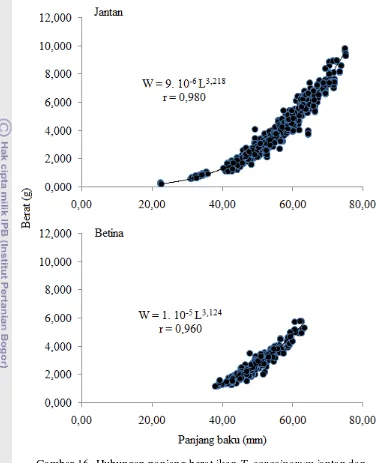
ukuran ... 47 14. Nisbah kelamin T. sarasinorum menurut lokasi sampling ... 54 15. Nisbah kelamin T. sarasinorum menurut waktu sampling ... 56 16. Hubungan panjang berat ikan T. sarasinorum jantan (atas) dan
betina (bawah) ... 57 17. Faktor kondisi relatif ikan T sarasinorum jantan dan betina
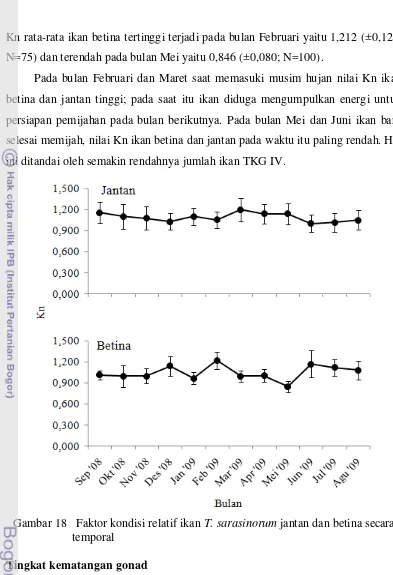
secara spasial ... 58 18. Faktor kondisi relatif ikan T. sarasinorum jantan dan betina
secara temporal ... 59 19. Struktur histologis gonad ikan T. sarasinorum jantan ... 60 20. Struktur histologis gonad ikan T. sarasinorum betina ... 62 21. Persentase TKG ikan T. sarasinorum jantan (atas) dan betina
23. Persentase TKG ikan T. sarasinorum jantan (atas) dan betina
(bawah) menurut kelas ukuran ... 66 24. Nilai IKG rata-rata ikan T. sarasinorum jantan dan betina
secara spasial ... 69 25. Nilai IKG rata-rata ikan T. sarasinorum jantan dan betina
secara temporal ... 70
26. Hubungan fekunditas dan panjang baku ikan T. sarasinorum ... 71
27. Sebaran diameter telur ikan T. sarasinorum pada TKG III,
1. Deskripsi lokasi penelitian ... 109 2. Pohon tambeua (Myrtacea) ... 112 3. Curah hujan rata-rata harian wilayah selama penelitian ... 113 4. Sebaran spasial ikan T. sarasinorum jantan menurut kelas
ukuran panjang ... 114 5. Sebaran spasial ikan T. sarasinorum betina menurut kelas
ukuran panjang ... 115 6. Sebaran temporal ikan T. sarasinorum jantan menurut
kelas ukuran panjang ... 116 7. Sebaran temporal ikan T. sarasinorum betina menurut kelas
ukuran panjang ... 117 8. Ikan T. sarasinorum jantan kuning (a), biru (b), biru kuning
(c), abu-abu (d) dan abu-abu kuning (e), dan betina (f) ... 118 9. Variasi spasial nisbah kelamin ikan di Danau Matano ... 119 10. Variasi temporal nisbah kelamin ikan di Danau Matano ... 120 11. Deskripsi karakter morfologis dan histologis gonad T. sarasinoum
jantan pada berbagai tingkat perkembangan ... 121 12. Deskripsi karakter morfologis dan histologis gonad T. sarasinoum
1 PENDAHULUAN
Latar Belakang
Pulau Sulawesi telah lama dikenal sebagai pusat biodiversitas karena tingginya derajat endemisme di antara fauna aslinya (Whitten et al. 2002). Sekitar 76% ikan asli di Sulawesi adalah endemik (Kottelat et al. 1993). Ikan endemik adalah ikan yang sebarannya sangat terbatas dan tidak terdapat di tempat lain di dunia. Penelitian mengenai keberadaan ikan-ikan endemik dari perairan tawar Sulawesi menunjukkan bahwa ikan-ikan ini hanya hidup di danau-danau dan sungai-sungai yang terletak di bagian tengah Sulawesi dan tidak menyebar ke perairan tawar yang ada di Sulawesi Utara maupun Sulawesi Selatan (Soeroto & Tungka 1991; Soeroto et al. 1992; Soeroto & Tungka 1996).
Danau Matano adalah salah satu danau tua di dunia yang terdapat di Sulawesi. Danau tua, yaitu danau yang telah ada sejak masa sebelum Pleistosin (Wilke et al. 2008). Danau Matano berada pada bagian hulu dari rangkaian sistem danau Malili yaitu Matano, Mahalona dan Towuti. Tiga danau utama telah ditetapkan sebagai kawasan konservasi Taman Wisata Alam (TWA) melalui Surat Keputusan Mentan No. 274/Kpts/Um/4/1979 tanggal 24 April 1979. Luas kawasan ini mencapai 98.500,00 ha (termasuk perairan danau), dengan rincian luas masing-masing yaitu: TWA Matano 30.000 ha (perairan danau 16.408 ha), TWA Towuti 65.000 ha (perairan danau 56.108 ha) dan TWA Mahalona 3.500 ha (perairan danau 2.440 ha).
Walaupun dihubungkan oleh sistem aliran dari dua danau di hilirnya, danau ini terisolasi dari sistem hilir oleh rintangan-rintangan fisik bagi penyebaran biota akuatik. Danau ini memiliki luas 164 km2, kedalaman 590 m (terdalam ke-8 di dunia), airnya sangat jernih (kecerahan >20 m), memiliki pinggiran yang curam dengan daerah litoral yang terbatas, dan terkenal sebagai danau oligotrofik (Haffner et al. 2001).
melaporkan adanya satu spesies baru dari famili Telmatherinidae Danau Matano yaitu T. albolabiosus.
Menurut Haffner et al. (2001), interaksi antara perubahan iklim dan aktivitas manusia dapat menjadikan fauna endemik Danau Matano Sulawesi berada dalam bahaya. Habitat utama ikan-ikan endemik Danau Matano terus mendapat tekanan dari berbagai aktivitas yang berlangsung di sekitar danau. Beberapa aktivitas yang merupakan ancaman potensial bagi ekosistem danau dengan fauna akuatik endemiknya adalah (1) pembukaan lahan pertambangan, permukiman, jalan, perkebunan, (2) perubahan sistem hidrologi danau dan sungai Petea akibat pelurusan dan pembendungan sungai, (3) limbah rumah tangga dan minyak dari mesin perahu, dan (4) introduksi ikan.
Studi terkini mengenai ikan-ikan Telmatherinidae dari sistem danau-danau Malili yang sudah dilakukan adalah: keragaman dan evolusi (Herder et al. 2006); radiasi adaptif dan genetika populasi (Heath et al. 2006); pemeliharaan polimorfisme warna jantan pada telmatherinid (Gray et al. 2006); deskripsi perbandingan tingkah laku kawin ikan telmatherinid dari danau-danau Malili (Gray & McKinnon 2006). Para peneliti menunjukkan kekhawatiran dan perhatian terhadap keselamatan biodiversitas fauna akuatik endemik yang ada dalam sistem sungai-sungai ini, sebagai akibat pengelolaan danau yang dinilai buruk. Penelitian-penelitian selama ini tidak ada yang memusatkan perhatian pada reproduksi telmatherinid sehubungan dengan habitat pemijahannya.
Aksi manajemen yang komprehensif perlu diterapkan di danau ini, jika tidak maka dapat dipastikan spesies akuatik khususnya yang endemik akan punah seiring dengan rusak dan hilangnya habitat di danau Matano. Kelompok ikan endemik yang diprediksi mendapat tekanan pertama adalah ikan-ikan telmatherinid, salah satunya T. sarasinorum, karena ikan ini memanfaatkan habitat litoral untuk menjalankan fungsi-fungsi hidupnya.
dengan akar-akar pohon menjurai atau pada batang/ ranting pohon tumbang yang diselimuti alga (fitofil). Pada lokasi habitat pertama telur diletakkan di substrat berpasir di antara batuan, sedangkan pada habitat kedua telur diletakkan pada substrat beralga. Perairan di habitat pemijahan kedua ini diamati memiliki dasar yang relatif berlumpur atau tertutup sedimen.
Latar belakang yang dikemukakan di atas menjadi dasar rencana penelitian yang diarahkan pada telaahan keterkaitan antara strategi reproduksi T. sarasinorum sehubungan dengan preferensi habitat dalam ekosistem danau yang sedang mengalami perubahan. Penggunaan spesies T. sarasinorum dalam penelitian ini didasarkan pada beberapa alasan yaitu: (1) merupakan salah satu spesies dominan dari kelompok telmatherinid penghuni Danau Matano, (2) menempati habitat litoral sebagai daerah pemijahan dan berbagai aktivitas biologi lainnya, dan (3) ditemukan memiliki dua habitat pemijahan berbeda (fitofil dan litofil).
Klasifikasi dan Ciri Taksonomik Ikan
Deskripsi taksonomi beberapa spesies anggota Telmatherinidae pertama kali dipublikasikan oleh Boulenger (1897), yang kemudian direvisi oleh Aarn dan Kottelat (1998) dan Kottelat (1990, 1991).
Klasifikasi T. sarasinorum menurut Kottelat (1991) adalah sebagai berikut: Kingdom : Animalia
sirip ventral I,5. Sisik pada baris longitudinal 32–34. Sisik pada baris transversal ½8½ (pada satu spesimen ½7½). Sisik predorsal 12–14. Sisik preoperkulum 4. Sisik operkulum 5–7. Gillraker pada lengkungan pertama 20–25. Warna ikan hidup, jantan bentuk kuning, tubuh keabu-abuan atau hijau kekuningan sampai kuning cerah, sirip-sirip kuning, dan sirip pektoral transparan. Ikan betina berwarna kuning kecoklatan (Kottelat 1991). Gray et al. (2003, 2006) mengidentifikasi lima bentuk warna jantan pada T. sarasinorum yaitu kuning, biru, biru kuning, abu, dan abu kuning, sedangkan betina berwarna abu-abu pasir. Heath et al. (2006) mengidentifikasi tiga warna dominan pada Telmatherinidae dari Danau Matano, yaitu biru, kuning dan coklat, serta dua warna transisi, yaitu kuning-biru dan kuning-hijau. Analisis filogenetik pada telmaterinid dari sistem Danau Malili menunjukkan suatu asal monofiletik kuno (Martens 1997; McKinnon 2002).
Penyebaran dan Habitat T. sarasinorum
Sebagai ikan endemik Danau Matano, menurut Kottelat (1991), wilayah penyebaran T. sarasinorum hanya terbatas di daerah dangkal sepanjang tepian danau, tetapi lebih sering terdapat di sepanjang tepian danau bagian selatan. Ikan menempati perairan dengan dasar berpasir dan hampir datar, serta berada pada perairan yang relatif dangkal (<1,5 m). Sehubungan dengan wilayah penyebaran dan habitatnya telah dilakukan pengamatan lapangan untuk studi pendahuluan, dengan melakukan snorkeling di 9 lokasi (27 November-1 Desember 2006), dan di 6 lokasi (5 -19 Januari 2007). Berdasarkan hasil pengamatan tersebut disimpulkan bahwa penyebarannya konsisten dengan laporan Kottelat (1991). Ikan T. sarasinorum dapat ditemukan mulai kedalaman 0,35 m sampai dengan <3m, dan tidak ditemukan pada habitat dangkal daerah berarus di in-let maupun
oleh Kottelat (1991); Gray et al. (2006); serta Gray & McKinnon (2006). Ikan T. sarasinorum memiliki alternatif habitat pemijahan, pertama di habitat pantai dangkal (kedalaman 1-2 m) dengan substrat terdiri dari kerikil kecil dan pasir, serta kedua pada habitat pantai relatif lebih dalam dan curam (kedalaman 1-3 m) pada garis pantai yang memiliki cabang-cabang bergelantungan yang ditutupi oleh algae (Gray & McKinnon 2006). Komposisi substrat dan kedalaman habitat juga merupakan penentu yang signifikan yang menentukan keberadaan ikan (Nilawati
et al. 2010).
Ikan-ikan yang ada di dalam sistem Danau Matano terutama ditemukan di daerah-daerah litoral. Tidak ditemukan ikan di kedalaman 30-40 m (Roy et al. 2004). Danau Matano mempunyai air yang sangat jernih (kecerahan > 23 m) dan biomassa fitoplankton rendah (< 52 ug/l berat basah) terutama terdiri dari cyanobacteria. Danau ini tidak mempunyai predator piscivora puncak dan jelas berada dalam kondisi yang sangat oligotrofik (Heath et al. 2006).
Indikator Reproduksi dan Tingkat Kebugaran Ikan
Studi reproduksi ikan, seperti penilaian ukuran pada kematangan, lamanya musim pemijahan, pemijahan dan fekunditas, memerlukan pengetahuan tentang tahap perkembangan gonad pada individu ikan. Metoda yang digunakan berkisar dari histologi hingga pentahapan secara visual berdasarkan penampilan eksternal. Ada metode lain di antara kedua metoda yang ekstrim ini, yaitu penentuan ukuran telur, pentahapan berdasarkan penampilan keseluruhan telur, dan indeks-indeks gonad (West 1990). Histologi gonad sering digunakan secara sendiri, atau bersama-sama dengan indeks kematangan gonad (IKG), untuk mengkonfirmasi fenotip gonad, menentukan kondisi perkembangan seksual, atau bahkan meneliti kerusakan reproduksi (Schmitt & Dethloff 2000).
lingkungan (misalnya perubahan musiman) atau tekanan-tekanan eksogen (misalnya paparan kontaminan) (Schmitt & Dethloff 2000).
Faktor kondisi adalah respon tingkat organisme, dengan faktor-faktor seperti status nutrisi atau pengaruh patogen yang menyebabkan berat melebihi atau kurang dari normal. Umumnya faktor kondisi bervariasi langsung dengan nutrisi. Faktor kondisi juga bisa bervariasi dengan musim, mungkin disebabkan perubahan ketersediaan makanan atau metabolisme, dan dengan perubahan status gonad. Faktor kondisi bervariasi di antara jenis-jenis ikan yang berbeda; juga bervariasi di antara ikan yang terdapat di lokasi berbeda (Ekanem 2000).
Polikromatisme dan Tingkah Laku Pemijahan
Telmatherinidae terkenal karena polikromatisme pada jantan yang ditunjukkan oleh setidaknya setengah dari spesies yang dideskripsikan oleh Kottelat (1990; 1991). Polikromatisme pada jantan telah lama dipelajari untuk memahami proses-proses yang memelihara keragaman di alam. Konservasi keragaman intraspesifik di alam adalah penting karena variasi tersebut merupakan awal dari keragaman di masa depan (Gray et al. 2006). Radiasi ikan yang menunjukkan polikromatisme warna jantan telah dipelajari secara luas oleh para ahli ekologi evolusi (Seehausen et al. 1999a; Seehausen et al. 1999b, Seehausen 2004; Turner et al. 2001; Fuller & Travis 2004; Joyce et al. 2005).
Tingkah laku pemijahan adalah aktivitas yang berhubungan langsung dengan produksi individu baru. Tingkah laku demikian kadang-kadang cukup sederhana, tetapi pada banyak spesies tingkah laku pemijahan bisa sangat kompleks dan meliputi pertunjukan dan gerakan yang menakjubkan.
Kompetisi untuk mendapatkan pasangan telah ditunjukkan dalam banyak penelitian. Ornamen pada jantan – baik ukuran sirip maupun warna– digunakan untuk menarik perhatian betina. Hal ini dapat terjadi pada spesies ikan yang mengalami kompetisi untuk mendapatkan pasangan (Andersson 1994). Selain itu, pada spesies yang tidak melakukan penjagaan teritori biasanya terjadi kompetisi antar jantan (Pyron 1996b).
Sistem pemijahan ikan T. sarasinorum dijelaskan oleh Gray & McKinnon (2006); T. sarasinorum adalah pemijah di substrat yang tidak melakukan penjagaan terhadap telur maupun larvanya. Kondisi lingkungan juga penting bagi jantan untuk memutuskan taktik perkawinan. Jantan kecil atau muda umumnya bisa berhasil kawin dengan menggunakan taktik berbeda dengan yang digunakan oleh jantan besar, kadang-kadang mengadopsi bentuk-bentuk alternatif tingkah laku pemijahan seperti menyerupai/mimikri betina dan tingkah laku mengganggu pembuahan (sneaking) (Pauers et al. 2004; Ito & Yanagisawa 2006).
Dimorfisme seksual
Dimorfisme ukuran seksual adalah berbeda menurut sistem kawin (Pyron 1996a). Dimorfisme ukuran seksual pada ikan berhubungan dengan tiga hal dalam sistem perkawinan yaitu: kompetisi sperma, penjagaan teritori oleh ikan jantan dan kontes/pertunjukan jantan-jantan.
Banyak spesies yang bersarang di substrat mempunyai ukuran jantan besar; ini berhubungan dengan keberhasilan menjaga dan mempertahankan sarang. Diantara spesies oportunistik, ikan jantan bisa lebih besar dan lebih menonjol (misalnya killifish Cyprinodontidae) atau lebih kecil dan lebih menonjol (guppy Poeciliidae) daripada betina. Perbedaan ukuran berkaitan dengan sistem sosial dan kompetisi di antara ikan-ikan jantan untuk mendapatkan habitat berkualitas tinggi. Dampak Antropogenik dan Kualitas Habitat
badan air. Keempat, pembuangan langsung atau tidak langsung bahan-bahan sintetik, baik organik maupun anorganik.
Hampir semua jenis aktivitas (pertanian, urbanisasi, komunikasi, pertambangan, industri) menyebabkan meningkatnya materi tersuspensi di permukaan perairan, terutama disebabkan oleh deforestasi (Elvira 1995). Konstruksi jalan menyebabkan peningkatan materi tersuspensi 5-20 kali. Operasi tambang, umumnya strip mining, menyebabkan tingginya materi tersuspensi di sungai. Operasi pengerukan di sungai baik yang bertujuan untuk memperdalam saluran atau untuk mengambil pasir, biasanya meningkatkan jumlah materi tersuspensi (Meybeck et al. 1989).
Degradasi kualitas air telah diketahui menyumbang secara langsung terhadap kehilangan habitat ikan dan penurunan manfaat-manfaat lain dari sumber daya perairan. Kualitas air yang baik merupakan persyaratan yang diperlukan untuk semua pemanfaatan sumber daya saat ini, termasuk pemeliharaan biodiversitas. Penetrasi cahaya di danau dikontrol oleh sedimen-sedimen tersuspensi dari aliran-aliran sungai dan hamparan danau oleh kelimpahan fitoplankton yang mengabsorpsi cahaya untuk fotosintesis (Anderson 1996).
Pengaturan aliran yang disebabkan oleh operasi dam dapat menurunkan keragaman biota (Agostinho et al. 2000). Misalnya, Cichla monoculus adalah spesies yang jarang di habitatnya sebelum pembangunan dam di Sungai Tocantins. Setelah dam dibangun spesies ikan ini menjadi berlimpah dan tersebar luas di dam. Kolonisasi spesies Cichla yang cepat dilaporkan setelah pembangunan dam di berbagai sungai di wilayah Brazil (Novaes et al. 2004). Status Konservasi
persaingan, dan (6) pemanasan global (Reid & Miller 1989; Moyle & Leidy 1992; Ogutu-Ohwayo et al. 1997).
Tujuan dan Manfaat Penelitian Tujuan penelitian adalah untuk :
1. Menganalisis karakteristik habitat yang menjadi aspek penentu preferensi pemijahan ikan T. sarasinorum di Danau Matano.
2. Menganalisis distribusi spasial dan temporal ikan T. sarasinorum di Danau Matano.
3. Menganalisis reproduksi ikan T. sarasinorum yang berada pada habitat pemijahan di Danau Matano.
Manfaat penelitian ini adalah mendapatkan konsep pengelolaan dan pemanfaatan ikan endemik rawan punah melalui konservasi spesies dan habitat untuk menjaga kestabilan keanekaragaman hayati di Danau Matano.
Kebaruan
Kebaruan penelitian ini meliputi: pertama, penggunaan T. sarasinorum
dalam penelaahan strategi reproduksi sehubungan dengan preferensi habitat dalam sebuah ekosistem danau tua; kedua, karakter bioekologi T. sarasinorum yang akan terungkap dari hasil penelitian ini dapat menjadi informasi baru yang dapat dijadikan dasar dalam konservasi ikan endemik di Danau Matano.
Ruang Lingkup Penelitian
2 HABITAT PEMIJAHAN IKAN
T. sarasinorum
DI DANAU MATANO
Pendahuluan
Penelitian mengenai habitat pemijahan ikan air tawar endemik Sulawesi yang dikaitkan dengan preferensi arena pemijahan belum pernah dilakukan dan ini adalah penelitian pertama. Penelitian dilakukan di Danau Matano, salah satu danau tua di dunia yang terdapat di bagian tengah Pulau Sulawesi. Pulau Sulawesi merupakan pulau terbesar di kawasan Wallacea, yang merupakan habitat campuran dari fauna Asia dan Australia, serta menjadi arena evolusi berbagai fauna endemik. Keunikan flora fauna Sulawesi tidak terlepas dari sejarah geologinya; pertemuan daratan Asia dengan daratan Australia menyebabkan terjadinya perpindahan flora dan fauna dari kedua daratan tersebut. Oleh karena itu kekhasan ekosistem perairan umum dengan keanekaragaman fauna khasnya di Sulawesi tidak terlepas dari peristiwa sejarah terbentuknya Pulau Sulawesi.
Whitten et al. (2002) mencatat 76% spesies ikan yang ditemukan di Sulawesi tidak ditemukan dimana pun di dunia. Danau Matano, Mahalona dan Towuti yang berada di bagian tengah Sulawesi merupakan salah satu tempat yang memiliki spesies ikan endemik. Ketiga danau ini berdekatan dan membentuk
suatu sistem danau yang dikenal dengan sebutan ―Kompleks Danau-danau
Malili‖. Ada dua danau lain yang termasuk dalam kompleks danau-danau ini yaitu danau Wawontoa dan Masapi.
Danau Matano merupakan hulu dari rangkaian sistem kompleks danau-danau Malili. Walaupun ketiga danau-danau ini dihubungkan oleh sistem aliran dari dua danau yang ada di hilirnya, namun danau ini terisolasi dari sistem hilir oleh rintangan fisik bagi penyebaran biota akuatik.
anggota (Glossogobius matanensis, G. intermedius, Mugilogobius adeia, M. latifrons), Adrianichthyidae, dan Hemirhamphidae masing-masing satu anggota yaitu Oryzias matanensis dan Dermogenys weberi (Tantu & Nilawati 2008). Menurut Soeroto (1997), mayoritas ikan air tawar Sulawesi adalah ikan sekunder, yaitu ikan yang mampu mentolerir kandungan garam; juga anggota dari famili ikan laut. Contoh ikan-ikan sekunder adalah Oriziidae dan Adrianichthyidae, sedangkan anggota dari famili ikan laut adalah Gobiidae, Eleotridae, Atherinidae dan Hemiramphidae.
Kelompok Telmatherinidae merupakan kelompok yang memiliki anggota paling banyak yang menghuni Danau Matano, dan T. sarasinorum adalah salah satu jenis dominan yang menempati daerah litoral danau. Studi mengenai T. sarasinorum belum banyak dilakukan, secara spesifik misalnya Gray et al. (2006) mempelajari tingkah laku kawin dari ikan ini. Umumnya studi yang berkaitan dengan Telmatherinidae ditujukan pada aspek keragaman dan evolusi, radiasi adaptif, genetika populasi, pemeliharaan polimorfisme warna jantan, dan perbandingan tingkah laku kawin (Herder et al. 2006; Heath et al. 2006; Gray et al. 2006; Gray & McKinnon 2006; Nilawati & Tantu 2007; Tantu & Nilawati 2007; Tantu & Nilawati 2008). Belum ada penelitian yang secara khusus mempelajari habitat pemijahan ikan endemik di Danau Matano, dan penelitian ini merupakan penelitian pertama dengan obyek ikan T. sarasinorum.
Penelitian ini menguraikan aspek habitat yang berkaitan dengan faktor-faktor fisik kimiawi perairan. Tujuan penelitian adalah menganalisis profil habitat pemijahan yang meliputi substrat pemijahan dan aspek fisik kimiawi perairan yang mendukung keberadaan strategi reproduksi. Diharapkan hasil penelitian ini dapat membantu upaya-upaya konservasi ikan dan habitatnya di Danau Matano.
Bahan dan Metode
pinggiran danau dan aktivitas masyarakat yang diduga bermakna bagi habitat ikan. Penilaian habitat pemijahan dilakukan melalui pengamatan bawah air dengan melakukan snorkeling untuk menentukan keadaan habitat dan substrat pemijahan. Habitat pemijahan ditentukan melalui penilaian adanya aktivitas pemijahan yang dilakukan oleh kelompok-kelompok ikan dalam habitat spesifik. Gambaran utuh mengenai keadaan habitat diperoleh dengan mengukur parameter fisik kimiawi perairan secara in situ (suhu, oksigen terlarut, pH, kecerahan dan transparansi air) dan ex situ (padatan tersuspensi total dan padatan terlarut total).
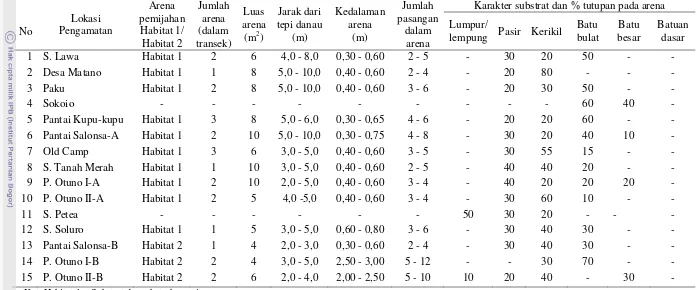
Penentuan lokasi pemijahan didasarkan pada hasil pengamatan bawah air oleh pengamat yang melakukan renang snorkeling dengan kecepatan lambat pada transek sepanjang 50 meter pada kedalaman antara 0,5 sampai dengan 2 meter sejajar garis pantai. Pengamat mengidentifikasi lokasi-lokasi yang secara khusus dijadikan sebagai arena pemijahan. Arena pemijahan didefinisikan sebagai area terbatas yang menjadi tempat berlangsungnya aktivitas kawin yang ditunjukkan oleh adanya pertunjukan pasangan jantan-betina, persaingan jantan-jantan untuk mendapatkan betina, aktivitas berpasangan, dan aktivitas kawin pasangan T. sarasinorum (Nilawati et al. 2010). Pengamat mencatat jumlah arena pemijahan yang terdapat dalam garis transek dan menilai persentase luas tutupan materi yang menyusun substrat pemijahan. Klasifikasi ukuran materi substrat mengikuti Wolman (1954) dalam Tabel 1.
Catatan: Komposisi substrat menurut Wolman (1954)
pengawet. Kecerahan diukur sebagai ukuran jarak pandang pengamat di dalam air terhadap benda berwarna putih berukuran 30 cm x 30 cm. Tinggi muka air danau dan curah hujan wilayah diperoleh berdasarkan data yang dikoleksi selama periode sampling dari stasiun pengamatan PT. INCO Sorowako.
Analisis statistik
Analisis keragaman satu arah dilakukan untuk menilai perbedaan antara parameter-parameter lingkungan yang diamati di dalam suatu lokasi dengan menggunakan fasilitas yang terdapat dalam perangkat lunak Minitab versi 14. Untuk mengetahui apakah terdapat korelasi di antara parameter-parameter lingkungan yang diukur dilakukan penghitungan koefisien korelasi Pearson r
dengan alat bantu perangkat lunak Minitab 14. Nilai uji statistik r bisa positif atau negatif tetapi selalu terletak di antara -1 dan +1. Nilai mendekati +1 menunjukkan korelasi positif yang kuat, sedangkan nilai mendekati -1 menunjukkan korelasi negatif yang kuat. Nilai 0 menunjukkan kurangnya korelasi, walaupun hal ini juga dapat berarti bahwa terdapat suatu korelasi yang lebih kompleks (McCleery et al. 2007).
Gambar 1 Sketsa lokasi sampling di Danau Matano
Ket.: 1) S. Lawa, 2) Desa Matano, 3) Paku, 4) Sokoio, 5) Pantai Kupu-kupu, 6,13) Pantai Salonsa, 7) Pantai Old Camp, 8) S. Tanah Merah,
Hasil dan Pembahasan
Deskripsi lokasi sampling
Sebanyak 15 lokasi di daerah litoral Danau Matano dijadikan sebagai titik pengamatan. Kelima belas lokasi ini menyebar dari bagian barat danau sampai ke bagian timur danau. Secara spasial lokasi-lokasi ini dapat dikelompokkan ke dalam 3 zona yaitu: zona 1 adalah lokasi sampling yang terletak di bagian barat danau (S. Lawa, Pantai Desa Matano, Pantai Paku dan Pantai Sokoio), zona 2 di bagian tengah danau (Pantai Kupu-kupu, Pantai Salonsa I-A, Pantai Salonsa I-B, Pantai Old-Camp, dan Pantai S. Tanah Merah) dan zona 3 di bagian timur danau (P. Otuno I-A, P. Otuno I-B, P. Otuno II-A, P. Otuno II-B, Pantai Sungai Petea, dan Pantai Sungai Soluro). Masing-masing lokasi secara ringkas dideskripsikan sebagaimana ditampilkan pada Lampiran 1. Beberapa vegetasi yang ditemukan di lokasi adalah Myrtacea (Lampiran 2), pohon sagu, Nephentes, pohon mangga dan Pandanus sp.
Menurut kontur dasar perairan kelima belas lokasi penelitian ini memiliki dua bentuk kontur dasar yaitu, kontur dasar landai dan kontur dasar curam (Gambar 2). Kontur dasar landai umumnya memiliki substrat dasar batu-batuan dan pasir.
Arena pemijahan
Pengamatan bawah air berhasil mengidentifikasi adanya habitat spesifik yang digunakan oleh T. sarasinorum untuk melakukan aktivitas perkawinan. Habitat pemijahan spesifik ini dalam penelitian ini disebut sebagai arena pemijahan, yang didefinisikan sebagai tempat berlangsungnya peristiwa perkawinan atau pemijahan kelompok ikan T. sarasinorum. Terdapat dua tipe arena pemijahan T. sarasinorum di Danau Matano (Gambar 3). Pertama arena pemijahan yang terletak di dasar perairan dengan kontur dasar landai yang substrat dasarnya terdiri dari pasir dan kerikil yang berada di antara batuan, atau ruang berbentuk kolam-kolam pasir bercampur kerikil. Kedua arena perakaran, ranting atau batang pohon yang terendam di perairan.
Sebuah arena dapat diidentifikasi dengan mengamati adanya peristiwa pemijahan yang ditandai oleh adanya rangkaian aktivitas kelompok ikan. Aktivitas itu berupa adanya beberapa ikan jantan yang sedang berusaha mendapatkan seekor betina yang ada di dalam arena pemijahan; adanya ikan betina yang berpasangan; serta ikan betina melakukan gerakan pelepasan telur dan dalam waktu yang hampir bersamaan ikan jantan melepaskan sperma yang ditandai oleh terlihatnya pasangan ikan jantan dan betina secara bersama-sama menekan abdomennya ke dasar substrat.
Secara keseluruhan ada 15 lokasi sampling yang diamati dan di dalamnya ditemukan 24 arena pemijahan yang terdiri atas 19 arena pemijahan pada substrat dasar perairan dan 5 arena pemijahan di perakaran. Luas keseluruhan arena pemijahan yang diamati dalam lokasi sampling kurang lebih 90 m² (Tabel 2). Susunan substrat dasar perairan di arena pemijahan bervariasi mulai dari substrat halus lumpur sampai dengam substrat batuan dasar. Tidak semua lokasi sampling memiliki arena pemijahan. Pantai Sokoio dan Sungai Petea tidak memiliki arena pemijahan. Tidak adanya arena pemijahan di Pantai Sokoio diduga berkaitan dengan tutupan substrat yang didominasi oleh batu-batuan besar yang tidak memiliki jebakan pasir atau kolam-kolam pasir. Di sekitar S. Petea substrat dasar perairannya banyak mengandung lumpur.
Penelitian ini juga berhasil mengidentifikasi bahwa arena pemijahan selalu berada di dekat tepian danau pada daerah bervegetasi pohon, dan berada dalam area bayang-bayang atau teduhan yang berasal dari pohon di tepi danau atau dari batu besar – batuan dasar di dekatnya. Umumnya jarak arena dari tepian berkisar antara 2 – 10 m, dan kedalaman arena berkisar antara 0,30 – 3,00 m.
kompetitor pembuahan. Sejauh ini tidak ditemukan penelitian mengenai pengaruh bayangan pohon atau benda lain di dalam habitat pemijahan ikan terhadap preferensi habitat pemijahan di dalam sistem lain.
Pemilihan tempat pelepasan telur (oviposition) dapat berdampak pada kelangsungan hidup keturunan (Smith et al. 2001). Tempat pelepasan telur dapat berbeda-beda kualitasnya. Pada spesies yang ikan jantannya melakukan pengasuhan terhadap anak, pemilihan pasangan oleh betina sering dipengaruhi oleh karakteristik posisi sarang, selain memperhatikan resiko predasi dan kondisi oksigen yang ada (Spence et al. 2007). Pemilihan tempat pelepasan telur adalah salah satu cara betina menjamin kelangsungan hidup anak. Seperti halnya ikan T. sarasinorum, ikan zebrafish betina adalah pemilih tempat pelepasan telur (Smith
et al. 2001). Jadi jika ikan betina aktif memilih tempat pelepasan telur, ikan jantan bisa meningkatkan keberhasilan pemijahannya dengan mengamankan telur yang telah dibuahi. Hal ini tampak pada ikan T. sarasinorum; ikan jantan tampak berusaha menutupi telur yang telah dibuahinya dengan alga atau pasir. Dengan demikian usaha-usaha pemilihan tempat pelepasan telur dan perlindungan telur yang baru dilepaskan oleh ikan T. sarasinorum sesungguhnya adalah bentuk kepedulian induk (parental care).
Tabel 2 Lokasi pengamatan, jumlah arena pemijahan dan deskripsi karakter substrat pemijahan
Karakter substrat dan % tutupan pada arena
lebar daripada bagian utara. Selain itu, pada sisi utara Danau Matano banyak terdapat batuan besar yang tidak disertai pasir.
Sementara itu, vegetasi adalah gambaran umum dari habitat alami zebrafish (Spence et al. 2007). Larvae zebrafish mempunyai perlengkapan untuk melekat dan mampu menempel pada tumbuhan setelah menetas sampai pecahnya gelembung renang. Penjelasan adaptif untuk preferensi ini ditunjukkan dengan meningkatnya kelangsungan hidup telur-telur di dalam kerikil. Telur-telur yang diinkubasi di dalam kerikil menunjukkan kelangsungan hidup yang secara signifikan lebih tinggi daripada di dalam lumpur. Pengaruh ini mungkin karena telur-telur yang diinkubasi di dalam kerikil mengalami oksigenasi yang lebih baik dibandingkan dengan yang diinkubasi di dalam lumpur; di dalam lumpur telur mungkin menghadapi resiko lebih besar terhadap mati lemas/kekurangan nafas atau infeksi oleh mikroorganisme (Wootton 1990). Unsur oksigen sangat diperlukan untuk perkembangan embrio.
Keuntungan lain dari kerikil sebagai substrat pemijahan adalah telur-telur jatuh di antara kerikil dan dengan demikian terlindung dari gangguan atau kanibalisme (Spence et al. 2007). Ikan T. sarasinorum yang memijah mencari daerah substrat yang lebih berpori berupa alga pada perakaran dan pasir. Pemilihan arena pemijahan di daerah yang ternaungi mungkin lebih berhubungan dengan kondisi intensitas cahaya yang lebih rendah (Goddard & Mathis 1997). Adanya preferensi tempat pemijahan demikian penting dalam menentukan manajemen habitat.
Keadaan hidrologis dan parameter fisik kimiawi perairan Danau Matano Curah hujan dan fluktuasi muka air
Tinggi muka air danau jika dilihat mulai dari Februari – Agustus 2009 tampak periode ini merupakan periode pengisian air danau. Hal ini karena pola yang terbentuk adalah peningkatan tinggi muka air secara terus-menerus tanpa fluktuasi mengikuti periode pola curah hujan. Puncak-puncak muka air danau tampaknya berada dalam periode bulan Agustus - Desember. Keadaan muka air terendah pada bulan Februari berada pada ketinggian 391,46 m dpl, sedangkan muka air tertinggi berada pada bulan September dengan ketinggian 392,65 m dpl. Gambar 4 memberikan informasi bahwa terdapat penurunan air yang terjadi sangat drastis pada periode bulan Desember - Januari dan titik terendahnya terjadi pada bulan Februari. Data ini menunjukkan bahwa tidak terjadi fluktuasi alami pada tinggi muka air danau. Data yang ditampilkan pada periode Desember, Januari dan Februari menunjukkan ada kegiatan pelepasan massa air Danau Matano.
Pola tersebut dikhawatirkan akan mengganggu biota perairan yang memanfaatkan daerah litoral sebagai tempat bereproduksi. Hal ini disebabkan pada saat muka air terendah sebagian daerah litoral menjadi kering. Pada saat itu ikan berpindah ke daerah yang masih tergenangi air dengan kedalaman sekitar 0,50 m.
Fluktuasi muka air memengaruhi proses dan pola ekologis danau dalam beberapa hal (Wantzen et al. 2008). Pada tahun 2003 dibangun dam di S. Petea yang dimaksudkan untuk pengadaan listrik bagi keperluan industri. Dam tersebut selesai dibangun dan mulai beroperasi pada tahun 2005. Pada waktu pintu air dam ditutup, massa air Danau Matano meningkat. Daerah pinggir danau yang tergenang menjadi bertambah luas. Naiknya muka air menyebabkan daerah-daerah yang tadinya tidak tergenangi menjadi tergenang. Pada saat itu pinggiran danau mengalami abrasi yang menyebabkan tanah bagian atas turun ke perairan menutupi daerah litoral yang menjadi tempat hidup ikan. Sedimentasi ini dapat memengaruhi keberadaan ikan di daerah tersebut.
Pada waktu pintu air di dam tersebut dibuka, massa air danau keluar menuju danau-danau di bagian hilir yaitu Danau Mahalona dan Towuti. Hal tersebut menyebabkan penurunan muka air secara tiba-tiba. Daerah-daerah litoral yang tadinya tergenang menjadi kering. Habitat pemijahan ikan asli danau itu yang umumnya terdapat di daerah litoral tiba-tiba menjadi kering. Telur-telur ikan yang baru dilepaskan diduga menjadi mati. Hal ini tentu berpengaruh terhadap keberadaan spesies ikan di habitatnya.
Secara alami, pada waktu air surut daerah litoral yang biasanya mengering dimanfaatkan oleh organisme terestrial seperti serangga untuk menjalankan sebagian siklus hidupnya untuk bereproduksi. Daun-daun dari pinggiran danau yang jatuh ke daerah litoral mengering. Pada waktu muka air meningkat, daerah tersebut tergenang air dan daun-daun mengalami dekomposisi. Serangga di perairan dan hasil dekomposisi tumbuhan itu menjadi sumber makanan bagi ikan-ikan di Danau Matano.
Tetapi perlu ditekankan bahwa fluktuasi muka air adalah pola alami yang dibutuhkan untuk kelangsungan hidup banyak spesies. Fluktuasi muka air alami di danau menjamin produktivitas dan biodiversitas. Hanya banjir dan kekeringan yang ekstrim dan yang bukan waktunya memberikan pengaruh berbahaya bagi biota.
luas perairan daratan di negara itu, telah mengalami pengaturan oleh manusia (Aroviita & Hämäläinen 2008). Fluktuasi muka air yang diubah adalah sebagian dari pengaruh antropogenik besar di dalam ekosistem danau dan sungai. Konstruksi dam dan reservoir untuk produksi listrik tenaga air dan pengontrol banjir, pemindahan air untuk irigasi, bersama-sama dengan penggunaan air lainnya, telah mengubah rejim alami hidrologis perairan tawar di seluruh dunia. Bagian perairan yang terutama rentan terhadap perubahan muka air adalah zona litoral danau. Organisme di daerah litoral danau mungkin terpengaruh langsung oleh kekeringan, dan terpengaruh tidak langsung oleh berkurangnya ketersediaan habitat dan sumber-sumber makanan.
Banyak faktor abiotik terutama di zona litoral danau-danau besar, yang dapat memengaruhi interaksi biotik pada berbagai kedalaman air: tekanan hidrolik pada organisme yang disebabkan oleh aksi gelombang lebih rendah di perairan yang lebih dalam (Baumgärtner et al. 2008). Stabilitas habitat, ukuran partikel substrat dan tempat berlindung makroavertebrata berubah dengan kedalaman air. Sampainya cahaya dan spektrum cahaya menjadi lebih sempit dengan kedalaman, dan komunitas alga epilitik—sumber makanan bagi peramban—berubah. Suhu dan kisaran suhu harian bervariasi bergantung pada kedalaman air dan musim.
Fluktuasi muka air jangka panjang berhubungan dengan pergeseran garis pantai yang besar, oleh karena itu dampak utama fluktuasi muka air jangka panjang terhadap ekositem danau adalah perubahan habitat (Hofmann et al. 2008). Fluktuasi muka air jangka panjang membanjiri/menggenangi daerah pinggiran yang sebelumnya kering atau permukaan sedimen di bawah permukaan air yang terpapar pada atmosfir. Jadi, fluktuasi muka air jangka panjang berperan dalam pemilihan spesies sessil yang beradaptasi pada kondisi kering atau basah. Selain itu, fluktuasi muka air jangka panjang menghasilkan suatu perubahan sifat permukaan sedimen di zona litoral, karena ukuran butiran di wilayah pantai tidak tersebar secara homogen tetapi berubah dari besar ke kecil menuju wilayah yang lebih dalam di danau.
ditandai oleh tingginya keragaman dan kelimpahan spesies dan penting untuk reproduksi dan siklus hidup banyak spesies ikan. Fluktuasi muka air dan curah hujan tampak tidak memengaruhi jumlah ikan T. sarasinorum di habitatnya (Bab 3). Kondisi berbeda dapat dilihat pada pemijahan ikan; puncak pemijahan ikan tampak dipengaruhi oleh fluktuasi muka air dan curah hujan (Bab 4).
Dinamika karakteristik fisik kimiawi perairan
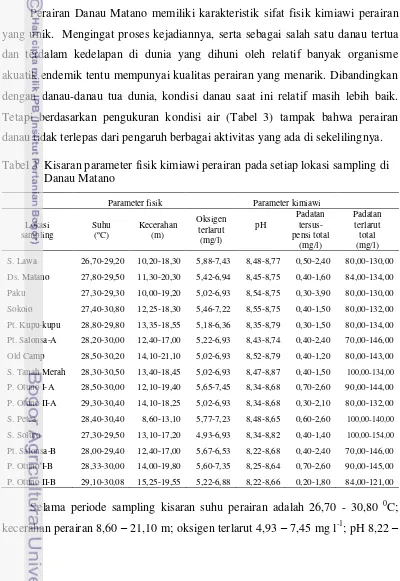
Perairan Danau Matano memiliki karakteristik sifat fisik kimiawi perairan yang unik. Mengingat proses kejadiannya, serta sebagai salah satu danau tertua dan terdalam kedelapan di dunia yang dihuni oleh relatif banyak organisme akuatik endemik tentu mempunyai kualitas perairan yang menarik. Dibandingkan dengan danau-danau tua dunia, kondisi danau saat ini relatif masih lebih baik. Tetapi berdasarkan pengukuran kondisi air (Tabel 3) tampak bahwa perairan danau tidak terlepas dari pengaruh berbagai aktivitas yang ada di sekelilingnya. Tabel 3 Kisaran parameter fisik kimiawi perairan pada setiap lokasi sampling di
Danau Matano
S. Lawa 26,70-29,20 10,20-18,30 5,88-7,43 8,48-8,77 0,50-2,40 80,00-130,00
Ds. Matano 27,80-29,50 11,30-20,30 5,42-6,94 8,45-8,75 0,40-1,60 84,00-134,00
Paku 27,30-29,30 10,00-19,20 5,02-6,93 8,54-8,75 0,30-3,90 80,00-130,00
Sokoio 27,40-30,80 12,25-18,30 5,46-7,22 8,55-8,75 0,40-1,50 80,00-132,00
Pt. Kupu-kupu 28,80-29,80 13,35-18,55 5,18-6,36 8,35-8,79 0,30-1,50 80,00-134,00
Pt. Salonsa-A 28,20-30,00 12,40-17,00 5,22-6,93 8,43-8,74 0,40-2,40 70,00-146,00
Old Camp 28,50-30,20 14,10-21,10 5,02-6,93 8,52-8,79 0,40-1,20 80,00-143,00
S. Tanah Merah 28,30-30,50 13,40-18,45 5,02-6,93 8,47-8,87 0,40-1,50 100,00-134,00
P. Otuno I-A 28,50-30,00 12,10-19,40 5,65-7,45 8,34-8,68 0,70-2,60 90,00-144,00
P. Otuno II-A 29,30-30,40 14,10-18,25 5,02-6,93 8,34-8,68 0,30-2,10 80,00-132,00
S. Petea 28,40-30,40 8,60-13,10 5,77-7,23 8,48-8,65 0,60-2,60 100,00-140,00
S. Soluro 27,30-29,50 13,10-17,20 4,93-6,93 8,34-8,82 0,40-1,40 100,00-154,00
Pt. Salonsa-B 28,00-29,40 12,40-17,00 5,67-6,53 8,22-8,68 0,40-2,40 70,00-146,00
P. Otuno I-B 28,33-30,00 14,00-19,80 5,60-7,35 8,25-8,64 0,70-2,60 90,00-145,00
P. Otuno II-B 29,10-30,08 15,25-19,55 5,22-6,88 8,22-8,66 0,20-1,80 84,00-121,00
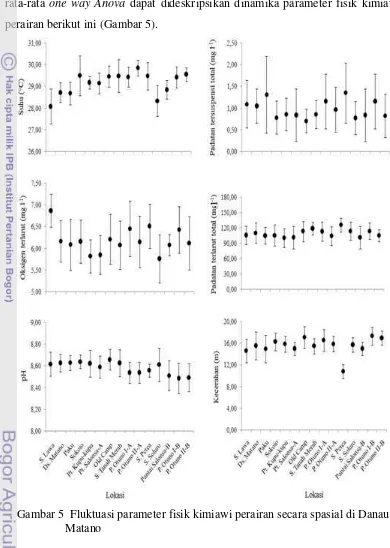
8,87; padatan tersuspensi total 0,20 - 3,90 mg l-1; dan padatan terlarut total 70 – 154 mg l-1. Secara spasial terdapat perbedaan nilai hasil pengukuran parameter fisik kimiawi antara lokasi sampling (Tabel 3). Berdasarkan Tabel 3 dan hasil uji rata-rata one way Anova dapat dideskripsikan dinamika parameter fisik kimiawi perairan berikut ini (Gambar 5).
Gambar 5 Fluktuasi parameter fisik kimiawi perairan secara spasial di Danau Matano
Sungai Lawa merupakan sungai utama yang mengalirkan air masuk ke dalam danau. Lokasi sampling di S. Lawa mempunyai naungan dari pohon di pinggiran danau.
Keadaan suhu perairan menurut lokasi sampling menunjukkan adanya perbedaan nyata rata-rata nilai pengukuran suhu di antara lokasi sampling. Suhu terendah berada di S. Lawa (28,08 0C) dan tertinggi di P. Otuno II-A (29,86 0C). Walaupun suhu air di S. Lawa merupakan suhu air terendah dibandingkan dengan suhu air di lokasi-lokasi lain di Danau Matano, hasil uji statistik one way Anova
menunjukkan tidak berbeda nyata dengan suhu air di Desa Matano, Pantai Paku dan S. Soluro. Daerah S. Lawa, Desa Matano dan Pantai Paku terletak di bagian hulu danau, sedangkan lokasi sampling di S. Soluro terletak dekat dengan sumber air masuk Danau Matano. Jika dilihat menurut zona, maka tampak bahwa suhu rata-rata perairan di lokasi-lokasi penelitian dalam zona 1 memiliki nilai lebih rendah daripada dua zona yang ada di bagian hilir. Tampak bahwa semakin ke bagian timur danau suhu rata-rata relatif semakin tinggi. Penelitian yang dilakukan oleh Haffner et al. (2001) menemukan bahwa secara vertikal suhu permukaan berkisar antara 27,53 0C – 29,06 0C, dan suhu permukaan dengan suhu pada kedalaman 40 meter hanya memiliki selisih 0,030C (27,53˚C di permukaan dan 27,56 0C untuk pengukuran pukul 09.00). Pengukuran pada waktu menjelang sore hari (pukul 15.00) suhu di permukaan lebih tinggi 1,47 0C daripada keadaan suhu pada kedalaman 20 m (29,06 0C di permukaan dan 27,59 0C di kedalaman 20 m). Menurut Haffner et al. (2001), perairan Danau Matano mengalami percampuran secara vertikal, dan terjadi proses penghangatan secara vertikal setiap hari.
Soluro (5,76 mg l-1 ± 0,56), berbeda nyata dengan oksigen terlarut rata-rata di S. Lawa, P. Otuno I-A, S. Petea, dan P. Otuno I-B.
Secara keseluruhan nilai pH perairan di lokasi-lokasi sampling tidak bervariasi lebar, dan tidak ada perbedaan yang nyata. Walaupun demikian, nilai pH air tertinggi ditemukan di lokasi S. Tanah Merah yaitu 8,87 pada bulan Juni. Nilai pH air terendah ditemukan di Pantai Salonsa B (8,22) pada bulan September, dan di P. Otuno II-B (8,22) pada bulan Januari. Berdasarkan pengamatan di bagian hulu S. Lawa terdapat penebangan kayu dan pengambilan rotan yang dilakukan oleh penduduk sekitar danau. Nilai pH di lokasi sampling S. Lawa masih berada dalam kisaran basa dan tidak berbeda jauh dengan kisaran pH di lokasi lain, sehingga diduga aktivitas penduduk tersebut tidak berpengaruh terhadap nilai pH perairan setempat; diamati bahwa penduduk tidak membuang sisa gergajian ke dalam perairan. Kisaran pH perairan Danau Matano berada dalam kisaran basa. Hal ini berkaitan dengan proses kejadian danau; Danau Matano merupakan perairan laut yang terperangkap saat pembentukan pulau Sulawesi. Daerah ini merupakan daerah karst. Batuan yang terdapat di dasar perairan dilapisi antara lain batuan kapur - terutama kalsium karbonat dari cangkang hewan (Whitten et al. 2002).
Nilai padatan tersuspensi total rata-rata antar lokasi sampling secara statistik tidak berbeda nyata. Padatan tersuspensi total rata-rata antar lokasi berkisar antara 0,71 – 1,35 mg l-1. Padatan tersuspensi total rata-rata relatif tinggi di S. Petea. Padatan tersuspensi total rata-rata antar lokasi menunjukkan beda nyata antar lokasi. Padatan tersuspensi total rata-rata tertinggi ditemukan di S. Petea (126,00 mg l-1 ± 13,58). Kandungan padatan tersuspensi total rata-rata di S. Petea berbeda nyata dengan di Pantai Kupu-kupu, Pantai Salonsa-A, P. Otuno II-A, dan Pantai Salonsa-B. Padatan tersuspensi total dan padatan terlarut total di daerah S. Petea paling tinggi dibandingkan dengan yang terdapat di lokasi-lokasi pengamatan lain. Hal ini mungkin berhubungan dengan letak S. Petea di bagian timur danau, tepatnya pada out-let danau.
berkaitan dengan aktivitas ikan dalam mencari makan dan pemijahan. Ikan-ikan di S. Petea tidak menggunakan daerah ini sebagai lokasi memijah karena tidak terdapat habitat untuk memijah di lokasi ini. Kecerahan paling rendah di S. Petea (8,60 – 13,10 m) karena sungai ini merupakan out-let danau; semua materi yang keluar danau harus melalui sungai ini. Sungai ini berarus kuat yang dapat membawa materi keluar danau.
Lokasi pemijahan ikan di S. Lawa yang kandungan oksigen terlarutnya tinggi mempunyai ikan-ikan yang panjang bakunya paling tinggi dibandingkan dengan lokasi lain. Hal sebaliknya tampak di lokasi S. Petea yang mempunyai ikan-ikan dengan panjang baku yang rendah. Oksigen terlarut di S. Petea paling rendah dibandingkan dengan lokasi lain. Dalam kasus ini diduga oksigen berperan dalam proses pertumbuhan ikan. Menurut Tom (1998), oksigen penting dalam proses respirasi dan metabolisme ikan. Laju metabolisme dipengaruhi oleh konsentrasi oksigen di dalam air. Jika konsentrasi oksigen terlarut menurun, aktivitas respirasi dan makan juga berkurang. Akibatnya laju pertumbuhan menurun dan kemungkinan serangan penyakit meningkat.
Tabel 4 Kisaran parameter fisik kimiawi perairan pada setiap bulan di Danau Matano
Sep '08 27,50-30,10 10,60-16,00 5,65-6,94 8,22-8,82 0,30-3,90 84,00-146,00 Okt '08 26,80-30,08 9,85-21,10 5,65-7,22 8,36-8,79 0,40-2,40 80,00-125,00 Nov '08 28,30-30,30 9,45-19,15 5,46-6,93 8,33-8,74 0,40-2,40 80,00-133,00 Des '08 27,60-30,20 11,75-16,50 5,18-7,22 8,29-8,67 0,40-2,40 80,00-140,00 Jan '09 26,70-30,50 10,20-18,55 5,18-7,45 8,22-8,78 0,50-1,50 90,00-138,00 Feb '09 28,00-30,20 12,40-20,30 4,93-6,75 8,35-8,58 0,40-2,60 80,00-140,00 Mar '09 27,40-30,30 10,35-17,00 5,02-7,02 8,34-8,79 0,40-2,10 80,00-134,00 Apr '09 27,30-29,60 8,60-19,80 5,02-7,03 8,25-8,72 0,40-2,10 70,00-134,00 Mei '09 28,30-30,20 12,10-19,00 5,43-7,23 8,44-8,75 0,20-1,10 80,00-154,00 Jun '09 27,30-30,30 10,50-17,10 5,22-6,93 8,54-8,87 0,40-1,50 90,00-132,00 Jul '09 27,70-30,80 11,10-18,40 5,35-7,43 8,54-8,72 0,50-1,40 90,00-143,00 Agu '09 28,40-30,30 10,15-19,35 5,02-7,35 8,49-8,75 0,40-2,10 80,00-145,00
secara temporal disajikan dalam Tabel 4 dan Gambar 6, dan dideskripsikan sebagai berikut.
Gambar 6 Fluktuasi parameter fisik kimiawi perairan secara temporal di Danau Matano
menjadi lebih hangat. Selain itu di lokasi sampling tidak terdapat naungan; daerah ini merupakan daerah perairan terbuka. Uji rata-rata dengan menggunakan one way Anova dengan fasilitas Minitab versi 14 pada tingkat kepercayaan 95% diperoleh hasil beda nyata antar waktu sampling (p < 0,05). Suhu pada bulan September 2008 berbeda nyata dengan suhu bulan November 2008, Januari 2009, Juli 2009 dan Agustus 2009. Sementara itu suhu bulan Juli 2009 berbeda nyata dengan suhu rata-rata bulan September 2008, Oktober 2008 dan Maret 2009.
Kandungan oksigen terlarut rata-rata perairan dengan nilai terendah pada bulan Juni 2009 yaitu 6,29 mg l-1 (± 0,58) dan tertinggi pada bulan Mei 2009 yaitu 6,37 mg l-1 (± 0,57). Oksigen terlarut rata-rata selama waktu penelitian September 2008 – Agustus 2009 tidak berbeda nyata antar waktu sampling.
Nilai pH perairan danau terendah pada bulan September dan Januari yaitu 8,22 dan tertinggi pada bulan Juni 2009 yaitu 8,87. Nilai pH perairan relatif tidak bervariasi jauh antar bulan selama penelitian.
Kandungan padatan tersuspensi total rata-rata perairan danau terendah pada bulan Mei 2009 yaitu 0,59 mg l-1 (± 0,24) dan tertinggi pada bulan September 2008 yaitu 1,27 mg l-1 (± 1,02). Padatan tersuspensi total rata-rata bulan Mei 2009 berbeda nyata dengan kandungan rata-rata bulan September dan Oktober 2008.
Kandungan padatan terlarut total rata-rata adalah terendah pada bulan Februari 2009 yaitu 100,20 mg l-1 (± 19,50) dan tertinggi pada bulan Juni 2009 yaitu 118,13 mg l-1 (± 15,25). Padatan terlarut total rata-rata setiap bulan selama penelitian tidak berbeda nyata.
Puncak musim pemijahan ikan T. sarasinorum terjadi pada akhir musim kemarau dan awal musim hujan (Bab 4). Pada waktu-waktu tersebut suhu cenderung meningkat menjadi lebih hangat daripada bulan-bulan sebelumnya.
S. P
Gambar 7 Pengelompokan habitat pemijahan ikan T. sarasinorum di Danau Matano
Kondisi fisik kimiawi perairan di semua lokasi sampling tidak bervariasi jauh, sehingga pengelompokan lokasi sampling dilakukan berdasarkan tingkat kesamaan karakter substrat, luas arena pemijahan, jarak dari tepian danau dan kedalaman arena di masing-masing lokasi (Gambar 7). Hasil pengelompokan lokasi diperoleh tiga kelompok lokasi yang memiliki kesamaan karakter substrat pada selang kepercayaan 85%. Kelompok pertama (S. Lawa, Desa Matano, Paku, Pantai Kupu-kupu, Pantai Salonsa A, Pantai Salonsa B, Old Camp, S. Tanah Merah, P. Otuno I-A, P. Otuno I-B, P. Otuno II-A, P. Otuno II-B, S. Soluro), kelompok kedua (Sokoio), dan kelompok ketiga (S. Petea). Kualitas perairan di lokasi-lokasi tersebut tidak berbeda jauh.
batu besar dengan dinding batuan dasar. Lokasi dalam kelompok kedua ini tidak mempunyai pasir; sehingga tidak ada arena pemijahan ikan. Kelompok ketiga mempunyai substrat dasar dengan kandungan lumpur tinggi, dan tidak terdapar arena pemijahan. Dengan demikian, dapat dikatakan bahwa substrat dasar di lokasi menentukan perbedaan setiap lokasi.
Efektivitas percampuran vertikal di dalam kolom air juga dikonfirmasi terjadi pada pH dalam kisaran basa (7,4) dan konduktivitas tidak berubah menurut kedalaman (Haffner et al. 2001). Jumlah fitoplankton sangat kecil (< 10 sel ml-1) dan kelimpahan spesies tidak berubah di dalam kolom air. Kedalaman eufotik adalah 40 m dan 32 m pada tahun 1993 dan 1995. Apabila sel-sel fitoplankton tercampur merata di dalam zona eufotik adalah mungkin bahwa percampuran vertikal membatasi produksi ototrofik. Tetapi pengamatan mikroskopis menunjukkan bahwa detritus C lebih berlimpah daripada biomassa alga, sehingga adalah mungkin bahwa jaring makanan Danau Matano disebabkan oleh masukan detritus organik yang berasal dari sekeliling danau.
Profil oksigen menunjukkan profil fotosintesis lemah hingga ke bawah kira-kira 40 m, dan setelah kedalaman itu oksigen berkurang hingga di bawah 1% kejenuhan. Danau Matano adalah oligotrofik dalam hal jumlah total nitrogen dan total fosfor di atas 100 m dari kolom air. Konsentrasi nitrogen dan fosfor sangat rendah dan sering tidak terdeteksi. Tingginya kecerahan perairan menunjukkan sistem dengan sangat sedikit bahan biotik dan abiotik tersuspensi. Profil oksigen menunjukkan kurva fotosintetik yang meluas ke bawah hingga 40 m, sama dengan prediksi kedalaman eufotik (Crowe et al. 2008). Setelah 40 m konsentrasi oksigen berkurang; penurunan ini merupakan respon terhadap laju respirasi mikroba yang tinggi yang berhubungan dengan metabolisme bahan organik yang mengalami sedimentasi.
Selain itu, sel-sel alga yang tertinggal kecil kemungkinannya untuk kembali ke lapisan epilimnion.
Faktor-faktor fisik seperti ketersediaan cahaya dan percampuran bukan merupakan faktor pembatas, walaupun sedikit pertukaran nutrien melalui lapisan termal yang dalam bisa berperan penting. Sedimentasi bahan biologis dan geologis dapat secara terus-menerus menurunkan cadangan nutrien pada permukaan air bagian atas.
Hampir semua jenis aktivitas (pertanian, urbanisasi, komunikasi, pertambangan, industri) menyebabkan peningkatan materi tersuspensi di permukaan perairan, terutama disebabkan oleh deforestasi (Meybeck et al. 1989). Salah satunya adalah perubahan siklus hidrologis yang mengubah intensitas dan kemampuan percampuran badan air dan keseimbangan hidrologis. Namun demikian hasil pengukuran parameter kualitas air di lokasi-lokasi sampling menunjukkan tidak terdapat perubahan nilai yang mencolok antar bulan. Dengan demikian dapat dipandang bahwa kualitas perairan saat ini tidak menyebabkan perubahan keberadaan ikan; jumlah ikan pada setiap bulan sampling tidak banyak bervariasi (Bab 3).
Kesimpulan
Habitat pemijahan ikan T. sarasinorum tersebar di sepanjang daerah litoral danau. Terdapat dua tipe habitat pemijahan dalam penelitian ini yaitu habitat batu berpasir dan habitat perakaran. Dari 12 lokasi penelitian ditemukan tiga lokasi yang mempunyai dua tipe habitat pemijahan (habitat batu berpasir dan habitat perakaran) yaitu Pantai Salonsa, P. Otuno I, dan P. Otuno II. Lokasi lainnya hanya memiliki satu tipe habitat, yaitu habitat batu berpasir. Tempat pemijahan pada setiap habitat berbentuk arena. Arena ini secara spesifik berada pada daerah yang mendapat naungan bayang-bayang yang berasal dari pohon di tepian danau dan atau dari batu-batu besar yang ada di sekitarnya.
3 SEBARAN SPASIAL-TEMPORAL IKAN
T. sarasinorum
DI DANAU MATANO
Pendahuluan
Sebaran ikan T. sarasinorum di Danau Matano pertama kali dilaporkan oleh Kottelat (1991). Hingga saat ini diketahui terdapat sembilan jenis ikan yang termasuk dalam famili Telmatherinidae yang hidup di Danau Matano. Ikan T. sarasinorum adalah salah satunya yang diketahui endemik Danau Matano dan salah satu spesies yang dominan. Menurut Kottelat (1991), ikan ini memiliki penyebaran yang terbatas di daerah litoral sepanjang tepian danau dan lebih sering ditemukan di bagian selatan dari danau. Ikan T. sarasinorum umumnya menyebar di sisi selatan danau yaitu di dekat pulau-pulau sebelah timur danau sampai ke bagian barat danau. Ikan ini umumnya menempati habitat perairan dengan dasar berpasir yang hampir datar, pada perairan yang relatif dangkal (<1,5m).
Kondisi fisik kimiawi lingkungan dan ketersediaan makanan serta tempat berlindung merupakan faktor utama yang menentukan sebaran ikan (Mazzoni & Iglesias-Rios 2002). Pemanfaatan habitat oleh spesies ikan berhubungan dengan karakter morfologi dan fisiologi termasuk ukuran, dan bentuk serta posisi sirip-sirip (Wootton 1990) serta kondisi trofik dan kebutuhan reproduksi (Balon 1975). Kejadian dan kelimpahan spesies ikan di dalam sistem Ubatiba berhubungan dengan trofik, yaitu pemilihan mikrohabitat sehingga memaksimalkan kemampuan mereka untuk memanfaatkan secara efisien suplay makanan dan persyaratan anti-predator. Sebaran suatu spesies ikan berhubungan dengan faktor-faktor yang berkaitan dengan habitat makan, reproduksi dan perlindungan dari predator. Sebaran juga dipengaruhi oleh musim.