PENGEMBANGAN PADI TOLERAN SALINITAS
MELALUI KULTUR ANTERA
HENI SAFITRI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa disertasi berjudul “Pengembangan Padi Toleran Salinitas melalui Kultur Antera” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2016
Heni Safitri
RINGKASAN
HENI SAFITRI. Pengembangan Padi Toleran Salinitas melalui Kultur Antera. Dibimbing oleh BAMBANG SAPTA PURWOKO, SINTHO WAHYUNING ARDIE dan ISWARI SARASWATI DEWI.
Kebutuhan beras di Indonesia terus meningkat seiring dengan meningkatnya jumlah penduduk, sehingga peningkatan produksi padi harus terus dilakukan untuk mendukung ketahanan pangan nasional. Berkurangnya lahan sawah irigasi yang merupakan pendukung utama produksi beras nasional ditambah dengan perubahan iklim global merupakan kendala dalam peningkatan produksi padi. Perubahan iklim dunia yang menyebabkan pemanasan global berakibat pada intrusi air laut ke daratan yang memicu salinitas di sepanjang lahan di daerah pantai.
Usaha meningkatkan produksi beras nasional dapat dilakukan melalui berbagai pendekatan antara lain dengan pemanfaatan lahan marginal, khususnya lahan salin di sepanjang pantai dan dengan penggunaan varietas unggul yang berdaya hasil tinggi dan toleran salinitas. Potensi lahan salin di Indonesia yang cukup luas dapat dimanfaatkan untuk penanaman padi, mengingat tanaman padi merupakan salah satu tanaman serealia yang potensial dibudidayakan pada lahan salin karena kemampuannya untuk hidup di lahan tergenang.
Perakitan varietas unggul secara konvensional memerlukan waktu yang cukup lama, terutama dalam menghasilkan galur-galur murni yang homozigos dan proses seleksi. Penggunaan kultur antera yang dikombinasikan dengan uji cepat dalam penapisan genotipe padi terhadap salinitas pada fase bibit diharapkan dapat mempersingkat siklus pemuliaan dalam menghasilkan varietas unggul baru toleran salinitas. Tujuan penelitian ini ialah untuk mendapatkan galur-galur padi dihaploid yang homozigos toleran salinitas melalui kultur antera, penapisan pada fase bibit dalam media hidroponik, penapisan di tanah salin dan evaluasi galur-galur dihaploid di lapangan.
Studi toleransi beberapa plasma nutfah padi terhadap salinitas pada fase bibit dengan menggunakan media hidroponik yang mengandung 120 mM NaCl mengidentifikasi lima genotipe toleran (skor 3) yaitu Dendang, Inpara 5, Inpari 29, IR77674 dan IR81493; lima genotipe moderat toleran (skor 5) yaitu Cilamaya Muncul, Inpari 30, IR64, IR78788 dan Siak Raya; dua genotipe peka (skor 7) yaitu Banyuasin dan Mendawak; dan satu genotipe sangat peka (skor 9) yaitu Inpara 4. Inpara 4 merupakan genotipe sangat peka, sebanding dengan IR29, sehingga Inpara 4 dapat digunakan sebagai verietas pembanding peka pada penelitian-penelitian selanjutnya. Penentuan toleransi suatu genotipe terhadap salinitas dengan menggunakan skoring visual berdasarkan kerusakan daun pada bibit tanaman lebih dapat membedakan antara genotipe toleran dan peka dibandingkan dengan apabila penentuan toleransi menggunakan karakter agronomi bibit lainnya.
mekanisme toleransi terhadap salinitas yang bersifat eksklusi, sedangkan Dendang dan Pokkali mempunyai mekanisme inklusi.
Pembentukan populasi F1 dengan melakukan persilangan antara tetua padi berdaya hasil tinggi dengan tetua toleran salinitas menghasilkan 12 kombinasi persilangan yang terdiri atas enam persilangan antara tetua toleran/toleran dan enam persilangan antara tetua moderat/toleran. Tanaman F1 digunakan sebagai sumber antera dalam pembentukan galur-galur dihaploid padi melalui kultur antera. Teknik kultur antera telah menghasilkan 125 galur dihaploid, 60 galur (43%) diantaranya diuji lebih lanjut karakter agronomi dan toleransinya terhadap salinitas.
Penapisan 60 galur padi dihaploid terhadap salinitas di kultur hara (hidroponik) yang mengandung 120 mM NaCl pada fase bibit menghasilkan 31 galur toleran (skor 3), 25 galur moderat toleran (skor 5) dan 4 galur peka (skor 7); sedangkan penapisan 60 galur padi dihaploid di tanah salin (40 mM NaCl) pada umur tanaman 60 hari menghasilkan 12 galur toleran (skor 3), 6 galur moderat toleran (skor 5), 14 galur peka (skor 7) dan 28 galur sangat peka (skor 9). Korelasi antara skor toleransi genotipe-genotipe padi pada media hidroponik dan di tanah salin adalah sangat signifikan positif. Skor toleransi suatu genotipe di tanah salin dapat 1-2 level lebih besar dibandingkan dengan skor toleransi suatu genotipe di media hidroponik. Dengan demikian, penapisan galur-galur padi pada media hidroponik pada fase bibit dapat digunakan sebagai dasar pemilihan galur, dimana galur yang tergolong toleran dapat diuji lebih lanjut pada pengujian galur-galur padi di tanah atau lahan salin pada fase reproduktif.
Evaluasi karakter agronomi dan daya hasil galur-galur padi dihaploid di lahan sawah irigasi non-salin menghasilkan 21 galur padi berpotensi hasil tinggi (>8 ton ha-1), sebanding dan/atau lebih tinggi dibandingkan dengan Ciherang (8.1 ton ha-1). Berdasarkan percobaan ini terpilih 32 galur yang diteruskan pada uji daya hasil pendahuluan (UDHP).
Uji daya hasil pendahuluan terhadap 32 galur dihaploid dan 3 varietas pembanding mengalami kendala kekeringan di lapangan dimulai dari 8 minggu setelah tanam sehingga mengakibatkan penurunan pada penampilan agronomi dan hasil gabah. Rata-rata hasil gabah galur-galur dihaploid pada UDHP sebanding dengan Ciherang (3.4 ton ha-1). Tiga galur yaitu HS14-15-1-2, HS17-3-1-3 dan HS17-21-1-5 adalah galur toleran salinitas berdasarkan skor kerusakan daun pada uji pot di tanah salin (EC = 6.2 dS m-1). Galur-galur dihaploid ini perlu diuji kembali di lahan optimal dan lahan sub-optimal (lahan salin).
SUMMARY
HENI SAFITRI. Development of Salinity Tolerant Rice through Anther Culture. Supervised by BAMBANG SAPTA PURWOKO as chairman, SINTHO WAHYUNING ARDIE and ISWARI SARASWATI DEWI as members of the advisory committee.
Rice demand in Indonesia increased along with population growth. Rice production should be increased to support national food security. Global climate change and reduction in irrigated land, which is the main contributor of national rice production, are the limiting factors in increasing rice production. Global climate change caused global warming, resulted in sea water intrusion, triggering salinity along the land in coastal areas.
The effort to increase national rice production can be done through various approaches among others by cultivation of marginal lands, particularly saline area along the coast and the utilization of high yielding varieties tolerant to salinity. Saline land potential in Indonesia is widespread and can be used for rice production, as rice is a cereal crop that can grow on saline land because of its ability to grow in waterlogged area.
Conventional rice breeding requires considerable amount of time, especially in the selection process and in producing homozygous pure lines. The use of anther culture combined with rapid evaluation in screening rice genotypes to salinity tolerance at seedling phase is expected to shorten the breeding cycle for producing new salinity tolerant varieties. The purpose of this study was to obtain haploid rice lines tolerant to salinity through anther culture, to screen the doubled haploid lines in hydroponic media and saline soil, and to evaluate the doubled haploid rice lines in the field.
The study of tolerance to salinity at seedling phase using hydroponic media containing 120 mM NaCl resulted in five tolerant genotypes (score 3), namely Dendang, Inpara 5, Inpari 29, IR77674 and IR81493; five moderate tolerant genotypes (score 5) namely Cilamaya, Inpari 30, IR64, IR78788 and Siak Raya; two sensitive genotypes (score 7) namely Banyuasin and Mendawak; and a highly sensitive genotype (score 9) namely Inpara 4. Inpara 4 was highly sensitive genotype, similar to IR29. Inpara 4 can be used as a sensitive check variety in future studies. The determination of a genotype tolerant to salinity using visual score based on leaf damage of the seedling could distinguish between tolerant and sensitive genotypes compared to the use of other seedling characters.
The morpho-physiological study of some rice genotypes at several NaCl concentrations provides a screening method for salinity tolerance of rice in saline soils by using pot experiment containing soil and water (ratio 7: 3) and 40 mM NaCl (6.2 dS m-1). The result showed that the tolerant genotypes IR77674 and IR81493 have the exclusion mechanisms of salinity tolerance, while Dendang and Pokkali have the inclusion mechanisms.
through anther culture. From anther culture technique 125 doubled haploid lines were produced, 60 lines (43%) were further tested for agronomic characters and salinity tolerance.
The screening of 60 doubled haploid lines to salinity tolerance in nutrient culture (hydroponics) containing 120 mM NaCl at seedling phase resulted in 31 tolerant lines (score 3), 25 moderate tolerant lines (score 5) and 4 sensitive lines (score 7); while the screening of 60 doubled haploid lines to salinity tolerance in saline soils (40 mM NaCl) at the 60 day-old plant resulted in 12 tolerant lines (score 3), 6 moderate tolerant lines (score 5), 14 sensitive lines (score 7) and 28 highly sensitive lines (score 9 ). The correlation between the tolerance scores of the genotypes in hydroponic media and in saline soils was very significantly positive. The tolerance score of the genotype in saline soils could be 1-2 level higher than the scores of the genotype in hydroponics media. Therefore, the screening of rice lines in hydroponic media on seedling phase can be used as the basis for selecting the lines which were classified as tolerant to be further tested in the salinity tolerance screening of the rice lines in soil or saline land in the reproductive phase.
Evaluation of agronomic and yield characters of 60 doubled haploid rice lines in irrigated non-saline land produced 21 high yielding rice lines (> 8 ton ha-1 of yield potential), equal and/or higher than Ciherang (8.1 ton ha-1 of yield potential). This experiment succesfully selected 32 lines for the preliminary yield trials (PYT).
Preliminary yield trials of 32 doubled haploid rice lines and 3 check varieties experienced drought problems in the field starting at 8 weeks after planting, resulting the reduction in agronomic performance and grain yield. The average grain yield of all doubled haploid rice lines on PYT was equal to Ciherang (3.4 ton ha-1). Three lines namely HS14-15-1-2, 3-1-3 and HS17-21-1-5 were salinity tolerant lines based on leaf damage score in the saline soil experiment. The doubled haploid rice lines need to be further tested in the optimal and sub-optimal land (saline condition).
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
PENGEMBANGAN PADI TOLERAN SALINITAS
MELALUI KULTUR ANTERA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Penguji pada Ujian Tertutup: Dr Ir Agus Purwito, MSc Agr Dr Ir Trikoesoemaningtyas, MSc
Penguji pada Ujian Terbuka: Dr Buang Abdullah
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala nikmat dan karunia-Nya sehingga disertasi ini berhasil penulis selesaikan. Tujuan penelitian ini adalah mendapatkan galur-galur padi dihaploid hasil kultur antera yang toleran terhadap cekaman salinitas.
Terima kasih penulis ucapkan kepada Prof Dr Ir Bambang Sapta Purwoko, MSc, Dr Ir Iswari Saraswati Dewi dan Dr Sintho Wahyuning Ardie SP MSi selaku pembimbing yang banyak memberi arahan, saran dan tambahan wawasan kepada penulis selama penelitian dan dalam menyelesaikan penulisan disertasi. Terima kasih juga penulis ucapkan kepada Dr Ir Yudiwanti Wahyu EK, MS selaku koordinator Mayor Pemuliaan dan Bioteknologi Tanaman, Dr Ir Trikoesoemaningtyas, MSc selaku penguji pada ujian prakualifikasi lisan, ujian tertutup dan sidang promosi doktor, Dr Dewi Sukma SP MSi selaku penguji pada ujian prakualifikasi lisan, Dr Ir Agus Purwito MSc Agr selaku penguji luar komisi pada ujian tertutup, dan Dr Buang Abdullah selaku penguji luar komisi pada sidang promosi doktor.
Penulis juga menyampaikan terima kasih kepada Badan Litbang Pertanian yang telah memberikan kesempatan dan beasiswa pendidikan, Program Kerja sama Kemitraan Penelitian dan Pengembangan Pertanian Nasional (KKP3N) yang telah memberikan dana penelitian, staf dan teknisi BB Biogen (Pak Iman, Pak Inan, Pak Kohar, Yeni, Deny, dan Pak Hery), staf dan teknisi KP Padi Muara Bogor (Pak Sularjo, Pak Supartopo, Cahyono, Erna Herlina, Yusuf, Oma, Pak Darno (alm) dan Pak Indarjo), staf dan teknisi BB Padi Sukamandi dan IPB yang telah membantu pelaksanaan penelitian.
Terima kasih juga penulis sampaikan kepada teman-teman PBT 2011, Fitri Rachmawati, Anneke Pesik, Angelita P Lestari, Rini Hermanasari, Marlina M, dan Agus Zainuddin atas dukungan semangat dan persahabatan selama studi. Terima kasih kepada Dr Sri Suhesti, Dr Redy dan Dr Ismail Maskromo atas dukungan, saran dan masukan dalam penyusunan disertasi. Terima kasih kepada Dr Indrastuti R, Yuni Widyastuti MSi, Cucu Gunarsih MSi, Yullianida MSi dan semua teman-teman yang telah mendukung saya yang tidak bisa saya sebutkan satu persatu.
Penulis menyampaikan rasa hormat, terima kasih dan penghargaan kepada Bapak dan Ibu sebagai orang tua yang telah menanamkan dasar pendidikan yang baik dan berguna bagi penulis, seluruh keluarga besar atas segala doa dan dukungannya. Secara khusus penulis menyampaikan terima kasih kepada suami tercinta Edy Suwarna ST dan kedua anak kami tersayang, Naura Azizah dan Naufal Fauzi atas segala pengertian, doa, motivasi, bantuan, pengorbanan dan kesabarannya dalam mendampingi penulis selama ini. Terima kasih juga kepada bibi yang sudah menjaga dan mengasuh anak-anak sehingga penulis dapat beraktivitas dengan baik.
Akhirnya kepada semua pihak yang turut membantu selama penelitian hingga penulisan disertasi ini, penulis ucapkan terima kasih. Semoga disertasi ini bermanfaat.
Bogor, Agustus 2016
DAFTAR ISI
Karakteristik Tanaman Padi 7
Cekaman Salinitas pada Tanaman Padi 9
Kultur Antera dalam Pemuliaan Tanaman Padi 12
3 PENAPISAN PLASMA NUTFAH PADI TERHADAP
SALINITAS PADA FASE BIBIT 15
Abstrak 15
4 RESPON MORFO-FISIOLOGI BEBERAPA GENOTIPE PADI
PADA BEBERAPA KONSENTRASI NaCl 31
Abstrak 31 PADI HASIL PERSILANGAN TETUA BERDAYA HASIL
TINGGI DENGAN TETUA TOLERAN SALINITAS 45
Abstrak 45
6 PENAPISAN GALUR-GALUR PADI DIHAPLOID TERHADAP
SALINITAS PADA FASE BIBIT 59
Abstrak 59
Pendahuluan 61
Bahan dan Metode 62
Hasil dan Pembahasan 64
Simpulan 81
7 PENAPISAN GALUR-GALUR PADI DIHAPLOID TERHADAP
SALINITAS DI TANAH SALIN 83
Abstrak 83
Abstract 84
Pendahuluan 85
Bahan dan Metode 85
Hasil dan Pembahasan 86
Simpulan 93
8 EVALUASI KARAKTER AGRONOMI DAN DAYA HASIL
GALUR-GALUR PADI DIHAPLOID DI LAHAN NON-SALIN 95
Abstrak 95
Abstract 96
Pendahuluan 97
Bahan dan Metode 97
Hasil dan Pembahasan 98
Simpulan 104
9 UJI DAYA HASIL PENDAHULUAN GALUR-GALUR PADI
DIHAPLOID 105
Abstrak 105
Abstract 106
Pendahuluan 107
Bahan dan Metode 107
Hasil dan Pembahasan 108
Simpulan 113
10 PEMBAHASAN UMUM 115
11 SIMPULAN DAN SARAN 123
DAFTAR PUSTAKA 125
LAMPIRAN 133
DAFTAR TABEL
3.1 Materi genetik padi yang digunakan untuk penapisan terhadap
salinitas 18
3.2 Urutan kegiatan penapisan plasma nutfah padi terhadap salinitas
melalui uji deteksi dini pada fase bibit 19
3.3 Kriteria evaluasi terhadap cekaman salinitas pada fase bibit 19 3.4 Skor kerusakan daun padi beberapa plasma nutfah padi terhadap
salinitas pada fase bibit 20
3.5 Tinggi bibit dan penurunan tinggi bibit akibat salinitas pada
beberapa genotipe padi 22
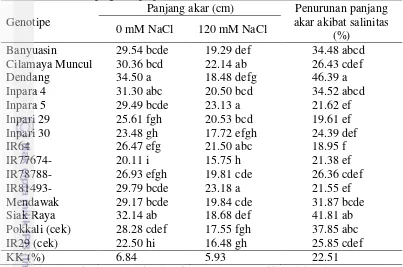
3.6 Panjang akar dan penurunan panjang akar akibat salinitas pada
beberapa genotipe padi 23
3.7 Jumlah anakan dan penurunan jumlah anakan akibat salinitas
pada beberapa genotipe padi 23
3.8 Jumlah daun dan penurunan jumlah daun akibat salinitas pada
beberapa genotipe padi 24
3.9 Bobot basah tajuk dan penurunan bobot basah tajuk akibat
salinitas pada beberapa genotipe padi 25
3.10 Bobot basah akar dan penurunan bobot basah akar akibat salinitas
pada beberapa genotipe padi 26
3.11 Bobot basah biomassa dan penurunan bobot basah biomassa
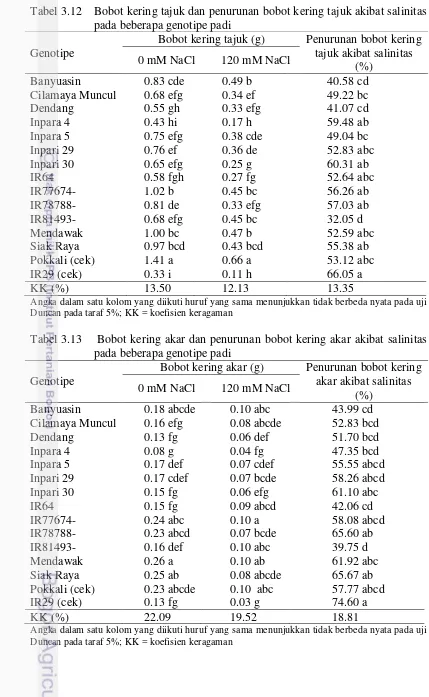
akibat salinitas pada beberapa genotipe padi 26 3.12 Bobot kering tajuk dan penurunan bobot kering tajuk akibat
salinitas pada beberapa genotipe padi 27
3.13 Bobot kering akar dan penurunan bobot kering akar akibat
salinitas pada beberapa genotipe padi 27
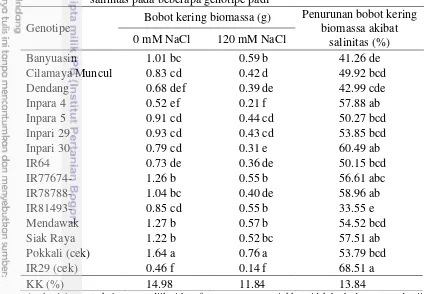
3.14 Bobot kering biomassa dan penurunan bobot kering biomassa
akibat salinitas pada beberapa genotipe padi 28 3.15 Koefisien korelasi Pearson antar karakter pada penapisan
beberapa genotipe padi pada fase bibit 29
4.1 Kuadrat tengah karakter agronomi dan hasil genotipe-genotipe
padi pada beberapa konsentrasi NaCl 35
4.2 Rata-rata hasil gabah per tanaman enam genotipe padi pada
beberapa konsentrasi salinitas 39
4.3 Kuadrat tengah karakter fisiologi enam genotipe padi pada
beberapa konsentrasi salinitas 40
5.1 Materi genetik padi yang digunakan sebagai tetua persilangan 48 5.2 F1 hasil persilangan antara tetua toleran salinitas dan varietas
unggul 51
5.3 Hasil induksi kalus beberapa persilangan padi untuk toleransi
terhadap salinitas 52
5.4 Hasil regenerasi tanaman beberapa persilangan padi untuk
toleransi terhadap salinitas 54
5.5 Efisiensi pembentukan kalus dan tanaman hijau pada kultur antera
5.6 Aklimatisasi dan tanaman dihaploid putatif yang dihasilkan dari kultur antera beberapa persilangan padi untuk pengembangan padi
toleran salinitas 56
6.1 Kriteria evaluasi terhadap cekaman salinitas pada fase bibit 63 6.2 Nilai tengah dan kisaran populasi beberapa karakter agronomi
bibit galur-galur padi dihaploid dan pembandingnya 64 6.3 Skor visual toleransi 60 galur dihaploid padi terhadap salinitas
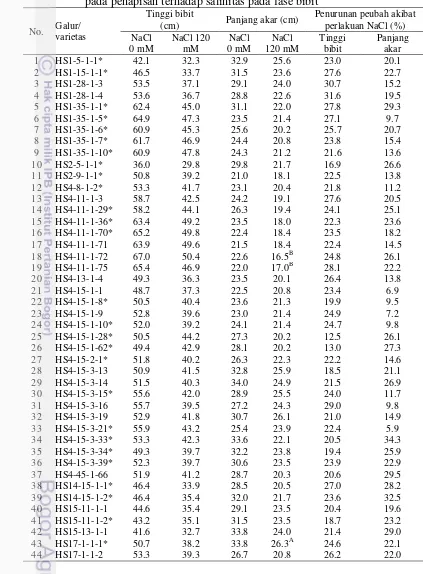
pada fase bibit 66
6.4 Tinggi bibit dan panjang akar galur-galur dihaploid yang diuji
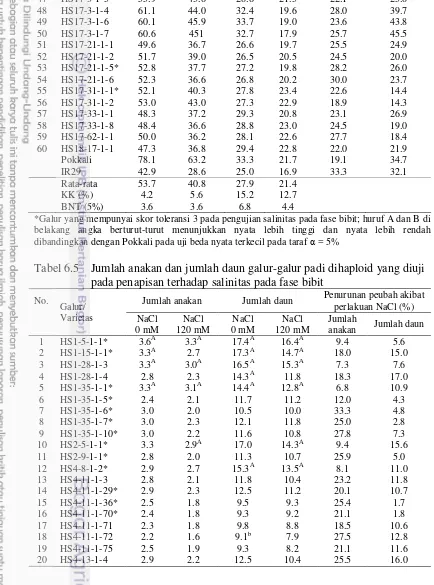
pada penapisan terhadap salinitas pada fase bibit 67 6.5 Jumlah anakan dan jumlah daun galur-galur dihaploid yang diuji
pada penapisan terhadap salinitas pada fase bibit 68 6.6 Bobot basah tajuk dan bobot basah akar galur-galur dihaploid
yang diuji pada penapisan terhadap salinitas pada fase bibit 70 6.7 Bobot kering tajuk dan bobot kering akar galur-galur dihaploid
yang diuji pada penapisan terhadap salinitas pada fase bibit 72 6.8 Bobot basah dan bobot kering biomassa galur-galur dihaploid
yang diuji pada penapisan terhadap salinitas pada fase bibit 73 6.9 Galur-galur padi dihaploid toleran salinitas pada fase bibit (skor
3) 75
6.10 Nilai komponen ragam karakter agronomi bibit pada kondisi
non-salin dan non-salin 76
6.11 Nilai duga heritabilitas arti luas karakter agronomi bibit
galur-galur padi dihaploid pada kondisi non-salin dan salin 77 6.12 Indeks sensitivitas galur-galur padi dihaploid terhadap cekaman
salinitas untuk karakter tinggi tanaman, panjang akar, jumlah
anakan, jumlah daun, bobot basah tajuk dan bobot kering tajuk 78 6.13 Indeks sensitivitas galur-galur padi dihaploid terhadap cekaman
salinitas untuk karakter bobot basar akar, bobot kering akar, bobot
basah biomassa dan bobot kering biomassa 79
6.14 Koefisien korelasi antara skor visual kerusakan daun dengan
indeks sensitivitas karakter agronomi bibit padi 81 7.1 Skor visual toleransi 60 galur padi dihaploid terhadap salinitas di
tanah salin 87
7.2 Tinggi tanaman dan jumlah anakan produktif galur-galur dihaploid yang diuji pada penapisan terhadap salinitas di tanah
salin 88
7.3 Jumlah gabah per malai galur-galur padi dihaploid yang diuji
pada penapisan terhadap salinitas di tanah salin 90 7.4 Hasil gabah per tanaman galur-galur padi dihaploid yang diuji
pada penapisan terhadap salinitas di tanah salin 92 8.1 Karakter agronomi galur-galur padi dihaploid pada sawah irigasi
di Cianjur, Jawa Barat, tahun 2015 101
8.2 Karakter hasil dan potensi hasil galur-galur padi dihaploid pada
sawah irigasi di Cianjur, Jawa Barat, tahun 2015 102 8.3 Nilai diferensial seleksi karakter agronomi galur-galur padi
9.1 Karakter agronomi galur-galur padi dihaploid pada uji daya hasil
pendahuluan di sawah irigasi di Sukabumi, Jawa Barat 2015 111 9.2 Komponen hasil dan hasil galur-galur padi dihaploid pada uji
daya hasil pendahuluan di sawah irigasi di Sukabumi, Jawa Barat
2015 112
10.1 Skor visual toleransi 60 galur padi dihaploid terhadap salinitas pada media hidroponik (fase bibit) pada 120 mM NaCl dan di
tanah salin (60 HST) pada 40 mM NaCl 118
10.2 Koefisien korelasi Pearson skor toleransi salinitas padi pada
media hidroponik (fase bibit) dan di tanah salin (60 HST) 118 10.3 Galur-galur padi dihaploid yang toleran dan moderat terhadap
DAFTAR GAMBAR
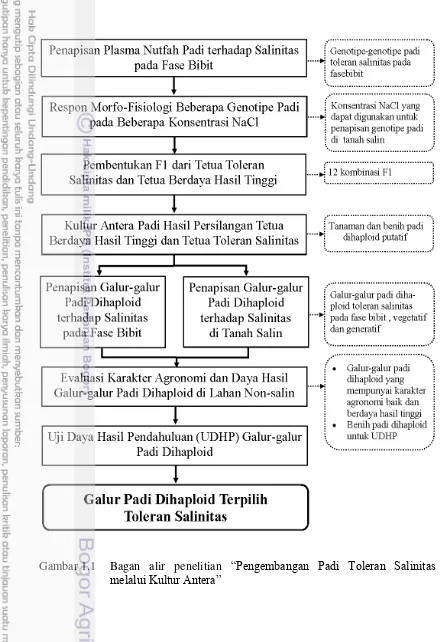
1.1 Bagan alir penelitian “Pengembangan Padi Toleran Salinitas
melalui Kultur Antera” 4
3.1 Penampilan genotipe toleran (A), moderat toleran (B), peka (C),
dan sangat peka (D) pada fase bibit 21
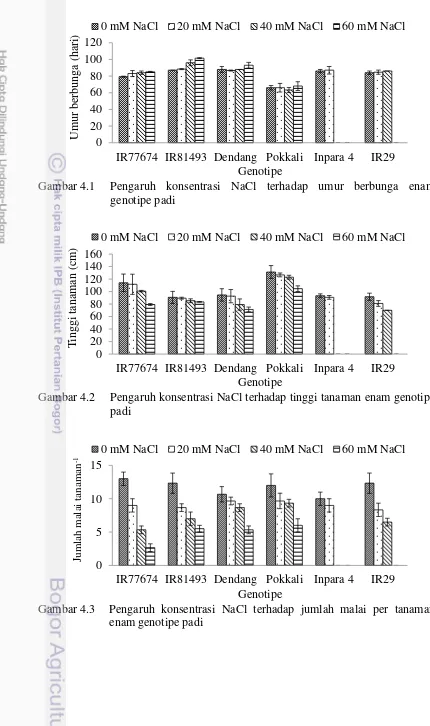
4.1 Pengaruh konsentrasi NaCl terhadap umur berbunga enam
genotipe padi 37
4.2 Pengaruh konsentrasi NaCl terhadap tinggi tanaman enam
genotipe padi 37
4.3 Pengaruh konsentrasi NaCl terhadap jumlah malai per tanaman
enam genotipe padi 37
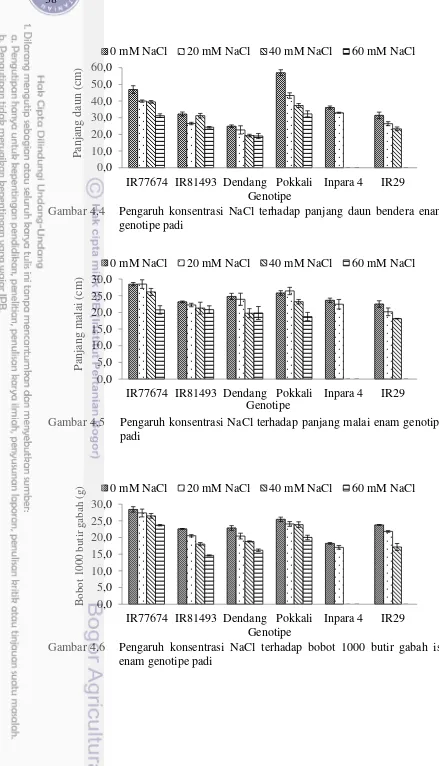
4.4 Pengaruh konsentrasi NaCl terhadap panjang daun bendera enam
genotipe padi 38
4.5 Pengaruh konsentrasi NaCl terhadap panjang malai enam
genotipe padi 38
4.6 Pengaruh konsentrasi NaCl terhadap bobot 1000 butir gabah isi
enam genotipe padi 38
4.7 Penurunan hasil relatif gabah per tanaman (%) dibandingkan dengan kontrol (0 mM NaCl) enam genotipe padi pada beberapa
konsentrasi salinitas 39
4.8 Pengaruh konsentrasi NaCl terhadap konsentrasi K+ dalam daun
enam genotipe padi 40
4.9 Pengaruh konsentrasi NaCl terhadap konsentrasi Na+ dalam daun
enam genotipe padi 41
4.10 Pengaruh konsentrasi NaCl terhadap konsentrasi Ca2+ dalam daun
enam genotipe padi 43
4.11 Pengaruh konsentrasi NaCl terhadap rasio K+/Na+ enam genotipe
padi 43
4.12 Pengaruh konsentrasi NaCl terhadap rasio Ca2+/Na+ enam
genotipe padi 44
5.1 Urutan kegiatan kultur antera padi mulai dari penanaman eksplan (a), seleksi spikelet (b), sterilisasi (c), pemotongan spikelet dan penanaman antera (d), inkubasi antera (e), regenerasi kalus (f) dan
perakaran (g) 50
5.2 Pembentukan kalus, regenerasi tanaman dan perakaran pada kultur antera padi: (A) kalus yang terbentuk dari antera yang dikulturkan, (B) hasil regenerasi kalus: 1. tanaman albino, 2. kalus yang tidak beregenerasi, dan 3. tanaman hijau, (C) tanaman hijau
pada media perakaran 53
5.3 Tanaman yang dihasilkan dari kultur antera padi: (A) tanaman
dihaploid dan (B) tanaman haploid 56
6.1 Penampilan galur padi dihaploid pada uji toleransi terhadap salinitas fase bibit: toleran (A), moderat toleran (B), peka (C),
Pokkali (D), IR29 (E) 65
7.1 Penampilan galur padi dihaploid toleran salinitas dan varietas pembanding: (A) galur HS1-35-1-5, (B) Pokkali, (C) IR29.
8.1 Penampilan galur-galur padi dihaploid di lahan sawah non-salin di Cianjur: (A) pelaksanaan percobaan di lapangan, dan (B) penampilan galur-galur dihaploid yang berasal dari satu
persilangan, tampak seragam dalam galur dan berbeda antar galur 100 9.1 Kondisi pertanaman padi pada 12 minggu setelah tanam: (A)
tanah kelihatan retak-retak karena kekeringan yang cukup lama,
(B) usaha pengairan yang dilakukan dengan memompa air 108 9.2 Pertanaman UHDP padi yang mendapat pengairan dengan
pompanisasi dan lahan petani yang dibiarkan puso 109 9.3 Penampilan galur-galur padi dihaploid dan varietas Ciherang pada
uji daya hasil pendahuluan di Sukabumi pada 105 HSS 113 10.1 Kondisi percobaan penapisan plasma nurfah padi terhadap
salinitas di lapangan: (A) lokasi Madura pada 65 HST, dan (B)
lokasi Serang, Banten pada 30 HST 116
10.2 Kondisi percobaan uji daya hasil pendahuluan (UDHP) padi di Indramayu, Jawa Barat: (A) saat tanam dan (B) pada saat 4
DAFTAR LAMPIRAN
1 Komposisi kimia larutan Yoshida 135
2 Komposisi kimia media dasar induksi kalus (N6) dan media dasar
regenerasi (MS) pada kultur antera padi 136
3 Deskripsi varietas dan galur padi yang digunakan sebagai tetua dan/atau varietas pembanding pada penelitian “Pengembangan
Padi Toleran Salinitas melalui Kultur Antera” 137 4 Hasil analisis tanah percobaan penapisan genotipe padi di tanah
salin pada perlakuan 0 mM NaCl dan 40 mM NaCl di
1 PENDAHULUAN
Latar Belakang
Padi (Oryza sativa L.) merupakan sumber bahan makanan pokok di Indonesia. Kebutuhan beras dalam negeri terus meningkat seiring dengan meningkatnya jumlah penduduk. Rata-rata konsumsi beras per kapita masyarakat Indonesia adalah 87 kg per tahun (Kementan 2016). Dengan pertambahan jumlah penduduk 1.49% per tahun, maka pada tahun 2020 penduduk Indonesia akan mencapai 275 juta jiwa (BPS 2015), sehingga meningkatkan kebutuhan beras secara signifikan. Luas pertanaman padi di Indonesia pada tahun 2013 yaitu 13.8 juta ha, dengan produksi padi nasional mencapai 71.3 juta ton dan produktivitas 5.3 ton ha-1 (Kementan 2016). Hal ini berarti peningkatan produksi padi harus terus dilakukan untuk mendukung ketahanan pangan nasional.
Perubahan iklim dunia yang menyebabkan pemanasan global (global warming) dalam beberapa dekade ini juga semakin menambah tantangan dalam produksi padi. Berbagai penelitian membuktikan hubungan pemanasan global dengan peningkatan kegaraman (salinitas) lahan (Dailidienė dan Davulien 2008; Sposito 2008). Perubahan iklim dan pengaruh salinitas berpengaruh pada pertumbuhan dan perkembangan produktivitas tanaman pangan di dunia termasuk Indonesia. Pengaruh salinitas pada produksi padi di Indonesia dapat mencapai 50% dari luas lahan sawah yang berada di sepanjang Pantai Utara Jawa (Hariadi et al. 2015). Selama ini sawah irigasi merupakan pendukung produksi beras yang utama. Dalam beberapa tahun terakhir, banyak terjadi konversi lahan sawah irigasi ke tujuan non-pertanian sehingga luas lahan pertanian subur berkurang.
Usaha untuk meningkatkan produksi padi di Indonesia dapat dilakukan dengan memanfaatkan lahan marginal, salah satunya adalah lahan salin. Potensi lahan salin di Indonesia mencapai 440 ribu ha, sebagian besar merupakan lahan pasang surut yang terletak di Sumatera, Jawa, Madura, Sulawesi, Maluku dan Papua (Alihamsyah 2004). Luas lahan salin di Indonesia terus bertambah terutama di daerah pesisir pantai. Selain intrusi air laut yang kejadiannya perlahan-lahan dalam waktu yang lama, salinitas dapat disebabkan secara tidak terduga akibat kejadian alam seperti tsunami yang melanda Aceh tahun 2004 (Subagyono et al. 2005).
Penggunaan varietas unggul toleran salinitas merupakan cara yang paling efisien dalam mengatasi masalah salinitas (Shrivastava dan Kumar 2015). Pengembangan padi di lahan salin terkendala oleh kurangnya ketersediaan varietas unggul toleran salinitas. Oleh karena itu perlu dilakukan penelitian untuk menghasilkan varietas baru yang toleran salinitas. Perakitan padi dengan pemuliaan konvensional melalui persilangan, proses seleksi dan penggaluran materi genetik memerlukan waktu yang cukup lama (>5 tahun). Teknik kultur antera dibantu uji cepat atau deteksi dini dapat mengurangi lamanya waktu seleksi dan penggaluran karena galur-galur yang akan diseleksi sudah merupakan galur murni homozigos yang langsung dihasilkan pada generasi pertama hasil kultur.
Teknik kultur antera F1 merupakan salah satu strategi untuk mengurangi lamanya waktu seleksi sehingga mempersingkat siklus pemuliaan dalam menghasilkan galur-galur murni homozigos. Pemuliaan konvensional membutuhkan 3 000–5 000 tanaman dalam generasi F2, sedangkan dengan kultur antera hanya dibutuhkan 60–70 tanaman (Senadhira et al. 2002). Beberapa galur hasil kultur antera sudah dihasilkan di Indonesia (Dewi dan Purwoko 2001; Sasmita et al. 2002; Safitri et al. 2010). Galur-galur tersebut merupakan galur padi sawah dan padi gogo, sehingga penelitian diarahkan untuk memperoleh varietas berdaya hasil tinggi, toleran terhadap cekaman kekeringan, naungan, toksisitas Aluminium dan blas. Penelitian perakitan padi toleran salinitas di Indonesia masih menggunakan sistem pemuliaan konvensional, sehingga penggunaan teknik kultur antera dalam pengembangan padi toleran salinitas diharapkan mampu menghasilkan galur-galur murni dihaploid (DH) yang toleran salinitas dalam waktu yang relatif cepat.
Perumusan Masalah
Lahan sawah irigasi yang semakin berkurang merupakan salah satu penyebab berkurangnya produksi beras nasional. Selain itu, perubahan iklim dan pemanasan global juga berpengaruh pada produktivitas padi di Indonesia, antara lain mempengaruhi salinitas lahan di sepanjang pantai. Penggunaan lahan salin merupakan salah satu upaya yang dapat dilakukan dalam peningkatan produksi padi. Saat ini varietas padi yang toleran salinitas masih sangat terbatas. Oleh karena itu, perlu dilakukan usaha perakitan varietas yang berdaya hasil tinggi toleran salinitas.
Perakitan varietas padi toleran salinitas dapat dilakukan melalui pemuliaan konvensional, penggunaan bioteknologi dan kombinasi keduanya. Pemuliaan konvensional membutuhkan waktu yang lama untuk menghasilkan galur murni yang homosigous. Penggunaan bioteknologi yang digabungkan dengan pemuliaan konvensional dapat mempersingkat proses pemuliaan dalam menghasilkan varietas padi.
Kultur antera merupakan salah satu metode yang mudah diterapkan pada tanaman padi. Penggunaan kultur antera yang dimodifikasi dengan pemuliaan konvensional, antara lain dalam pembentukan populasi melalui persilangan dan proses seleksi dapat menghasilkan galur murni padi yang homozigos dalam waktu yang relatif singkat.
dalam proses pemilihan galur-galur dihaploid toleran salinitas pada tahap awal, sehingga dapat mempercepat proses seleksi galur-galur dihaploid yang dihasilkan.
Tujuan Penelitian
Tujuan Umum
Tujuan penelitian ini adalah untuk mendapatkan galur-galur dihaploid yang mempunyai karakter agronomi baik, berdaya hasil tinggi dan toleran salinitas. Tujuan Khusus
Penelitian ini bertujuan untuk:
1. Mendapatkan plasma nutfah padi dengan berbagai tingkat toleransi terhadap salinitas melalui uji deteksi dini untuk digunakan sebagai tetua persilangan. 2. Mendapatkan konsentrasi NaCl kritis yang dapat digunakan untuk melakukan
penapisan genotipe padi di tanah salin di rumah kaca.
3. Mendapatkan F1 dari persilangan antara tetua berdaya hasil tinggi dengan tetua toleran salinitas.
4. Mendapatkan galur padi dihaploid melalui kultur antera.
5. Mendapatkan galur-galur dihaploid dengan berbagai tingkat toleransi terhadap salinitas pada fase bibit melalui pengujian pada media hidroponik.
6. Mendapatkan galur-galur dihaploid dengan berbagai tingkat toleransi terhadap salinitas di tanah salin melalui pengujian dengan menggunakan NaCl dan media tanah di rumah kaca.
7. Mendapatkan galur-galur padi dihaploid berpotensi hasil tinggi, mempunyai karakter agronomi baik dan toleran terhadap salinitas.
Hipotesis Penelitian
1. Terdapat plasma nutfah padi yang toleran salinitas berdasar pengujian pada fase bibit.
2. Terdapat konsentrasi NaCl yang dapat digunakan untuk melakukan uji penapisan genotipe padi di tanah salin di rumah kaca.
3. Terdapat F1 hasil persilangan antar tetua untuk perakitan padi toleran salinitas dengan daya kultur antera yang baik.
4. Terdapat galur-galur dihaploid yang bersifat homozigos melalui kultur antera. 5. Terdapat galur-galur dihaploid yang toleran salinitas berdasar pengujian pada
fase bibit pada media hidroponik.
6. Terdapat galur-galur dihaploid yang toleran salinitas berdasar pengujian pada di tanah salin.
7. Terdapat galur-galur dihaploid dengan karakter agronomi baik, berdaya hasil tinggi dan toleran terhadap salinitas.
Ruang Lingkup Penelitian
percobaan-percobaan dalam penelitian ini saling terkait. Penelitian ini terdiri atas tujuh percobaan, bagan alir penelitian disajikan pada Gambar 1.1.
Manfaat Penelitian
Hasil penelitian ini akan bermanfaat dalam menambah keragaman genetik padi toleran salinitas dengan galur/varietas yang dihasilkan. Penelitian ini juga bermanfaat dalam kegiatan penapisan terhadap genotipe padi di tanah salin melalui percobaan pot yang dilakukan dengan penambahan NaCl.
Kebaruan Penelitian
2 TINJAUAN PUSTAKA
Karakteristik Tanaman Padi
Padi (Oryza sativa L.) dalam taksonomi tumbuhan diklasifikasikan ke dalam divisio Spermatophyta, sub divisio Angiospermae, kelas Monocotyledoneae, ordo Poales, dan famili Gramineae (Grist 1959). Tanaman padi termasuk terna semusim yang mempunyai akar serabut, batang pendek serta daun berbentuk lanset dengan warna hijau muda hingga hijau tua dan berurat daun sejajar yang pada umumnya tertutupi oleh rambut-rambut yang pendek. Bunga padi tersusun majemuk dengan tipe malai bercabang. Satuan bunga disebut floret yang terletak pada satu spikelet yang duduk pada panikula. Tipe buah padi adalah kariopsis sehingga tidak dapat dibedakan mana buah dan bijinya. Bentuk biji padi bulat hingga lonjong dengan ukuran panjang 3 mm hingga 15 mm yang tertutup oleh palea dan lemma (sekam). Setiap bunga padi (floret) memiliki enam kepala sari (antera) dan kepala putik (stigma) bercabang dua. Kedua organ seksual ini umumnya siap bereproduksi dalam waktu yang bersamaan. Antera kadang-kadang keluar dari palea dan lemma jika telah masak. Dari segi reproduksi, padi merupakan tanaman menyerbuk sendiri, karena 95% atau lebih polen membuahi sel telur pada bunga di tanaman yang sama. Pada akhir perkembangan, sebagian besar bulir padi mengandung pati di bagian endosperm (IRRI 2004).
Genus Oryza memiliki lebih dari 20 spesies, tetapi yang banyak dibudidayakan di lima benua adalah Oryza sativa L., sedangkan Oryza glaberrima
Steund. hanya dibudidayakan terbatas di daerah Afrika Barat. Kedua spesies ini termasuk diploid (Gould 1968). Berdasar gambaran umum morfologi dan fisiologinya, Oryza sativa dibedakan menjadi tiga subspecies, yaitu indica,
japonica dan javanica (Chang dan Bardenas 1965). Subspesies indica mempunyai karakteristik: daun berukuran sempit berwarna hijau tua, gabah pendek dan agak bulat panjang, pada umumnya gabah tidak mempunyai ekor (awn), bulu sekam lebat dan panjang, gabah agak mudah rontok, jumlah anakan sedang, batang sedang-pendek, jaringan keras, tidak peka terhadap panjang hari atau agak peka, dan kandungan amilosa pada biji 10–24%. Subspesies japonica mempunyai karakteristik: daun berukuran sempit-lebar dan berwarna hijau muda, gabah panjang-pendek, umumnya gabah tidak mempunyai ekor (awn), bulu sekam jarang dan pendek, gabah mudah rontok, jumlah anakan banyak, batang sedang-tinggi, jaringan lunak, kepekaan terhadap panjang hari bervariasi, dan kandungan amilosa pada biji 23–31%. Subspesies javanica mempunyai karakteristik: daun berukuran lebar, kaku dan berwarna hijau muda, gabah panjang lebar dan tebal, gabah mempunyai ekor (awn) panjang atau tidak ada, bulu sekam panjang, gabah sulit rontok, jumlah anakan sedikit, batang tinggi, jaringan keras, agak peka terhadap panjang hari, dan kandungan amilosa pada biji 20–25% (Gupta dan O‟toole 1986).
periode minimum untuk berbunga umumnya panjang, biji sulit rontok, kurang toleran terhadap lingkungan yang tidak menguntungkan, dan hasil gabah relatif rendah. Padi cere mempunyai karakteristik: pada umumnya tidak mempunyai ekor pada ujung gabah, panjang malai sangat bervariasi, bentuk biji kecil dan panjang, daun kecil dan lembut, jumlah anakan banyak, batang pada umumnya kecil dengan panjang bervariasi, respon terhadap fotoperiode bervariasi, umumnya lebih responsif dibandingkan dengan padi bulu, periode minimum untuk berbunga bervariasi, umumnya lebih cepat dibandingkan dengan bulu, biji mudah rontok, mempunyai toleransi yang sedang-tinggi terhadap kondisi lingkungan yang tidak menguntungkan, dan hasil gabah relatif tinggi (Takahashi 1997).
Pertumbuhan tanaman padi dibedakan dalam tiga fase, yaitu fase vegetatif, generatif dan pematangan. Fase vegetatif dimulai dari awal pertumbuhan sampai pembentukan malai, fase reproduktif dari pembentukan malai sampai pembungaan dan fase pematangan dari pembungaan sampai gabah matang. Di daerah tropis, fase generatif berlangsung 35 hari dan fase pematangan 30 hari. Perbedaan masa pertumbuhan dibedakan berdasar lamanya fase vegetatif (Taslim dan Fagi 1989). IRRI (2004) mengelompokkan pertumbuhan tanaman padi dalam sembilan fase, yaitu: (0) perkecambahan (germination), (1) fase bibit (seedling), (2) fase pembentukan anakan (tillering), (3) fase pemanjangan batang (stem elongation), (4) fase inisiasi bunga sampai bunting (panicle initiation to booting), (5) fase
heading, (6) fase berbunga (flowering), (7) fase masak susu (milk grain), (8) fase masak tepung (dough grain), (9) fase biji masak (mature grain). Fase 0–3 merupakan fase vegetatif, fase 4–6 merupakan fase reproduktif, dan fase 7–9 merupakan fase pematangan (ripening).
Padi tergolong tanaman yang toleran terhadap kondisi pengairan. Harahap (1982) mengelompokkan padi menjadi tiga berdasar habitatnya yaitu padi gogo, padi sawah dan padi rawa. Menurut Taslim dan Fagi (1989), berdasarkan atas dalamnya air genangan, maka padi dapat digolongkan menjadi lima, yaitu:
1. Padi gogo, yaitu padi yang dalam budidayanya tidak pernah digenangi.
2. Padi sawah, yaitu padi yang seluruh waktu pertumbuhannya digenangi 5–25
Soepraptohardjo dan Suwardjo (1988) menyatakan bahwa dalam budidaya padi sawah dibutuhkan pengairan yang baik untuk mengatasi kendala pertumbuhan tanaman padi, sedangkan pada padi rawa atau padi rawa pasang surut diperlukan penguasaan teknologi budidaya yang tepat untuk mengatasi kondisi pertumbuhan tanaman agar tetap optimal. Sementara itu, untuk budidaya padi gogo masih sepenuhnya tergantung pada air hujan.
Cekaman Salinitas pada Tanaman Padi
Tanah Salin
Salinitas adalah kondisi tanah yang ditandai dengan konsentrasi garam terlarut tinggi. Tanah yang diklasifikasikan sebagai tanah salin adalah tanah dengan daya hantar listrik (Electrical Conductivity, EC) sebesar lebih dari 4 dS m-1, setara dengan 40 mM NaCl dan menghasilkan tekanan osmotik sekitar 0.2 MPa (Munns dan Tester 2008). Garam-garam yang menyebabkan salinitas adalah NaCl, Na2CO3, NaHCO3, Na2SO4, CaCl2, MgSO4 dan MgCl2. Di Indonesia, salinitas umumnya terjadi karena pengaruh air laut. Oleh sebab itu, NaCl merupakan penyebab utama terjadinya tanah salin (Suwarno 1985).
Lahan salin dibedakan menjadi dua, yaitu salinitas pada lahan pantai dan salinitas pada lahan di daerah arid (Ikehashi 1997). Sebagian besar dari lahan salin ini terjadi secara alamiah, dari akumulasi garam selama jangka waktu yang lama di zona arid dan semi arid. Pelapukan batuan tua melepaskan berbagai jenis garam terlarut, terutama klorida dari natrium, kalsium, dan magnesium, dan dalam jumlah yang lebih sedikit yaitu sulfat dan karbonat. NaCl adalah garam yang mudah larut dan paling banyak. Penyebab lain dari akumulasi garam adalah pengendapan garam laut yang dibawa oleh angin dan hujan. Dalam 1 kg air hujan dapat mengandung NaCl 6–50 mg; konsentrasinya menurun dengan jarak terjadinya hujan dari pantai (Munns dan Tester 2008).
Selain salinitas alami, salinitas sekunder juga menyumbang perubahan dari lahan pertanian menjadi salin secara signifikan melalui pembukaan lahan atau irigasi. Hal ini menyebabkan air tanah mengandung garam di zona perakaran. Dari 1 500 juta ha lahan pertanian di dunia berupa lahan kering, 32 juta ha (2%) dipengaruhi oleh salinitas sekunder dalam berbagai derajat. Dari 230 juta hektar lahan irigasi, sebanyak 45 juta ha (20%) dipengaruhi garam. Lahan irigasi hanya 15% dari total lahan untuk pertanian, tetapi produktivitas lahan irigasi dua kali produktivitas lahan tadah hujan sehingga mampu menyumbang produksi, sepertiga produksi bahan pangan dunia (Munns dan Tester 2008). Hal ini berarti produktivitas di lahan salin masih sangat rendah, sehingga perlu digunakan genotipe-genotipe yang toleran salinitas untuk meningkatkan produksi.
Pada budidaya tanaman padi, tanah salin dibedakan menjadi empat macam (Suwarno 1985): (1) tanah salin di daerah lebak yang dipengaruhi oleh musim, pada musim hujan tanah ini ditanami oleh padi toleran genangan dan pada musim kemarau tidak ditanami karena salinitas; (2) tanah salin yang disebabkan oleh pengaruh air laut di daerah pantai atau delta, dimana varietas padi toleran salinitas dapat ditanam pada musim hujan; (3) tanah salin yang terjangkau air laut pasang sehingga padi hanya dapat ditanam pada bagian yang tinggi dengan menggunakan air tawar untuk irigasi; (4) tanah salin di daerah beriklim kering yang terjadi akibat kurangnya drainase sehingga terjadi penumpukan garam.
Pengaruh Salinitas terhadap Tanaman
sehingga terjadi kekeringan fisiologis; (2) keseimbangan hara, karena salinitas menghambat penyerapan K, Ca dan Mg pada tanaman; dan (3) pengaruh racun, disebabkan oleh konsentrasi Na+ atau Cl- dalam jaringan yang tinggi.
NaCl mempengaruhi sifat kimia tanah dan selanjutnya berpengaruh terhadap pertumbuhan tanaman (Bhumbla dan Abrol 1978; Soepardi 1983). Persentase Na+ pada koloid tanah salin kurang dari 15% dan pada umumnya mempunyai pH kurang dari 8.5. Hal ini disebabkan garam yang terdapat dalam tanah adalah netral. Pada tanaman, salinitas berpengaruh pada proses-proses penting yang berhubungan dengan air dan ion (Sultana et al. 1999). Tingginya konsentrasi Na+ di dalam tanah menyebabkan hambatan penyerapan Ca2+, Mg2+, dan K+ bagi tanaman. Salinitas juga dapat menurunkan serapan P meskipun tidak sampai terjadi defisiensi. Meningkatnya kandungan Cl- diikuti pula oleh berkurangnya kandungan NO3- dalam tajuk (Quijano-Guerta dan Kirk 2002). Marschner (2012) menyatakan bahwa ion-ion Na+ dan Cl- yang terdapat pada tanah bergaram akan menghancurkan struktur enzim dan makromolekuler lain yang merusak organel sel, mengganggu fotosintesis dan respirasi, serta menghambat sintesis protein.
Salinitas pada padi berpengaruh pada terhambatnya perkecambahan, menurunnya pertumbuhan, luas daun, bobot kering, pembentukan biji (Khatun dan Flowers 1995) dan meningkatnya kehampaan (Asch et al. 1999). Gejala tanaman yang terpapar cekaman salinitas sangat sulit dibedakan dengan gejala kekeringan. Keracunan Na ditandai dengan mengeringnya daun (nekrosis) pada daun-daun tua, dimulai dari tepi dan ujung daun hingga akhirnya keseluruhan bagian daun mengering (Suwarno 1985; Tester dan Davenport 2003).
Menurut Suwarno (1985), pada fase perkecambahan tanaman padi toleran terhadap salinitas dan menjadi sangat peka pada saat fase awal bibit, kemudian toleransinya meningkat selama fase pertumbuhan vegetatif. Meskipun demikian, tidak terdapat korelasi antara toleransi pada fase berkecambah dengan toleransi pada fase bibit. Menurut Zeng et al. (2001), cekaman salinitas selama fase bibit dapat menurunkan bobot kering tanaman sebesar dua kali lipat dibandingkan dengan apabila cekaman tersebut terjadi pada fase pemasakan. Fase awal bibit dan fase reproduktif merupakan fase yang lebih peka terhadap salinitas dibandingkan dengan fase pembentukan anakan.
Pada kegiatan pertanian yang dilakukan di lahan pantai dimana aliran air laut selalu terjadi, kerusakan tanaman akibat NaCl merupakan masalah umum. Pada tanaman padi, NaCl berpengaruh pada penyerapan air dan nutrisi tanaman, dan juga pengambilan oksigen (respirasi) di akar. Pengaruh penghambatan oleh NaCl berturut-turut terjadi pada penyerapan H2O > K2O > P2O5 > O2 (respirasi) > NH4-. Urutan tersebut menunjukkan bahwa NaCl tidak secara langsung menghambat respirasi pada akar tanaman padi, tetapi menghambat penyerapan air dan transpirasi. Antagonisme antara ion-ion Na+ dan K+ juga mungkin terlibat dalam penurunan absorpsi K (Takenaga 1995).
Na+ dan Cl- secara efektif dieksklusi oleh akar tanaman. Halofit, flora alami pada tanah sangat salin, mampu mempertahankan eksklusi ini pada salinitas tinggi dibandingkan dengan glikofit (Munns dan Tester 2008).
Mekanisme Toleransi terhadap Salinitas
Mekanisme yang berperan dalam toleransi terhadap salinitas pada pada pertumbuhan tanaman ditentukan oleh pengaruh osmotik garam di dalam tanah, atau pengaruh toksik garam di dalam jaringan tanaman. Respon tanaman terhadap cekaman salinitas ditunjukkan oleh terjadinya penurunan pertumbuhan tajuk yang terjadi dalam dua fase, yaitu (1) respon yang cepat terhadap peningkatan tekanan osmotik eksternal, dan (2) respon yang lebih lambat karena akumulasi Na+ pada daun (Munns dan Tester 2008).
Fase pertama yang menyebabkan laju pertumbuhan tajuk menurun secara signifikan dimulai segera setelah konsentrasi garam di sekitar akar meningkat hingga konsentrasi ambang, yaitu 40 mM NaCl pada sebagian besar tanaman. Penurunan pertumbuhan daun ditandai dengan daun baru yang muncul lebih lambat dan perkembangan tunas lateral yang lebih lambat atau bahkan tidak terbentuk. Pada tanaman serealia, pengaruh utama salinitas yaitu menurunnya jumlah anakan dan penghambatan pertumbuhan daun (Munns dan Tester 2008).
Pada fase kedua, respon tanaman terhadap salinitas dimulai ketika akumulasi garam pada daun-daun tua mencapai konsentrasi toksik, yaitu konsentrasi meracuni yang pemanjangan daun tidak terjadi lagi sebelum kemudian daun-daun tersebut mati. Laju kematian daun yang lebih tinggi dibandingkan dengan laju pertumbuhan daun baru menyebabkan kapasitas fotosintesis tanaman tidak dapat mensuplai kebutuhan karbohidrat daun-daun muda sehingga laju pertumbuhan menurun (Munns dan Tester 2008).
Menurut Levitt (1980), tanaman beradaptasi terhadap kondisi cekaman salinitas melalui dua cara, yaitu penghindaran (avoidance) dan toleransi terhadap cekaman. Pada mekanisme penghindaran, tanaman berusaha mencegah atau mengurangi penetrasi cekaman ke dalam jaringan, misalnya melalui penebalan kutikula dan penutupan stomata. Mekanisme toleransi dilakukan dengan cara bertahan terhadap cekaman internal. Tanaman membiarkan cekaman masuk ke dalam jaringan, tetapi tanaman berusaha mengurangi atau menghilangkan pengaruh cekaman tersebut dengan cara menjaga keseimbangan osmotik dalam jaringan sehingga mencegah terjadinya kerusakan akibat keracunan garam.
Strategi tanaman dalam menghadapi salinitas tinggi dapat dibedakan menjadi:
1. Adaptasi morfologi
2. Adaptasi fisiologi
a. Pengaturan potensial osmotik (osmoregulasi)
Tanaman yang toleran salinitas dapat melakukan penyesuaian dengan menurunkan tekanan osmotik tanpa kehilangan turgor. Penyesuaian ini dilakukan dengan penyerapan atau akumulasi ion-ion dan sintesis solut organik di dalam sel. Senyawa yang disintesis tanaman berupa asam-asam organik, asam amino dan senyawa gula. Selain untuk penyesuaian osmotik, sintesis senyawa-senyawa tersebut juga bertujuan untuk melindungi enzim-enzim terhadap penonaktifan pada aktivitas air internal yang rendah (Harjadi dan Yahya 1988). Senyawa yang umum ditemukan pada tanaman yang tercekam salinitas adalah sukrosa, prolina, glisin betaina, asam oksalat, malat dan glutamat (Kumar et al. 2000; Munns dan Tester 2008). b. Eksklusi Na+
Eksklusi Na+ oleh akar memastikan bahwa natrium tidak terakumulasi pada konsentrasi yang toksik dalam daun. Kegagalan eksklusi Na+ oleh akar mengakibatkan keracunan setelah beberapa hari atau beberapa minggu, tergantung pada spesies tanaman, dan menyebabkan kematian dini daun-daun tua (Munns dan Tester 2008).
c. Toleransi jaringan (kompartementasi)
Toleransi jaringan dilakukan tanaman dengan cara mengakumulasi Na+ atau Cl-. Mekanisme ini membutuhkan kompartementasi Na+ dan Cl -pada level seluler dan interseluler untuk menghindari konsentrasi toksik dalam sitoplasma, terutama pada sel-sel mesofil daun. Keracunan terjadi pada saat konsentrasi Na+ sangat tinggi pada daun-daun tua (Munns dan Tester 2008).
3. Adaptasi biokimia
Adaptasi biokimia tanaman yang tercekam salinitas dilakukan dengan peningkatan produksi asam absisat (ABA) dan peningkatan aktivitas enzim (Sopandie 2014). ABA berperan penting dalam konduktansi stomata (Munns dan Tester 2008) dan memperbaiki rasio K+/Na+ dengan memacu ekskresi garam melalui pengguguran daun (Sopandie 2014).
4. Adaptasi molekuler
Tanaman menghadapi cekaman salinitas dengan mengaktifkan gen yang berhubungan dengan selektivitas transpor ion dan integritas membran. Konsentrasi K+ di dalam sel tanaman dapat dipertahankan dengan meningkatkan ekspresi gen yang mengendalikan potassium-specific cotransporter. Pada beberapa spesies, enzim yang mengendalikan transporter K+ hanya diinduksi pada kondisi stres salin (Sopandie 2014).
Kultur Antera dalam Pemuliaan Tanaman Padi
dalam pembangunan pertanian. Kultur jaringan merupakan salah satu penerapan bioteknologi, dimana dalam teknik ini dilakukan isolasi terhadap bagian dari tanaman seperti protoplasma, sel, kelompok sel, jaringan dan organ, serta menumbuhkannya dalam kondisi aseptik sehingga bagian-bagian tersebut dapat memperbanyak diri dan beregenerasi menjadi tanaman yang lengkap. Hal ini merupakan penerapan dan pembuktian dari teori „totipotensi sel‟, dimana setiap sel, jaringan atau organ mempunyai kemampuan untuk beregenerasi menjadi tanaman yang lengkap (Khumaida dan Efendi 2011).
Kultur antera merupakan salah satu teknik dalam kultur jaringan tanaman yang direkomendasikan untuk membantu program pemuliaan padi. Secara teoritis, kultur antera memiliki beberapa keuntungan, yaitu (1) memperpendek siklus pemuliaan dengan diperoleh homozigositas secara cepat, (2) menambah efisiensi seleksi, dimana seleksi diantara progeni dihaploid homozigos dapat lebih efisien dibandingkan dengan seleksi pada progeni bersegregasi dalam pemuliaan konvensional, (3) memperluas variabilitas genetik melalui produksi variasi gametoklonal, (4) gen resesif terekspresi lebih cepat, dan (5) sumber benih homozigos dapat tersedia lebih cepat dalam rangka peningkatan dan pelepasan kultivar baru. Di samping keuntungan-keuntungan tersebut, teknik kultur antera juga mempunyai beberapa kelemahan yaitu: (1) pelaksanaan teknik kultur antera memerlukan peralatan dan personil khusus, (2) frekuensi haploid tidak dapat diprediksi dalam populasi, (3) penampilan galur inbred turunan dihaploid mungkin lebih inferior dibandingkan dengan penampilan galur inbred hasil pemuliaan konvensional (Fehr 1987; Zapata 1990; Dewi et al. 1996; Masyhudi et al. 1997), (4) rendahnya tingkat regenerasi tanaman hijau, (5) banyaknya tanaman albino, (6) tidak semua genotipe responsif terhadap kultur antera, dan (7) ploidisitas tanaman yang dihasilkan (Masyhudi et al. 1997, Somantri et al. 2003).
Kultur antera merupakan salah satu aplikasi bioteknologi yang dapat mempersingkat siklus pemuliaan tanaman karena dapat menghilangkan sebagian besar dari kegiatan seleksi pada tiap generasi (6–8 generasi) yang umum dilakukan pada pemuliaan konvensional. Melalui teknik kultur antera, pembentukan galur homozigos tanaman padi dapat dipercepat. Galur murni dapat diseleksi dari populasi dihaploid yang homogen dan homozigos sehingga hasil rekombinasi dari persilangan difiksasi sebagai galur homozigos dan galur-galur harapan diseleksi berdasarkan keunggulan sifat-sifat agronominya. Populasi tanaman yang diseleksi juga akan lebih sedikit. Populasi dihaploid minimum yang diperlukan untuk evaluasi bervariasi tergantung dari jumlah gen untuk seleksi. Jika perbedaan pada tetua persilangan adalah sejumlah n gen dan diasumsikan tidak ada pautan, maka minimum sebanyak 2n tanaman harus ditanam agar semua genotipe homozigos dapat terwakili, sedangkan dengan pemuliaan konvensional diperlukan sebanyak 4n tanaman (Dewi dan Purwoko 2001). Jumlah populasi yang diperlukan untuk bahan seleksi tergantung kepada tujuan seleksi dalam arti bagaimana kontrol genetik karakter yang diinginkan tersebut. Makin banyak gen yang mengontrol karakter yang diinginkan maka jumlah anggota materi populasi untuk bahan seleksi akan semakin besar (Somantri et al. 2003), walaupun tetap lebih sedikit dibandingkan dengan bahan seleksi pada pemuliaan konvensional (Dewi dan Purwoko 2001).
generasi ke generasi sehingga seleksi dapat dilakukan langsung pada generasi kedua (DH1) yang berasal dari perbanyakan generasi awal (DH0) hasil kultur antera. Karakteristik agronomi seperti hasil dan kualitas biji serta toleransi terhadap cekaman biotik dan abiotik dikendalikan oleh gen minor sehingga dapat segera dievaluasi pada generasi DH1 dan DH2 (Fehr 1987, Chung 1992). Pembentukan tanaman dihaploid spontan tersebut sangat menguntungkan sebagai bahan seleksi karena tidak perlu menggandakan kromosom tanaman haploid terlebih dahulu seperti pada kultur mikrospora. Pada umumnya, tanggap in vitro dari mikrospora seringkali terjadi pada saat mikrospora masih berada di dalam antera (Dewi dan Purwoko 2011).
Keberhasilan kultur antera dipengaruhi oleh berbagai faktor, yaitu genotipe tanaman, komposisi media, praperlakuan antera sebelum dikulturkan, fase pembentukan mikrospora pada saat antera diambil, kondisi tumbuh lingkungan tanaman yang akan diambil anteranya dan lingkungan pada saat pengambilan sampel malai (Chu 1978, Gupta dan Borthakur 1987, Cowen et al. 1992, Raina dan Zapata 1997, Lee et al. 2004).
Dewi et al. (2004, 2007) melaporkan bahwa penambahan poliamin, yaitu 1 mM putresin pada media induksi kalus dan regenerasinya dalam kultur antera padi menghasilkan induksi kalus dan regenerasi tanaman hijau yang lebih baik. Poliamin merupakan zat pengatur tumbuh yang berperan dalam pertumbuhan dan perkembangan tanaman. Poliamin yang umum ditemukan pada tanaman adalah putresin (butan-1, 4-diamin), spermidin [(N-3-aminopropil) butan-1, 4-diamin] dan spermin [NN‟-bis-(3-aminopropil) butan-1, 4-diamin]. Putresin merupakan prekursor untuk sintesis spermidin dan spermin berturut-turut melalui penambahan satu dan dua gugus aminopropil. Santos et al. (1996), menyatakan bahwa poliamin berperan dalam morfogenesis polen jagung pada kultur antera jagung karena dapat meningkatkan pertumbuhan dan perkembangan kalus mikrospora serta regenerasinya. Metabolisme poliamin melalui lintasan arginin dekarboksilase (ADC) diketahui mempengaruhi pertumbuhan dan potensi embriogenik kalus asal embrio pada kultur jaringan padi. Penelitian Dewi et al. (2007) menunjukkan bahwa aplikasi 1 mM putresin berhasil meregenerasikan tanaman hijau pada kultur antera padi indica yang biasanya sukar menghasilkan tanaman hijau atau rekalsitran in-vitro.
3 PENAPISAN PLASMA NUTFAH PADI TERHADAP
SALINITAS PADA FASE BIBIT
Abstrak
Toleransi terhadap salinitas pada tanaman padi merupakan tujuan penting dalam pemuliaan padi, oleh sebab itu penapisan terhadap plasma nutfah padi perlu dilakukan, salah satunya adalah penapisan pada fase bibit. Penelitian dilaksanakan di Rumah Kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik (BB Biogen), Bogor pada bulan April–Mei 2013. Materi yang digunakan adalah 13 plasma nutfah dan dua pembanding, yaitu Pokkali (toleran salinitas) dan IR29 (peka salinitas). Percobaan hidroponik menggunakan rancangan kelompok lengkap teracak (RKLT) tiga ulangan dengan dua faktor yaitu konsentrasi NaCl (0 mM dan 120 mM) dan genotipe tanaman padi. Hasil penelitian menunjukkan bahwa salinitas yang diinduksi oleh 120 mM NaCl menyebabkan penurunan pertumbuhan bibit pada semua genotipe padi hingga menyebabkan kematian pada genotipe yang sangat peka. Skoring visual berdasarkan gejala kerusakan daun pada bibit padi yang tercekam salinitas karena pemberian 120 mM NaCl menghasilkan lima genotipe toleran yaitu Dendang, Inpara 5, Inpari 29, IR77674-3B-8-2-2-14-4-AJY2, dan IR81493-B-B-B-6-B-2-1-2. Inpara 4 tergolong sebagai genotipe yang sangat peka, sebanding dengan IR29, sehingga varietas Inpara 4 dapat dijadikan sebagai pembanding peka pada penelitian selanjutnya.
Abstract
Salinity significantly reduced the growth and yield of rice especially in coastal areas. Early seedling phase of rice growth is the most sensitive phase to salinity, therefore the screening of rice genotypes at the seedling phase need to be done. The greenhouse experiment was conducted at Indonesian Center for Agricultural Biotechnology and Genetic Resources Research and Development (ICA BIOGRAD), Bogor, Indonesia from April to May 2013. The material used was 13 genotypes and two check varieties, namely Pokkali (salt tolerant) and IR29 (salt sensitive). Nutrient culture (hydroponic) experiment was conducted using a randomized block design with three replications and two factors, namely the concentrations of NaCl (0 mM and 120 mM) and the genotypes of rice plants. The results showed that the salinity induced by 120 mM NaCl caused seedling growth reduction in all rice genotypes, and at the end the highly sensitive genotypes were not survived. Based on visual injury symptoms of rice seedling leaves, five genotypes were found to be tolerant at 120 mM salinity level, i.e. Dendang, Inpara 5, Inpari 29, IR77674-3B-8-2-2-14-4-AJY2, and IR81493-BBB-6-B- 2-1-2. Inpara 4 was classified as highly salt sensitive similar to IR29. This variety can be used as a salt sensitive check.
Pendahuluan
Salinitas merupakan salah satu masalah utama dalam budidaya tanaman padi di seluruh dunia (Hosseini et al. 2012; Abbas et al. 2013). Tanaman padi diklasifikasikan peka terhadap salinitas, namun padi sawah merupakan salah satu yang direkomendasikan ditanam di tanah salin, karena padi sawah mempunyai kemampuan untuk tumbuh di tanah tergenang (Sankar et al. 2011; Aref dan Rad 2012). Kategori kondisi lahan salin dibagi menjadi tiga, yaitu salinitas rendah (2-6 dS m-1), salinitas sedang (6-10 dS m-1) dan salinitas tinggi (>10 dS m-1) (Zeng dan Shannon 2000). Kehilangan hasil padi pada kondisi tanah salin sedang sampai tinggi tinggi (>6 dS m-1) mencapai 50–100% (Rad et al. 2012).
Pengaruh salinitas pada tanaman dimulai ketika akumulasi garam pada daun-daun tua mencapai konsentrasi meracuni, ditandai dengan berhentinya pemanjangan daun dan kemudian daun-daun tersebut mati. Laju kematian daun lebih tinggi dibandingkan dengan laju pertumbuhan daun baru akan menyebabkan kapasitas fotosintesis tanaman menurun dan tidak akan dapat mensuplai kebutuhan karbohidrat daun-daun muda sehingga laju pertumbuhan menurun (Munns dan Tester 2008).
Garam yang menyebabkan salinitas pada tanah didominasi oleh NaCl karena NaCl adalah garam yang mudah larut dan paling banyak. Pengaruh salinitas pada padi sawah menyebabkan gagal atau terhambatnya perkecambahan, menurunnya pertumbuhan, luas daun, produksi bobot kering dan pembentukan biji (Khatun dan Flowers 1995) serta meningkatnya kehampaan (Asch et al. 1999). Keracunan Na ditandai dengan mengeringnya tepi bagian ujung daun, demikian juga keracunan Cl. Gejala tersebut sangat sulit dibedakan dengan gejala kekeringan. Keparahan dari pengaruh salinitas tergantung pada faktor berikut: (a) intensitas cekaman salinitas, (b) kondisi iklim dan (c) tingkat toleransi genotipe yang ditanam (Suwarno 1985).
Respon tanaman padi terhadap salinitas bervariasi menurut stadia pertumbuhan. Pada sebagian besar padi budidaya, fase awal bibit merupakan fase yang paling peka terhadap salinitas (Suwarno 1985; Zeng dan Shannon 2000; Zeng et al. 2001; Haq et al. 2009). Toleransi tanaman terhadap salinitas yang diamati pada fase bibit dapat digunakan dalam menentukan pertumbuhan tanaman dan toleransi akhir terhadap salinitas pada spesies tanaman (Haq et al. 2009). Hosseini et al. (2012) menyatakan bahwa penapisan toleransi terhadap salinitas pada fase bibit dilakukan karena variasi pada fase ini dikendalikan secara genetik. Menurut Zeng et al. (2001), cekaman salinitas selama fase bibit dapat menurunkan bobot kering tanaman sebesar dua kali lipat dibandingkan dengan apabila cekaman tersebut terjadi pada fase pemasakan. Fase awal bibit dan fase reproduktif merupakan fase yang lebih peka terhadap salinitas dibandingkan dengan fase pembentukan anakan. Menurut Suwarno (1985), pada fase perkecambahan tanaman padi toleran terhadap salinitas dan menjadi sangat peka pada saat fase awal bibit, kemudian toleransinya meningkat selama fase pertumbuhan vegetatif. Meskipun demikian, tidak terdapat korelasi antara toleransi pada fase berkecambah dengan toleransi pada fase bibit.
Bahan dan Metode
Tempat dan Waktu
Penelitian dilaksanakan di Rumah Kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian (BB Biogen), Bogor pada bulan April-Mei 2013.
Bahan dan Metode
Materi yang digunakan adalah 13 genotipe padi yang berasal dari IRRI dan Balai Besar Penelitian Tanaman Padi (BB padi), dengan pembanding Pokkali (toleran salinitas) dan IR29 (peka salinitas) (Tabel 3.1). Percobaan menggunakan rancangan kelompok lengkap teracak (RKLT) dengan dua faktor dan tiga ulangan. Faktor pertama adalah konsentrasi NaCl (0 mM dan 120 mM atau setara dengan 0 dS m-1 dan 11 dS m-1), sedangkan faktor kedua adalah genotipe tanaman padi. Setiap satu satuan percobaan terdiri atas empat bibit tanaman padi. Penapisan dilakukan dengan metode Egdane et al. (2007) yang dimodifikasi (Lampiran 1), yaitu dengan menggunakan media hidroponik (Yoshida et al. 1976) yang mengandung NaCl 120 mM atau Electrical Conductivity (EC) + 11 dS m-1.
Tabel 3.1 Materi genetik padi yang digunakan untuk penapisan terhadap salinitas
No. Genotipe Asal atau persilangan
1 Banyuasin Cisadane/Kelara
2 Cilamaya Muncul Pelita I-1/B2388
3 Dendang Osok/IR5657-33-2
4 Inpara 4 (Swarna sub-1) Introduksi IRRI (IR05F101) 5 Inpara 5 (IR64 sub-1) Introduksi IRRI (IR07F101)
6 Inpari 29 IR69502/KAL9418// Pokkali/Angke
7 Inpari 30 (Ciherang sub-1) Introduksi IRRI
8 IR64 IRRI (IR5657/IR2061)
9 IR77674-3B-8-2-2-14-4-AJY2 IRRI (IR71730-51-2/ IRRI 128)
10 IR78788-B-B-10-1-2-4-AJY1 IRRI (BR41/IRRI 128), dilepas dengan nama Inpari 34 Salin Agritan
11 IR81493-B-B-B-6-B-2-1-2 IRRI (IR75000/IR71684)
12 Mendawak Mahsuri/Kelara
13 Siak Raya Batang Ombilin/Kelara
14 Pokkali (pembanding toleran) 15 IR29 (pembanding peka)
Bibit berumur 7 hari dicabut dari media pesemaian dan dipindahtanamkan ke dalam kultur hara dengan menggunakan nampan styrofoam yang telah dilubangi dengan diameter 18 mm, jarak antar lubang 50 mm x 40 mm. Penanaman bibit dilakukan dengan cara menggulung batang bibit dengan lembaran busa tipis dan meletakkannya ke lubang yang sudah dibuat pada
kemudian setelah dua hari ditingkatkan menjadi 120 mM NaCl. Larutan dipertahankan pada pH 5.0–5.1 dengan menambahkan NaOH atau HCl 1 N setiap dua hari sekali. Media kultur hara diganti satu minggu sekali. Perlakuan bibit dilakukan selama 14 hari atau hingga varietas pembanding peka telah mati. Urutan kegiatan percobaan dapat dilihat pada Tabel 3.2.
Pengamatan yang dilakukan meliputi skoring toleransi bibit terhadap salinitas sesuai dengan standar evaluasi dari IRRI (2003) yang disajikan pada Tabel 3.3. Selain itu dilakukan pengamatan terhadap karakter agronomi bibit yaitu tinggi bibit, panjang akar terpanjang, jumlah anakan, jumlah daun, bobot basah tajuk, bobot kering tajuk, bobot basah akar dan bobot kering akar. Data agronomi bibit dianalisis ragam, apabila terdapat beda nyata maka diteruskan dengan uji jarak berganda Duncan pada taraf α = 5%.
Tabel 3.2 Urutan kegiatan penapisan plasma nutfah padi terhadap salinitas melalui uji deteksi dini pada fase bibit
Umur bibit
(hari) Perlakuan Keterangan
0 Semai benih tahap 1 Dalam cawan petri 2 Semai benih tahap 2 Dalam bak pembibitan 7 Pindah bibit ke dalam
media hidroponik (tanpa NaCl)
Bibit ditanam mennggunakan nampan
styrofoam
21 Perlakuan NaCl Pemberian NaCl dilakukan bertahap dari 0 mM menjadi 60 mM dan 120 pengukuran tinggi tanaman, jumlah anakan, panjang akar, berat basah dan berat kering tanaman
Tabel 3.3 Kriteria evaluasi terhadap cekaman salinitas pada fase bibit
Skor Deskripsi Kategori toleransi
1 Pertumbuhan normal, beberapa daun tua menunjukkan gejala berwarna putih pada ujung daun. Tidak ada gejala terlihat pada daun-daun muda.
Sangat toleran
3 Pertumbuhan agak normal, ujung daun tua kering kecoklatan, ujung daun muda berwarna putih.
Toleran
5 Pertumbuhan terhambat, beberapa daun sudah mengering, titik tumbuh masih masih dapat tumbuh dan memanjang.
Moderat toleran