ISOLASI, SELEKSI, DAN IDENTIFIKASI BAKTERI
ENDOFIT SEBAGAI AGENS PENGINDUKSI KETAHANAN
TANAMAN PADI TERHADAP PENYAKIT HAWAR DAUN
BAKTERI
IDA PARIDA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Isolasi, Seleksi, dan Identifikasi Bakteri Endofit sebagai Agens Penginduksi Ketahanan Tanaman Padi terhadap Penyakit Hawar Daun Bakteri adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
IDA PARIDA. Isolasi, Seleksi, dan Identifikasi Bakteri Endofit sebagai Agens Penginduksi Ketahanan Tanaman Padi terhadap Penyakit Hawar Daun Bakteri. Dibimbing oleh GIYANTO dan TRI ASMIRA DAMAYNTI.
Hawar daun bakteri (HDB) yang disebabkan Xanthomonas oryzae pv. oryzae menjadi salah satu masalah besar dalam produksi padi di Indonesia. Salah satu upaya pengendalian yang dapat dilakukan yaitu dengan pemanfaatan agens hayati seperti bakteri endofit. Bakteri endofit merupakan bakteri yang dapat mengolonisasi jaringan internal tanaman tanpa menyebabkan kerusakan pada inangnya. Bakteri ini memiliki mekanisme induksi ketahanan tanaman atau dikenal dengan induced systemic resistance (ISR) dalam pengendalian penyakit. Namun potensinya sebagai agens penginduksi ketahanan tanaman terhadap penyakit hawar daun bakteri serta pengaruhnya terhadap pertumbuhan dan hasil panen padi sejauh ini belum banyak diketahui.
Penelitian ini bertujuan mengisolasi, menyeleksi, dan menguji kemampuan bakteri endofit asal tanaman padi sebagai agens penginduksi ketahanan tanaman serta mengetahui pengaruhnya terhadap pertumbuhan dan hasil panen padi. Selain itu untuk mengidentifikasi bakteri endofit yang potensial sebagai agens penginduksi ketahanan tanaman padi terhadap penyakit HDB. Penelitian dilaksanakan di Laboratorium Bakteriologi Tumbuhan, Departemen Proteksi Tanaman, dan Rumah Kaca Cikabayan, IPB mulai bulan Juni 2013 sampai Desember 2014. Bakteri endofit yang digunakan merupakan hasil isolasi dari tanaman padi sehat yang berada diantara tanaman sakit akibat HDB. Isolat bakteri endofit hasil isolasi diseleksi berdasarkan viabilitas dan morfologi koloni, pengaruhnya terhadap induksi ketahanan dan pertumbuhan padi di pembibitan, serta uji reaksi hipersensitif pada tanaman tembakau. Bakteri endofit hasil seleksi selanjutnya diuji pada percobaan rumah kaca. Parameter yang digunakan meliputi ekspresi gen PR1 dan PBZ1, aktivitas enzim peroksidase, periode inkubasi, dan perkembangan penyakit HDB. Selain itu juga diamati pengaruhnya terhadap pertumbuhan dan hasil panen padi dengan parameter daya berkecambah, indeks vigor benih, pertambahan tinggi tanaman, jumlah anakan maksimum, jumlah anakan produktif, bobot kering, dan kadar air gabah. Perlakuan bakteri endofit di rumah kaca terdiri dari W1 (aplikasi bakteri endofit pada benih saja), W2 (aplikasi bakteri endofit pada benih dan 4 MST), dan W3 (aplikasi bakteri endofit pada benih, 4, dan 6 MST). Bakteri endofit yang potensial sebagai agens penginduksi ketahanan tanaman padi dan berpengaruh baik terhadap pertumbuhan dan hasil panen padi selanjutnya diidentifikasi menggunakan teknik molekuler berdasarkan sekuen 16S rRNA.
menyebabkan reaksi hipersensitif pada tanaman tembakau. Tujuh isolat bakteri endofit tersebut adalah EA2 154, EB3 307, EB4 451, EB4 452, EB6 748, ED1 63, dan ED4 467. Hasil pengujian di rumah kaca menunjukkan bahwa ketujuh isolat yang diujikan mampu menginduksi ekspresi gen PR1 dan PBZ1, dan 4 diantaranya mampu meningkatkan aktivitas enzim peroksidase (EB4 451, EB4 452, EB6 748, dan ED4 467). Selain itu 4 isolat mampu memperpanjang periode inkubasi (EA2 154, EB4 451, EB6 748, dan ED4 467) dan 2 isolat mampu menghambat perkembangan penyakit HDB (EB4 451 dan ED1 63). Secara umum perlakuan bakteri endofit tidak berpengaruh nyata terhadap pertumbuhan dan hasil panen padi, kecuali terhadap daya berkecambah, yaitu EB6 748 yang mempengaruhi daya berkecambah lebih rendah dibandingkan kontrol dan perlakuan lain. Diantara 7 isolat yang diuji, hanya isolat EB4 451 yang konsisten mampu menginduksi ekspresi gen PR1 dan PBZ1, meningkatkan aktivitas enzim peroksidase, memperpanjang periode inkubasi, menekan perkembangan penyakit, serta berpengaruh baik terhadap pertumbuhan dan hasil panen padi. Hasil identifikasi dengan teknik molekuler diketahui bahwa EB4 451 memiliki persen homologi sebesar 98% dengan Bacillus subtilis.
Berdasarkan penelitian ini bakteri endofit yang diisolasi dari bagian tanaman padi berpotensi sebagai agens penginduksi ketahanan tanaman dan bermanfaat dalam upaya pengendalian penyakit HDB. B. subtilis merupakan salah satu bakteri endofit yang secara konsisten mampu menginduksi ketahanan tanaman padi. Hasil penelitian ini menunjukkan bahwa bakteri endofit sebagai agens hayati dapat dimanfaatkan untuk pengendalian HDB.
SUMMARY
IDA PARIDA. Isolation, Selection and Identification of Endophytic Bacteria as Inducers Sistemic Resistance Agents to Bacterial Leaf Blight Disease on Rice. Supervised by GIYANTO and TRI ASMIRA DAMAYNTI.
Bacterial leaf blight (BLB) caused by Xanthomonas oryzae pv. oryzae become one of the major problems on rice production in Indonesia. One of the control method is by utilizing endophytic bacteria as biocontrol. Endophytic bacteria is a bacteria which colonize the internal tissues of plants without causing damage to the host. These bacteria play a role as plant resistance inducer, known as induce systemic resistance (ISR) in controlling the disease. However, its potential as inducer of plant resistance agents to bacterial leaf blight disease, its influence on the growth and yield of rice so far is not much known.
This study aims to isolate, select and test the ability of endophytic bacteria from rice plant as inducers plant resistance agents and their influence on the growth and yield of rice, besides to identifying potential endophytic bacteria as inducers plant resistance agents to BLB on rice. Research conducted at the Laboratory of Plant Bacteriology, Department of Plant Protection, and Cikabayan Greenhouse, IPB, started in June 2013 until December 2014. The endophytic bacteria are isolated from healthy rice plants. Endophytic bacteria isolates were selected based on the viability and morphology of the colony, its influence on the induction of resistance and plant growth in the nursery, and hypersensitive reaction test on tobacco plants. Endophytic bacteria of selection results were tested in greenhouse trials with parameters PR1 and PBZ1 gene expression, peroxidase enzyme activity, the incubation period, and disease progression. And than they influence on the growth and yield of rice with parameters germination, seed vigor index, increase plant height, number of maximum tillers, number of productive tillers, dry weight, and water content of grain. The treatment of endophytic bacteria consisted of W1 (application endophytic bacteria on the seed alone), W2 (application endophytic bacteria on the seed and 4 week after plants), and W3 (application endophytic bacteria on the seed, 4, and 6 week after plants). Identification of potential endophytic bacteria as inducer plant resistance and have better effect on growth and yield of rice was conducted by sequencing of the 16S rRNA.
the incubation period (EA2 154, EB4 451, EB6 748, and ED4 467) and 2 isolates could inhibit the disease progression (EB4 451 and ED1 63). In general, the application of endophytic bacteria had no significant effect on the growth and yield of rice, except for germination, ie EB6 748 affecting lower germination than control and other treatment. Among 7 isolates tested, only EB4 451 were consistent to induce PR1 and PBZ1 gene expression, increases the peroxidase enzyme activity, prolong the incubation period, suppress the disease progression, and good influence on the growth and yield of rice. Results of identification by molecular techniques known that EB4 451 has 98% homology with Bacillus subtilis.
Endophytic bacteria isolated from the rice plant is potentially as inducer plant resistance agents to control BLB. B. subtilis is one of endophytic bacteria was consistently able to induce plant resistance related PR1 and PBZ1 gene expression, peroxidase enzyme activity, prolong incubation period, lower disease development, and having better effect on plant growth and yield of rice. Based on the present results showed that utilizing endophytic bacteria might be considerated as biocontrol agent of BLB.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
ISOLASI, SELEKSI, DAN IDENTIFIKASI BAKTERI
ENDOFIT SEBAGAI AGENS PENGINDUKSI KETAHANAN
TANAMAN PADI TERHADAP PENYAKIT HAWAR DAUN
BAKTERI
IDA PARIDA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Fitopatologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT yang telah memberikan rahmat dan hidayah sehingga penulis dapat menyelesaikan tesis dengan judul
“Isolasi, Seleksi, dan Identifikasi Bakteri Endofit sebagai Agens Penginduksi Ketahanan Tanaman Padi terhadap Penyakit Hawar Daun Bakteri”.
Penulis menyadari bahwa banyak pihak yang telah berjasa dalam memberikan bimbingan, dukungan, arahan dan bantuan dalam menyelesaikan tesis ini. Penulis sampaikan ucapan terima kasih terutama kepada Dr Ir Giyanto, MSi dan Dr Ir Tri Asmira Damayanti, MAgr yang telah memberikan bimbingan dan saran dengan penuh kesabaran selama pengerjaan tesis. Terima kasih kepada Program KKP3N, Badan Penelitian dan Pengembangan Pertanian, Kementrian Pertanian, Indonesia atas pendanaan penelitian ini sebagai bagian dari kerjasama dengan nomor kontrak: 50/PL.220/I.1/2014.K10, serta beasiswa pendidikan dari Provinsi Jawa Barat dengan nomor kontrak: 073/08/Yansos dan 1683/IPB/DL/2013. Terima kasih kepada Dr Ir Abdjad Asih Nawangsih, MSi selaku penguji luar komisi, dan Prof Dr Ir Sri Hendrastuti Hidayat, MSc yang telah memberikan saran dalam penulisan tesis.
Penulis sampaikan rasa terima kasih yang sebesar-besarnya kepada keluarga, terutama Ibu Titi Rositi dan Bapak Ai Ruhaeman SPd.I (almarhum) yang selama hidupnya selalu memberikan kasih sayang, dukungan, doa, dan semangat belajar. Terima kasih kepada suami tercinta Apan Iskandar SPd atas saran, dukungan, doa, dan semangat dalam pengerjaan tesis, dan terima kasih kepada anak tercinta Muhammad Sajjad Iskandar yang selalu menjadi penyemangat bagi penulis. Selain itu penulis sampaikan terima kasih kepada semua pihak yang telah memberikan dukungan baik moril maupun materil, terutama para sahabat Rita Kurnia Apindiati, Lutfi Afifah, Tatit Sastrini, Risma Junita, Ima Karimah, dan Rindu Dwi Malateki. Terima kasih kepada keluarga besar Laboratorium Bakteriologi Tumbuhan PTN dan keluarga besar Fitopatologi 2012 atas dukungan saran dan semangat.
Semoga tesis ini bermanfaat bagi penulis maupun pembaca. Penulis menyadari bahwa dalam tesis ini masih banyak kekurangan. Sebab itu penulis mengharapkan saran dan kritik yang bersifat membangun.
Bogor, April 2016
DAFTAR ISI
Halaman
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 3
Hipotesis 3
Alur Penelitian 4
2 TINJAUAN PUSTAKA 5
Penyakit Hawar Daun Bakteri Pada Padi 5
Peran Bakteri Endofit Sebagai Agens Hayati 6
Induksi Ketahanan Tanaman Padi 7
3 METODE PENELITIAN 10
Tempat dan Waktu 10
Bahan dan Alat 10
Metode 10
Pengambilan Sampel Tanaman Padi untuk Isolasi Bakteri Endofit 10
Isolasi Bakteri Endofit 10
Penyiapan Isolat X. oryzae pv oryzae 11
Seleksi Isolat Bakteri Endofit Potensial 11
Uji viabilitas dan morfologi koloni 11
Uji induksi ketahanan dan pemacu pertumbuhan di pembibitan 12
Uji reaksi hipersensitif (HR) 12
Pengaruh Aplikasi Bakteri Endofit terhadap Induksi Ketahanan dan Pertumbuhan serta Hasil Padi pada Percobaan Rumah Kaca 12
Penyiapan tanaman uji 12
Perlakuan bakteri endofit 12
Peubah pengamatan 12
Rancangan percobaan 16
Identifikasi Bakteri Endofit Potensial sebagai Agens Penginduksi Ketahanan Tanaman Padi terhadap Infeksi X. oryzae pv oryzae 16
Identifikasi berdasarkan karakter morfologi dan fisiologi 16 Identifikasi berdasarkan teknik molekuler 16
Analisis Data 17
4 HASIL DAN PEMBAHASAN 18
Sampel Tanaman Padi untuk Isolasi Bakteri Endofit 18
Bakteri Endofit Hasil Isolasi 19
Isolat X. oryzae pv. oryzae 20
Seleksi Bakteri Endofit 21
Seleksi Bakteri Endofit Berdasarkan Viabilitas dan Morfologi
Seleksi Berdasarkan Kemampuan Induksi Ketahanan dan Pemacu
Pertumbuhan Padi di Pembibitan 21
Bakteri Endofit Hasil Seleksi Berdasarkan Uji HR 23 Pengaruh Aplikasi Bakteri Endofit terhadap Induksi Ketahanan dan
Pertumbuhan serta Hasil Panen Padi di Rumah Kaca 23 Pengaruh Aplikasi Bakteri Endofit terhadap Ekspresi Gen PR1 dan
PBZ1 23
Pengaruh Aplikasi Bakteri Endofit terhadap Aktivitas Enzim
Peroksidase 27
Pengaruh Aplikasi Bakteri Endofit terhadap Periode Inkubasi HDB 29 Pengaruh Aplikasi Bakteri Endofit terhadap Perkembangan penyakit
HDB 30
Pengaruh Aplikasi Bakteri Endofit terhadap Perkecambahan, Tinggi Tanaman, Anakan Maksimum, Anakan Produktif, Bobot Kering, dan
Kadar Air Gabah 31
Hasil Identifikasi Bakteri Endofit yang Berpotensi sebagai Agens
Penginduksi Ketahanan Tanaman Padi terhadap X. oryzae pv. oryzae 35
6 PEMBAHASAN UMUM 39
5 SIMPULAN DAN SARAN 41
Simpulan 41
Saran 41
7 DAFTAR PUSTAKA 42
LAMPIRAN 49
RIWAYAT HIDUP 59
DAFTAR TABEL
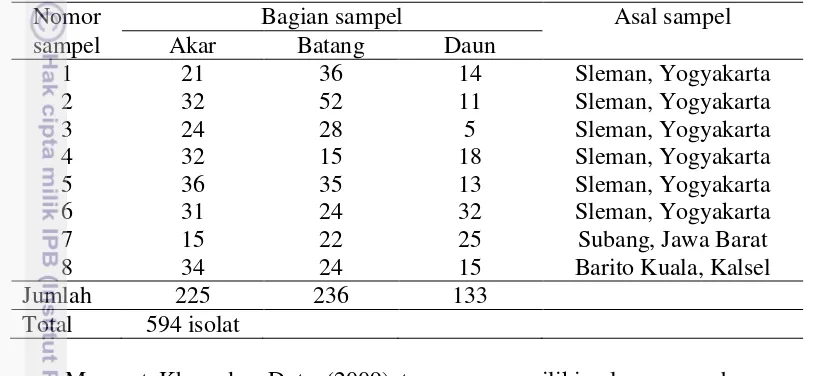
1 Distribusi isolat bakteri endofit 19
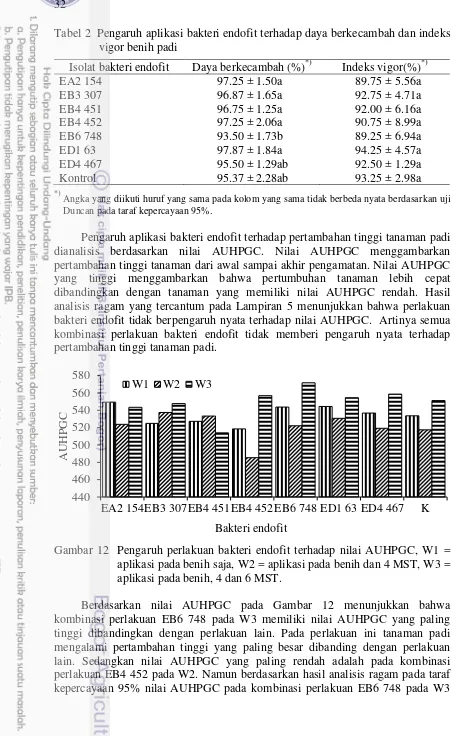
2 Pengaruh aplikasi bakteri endofit terhadap daya berkecambah dan
indeks vigor benih 32
3 Ciri morfologi isolat bakteri endofit kandidat agens penginduksi
ketahanan tanaman padi 36
4 Tingkat kemiripan basa nukleotida 16S rRNA isolat EB4 451 dengan isolat B. subtilis yang terdaftar pada GenBank berdasarkan hasil
DAFTAR GAMBAR
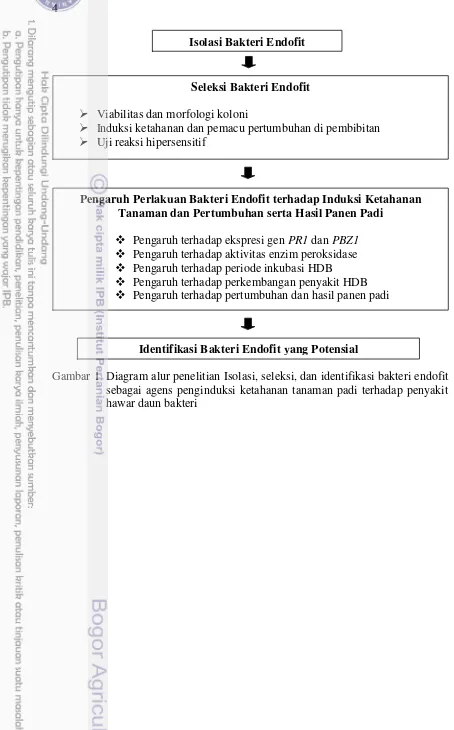
1 Diagram alur penelitian isolasi, seleksi, dan identifikasi bakteri endofit sebagai agens penginduksi ketahanan tanaman padi terhadap penyakit
hawar daun bakteri 4
2 Komposisi isolat bakteri endofit berdasarkan asal bagian tanaman 19 3 Koloni isolat X. oryzae pv oryzae patotipe IV (A), hasil uji reaksi
hipersensitif isolat X. oryzae pv oryzae patotipe IV (B), hasil uji patogenisitas isolat X. oryzae pv oryzae patotipe IV pada tanaman padi
(C) 20
4 Pengaruh perlakuan bakteri endofit terhadap rata-rata panjang hawar
daun bakteri pada tanaman padi di pembibitan 21
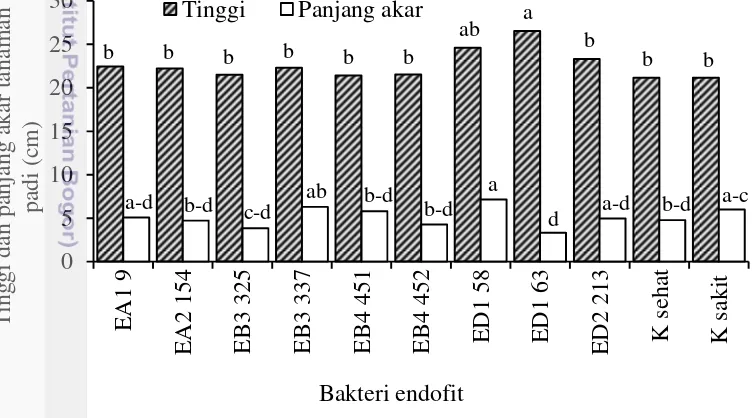
5 Pengaruh perlakuan bakteri endofit terhadap tinggi dan panjang akar
tanaman padi di pembibitan 22
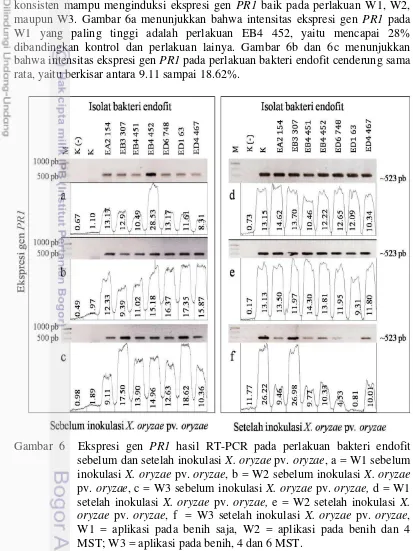
6 Ekspresi gen PR1 hasil RT-PCR pada perlakuan bakteri endofit sebelum dan setelah inokulasi X. oryzae pv. oryzae, a = W1 sebelum inokulasi X. oryzae pv. oryzae, b = W2 sebelum inokulasi X. oryzae pv. oryzae, c = W3 sebelum inokulasi X. oryzae pv. oryzae, d = W1 setelah inokulasi X. oryzae pv. oryzae, e = W2 setelah inokulasi X. oryzae pv. oryzae, f = W3 setelah inokulasi X. oryzae pv. oryzae, W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4 MST; W3 = aplikasi
pada benih, 4 dan 6 MST 24
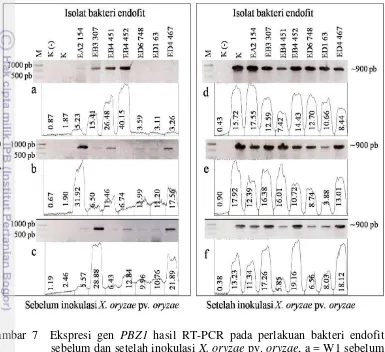
7 Ekspresi gen PBZ1 hasil RT-PCR pada perlakuan bakteri endofit sebelum dan setelah inokulasi X. oryzae pv. oryzae, a = W1 sebelum inokulasi X. oryzae pv. oryzae, b = W2 sebelum inokulasi X. oryzae pv. oryzae, c = W3 sebelum inokulasi X. oryzae pv. oryzae, d = W1 setelah inokulasi X. oryzae pv. oryzae, e = W2 setelah inokulasi X. oryzae pv. oryzae, f = W3 setelah inokulasi X. oryzae pv. oryzae, W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4 MST; W3 = aplikasi
pada benih, 4 dan 6 MST 25
8 Aktivitas enzim peroksidase pada 3 waktu aplikasi bakteri endofit. W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4 MST, W3 =
aplikasi pada benih, 4 dan 6 MST 28
9 Pengaruh perlakuan bakteri endofit terhadap periode inkubasi HDB. W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4 MST,
W3 = aplikasi pada benih, 4 dan 6 MST 29
10 Gejala hawar daun bakteri setelah inokulasi X. oryzae pv oryzae. (A)
gejala awal dan (B) gejala lanjutan 30
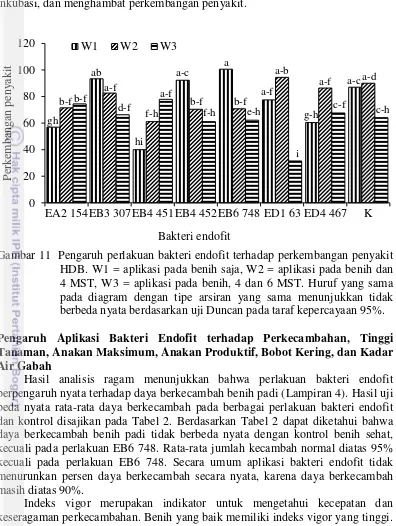
11 Pengaruh perlakuan bakteri endofit terhadap perkembangan penyakit HDB. W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4
MST, W3 = aplikasi pada benih, 4 dan 6 MST 31
12 Pengaruh perlakuan bakteri endofit terhadap nilai AUHPGC, W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4 MST, W3 =
aplikasi pada benih, 4 dan 6 MST 32
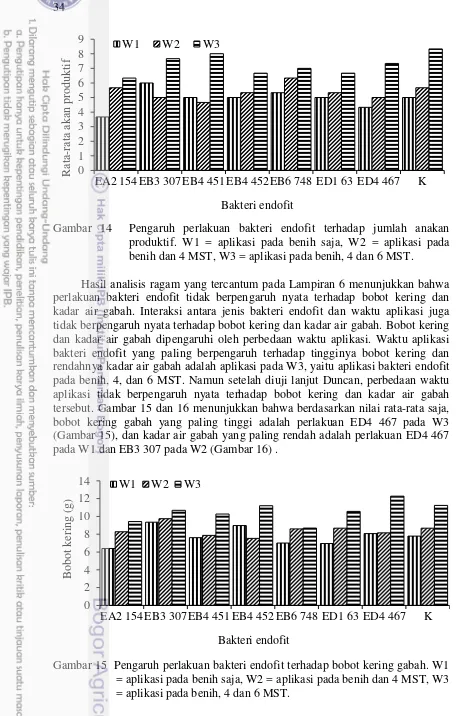
14 Pengaruh perlakuan bakteri endofit terhadap jumlah anakan produktif. W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4 MST,
W3 = aplikasi pada benih, 4 dan 6 MST 34
15 Pengaruh perlakuan bakteri endofit terhadap bobot kering gabah. W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4 MST, W3 =
aplikasi pada benih, 4 dan 6 MST 34
16 Pengaruh perlakuan bakteri endofit terhadap kadar air gabah. W1 = aplikasi pada benih saja, W2 = aplikasi pada benih dan 4 MST, W3 =
aplikasi pada benih, 4 dan 6 MST 35
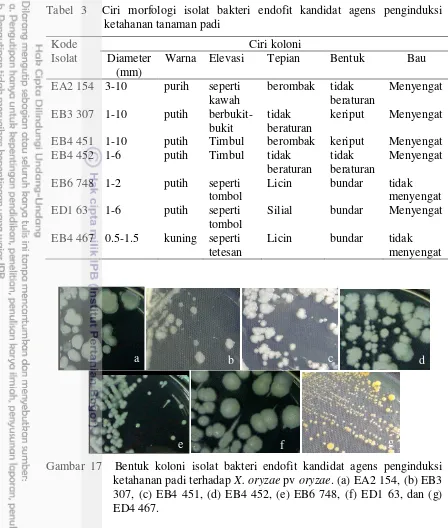
17 Bentuk koloni isolat bakteri endofit kandidat agens penginduksi ketahanan padi terhadap X. oryzae pv oryzae. (a) EA2 154, (b) EB3 307, (c) EB4 451, (d) EB4 452, (e) EB6 748, (f) ED1 63, dan (g) ED4
467 36
18 Hasil pewarnaan Gram isolat EB4 451 yang menunjukkan hasil sebagai
Gram positif 37
19 Hasil pewarnaan endospora isolat EB4 451 yang menunjukkan dapat
menghasilkan endospora 37
DAFTAR LAMPIRAN
1 Analisis ragam pengaruh aplikasi bakteri bakteri endofit terhadap
induksi ketahanan tanaman padi di pembibitan 51
2 Analisis ragam pengaruh aplikasi bakteri endofit terhadap pertumbuhan
tanaman padi di pembibitan 51
3 Analisis ragam pengaruh aplikasi bakteri endofit terhadap periode
inkubasi dan perkembangan penyakit HDB 52
4 Analisis ragam pengaruh perlakuan bakteri endofit terhadap daya
berkecambah (DB) dan indeks vigor (IV) benih 52
5 Analisis ragam pengaruh perlakuan bakteri endofit terhadap pertambahan tinggi (AUDPC), jumlah anakan maksimum, anakan
produktif 53
6 Analisis ragam pengaruh perlakuan bakteri endofit terhadap bobot
kering dan kadar air gabah 53
7 Urutan basa nukleotida isolat EB4 451 (teridentifikasi sebagai B.
subtilis) 54
8 Penjajaran basa nukleotida isolat EB4 451 dengan isolat B. subtilis yang terdaftar pada GenBank dengan nomor aksesi HM753632.1,
1 PENDAHULUAN
Latar Belakang
Salah satu penyakit tanaman yang menjadi masalah besar dalam produksi padi adalah penyakit hawar daun bakteri (HDB) yang disebabkan Xanthomonas oryzae pv. oryzae. Penyakit ini dapat menyebabkan kehilangan hasil antara 20 sampai 30%, bahkan dapat mencapai 50% (Verdier et al. 2011). Penyakit ini dapat menyerang mulai dari fase vegetatif sampai generatif. Sembiring (2011) menjelaskan bahwa penyakit ini menghasilkan dua gejala yang khas, yaitu kresek dan hawar. Kresek adalah gejala yang terjadi pada tanaman yang berumur kurang dari 30 hari dan biasanya muncul pada persemaian atau tanaman baru pindah tanam. Gejala ini berupa daun berwarna hijau kelabu, melipat, dan menggulung. Sedangkan hawar merupakan gejala yang umum terjadi pada fase tumbuh anakan sampai pemasakan. Gejala diawali dengan bercak kuning kemudian berubah menjadi kelabu dan putih jerami pada ujung dan tepian daun pada daun yang telah berkembang sempurna. Bercak meluas sepanjang tulang daun, bergabung, mengalami klorosis, kemudian menjadi nekrosis dan mengering.
Upaya pengendalian HDB sejauh ini mencakup kultur teknis, perakitan varietas tahan, penggunaan bakterisida, dan penggunaan agens hayati. Pengaturan jarak tanam dan pemupukan yang seimbang berperan penting dalam upaya pengendalian penyakit. Beberapa galur tahan HDB telah dikembangkan seperti yang dijelaskan Dewi et al. (2007). Varietas tahan seperti Angke, Conde, Cigeulis, Cibodas, Kanawe, dan Singkil juga mulai dirilis (Suprihanto et al. 2010). Namun ada kelemahan dari penggunaan varietas tahan dalam pengendalian penyakit HDB, yaitu ketahanan mudah terpatahkan oleh munculnya patotipe baru dari X. oryzae pv. oryzae. Suparyono dan Suprihanto (2004) menjelaskan bahwa sejauh ini di Indonesia terdapat 12 patotipe X. oryzae pv. oryzae. Patotipe yang mendominasi diantaranya adalah patotipe III, IV, dan VIII. Di lain pihak penggunaan bakterisida juga mulai dibatasi karena memiliki dampak negatif seperti terjadinya resistensi organisme sasaran terhadap antibiotik yang merupakan bahan dasar bakterisida serta terjadinya pencemaran lingkungan.
isolat bakteri endofit dari Sophora alopecuroides yang dapat menekan Verticillium sp. (Lin et al. 2013), dan masih banyak yang lainnya. Selain itu beberapa bakteri endofit juga dilaporkan memiliki kemampuan memacu pertumbuhan tanaman seperti bakteri endofit asal padi gogo yang mampu memacu pertumbuhan padi (Munif et al. 2012), bakteri endofit pemacu pertumbuhan tomat (Munif et al. 2000), dan bakteri endofit pemacu pertumbuhan cabai (Sundaramoorthy et al. 2012).
Salah satu mekanisme bakteri endofit yang dapat dimanfaatkan dalam pengendalian penyakit tanaman adalah melalui induksi ketahanan tanaman atau induced systemic resistance (ISR). ISR adalah fenomena dimana terjadi peningkatan ketahanan tanaman terhadap infeksi patogen setelah terjadinya rangsangan baik oleh faktor biotik maupun abiotik. Rangsangan tersebut menyebabkan kondisi fisiologis yang mengatur sistem ketahanan yang dimiliki tanaman menjadi aktif. Secara alami tanaman memiliki beragam respon pertahanan terhadap serangan patogen. Reaksi pertahanan ini meliputi penebalan dinding sel, akumulasi metabolit sekunder antimikrob dan ekspresi pathogenesis related (PR) protein (Kim et al. 2001). Induksi ketahanan oleh bakteri endofit dapat meningkatkan ketahanan dasar yang dimiliki tanaman terhadap serangan patogen. Bakteri endofit diindikasi dapat menginduksi ketahanan tanaman pertama kali diketahui pada tahun 1991 yaitu Pseudomonas fluoresens G8-4 yang ditemukan dalam jaringan tanaman dan dapat menginduksi ketahanan mentimun terhadap penyakit antraknosa (Wei et al. 1991). Pada tahun 1996 dilaporkan bahwa Bacillus pumilus INR7 yang diisolasi dari tanaman mentimun dapat meningkatkan ketahanan tanaman terhadap penyakit layu yang disebabkan Erwinia tracheiphila (Wei et al. 1996). Selain itu B. subtilis juga dilaporkan dapat menginduksi ketahanan tomat terhadap Cucumber mosaic virus (CMV) (Zehnder et al. 2000) dan beberapa bakteri endofit dapat meningkatkan ketahanan bawang merah terhadap penyakit HDB (Resti et al. 2013). Pada padi, induksi ketahanan tanaman terhadap HDB baru diteliti menggunakan rizobakteri sekitar perakaran (Khaeruni et al. 2014). Penggunaan bakteri endofit sebagai agens penginduksi ketahanan tanaman padi terhadap HDB belum banyak diketahui. Berdasarkan informasi tersebut penelitian yang telah dilaksanakan ini bertujuan mendapatkan isolat bakteri endofit asal tanaman padi, menyeleksi dan menguji kemampuan isolat bakteri endofit sebagai agens penginduksi ketahanan tanaman serta pengaruhnya terhadap pertumbuhan dan hasil panen padi, kemudian mengetahui bakteri endofit yang potensial sebagai agens penginduksi ketahanan tanaman padi terhadap penyakit HDB.
Tujuan Penelitian
Penelitian ini dilaksanakan dengan tujuan: 1) Mengisolasi bakteri endofit asal tanaman padi,
2) Menyeleksi dan menguji kemampuan isolat bakteri endofit sebagai agens penginduksi ketahanan,
4) Mengidentifikasi bakteri endofit yang potensial sebagai agens penginduksi ketahanan tanaman terhadap penyakit HDB.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberi manfaat berupa infomasi bakteri endofit yang potensial sebagai agens penginduksi ketahanan tanaman terhadap X. oryzae pv. oryzae dan pengaruhnya terhadap sifat agronomi padi yang dapat dimanfaatkan dalam pengendalian HDB.
Hipotesis
Hipotesis dalam penelitian ini adalah:
1) Bakteri endofit dapat diisolasi dari bagian daun, batang, dan akar tanaman padi,
2) Aplikasi bakteri endofit hasil isolasi dapat menginduksi ketahanan tanaman terhadap X. oryzae pv. oryzae,
3) Semakin sering bakteri endofit diaplikasikan, maka semakin tinggi pengaruhnya terhadap penekanan X. oryzae pv. oryzae,
Gambar 1 Diagram alur penelitian Isolasi, seleksi, dan identifikasi bakteri endofit sebagai agens penginduksi ketahanan tanaman padi terhadap penyakit hawar daun bakteri
Isolasi Bakteri Endofit
Seleksi Bakteri Endofit
Viabilitas dan morfologi koloni
Induksi ketahanan dan pemacu pertumbuhan di pembibitan Uji reaksi hipersensitif
Identifikasi Bakteri Endofit yang Potensial
Pengaruh Perlakuan Bakteri Endofit terhadap Induksi Ketahanan Tanaman dan Pertumbuhan serta Hasil Panen Padi
Pengaruh terhadap ekspresi gen PR1 dan PBZ1 Pengaruh terhadap aktivitas enzim peroksidase Pengaruh terhadap periode inkubasi HDB
2 TINJAUAN PUSTAKA
Penyakit Hawar Daun Bakteri pada Padi
Hawar daun bakteri (HDB) merupakan penyakit yang menyerang pembuluh dan menyebabkan infeksi sistemik pada tanaman padi. Patogen hidup pada jaringan pembuluh terutama xilem, melakukan multiplikasi, kemudian menyebar ke seluruh jaringan tanaman (Nino-Liu et al. 2006). Penyakit HDB menghasilkan dua gejala yang khas, yaitu kresek dan hawar. Kresek adalah gejala yang terjadi pada tanaman yang berumur kurang dari 30 hari dan biasanya muncul pada persemaian atau tanaman baru pindah tanam. Gejala ini berupa daun berwarna hijau kelabu, melipat, dan menggulung. Sedangkan hawar merupakan gejala yang umum terjadi pada fase tumbuh anakan sampai pemasakan (Sembiring 2011). Gejala diawali dengan bercak kuning kemudian berubah menjadi kelabu dan putih jerami pada ujung dan tepi daun pada daun yang telah berkembang sempurna. Bercak meluas sepanjang tulang daun, bergabung, mengalami klorosis, kemudian menjadi nekrosis yang disebut hawar dan kemudian mengering.
Penyebab penyakit HDB pada padi adalah bakteri X. oryzae pv. oryzae (Swings et al. 1990). Semua spesies Xanthomonas merupakan patogen dan ditemukan berasosiasi dengan tumbuhan atau bahan tumbuhan (Agrios 2005). Bakteri ini berbentuk batang lurus dengan panjang 1.2 sampai 3.0 µm dan lebar 0.4 sampai 1.0 µm, bergerak dengan satu bulu cambuk polar, dan termasuk Gram negatif. Koloni bakteri pada media YDCA berwarna kuning, cembung, dan mukoid. Warna kuning dikarenakan bakteri memproduksi pigmen xanthomonadin (Schaad et al. 2000).
Bakteri X. oryzae pv. oryzae menginfeksi bagian daun dengan cara masuk melalui hidatoda pada ujung dan tepi daun (Nino-Liu et al. 2006). Bakteri dapat pula menginfeksi melalui luka-luka pada daun akibat pemotongan, luka akibat gesekan antar daun, dan luka akibat serangga. Menurut Singh dan Mathur (2004), bakteri X. oryzae pv. oryzae dapat terbawa benih dan bertahan dalam waktu yang cukup lama karena bakteri berada pada fase dorman ketika berada dalam benih. Bakteri ini terdapat dalam endosperma benih dan dapat bertahan di benih selama 0.16 sampai 0.9 tahun (Agarwal & Sinclair 1996) atau 2 sampai 6 bulan (Singh & Mathur 2004). Selain itu, penyebaran X. oryzae pv. oryzae di lapangan dapat dibantu oleh angin, gesekan antara daun, dan melalui percikan air hujan (Nino-Liu et al.2006)
Pengendalian penyakit kresek umumnya adalah dengan menanam varietas padi yang tahan (Hifni & Kardin 1998). Selain itu, juga dengan tidak melakukan pemotongan ujung daun pada bibit padi yang dipindah dari persemaian, dan pemupukan yang seimbang. Penggunaan senyawa kimia atau bakterisida, seperti nickel dimethyl dithiocarbamate, dithianone, phenazine, dan yang mangandung tembaga dapat digunakan untuk pengendalian jika benar-benar diperlukan (Gnanamanickam et al. 1994).
Peran Bakteri Endofit sebagai Agens Hayati
Mikroorganisme yang berada dalam jaringan tanaman memiliki peranan dalam mempengaruhi pertumbuhan, perkembangan, serta pertahanan tanaman. Mikroba tersebut dikenal dengan bakteri atau cendawan endofit. Bakteri endofit merupakan bakteri yang mengolonisasi bagian internal jaringan tanaman. Bakteri endofit dapat mengolonisasi relung ekologi yang sama dengan patogen tanaman, tetapi tidak menyebabkan kerusakan pada inangnya. Beberapa bakteri endofit dilaporkan dapat memacu pertumbuhan, meningkatkan kebugaran tanaman (Kirchhof et al. 2001) dan dapat menjadi agens biokontrol (Shimizu et al. 2009).
Beberapa laporan menunjukkan bahwa bakteri endofit banyak memberi keuntungan terhadap inangnya. Keberadaan bakteri endofit di dalam jaringan tanaman berperanan dalam perbaikan pertumbuhan tanaman karena kemampuannya menghasilkan zat pemacu pertumbuhan, memfiksasi nitrogen, memobilisasi fosfat, dan juga berperanan dalam kebugaran tanaman (plant health promotion). Bakteri endofit diduga mampu meningkatkan sistem pertahanan tanaman terhadap gangguan penyakit karena kemampuannya untuk memproduksi senyawa antimikroba, enzim, asam salisilat, etilen dan senyawa sekunder lainnya yang berperanan dalam menginduksi ketahanan tanaman (Backman & Sikora 2008).
Beberapa bakteri endofit telah dilaporkan dapat mengendalikan penyakit tanaman seperti Steptomyces spp. yang dapat mengendalikan X. oryzae pv. oryzae (Hastuti et al. 2012), kelompok Flexibacter-Cytophaga-Bacteroides yang dapat menekan Erwinia carotovora subsp. atroseptica pada tanaman kentang (Reiter et al. 2002), isolat bakteri endofit dari Sophora alopecuroides yang dapat menekan Verticillium sp. (Lin et al. 2013), dan masih banyak yang lainnya. Bakteri endofit dari beberapa genus seperti Pseudomonas, Bacillus dan Azospirillum, dilaporkan mampu meningkatkan pertumbuhan tanaman, menguraikan dinding sel patogen, dan menghambat pertumbuhan patogen dengan menghasilkan senyawa antimikroba seperti siderofor (Chandrashekhara et al. 2007). Bakteri endofit dapat melindungi inang dengan melawan patogen melului induksi pertahanan tanaman, sekresi zat yang bersifat antagonis terhadap patogen atau melalui kompetisi untuk memperoleh situs kolonisasi dan nutrisi (Reinhold-Hurek & Hurek 2011).
Potensi bakteri endofit dalam mengendalikan penyakit tanaman menyebabkan bakteri endofit diperhitungkan sebagai agens hayati. Bakteri endofit diindikasi dapat menginduksi ketahanan tanaman pertama kali diketahui pada tahun 1991 yaitu Pseudomonas fluoresens G8-4 yang ditemukan dalam jaringan tanaman dan dapat menginduksi ketahanan mentimun terhadap penyakit antraknosa (Wei et al. 1991). Pada tahun 1996 dilaporkan bahwa Bacillus pumilus INR7 yang diisolasi dari tanaman mentimun dapat meningkatkan ketahanan tanaman terhadap penyakit layu yang disebabkan Erwinia tracheiphila (Wei et al. 1996). Selain itu B. subtilis juga dilaporkan dapat menginduksi ketahanan tomat terhadap Cucumber mosaic virus (CMV) (Zehnder et al. 2000) dan beberapa bakteri endofit seperti B. cereus, Bacillus sp., dan Serratia marcescens dapat meningkatkan ketahanan bawang merah terhadap penyakit HDB yang disebabkan X. axonopodis pv. allii (Resti et al. 2013).
tanaman, lebih terlindungi dari stres akibat faktor abiotik, menempati relung yang sama dengan patogen pada umumnya, dan proses translokasi senyawa metabolit ke dalam jaringan tanaman yang lebih baik (Hallmann et al. 1997). Selain sebagai agens pengendali patogen, bakteri endofit juga memiliki kemampuan memacu pertumbuhan tanaman. Beberapa bakteri endofit yang dilaporkan dapat memacu pertumbuhan tanaman adalah bakteri endofit asal padi gogo yang mampu memacu pertumbuhan padi (Munif et al. 2012), tomat (Munif et al. 2000), dan cabai (Sundaramoorthy et al. 2012).
Induksi Ketahanan Tanaman Padi
Tanaman akan bereaksi terhadap kehadiran suatu patogen dengan mengaktifkan suatu sistem pertahanan. Pengaktifan sistem pertahanan diawali dengan sinyal transduksi yang mengubah stimulus menjadi bentuk lain dengan melibatkan urutan reaksi biokimia di dalam sel yang dilaksanakan oleh enzim dan berhubungan melalui second messenger (Voet & Donald 1995). Ada tiga mekanisme utama dalam proses penerimaan sinyal dan inisiasi transduksi sel dalam merespon sinyal dari lingkungan, yaitu penerimaan sinyal oleh sel target, penguatan sinyal, dan respon seluler terhadap sinyal. Sebagai contoh tanaman padi yang terinduksi oleh serangan patogen blas akan mengaktifkan kelompok gen yang menyandi PR protein (Gee et al. 2001).
Induksi ketahanan tanaman merupakan ketahanan yang berkembang setelah tanaman diinokulasi dengan agens biotik, senyawa kimia, atau perlakuan fisik. Tingkat ketahanan tanaman menjadi meningkat terhadap berbagai serangan patogen. Sebagai contoh infeksi Tobacco mosaic virus (TMV) pada tanaman tembakau menyebabkan ketahanan sistemik terhadap patogen itu sendiri dan terhadap beberapa jenis virus lain, terhadap Phytophthora sp., terhadap Pseudomonas tabaci, serta terhadap serangan kutu daun (Agrios 2005). Induksi ketahanan juga dapat dilakukan dengan cara menginokulasi tanaman dengan spora cendawan atau bakteri yang telah dimatikan dengan perlakuan panas, dan inokulasi patogen pada fase tanaman belum rentan terhadap patogen. Agar ketahanan terinduksi dapat muncul, maka harus ada lag period antara pemberian agens penginduksi dan inokulasi tantangan (challenge inoculation). Waktu tersebut dibutuhkan untuk menyintesis dan mendistribusikan zat-zat secara sistemik dari bagian tanaman yang diberi perlakuan agens penginduksi ke bagian lain tanaman tersebut. Ketahanan terinduksi yang bersifat lokal biasanya berkembang 2 sampai 3 hari setelah perlakuan, sedangkan ketahanan terinduksi yang bersifat sistemik berkembang 7 hari setelah perlakuan atau bahkan 3 sampai 5 minggu. Ketahanan terinduksi dapat berkembang jika sel-sel inang mampu melakukan transkripsi dan menghasilkan enzim atau protein yang mengaktifkan gen yang bertanggung jawab dalam mekanisme pertahanan tanaman tersebut. Peningkatan aktivitas enzim peroksidase, fenilalanin amonia liase, fitoaleksin, lignifikasi, dan proteinase inhibiting juga ditemukan pada beberapa tanaman yang mengalami peningkatan pertahanan terinduksi (Agrios 2005).
maupun secara tidak langsung (induksi ketahanan) (Janse 2005). Bakteri nonpatogenik dapat menginduksi respons induce systemic resistance (ISR) pada tanaman sebagaimana patogen dapat menginduksi sistem pertahanan systemic acquired resistance (SAR). ISR terinduksi oleh bakteri nonpatogenik yang bergantung pada respon dari asam jasmonat dan etilen. ISR efektif melawan patogen dengan dibatasi oleh asam jasmonat dan etilen sebagai mekanisme pertahanan dasar. Adanya induksi ketahanan dapat meningkatkan ketahananan dasar inang dalam menghadapi patogen (Chaudary et al. 2007).
ISR pada dasarnya memiliki kesamaan dengan SAR. Perbedaan antara ISR dengan SAR adalah ISR tidak menyebabkan adanya gejala tampak seperti lesio nekrotik (Compant et al. 2005). Ramamoorthy et al. (2001) memaparkan bahwa mekanisme ISR terjadi sebagai akibat perubahan fisiologi tanaman yang kemudian menstimulasi terbentuknya senyawa kimia yang berguna dalam pertahanan terhadap serangan patogen. Perubahan fisiologi tersebut dapat berupa modifikasi struktural dinding sel atau perubahan reaksi biokimia pada tanaman inang. Beberapa faktor yang dapat menyebabkan adanya induksi ketahanan sistemik oleh bakteri yaitu adanya sumbangan lipopolisakarida oleh bakteri, produksi siderofor, dan produksi asam salisilat yang dapat terjadi secara langsung oleh bakteri ataupun secara tidak lansung (Van Loon et al.1998).
Gen PR1 dan PBZ1 merupakan PR protein, yaitu gen-gen yang aktif sebagai reaksi tanaman terhadap infeksi patogen. PR protein ini dapat menghambat patogenesis, mendegradasi dinding sel patogen yang tersusun oleh polisakarida, meningkatkan ketahanan terhadap beberapa patogen tertentu, dan bersifat anti mikroba (Gee et al. 2001). Menurut Agrawal et al. (2001) etilen terlibat dalam ekspresi gen PR yang terdapat pada tanaman dikotil. Etilen juga mempengaruhi tiga gen PR penting pada padi (kultivar Nipponbare) seperti gen PR1, PR5, dan PR10 terhadap perkecambahan. Hasil ini menunjukkan adanya suatu peran etilen terhadap induksi gen PR pada tanaman padi.
PR1 merupakan gen yang pertama kali berhasil diidentifikasi dan merupakan kelompok yang paling dominan dari PR protein. Protein ini berfungsi sebagai anticendawan (Selitrennikoff 2001). Protein PR1 terakumulasi oleh infeksi patogen, induksi bahan kimia seperti salicylic acid (SA) dan benzo (1,2,3) thiadiazole-7-carbothioic acid S-methyl ester (BTH) (Kim et al. 2002). Protein PR1 terakumulasi dalam jumlah yang tinggi setelah terjadi infeksi oleh patogen. Gen PR1 ini ditemukan di padi, gandum, tomat, tembakau, Arabidopsis thaliana, barley dan beberapa tanaman lainnya. PR1 terakumulasi oleh infeksi patogen diantaranya Uromyces fabae, Phytophthora infestans, Erysiphe graminis (Selitrennikoff 2001). Pada tanaman padi gen PR1 diinduksi oleh Bipolaris sorokiniana, P. syringae pv. syringae (Schweizer et al. 1997) dan Magnaporthe grisea (Suharsono et al. 2002). Protein ini juga terinduksi oleh bahan kimia seperti salicylic acid (SA), methyl 1-2,6-dichloroisonicotinic acid (INA) dan benzo (1,2,3) thiadiazole-7-carbothioic acid S-methyl ester (BTH) (Kim et al. 2001).
pada tanaman, seperti peroksidase, polyphenoloxidase, ammonia-lyase dan catechol-O-methyltransferase serta asam α-linolenik yang berfungsi sebagai penghambat perkecambahan konidia cendawan (Nakashita et al. 2001). Gen PBZ1 pada tanaman padi juga dapat diinduksi oleh infeksi patogen M. grisea, namun tidak oleh pelukaan biasa saja (Midoh & Iwata 1996).
3 METODE PENELITIAN
Tempat dan Waktu
Penelitian dilaksanakan di Laboratorium Bakteriologi Tumbuhan Departemen Proteksi Tanaman dan rumah kaca Cikabayan, IPB, mulai Juni 2013 sampai Desember 2014.
Bahan dan Alat
Bahan dan alat yang digunakan dalam penelitian ini adalah sampel tanaman padi sehat sebagai sumber bakteri endofit, media agar dan cair untuk isolasi serta pertumbuhan bakteri endofit, isolat X. oryzae pv. oryzae patotipe IV, benih padi sehat varietas Ciherang, media tanam, bahan untuk analisis ekspresi gen PR1 dan PBZ1, bahan untuk pengujian aktivitas enzim peroksidase, primer universal untuk identifikasi bakteri endofit, serta alat dan bahan untuk uji Gram dan pewarnaan endospora.
Metode
Pengambilan Sampel Tanaman Padi untuk Isolasi Bakteri Endofit
Pengambilan sampel tanaman padi dilakukan di tiga lokasi berbeda yang mewakili 3 tipe sawah di Indonesia, yaitu lahan sawah tadah hujan, lahan sawah irigasi, dan lahan sawah rawa pasang surut. Sampel tanaman padi yang diambil merupakan tanaman sehat yang berada diantara tanaman padi yang terserang penyakit hawar daun bakteri. Lokasi pengambilan sampel untuk lahan sawah tadah hujan dilakukan di dua lokasi yaitu di Desa Widodomartani, Kecamatan Ngemplak dan Desa Harjobinangun, Kecamatan Tratas, Kabupaten Sleman, Yogyakarta. Pengambilan sampel untuk lahan sawah irigasi dilakukan di Desa Gempol Sari, Kecamatan Patok Beusi, Kabupaten Subang, Jawa Barat. Lokasi pengambilan sampel terakhir adalah di Desa Karang Indah, Kecamatan Mandastana, Kabupaten Barito Kuala, Kalimantan Selatan.
Isolasi Bakteri Endofit
1 L) sebagai kontrol. Sterilisasi permukaan dianggap berhasil apabila kontrol tidak ditumbuhi oleh koloni bakteri.
Akar, batang, dan daun digerus secara terpisah dengan mortar dan ditambahkan 9 mL larutan fisiologis (0.85% NaCl). Ekstrak akar, batang dan daun diencerkan sampai pengenceran 10-4. Sebanyak 0.1 mL ekstrak dari pengenceran 10-3 dan 10-4 disebar pada media Tryptic Soy Agar (TSA) 5% (0.75 g pancreatic digest of casein, 0.25 g enzymatic digest of soybean meal, 0.75 g sodium chloride, 15 g agar, dan akuades sampai dengan volume 1 L), TSA 100% (15 g pancreatic digenst of casein, 5 g enzymatic digest of soybean meal, 15 g sodium chloride, 15 g agar, dan akuades sampai dengan volume 1 L), NA, King’s B 100% (20 g protease peptone, 1.5 g K2HPO4, 1.5 g MgSO4.7H2O, 15 ml gliserol, 15 g agar, dan akuades sampai volume 1 L), water-yeast extract-agar (WYE) (0.25 g yeast extract, 0.5 g K2HPO4, 15 g agar, dan akuades sampai volume 1 L), dan casamino acids-yeast extract-glucose agar (YCED) (0.3 g yeast extract, 0.3 g casamino acid, 0.3 g D-glukose, 2 g K2HPO4, dan akuades samapi volume 1 L) dan diinkubasi pada suhu ruang selama 48 jam. Koloni bakteri yang tumbuh dimurnikan kemudian disimpan dalam akuades steril.
Penyiapan Isolat X. oryzae pv. oryzae
Isolat bakteri X. oryzae pv oryzae patotipe IV diperoleh dari Balai Besar Penelitian Padi. Isolat diremajakan pada media Wakimoto (kaldu dari 300 g kentang, 7 g bacto peptone, 17 g sukrosa, 0.5 g Ca(NO3)2.4H2O, 1 g Na2HPO4.12 H2O, 17 g agar, dan akuades sampai volume 1 L). Uji reaksi hipersensitif (HR) pada tanaman tembakau dilakukan sebagai konfirmasi bahwa isolat X. oryzae pv oryzae yang digunakan merupakan bakteri patogen tumbuhan penyebab hawar daun bakteri.
Uji HR dilakukan pada tanaman tembakau sehat mengikuti metode yang digunakan Wahyudi et al. (2011). Suspensi X. oryzae pv oryzae patotipe IV dibuat dengan cara mengambil bakteri sebanyak satu ujung jarum ose ke dalam 5 mL media Wakimoto cair. Inokulasi dilakukan dengan cara menyuntikkan suspensi menggunakan jarum suntik volume 1 mL ke bagian permukaan bawah daun tanpa menembus lapisan daun bagian atas. Gejala diamati maksimal 24 jam setelah inokulasi.
Uji patogenisitas dilakukan dengan cara menyiapkan tanaman padi (IR64) sehat beserta suspensi bakteri X. oryzae pv oryzae patotipe IV. Inokulasi dilakukan dengan cara menggunting bagian daun menggunakan gunting steril yang sebelumnya dicelupkan ke dalam suspensi bakteri X. oryzae pv oryzae patotipe IV. Tanaman padi disungkup untuk menjaga kelembapan. Gejala diamati setiap hari sampai 1 minggu setelah inokulasi.
Seleksi Isolat Bakteri Endofit Potensial
Uji induksi ketahanan dan pemacu pertumbuhan di pembibitan. Bakteri endofit hasil seleksi berdasarkan viabilitas dan morfologi koloni diseleksi kembali berdasarkan pengaruhnya terhadap induksi ketahanan dan pemacu pertumbuhan padi di pembibitan. Isolat bakteri yang akan diuji ditumbuhkan pada media NA. Biakan yang berumur 24 jam kemudian disuspensikan dalam media luria bertani broth (LB) (10 g tryptone, 5 g NaCl, 5 g yeast extract, dan akuades sampai volume 1 L). Benih padi varietas Ciherang yang telah disterilisasi permukaan pada suhu 55 oC selama 20 menit dan direndam dalam suspensi bakteri endofit selama 13 jam ditanam pada media pasir dan kompos (1:1), kemudian ditempatkan dalam growth chamber. Setelah berumur 3 minggu, bagian ujung daun kedua dipotong menggunakan gunting yang telah dicelupkan dalam suspensi X. oryzae pv oryzae patotipe IV dengan kerapatan 108 sel mL-1. Peubah yang diamati terdiri atas panjang hawar, tinggi tanaman, serta panjang akar. Rancangan penelitian menggunakan Rancangan Acak Lengkap. Setiap perlakuan sebanyak 3 ulangan, dan setiap ulangan sebanyak 1 unit tanaman.
Uji HR. Isolat bakteri endofit hasil seleksi di pembibitan diseleksi kembali berdasarkan uji HR pada daun tembakau. Pengujian mengikuti metode yang digunakan oleh Wahyudi et al. (2011). Uji HR dilakukan dengan cara mengambil bakteri sebanyak 1 ujung jarum ose ke dalam 5 mL media Wakimoto cair, kemudian diinkubasi selama 24 jam. Inokulasi dilakukan dengan cara menyuntikan suspensi menggunakan jarum suntik volume 1 mL pada bagian permukaan bawah daun tanpa menembus lapisan daun bagian atas. Gejala diamati maksimal 24 jam setelah inokulasi.
Pengaruh Aplikasi Bakteri Endofit terhadap Induksi Ketahanan dan Pertumbuhan serta Hasil Padi pada Percobaan Rumah Kaca
Penyiapan tanaman uji. Bibit padi yang berumur 2 minggu dipindah tanam pada tanah sawah steril dalam ember berukuran 13.5 cm x 11.5 cm. Pemupukan dilakukan berdasarkan dosis anjuran mengikuti metode Sasmita et al. (2006), yaitu 250 kg ha-1 urea, 100 kg ha-1 SP-36, dan 100 kg ha-1 KCl.
Perlakuan bakteri endofit. Bakteri endofit diaplikasikan pada 3 waktu aplikasi yang berbeda, yaitu W1 (aplikasi bakteri endofit pada benih saja), W2 (aplikasi bakteri endofit pada benih dan 4 MST), dan W3 (aplikasi bakteri endofit pada benih, 4, dan 6 MST). Inokulasi X. oryzae pv. oryzae dilakukan pada akhir masa vegetatif dengan cara yang sama seperti pada percobaan di pembibitan. Jumlah anakan yang digunting untuk setiap unit tanaman adalah 5 anakan, dan selanjutnya tanaman disungkup selama 3 hari. Setiap perlakuan sebanyak 3 ulangan, dan setiap ulangan sebanyak 1 unit tanaman. Kerapatan suspensi bakteri endofit pada waktu aplikasi perendaman benih maupun pada 4 dan 6 MST berkisar antara 107 sampai 109 sel mL-1. Aplikasi pada 4 dan 6 MST dilakukan dengan cara penyiraman di sekitar perakaran dan penyemprotan pada seluruh permukaan tanaman. Volume suspensi bakteri yang digunakan untuk penyiraman sebanyak 30 mL dan yang digunakan untuk penyemprotan sebanyak 20 mL.
vigor benih, pertambahan tinggi tanaman, jumlah anakan maksimum, jumlah anakan produktif, bobot kering, dan kadar air gabah.
Ekspresi gen PR1 dan PBZ1 dianalisis dengan teknik RT-PCR mengacu pada metode Kurnianingsih (2008). Isolasi RNA total dilakukan dari tanaman yang diberi perlakuan bakteri endofit sebelum dan setelah diinokulasi X. oryzae pv. oryzae menggunakan peqGOLD Plant RNA kit (peqlab). Sebanyak 0.1 g sampel padi digerus dalam keadaan dingin, kemudian ditambahkan 450 µl RNA lysis buffer T. Inkubasi dilakukan pada suhu ruang selama 30 menit dan setiap 10 menit dihomogenkan menggunakan vorteks selama 10 detik. Larutan dimasukkan dalam tabung mikro volume 2 mL yang sudah dipasang DNA removing column. Sentrifugasi dilakukan selama 2 menit dengan kecepatan 10 000 g. DNA removing column dibuang, sedangkan tabung mikro disimpan dan ditambahkan 70% etanol sebanyak volume yang sama dengan larutan. Larutan kemudian dimasukkan dalam perfect bind RNA column. Sentrifugasi dilakukan dengan kecepatan 10 000 g selama 1 menit. Perfect bind RNA column dimasukkan pada tabung mikro baru dan ditambahkan 500 µL RNA wash buffer I, kemudian disentrifugasi dengan kecepatan 10 000 g selama 1 menit. Larutan dibuang, sedangkan tabung mikro dan saringan digunakan kemabli. Sebanyak 650 µL RNA wash buffer II ditambahkan dan Perfect bind RNA column disentrifugasi selama 1 menit dengan kecepatan 10 000 g. Larutan dibuang kemudian dimasukkan kembali 650 µL RNA wash buffer II. Perfect bind RNA column disentrifugasi selama 1 menit dengan kecepatan 10 000 g dan cairan dibuang kembali. Column ditempatkan pada tabung mikro yang sama dan disentrifugasi selama 2 menit pada kecepatan 10 000 g, selanjutnya ditempatkan dalam 1.5 mL tabung mikro baru. Sebanyak 30 sampai 80 µL RNAse free water kemudian diinkubasi selama 1 menit. Tahap terakhir adalah sentrifugasi selama 1 menit dengan kecepatan 10 000 g.
Amplifikasi gen PR1 dan PBZ1 menggunakan TransScriptTM II One-Step RT-PCR SuperMix (TransGen Biotech). Amplifikasi gen PR1 menggunakan primer spesifik PR1 forward (5’-TAACTATGGAGGTATCCAAGCTGCC-3’) dan primer reverse (5’-CCAGTACGTACGCCCGTGTGTATAA-3’) dengan target amplikon berukuran ± 523 pb (Kurnianingsih 2008). Amplifikasi gen PBZ1
menggunakan primer spesifik PBZ1 forward (5’
-CAGTGGTCAGTAGAGTGATC-3’) dan primer PBZ1 reverse (5’ -CTGGATAGAGGCAGTATTCC-3’) dengan target amplikon berukuran ± 900 pb (Midoh & Iwata 1996).
Program PCR yang digunakan mengacu pada Kurnianingsih (2008) yaitu reverse transcriptase pada 42 ºC selama 60 menit, predenaturasi pada suhu 94 ºC selama 5 menit, denaturasi pada suhu 94 ºC selama 30 detik, annealing pada suhu 55 ºC selama 30 detik, dan ekstensi pada suhu 72 ºC selama 2 menit, siklus denaturasi-ekstensi diulang sebanyak 39 kali, pasca PCR 72 ºC selama 5 menit dan pendinginan pada suhu 25 ºC selama 10 menit. Program PCR untuk deteksi gen PBZ1 sama dengan program PCR untuk deteksi gen PR1 kecuali tahap annealing yaitu pada suhu 56 ºC.
Hasil PCR diseparasi pada gel agarosa 1% (b v-1) di dalam larutan penyangga TAE 1x [(4.84 g Tris base, 1.142 mL glacial acetic acid dan 2 mL 0.5 M EDTA (pH 8.0)].
menganalisis gambar secara kuantitatif sehingga dapat mengilustrasikan perbandingan intensitas DNA gen PR1 dan PBZ1 antara satu perlakuan dengan perlakuan lainnya.
Analisis aktivitas enzim peroksidase dilakukan pada 2 hari setelah inokulasi X. oryzae pv. oryzae. Analisis menggunakan metode yang digunakan oleh Damayanti et al. (2007). Sebanyak 0.5 g sampel yang diambil secara komposit dari masing-masing perlakuan ditambahkan dengan 1.5 mL dari 0.1 M buffer fosfat pH 7.0 pada suhu 4 oC dan digerus menggunakan mortar dalam kondisi dingin. Suspensi diambil dan dimasukkan pada tabung berukuran 1.5 mL, kemudian disentrifugasi dengan kecepatan 16 000 g selama 15 menit. Supernatan yang terbentuk digunakan sebagai sumber enzim.
Sebanyak 1.5 mL dari 5 molal pyrogallol dan 0.5 mL dari 1% hydrogen peroxide (H2O2) dimasukan dalam supernatan. Campuran tersebut kemudian diinkubasi pada suhu ruang dan dihitung aktivitas enzim dengan menggunakan spektrofotometer pada penjang gelombang 420 nm dengan interval 30 detik sampai 3 menit. Aktivitas enzim peroksidase terekspresi sebagai perubahan nilai absorban min-1mg-1 protein. Total protein diukur dengan menggunakan Bradford reagent dengan bovine serum albumin (BSA; Sigma Aldrich, USA) sebagai standar.
Periode inkubasi diamati setiap hari setelah inokulasi patogen. Pengamatan berhenti setelah muncul gejala pertama pada setiap unit percobaan. Perkembangan penyakit diamati dengan menghitung panjang hawar setiap hari dan berhenti setelah gejala hawar sampai pada pangkal daun, selanjutnya dihitung keparahan penyakit dengan menggunakan rumus:
KP = keparahan penyakit
n = jumlah daun dari setiap kategori serangan v = kategori serangan
N = jumlah daun yang diamati V = nilai kategori serangan tertinggi
Nilai keparahan penyakit dihitung dengan skor kerusakan daun berdasarkan sistem evaluasi baku dari Standard Evaluation System for Rice (IRRI 1996). Kategori serangan X. oryzae pv. oryzae yang digunakan yaitu
Data seluruh keparahan penyakit dianalisis menggunakan formula Area Under Disease Progress Curve (AUDPC) (Van der Plank 1963), yaitu:
Ri = keparahan penyakit waktu i; ti = waktu ke-i
Pengujian pengaruh bakteri endofit terhadap daya berkecambah dan indeks vigor benih menggunakan metode uji kertas digulung dan didirikan dalam plastik (UKDdp) berdasarkan standar International Rules for Seed Testing Asosiation (ISTA 2010). Sebanyak 100 benih ditumbuhkan pada media kertas yang telah dibasahi dengan akuades steril dan dilapisi dengan plastik. Kertas kemudian digulung dan disimpan dalam germinator dengan kelembapan 80 sampai 90% dan suhu sekitar 26 sampai 30 oC. Indeks vigor dihitung berdasarkan jumlah kecambah normal pada 5 HST, sedangkan daya kecambah dihitung berdasarkan jumlah kecambah normal pada 7 dan 14 HST. Rumus yang digunakan untuk menghitung persen daya berkecambah (DB) adalah:
Sedangkan rumus untuk menghitung indeks vigor benih (IV) adalah:
DB = daya berkecambah IV = indeks vigor benih KN = kecambah normal
Pengamatan terhadap pertambahan tinggi tanaman dilakukan setiap minggu sampai 16 MST. Nilai pertambahan tinggi tanaman dianalisis menggunakan formula area under high of plant growth progress curve (AUHPGC) menggunakan rumus Van der Plank (1963) yang dimodifikasi, yaitu:
AUHPGC
Yi = pertambahan tinggi tanaman waktu i; ti = waktu ke-i
dengan cara gabah dikeringkan dalam oven pada suhu 60 oC selama 3 hari. Kadar air gabah ditentukan menggunakan rumus:
Kadar air gabah = x 100%
Rancangan percobaan. Rancangan yang digunakan dalam pengujian ini adalah rancangan 2 Faktorial dalam Rancangan Acak Lengkap. Faktor pertama adalah jenis isolat bakteri endofit yang terdiri atas 8 taraf, sedangkan faktor yang kedua adalah waktu aplikasi bakteri endofit terdiri atas 3 taraf. Kombinasi perlakuan yang diujikan sebanyak 24 kombinasi perlakuan. Masing-masing perlakuan terdiri atas 3 ulangan, dan masing-masing ulangan terdiri atas 1 unit tanaman. Pengujian daya berkecambah Rancangan penelitian menggunakan Rancangan Acak Lengkap. Masing-masing perlakuan terdiri atas 4 ulangan, masing-masing ulangan terdiri atas 100 unit, dan masing-masing unit terdiri atas 1 butir benih padi.
Identifikasi Bakteri Endofit Potensial sebagai Agens Penginduksi Ketahanan Tanaman Padi terhadap Infeksi X. oryzae pv oryzae
Identifikasi berdasarkan karakter morfologi dan fisiologi. Identifikasi dilakukan berdasarkan karakter morfologi koloni, pewarnaan Gram dan endospora. Morfologi koloni yang diamati meliputi diameter, warna, elevasi, tepian, bentuk, dan bau. Pewarnaan Gram dan endospora menggunakan metode yang digunakan Schoenhard (1978). Pewarnaan Gram dilakukan dengan cara sel bakteri endofit dioleskan pada permukaan kaca preparat, selanjutnya difiksasi panas di atas api sedang. Olesan bakteri digenangi pewarna primer yaitu kristal ungu selama 1 menit. Kelebihan pewarna dibuang, lalu dibilas menggunakan air. Kaca preparat ditiriskan kemudian digenangi iodium Gram selama 1 menit. Kelebihan pewarna dibuang, kemudian dibilas menggunakan air. Olesan bakteri dicuci dengan pemucat warna etanol 95% selama 30 detik, selanjutnya dicuci dengan air dan ditiskan. Olesan bakteri kembali digenangi dengan pewarna tandingan safranin selama 30 detik, kemudian dibilas menggunakan air. Kaca preparat ditiriskan, kemudian diamati dibawah mikroskop compound dengan perbesaran 1000 kali.
Pewarnaan endospora dilakukan dengan cara sel bakteri endofit dioleskan pada permukaan kaca preparat, selanjutnya difiksasi panas di atas api sedang. Olesan bakteri digenangi dengan hijau malakit. Masuknya pewarna ke dalam endospora dibantu dengan cara dipanaskan dengan api dari pembakar Bunsen sampai beruap selama 10 menit. Kelebihan pewarna dicuci dengan air. Olesan bakteri selanjutnya digenangi dengan safranin selama 1 menit. Safranin dicuci dengan air mengalir, selanjutnya kaca preparat ditiriskan kemudian diamati dibawah mikroskop compound dengan perbesaran 1000 kali.
Identifikasi bakteri endofit potensial dengan teknik molekuler. Identifikasi bakteri endofit potensial yang mampu menginduksi ketahanan tanaman padi terhadap X. oryzae pv. oryzae dilakukan berdasarkan runutan gen 16S rRNA. Isolasi DNA total menggunakan GeneJET Genomic DNA purification Kit (Thermo Scientific). Isolasi DNA diawali dengan penyiapan kultur bakteri
pada media cair. Sebanyak 1.5 mL biakan bakteri dimasukkan dalam tabung mikro kemudian disentrifugasi pada kecepatan 10 000 g selama 5 menit pada suhu ruang. Pelet yang terbentuk disuspensikan dengan 1800 µL lysis buffer A yang mengandung 20 mg ml-1 lysozim. Suspensi diinkubasi pada suhu 37 oC selama 30 menit menggunakan shaker water bath.
Sebanyak 200 µL lysis solution dan 20 µL proteinase K ditambahkan, kemudian dihomogenkan dengan menggunakan shaker water bath pada suhu 56 o
C selama 30 menit. Sebanyak 20 µL RNAse A solution ditambahkan kemudian dihomogenkan dengan menggunakan vorteks. Inkubasi dilakukan selama 10 menit, kemudian ditambahkan 400 µL etanol 50% dan dihomogenkan dengan menggunakan vortex. Suspensi dimasukan dalam GeneJET Genomic DNA purification column baru. Sebanyak 500 µL buffer wash I ditambahkan, kemudian disentrifugasi selama 3 menit pada kecepatan 12 000 g. GeneJET Genomic DNA purification column dalam tabung mikro ukuran 1.5 mL. Sebanyak 200 µL buffer ulusi ditambahkan kemudian diinkubasi selama 2 menit pada suhu ruang. Tahap terakhir adalah tabung mikro disentrifugasi selama 1 menit pada kecepatan 8 000 g, kemudian disimpan pada suhu -20 oC.
Amplifikasi DNA kromosom dengan teknik PCR menggunakan sepasang primer general 16S rRNA untuk prokariot yaitu 27F forward (5’ -AGAGTTTGATCCTGGCTCAG-3’) dan 1492R reverse (5’-GGT TACCTTACGACTT-3’) yang digunakan Lane (1991) dengan target amplikon ± 1500 pb. Reaksi PCR dilakukan pada volume total 25 µ L. Bahan-bahan PCR dicampur dalam tabung mikro 1.5 mL pada kondisi dingin di atas es. Sebanyak 1 µL DNA template dimasukkan ke dalam tabung PCR. Bahan-bahan PCR yang telah dicampurkan dimasukkan ke dalam tabung PCR yang berisi DNA template. Selanjutnya tabung ditempatkan pada mesin PCR. Program PCR yang digunakan yaitu denaturasi awal pada suhu 92 oC selama 1 menit, denaturasi pada suhu 95 oC selama 1 menit, annealing pada suhu 55 oC selama 1 menit, ekstensi pada suhu 72 o
C selama 1.5 menit, ekstensi akhir pada suhu 72 oC selama 10 menit, dan dilakukan sebanyak 35 siklus.Visualisasi hasil PCR diseparasi pada 1.5% agarosa dengan voltase 75 volt, selama 30 menit.
Sekuen parsial nukleotida yang diperoleh dibandingkan dengan sekuen di NCBI genbank database dengan software Basic Local Alignment Search Tool (BLAST) (http://www.ncbi.nlm.nih.gov/BLAST/).
Analisis Data
4 HASIL DAN PEMBAHASAN
Sampel Tanaman Padi untuk Isolasi Bakteri Endofit
Hasil pengambilan sampel diperoleh 8 sampel tanaman padi yang mewakili tiga lokasi berbeda. Sampel diberi kode dengan angka 1 sampai 8. Sampel nomor 1 sampai 6 mewakili lahan sawah tadah hujan di Desa Widodomartani, Kecamatan Ngemplak dan Desa Harjobinangun, Kecamatan Tratas, Kabupaten Sleman, Yogyakarta. Sampel nomor 7 mewakili lahan sawah irigasi dari Desa Gempol Sari, Kecamatan Patok Beusi, Kabupaten Subang, Jawa Barat. Sedangkan sampel nomor 8 mewakili lahan rawa pasang surut dari Desa Karang Indah, Kecamatan Mandastana, Kabupaten Barito Kuala, Kalimantan Selatan.
Lahan sawah tadah hujan di Kabupaten Sleman, Yogyakarta, memiliki pola tanam dua kali tanam padi dalam setahun dan dirotasi dengan tanaman palawija seperti kacang-kacangan. Petani di daerah tersebut mengembalikan jerami ke lahan sawah mereka. Menurut Cho dan Kobata (2000) jerami merupakan sumber bahan organik yang potensial dalam pengembangan usaha tani. Jerami padi merupakan sumber kalium (K) yang mudah tersedia di lahan sawah. Setiap 5 ton jerami mengandung K setara dengan 50 kg KCl. Sekitar 80% K yang diserap tanaman padi berada dalam jerami. Dengan demikian, mengembalikan jerami ke tanah sawah dapat memenuhi sebagian hara K yang dibutuhkan tanaman. Selain itu jerami memiliki fungsi biologi sebagai sumber energi dan makanan bagi mikroba dan mesofauna tanah. Bahan organik yang cukup tersedia dapat menunjang aktivitas organisme tanah dalam memperbaiki ketersediaan hara, siklus hara, dan pembentukan pori mikro dan makro tanah.
Lahan sawah irigasi di Kabupaten Subang, Jawa Barat memiliki pola tanam tiga kali tanam padi tanpa dirotasi dengan palawija. Ketersediaan air sepanjang tahun menyebabkan intensitas penanaman padi menjadi lebih sering. Pola tanam ini memiliki kelemahan yaitu selalu tersedianya makanan serta tempat hidup bagi hama dan penyakit tanaman, sedangkan lahan rawa di Kabupaten Barito Kuala, Kalimantan Selatan memiliki pola tanam satu kali tanam padi dalam setahun. Padi yang ditanam di daerah ini memiliki umur panjang dan tahan terhadap genangan. Pengambilan sampel dari tiga tipe lokasi berbeda bertujuan memperkaya keragaman bakteri endofit yang diisolasi, sehingga memperbesar kemungkinan untuk mendapatkan bakteri endofit yang berpotensi sebagai penginduksi ketahanan tanaman padi.
Bakteri Endofit Hasil Isolasi
Hasil isolasi diperoleh 594 isolat bakteri endofit. Isolasi dilakukan dari bagian akar, batang, dan daun tanaman padi. Isolat bakteri endofit dari bagian akar sebanyak 225 isolat, dari bagian batang sebanyak 236 isolat, dan dari bagian daun sebanyak 133 isolat (Tabel 1). Isolat bakteri endofit yang berasal dari bagian akar diberi kode EA (endofit akar), dari bagian batang diberi kode EB (endofit batang), dan dari bagian daun diberi kode ED (endofit daun).
Tabel 1 Distribusi isolat bakteri endofit hasil isolasi
Nomor Bagian sampel Asal sampel
sampel Akar Batang Daun
1 21 36 14 Sleman, Yogyakarta
2 32 52 11 Sleman, Yogyakarta
3 24 28 5 Sleman, Yogyakarta
4 32 15 18 Sleman, Yogyakarta
5 36 35 13 Sleman, Yogyakarta
6 31 24 32 Sleman, Yogyakarta
7 15 22 25 Subang, Jawa Barat
8 34 24 15 Barito Kuala, Kalsel
Jumlah 225 236 133
Total 594 isolat
Menurut Khan dan Doty (2009) tanaman memiliki relung yang beragam bagi keberadaan bakteri endofit. Beberapa penelitian telah dilakukan untuk mengetahui posisi bakteri endofit pada bagian tanaman. Menurut Benhizia et al. (2004) bakteri endofit ditemukan pada bagian tanaman seperti akar, batang, daun, biji, umbi, dan ovul. Berdasarkan Gambar 2 komposisi isolat bakteri endofit asal padi lebih banyak diisolasi dari bagian akar dan batang, serta ada kecenderungan isolat asal batang lebih banyak dibandingkan asal akar dan daun.
Gambar 2 Komposisi isolat bakteri endofit berdasarkan asal bagian tanaman padi.
Menurut Rosenblueth dan Martinez-Romero (2004) akar umumnya merupakan bagian yang paling banyak dikolonisasi bakteri endofit dibandingkan dengan bagian tanaman lain yang berada diatas permukaan tanah. Reinhold-Hurek dan Hurek (1998) juga menjelaskan bahwa selain akar, batang juga merupakan bagian yang banyak dikolonisasi bakteri endofit terutama pada bagian pembuluh xilem. Hal ini diduga karena bakteri endofit mengolonisasi jaringan inang melalui
Akar 38% Batang
40% Daun
celah atau luka yang terbentuk saat munculnya akar lateral atau zona pemanjangan akar serta diferensiasi akar dan selanjutnya menyebar ke bagian tanaman yang lain seperti yang telah dijelaskan Rosenblueth dan Martinez-Romero (2006).
Isolat X. oryzae pv. oryzae
Isolat bakteri X. oryzae pv oryzae diperoleh dari Balai Besar Penelitian Padi Sukamandi, Subang, Jawa Barat. Isolat yang digunakan dalam penelitian ini adalah isolat X. oryzae pv oryzae patotipe IV. Patotipe ini merupakan salah satu patotipe yang dominan di Indonesia dan memiliki virulensi tinggi dibandingkan dengan patotipe lainnya (Rahim et al. 2012). Isolat X. oryzae pv oryzae patotipe IV memiliki ciri koloni berbentuk bundar, tepian licin, elevasi cembung, dan berwarna kuning terang (Gambar 3A).
Gambar 3 Koloni isolat X. oryzae pv oryzae patotipe IV (A), hasil uji reaksi hipersensitif isolat X. oryzae pv oryzae patotipe IV (B), hasil uji patogenisitas isolat X. oryzae pv oryzae patotipe IV pada tanaman padi (C),
Berdasarkan uji reaksi hipersensitif (HR) pada tanaman tembakau menunjukkan bahwa isolat X. oryzae pv oryzae patotipe IV mampu menimbulkan gejala nekrotik pada daun (Gambar 3B). Reaksi hipersensitif merupakan salah satu mekanisme pertahanan yang sangat penting pada tanaman. Menurut Agrios (2005) hal ini terjadi hanya pada kombinasi yang tidak cocok antara tanaman inang dengan patogen. Sel tanaman yang terinfeksi cepat kehilangan turgor, warna daun berubah menjadi coklat, dan mengalami kematian jaringan. Perubahan fisiologi pada bagian sel yang terinfeksi dan bagian di sekelilingnya meliputi hilangnya permeabilitas membran sel, peningkatan respirasi, akumulasi dan oksidasi senyawa fenolik, serta produksi fitoaleksin.
Selain hasil uji HR, hasil uji patogenisitas pada tanaman padi juga menunjukkan bahwa isolat ini menghasilkan gejala nekrotik pada ujung daun padi dan berkembang menjadi hawar (Gambar 3C). Patogen masuk melalui luka bekas guntingan dan menyebar ke bagian jaringan tanaman yang lain. Gejala yang muncul diawali dengan warna daun menjadi keabuan, ujung daun menggulung, dan mengalami nekrosis. Hasil uji HR dan patogenisitas membuktikan bahwa
isolat X. oryzae pv oryzae patotipe IV bersifat patogen pada tanaman khususnya padi.
Seleksi Bakteri Endofit
Seleksi Bakteri Endofit Berdasarkan Viabilitas dan Morfologi Koloni
Sebanyak 370 isolat dari 594 isolat bakteri endofit menunjukkan viabilitas yang baik dan memiliki ciri morfologi yang berbeda antara satu isolat dengan isolat lainnya. Isolat tersebut dapat ditumbuhkan kembali pada media agar dengan kecepatan tumbuh berkisar antara 24 sampai 72 jam dan memiliki bentuk, warna, tepian, serta elevasi koloni yang berbeda.
Seleksi Bakteri Endofit Berdasarkan Kemampuan Induksi Ketahanan dan Pemacu Pertumbuhan Padi di Pembibitan
Beberapa bakteri endofit dilaporkan dapat meningkatkan kebugaran tanaman, memacu pertumbuhan (Kirchhof et al. 2001) dan dapat menjadi agens hayati (Shimizu et al. 2009). Menurut Rosenblueth dan Martinez-Romero (2006) peran bakteri endofit bagi inangnya dapat sebagai pemacu pertumbuhan dan meningkatkan hasil panen, sebagai agens hayati, membantu melarutkan posfat, dan berkontribusi dalam ketersediaan nitrogen bagi tanaman. Dalam penelitian ini bakteri endofit diseleksi untuk mendapatkan kandidat yang berpotensi sebagai agens hayati terhadap X. oryzae pv oryzae, khususnya yang memiliki mekanisme menginduksi ketahanan dan berpengaruh baik terhadap pertumbuhan tanaman. Berdasarkan hasil seleksi di pembibitan terhadap 370 isolat bakteri endofit, diperoleh 8 isolat bakteri endofit yang dapat menginduksi ketahanan tanaman (Gambar 4) dan 9 isolat yang dapat memacu pertumbuhan tanaman padi (Gambar 5).
Gambar 4 Pengaruh perlakuan bakteri endofit terhadap rata-rata panjang hawar daun bakteri pada tanaman padi di pembibitan. Huruf yang sama menunjukkan tidak berbeda nyata berdasarkan uji Duncan pada taraf kepercayaan 95%.
Berdasarkan hasil analisis ragam menunjukkan bahwa perlakuan bakteri endofit berpengaruh nyata terhadap induksi ketahanan tanaman padi (Lampiran
0
EA2 154 EA5 595 EB3 307 EB4 451 EB4 452 EB6 748 EB5 606 ED4 467 K sehat K sakit