ABSTRAK
SELEKSI KARAKTER KETAHANAN TERHADAP SOYBEAN MOSAIC VIRUS DAN KARAKTER AGRONOMI KEDELAI GENERASI F2 HASIL
PERSILANGAN TANGGAMUS DAN TAICHUNG
Oleh Nurrul Aslichah
Penelitian ini bertujuan untuk melakukan seleksi berdasarkan informasi tentang keragaman genetik dan heritabilitas serta nilai tengah karakter keparahan penyakit tanaman kedelai untuk ketahanan terhadap SMV dan karakter agronomi pada generasi F2 hasil persilangan Tanggamus dan Taichung. Penelitian ini
dilaksanakan di Laboratorium Lapangan Terpadu Fakultas Pertanian, Universitas Lampung dari bulan September 2013 sampai dengan Januari 2014. Pengamatan setelah panen dilanjutkan di Laboratorium Benih dan Pemuliaan Tanaman
menunjukkan bahwa karakter ketahanan dan sebagian besar karakter agronomi yang diamati mempunyai nilai keragaman fenotipe maupun genotipe yang luas dan nilai heritabilitas yang tinggi sehingga seleksi efektif dilakukan dengan memanfaatkan informasi tentang keragaman genetik dan heritabilitas. Terdapat delapan tanaman yang memiliki nilai keparahan penyakit rendah sebesar 25% dikategorikan tahan dan bobot biji pertanaman yang lebih tinggi dari kedua tetua serta rata-rata F2 keseluruhan.
SELEKSI KARAKTER KETAHANAN TERHADAP SOYBEAN MOSAIC VIRUS DAN KARAKTER AGRONOMI KEDELAI GENERASI F2 HASIL
PERSILANGAN TANGGAMUS DAN TAICHUNG
Oleh
NURRUL ASLICHAH
Skripsi
Sebagai Salah Satu Syarat untuk Mencapai Gelar SARJANA PERTANIAN
Pada
Jurusan Agroteknologi
Fakultas Pertanian Universitas Lampung
FAKULTAS PERTANIAN UNIVERSITAS LAMPUNG
RIWAYAT HIDUP
Penulis dilahirkan di desa Bandar Agung kecamatan Terusan Nunyai kabupaten Lampung Tengah, pada tanggal 8 Juni 1992, sebagai anak pertama dari pasangan Bapak Solichin dan Ibu Sumiyati. Penulis menyelesaikan pendidikan Sekolah Dasar (SD) Negeri 2 Gunung Agung pada tahun 2004, Sekolah Menengah Pertama (SMP) Negeri 3 Terusan Nunyai pada tahun 2007, dan Sekolah Menengah Atas (SMA) Negeri 1 Terbanggi Besar pada tahun 2010.
Penulis terdaftar sebagai mahasiswa Jurusan Agroteknologi Fakultas Pertanian Universitas Lampung pada tahun 2010, melalui jalur SNMPTN (Seleksi Nasional Masuk Perguruan Tinggi Negeri). Bulan Juli sampai Agustur 2013, penulis melaksanakan Praktik Umum di Taman Bogo, Kecamatan Purbolinggo, Kabupaten Lampung Timur. Bulan Januari sampai Maret 2014 penulis
MOTO
Mengeluh hanya menyebabkan hidup dan pikiran jadi lebih berat. Nikmati ritme masalah yang dihadapi.
Tidak ada masalah, tidak hidup.
Tidak ada masalah, tidak belajar (Mario Teguh).
PERSEMBAHAN
“Kepada ALLAH SWT dan Nabi Muhammad SAW, hanya karena rahmat-Mu
hamba dapat menyelesaikan skripsi ini, dan hamba persembahkan skripsi ini untuk Bapak dan Ibu tercinta yang selalu memberikan motivasi & do’a dalam
SANWACANA
Alhamdulillahhirobbil’alaamiin segala puji bagi ALLAH SWT, Rabb yang telah
melimpahkan rahmat dan kasih sayang-Nya sehingga penulis dapat
menyelesaikan skripsi ini. Skripsi dengan judul “Seleksi Karakter Ketahanan Terhadap Soybean Mosaic Virus Dan Karakter Agronomi Kedelai Generasi F2
Hasil Persilangan Tanggamus Dan Taichung” adalah salah satu syarat untuk memperoleh gelar Sarjana Pertanian di Universitas Lampung.
Penulis berharap, skripsi yang merupakan wujud dari kerja keras, dan do’a serta didukung dengan bantuan dan keterlibatan berbagai pihak ini akan bermanfaat dikemudian hari. Oleh karena itu, penulis ingin mengucapkan terima kasih kepada:
1. Dr. Ir. Nyimas Sa’diyah, M.P., selaku pembimbing utama yang telah memberikan perhatian, pemikiran, dan bimbingan yang sangat membangun selama penulis melakukan perkuliahan, penelitian, dan penyelesaian skripsi.. 2. Prof. Dr. Ir. Hasriadi Mat Akin, M.S., selaku pembimbing kedua yang telah
memberikan ilmu pengetahuan, saran, kritik, semangat, dan kesabaran yang tak terhingga saat membimbing dalam penelitian ini..
vii 4. Dr. Ir. Kuswanta Futas Hidayat. M.P., selaku Ketua Jurusan Agroteknologi
Fakultas Pertanian Universitas Lampung.
5. Prof. Dr. Ir. Wan Abbas Zakaria, M.S., sekalu Dekan Fakultas Pertanian Universitas Lampung.
6. Orang tua tercinta Bapak Solichin dan Ibu Sumyati, Papah Saji, dan Mamah Siti Sukaesih serta sahabat tercinta Yuni Susilawati yang selalu memberikan motivasi dan memberikan masukan kepada penulis .
7. Teman-teman satu penelitian Nidya Wanda, Riza Aprianti, S.P., Tety Maryenti, S.P., Noviaz Adriani, Aulia Meydina, Christian Raymon, Jefri Julkarnain, Dimas Nugroho, Miandri Sabli P., S.P., Ria Putri, S.P., Risa Jamil, S.P., Yurida Sari, S.P, Lilis Wantini, S.P., Puput, dan Tisa Wulandari, S.P. yang terlibat dalam penelitian dan memberikan masukan dalam
pembuatan skripsi ini.
8. Sahabat tercinta Widiana E.W., Senja A., Tiya O., Rendina D.P., Sri Mulyani dan Tri Purnama S., Wika M.F, dan Tika Mutiasari, dan Dewi Kartika C., yang telah membantu dalam penelitian, memberi motivasi dan memberi masukan kepada penulis dalam menyelesaikan skripsi ini.
Penulis menyadari bahwa skripsi masih belum sempurna karena kesempurnaan hanya milik ALLAH dan semoga ini dapat bermanfaat bagi semua. Aamiin
Bandar Lampung, Agustus 2014 Penulis
DAFTAR ISI
Halaman
DAFTAR TABEL ... xi
DAFTAR GAMBAR ... xii
I. PENDAHULUAN ... 1
1.1 Latar Belakang Masalah ... 1
1.2 Rumusan Masalah ... 6
1.3 Tujuan Penelitian ... 7
1.4 Kerangka Pemikiran ... 7
1.5 Hipotesis ... 11
II. TINJAUAN PUSTAKA ... 12
2.1 Tanaman Kedelai ... 12
2.1.1 Sejarah Singkat Tanaman Kedelai ... 12
2.1.2 Morfologi Tanaman Kedelai ... 13
2.1.3 Syarat Tumbuh Tanaman Kedelai ... 14
2.2 Penyakit Mosaik Tanaman Kedelai ... 15
2.2.1 Penyebab Penyakit ... 15
2.2.2 Gejala Penyakit Mosaik Tanaman Kedelai ... 15
ix
2.3 Ketahanan Terhadap Penyakit ... 17
2.3.1 Ketahanan Horizontal ... 17
2.3.2 Ketahanan Vertikal ... 18
2.4 Keragaman Genetik dan Heritabilitas ... 20
2.4.1 Keragaman Genetik ... 20
2.4.2 Heritabilitas ... 22
III. BAHAN DAN METODE ... 24
3.1 Tempat dan Waktu Penelitian ... 24
3.2 Alat dan Bahan ... 24
3.3 Metode Penelitian ... 25
3.4 Analisis Data ... 25
3.5 Pelaksanaan Penelitian ... 27
3.5.1 Pembuatan Larutan Buffer Fosfat ... 27
3.5.2 Perbanyakan Inokulum soybean mosaik virus ... 27
3.5.3Persiapan Lahan ... 28
3.5.4 Penanaman ... 28
3.5.5 Pemupukan ... 29
3.5.6 Inokulasi soybean mosaic virus di Lapangan ... 30
3.5.7 Pelabelan ... 30
3.5.8 Perawatan dan Pemeliharaan Tanaman ... 31
3.5.9 Pemanenan ... 31
3.5.10 Pengamatan ... 31
IV. HASIL DAN PEMBAHASAN ... 35
x
4.2 Pembahasan... 38
V. KEIMPULAN DAN SARAN ... 45
VI.Kesimpulan ... 45
6.1 Saran ... 46
PUSTAKA ACUAN ... 47
DAFTAR TABEL
Tabel Halaman
1. Ragam dan kriteria keragaman genotipe dan fenotipe populasi
F2 hasil persilangan Tanggamus dan Taichung. ... 36
2. Heritabilitas arti luas populasi F2 hasil persilangan Tanggamus
dan Taichung. ... 37
3. Peringkat genotipe hasil persilangan F2 Tanggamus dan
Taichung berdasarkan keparahan penyakit (%) dan bobot biji (g). . ... 38
4. Data asli jumlah populasi F2 persilangan Tanggamus x Taichung yang
Hidup. ... 51
5. Data asli jumlah tetua Tanggamus yang hidup. ... 56 6. Data asli jumlah tetua Taichung yang hidup. ... 57 7. Ragam fenotipe populasi F2 hasil persilangan Tanggamus dan
Taichung. ... 58
8. Ragam genotipe populasi F2 persilangan Tanggamus dan
Taichung. ... 59
9. Peringkat genotipe hasil persilangan F2 Tanggamus dan Taichung
berdasarkan keparahan penyakit (%), bobot biji (g), bobot biji sehat,
DAFTAR GAMBAR
Gambar Halaman
1. Tata letak penanaman benih kedelai hasil persilangan Tanggamus x
Taichung dan kedua tetuanya. ... 29
2. Tahap-tahap inokulasi soybean mosaic virus di lapangan. ... 30
3. Skor gejala penyakit. ... 32
1.PENDAHULUAN
1.1 Latar Belakang Masalah
Kedelai merupakan tanaman asli daratan Cina dan telah dibudidayakan sejak 2500 SM. Sejalan dengan makin berkembangnya perdagangan antarnegara yang terjadi pada awal abad ke-19, menyebabkan tanaman kedelai juga ikut tersebar ke
berbagai negara tujuan perdagangan tersebut, yaitu Jepang, Korea, Indonesia, India, Australia, dan Amerika. Kedelai mulai dikenal di Indonesia sejak abad ke-16 dan penyebarannya yaitu di Pulau Jawa, kemudian berkembang ke Bali, Nusa Tenggara, dan pulau-pulau lainnya sebagai tanaman makanan dan pupuk hijau (Irwan, 2006).
2
Rendahnya produksi kedelai di Indonesia disebabkan oleh beberapa faktor di antaranya yaitu perbedaan kondisi lingkungan. Pada negara asalnya kedelai merupakan tanaman subtropis sedangkan di Indonesia kedelai merupakan tanaman tropis. Oleh karena itu, adanya perbedaan lingkungan yang
menyebabkan produksi kedelai di Indonesia tidak setinggi di negara asalnya. Perbedaan kondisi lingkungan tersebut antara lain yaitu adanya perbedaan intensitas cahaya matahari. Intensitas cahaya matahari mempengaruhi pertumbuhan tinggi tanaman kedelai, karena intensitas cahaya matahari mempengaruhi proses fotosintesis.
Menurut Martin dkk. (2006), kedelai termasuk tanaman berhari pendek yang sangat sensitif terhadap fotoperiode. Kedelai memerlukan lama penyinaran 13,5 jam sehari untuk berbunga. Benih kedelai yang diproduksi di bawah kondisi suhu tinggi cenderung menghasilkan kualitas yang rendah. Menurut Gardner dkk. (1991), kultivar kedelai yang beradaptasi pada daerah subtropis mempunyai ruas-ruas yang lebih sempit, lebih pendek, serta berbunga lebih awal apabila
ditumbuhkan pada daerah tropis. Pada daerah subtropis pertumbuhan vegetatif tanaman kedelai lebih lama. Pada daerah tropis periode pertumbuhan vegetatif tanaman kedelai lebih singkat.
3
lapangan. Karena itu, patogen dan serangga vektor menjadi selalu tersedia dan terakumulasi tingkat populasinya dari tahun ke-tahun (Akin, 2003).
Salah satu penyakit yang menyerang pertanaman kedelai di Indonesia yang disebabkan oleh virus yaitu penyakit mosaik tanaman kedelai. Penyakit mosaik tanaman kedelai disebabkan oleh soybean mosaic virus (SMV). SMV merupakan salah satu jenis virus penting pada tanaman kedelai. Penyakit ini tersebar di beberapa sentra produksi kedelai di Indonesia dan menyebabkan penurunan hasil. Menurut Kameya (2001) serta Ooffei dan Albrechtsen (2005) yang dikutip
Prayogo (2012), penurunan hasil akibat SMV dapat mencapai 25% apabila
penularan terjadi pada fase vegetatif, namun kehilangan hasil dapat mencapai 90% apabila tanaman terinfeksi sejak fase awal pertumbuhan.
Infeksi virus yang terjadi dalam sel akan mempengaruhi sintesis protein dan asam nukleat tanaman. Infeksi virus juga akan mempengaruhi jumlah dan bentuk sel serta organel, seperti mitokondria dan kloroplas. Gangguan fisiologis tanaman mengakibatkan tanaman inang menunjukkan gejala di seluruh bagian tanaman seperti tanaman menjadi kerdil, perubahan warna daun, ukuran, dan bentuk buah yang dihasilkan (Akin, 2006).
4
tangkai daun menjadi berwarna coklat, tunas-tunas penuh bercak, daun cepat rontok, dan akhirnya tanaman mati (Semangun, 1993).
Salah satu cara yang dapat dilakukan dalam pengendalian penyakit mosaik tanaman kedelai adalah penggunaan kultivar unggul yang tahan terhadap SMV dengan produktivitas tinggi. Menurut Putri (2013), varietas Tanggamus, Yellow bean, zuriat Tanggamus dan Yellow bean, dan zuriat Tanggamus dan Taicung pada populasi F1 tahan terhadap SMV. Orba merupakan varietas yang rentan
terhadap SMV.
Benih yang digunakan pada penelitian ini merupakan benih generasi F2 hasil
persilangan antara Tanggamus dan Taichung. Hasil persilangan antara
Tanggamus dan Taichung populasi F1 ini merupakan hasil persilangan Maimun
Barmawi menggunakan metode dialel setengah dengan lima tetua yaitu Tanggamus, Yellow bean, Taichung, B3570, dan Orba dan menghasilkan 10 kombinasi persilangan. Kemudian penelitian tersebut diteruskan oleh Ria Putri dan Risa Jamil pada populasi F1untuk menguji ketahanan tanaman kedelai
terhadap SMV pada generasi F1. Benih Tanggamus dan Taichung generasi F2 dari
generasi F1 genotipe No 8 tersebut dipilih dengan pertimbangan mempunyai
jumlah biji sehat sebanyak 423 butir, jumlah biji sakit sebanyak 29 butir, dan persentase keparahan penyakit (KP) sebesar 25 % dan tergolong tahan. Perlu dilakukan penelitian untuk melihat ketahanan tanaman kedelai terhadap infeksi SMV pada generaasi F2. Dari hasil penelitian Putri (2013) menunjukkan bahwa
5
Menurut Aprianti (2014), ketahanan suatu tanaman terhadap penyakit tidak
berkorelasi positif dengan hasil tanaman. Tanaman yang rentan terhadap penyakit khususnya yang disebabkan oleh virus tidak akan menurunkan hasil produksi.
Seleksi merupakan salah satu proses pemuliaan tanaman dan merupakan dasar dari seluruh perbaikan tanaman untuk mendapatkan kultivar unggul baru. Keberhasilan seleksi tergantung pada kemampuan pemulia untuk memisahkan genotipe yang tidak diinginkan dari genotipe yang diinginkan. Heritabilitas dan keragaman genetik merupakan salah satu parameter genetik yang dapat digunakan sebagai pertimbangan agar seleksi efektif dan efisien. Keragaman dan
heritabilitas dapat diamati pada karakter agronomi tanaman. Karakter agronomi merupakan karakter-karakter yang berperan dalam penentuan hasil suatu tanaman.
Menurut Barmawi dkk. (2013), seleksi yang efektif terjadi jika populasi memiliki keragaman genotipe dan fenotipe yang luas. Keragaman genetik yang luas, baik keragaman fenotipe maupun genetik menunjukkan adanya peluang yang besar untuk menyeleksi sifat-sifat yang diinginkan. Keragaman yang sempit dalam suatu populasi menunjukkan bahwa individu dalam populasi tersebut cenderung homogen dan proses seleksi untuk karakter tersebut tidak akan berjalan efektif (Racmadi, 2000). Populasi F2 merupakan populasi yang mengalami segregasi
secara bebas yang mengakibatkan 50% dari populasi merupakan genotipe
heterozigot. Oleh karena itu, populasi F2 merupakan populasi yang memiliki
keragaman tertinggi. Menurut Belanger dkk.(2003), besarnya keragaman pada
populasi F2 disebabkan semakin banyak gen yang mengendalikan sehingga
6
Heritabilitas merupakan salah satu faktor yang menentukan dalam keberhasilan seleksi karena dengan nilai heritabilitas dapat diketahui bahwa suatu sifat lebih dipengaruhi oleh faktor genetik atau faktor lingkungan. Menurut Sa’diyah dkk. (2013), heritabilitas digunakan untuk mengetahui seberapa besar pengaruh faktor genetik pada suatu karakter. Rendahnya nilai duga heritabilitas karena pengaruh faktor lingkungan lebih besar daripada faktor genetik sehingga seleksi menjadi kurang efektif. Seleksi terhadap karakter yang heritabilitasnya tinggi dapat dilakukan pada generasi awal (F2 dan F3).
Berdasarkan latar belakang maka dibuat perumusan masalah sebagai berikut: 1. Bagaimana efektivitas seleksi berdasarkan keragaman genetik dan
heritabilitas karakter ketahanan dan agronomi kedelai terhadap infeksi SMV generasi F2 hasil persilangan Tanggamus dan Taichung?
2. Bagaimana keragaman genetik karakter ketahanan dan agronomi kedelai terhadap infeksi SMV generasi F2 hasil persilangan Tanggamus dan
Taichung?
3. Berapa besaran nilai duga heritabilitas karakter ketahanan dan agronomi kedelai terhadap infeksi SMV generasi F2 hasil persilangan Tanggamus dan
Taichung?
4. Berapa besaran nilai tengah keparahan penyakit karakter ketahanan kedelai terhadap infeksi SMV generasi F2 hasil persilangan Tanggamus dan
7
1.2 Tujuan Penelitian
Berdasarkan identifikasi dan perumusan masalah yang telah dikemukakan, maka disusun tujuan penelitian sebagai berikut:
1. Mengetahui kefektifan seleksi dengan memanfaatkan informasi berdasarkan keragaman genetik dan heritabilitas karakter ketahanan dan agronomi kedelai terhadap infeksi SMV pada generasi F2 hasil persilangan Tanggamus dan
Taichung.
2. Mengetahui keragaman genetik karakter ketahanan dan agronomi kedelai terhadap infeksi SMV pada generasi F2 hasil persilangan Tanggamus dan
Taichung.
3. Mengetahui nilai duga heritabilitas karakter ketahanan dan agronomi kedelai terhadap infeksi SMV pada generasi F2 hasil persilangan Tanggamus dan
Taichung.
4. Mengetahui nilai tengah keparahan penyakit karakter ketahanan kedelai terhadap infeksi SMV pada generasi F2 hasil persilangan Tanggamus dan
Taichung.
1.3 Kerangka Pemikiran
Untuk menjelaskan perumusan masalah dalam penelitian ini, maka disusun kerangka pemikiran sebagai berikut:
8
daerah subtropis mempunyai periode vegetatif tanaman yang lebih lama
dibandingkan kedelai di negara tropis lebih pendek. Periode vegetatif yang lama menyebabkan hasil fotosintesis tanaman banyak digunakan untuk pertumbuhan batang yang akan mempengaruhi jumlah buku pada tanaman. Jika jumlah buku pada tanaman banyak maka polong yang dihasilkan juga banyak, karena polong pada kedelai terdapat pada buku dan juga akan meningkatkan hasil produksi.
Perbedaan kondisi lingkungan tersebut juga menybabkan timbulnya berbagai penyakit. Umumnya kondisi lingkungan di negara tropis mendukung bagi perkembangan penyakit. Di negara tropis tidak mempunyai musim winter yang dapat menekan perkembangan penyakit. Salah satu penyakit yang menyerang tanaman kedelai adalah penyakit mosaik tanaman kedelai yang disebabkan oleh soybean mosaic virus (SMV). Penyakit mosaik tanaman kedelai merupakan
penyakit penting yang menjadi kendala dalam budidaya tanaman kedelai.
Virus merupakan mikroorganisme yang bersifat sistemik yang artinya apabila virus masuk ke dalam tanaman maka virus ini akan menyebar ke seluruh tubuh tanaman melalui pembuluh xylem maupun pembuluh floem. Oleh karena itu, tanaman yang telah terinfeksi virus akan mengalami gangguan proses
metabolisme yang menyebabkan penurunan hasil. Hasil metabolisme tanaman lebih banyak digunakan untuk sintesis virus sehingga tanaman akan mengalami kahat metabolit. Pada tanaman yang terserang virus pertumbuhan dan
9
agronomi tanaman tersebut. Tanaman yang terserang SMV ini memiliki ciri-ciri yaitu pertumbuhan tanaman terhambat atau tanaman menjadi kerdil, adanya gejala mosaik, dan permukaan daun menjadi keriting.
Penyakit yang menyerang tanaman dipengaruhi oleh tiga faktor yaitu inang, virus dan lingkungan. Virus dapat berkembang dengan cepat apabila strain virus yang menyerang bersifat virulen, tanaman yang diserang rentan, dan kondisi
lingkungan mendukung untuk perkembangan penyakit. Oleh karena itu, salah satu cara yang dapat dilakukan untuk mengurangi penurunan hasil akibat serangan penyakit ini adalah dengan menggunakan varietas yang tahan. Tanaman
dikategorikan tahan apabila tanaman tersebut hanya mengalami sedikit infeksi atau infeksi yang terbatas. Ketahanan terhadap infeksi virus dapat ditingkatkan apabila tanaman tersebut mempunyai gen pengendali ketahanan.
Generasi F2 yang digunakan dalam penelitian ini merupakan hasil persilangan
antara Tanggamus dan Taichung. Varietas Tanggamus mempunyai daya hasil yang cukup tinggi, dan rentan terhadap virus SMV. Taichung memilki daya hasil dan kualitas rendah, namun tahan terhadap virus SMV. Generasi F2 mempunyai
keragaman genetik yang luas pada seluruh karakter agronomi yang diamati. Oleh karena itu generasi ini mempunyai nilai keparahan penyakit yang sangat beragam. Generasi F2 merupakan generasi yang bersegregasi paling tinggi karena memiliki
heterogenitas genetik tertinggi. Benih F2 memiliki gabungan karakter dari kedua
tetuanya.
10
penampilan populasi tanaman. Ragam genetik yang besar dalam suatu populasi menunjukkan bahwa semakin besar peluang untuk memperoleh genotipe yang diinginkan yaitu mendapatkan kultivar unggul yang tahan terhadap SMV dan berdaya hasil tinggi. Keragaman dalam suatu tanaman ditentukan oleh faktor lingkungan dan faktor genetik. Faktor lingkungan yang mempengaruhi seperti iklim, kesuburan tanah, cahaya matahari, dan lain sebagainya. Faktor genetik terjadi dari pewarisan kedua tetuanya. Keragaman genetik dapat terlihat jika terdapat variasi genotipe ditanam pada lingkungan yang sama.
Estimasi heritabilitas perlu dilakukan untuk mendapatkan informasi genotipe populasi F2 hasil persilangan Tanggamus dan Taichung yang menghasilkan nilai
11
1.4 Hipotesis
Berdasarkan kerangka pemikiran yang telah dikemukakan, dapat disusun hipotesis sebagai berikut:
1. Seleksi efektif dilakukan dengan memanfaatkan informasi tentang keragaman genetik dan heritabilitas karakter ketahanan dan agronomi tanaman kedelai generasi F2 hasil persilangan Tanggamus dan Taichung.
2. Karakter ketahanan dan agronomi tanaman kedelai generasi F2 hasil
persilangan Tanggamus dan Taichung memiliki keragaman genetik yang luas. 3. Karakter ketahanan dan agronomi tanaman kedelai generasi F2 hasil
persilanganTanggamus dan Taichung mempunyai nilai heritabilitas yang tinggi.
4. Karakter ketahanan kedelai generasi F2 hasil persilangan Tanggamus dan
II. TINJAUAN PUSTAKA
2.1 Tanaman Kedelai
2.1.1 Sejarah Singkat Tanaman Kedelai
Kedelai (Glycine max L) merupakan salah satu tanaman pangan pengasil protein nabati. Tanaman ini berasal dari daratan Cina Pusat dan Cina Utara. Hal ini didasarkan pada penyebaran Glycine ussuriensis, spesies yang diduga sebagai tetua Glycine max. Penyebaran kedelai di kawasan Asia, seperti Jepang,
Indonesia, Filipina, Vietnam, Thailand, Malaysia, Birma, Nepal, dan India yang dimulai sejak abad pertama setelah masehi sampai abad ke-15 hingga abad ke16 (Adie dan Krisnawati (2007) yang dikutip Wardoyo (2009)). Kedudukan tanaman kedelai dalam sistematik tumbuhan (taksonomi) diklasifikasikan sebagai berikut: Kingdom : Plantae
Divisi : Spermatophyta Sub Divisi : Angiospermae Kelas : Dycotiledonae Ordo : Papilionoideae Famili : Leguminosae Genus : Glycine
13
2.1.2 Morfologi Tanaman Kedelai
Kedelai adalah tanaman tahunan setahun yang tumbuh tegak dengan ketinggian 70–150 cm, berbatang menyemak, berbulu halus, dan mempunyai sistem
perakaran yang luas. Daunnya majemuk beranak-daun-tiga dan berselang-seling. Tanaman ini menyukai tanah bertekstur ringan hingga sedang dan berdrainase baik (Rubatzky dan Yamaguchi, 1997). Tipe pertumbuhan tanaman kedelai dapat dibedakan menjadi 3 macam yakni determinit, indenterminit, dan semi determinit. Tipe semi determinit merupakan tipe antara indeterminit dan determinit dan varietas orba termasuk tipe pertumbuhan semi determinit (Suprapto, 2001).
Tanaman kedelai memiliki bunga sempurna, yakni dalam setiap bunga terdapat alat kelamin betina (putik) dan kelamin jantan (benang sari). Penyerbukan bunga kedelai terjadi saat mahkota bunga masih tertutup, sehingga kemungkinan
terjadinya perkawinan silang alami sangat kecil (Suprapto, 2001). Bunganya berwarna putih, ungu pucat, atau ungu. Polong berkembang dalam kelompok dan biasanya mengandung 2–3 biji yang berbentuk bundar atau pipih dan sangat kaya akan protein (Rubatzky dan Yamaguchi, 1997).
Kedelai mempunyai biji berkeping dua yang terbungkus oleh kulit biji.
14
mengikat nitrogen dari udara bebas yang dapat digunakan untuk petumbuhan tanaman (Suprapto, 2001).
2.1.3 Syarat Tumbuh Tanaman Kedelai
Tanaman kedelai tumbuh baik pada jenis tanah berstruktur lempung berpasir atau liat berpasir. Suhu tanah yang optimal dalam proses perkecambahan yaitu 30 °C. Bila tumbuh pada suhu tanah yang rendah (<15 °C), proses perkecambahan menjadi sangat lambat, bisa mencapai 2 minggu. Hal ini karena perkecambahan biji tertekan pada kondisi kelembaban tanah tinggi. Sementara pada suhu tinggi (>30 °C), banyak biji yang mati akibat respirasi air dari dalam biji yang terlalu cepat. Suhu lingkungan optimal untuk pembungaan tanaman kedelai yaitu 24–25 °C.
15
2.2 Penyakit Mosaik Tanaman Kedelai
2.2.1 Penyebab Penyakit
Penyakit mosaik pada tanaman kedelai disebabkan oleh SMV. Menurut Sudjono dkk. (1993) yang dikutip oleh Mulia (2008), SMV tergolong genus potyvirus yang berbentuk batang lentur dengan panjang rata–rata berukuran 750 nm dan lebar rata-rata 15–18 nm. Virion yang paling infektif berukuran panjang > 656 nm. Infektifitas SMV menurun bila terkena sinar ultraviolet atau berada dalam larutan dengan pH<4 atau pH>9. Translokasi dan replikasi virus cepat terjadi pada suhu 26 °C, tetapi pada suhu dibawah 10°C translokasi virus terhenti. Menurut Matthews (1992) yang dikutip oleh Mulia (2008), genom SMV terdiri atas RNA utas tunggal yang berukuran sekitar 10 kb dan poli-A pada ujung tiganya. Genom SMV menyandikan delapan protein yang pada awalnya merupakan satu protein besar yang kemudian mengalami pemotongan (Posttranslationally processed) menjadi protein virus.
2.2.2 Gejala Penyakit Mosaik Tanaman Kedelai
Gejala penyakit mosaik tanaman kedelai yang disebabkan oleh SMV bervariasi tergantung dari kerentanan tanaman. Gejala awal penyakit ini adalah tulang daun pada anak daun yang masih muda berubah warna menjadi kuning jernih.
16
Kemudian bintil akar yang dihasilkan pada tanaman sakit menjadi lebih sedikit dan lebih kecil (Semangun, 1993).
Terjadi gejala nekrotik pada beberpa varietas yang disertai dengan perubahan warna dari hijau menjadi coklat pada batang dan tulang daun, kemudian daun menguning, tanaman menjadi kerdil, tunas-tunas penuh dengan bercak, daun cepat rontok, dan mati. Perubahan warna belang di sekitar tulang daun disebabkan oleh berkurangnya klorofil daun akibat adanya infeksi SMV (Prayogo, 2012).
Menurut Kholidah dkk. (2013), pada tanaman kedelai yang diinokulasi SMV memiliki rata-rata tinggi tanaman lebih rendah yaitu sebesar 67,33–74,25 cm dan masa inkubasi SMV pada tanaman kedelai hitam varietas Detam 1
menunjukkan bahwa gejala muncul rata-rata 14,67–1,.67 hari setelah inokulasi.
Menurut Kameya (2001) dan Jones (2003) yang dikutip Prayogo (2012), infeksi virus dengan gejala mosaik pada tanaman menyebabkan terjadinya peningkatan respirasi, penurunan fotosintesis, keseimbangan hormon yang tidak normal, penurunan kandungan air pada tanaman, sedangkan tanaman yang sehat tidak menunjukkan gejala tersebut.
2.2.3 Pengendalian Penyakit Mosaik Tanaman Kedelai
Menurut Sudjono dkk. (1983) yang dikutip oleh Semangun (1993), pengendaliaan penyakit mosaik tanaman kedelai yang disebabkan oleh SMV dapat dilakukan dengan dengan berbagai cara antara lain:
1. Menanam benih yang bebas virus.
17
3. Menanam varietas kedelai yang tahan terhadap infeksi virus.
4. Jika perlu menggunakan insektisida untuk mengendalikan kutu daun yang menjadi vektor virus.
5. Membasmi tumbuhan inang virus mosaik kedelai.
2.3 Ketahanan Tanaman Terhadap Penyakit
Menurut Van der Plank (1963) yang dikutip Semangun (2006), ketahanan tanaman terhadap penyakit dibagi menjadi dua yaitu ketahanan horizontal dan ketahanan vertikal.
2.3.1 Ketahanan Horizontal
Menurut Oka (1993), tanaman yang memiliki ketahanan secara alamiah itu
bersifat poligenik yang dikendalikan oleh sejumlah gen atau disebut juga tanaman yang memiliki ketahanan horizontal, ketahanan lapangan, atau ketahanan umum.
Sifat ketahanan horizontal yaitu sebagai berikut:
1) Ketahananan yang dikendalikan oleh sujumlah gen; 2) Reaksinya tidak diferensial;
3) Tahan terhadap semua ras dari satu spesies patogen, terhadap spesies patogen berbeda, atau genus;
18
Menurut plank (1968) yang dikutip oleh Semangun (2006), ketahanan horizontal memberikan ketahanan yang lebih rendah tingkatannya dibandingkan dengan ketahanan vertikal. Ketahanan ini juga diwariskan secara poligenik yang diperkirakan bahwa banyak gen yang terkait dalam ketahanan ini. Oleh karena itu, ketahanan ini sulit ditangani oleh pemulia.
2.3.2 Ketahanan Vertikal
Selain memiliki ketahanan horizontal tanaman biasanya memiliki ketahanan vertikal disebut juga ketahanan spesifik. Tanaman yang memiliki ketahanan vertikal ini merupakan tanaman yang benar-benar tahan dalam menghadapi gen virulen dari patogen. Jadi interaksinya adalah gen tahan tanaman melawan gen virulen patogen.
Sifat – sifat ketahanan verikal adalah sebagai berikut: 1) Ketahannya dikendalikan oleh satu gen utama (mayor); 2) Reaksinya diferensial;
3) Tahan terhadap satu ras dari suatu spesies patogen, 4) Mengikuti nisbah Mendel;
5) Gennya dapat diidentifikasi;
Ketahanannya sedikit menurun apabila menghadapi patogen yang bersifat mutabilitas tingggi (Oka, 1993).
Menurut Semangaun (2006), timbulnya penyakit dipengaruhi oleh tiga faktor utama yaitu tanaman inang, virus dan lingkungan. Penyakit tidak akan terjadi jika patogen yang virulen bertemu dengan tanaman yang rentan tetapi kondisi
19
interaksi dengan tumbuhan inang. Patogen melakukan aksi sedangkan tumbuhan inang melakukan reaksi. Lingkungan yang mempengaruhi patogen maupun tumbuhan inang yaitu unsur hara, kelembaban, suhu, dan sinar matahari. Berikut ini adalah model segitiga penyakit menurut Semangun (2006):
Menurut Flor (1942) yang dikutip Semangun (2006), konsep ketahanan terhadap penyakit diungkapkan melalui hipotesis gene for gene. Dalam konsep tersebut dikemukakan bahwa setiap gen yang mengendalikan sifat tahan pada tanaman inang memiliki pasangan gen komplementer yang mengendalikan sifat virulensi pada patogen. Tanaman inang menunjukkan reaksi tahan jika gen yang
mengendalikan sifat tahan pada tanaman inang berpasangan dengan gen avirulen patogen. Bila patogen memilki gen virulen pasangan tersebut, maka inang akan menunjukkan reaksi rentan.
Tumbuhan Patogen
20
Menurut Flor (1956) yang dikutip Crowder (1990), konsep gene for gene dapat digambarkan sebagai berikut:
Patogen Keterangan:
V v R = Tahan
R r = rentan
Tanaman inang V = avirulen
r v = virulen
- = tidak ada pertumbuhan + = patogenitas
2.4 Keragaman Genetik dan Heritabilitas
2.4.1 Keragaman Genetik
Seleksi merupakan dasar dalam perbaikan tanaman untuk mendapatkan varietas unggul baru. Dalam perakitan varietas unggul, keragaman genetik memegang peranan yang sangat penting. Keragaman genetik adalah suatu besaran yang mengukur variasi penampilan yang disebabkan oleh komponen-komponen genetik. Penampilan suatu tanaman dengan tanaman lainnya akan berbeda dalam beberapa hal. Keragaman (variabilitas) suatu penampilan tanaman dalam populasi dapat disebabkan oleh variabilitas genetik penyusun populasi, variabilitas
lingkungan, dan variabilitas interaksi genotipe x lingkungan (Rachmadi, 2000). Untuk mengetahui keragaman tanaman perlu dilakukan pengamatan karakter tanaman. Karakter tanaman, seperti tinggi tanaman, potensi hasil, dan lain-lain yang secara umum terbagi menjadi dua, yaitu karakter kualitatif dan karakter kuantitatif.
21
Karakter kualitatif adalah karakter-karakter yang perkembangannya dikendalikan oleh aksi gen atau gen-gen yang memiliki sebuah efek yang kuat atau
dikendalikan oleh sedikit gen, seperti warna bunga, bentuk bunga, bentuk buah, bentuk daun, dan bagian tanaman lain. Karakter kuantitatif merupakan karakter yang dikendalikan oleh banyak gen-gen yang masing-masing berkontribusi terhadap penampilan atau ekspresi karakter kuantitatif tertentu, seperti tinggi tanaman, jumlah butir benih, hasil, dan lain sebagainya (Baihaki, 2000). Semakin tinggi keragaman genetik semakin tinggi peluang untuk mendapatkan sumber gen bagi karakter yang akan diperbaiki (Martono, 2009).
Ragam fenotipe yang luas pada tanaman kacang panjang terdapat pada karakter yaitu umur berbunga, umur panen polong segar, umur panen polong kering, jumlah tangkai bunga, jumlah polong tanaman, rata-rata jumlah polong tanaman, rata-rata panjang polong tanaman, rata-rata jumlah lokul tanaman, rata-rata panjang lokul, jumlah benih total, bobot benih, dan bobot 100. Keragaman genotipe yang luas terdapat pada variabel umur berbunga, umur panen polong kering, jumlah polong tanaman, rata-rata jumlah polong tanaman, rata-rata
panjang lokul, jumlah benih total, dan bobot benih tanaman (Sa’diyah dkk., 2013).
22
bahwa karakter penampilannya lebih ditentukan oleh faktor genetik (Sa’diyah dkk., 2013).
Menurut Anderson dan Bancrof (1952) yang dikutip Wahdah (1996), keragaman fenotipe dikatakan luas apabila keragaman fenotipenya lebih besar dua kali lipat standar deviasinya. Sedangkan keragaman fenotipe dikatakan sempit apabila keragaman fenotipenya lebih kecil dua kali lipat standar deviasinya.
2.4.2 Heritabilitas
Heritabilitas merupakan suatu parameter yang digunakan untuk mengukur kemampuan suatu genotipe populasi tanaman dalam mewariskan karakteristik yang dimiliki. Pendugaan nilai heritabititas suatu karakter sangat terkait dengan faktor lingkungannya. Faktor genetik tidak akan mengekspresikan karakter yang diwariskan apabila faktor lingkungan tidak mendukung. Sebaliknya, sebesar apapun manipulasi yang dilakukan terhadap faktor lingkungan tidak akan mempu mewariskan suatu karakter yang diinginkan apabila gen pengendali karakter tersebut tidak ada (Rachmadi, 2000).
23
Menurut Barmawi dkk. (2013), nilai duga heritabilitas (daya waris) tanaman kedelai tinggi terdapat pada karakter umur berbunga, umur panen, tinggi tanaman, jumlah polong per tanaman, dan bobot biji per tanaman. Nilai duga heritabilitas tanaman kedelai rendah terdapat pada karakter jumlah cabang produktif, dan bobot 100 butir menunjukkan nilai duga heritabilitas yang sedang.
Nilai duga heritabilitas arti luas merupakan perbandingan antara ragam genetik dan ragam fenotipe yang menunjukkan besarnya proporsi faktor genetik dalam fenotipe suatu karakter. Heritabilitas arti sempit memberikan indikasi derajat kemiripan antar tetua dengan keturunannya atau mengukur proporsi ragam genetik yang diwariskan pada keturunannya (Fehr, 1987).
Menurut Rachmadi (2000), nilai duga heritabilitas berkisar antara 0-1. Nilai duga heritabilitas 1 menunjukkan bahwa varians penampilan dari suatu tanaman
dilebabkan oleh faktor genetik. Sedangkan nilai duga heritabilitas 0 menunjukkan bahwa tidak satupun varians dari suatu tanaman yang muncul disebabkan oleh faktor genetik. Hal tersebut berarti bahwa tidak ada karakteristik suatu genotipe yang diwariskan kepada generasi selanjutnya. Pada kondisi seperti itu, perbaikan karakter melalui kegiatan pemulia tidak akan memberikan harapan kemajuan secara genetik.
Berikut ini adalah kriteria nilai heritabilitas menurut Mendez-Natera dkk. (2012): 1. Heritabilitas tinggi apabila H ≥ 50% atau ≥ 0,5
III. BAHAN DAN METODE
3.1 Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan di Labolatorium Lapangan Terpadu Fakultas Pertanian, Universitas Lampung dari bulan September 2013 sampai dengan Januari 2014, kemudian pengamatan dilanjutkan Laboratorium Benih Tanaman Universitas Lampung. Perbanyakan virus dilakukan di Kampung Baru, Bandar Lampung.
3.2 Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah alkohol 70%, zeolit, air, Furadan 3g, fungisida berbahan aktif mancozeb 80%, insektisida berbahan aktif delhtametrin 25 g/l aquades, buffer fosfat, Urea 50 kg/ha, SP36 100 kg/ha, KCl
100 kg/ha dan pupuk organik (kompos) 10 g/tanaman dan pupuk kandanag 10 ton/ha. Benih yang digunakan yaitu 100 tanaman dari 1 populasi F2 hasil
persilangan Tanggamus x Taichung dan 20 tetua kedelai yang terdiri atas Varietas Tanggamus dan Taichung. Tanggamus dan Taichung merupakan hasil
25
yang digunakan dalam penelitian ini yaitu mortal, alu, hand sprayer, mistar, gunting, sungkup, cangkul, sabit, koret, golok, knapsack sprayer, polybag, cotton bud, botol aqua, gelas ukur, timbangan analitik, sabit, jaring, bambu, gembor,
kantung, dan tali rafia.
3.3 Metode Penelitian
Untuk menjawab pertanyaan dalam perumusan masalah dan untuk menguji hipotesis maka rancangan perlakuan yang digunakan yaitu rancangan perlakuan tunggal terstruktur bersarang. Dalam penelitian ini seluruh tanaman yang diuji diamati.
3.4 Analisis Data
Analisis ragam fenotipe ( ), ragam lingkungan ( ), dan ragam genetik (σ2g)
berdasarkan rumus Suharsono dkk. (2006) :
σ2 f2 =
∑
σ2e =
σ σ
σ
2g = σ2
p - σ2e
keterangan:
σ2 f = varians fenotipe,
Xi = nilai pengamatan tanaman ke –i, µ = nilai tengah populasi,
N = jumlah tanaman yang diamati, σp1 = simpangan baku tetua 1,
σp2 = simpangan baku tetua 2,
n1+n2 = jumlah tanaman tetua, σ2
p = ragam fenotipe,dan σ2
e = ragam lingkungan
26
nol. Ragam lingkungan tetua sama dengan ragam lingkungan populasi keturunan jika tetua dan populasi keturunannya ditanam pada lingkungan yang sama.
Menurut Anderson dan Bancrof (1952) yang dikutip Wahdah (1996), keragaman fenotipe dikatakan luas apabila keragaman fenotipenya lebih besar dua kali lipat standar deviasinya. Keragaman fenotipe dikatakan sempit apabila keragaman fenotipenya lebih kecil dua kali lipat standar deviasinya.
Rumus penghitungan simpangan baku ( ) berdasarkan Walpole (1992) :
= ∑
Keterangan:
= simpangan baku, Xi = nilai pengamatan ke –i, µ = nilai tengah populasi,
dan N = jumlah tanaman yang diamati
Pendugaan heritabilitas dalam arti luas (H) dengan menggunakan rumus :
H=
(Suharsono dkk., 2006) Keterangan :
H= heritabilitas arti luas, = ragam genotipe, dan = ragam fenotipe
Kriteria nilai heritabilitas menurut Mendez-Natera dkk. (2012) adalah sebagai berikut:
1. Heritabilitas tinggi apabila H ≥ 50% atau H ≥ 0,5
27
Pengamatan yang dilakukan dalam penelitian ini yaitu pada tiap individu tanaman, tidak menggunakan sampel, karena benih yang digunakan masih mengalami segregasi (Baihaki, 2000). Setiap tanaman memiliki ciri dan karakteristik yang berbeda dari tanaman lainnya. Hal ini juga dapat menjadi alasan pengamatan dilakukan pada tiap individu tanaman untuk melihat keragaman dari masing-masing tanaman.
3.5 Pelaksanaan Penelitian
3.5.1 Pembuatan larutan bufer fosfat
Alat yang digunakan adalah timbangan elektrik, dua buah gelas ukur berukuran berukuran 1000 ml dan satu buah berukuran 500 ml, pengaduk, dan botol berukuran 2 liter. Bahan pembuatan larutan bufer fosfat terdiri atas KH2PO4
(larutan A: 1,36 g), Na2HPO4 . 2H2O (larutan B: 1,78 g) dan akuades sebanyak
dua liter. Pembuatan bufer fosfat dapat dilakukan dengan meninmbang 1,36 g KH2PO4 dan 1,78 gr Na2HPO4 . 2H2O. Pembuatan larutan A dilakukan dengan
menimbang 1,36 g KH2PO4 dan melarutkannya ke dalam satu liter akuades.
Pembuatan larutan B dilakukan dengan menimbang 1,78 g Na2HPO4 . 2H2O,
kemudian dilarutkan ke dalam satu liter akuades. Satu liter bufer fosfat diperoleh dengan cara mencampurkan 510 ml larutan A dan 490 ml larutan B, kemudian dimasukkan ke dalam botol yang telah disediakan dan ditutup rapat.
3.5.2 Perbanyakan Inokulum SMV
28
terinfeksi sebanyak 5g dengan menggunakan mortal dan alu yang diencerkan dengan buffer fosfat pH 7 sebanyak 5 ml. Inokulasi secara mekanik dilakukan sesuai dengan prosedur Akin (2006) setelah daun berjumlah lebih dari 4 helai atau berumur ˃ 10 hari. Caranya yaitu dengan mengoleskan sap pada permukaan daun tanaman yang yang telah ditaburi zeolit. Setelah sap dioleskan, dilakukan
pencucian menggunakan aquades dengan cara disemprot menggunakan hand sprayer. Benih kedelai yang digunakan untuk perbanyakan SMV yaitu benih
varietas Orba karena merupakan benih yang rentan terhadap virus.
3.5.3 Persiapan Lahan
Lahan diolah dengan menggunakan cangkul untuk memperbaiki sifat fisik tanah dan untuk membersihkan gulma. Kemudian tanah tersebut dicampur dengan pupuk kandang secara merata untuk meningkatkan kesuburan tanah.
3.5.4 Penanaman
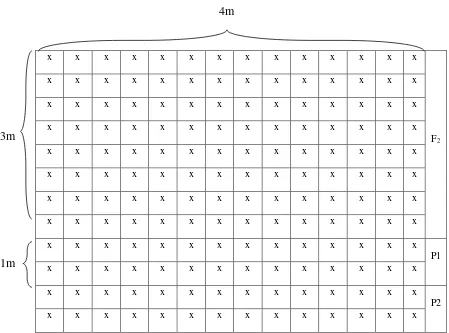
Penelitian ini dilakukan dengan menanam 100 benih F2 hasil persilangan
Tanggamus x Taichung pada petak percobaan berukuran 3m x 4m. Tanaman tersebut ditanam dengan jarak tanaman 20cm x 50cm. Jarak antar baris 50 cm dan jarak tanaman dalam baris 20 cm. Untuk benih tetua ditanam dengan ukuran petak 1m x 4m. Tata letak penanaman kedelai F2 hasil persilangan Tanggamus x
29
x x x x x x x x x x x x x x
F2
x x x x x x x x x x x x x x
x x x x x x x x x x x x x x
x x x x x x x x x x x x x x
x x x x x x x x x x x x x x
x x x x x x x x x x x x x x
x x x x x x x x x x x x x x
x x x x x x x x x x x x x x
x x x x x x x x x x x x x x P1 x x x x x x x x x x x x x x
[image:45.595.78.527.90.423.2]x x x x x x x x x x x x x x P2 x x x x x x x x x x x x x x
Gambar 1. Tata letak penanaman benih kedelai hasil persilangan Tanggamus x Taichung dan kedua tetuanya
Keterangan
P1 = Tetua Tanggamus, P2 = Tetua Taichung, dan
F2 = Persilangan Tanggamus x Taichung
3.5.5 Pemupukan
Pemupukan dilakukan dua kali yaitu pada awal tanam dan pada fase generatif. Pupuk yang diaplikasikan yaitu KCl 100 kg/ha, SP-36 100 kg/ha, dan Urea 50 kg/ha. Pupuk diaplikasikan dengan jarak 5 cm dari lubang tanam tanaman kedelai.
3m
4m
30
3.5.6 Inokulasi SMV di Lapangan
[image:46.595.112.513.196.448.2]Tanaman kedelai yang sudah memiliki daun terbuka sempurna (7 – 10 HST) dapat diinokulasi dengan sap SMV yang sebelumnya telah ditaburi zeolit.
Gambar 2. Tahap-tahap inokulasi SMV di lapangan. (1) Sterilkan mortar dan alu dengan cara menyemprotkan alkohol 70% lalu dibakar
menggunakan korek api. (2-3) Satu lembar (± 5 g) daun tanaman yang terserang SMV (tanaman sakit) dipotong kecil- kecil. (4) Tambahkan larutan buffer fosfat ± 5ml. (5) Campuran daun kedelai terinfeksi SMV dan larutan buffer fosfat dihaluskan dengan
menggunakan mortar dan alu (sap). (6-7) Taburkan zeolit diatas daun tanaman sehat yang akan di inokulasi. (8) Dioleskan sap pada permukaan daun tanaman kedelai yang sehat dengan menggunakan cutton but.
3.5.7 Pelabelan
31
3.5.8 Perawatan dan Pemeliharaan Tanaman
Perawatan dan pemeliharaan tanaman meliputi penyiraman, penyiangan gulma, penyulaman tanaman yang mati, dan pengendalian hama dan penyakit.
Penyiangan gulma dilakukan secara mekanis yaitu menggunakan koret. Penyemprotan dengan insektisida dan fungisida dilakukan sesuai dengan
kebutuhan. Insektisida yang digunakan yaitu Decis dan fungisida yang digunakan yaitu Dithane. Penyiraman dilakukan pada sore hari dengan menggunakan
gembor dan selang.
3.5.9 Pemanenan
Tanaman kedelai yang siap panen memiliki cirri-ciri yaitu polong berwarna kuning kecoklatan secara merata dan matang. Tanaman kedelai secara utuh dicabut satu persatu kemudian di masukkan ke dalam kantong panen yang telah diberi label.
3.5.10 Pengamatan
Pengamatan yang dilakukan pada penelitian ini terdiri dari pengamatan sebelum panen dan pengamatan setelah panen.
Pengamatan sebelum panen yaitu:
a. Periode inkubasi. Periode inkubasi dihitung dari waktu inokulasi sampai dengan timbulnya gejala.
32
Keparahan penyakit dihitung dengan rumus Campbell dan Madden (1990) yang dikutip Mulia (2008) :
KP =
x 100% Keterangan:
KP: Keparahan penyakit,
N : Jumlah sampel yang diamati, Z : Nilai skor tertinggi
n : Jumlah sampel untuk kategori serangan, dan V : Nilai skor untuk kategori serangan
[image:48.595.115.511.323.582.2]Menurut Akin (2006), gejala serangan setiap jenis virus yang muncul memiliki rincian sebagai berikut:
Gambar 3. Skor Gejala Penyakit .
Tidak bergejala = 0 (A), klorosis dan tulang daun memucat = 1 (B), mosaik dengan klorosis pada tulang daun dan permukaan daun = 2 (C), mosaik berat, klorosis dan terjadi pembengkokan pada permukaan daun, daun melengkung ke bawah atau ke atas = 3 (D), dan
malformasi daun = 4 (E).
A B C
33
Kategori ketahanan keparahan penyakit (%) (Akin, 2014 komunikasi pribadi) : 0 – 10 = Sangat tahan,
11 – 25 = Tahan, 26 – 35 = Agak tahan, 36– 50 = Agak rentan, 51 – 75= Rentan, dan 76-100 = Sangat rentan
Pengamatan yang dilakukan setelah panen meliputi:
a. Tinggi tanaman, diukur dari pangkal batang sampai titik tumbuh tanaman. Pengukuran dilakukan setelah panen;
b. Cabang produktif, dihitung berdasarkan jumlah cabang yang dapat menghasilkan polong;
c. Total jumlah polong, dihitung berdasarkan jumlah polong per tanaman; d. Jumlah polong bernas, dihitung berdasarkan jumlah polong bernas per
tanaman;
e. Persentase jumlah polong bernas, (jumlah polong bernas : jumlah polong) x 100%
f. Jumlah polong hampa, dihitung berdasarkan jumlah polong hampa per tanaman;
g. Total jumlah biji, dihitung berdasarkan jumlah total biji per tanaman; h. Jumlah biji sehat, dihitung berdasarkan jumlah biji sehat per tanaman;
34
k. Bobot 10 butir biji sehat, diamati setelah dikeringanginkan sekitar 3 minggu setelah panen (g);
l. Bobot biji, dengan cara menimbang biji per tanaman;
V. KESIMPULAN DAN SARAN
5.1 Kesimpulan
Adapun kesimpulan dari penelitian ini adalah :
1. Seleksi pada generasi F2 hasil persilangan Tanggamus dan Taichung efektif
dilakukan dengan memanfaatkan informasi tentang keragaman genetik dan heritabilitas.
2. Karakter keparahan penyakit dan karakter tinggi tanaman, total jumlah polong, jumlah polong bernas, persentase polong bernas, jumlah polong hampa, total jumlah biji, jumlah biji sehat, persentase biji sehat, jumlah biji sakit, bobot biji per tanaman, bobot biji sehat,persentase bobot biji sehat, bobot biji sakit, dan umur panen mempunyai nilai keragaman genotipe yang luas.
3. Karakter keparahan penyakit dan karakter jumlah cabang produktif, tinggi tanaman, total jumlah polong, jumlah polong bernas, persentase polong bernas, total jumlah biji, jumlah biji sehat, persentase biji sehat, bobot biji pertanaman, bobot biji sehat, persentase bobot biji sehat, dan umur panen mempunyai nilai duga heritabilitas yang tinggi.
4. Generasi F2 hasil persilangan Tanggamus dan Taichung mempunyai nilai
46
5.2 Saran
Perlu dilakukan penelitian lebih lanjut untuk delapan genotipe tanaman unggul yang dipilih berdasarkan keparahan penyakit yang dikategorikan tahan dan bobot biji per tanaman yang lebih tinggi daripada kedua tetuanya dan rata-rata F2
PUSTAKA ACUAN
Akin, H. M. 2006. Virologi Tumbuhan. Yogyakarta. Kanisius. 187 hlm.
Akin, H. M. 2003. Respon beberapa genotipe kedelai terhadap infeksi CPMMV (Cowppea Mild Mottle Virus). J. Hama dan Penyakit Tumbuhan Tropika. 3(2) : 40 – 44.
Aprianti, R. 2014. Keragaman dan uji korelasi karakter ketahanan kedelai (Glycine max [l.]Merrill) terhadap Soybean mosaic virus generasi F2
persilangan Tanggamus x B3570. Skripsi. Universitas Lampung. Bandar lampung.
BPS. 2013. Data produksi tanaman kedelai. Katalog. Jakarta. BPS 521. Baihaki, A. 2000. Teknik Rancangan dan Analisis Penelitian Pemuliaan.
Universitas Padjajaran. Bandung. 91 hlm.
Barmawi, M., A. Yushardi, N. Sa’diyah. 2013. Daya waris dan harapan kemajuan seleksi karakter agronomi kedelai generasi F2 hasil persilangan antara
Yellow bean dan Taichung. J. Agrotek Tropika. 1(1):20-24.
Belanger, F.C., K.A. Plumley, P.R. Day, and W.A. Meyer. 2003. Interspecific
hybridization as a potential method for improvement of Agrostis species.
43(6): 2172-2176
Crowder, L. V. 1990. Ketahanan Penyakit pada Tanaman. Dalam Genetika Tumbuhan. Diedit oleh Soetarsono. Diterjemahkan oleh Kusdiarti, L. Yogyakarta. Gadjah mada University press. 499 hlm..
Fehr, W. R. 1987. Principles of Cultivar Development. Vol.1. Macmillan Publ. Co. New York. 536 hlm.
Gardner, F. P., R. B. Pearce, dan R. L. Mitchell. 1991. Fisiologi Tanaman
Budidaya. Diterjermahkan oleh Herawati, S. Jakarta. Universitas Indonesia Press. 428 hlm.
48
Irwan, A.W. 2006. Budidaya Tanaman Kedelai (Glycine max [L.] Merril).
http://pustaka.unpad.ac.id/wp-content/uploads/2009/03/budidaya_tanaman_kedelai.pdf. Diunduh tanggal 14 Desember 2013.
Kholida, L. N., T. Hadiastono, M. Martosudiro. 2013. Pengaruh dosis pupuk organik cair terhadap infeksi Soybean mosaic virus (SMV), pertumbuhan dan produksi tanaman kedelai hitam (Glycine max (l.) Merr.) Varietas Detam-1. Jurnal HPT.1(3):50-59.
Oka, I. N. 1993. Pengantar Epidemiologi Penyakit Tanaman. Yogyakarta. Gadjah mada University press. 92 hlm.
Martin, J. H., R. P. Waldren, dan D. L. Stamp. 2006. Soybean4nd Edition. In Principles Of Field Crop Productio. United State. 613-630 page.
Martono, B. 2009. Keragaman genetik, heritabilitas dan korelasi antar karakter kuantitatif nilam (Pogostemon Sp.) hasil fusi protoplas. Jurnal
Littri.15(1):9-15.
Mendez-Natera, J. R., A. Rondon, J. Hernandez, and J. F. Morazo-Pinoto. 2012. Genetic studies in upland cotton. III. Genetic parameters, correlation and path analysis. Sabrao J. Breed. Genet. 44(1):112-128.
Mulia, Y. 2008. Uji daya gabung karakter ketahanan beberapa genotipe kedelai [Glycine max (l.) merril]. Tesis. Universitas lampung. 65 hlm.
Prayogo, Y. 2012. Keefektifan cendawan entomopatogen Lecanicillium lecanii (zare & gams) terhadap bemisia tabaci gen. Sebagai vektor Soybean mosaic virus (SMV) pada tanaman kedelai. Superman: Suara Perlindungan
Tanaman. 2 (1):11-21.
Putri, R. 2013. Estimasi nilai heritabilitas dan nisbah potensi karakter ketahanan dan agronomi terhadap infeksi Soybean mosaic virus. Skripsi. Universitas Lampung. Bandar lampung. 78 hlm.
Rachmadi, M. 2000. Pengantar Pemuliaan Tanaman Membiak Vegetatif. Universitas Padjajaran : Bandung. 159 hlm.
Rubatzky, E.R., dan M. Yamaguchi. 1997. Sayuran Dunia 2. Diterjemahkan oleh Catur, H. Insitut Teknologi Bandung: Bandung. 320 hlm.
49
Sa’diyah, N., M. Widiastuti, dan Ardian. 2013. Keragaan, keragaman, dan heritabilitas karakter agronomi kacang panjang (Vigna unguiculata) generasi F1hasil persilangan tiga genotipe. J. Agrotek Tropika. 1(1):32-37.
Semangun, H. 1993. Penyakit-Penyakit Tanaman pangan di Indonesia. Yogyakarta. Gadjah Mada University Press.848 hlm.
Semangun, H. 2006. Pengantar Ilmu Penyakit Tumbuhan. Yogyakarta. Gadjah Mada University Press.850 hlm.
Suharsono, M. Jusuf, dan A.P. Paserang. 2006. Analisis ragam, heritabilitas, dan pendugaan kemajuan seleksi populasi F2 dari persilangan kedelai kultivar
Slamet dan Nokonsawon. Jurnal Tanaman Tropika. XI (2) : 86-93.
Sulistyo, A., dan Yuliasti. 2012. Nilai duga heritabilitas galur-galur mutan kacang hijau (Vigna radiata). Seminar Nasional Pangan. Hlm13-16.
Sukur, M., S. Sujiprihati, R. Yunianti, dan K. Nida. 2010. Pendugaan komponen ragam, heritabilitas dan korelasi untuk menentukan kriteria sleksi cabai (Capsicum annuum L.) populasi F5. J. Horti. Indonesia. 1(3):74-80.
Suprapto. 2001. Bertanam Kedelai. Jakarta. Penebar Swadaya. 76 hlm.
Ujianto, L., Idris dan U. Yakop. 2006. Evaluasi ketahanan terhadap kekeringan 15 galur hasil seleksi kacang tanah varietas lokal bima. Jurnal Penelitian. Universitas Mataram. 2(3).
Wahdah, R. 1996. Variabilitas dan pewarisan laju akumulasi bahan kering pada biji kedelai. Disertasi. Program Pascasarjana Universitas Padjadjaran. Bandung.
Wardoyo, S. D.Y. 2009. Uji daya hasil lanjutan galur - galur harapan kedelai (Glycine Max (L.) Merr.) berdaya hasil tinggi. Skripsi. Institut Pertanian Bogor. 70 hlm.



![POLA SEGREGASI KARAKTER KETAHANAN TANAMAN KEDELAI (GLYCINE MAX[L.] MERRILL) TERHADAP INFEKSI SOBEAN MOSAIC VIRUS POPULASI F2 KETURUNAN TAICHUNG x TANGGAMUS](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)