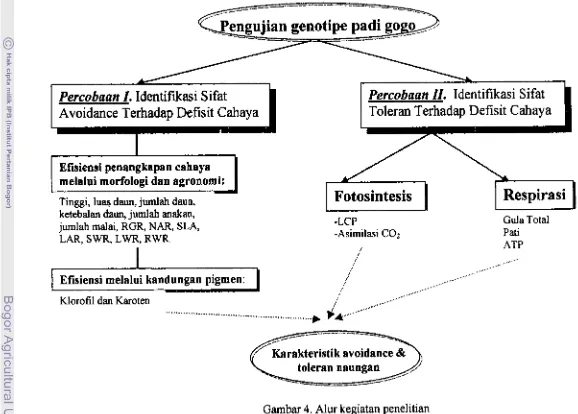
FISIOLOGI TOLEFUNS1 PAD1
GOGO
TERHADAP NAUNGAN:
TINJAUAN KARAKTERISTlK FOTOSINTESIS
DAN RESPIRASI
Oleh
BAMBANG SUPRWONO LAUTT
PROGRAM PASCA SARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
BAMBANG
SUPRIYONO LAUTT. Fisiolog Toleransi Padi Gogo Terhadap Naungan: Tinjauan Karakteristik Fotosintesis dan Respirasi. (Di bawah birnbinganDIDY SOPANDIE, MUHAMMAD ACHMAD CHOZIN, LATTFAH K.
DAICUSMAN, KUKUH SE'IAWAN).
Penanaman padi gogo sebagai tanaman sela pa& lahan perkebunan dan
unit rehabilitasi lahan sociui forestry (SF) perlu diintensifkan agar lebih
meningkatkan produktivitas lahan serta mengurangi impor berm. Salah satu ken&la yang dihadapi dalarn pengembangan tanaman sela adalah rendahnya
intensitas cahaya. Dilain pihak berbagai genotipe mernberikan respon spesifik
&lam beradaptasi terhadap intensi tas cahaya rendah. Adaptasi dilakukan melal ui
dua cara yaitu penghindaran (avoidance) dm toleran. Oleh karena itu perlu
dianalisis karakter fotosintetik
dm
respirasi tanaman yang berkaitan denganadaptasi terhadap intensitas cahaya rendah.
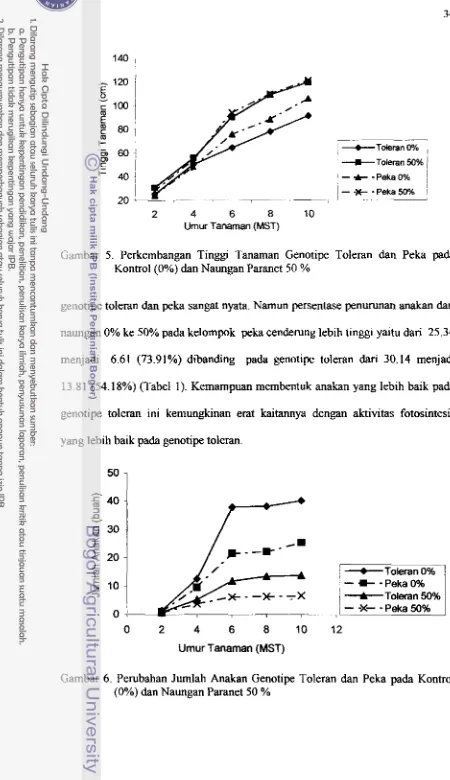
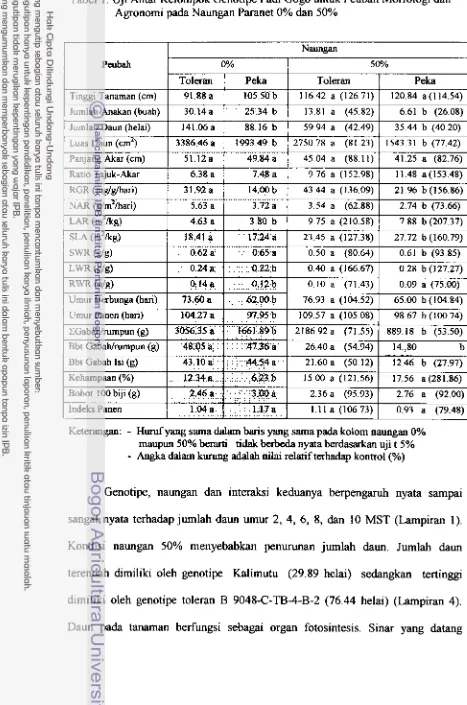
Hasil penelitian menunjukkan pada namgan 50%, tanaman toleran
memperlihatkan peningkatan luas dam yang lebih besar dibanding genotipe peka.
Selanjutnya genotipe peka memiliki kehijauan daun yang lebih ti- dibanding
genotipe toleran. Tingginya kehijauan dam pada genotipe peka terkait dengan
tingginya ketebalan dam. B e r h k a n analisis klorofil, genotipe toleran clan peka
memperlihatkan peningkatan kandmgan klorofil a, klorofil b dan total klorofil,
namun terjadi penurunan nisbah klorofil ah. Peningkatan iuas dam dm klorofil merupakan mekanisme avoidance tanaman terhadap intensitas cahaya rendah.
Kandungan total karoten
dan
P-karoten menurun pada naungan 50%.Narnun terdapat kecenderungan bahwa penurunan P-karoten p d a genotipe peka
lebih besar yaitu 1 3,17% dibanding genotipe toleran sebesar 4,3 1%.
Total
karotendiduga bukan merupakan mekanisrne avoidance terhadap intensitas cahaya
rendah.
Pada kondisi ternaungi, genotipe toleran Jatiluhur memiliki titik
kompensasi cahaya (LCP) terendah yaitu 19 sedangkan Kalimutu 25
pmo~rn2/detik. Rendahnya titik kompensasi cahaya pada kondisi naungm 50%
rnenandakan rendahnya respirasi p d a genotipe toleran.
Genotipe toleran I39048 C-TB-4-B-2 memiliki kandungan pati tertinggi sedang terendah adalah Kalimutu dengm nilai masing-masing 20,94; dan 14,77
mglg. Selanjutnya, penurunan gula total lebih tinggi pada genotipe peka. Tingginya penurunan rneqgndi kasi
kan
bahwa pada genot ipe peka penggunaan gula sebagai substrat respirasi lebih tinggi. Sebaliknya, tingginya sisa kandungangula total setelah periode 3
hari
gelap pada genotipe toleran Jatiluhurmenunjukkan rendahnya respirasi yang te jadi pada tanaman tersebut. Rendahnya LCP dan respirasi pa& genotipe toleran merupakan m e h i s m e toleransi t e h d a p
cahaya rendah.
.
Padi gogo yang toleran terhadap cahaya rendah memkrikan produksibobot gabah yang lebih baik (31,38
-
84,78% kontrol) dibanhngkan dengangenotipe peka (27,77 - 34,98% kontrol).
ABSTRACT
BAMBANG SLTPRIYONO LAU'TT. Physiology of Tolerance to Shading in Upland Rice: Study of Photosynthesis and Respiration Characteristics. (Under Supervision of D D Y SOPANDIE, MWAMMAD ACHMAD CHOZIN,
LATIFAH
K.
D A R U S W , KUKUH SETLAWAN).Upland rice planting as intercrops in plantation and "unit rehabilitation social forestry (SF)" areas must be continued to increase land productivity and to
decrease rice import. One among constraints in this prograrne is low light intensity. In the other hands, many genotypes have spesific response in adaptation
to low light intensity. Plant adaptation to low light intensity pass throught two manner i.e. avoidance and tolerance. Therefore, it is needed to analyze
photosynthetic and respiration characters in relation to low light adaptation. The result of this research showed that plant morphologcal character involved in avoidance to low light intensity are leaf area and chlorophyll content. Tolerant plant showed that leaf area is higher in 50% shadng. Furthermore, sensitive genotype have higher greeness compare to that of tolerance genotypes. Higher greeness in sensitive genotype is related to higher thickness of leaf. Based on laboratory chlorophyll analysis, tolerance and sensitive genotypes exhibited in increasing chlorophyll a, b and total chlorophyll, but decreasing ratio of chlorophyll ah.
Total carotens and pcarotens contents decreased in 50% shading. However, the reduction of j3-carotens
in
sensitive genotypes is higher i.e. 13.17% compare to that of tolerance one i.e. 4.3 1%. It is assumed that does total carotens not belong to avoidance mechanism to low light intensity.Zn shading condition, tolerance genotypes Jatiluhur have the lower light
compensation point (LCP) i.e. 19 pmonol/m2/second whereas Kalimutu was 25 pmol/rn2/second. Lower
LCP
indicated lower respiration rate.Tolerance genotype B9048C-TB-4-B-2 have highest s&rch contens while the lowest is in Kalimutu
with
value of 20.94 and 14.77 rnglg. Furthermore, reduction of total sugar is higher in sensitive genotypes. Higher reduction in sensitive genotypes indicated that sugar utilization for respiration substrat was highest. On the contrary, higher total sugar residue after 3 days in dark periods showed lowest respiration in tolerance genotypes. Lowest LCP and respiration rate in tolerant genotypes act as tolerank mechanism to low light intensity.Upland rice tolerance give a good yield in 50% shading i.e. (31.38 - 84.78 % of control) while sensitive genotypes (27.77 - 34.98% of control).
Key words.: S w i n g , t i p W rice, phoiogwt~ksi.~, repiratim, c k l o ~ o p ~ f , carolenoid, total sugar,
PERNYATAAN
Saya menyatakan dengan sebenar-benamya bahwa segala pernyataan dalam disertasi saya yang berjudul:
FISIOLOGI TOLERANSI PAD1
GOGO
TERHADAP
NAUNGAN: TINJAUAN
KARAKTERISTIK
FOTOSINTESIS DAN RESPIRASI
adalah gagasan atau hasil penelitian disertasi saya sendiri dengan bimbingan Komisi Pembimbing, kecuali yang dengan j elas ditunj ukkan ruj ukannya. Disertasi ini belum pernah diajukan
untuk
memperoleh gelar pada program sejenis di Perguruan Tinggi lain.Semua data dan infarmasi yang dgunakan telah dinyatakan dengan jelas dan dapat diperi
ksa
kebenarannyaBAMBAM
SUPIUYONO LAUTTFlSIOLOGI TOLERANST PAD1
GOGO
TERHADAP NAUNGAN:
TlNJAUAN KARAKTERISTTK FOTOSINTESIS
DAN mSPIRASI
Oleh:
BAMBANG SUPIUYONO LAUTT
9950391AGR
Disertslsi
sebagai stlab satu syarat untuk memperoleh gelar Jhktor pada
Program Studi Agronomi
PROGRAM PASCA SARJANA
INSTITUT PERTAIVIGN BOGOR
Judul disertasi : Fisiologi Toleransi Padi Gago Terhadap Naungan:
Tinjauan Karakteristik Fotosintesis dan Respirasi
Nama mahasiswa : BAMBANG SUPRIYONO
LAUTT
Nomor pokok : 995039
Program studi : AGRONOMI
Menyetuj u i 1 . Komisi Pembimbing
1
Dr. Ir. Didy Ketua Sopandie, M.Agr.I
/Prof. Dr. Ir.
A.
A. Chozin, M.Agr. Prof Dr. Tr. Latifah K. Darusman,MS.Anggota Anggota
Dr.
IT.
Kukuh Setiawan, M.Sc.Anggota
2. Program Stud1 Agronomi Ketua,
5
:
Dr. Ir. Hajrial Aswidinnoor,
M.
RIWAYAT HiDUP
Penulis dilahirkan di Palangka Raya, Kalimantan Tengah, tanggal 25 Juli 1963, merupakan
anak
ke empt dari delapan bemudara dari ayah Joseph Lautt dan ibu Nellie Rasan (Alrn). Penulis menikah dengan Hana Pertiwi, SPd padatahun 199 1 dan telah dikaruniai 2 orang anak: Jepisko Tabengan Asi b u t t dan Episcia Puspita Lautt.
Penulis menyelesaikan pendidikan Sekolah Dasar "Don Bosw" di Palangka Raya tahun 1975, Sekolah Menengah Pertama Negeri I di Palangka h y a tahun 1979
dsn
Sekolah Menengah Atas Negeri I di Palangka Raya tahun 1982. Tahun 1 9 88 menyelesaikan pendidi kan Saq m a pa& Jurusan Budidaya Pertanian, Universitas Udayana di Denpasar, Mi. Tahun 1996 mengikuti program Magister Sains pada Program Studi Agronomi, Fakultas Pascasa janaPB,
lulus Pebruari 1999. Kesempatan untuk meianjutkanke
progam Doktor pada program studi danperguruan tin= yang sama diperoleh pada bulan Agustus tahun 1999. Beasiswa pendidikan pascasarj ana diperoleh dari Departemen Pendidikan NasionaI
Republik Indonesia.
Pada tahun 1 988 diangkat sebagu pegawai negeri si pi1 pada Fakultas Non-
Gelar Teknologi Universitas Palangka Raya (selanj utnya berubah menjadi Fakultas Pertanian tahun 1993). Mngga saat ini, penulis adalah tenaga pengdjar
PRAKATA
Puji syukw pnulis panjatkan kehadirat Tuhan Yang Maha Kuasa karena atas limpahan rahmat-Nya, penulis &pat menyelesai kan di sertasi ini. Disertasi ini
bertuj uan untuk menganal i sis karakter-karakter fotosintetik dan respirasi yang
diduga berkaian erat dengan penghindaran dan toleransi terhadap naungan.
Diharapkan inforrnasi tentang karakter fotosintetik
dan
respirasi &pat dijadikansebagai masukan bagi para fisiologis untuk mengelaborasi lebih lanjut karakter
fisiolog~ tanaman yang berguna &lam pembentukan varietas unggul toleran
naungan.
Selesainya disertasi ini tidak terlepas dari bantuan dan arahan dari
berbagai pihak. Oleh karena itu, pada kesempatan ini penulis menyarnpaikan rasa terima kasih yang tulus kepada Bapak Dr. Ir. Didy Sopandie, M.Agr, sebagai
ketua komisi pembimbing yang mengarahkan berpikir secara sistematis, analisis dan komprehensif sehingga memperkaya wawasan Mam perbaikan disertasi dan
memberikan "solusi terbaik" dalam berbagai masalah selama penelitian. Penulis hanya dapat mendoakan semoga Tuhan selalu melindungi Bapk.
Ucapan terima kasih dan honnat yang sangat dalam kepada Bapak Prof.
Dr. Ir. Muhammad Achmad Chozin, M.Agr. selaku anggota komisi pembimbing
yang yang Uarn kesibukamya sebagsu pejabat di lingkungan IPB, selalu dengan rarnah clan senyurn menerima penulis, memberi wawasan tentang atmosfer
akademik serta tuntunan berfikir secara sistematis dan konsisten sehingga
Terima kasih yang tulus juga penulis sarnpaikan kepada
Ibu
Prof. Dr. Lr.Latifah K. Darusman, MS. selaku anggota komisi pembimbing yang memberi kan pengarahan dan memperkaya pengetahuan penulis terutama dalam analisis
laboratorium hingga penyajian hasil penelitian, juga memberikan darongan moril
agar dapat rnenyelesai kan stud tepat waktu.
Ucapan terima kasih yang tulus juga penulis satnpaikan
kepada
Bapak Dr. h. Kukuh Setiawan, MSc. selaku anggota komisi pembimbing yang telahmenyediakan dana penelitian melalui DCRG serta bimbingan dan masukan
berharga baik selama penelitian maupun penulisan disertasi. Juga kepada Dr. Ir.
M. Karnal, MSc. serta Ir. Syamsul Hadi, MSc. atas kesediaan berdiskusi dan
kerjasama yang baik selama ini.
U c a p terima kasih dan penghargaan yang tulus penulis sarnpaikan
kepada:
1. Relctor Universitas Palangka Raya, Rektor Institut Pertanian Bogor, Direktur
Program Pascasarjana IPB yang telah memberikan kesempatan kepada penulis unhrk mengikuti pendidikan Program Pascasarj ma di Institut Perkmian Bogor.
2. Direktur Proyek DCRG dan Staf, Pemerintah Provinsi Kalimantan Tengah, Pemko. Palangka Raya, Pemkab. Katingan, Pemkab Lamandau, HPH Gunung Meranti, Direktur RUBRBIPB, Direktur Supersemar, atas beasiswa clan dana
bantuan penelitian yang telah diberi kan.
3. Kepala Kebun Percobaan PSPT-Darmaga dm staf serta Balai Penelitian Padi
Muara, yang telah mernberikan dukungan rnoril
dan
fasilitas pelaksanaan4. Staf Laboratorium Kimia Pangan Bioteknologi-PB, Wratorium PSPT-IPB,
Laboratorium Balitbio dm Laboratorium Balitro, yang telah rnemfasilitasi dalam analisis bboratorium.
5. Rekan-rekan sesarna mahasiswa, khususnya program Studi Agronomi IPB,
atas dorongan m o i l clan bantuan yang telah diberikan selama penelitian. Ucapan terima kasih yang sangat dalam dan penuh kasih penulis tujukan
buat isteri tercinta Hana Pertiwi, SPd. yang dengam sangat sabar dm tak henti-
hentinya memotivasi serta memberikan dorongan untuk penyelesaian studi. Ucapan yang sama ditujukan buat anak-anak kami Jepisco Takngan Asi Lautt
dan Episcia Puspita Lautt yang sangat mengerti dengan kondisi orang tua yang
"berstatus mahasiswa" sib& dengan kegiatan perkuliahan dan penelitian. Ucapan
terima kasih yang tak terhingga buat lbunda tercinta (Nelly Rasan-Alm) dan
Ayahnda Joseph b u t t yang walaupun dalam kondisi tubuh =kit dan lemah, tetap
memberikan semangat dan dorongan &lam penyelesaian studi, serta buat mertua
Drs. Monon Sahadan, Kakak-kakak, Adik-adik clan seluruh keluarga, atas segala
doa
dan
kasih sayangnya.Akhirnya, kepada sernua pihak yang turut
membantu
dalam penelitianhingga penulisan disertasi ini, penulis sampaikan terima kasih, semoga chsertasi
ini bermanfaat bagi pengembangan Ilmu Pertanian khususnya bidang Fisiologi
Tanaman.
DAlFTAR TABEL
1.
Uj
i Antar Kelompok Genotipe Padi Gogo untuk Peubah Morfologidan Agronomi Pada Naungan Paranet 0% dan 50% ... 35
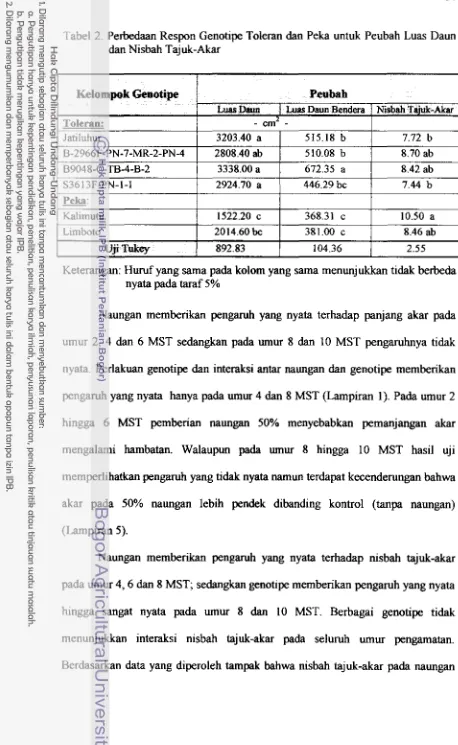
2 , Perbedaan Respon Genotipe Toleran dan Peka untuk Peubah Luas
Daun
dan
Nisbah Tajuk-Akar ... 373. Perbedaan Respon Genotipe Toleran dan Peka terhadap Laju Pertum-
buhan Relatif (RGR), Laj u Asirnilasi Bersih
WAR)
dan Nisbah LuasDaun (LAR) ...
..
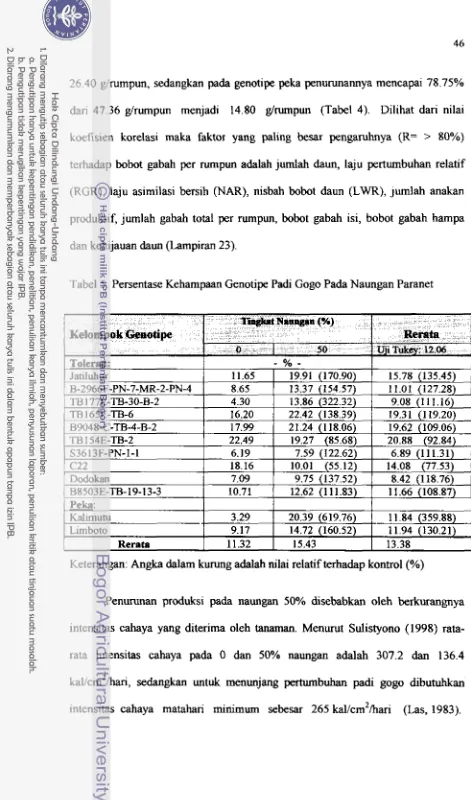
... 4 14. Bobot Gabah Total Genotipe Padi Gogo Pada Naungan Paranet ....
45
5 . Persentase Kehampaan Genotipe Padi Gogo Pada Naungan Paranet 46
6. Bobot 100 Bij i Genotipe Padi Gogo Pada Naungan Paranet
...
48 7. Uji Antar Kelompok Genotipe Padi Gogo Terhadap Peubah FisiologiTanaman pa& Naungan Paranet 0% dan 50% ... 52
8. Hasil Analisis Komponen Utama Karakter Morfologi dan Perhun-
buhan dari 1 2 Genotipe Padi Gogo pada Kondisi Naungan Paranet
DAFTAR GAMBAR
Halaman
1. Adaptasi Tanaman Naungan yang Berperanan Penting dalam
Avoidance Terhadap Defisit Cahaya ...
.
.
.
...
112. Adaptasi Tanaman Naungan yang Berperanan Penting &lam
Toleransi Defisit Cahaya ... 12
3 . Skema Pembentukan Klorofil ... 15
4. Alur Kegiatan Penelitian ... 27
5 . Perkembangan Tinggi Tanaman Genotipe Toleran dan Peka pada
Kontrol (0%) dan Naungan Paranet 50% ... 34
6. Pertambahan Luas Daun Genotipe Toleran dan Peka pada Kontrol
(0%) dm Naungan Paranet 50% ... 34
7. Laju Pertumbuhan Relatif (RGR) Tanaman Genotipe Toleran dan
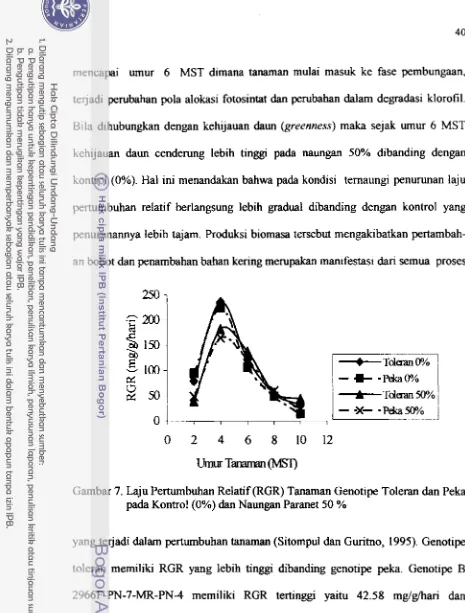
Peka pada Kondisi KontTol (0%) dan Naungan Paranet 50% ... 40
8. Pembahan Tingkat Kehijauan Daun Padi Gogo yang
Diukur
Dengan Alat Klorofil Meter SPAD ... 50
9. Regresi Linier antara Tingkat Kehijauan Daun dari Hasil Pengamatan Menggunakan Klorofil Meter SPAD dengan
Kandungan Klorofil Total Hasil Analisis Laboratorium ... 53
10. Perubahan Kandungan Klorofil Total Daun Padi Gogo pada Perlakuan GelapTerang: (0
t)
= Awal perlakuan umur 42 HST, terang; (3 D) = tiga hari pada ruang gelap; ( 6 L) = tiga hari padacahaya penuh; (9 S) = tiga hari pa& naungan 50% ... 54
1 1. Perubahan Kandungan Klorofil a dan
b
Dam Padi Gogo Perlakuan"Uj i Cepat" pada Naungan 5 0% ... 55
12. Perubahan
Kandungan
Total Karoten Daun Padi Gogo pada Perlakuan Gelap-Terang: (0 L) = Awd perlakuan umur 42 HST,terang; (3 D) = tiga hari pada ruang gelap; ( 6 L) = tiga hmi pada
cahaya penuh; (9 S) = tiga hari pada naungan 50%
...
571 3. Perubahan Kandungan 0-Karoten
Daun
Padi Gogo pada Genotipe14. Hubungan antara Asirnilasi C 0 2 dengan I'huton I g ' I u x IJemsity (PFD) Daun Padi Gogo pada Genotipe Toleran Jatiluhur dan Peka
...
Kalimutu 60
15. Hubungan antara Konduktans Stomata dengan Photon Flux Densily (PFD) Daun Padi Gogo pada Genotipe Toleran Jatiluhur dan Peka
...
Kalimutu 6 1
16. Kandungan Gula Total pada Perlakuan "Uji Cepat" Naungan 50%
...
pada Padi Gogo 65
17. Perubahan Kandungan Pati Daun Padi Gogo pada Perlakuan Gelap-Terang: (0 L) = A d perlakuan umur 42 HST, terang;
(3
D)
= tiga hari pada ruang gelap; (6L)
= tiga hari pada cahayapenuh; (9 S) = tiga hari pada naungan 50% ... 66
f 8. Perubaham Kandungan Gula Total Daun Padi Gogo pada Perlakuan Gelap-Terang: (0 L) = Awal perlakuan umur 42 HST, terang;
(3 D) = tiga hari pada m ggeiap; (6 L) = tiga harr pa& cahaya
penuh; (9 S) = tiga hari pada naungan 50% ... 68
1 9. Dendogram 5 Genotipe Padi Gogo dengm 30 Variabel Pengamatan pada Naungan 50%. (1) = Jatiluhur, (2) = B-2966 F-PN-7-MR-PN-4
DAFTAR LAMPIRAN
1 . Tabel Analisis Ragam Respon Padi Gogo pada Naungan Paranet 2. Tabel Tinggi Tanarnan Maksimum Genotipe Padi Gogo pada
Naungan Paranet . . .
. . . .
. . .. . .. . .
. . .3. Tabel Jumlah Anakan Maksimum Genotipe Padi Gogo pada Naungan Paranet . . .
. .
.. .
..
. . . .. . .. . . .
..
. . .4. Takl Jumlah Daun Genotipe Padi Gogo pada Naungan Paranet
5 . Tabel Panjang Akar Genoti pe Padi Gogo pada Naungan Paranet
6 . Tabel Jumlah Anakan Produktif Genotipe Padi Gogo pada Naungan Paranet . .
. .
. . ..
..
,.
7. Tabel Umur Berbunga Genotipe Padi Gogo pada Naungan Paranet8. Tabel
Umur
Panen Genotipe Padi Gogo pa& Naungan Paranet9. Tabel Luas Daun Spesifik (SLA) Maksimum Genotipe Padi Gogo
pada Naungan Paranet .
. . .
. . ..
..
.
.
. . .10. Tabel Nisbah Bobot Dam (LWR) Maksimum Genotipe Padi Gogo pada Naungan Paranet
.
. . .. . .
..
. . .1 1 . Tabel Nisbah Bobot Batang (SWR) Maksimum Genotipe Padi Gogo pa& Naungan Paranet ... . .
. .
. . .. . . .
, 1 2. Tabel Jumlah Gabah Total Genotipe Pad Gogo pa& Naungan Paranet . . .. . .
. . .. .
. . .. . .
. . .1 3. Tabel Bobot Gabah Isi Genotipe Padi Gogo pa& Naungan
Paranet . . .
. . . .
. . ..
. ..
. . .. . . .
,. . . .
. . . ..
.1 4. Tabel lndeks Panen Genoti pe Padi Gogo pa& Naungan Paranet
1 5. Tabel Kehijauan Daun (SPAD) Genotipe Padi Gogo pada Naungan Paranet .
. . .
. . ..
. . .16. Tabel Kandungan Klorofil
a
Fase Pengsian Biji Genotipe Padi Gogo pada Naungan Paranet . . . . .. . . .. . . .. . . .. . .Halaman
1 7. Tabel Kandungan KlorofiI b Fase Pengisian Biji Genotipe Padi Gogo pada Naungan Paranet . . .
.
. . . .. . .1 8. T a k l Kandungan Total Klorofil Fase Pengisian Biji Genotipe Padi Gogo pada Naungan Paranet .. . .
.
.
. . . ...19. Tabel Kandungan Total Karoten Fase Vegetatif Aktif dan Pengisian Bij i pada Naungan Paranet . . .
. . .
. . .20. Tabel Hasil Pengukuran Asimilasi CO1 pa& Padi Gogo yang Toleran dan Peka terhadap Naungan pada Berbagai PFD . .
. .. . .
.2 1 . Tabel Kandungan GulaTotal Fase Pengisian Biji Genotipe Padi Gogo pada Naungan Paranet .. . .
.
.. . . .
22. Tabel Hasil Analisis Tanah Sebelum dan Sesudah Percobaan di-
Kebun Percobaan Pusat Studi Pernuliaan Tanaman (PSPT) Cikabayan
.
.. . .
. . ..
..
. . .. . . .
. . ..
.. . .
23. Tabel Koefisien Korelasi Karakter Perturnbuhan dan Fisiologi Genotipe Padi Gogo pada Naungan 50%
. .
. . .. . . .
..
. . . 24. Prosedur Keja Penentuan Kandungan Klorofil Daun . . .. . .
,. .
.25. Prosedur Kerja Analisis Total Karoten
...
. . .. . ...
. . .26. Prosedur Kerja Penentuan Gula dm Pati Jaringan Tanaman . . .
Penanaman padi gogo sebagai tanarnan sela pada lahan perkebunan dapat memberikan beberapa manfaat yai tu: ( 1 ) meningkatkan efisiensi penggunaan lahan, (2) tersedianya produksi padi secara "in situ", (3) meningkatkan penciapatan petani, (4) res~du pupuk bisa dimanfaatkan oleh tanaman pokok, (5) mengurang biaya penanggulangan gulma dan tejaganya kebersihan lahan, ( 6 ) rneningkatkan aktivitas buruh tani, (7) mengurangt kasus kebakaran lahan. Narnun program agroforesfry ini perlu dipelajari dan diteliti sebab tanaman padi secara relatif sangat sensitif terhadap intensitas cahaya yang rendah.
Peningkatan daya adaptasi padi gogo terhadap intensitas cahaya rendah merupakan saIah satu prioritas utama program
TRRI
(Herdt, 1 998). Beberapa studi telah dilakukan untuk mendapatkan padi toleran naungan. Sahardi ef a]. (2000) mendapatkan bahwa dari 200 genotipe padi gogo yangd~
uj i, terdapat 10 genotipe toleran naungan yang potensial untuk dicobakm pada bexbagai daerah.Selanj utnya dikatakan bahwa mekanisme toleransi terhadap naungan pa& tanaman pad^ dengan cara menurunkan tingkat ketebalan dam, meningkatkan kandungan klwofil a
dan
nisbah klorofil alb serta rnengurangi pningkatan klorofil b. Lautt et al. ( 2000) menyatakan bahwa genotipe padi yang toleran naungan memperli hatkan tingginy a aktivitas enzim sukrosa fosfat sintase (SPS) dan kandungan pati batang pada kondisi naungan 50%. Selanjutnya Sopandie ermeningkatkan kandungan pati pada daun dan batang dibandingkan dengan
genotipe yang peka.
Meski pun telaah tentang fisiologi clan morfologi padi toleran naungan telah ditulis oleh beberapa peneliti namun informasi tentang toleransi terhadap naungan pada padl gogo rnasih banyak yang belurn diulas. Telaah fisiologi padi gogo toleran naungan yang telah dilakukan sebagan hanya rnengulas tentang
mekanisme penghindaran (moidunce), namun yang menyangkut mekanisme toleransi belurn banyak dilakukan. Ada 2 mekanisme toleransi terhadap defisit cahaya yaitu menurunkan titik kompensasi cahaya (Light Uompensution Point =
LCP) dan menurunkan respirasi (Levitt, 1980). Kedua ha1 tersebut sampai saat ini
masih be1 urn banyak di pelajari . Selain itu, dalam kaitannya dengan penghindaran terhadap defisit cahaya, keberadaan pigmen terutama klorofil dan karoten j uga perlu hteliti. Variabel-variabel tersebut akan membantu menjelaskan mekanisme penghmdaran clan toleransi padi gogo terhadap naungan secara komprehensi f.
Studi tentang karakteristik fotosintesis sebagan besar telah dilakukan, namun analisis tentang pertumbuhan tanaman belum dlakukan. Analisis pertumbuhan tidak hanya menglwtung atnbut pertumbukn seperti laju
Tujuan Penelitian Tujuan Umum:
Memperoleh inforrnasi tentang mekanisme adaptasi tanaman terhadap intensitas cahaya rendah khususnya yang berkaitan dengan penghindaran
(avo~dance) dan toleransi.
Tujuan Khusus:
I. Memperoleh informasi tentang karakter morfologi
dan
agronomi padi gogo yang rnenentukan sifat uvoidance terhadap defisit cahaya2. Memperoleh informasi tentang analisis perturnbuhan pdi gogo pada kondisi defisit cahaya
3. Memperoleh informasi tentang kandungan pigrnen khususnya klorofil dan karoten pada padi y ang toleran terhadap defisit cahay a
4. Menentukan tit i k kompensasi cahaya (LCP) dalarn kaitannya dengan toleransi terhadap defisit cahay a
5. Memperoleh informasi tentang kandungan karbohidrat (gula total dm pti)
dalam kaitannya dengan respirasi tanaman
Kerangka Pernibran
cahaya yang direfleksi jdipantulkan), ditmsmisi (diteruskan) serta meniadakan substansi pengabsorbsi cahaya yang tidak dapat mentransper energi cahaya ke pusat reaksi fotosintetik. Transmisi cahaya dikurangi hingga minimum dengan meningkatkan konsentrasi kforoplas dan pigmen kloroplas per unit sel mesofil.
Tanaman tingkat tingg akan menyesuaikan pigmennya terhadap perubahan i ntensitas cahaya pada kondisi naungan melalui penurunan rasi o klorofil a terhadap klorofil b. Dalam rangka meningkatkan penangkapan cahaya per unit area fotosintetik, daun tanaman yang ternaungi akan meniadakan pigmen non kloroplas yaitu antosianin, Hopkins (1995) menyatakan bahwa pigmen karotenoid juga terdapat pada daun yang hijau. Hasil penelitian Johnston dan
On wueme ( 1 998) pada tanaman taro mem perlihatkan bahwa kandungan
karotenoid per unit bobot segar daun dalam kondisi naungan lebih besar bila dibandingkan dengan tanpa naungan. Keberadaan pigmen terutama klorofil dan karoten masih perlu diteliti lebih lanjut pada tanaman padi yang ditanarn pada kondisi ternaungi. Selain i t y hasil penelitian Soverda (2000) rnemperlihatkan bahwa te rjadi peningkatan kandmgan klorofil tanaman padi pada kondisi ternaungi. Kapan tejadinya peningkatan juga perlu dipelajari. Penggunaan
klorofil meter SPAD untuk mengukur kehijauan (greenness) secara kontmyu akan dilakukan dalam penelitian ini.
Tanaman yang toleran naungan ditandai oleh rendahnya titik kompensasi cahaya (LCP) sehingga dapat mengakumulasi produk fotosintetik pada tingkat cahaya yang rendah dibanding dengan tanaman cahaya penuh (Levitt, 1980).
menggunakan alat "ivlfra red gus analyzer" (IRGA-CIRAS 1). Pendugaan nilai LCP untuk padi gogo masih belurn dilakukan.
Faktor toleransi yang paling penting &lam resistensi kekurangan cahaya
adalah penurunan laju respirasi (Levitt, 1980). Salah satu cara untuk melihat besarnya aktivitas respirasi adalah dengan mengukur jumlah karbohidrat yang digunakan selarna proses respirasi berlangsung. Oleh karena itu perlu dflakukan perlakuan "uji c e p f " untuk melihat pembentukan gula dan pati pada padi gogo tersebut. Pengenaan tanaman pa& kondsi gelap akan &pat menjawab pertanyaan tersebut. Selain itu jumlah CO2 yang dikeluarkan dalam proses respirasi juga
dapat digunakan sebagai indikator t i n m respirasi yang te rj adi pa& tanaman. Kandungan adenasin trifosfat (ATP) juga &pat menjadi salah satu indikator proses respirasi karena ATP yang tinggi dapat mengharnbat proses respimi. Dengan demikian maka penelitian ini akan banyak mengelaborasi teori Levitt.
Hipotesis
Hipotesis yang diajukan dalam penelitian ini :
2 . Genotipe padi gogo yang toleran naungan akan memiliki karakter morfologi dan agronomi yang berbeda dibanding dengan yang peka.
2 Genotipe yang toleran naungan akan memiliki karakter tumbuh yang lebih baik dibanding genotipe peka.
3. Terdapat perbedaan kandungan pigmen klorofil dan karoten antara genoti pe yang peka dan toleran terhadap naungan.
4. Genotipe yang toleran naungan akan memiliki aktivitas fotosintetik serta meng- akumulasi bahan kering y ang lebih tinggi dibanding genotipe y ang peka.
TfNJAUAN PUSTAKA
Fotosintesis pada Kondisi Cahsya Rendah
Proses fotosintesi s memegang peranan kunci &lam si klus hidup tanaman.
Menurut Noggle dan Fritz (1 979) i ntensitas radiasi matahari merupakan
gelombang elektromagnetik yang rnengandung energi foton yang memiliki efek panas, fotosintesis dan morfogenetik terhadap pertumbuhan tanaman. Radasi gelombang panj ang dapat menimbul kan efek panas yang berguna untuk evaptranspirasi, sedangkan radiasi dengan panjang gelombang 400 - 700 nrn berguna untuk fotosintesis. Pa& intensitas cahaya yang rendah, laju asimilasi karbon akan rendah.
Fotosintesis &pat dibagi ke &lam 3 bagan yang terpisah: (i) reaksi terang, dimana energi radiasi diabsorbsi dan digunakan untuk menghasilkan senyawa berenergi tinggi ATP dan NADPH; (ii) reaksi gelap, meliputi reduksi biokimia COz menjadi gula menggunakan senyawa berenergi tinggi yang dihasilkan pada reaksi terang dan (iii) suplai COz dari udara ke tempat reduksi di
kloroplas (Jones, 1992).
Secara umum fotosintesis dipenganh oleh umw daun, genotipe tanaman,
besarnya kebutuhan has11 asimilat oleh sink, dm pengaruh lingkungan seperti kandungan COz, kelembaban, suhu
dan
cahaya. Dalam kondisi tanpa stres, intensitas radiasi merupakan faktor lingkungan terpenting yang menyebabkan perbsdaan laju fotosintesis (Sindair dan Torie, 1989).Hubungan antara kecepatan fotosintesis dan konduktans stomata
rnemperlihatkan bahwa kscepatan fotosintesis pada 20" C leb~h tinggi 20 hingga 25% pada tanaman yang ditumbuhkan pada radiasi yang rendah dibandingkan pada tanaman yang ditumbuhkan pada radiasi y ang tinggi . Konduktans stomata
yang diukur pada 25 dan 20°C lebih rendah pada tanaman yang ditumbuhkan pada radiasi cahaya yang rendah (Ohashi et al. 1 998).
Tanaman yang ditumbuhkan pada kondisi radiasi rendah &pat memacu kapasitas regenerasi RuBP; sehingga menyebabkan relatif tinggnya fotosintetik pada level COz yang normal (Ohashi el al. 1998). Stimulasi kapasitas regenerasi RuBP oleh suhu dan radiasi rendah mungkin disebabkan oleh modifikasi atau perubahan keadaan enzim atau komponen yang berhubungan dengan regenerasi
RuBP dan perubahan dalarn level rnetabolit fotosintetik.
Meskipun terdapat tendensi bahwa fotosintesis neto (Pn) pada tanaman
Cd
cenderung meningkat pa& radiasi yang tin@ daripada tanaman C3; terdapat perbedaan yang ksar antar spesies tanaman cahaya penuh dengan naungan atau antara dam dari satu spesies yang tumbuh @a radiasi yang berbeda. Pada s p i e s tanaman naungan atau daun yang tumbuh pada kondisi ternaungi,Pn
munglunmencapai kejenuhan pada PAR kurang dari 100 pnol m-2 s-' atau mendekati 5%
dari cahaya penuh. Titik kompensasi cahaya juga bewariasi dari 0.5
-
2.0 pmol rn-'s-' pada s p i e s yang ternaungi seperti Al/ocasra macrorrhiza yang tumbuh-2 - 1
pada hutan Queensland hmgga lebih dari 40 prnol rn s pada tanaman cahaya
penuh (Jones, 1992).
cahaya hanya dengan meningkatkan efisiensi penangkapan cahaya (Gambar 1). Ada 2 cara yang tersedia yaitu: (i) meningkatkan total intersepsi cahaya rnelalui
Meningkatkan efisiensi penangkapan cahaya
I,
Meningkat an area Meningkatkan penangkapan
I
cahayapenangkapan cahay a per unit area fotosintetik
c
Refleksi Transmisi Avoidance "waste"
I
Meningkat nya Meningkatnya
1
kandungan Kandungan pigmen
per kloroplas
I
Meningkatkan proporsi avoidance avoidance &sohsi
1
Meningkatnya
fotosintetik area
I
Hilangnya kl~tikul& lilin
dan rambut
pada perrnu-
kaan daun
kandungan per sel epidermis
mesofil
I
Hilangnya pigmen
non-kloroplas
(ex. antosianin)
Garnbar 1. Adaptasi Tanaman Naungan yang Berperanan Penting Man Avoidance terhadap Defisit Cahaya (Levitt, 1980).
peningkatan luas daun; (ii) meningkatkan persentase cahaya yang digunakan dalam fotosintesis rnelalui p e n m a n j umlah cahaya yang direfleksi d m
ditransmisi jlevitt, 1980; Lawlor, 1987). Penghindaran defisit cahaya dilakukan
dengan cara tidak mengembangkan kutikula, lilin dan bulu-bul u rarnbut pada -
Toleran defisit cahay a
Menumnkan LCP Menurunkan kecepatan respirasi
dibawah LCP
Menghindari kerusakan sistem Menurunkan kecepatan respirasi
fotosintet ik mendekati LCP
Menghindari pnurunan Menghindari Menurunkan substansi Menurunkan sistem
aktivitas enzim kerusakan pigmen respirasi respiratory (mito-
[image:164.616.54.529.48.824.2]kondria & enzirn)
Gambar 2. Adaptasi Tanaman Naungan yang Berperanan Penting terhadap Toleransi
Defisit Cahaya (LCP= lighi compensation point) (Levitt, 1980)
Asimilasi bersih COz no1 (zero) terjad pada titik kompensasi cahaya
(LCP) yaitu cahaya pa& permukaan daun yang rnengnduksi kecepatan asimilasi
COz aktual sama dengan kecepatan evolusi C 0 2 respirasi. Tanaman naungan ditandai oleh rendahnya titik kompensasi cahaya sehingga dapat mengakumulasi produk fotosintetik pada tingkat cahaya yang rendah dibanding tanaman cahaya
penuh. Disamping itu tanaman naungan juga memperlihatkan kejenuhan cahaya
pada level cahaya rendah.
Titik kompensasi pada tanaman yang beradaptasi adalah sekitar 10 w/m2 dan pada tamman yang tidak kradaptasi adalah sekitar 18 w/m2. P e n m a n cahaya dari 90 ke 4.5 wlm2 menumnkan titik kompensasi cahaya dari 1 1.1 ke 1.9
d m 2 . Adaptasi ini membutuhkan waktu sekitar 8
hari
(Levitt, 1980).Titik kompnsasi cahaya pada daun gandum dewasa juga meningkat selama slang hari ; berkorelasi dengan kecepatan respirasi. Hubungan ekstrapolasi ini pada awalnya mengisyaratkan bahwa respirasi merupakan komponen u b n a
Respirasi pada Tanaman Naungan
Respirasl daun menyediakan ATP, senyawa pereduksi dan rangka karbon yang di butuhkan untuk reaksi bi osinteti k . Lingkungan cahaya rnempengaruhi kecepatan respirasi tanaman dan spesies yang &pat toleran terhadap naungan memiliki kecepatan respirasi yang lebih rendah daripada tanaman cahaya penuh (Levit, 1980).
S pesies tanaman ternaung memperli hatkan dam y ang memili ki kecepatan respirasi dan fotosintesis y ang lebih rendah dibandingkan tanaman cahaya penuh (Fitter
dan
Hay, 1989). Dikatakan bahwa kecepatan respirasi pnda spesies ternaung merupakan sesuatu yang knti kal agar te rjadi keseimbanganM
n
yang positif pada lingkungan yang temaung, seperti pada hutan.Kecepatan respirasi pada j aringan dewasa tanaman tingkat tinggi dapat dipengaruhi oleh: (i) kapasitas mesin respirasi (enzim dan transporter), (ii) konsentrasi substrat respirasi (misal karbohdrat) atau (iii) kecepatan dnnana ATP dan NAD(P)H dikansumsi. Dari segi ekolog i nyata bahwa kecepatan respirasinya kecil ketergantungannya pada ketersediaan h a y a. Tanaman dapat mencapai keadaan ini dengan memili ki sistem respirasi yang hregulasi oleh kebutuhan akan ATP (Noguchi et al. 1 996).
Regulasi respirasi pa& daun dewasa berbeda antara S. oleraceae (spesies matahari)
dan
A. d o r a (spesies naungan). Dam S. oleraceae mempedihatkan tingginya kecepatan efluks COzdan
pengambilan 0 2 dibandingkan A. odoracahaya penuh mungkin membutuhkan Iebih hanyak energi untuk rnemelihara
tingginya kecepatan fotosintesis sehingga memiliki kecepatan respirasi yang tinggi.
Pada daun dewasa, ATP yang dihasilkan oleh respirasi digunakan untuk translokasi metabolit, pernbuatan kembali struktur yang ada (protein dan membran) dan transpor intraseluler metabolit untuk meniadakan kebocoran melalui mernbran (Amthor, 1994). Ihantara proses konsumsi ATP, turn over protein kelihatannya sangat penting. Diduga bahwa 24 - 48% respirasi pemeli haraan digunakan untuk turn over protein pada akar Dactylis glomeratu.
Kecepatan respirasi gelap juga dipengarhi oleh radiasi; dlbawah kisaran 4 2 - 1
pg mV2 5.' pads tanaman ternaungi dibandingkan 50 - 150 pg rn- s pada daun
cahaya penuh. Perbedaan ini memberikan kontribusi terhadap fotosintesis neto
yang menguntungkan pa& cahaya rendah yang sering diperlihatkan oleh daun
ternaungi (Jones, 1992).
Respirasi di rnitokondria terhri atas daur Krebs, sistem pengangkutan elektron dan fosforilasi oksidatif. Terdapat berbagai kemungkinan titik pengendalian &lam ke tiga proses
di
atas. Salah satu enzim pengatur pa&tahap
pemma daur krebs adalah kinase, yang menggunakan ATP u t u k rnemfosforilasi gugus hidroksil dari berbagai gugus residu asam amino treonin pada bagtan
atau lambat sehingga melambath semua proses respirasi berikutnya di mitokondria (Salisbury dan Ross, 1995).
Pembentukan Pigmen
Prekursor dalam pernbentukan klorofil adalah senyawa organik yang merupakan senyawa intermediet hasil metabolisme. Pada tanaman tingkat tinggi,
Succinyl-CoA dan Glycine diyakini sebagai prekursor (Lawlor, 1987). Penemuan
baru-baru ini mengungkapkan bahwa prekursornya adalah glutamat yang
mengalami dearni nasi menghasilkan a-ketoglutarat yang kemudian direduksi
menjadi y,&dioxovalerate dan mengalami transaminasi menjadi &amino-
laevulinic. Sintesis ini memerlukan ATP dan NADPH.
Glutamate + a-ketoglutarate succinate + gl ycine
J
-
aminolaevulinic acidglutamate
-,
glutarnic/
2 -2H.O1 -phosphate semialdeh yde
prphobilinogen
Protochlorophyll
it
wprophorphyrinogenMg2+-adenosylmethionine
H 2 0
Protochlorophyllide
- 6 H Mg-protoporphyirin
- 4 monomethylester
gerany l gerany Ipyrophospate
Chlorophyll a .+b Chlorophyl b
[image:167.612.44.523.10.797.2]- 2 H
Pelepasan air dari 2 molekul asam aminolaevulinic menghasilkan porphobi linogen yang mengandung struktur cincin pyrrole. Reaksi seianj utnya adalah pelepasan NH3 dart COz yang kemudian mernbentuk protoporphyrinogen.
Penambahan M ~ ~ + dan adenosylmethonine pada protoporphyrin menghasilkan Mg-protoporphyrin monometylester. Mg pada klorofil berfungsi sebagai penyetel absorbsi spektrum. Mg-protoporphyrin monomet ylester mengalami dehdrasi dan reduksi menghasil kan protwhlorophyll ide. Penambahan proton akan menghasilkan Chlorofillide a. Cahaya yang diabsorbsi oleh protochlorophyllide akan mereduksi menjab klorofil a. Klorofil
b
mempakan "benruk spestar' dari klorofil a yang &lam reaksinya membutuhkan O2 dan NADPH2 (Gambar 3).Dengan demikian pembentukan klorofil a sangat dipenganrhi oleh keberadaan cahaya (Lawlor, 1987).
Klorofil menyusun sekitar 4% dari bobot kering kloroplas, tidak larut dalam air tetapi larut dalarn pelarut organik. Klorofil a berwarna hijau kebiruan, sedangkan klorofil b berwarna hijau kekuningan. Klorofil b be jumlah 113 dari kloro fil a (Hall
dan
Rao, 1 999)Perbedaan utama antam tanaman yang tidak temaungi dengan tanaman
ternaungi terktak pada kloroplasnya. Pada dam-daun muda,
tanaman
ternaungimerniliki turnpkan grana yang besar, sekitar 100 thylakoid per granum yang terletak tidak teratur &lam kloroplas. Terdapat proporsi lamella pembentuk grana yang lebih besar dan nisbah membran thylakoid terhadap stroma yang lebih tinggi
genotipe Pangrango dan B 613 yang ditumbuhkan pada naungan 50% memperlihatkan grana dan butir pati yang lebih berkemhg, dan lebih banyak thylakoid ddam masing-masing grana, jumlah stroma yang lebih sedikit dan ukuran kloroplas yang lebih kscil.
Hal yang agak berbeda terjadi pada dam yang mengalami tahap senesen. Klorofil yang berhubungan dengan protein dalam thylakoid banyak mengalami perubahan. Hidema el ul(1992) mendapatkan bahwa LHCII protein dan pigmen klorofil relatif lebih stab11 pada daun padi yang mengalami senesen pada kondisi
ternaung. Pada senesen pigmen klorofil protein seluruhnya ditahan, semen- rubisco, fosfon'lasi dan keseluruhan kapasitas fiksasi COz mengalami penurunan.
Sifat labil yang luar biasa dari penurunan klorofil daun juga bisa diamati pa& perubahan warm pi sang menjacb kuning pada kenaikan temperatur yang memperlihatkan hasil bahwa terdapat jejas kerusakan spesifik pada pigmen. Dari kenyataan ini
dan
hasil pengamatan yang lain menunjukkan bahwa sejumlah kecil aktivi tas kontrol protein-klorofil dapat rusak akibat sensitivi tasnya terhadap perubahan lingkungan, misalnya tingkat penyinaran. Dengan demikian bahwabiosintesis dan katabolisme dan protein thylakoid langsung atau ti&
Klorofil merupakan suatu komponen integral kompleks protein pemanen cahaya, klorofil a h (LHC) dan klorofil a-binding protein pada kompleks pusat reaksi PS I dan PS II yang ada di membran thylakoid. Jadi pengurangan klorofil menunj ukkan pemecahan (breakdown) kompieks protein atau degradasi membran thylakoid (Okada el a!, 1992). Pemecahan klorofil &lam daundaun padl dapat
ditekan secara komplit dengan adanya 1 pMcyclohexarnide. Degradasi dari
protein membran thylakoid juga sangat kuat dihambat oleh cyclohexamide pada konsentrasi tersebut. Kehilangan klorofil sedikit dirangsang S a r a lemah oleh adanya chlorampenicol. Kekuatan melawan terhadap de-i klorofil pada tanarnan terlindung dilakukan dengan meningkatkan konsentrasi klorofil
.
Nisbah klorofil a h menurun baik pada daun tua maupun pada dam muda. Menurut Anderson dm Osmond (1987), daun yang tidak temaungi mempunyai rasio klorofil
ah
yang lebih tingg (3.5) dibandingkan dengan permukaan yang temaungi dengan nisbah klorofilah
(2.5). P e n m a n nisbah pada kondisi ternaungi disebabkan oleh perubahan dalam isi LHCII. Pada tahap perkernbangan dam penuh, klorofil berikatan dengan LHCII mempunyai 57% dari total klorofil daun. Tanaman temaungi biasanya memiliki lebih banyak klorofil b dan LHCII, dengan rendahnya level pusat reaksiPS
11 dan inti klorofil (Hidema et al. 1992).Karotenoid disintesis dari acetyl-CoA melalui asam mevalonat, kemudan geranyl pyrophosphate, farnesyl pyrophosphate clan geranyl-geranyl pyrophosphate membentuk phytoene. Proses tersebut rneningkatkan panjang rantai rnenghasilkan stnrktur ikatan ganda yang disebut unit isoprene (Lawlor, 1987). Menurut Taiz dan Zeiger (1995), karotmoid atau turunannya rnerupakan senyawa hidrokarbon yang terdiri dari beberapa unit isoprene seperti y ang terli hat berikut ini:
Karotenoid adalah golongan zat yang mempunyai bentuk senyawa hidrokarbon
((240) dengan shuktur dasar tetra terpenoid (Goodwin dan Mercer, 1991). Karotenoid merupakan pigmen yang berwama kuning, jingga, merah jingga serta larut dalam rninyaknipida. Karotenoid terdapat dalam kloroplas (0.5%) bersama- sama dengan kiorofil (9.3%), terutama pada bagran atas perrnukaan dam, debt dengan dinding palisade. Salah satu fmgsi karotenoid & &un yaitu berperan dalam fotosintesis (Taiz dm Zeiger, 199 1).
Sifat karotenoid yang tidak stabil, mudah teroksidasi dan mudah membentuk isomer cis-trans, perlu Qperhatikan bila melakukm isolasi. Larutan karotenoid hams disimpan &lam gelap, pada suhu rendah, dibawah gas nitrogen (Hall dan Rao, 1999).
Meningkatnya stres cahaya tingg juga berhubungan dengan peningkatan nisbah klorofil
ah
dan meningkatnya ukuran pool siklus xanthophyll. Respon seluruh karotenoid yang lain terhadap stres cahaya sedikit nyata dibandingkanmeningkat dengan meningkatnya stres cahaya tinggi, nisbah neoxanthin terhadap
klorofi 1 tetap konstan. Hanya karotenoid lactucaxanthin dan a-karoten, relatif
menurun terhadap klorofil dengan meningkatnya stres cahaya tinggi (Demings-
Adams & Adams, 1996).
Level
P-
karoten kadang-kadang di temukan lebih tinggi pa& daun yangkena cahaya dibandingkan daun ternaungi, sementara pa& kasus yang lain
nilainya bisa sama atau lebih rendah (Deming-Adarns et al. 1995). Hasil
penelitian Johnston dan Onwueme (1998) pada tanarnan taro, tania, dan yam
memperl i hatkan bah wa tanaman yang diperlakukan pada kondisi ternaungr mem perli hatkan kandungan total karotenoid per rng bobot basah cenderung mengalami peningkatan.
Analisis Pertum buhan Tanaman
Spesies tanaman yang tumbuh di bawah kondrsi sub optimal akan mernperlihatkan variasi yang luas &lam laju perturnbuhan relatif (RGR) yaitu kecepatan peningkatan bahan kering per unit bahan kering yang ada (Chapin,
1980; Grime dan Hunt, 1975; Poorter, 1989 b). Hasil analisis pertumbuhan pada
24 spesies herbaceous memperlihatkan perbedaan dalam RGR temtarna disebabkan oleh variasi dalarn Iuas daun spesifik (SLA) dan nisbab bobot daun (LWR). Hasil survey pa& 63 publikasi yang rnembandingkan percobaan pertumbuhan pa& spesies herbaceous menyimpulkan bahwa SLA merupakan faktor yang sangat dominan menerangkan variasi dalarn RGR (Lambers dm
Poorter, 1992).
Laju asimilasi bersih WAR, laju peningkatan bobot tanaman per unit luas daun) clan laju fotosintesis per unit has daun tidak bervariasi secara sistematik dengan RGR (Pwrter ef al. 1 990). Perbedaan dalam laj u res pirasi pucuk &n akar; yang diekspresikan per unit bobot pucuk dan akar dsebabkan oleh perbedaan
dalarn kompsisi kimia (Poorter dan Bergkotte, 1992).
Tanaman dengan kecepatan pertumbuhan relatif (RGR) yang lebih tin&
kecepatan perturnbuhan relatif (RGR) dan komponen-komponennya seperti
kecepatan asimilasi bersih
(NAR),
nisbah luas daun (LAR), nisbah bobot daun (LWR) dan luas daun spesi fik (SLA) dapat rnembantu untuk menjelaskan variasidalam perturnbuhan clan perkembangan.
Tanaman Therrnop.srs montana yang diturnbuhkan pada ruang dengan cahaya tinggi memiliki daun 34% lebih tebal dibandingkan dalam ruang dengan cahaya rendah. Pada cahaya tinggi, daun mengalami penambahan 1 - 2 lapisan palisade parenkirn dibawah permukaan adaxial dan 1 lapisan sel palisade pendek dibawah epidermis abaxial. Sel palisade dari T. montana yang tumbuh pa& cahaya rendah panjangnya berkurang dan berbentuk kolom. Daun T. montma yang tumbuh pada cahaya tin& luasnya lebih kecil daripada daun pada cahaya rendah; semen- daun S. steilatu tidak mempunyai pehedaan pada kedua kondisi itu (Myers et al. 1 997).
Studi sebelumnya memperlihatkan bahwa kultivar padi mempunyai perbedaan yang besar dalam kemampuan memkntuk anakan (Wu et al. 1998). Kemampuan membentuk anakan ini mentpakan ciri penting pa& karena kemampuan ini mtinya mempunyai p e n g a d pa& pembentukan malai (Mller er a!. 1991) yang mempunyai korelasi yang tinggi terhadap hail biji (Wu et al.
1998).
Tanaman padi yang toleran terhadap naungan daunnya cenderung memanjang serta meningkatkan t i n u tanaman,
jwnlah
dan luas dam (Cabuslaykecepatan asirnilasi COz dan rendahnya alokasi biomasa untuk respirasi jaringan akan sangat menguntungkan untuk pertumbuhan (Bogaard el ui. 1996).
Varietas Teqing menghasilkan jumlah anakan lebih besar dibandingkan Gul fmont dan Rosemont; diaki batkan oleh tingginya kecepatan produksi nodus dan besamya jumlah nodus yang dapat menghasilkan anakan. Meskipun Teqing mempunyai kemarnpuan rnembentuk anakan yang lebih besar serta mempunyai
j umlah anakan yang lebi h besar, varietas ini memil iki kecepatan mortalitas anakan
yang lebih besar dibandingkan Gulfmont dan Rosemont. Tingginya hasil pada Teqing kelihatannya lebih merupkan hasil dari besarnya kemampuan membentuk anakan dan besarnya kepadatan malai. Kdtivar dan kelompok
anakan
secara signi fikan rnempengaruhi kepadatan gabah. Meskipun Teqing memildci j umlahrnalai dan kepadatan gabah yang tinggi namun rnemiliki persentase gabah berisi
yang rendah (72%); diikuti Rosemont(79%) dm Gulfmant (86%) (Wu et 01.
1998).
Cabuslay et al. ( 1995) melakukan penguj ian toleransi naungan terhadap 14 galur p d i . Persemaian diturnbuhkan &lam nunah kaca phytotron selama 2 minggu clan dinaungi, selanjutnya dibiarkan tumbuh pada kondisi 100 % ftanpa naungan), 50% dan 20% cahaya matahari sampai panen. Pemberian naungan 50% mengurangi bobot kering total tanaman rata-rata per galur sekitar 34.13%, sedangkan pada cahaya matahari 20% berkurang sekitar 68.92%.
Murty
dan
Dey (1 992 a) dalarn percobaan toleransi cahaya rendah pada kultivar hibrida menunj ukkan bahwa intensitas cahaya rendah dapat menunmkan hasil bahan kering tanaman pad yakni rata-rata 47% dan hasil persatuan luas70% pada hasil per satuan luas. Selanjutnya dalarn penelitian lain (Murty dan Dey, 1992 b) menunjukkan bahwa pda kultivar hibrida, laju fotosintesis dan indeks luas daun (LAI) umumnya lebih tinggi daripada hibrid 54752 terutama
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Peneli tian lapangan dilaksanakan di kebun percobaan
PB,
Ci kabayan- Darmaga, Bogor. Analisis Laboratorium dilaksanakan di Pusat studi Pemuliaan Tanaman (PSPT) IPB, Lab. Kimia Pangan-Bioteknologi IPB, Lab. Balai Penel itian B iotek-nologi (Bali tbio) d