PENCIRI FISIOLOGI KECEPATAN PERTUNASAN UMBI
GARUT (
Maranta arundinaceae
L.)
RENA UKHRAENAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Penciri Fisiologi Kecepatan Pertunasan Umbi Garut (Maranta arundinaceae L.) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
RENA UKHRAENAH. Penciri Fisiologi Kecepatan Pertunasan Umbi Garut (Maranta arundinacea L.). Dibimbing oleh TRIADIATI dan RITA MEGIA.
Maranta arundinacea L. (garut, Marantaceae) adalah tanaman yang berasal dari daerah Amerika tropis yang telah dibudidayakan secara luas termasuk di Indonesia. Umbi garut yang berasal dari daerah berbeda mempunyai kecepatan bertunas yang berbeda. Hal ini terjadi karena pertunasan dipengaruhi oleh beberapa faktor diantaranya faktor fisiologis. Tujuan penelitian ini adalah menganalisis penciri fisiologi yang meliputi kadar ZPT internal dan enzim α -amilase yang mempengaruhi kecepatan pertunasan umbi garut pada aksesi yang berbeda. Umbi garut aksesi Bantul dan Krajan yang digunakan dalam penelitian ini ditanam dengan rancangan acak kelompok faktorial. Faktor perlakuan yang digunakan yaitu kelembapan media tanam, penyinaran, dan aksesi. Hasil penelitian menunjukkan bahwa aksesi tidak mempengaruhi kecepatan pertunasan. Kandungan α-amilase umbi garut aksesi Krajan lebih tinggi daripada aksesi Bantul. Faktor fisiologi berupa hormon tidak mempengaruhi kecepatan pertunasan. Kata kunci: α-amilase, gelap, GA3, IAA, pertunasan, umbi Garut.
ABSTRACT
RENA UKHRAENAH. Physiological Markers of Shoot Initiation Rate on Arrowroot (Maranta arundinacea L.). Supervised by TRIADIATI and RITA MEGIA.
Maranta arundinacea L. (arrowroot, Marantaceae) is a native of tropical America and widely cultivated in many countries including Indonesia. Arrowroot tubers from different region have different shoot initiation rate. It was influenced by several factors, including physiological factors i.e. plant growth regulator (PGR) content and amylase enzyme. The aim of this study was to analyze the physiology markers i.e. PGR and α-amylase enzyme content that affect the shoot initiation rate of arrowroot tubers from different accession. Arrowroot tubers from Bantul and Krajan accessions this study were planted with a factorial randomized block design. Treatments were used in this study i.e. humidity of planting medium, light, and accessions. The results showed that the arrowroot accession did not affect the shoot initiation rate. The level of α-amylase of arrowroot tubers Krajan accession was higher than those of Bantul accession. Physiological factors such as hormone did not affect the shoot initiation rate.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains pada
Departemen Biologi
PENCIRI FISIOLOGI KECEPATAN PERTUNASAN UMBI
GARUT (
Maranta arundinaceae
L.)
RENA UKHRAENAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PRAKATA
Alhamduulillahirabbal’alamiin. Puji syukur kehadirat Allah SWT atas limpahan rahmat dan karunia-Nya, sehingga saya dapat menyelesaikan karya ilmiah yang berjudul Penciri Fisiologi Kecepatan Pertunasan Umbi Garut (Maranta arundinaceae L.)
Penulis ucapkan terima kasih kepada Ibu Dr Triadiati MSi dan Ibu Dr Rita Megia DEA selaku pembimbing atas pengarahan, teladan, kesabaran, waktu, serta nasehat yang diberikan kepada penulis selama penelitian hingga penulisan karya ilmiah ini. Ucapan terima kasih kepada penguji wakil Departemen Bapak Dr Berry Juliandi MSi. Terima kasih juga disampaikan kepada Ibu Suhaemi, Ayah Busro, Kakak penulis Ukit Nuryakin dan Uhmayudin, serta Adik Dede Rina Utariah atas segala doa dan dukungannya. Tak lupa penulis ucapkan terima kasih kepada Wiwi Uliyati SE, Shara Zen, Suci Wulandari, teman-teman Biologi 48 (khususnya Jeane Siswitasari Mulyana SSi, Rizky Dwi Satrio SSi, Miftahul Huda Fendiyanto SSi, Suci Dwi Rahmawati SSi, Yulianingsih, Muhammad Fadhil Amin, Rina Pangastuti), LBD Crew, keluarga Wisma QQ, keluarga Serum-G, dan BEM KM 2014 atas kerjasama, dukungan, dan semangatnya. Penulis juga ucapkan kepada Bapak Asep, Kakak Laksmi, dan Kakak Pinta atas bantuan yang diberikan selama penelitian.
Semoga karya ilmiah ini dapat menambah wawasan dan pengetahuan bagi kita semua.
DAFTAR ISI
DAFTAR TABEL xii
DAFTAR GAMBAR xii
DAFTAR LAMPIRAN xii
PENDAHULUAN 1
Latar Belakang 1
Tujuan 2
BAHAN DAN METODE 2
Waktu dan Tempat 2
Alat dan Bahan 2
Metode 2
HASIL 4
PEMBAHASAN 7
SIMPULAN 8
DAFTAR PUSTAKA 8
LAMPIRAN 11
DAFTAR TABEL
1 Rata-rata kandungan ZPT pada aksesi berbeda 6
DAFTAR GAMBAR
1 Rata-rata kecepatan pertunasan umbi garut (jumlah tunas) pada perlakuan penyinaran dan kelembapan media 50% 4 2 Rata-rata kecepatan pertunasan umbi garut (jumlah tunas) pada perlakuan penyinaran dan kelembapan media 90% 4 3 Rata-rata kecepatan pertunasan umbi garut (tinggi tunas) pada perlakuan yang
berbeda 5
4 Rata-rata pertambahan jumlah tunas minggu I dan minggu VIII pengamatan 5 5 Rata-rata tinggi tunas umbi garut pada minggu VIII 6 6 Rata-rata kandungan α-amilase pada perlakuan aksesi yang berbeda 6
DAFTAR LAMPIRAN
1 Hasil uji ANOVA jumlah tunas 11
1
PENDAHULUAN
Latar Belakang
Garut (Maranta arundinacea L.) termasuk anggota famili Marantaceae, berasal dari daerah Amerika tropis yang telah dibudidayakan secara luas di negara-negara seperti India, Srilanka, Australia, Indonesia, dan Filipina. Tanaman ini dapat tumbuh pada ketinggian 0-900 m dpl yang memiliki tanah lembap dan terlindung dari sinar matahari langsung (Sastrapradja et al. 1977). Daerah persebaran tanaman garut di Indonesia meliputi Sumatra, Madura, Jawa hingga Ternate. Di daerah Jawa, tanaman ini tumbuh pada tempat yang memiliki ketinggian tempat yang bervariasi, seperti di Bantul (60 m dpl), Krajan (320 m dpl), Kendalsari (680 m dpl), dan Begawat (850 m dpl). Garut berpotensi sebagai alternatif pangan fungsional dalam terapi diet bagi penderita diabetes. Marsono et al. (2005) menyatakan bahwa umbi garut mengandung banyak pati dan senyawa lainnya dengan jumlah serat yang tinggi. Selain itu, umbi garut juga dapat dijadikan sebagai obat luka (Kay 1973). Tepung garut dapat digunakan sebagai pemadat media kultur jaringan, dengan hasil pertumbuhan tanaman yang tidak berbeda nyata dengan agar/gelrite yang biasa digunakan (Gonzales dan Sosa 2006).
Perbanyakan garut dilakukan dengan umbi yang mempunyai titik tumbuh untuk tunas. Pertunasan pada umumnya dipengaruhi oleh zat pengatur tumbuh yaitu auksin, giberelin, dan sitokinin. IAA merupakan hormon Auksin yang mempunyai efek pada pertumbuhan dan perkembangan tanaman (Zhao 2010). Auksin diantaranya berperan pada pembentukan akar (Casimiro et al. 2001) dan dominansi apikal tajuk (Booker et al. 2003). Giberelin merupakan salah satu fitohormon penting yang mempunyai fungsi esensial dalam pertumbuhan dan perkembangan tanaman (Auzanneau et al. 2011; Liu et al. 2011), berperan dalam pemanjangan sel, pembelahan sel, dan pertumbuhan beberapa jenis tanaman (Mahmoody dan Noori 2014). Giberelin juga diketahui berperan dalam memacu perkecambahan (Grappin et al. 2000; Kaur et al. 2000) dan merupakan sinyal untuk ekspresi gen enzim α-amilase (Jacobsen dan Chandler 1987; Nolan dan Ho 1988). Enzim ini mengkatalisis perombakan pati menjadi glukosa yang menyediakan energi untuk pertumbuhan akar dan pertunasan (Akazawa dan Hara-Mishimura 1985; Beck dan Ziegler 1989). Sitokinin merupakan fitohormon yang berperan dalam perbanyakan sel, morfogenesis tunas dan akar, pengambilan nutrient, perkembangan vaskular, respon terhadap cahaya, dan senesensi (Taiz dan Zeiger 2010).
2
gandum yang diberi perlakuan GA3 memiliki kecepatan bertunas yang lebih tinggi dibandingkan dengan yang tidak diberi perlakuan. Selain itu, telah dilaporkan bahwa peningkatan aktifitas α-amilase meningkatkan pemanjangan tunas pada tulip (Lambrechts et al. 1994; Komiyama et al. 1997). Pengaruh faktor internal terhadap kecepatan pertunasan pada umbi garut belum pernah dilaporkan. Untuk itu, penelitian lebih lanjut perlu dilakukan untuk mengetahui faktor fisiologi yang mempengaruhi kecepatan pertunasan umbi garut.
Tujuan
Penelitian ini bertujuan menganalisis penciri fisiologi yang mempengaruhi kecepatan pertunasan umbi garut (Maranta arundinacea L.) dari aksesi yang berbeda.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini dilakukan di Laboratorium Fisiologi dan Genetika Molekuler dan Rumah kaca Departemen Biologi FMIPA, IPB Bogor dari Bulan Februari hingga Juli 2015.
Alat dan Bahan
Bahan penelitian terdiri dari umbi garut aksesi Bantul dan aksesi Krajan, Soluble starch (Merck), DNS (3,5 dinitro salicilic acid), buffer asetat, metanol, kloroform, 2N ammonium hidroksida, akuades, HCL 7N, NaOH 7N, dan etilasetat. Alat yang digunakan diantaranya: spektrofotometer UV-VIS Double Beam, spektronic 20D+, centrifuge, dan Inkubator
Metode
Kecepatan Pertunasan
Umbi garut ditanam pada tanah biasa dalam polybag di rumah kaca. Perlakuan gelap dilakukan dengan sungkup berwarna hitam. Rancangan penelitian yang dilakukan dengan menggunakan Rancangan Acak Kelompok (RAK) faktorial. Faktor pertama adalah penyinaran dan terdiri dari dua taraf yaitu sinar matahari (PI) dan gelap (PII). Faktor yang kedua adalah kelembapan media yang terdiri dari dua taraf yaitu 50% (A1) dan 90% (A2). Faktor yang ketiga yaitu aksesi terdiri dari dua taraf yaitu Bantul (B) dan Krajan (K). Setiap unit percobaan diulang sebanyak 5 kali. Parameter yang yang diukur adalah kecepatan pertunasan (tinggi dan jumlah tunas) diukur setiap minggu selama 8 minggu.
Analisis ZPT [IAA, GA3, dan Kinetin; Unyayar et al. (1996)]
3
yang diperoleh kemudian disaring dan ditambahkan akuades sebanyak 22.4 ml, kemudian dimasukkan ke dalam corong pemisah dan didiamkan selama 24 jam. Fase cair diambil, sedangkan fase kloroform dibuang. Selanjutnya pH pada fase cair diatur menjadi 2.5 dengan menambahkan HCl 7N, kemudian diekstraksi sebanyak 3 kali dengan menggunakan 15 ml etilasetat, kemudian didiamkan. Fase etilasetat dipisahkan dari fase cair. Fase etilasetat yang didapat diharapkan mengandung IAA dan GA3. Fase cair yang telah dipisahkan diatur pHnya menjadi 7 dengan menambahkan 7 N NaOH kemudian diekstraksi kembali sebanyak 3 kali dengan menggunakan 15 ml etilasetat. Ekstrak IAA, GA3, dan Kinetin yang diperoleh selanjutnya dievaporasi. Ekstrak kering yang diperoleh kemudian dilarutkan menggunakan 10 ml metanol. Kemudian diukur menggunakan spektroforometri UV-VIS pada panjang gelombang masing-masing 253 nm untuk GA3, 280 nm untuk IAA, dan 269 nm untuk kinetin. Percobaan diulang sebanyak 3 kali.
Analisis α-amilase (AOAC 1995)
Analisis α-amilase dilakukan pada umbi segar yang belum bertunas. Sebanyak 1 g umbi ditambahkan 5 mL buffer asetat 0,2 M pH 5. Umbi yang sudah hancur disimpan selama 10 menit sambil sekali-sekali dikocok kemudian dilakukan penyaringan dengan kapas. Filtrat yang dihasilkan disentrifugasi selama 20 menit dengan kecepatan 2000 rpm pada suhu 5 ºC. Supernatan (ekstrak enzim) yang dihasilkan diukur volumenya. Selanjutnya, sebanyak 1 mL filtrat enzim hasil ekstraksi ditambahkan dengan 1 mL larutan substrat (soluble starch), kemudian diinkubasi selama 3 menit pada suhu optimum 30 ºC. Reaksi enzim dilanjutkan dengan penambahan 2 mL DNS (3,5 dinitro salicilic acid) kemudian ditambahkan 20 mL akuades dan serapan diukur dengan Spektrofotometer pada panjang gelombang 550 nm. Percobaan diulang sebanyak 3 kali.
Analisis Statistik
4
HASIL
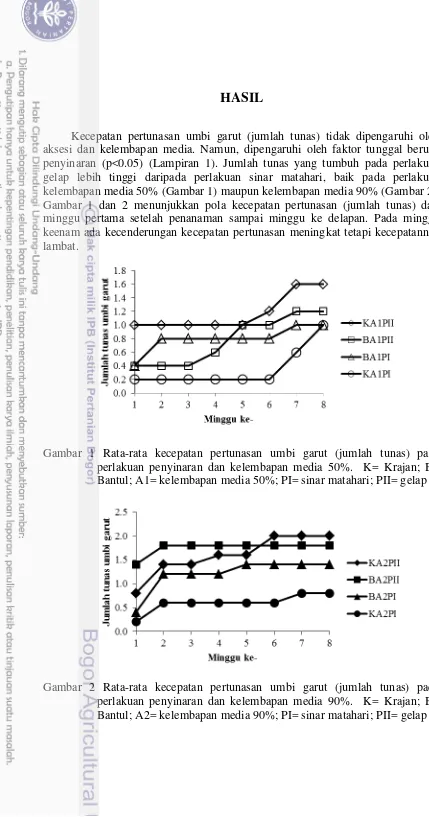
Kecepatan pertunasan umbi garut (jumlah tunas) tidak dipengaruhi oleh aksesi dan kelembapan media. Namun, dipengaruhi oleh faktor tunggal berupa penyinaran (p<0.05) (Lampiran 1). Jumlah tunas yang tumbuh pada perlakuan gelap lebih tinggi daripada perlakuan sinar matahari, baik pada perlakuan kelembapan media 50% (Gambar 1) maupun kelembapan media 90% (Gambar 2). Gambar 1 dan 2 menunjukkan pola kecepatan pertunasan (jumlah tunas) dari minggu pertama setelah penanaman sampai minggu ke delapan. Pada minggu keenam ada kecenderungan kecepatan pertunasan meningkat tetapi kecepatannya lambat.
Gambar 1 Rata-rata kecepatan pertunasan umbi garut (jumlah tunas) pada perlakuan penyinaran dan kelembapan media 50%. K= Krajan; B= Bantul; A1= kelembapan media 50%; PI= sinar matahari; PII= gelap
5
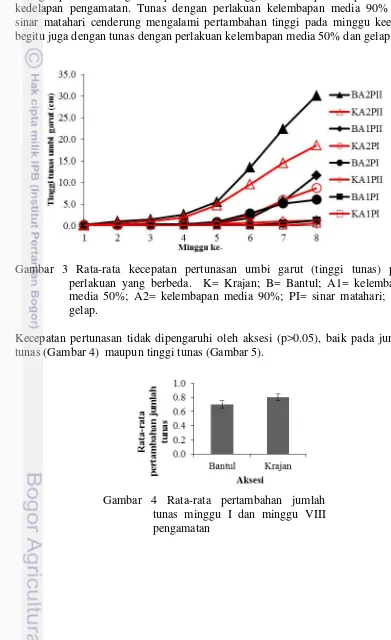
Tinggi tunas dipengaruhi oleh faktor tunggal kelembapan media atau penyinaran (p<0.05)(Lampiran 2). Gambar 3 menunjukkan pola kecepatan pertunasan (tinggi tunas) dari minggu pertama setelah penanaman sampai minggu ke delapan. Pada minggu keempat tunas yang diberi perlakuan kelembapan 90% dan gelap mulai mengalami pertambahan tinggi secara cepat sampai minggu kedelapan pengamatan. Tunas dengan perlakuan kelembapan media 90% dan sinar matahari cenderung mengalami pertambahan tinggi pada minggu keenam begitu juga dengan tunas dengan perlakuan kelembapan media 50% dan gelap.
Gambar 3 Rata-rata kecepatan pertunasan umbi garut (tinggi tunas) pada perlakuan yang berbeda. K= Krajan; B= Bantul; A1= kelembapan media 50%; A2= kelembapan media 90%; PI= sinar matahari; PII= gelap.
Kecepatan pertunasan tidak dipengaruhi oleh aksesi (p>0.05), baik pada jumlah tunas (Gambar 4) maupun tinggi tunas (Gambar 5).
6
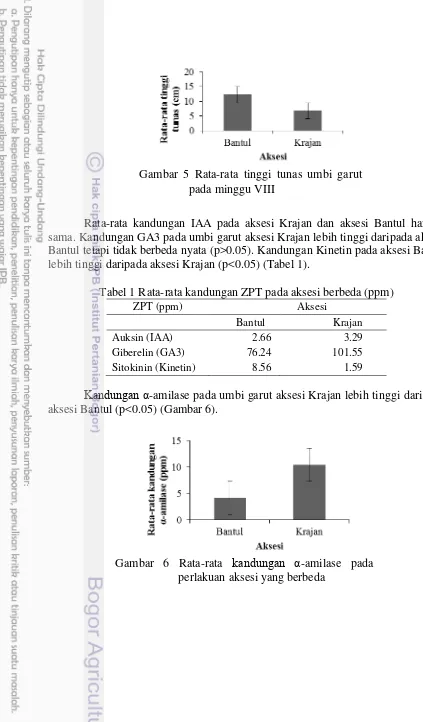
Rata-rata kandungan IAA pada aksesi Krajan dan aksesi Bantul hampir sama. Kandungan GA3 pada umbi garut aksesi Krajan lebih tinggi daripada aksesi Bantul tetapi tidak berbeda nyata (p>0.05). Kandungan Kinetin pada aksesi Bantul lebih tinggi daripada aksesi Krajan (p<0.05) (Tabel 1).
Tabel 1 Rata-rata kandungan ZPT pada aksesi berbeda (ppm)
ZPT (ppm) Aksesi
Bantul Krajan
Auksin (IAA) 2.66 3.29
Giberelin (GA3) 76.24 101.55
Sitokinin (Kinetin) 8.56 1.59
Kandungan α-amilase pada umbi garut aksesi Krajan lebih tinggi daripada aksesi Bantul (p<0.05) (Gambar 6).
Gambar 6 Rata-rata kandungan α-amilase pada perlakuan aksesi yang berbeda
7
PEMBAHASAN
Kecepatan pertunasan pada umbi garut yang diteliti tidak dipengaruhi oleh aksesi tetapi dipengaruhi oleh kelembapan media dan penyinaran secara terpisah. Umbi garut dengan perlakuan kelembapan media 90% memiliki tinggi tunas yang lebih tinggi daripada kelembapan 50%. Kelembapan media yang berbeda menunjukkan kadar air yang berbeda pada media. Air berperan penting bagi metabolisme tanaman (David dan Lawlor 2002), salah satunya yaitu mengaktifkan enzim yang berperan dalam pertunasan. Penelitian yang dilakukan oleh Chaves et al. (2002) menunjukkan bahwa kekurangan air menyebabkan pertumbuhan tunas terhambat. Kecepatan pertunasan pada umbi garut (jumlah tunas dan tinggi tunas) yang diberi perlakuan gelap lebih tinggi daripada sinar matahari. Hal ini karena kondisi gelap mempengaruhi pemanjangan sel yaitu dengan mengaktifkan beberapa enzim yang berperan dalam metabolisme (Perchorowicz et al. 1981)
Kandungan kinetin pada umbi garut aksesi Bantul lebih tinggi daripada Krajan. Kinetin yang merupakan hormon sitokinin, berperan untuk memacu pembelahan sel. Pertambahan tinggi tanaman merupakan hasil aktifitas meristem tajuk. Meristem tajuk merupakan bagian tanaman yang aktif membelah (Marta et al. 1995). Meskipun begitu kinetin tidak memberikan pengaruh nyata terhadap tinggi tunas umbi garut kedua aksesi. Kandungan IAA pada kedua aksesi hampir sama. Hal ini tercermin dari tinggi tanaman yang tidak berbeda pada kedua aksesi. Kandungan GA3 pada kedua aksesi juga tidak berbeda nyata. Kandungan GA3 pada kedua aksesi juga tidak memberikan pengaruh pada jumlah tunas yang tumbuh. Kandungan IAA dan GA3 di dalam sel dan jaringan garut tidak dipengaruhi oleh aksesi. Kandunguan ZPT dalam sel secara umum dipengaruhi oleh faktor internal berupa spesies, bagian tumbuhan, dan fase perkembangan (Salisbury dan Ross 1995). Di sisi lain pemberian ZPT secara eksternal dapat mempengaruhi pertumbuhan. Penelitian yang dilakukan oleh Lestari et al. (2008) menunjukkan bahwa tanaman garut yang diberi perlakuan GA3 dengan konsentrasi 50 ppm menghasilkan tinggi tanaman tertinggi, sedangkan yang diberi perlakuan GA3 dengan konsentrasi yang lebih tinggi mengakibatkan penurunan pertumbuhan pada tanaman garut. Kandungan α-amilase pada umbi garut aksesi Krajan lebih tinggi daripada aksesi Bantul.
8
dipengaruhi oleh faktor internal lain selain α-amilase. Taiz dan Zeiger (2010) menyatakan bahwa selain α-amilase, enzim yang berperan dalam pemecahan pati adalah β-amilase. Enzim β-amilase dalam hal ini diduga berperan lebih besar dalam perombakan pati untuk pertunasan pada umbi garut.
SIMPULAN
Kecepatan pertunasan umbi garut tidak dipengaruhi oleh aksesi tetapi dipengaruhi oleh kelembapan media dan penyinaran secara terpisah. Faktor fisiologi berupa hormon tidak mempengaruhi kecepatan pertunasan.
DAFTAR PUSTAKA
Akazawa T, Hara-Mishimura I. 1985. Topographic aspects of biosynthesis, extracellular section and intracellular storage of proteins in plant cells. J Annu. Rev. Plant Physiol 70: 441–472
AOAC. 1995. Official Methods of Analysis of the Association of Official Analytical Chemists. Washington DC (US): Association of Official Analytical Chemists, Inc.
Auzanneau J, Huyghe C, Escobar-Gutierrez AJ, Julier B, Gastal F, Barre P. 2011. Association study between the gibberellic acid insensitive gene and leaf length in a Lolium perenne L. synthetic variety. J BMC Plant Biology. 183(11): 1–13.
Beck E, Ziegler P. 1989. Biosynthesis and degradation of starch in higher plants. J Annu. Rev. Plant Physiol Plant Mol Biol 40: 95–117
Booker J, Chatfield S, Leyser O. 2003. Auxin acts in xylem-associated or medullary cells to mediate apical dominance. JPlant Cell 15(2): 495–507. Casimiro I, Marchant A, Bhalerao RP, Beeckman T, Dhooge S, Swarup R,
Graham N, Inze D, Sandberg G, Casero PJ, Bennett M. 2001. Auxin transport promotes Arabidopsis lateral root initiation. J Plant Cell. 13:843– 852.
Chaves MM, Pepeira JS, Maroco J, Rodrigues ML, Ricardo CPP, Osorio ML, Carvalho I, Faria T, Penheiro C. 2002. how plant cope with water stress in the field? photosynthesis and growth. J Annals of Botany. 89(7): 907-916 David w, Lawlor. 2002. Limitation to photosynthesis in water-stressed leaves:
stomata vs. metabolism and the role of ATP. J Annals of Botany 89:871-885 Gonzales HR, Sosa IH. 2006. Aloe vera (L.) Burm gel and sagu flour as a solid
support of culture medium for medicinal plants. J Rev. Cubana Plant Med. 11(1)
9
Jacobsen JV, Chandler PM. (1987). Gibberellin and abscisic acid in germinating cereals. In PJ Davies, ed. plant hormones and their role in plant growth and development. J Norwell, MA: Martinus-Nijhoff. 164-193
Kaur S, Gupta A K, Kaur N. 2000. Effect of GA3, kinetin and indole acetic acid on carbohydrate metabolism in chickpea seedlings germinating underwater stress. J Plant Growth Regul. 30(1):61-70
Kay DE. 1973. Root Crops. London (GB): The Tropical Product Institute.
Komiyama S, Yamazaki T, Hori E, Shida Y, Murayama A, Ikarashi T, Ohyama T. 1997. Degradation of storage starch in tulip bulb scales induced by cold temperature. J Soil Sci. Plant Nutrition. 68:2-29
Lambrechts H, Rook F, Kolloffel C. 1994. Carbohydrate status of tulip bulbs during cold-induced flower stalk elongation and flowering. J Plant Physiol. 104:515-520
Lestari GW, Solichatun, Sugiyarto. 2008. Pertumbuhan, kandungan klorofil, dan laju respirasi tanaman garut (Maranta arundinacea L.) setelah pemberian asam giberelat (GA3). J Bioteknologi. 5(1): 1-9
Liao X, Sun Q, Li X, Gao J. 2005. Effect of gibberellic acid and abscisic acid pretreatment on seedling growth and α-amylase activity in endosperms of wheat. Chinese Bul. Bot.1-12.
Liu Y, Wang QS, Ding YF, Li GH, Xu JX, Wang SH. 2011. Effects of external ABA, GA3 and NAA on the tiller bud outgrowth of rice is related to changes in endogenous hormones. J Plant Growth Regulation. 65(2): 247–254. Mahmoody M, Noori M. 2014. Effect of gibberellic acid on growth and
development plants and its relationship with abiotic stress. IJFAS. 3(20) 717–721.
Marsono Y, Wiyono P, Utomo Z. 2005. Indeks glikemik produk olahan garut (Maranta arundinacea L.) dan uji sifat fungsionalnya pada model hewan coba. Laporan Akhir Penelitian Rusnas Diversifikasi Pangan Pokok. Jakarta (ID): Kementrian Ristek.
Marta J, Laskowski, Mary E, William, Chad NH, Ian MS. 1995. Formation of root meristems is a two-stage process. J Development. 121: 3303-3310
Nolan RC, Ho TD. (1988). Hormonal regulation of α-amylase expression in barley aleurone layers: The effect of gibberellic acid removed and abscisic acid and phaseic acid treatments. J Plant Physiol. 88:588-593
Perchorowicz JT, Raynes DA, Jensen RG. 1981. Light limitation of photosynthesis and activation of ribulose bisphosphate carboxylase in wheat seedlings. J Proc. Nati. Acad. Sci. 78(5):2985-2989
Salisbury FB, Ross CW. 1995. Fisiologi tumbuhan. Jilid 1 Terjemahan Diah R. Lukman dan Sumaryo. Bandung (ID): ITB
Sastrapradja S, Soetjipto NW, Danimihardja S, Soejon (cek) R. 1977. Ubiubian. LBN 7, SDE 40. Proyek Sumber Daya Ekonomi. Bogor (ID): LBN-LIPI Taiz L, Zeiger E. 2010. Plant Physiology Fifth Edition. Sunderland (US): Sinauer
Associates, Inc.
10
absidic acid (ABA), and zeatin produced by Phanerochaeta chrysosporium ME446. J Plant Physiol. 22(3-4):105-110
11
LAMPIRAN
Lampiran 1 Hasil uji ANOVA jumlah tunas
Tests of Between-Subjects Effects
Dependent Variable: Jumlah_Tunas
Source Type III Sum
of Squares
df Mean Square F Sig.
Corrected Model 6.300a 7 .900 1.946 .094
Intercept 72.900 1 72.900 157.622 .000
Aksesi .000 1 .000 .000 1.000
kelembapan_media .900 1 .900 1.946 .173
Penyinaran 3.600 1 3.600 7.784 .009b
aksesi * kelembapan_media .400 1 .400 .865 .359
aksesi * penyinaran .900 1 .900 1.946 .173
kelembapan_media * penyinaran .400 1 .400 .865 .359
aksesi * kelembapan_media * penyinaran .100 1 .100 .216 .645
Error 14.800 32 .463
Total 94.000 40
Corrected Total 21.100 39
a. R Squared = .299 (Adjusted R Squared = .145)
12
Lampiran 2 Hasil uji ANOVA tinggi tunas
Tests of Between-Subjects Effects
Dependent Variable: tinggi
Source Type III Sum of
Squares
Df Mean Square F Sig.
Corrected Model 3751.936a 7 535.991 5.910 .000
Intercept 3626.168 1 3626.168 39.986 .000
Aksesi 300.030 1 300.030 3.308 .078
kelembapan_media 1391.810 1 1391.810 15.348 .000b
Penyinaran 1375.343 1 1375.343 15.166 .000b
aksesi * kelembapan_media .315 1 .315 .003 .953
aksesi * penyinaran 306.086 1 306.086 3.375 .075
kelembapan_media * penyinaran 373.627 1 373.627 4.120 .051
aksesi * kelembapan_media * penyinaran 4.727 1 4.727 .052 .821
Error 2901.938 32 90.686
Total 10280.043 40
Corrected Total 6653.874 39
a. R Squared = .564 (Adjusted R Squared = .468)
b. Berbeda nyata pada α = 0.05
13
RIWAYAT HIDUP
Penulis dilahirkan di Serang pada tanggal 09 Agustus 1993 dari ayah Busro dan ibu Suhaemi. Penulis merupakan anak ketiga dari empat bersadara. Tahun 2011 penulis lulus dari SMA Negeri 2 Kota Serang. Pada tahun yang sama, penulis lulus seleksi masuk IPB melalui jalur Seleksi Nasional Masuk Perguruan Tinggi Negeri (SNMPTN) tertulis di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam. Penulis mendapatkan beasiswa Badan Usaha Milik Negara (BUMN) pada tahun 2012-2014 dan Beasiswa Karya Salemba Empat (KSE) pada tahun 2014-2015.
Selama mengikuti perkuliahan, penulis menjadi asisten praktikum biologi dasar Tingkat Persiapan Bersama tahun ajaran 2014/2015 semester genap, asisten praktikum fisiologi tumbuhan tahun ajaran 2014/2015, dan asisten praktikum kultur jaringan tahun ajaran 2014/2015. Penulis juga mengikuti kepengurusan Serambi
Ruhiyah Mahasiswa FMIPA (SERUM-G) sebagai anggota divisi Syiar and Science
(SAS) pada tahun 2013 dan anggota divisi Class Rohis Management (CRM) pada
tahun 2014). Selain itu, pada tahun 2014 penulis juga menjadi bagian dari Badan Eksekutif Mahasiswa Keluarga Mahasiswa (BEM KM) IPB sebagai anggota Biro Badan Internal.