PRODUKSI SENYAWA AKTIF KAPANG ENDOFIT
UNTUK MENGHAMBAT PROLIFERASI SEL
KANKER PAYUDARA MCF-7 SECARA
IN VITRO
ERWAHYUNI ENDANG PRABANDARI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Aktivitas Antioksidan Ekstrak Rumput Laut Coklat (Sargassum echinocarpum) sebagai Pencegah Disfungsi Sel Endotelium Aorta Tikus Diabetes Melitus adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, 22 Nopember 2011
MUHAMAD FIRDAUS. Antioxidant activity of brown algae (Sargassum echinocarpum) as preventive on dysfunction of endothelium cell in aorta of
diabetes mellitus rats. Under supervision of MADE ASTAWAN as chairman, DEDDY MUCHTADI, TUTIK WRESDIYATI, SARWONO WASPADJI, and SETYAWATI S. KARYONO, as member committee.
Diabetes mellitus may cause oxidative stress. Chronic oxidative stress initiates the dysfunction of endothelium cell. Polyphenols are believed to have ability to prevent the dysfunction of endothelium cell. Marine brown algae contain polyphenol which is probably has good antioxidant activity. The aims of this study were to obtain the antioxidant activity and structure of phlorotannin, the lethal dose, the anti oxidative stress, and the anti endothelium cell dysfunction of
Sargassumechinocarpum extract. The Sargassumechinocarpum was collected in
April 2008 from the coastline of Talango island in Sumenep District of East Java, Indonesia and authenticated by the botanist, Research Centre of Oceanography, Indonesian Institute of Sciences. The antioxidant activity was assayed with DPPH radicals, the structure elucidation was assessed with spectroscopic methods, the lethal dose was investigated on mice, and the anti oxidative stress and the anti endothelium cell dysfunction were evaluated in diabetic rats. The experimental animals were divided into 5 groups, i.e.: (1) normal, (2) diabetic treated with 0, (3) 150, (4) 300, and (5) 450 mg/kg body weight of Sargassum echinocarpum extract,
respectively. Extracts were administered by oral gavages for 12 weeks. Diabetes was induced by single administration of streptozotocin (45 mg/kg, i.p.). Diabetes was confirmed ten days later in streptozotocin induced animals with blood glucose levels > 200 mg/dL. After the treatment period, the blood serum acquired was used for antioxidant enzymes assays while the thoracic aorta was used for vasodilatation assay. The result showed that the phlorotannin content of
Sargassum echinocarpum extract was 4.25 - 12.16 equivalent of mg
phloroglucinol/g extract. The strongest antioxidant activity obtained from the methanol extract of Sargassum echinocarpum, i.e. 67.77 %. The methanol extract
of Sargassum echinocarpum contained phloroglucinol and bifuhalol. The
methanol extract of Sargassum echinocarpum was relatively non-toxic. The mice
treated with 1250 mg/kg or more methanol extract of Sargassum echinocarpum
showed inhibition of body weight and necrosis on hepatocyte and tubules of kidney. It was affected by inhibition activity of gastrointestinal enzymes and initiation lipid peroxidation. The diabetic rats revealed the oxidative stress; meanwhile, the diabetic rats treated with 450 mg/kg extract showed the mild oxidative stress. It was caused by free radical scavenging and induction of antioxidant defense activity by extract. The diabetic rats exhibited the endothelial dysfunction; meanwhile, the diabetic rats treated with 450 mg/kgextract showed the mild endothelial dysfunction. It appeared that Sargassum echinocarpum
extract has the capability to preserve bioavailability of nitric oxide and protect the muscarinic receptor and endothelium cell against free radical damage.
MUHAMAD FIRDAUS. Aktivitas Antioksidan Rumput Laut Coklat (Sargassum echinocarpum) sebagai Pencegah Disfungsi Sel Endotelium Aorta Tikus Diabetes
Melitus. Di bawah bimbingan MADE ASTAWAN, selaku ketua serta DEDDY MUCHTADI, TUTIK WRESDIYATI, SARWONO WASPADJI, dan SETYAWATI S. KARYONO sebagai anggota.
Petanda awal terjadinya penyulit pada diabetes adalah didapatnya disfungsi sel endotelium. Tingginya kadar radikal bebas pada diabetes dapat mengakibatkan sel endotelium pembuluh darah menjadi lebih permeabel, adesif dan kontraktif. Pencegahan disfungsi sel endotelium pada diabetes dapat dilakukan dengan mengonsumsi antioksidan seperti vitamin C, E, dan polifenol. Antioksidan ini dapat mencegah disfungsi sel endotelium karena berkemampuan menurunkan radikal bebas dalam tubuh. Florotanin adalah polifenol yang terkandung dalam rumput laut coklat. Tujuan penelitian ini adalah mendapatkan aktivitas antioksidan, struktur florotanin, dosis kematian, anti stres oksidatif, dan anti disfungsi sel endotelium aorta tikus diabetes melitus ekstrak rumput laut coklat (Sargassum echinocarpum).
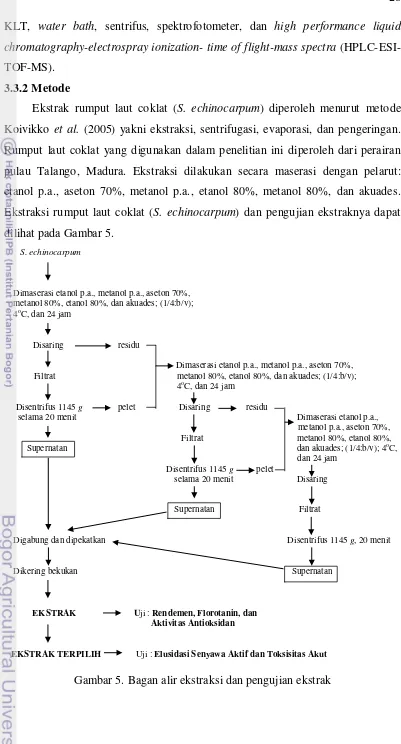
Sargassum echinocarpum diperoleh April 2008 dari perairan pulau Talango,
kabupaten Sumenep, Jawa Timur. Rumput laut coklat telah diidentifikasi oleh botanis Pusat Penelitian Oseanografi, Lembaga Ilmu Pengetahuan Indonesia. Rumput laut selanjutnya dikeringkan, ditepungkan, dimaserasi, dipekatkan dan dikering bekukan untuk mendapatkan ekstrak. Tepung rumput laut dimaserasi etanol p.a., metanol p.a., aseton 70%, etanol 80%, metanol 80% dan akuades. Kadar florotanin ekstrak ditentukan dengan spektrofotometer dan menggunakan floroglusinol sebagai standar. Uji aktivitas antioksidan ekstrak dilakukan dengan metode diphenyl picrylhydracyl (DPPH). Identifikasi struktur senyawa aktif dalam
ekstrak beraktivitas antioksidan terkuat dilakukan secara spektroskopik yaitu ultra ungu, infra merah, dan spektra massa.
Uji toksisitas akut ekstrak Sargassum echinocarpum dilakukan pada mencit
(Mus musculus) strain BALB/c berjenis kelamin jantan dan betina berumur 2
bulan. Ekstrak diberikan satu jam sebelum hewan uji diberi ransum dengan dosis 0 mg/kg BB, 625 mg/kg BB, 1250 mg/kg BB, 2500 mg/kg BB, dan 5000 mg/kg BB. Selama 14 hari pengujian dilakukan pengamatan berat badan dan jumlah kematian, sedang pada akhir masa uji diamati perubahan histopatologis hati dan ginjal.
Uji oksidatif stres dan disfungsi sel endotelium dilakukan pada tikus (Rattus norvegicus) strain Sprague-Dawley jantan berumur 2,5 bulan. Tikus diabetes
melitus didapat dengan cara menginjeksi streptozotocin dosis tunggal 45 mg/kg
untuk uji disfungsi sel endotelium.
Hasil menunjukkan kadar florotanin ekstrak Sargassum echinocarpum berkisar
antara 4,25 – 12,16 mg floroglusinol/g ekstrak. Ekstrak metanol Sargassum echinocarpum mempunyai aktivitas antioksidan paling kuat yaitu 67,77 %.
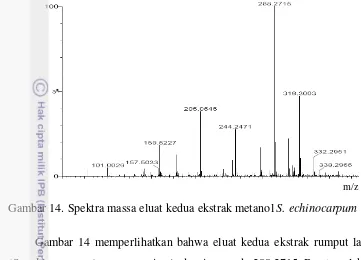
Berdasar analisis spektroskopi ultra ungu, infra merah, dan spektra massa pada kromatografi cair kinerja tinggi - Electrospray Ionization – Time of Flight – Mass Spectra mode ion positif menunjukkan bahwa senyawa aktif yang terkandung
dalam ekstrak metanol Sargassum echinocarpum adalah floroglusinol dan
bifuhalol.
Hasil uji toksisitas akut menunjukkan bahwa ekstrak metanol Sargassum echinocarpum tidak menyebabkan kematian hewan uji dan dapat digolongkan
sebagai bahan relatif tidak toksik. Ekstrak metanol Sargassum echinocarpum
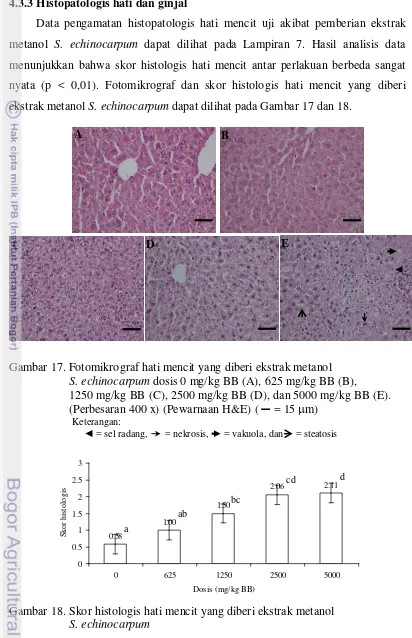
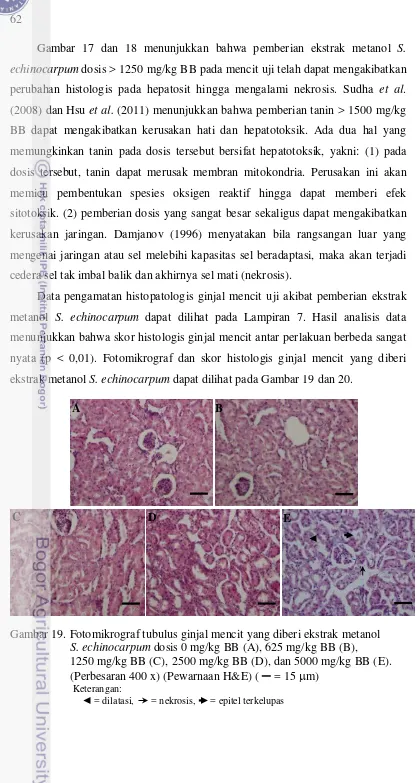
dosis 1250 mg/kg BB atau lebih dapat menghambat kenaikan berat badan. Hasil histopatologis hati dan ginjal menunjukkan pemberian dosis 1250 mg/kg BB atau lebih mengakibatkan nekrosis pada hepatosit dan tubulus ginjal. Hal ini menunjukkan ekstrak aman dikonsumsi pada dosis ≤ 625 mg/kg BB.
Hasil uji stres oksidatif menunjukkan berat badan tikus diabetes melitus mengalami penurunan hingga 48%, sementara itu tikus diabetes melitus yang diberi ekstrak metanol Sargassum echinocarpum khususnya 450 mg/kg BB dapat
mencegah penurunan berat badan dan mengalami kenaikan berat badan sebesar 6%. Hal ini menunjukkan ekstrak metanol Sargassum echinocarpum dapat
meningkatkan sintesis glikogen. Hasil uji menunjukkan kadar glukosa darah tikus diabetes melitus tetap di atas batas kadar glukosa darah tikus diabetes melitus terinduksi streptozotocin yaitu 200 mg/dL, sementara itu pemberian ekstrak
metanol Sargassum echinocarpum dosis 450 mg/kg BB dapat menurunkan kadar
glukosa darah hingga mencapai 196 mg/dL. Hal ini menunjukkan ekstrak metanol
Sargassum echinocarpum dapat memberikan efek hipoglikemik. Hasil uji
menunjukkan bahwa serum tikus diabetes melitus mengandung malondialdehid sebesar 6,76 nmol/mL, sedangkan tikus diabetes melitus yang diperlakukan dengan ekstrak metanol Sargassum echinocarpum dosis 450 mg/kg BB kadarnya
turun menjadi 0,98 nmol/mL. Hal ini menunjukkan bahwa ekstrak metanol
Sargassum echinocarpum dapat membersihkan radikal bebas yang terbentuk
akibat hiperglikemia. Hasil uji menunjukkan aktivitas antioksidan enzim serum tikus diabetes melitus (superoksida dismutase, katalase, dan glutation peroksidase)
sebesar 12,96 U/mL; 67,17 μmol H2O2/min/mL; dan 0,8 mU/min/mL, sementara
itu pemberian ekstrak metanol Sargassum echinocarpum dosis 450 mg/kg BB
pada tikus diabetes melitus dapat mempertahankan aktivitas superoksida dismutase tetap tinggi, yaitu 31,29 U/mL, katalase 162,40 μmol H2O2/min/mL,
dan glutation peroksidase 2,01 mU/min/mL. Hal ini menunjukkan bahwa ekstrak metanol Sargassum echinocarpum dapat meningkatkan ekspresi dan aktivitas
enzim tersebut. Hasil uji menunjukkan profil negatif kandungan Cu,Zn-SOD ginjal dan hati pada tikus diabetes melitus mencapai 64,49% dan 67,87%, namun tikus diabetes melitus yang diperlakukan ekstrak metanol Sargassum echinocarpum dosis 450 mg/kg BB dapat menurunkan profil negatif kandungan
metanol menunjukkan bahwa persentase vasorelaksasi tikus diabetes melitus sebesar 14,41%, namun dengan pemberian ekstrak metanol
Sargassum echinocarpum dosis 450 mg/kg BB dapat meningkatkan vasorelaksasi
aorta tikus diabetes melitus menjadi 67,84%. Hal ini menunjukkan bahwa ekstrak dapat meningkatkan ketersediaan nitrit oksida. Sensitivitas muskarinik reseptor sel endotelium pada tikus diabetes melitus sebesar -5,49 (log M), namun dengan pemberian ekstrak metanol Sargassum echinocarpum dosis 450 mg/kg BB
sensitivitas muskarinik reseptor sel endotelium tikus diabetes melitus meningkat menjadi -6,23 (log M). Hal ini berarti bahwa kerusakan konformasi muskarinik reseptor oleh radikal bebas dapat dicegah oleh ekstrak metanol Sargassum echinocarpum. Rasio sel endotelium aorta tikus diabetes melitus sebesar 65,36 %,
namun pemberian ekstrak metanol Sargassum echinocarpum dosis 450 mg/kg BB
dapat mencegah kerusakan morfologi aorta tikus diabetes melitus dan meningkatkan rasio sel endotelium menjadi 89,85%. Hal ini menunjukkan bahwa ekstrak metanol Sargassum echinocarpum mampu mencegah kerusakan sel
endotelium aorta oleh radikal bebas.
RUMPUT LAUT COKLAT (
Sargassum echinocarpum
)
SEBAGAI PENCEGAH DISFUNGSI SEL ENDOTELIUM
AORTA TIKUS DIABETES MELITUS
MUHAMAD FIRDAUS
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Hak Cipta milik IPB, tahun 2011
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
Penguji pada Ujian Tertutup : Dr. Ir. Sedarnawati Yasni, M. Agr. Dr. Ir. Nurjanah, M.Si.
Disfungsi Sel Endotelium Aorta Tikus Diabetes Melitus Nama : Muhamad Firdaus
NIM : F-226010061
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Made Astawan, M.S Ketua
.
Prof. Dr. Ir. Deddy Muchtadi, M.S.
Anggota Anggota
Prof. Dr. drh. Tutik Wresdiyati. PA.Vet
Prof. Dr. dr. Sarwono Waspadji, SpPD-KEMD Dr. dr. Setyawati S. K., M.Kes Anggota Anggota
.
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana Ilmu Pangan
Dr. Ir. Ratih Dewanti Hariyadi, M.Sc. Dr. Ir. Dahrul Syah, M.Agr.Sc.
Puji syukur penulis panjatkan kepada Allah SWT atas segala rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan disertasi ini. Judul penelitian ini adalah “Aktivitas Antioksidan Ekstrak Rumput Laut Coklat (Sargassum echinocarpum) sebagai Pencegah Disfungsi Sel Endotelium Aorta Tikus Diabetes
Melitus”.
Penulis mengucapkan terima kasih dan penghargaan sebesar-besarnya kepada Prof. Dr. Ir. Made Astawan MS. selaku ketua komisi pembimbing serta Prof. Dr. Ir. Deddy Muchtadi, MS., Prof. Dr. drh. Tutik Wresdiyati, PAVet., Prof. Dr. dr. Sarwono Waspadji, SpPD-KEMD, dan Dr. dr. Setyawati S. Karyono, MKes sebagai anggota komisi pembimbing atas segala pengarahan dan bimbingannya sehingga penulis dapat menyelesaikan penelitian dan penulisan ini dengan sebaik-baiknya.
Penulis juga mengucapkan terima kasih kepada Dr. Ir. Sugiyono, M.App.Sc sebagai penguji luar komisi dalam ujian kualifikasi Doktor, Dr. Ir. Sedarnawati Yasni, M.Agr. dan Dr. Ir. Nurjanah, M.Si., dalam ujian tertutup, dan Prof. Dr. Rachmaniar Rachmat, Apt. dan Dr. Ir. Nurheni Sri Palupi, M.Si. pada ujian terbuka atas segala masukannya dalam menyempurnakan rancangan penelitian, penulisan dan isi disertasi.
Penulis juga menyampaikan terima kasih kepada Rektor Universitas Brawijaya dan Dekan Fakultas Perikanan dan Ilmu Kelautan Universitas Brawijaya yang telah mengijinkan dan mendukung penulis dalam melanjutkan pendidikan Doktor ilmu pangan. Terima kasih juga kepada ketua Program Studi Ilmu Pangan, Kepala Lab. Farmakologi Fak. Kedokteran Univ. Brawijaya dan Lab. Histologi FKH IPB dan Dr. Ir. Anis Mahsunah, M.Sc. (Laboratorium Bioteknologi Industri, Balai Pengkajian Bioteknologi, BPPT) atas bantuan analisis spektra massa.
Terima kasih kepada Direktorat Jenderal Pendidikan Tinggi atas dukungan beasiswa BPPS dan hibah penelitian melalui program Hibah Bersaing (2008) dan Hibah Doktor (2009) sehingga penulis dapat menyelesaikan penelitian dengan baik.
Ungkapan terima kasih yang tulus disampaikan kepada kedua orang tua yaitu bapak H. Muhammad Ad dan ibu Hj. Chodijah serta mertua bapak Drs. H. Suroyo dan ibu Hj. Sri Wahyurini yang telah mendidik dan selalu mendoakan keberhasilan kepada penulis. Penulis ucapkan terima kasih kepada saudara-saudara dan istri tercinta Rina Hernugraheni S.P., M.P. dan anak-anakku tersayang Salsabila Firdausa, Fadila Ulil Albab, Ibnu Rasyid Ridlo, dan Hafizhuddin Hilmi yang sangat mendukung penulis menyelesaikan studi program Doktor.
Bogor, Nopember 2011
Penulis dilahirkan di Surabaya pada tanggal 19 September 1968 sebagai anak kedua dari pasangan H. Muhammad Ad dan Hj. Chodidjah. Pendidikan sarjana ditempuh di program studi Teknologi Hasil Perikanan, Fakultas Perikanan Universitas Brawijaya, lulus pada tahun 1992. Pada tahun 1998, penulis diterima di program studi Teknologi Hasil Pertanian pada program Pascasarjana Universitas Brawijaya dan menamatkannya pada tahun 2000. Kesempatan untuk melanjutkan ke program doktor studi Gizi Masyarakat dan Keluarga IPB diperoleh pada tahun 2001, namun pada semester kedua tahun ajaran 2001-2002, penulis pindah program studi Ilmu Pangan IPB. Beasiswa pendidikan pascasarjana diperoleh dari Departemen Pendidikan Nasional Republik Indonesia melalui Beasiswa Pendidikan PascaSarjana (BPPS).
Penulis bekerja sebagai staf pengajar di Fakultas Perikanan Universitas Hang Tuah Surabaya sejak tahun 1995 dan pada tahun 2005 diterima sebagai staf pengajar di Fakultas Perikanan dan Ilmu Kelautan Universitas Brawijaya Malang. Bidang penelitian yang menjadi perhatian peneliti adalah rumput laut: kimia dan bioaktivitasnya.
Selama mengikuti program S3 penulis telah menerbitkan dua artikel pada majalah terakreditasi Dirjen Dikti yaitu: (1) “Protective effects of Sargassum echinocarpum against endothelial dysfunction in diabetic rats” pada Medical
Halaman
DAFTAR TABEL ... iii
DAFTAR GAMBAR ………... iv
DAFTAR LAMPIRAN ... vii
SINGKATAN ... viii
1 PENDAHULUAN ... 1
1.1 Latar Belakang ………...………... 1
1.2 Tujuan Penelitian ………...………... 3
1.3 Hipotesis ... 4
2 TINJAUAN PUSTAKA ... 5
2.1 Diabetes Melitus ... 5
2.2 Stres Oksidatif ... 9
2.2.1 Stres Oksidatif dan Kerusakan Sel ... 12
2.2.2 Stres Oksidatif dan Membran Sel ... 13
2.2 3 Sistem Proteksi terhadap Radikal Bebas ... 14
2.3 Sel Endotelium ... 16
2.3.1 Struktur dan Fungsi ... 16
2.3.2 Disfungsi Sel Endotelium ... 19
2.4 Florotanin ... 22
3 BAHAN DAN METODE ... 27
3.1 Tempat dan Waktu ...……...….………... 27
3.2 Metode penelitian ... 27
3.3 Aktivitas antioksidan dan Elusidasi struktur ... 27
3.3.1 Bahan ... 27
3.3.2 Metode ... 28
3.3.3 Rendemen ... 29
3.3.4 Florotanin ... 29
3.3.5 Aktivitas antioksidan ... 29
3.3.6 Elusidasi struktur ... 30
3.4 Toksisitas akut ... 30
3.4.1 Bahan ... 30
3.4.2 Metode ... 30
3.5 Stres oksidatif dan Disfungsi sel endotelium ... 34
3.5.1 Bahan dan Alat ... 34
3.5.2 Metode ... 34
3.5.3 Glukosa darah ... 35
3.5.4 Hemoglobin A1c 3.5.5 Peroksida Lemak ... ... 35 35 3.5.6 Superoksida dismutase ... 36
3.5.7 Katalase ... 38
3.5.9 Cu,Zn-SOD hati dan ginjal secara imunohistokimia ... 41
3.5.10 Vasorelaksasi ... 42
3.5.11 Dosis efektif 50% ... 43
3.5.12 Rasio sel endotelium ... 43
3.6 Analisis data ... 45
4 HASIL DAN PEMBAHASAN ... 47
4.1 Identitas rumput laut coklat ... 47
4.2 Aktivitas antioksidan dan Identitas florotanin ... 48
4.2.1 Rendemen ... 48
4.2.2 Florotanin ... 49
4.2.3 Aktivitas antioksidan ... 50
4.2.4 Identitas florotanin ... 52
4.3 Toksisitas akut ... 59
4.3.1 Berat badan ... 59
4.3.2 LD50 ... 60
4.3.3 Histopatologis hati dan ginjal ... 61
4.4 Stres oksidatif ... 64
4.4.1 Berat badan ... 64
4.4.2 Glukosa darah ... 65
4.4.3 Hemoglobin A1c ... 67 4.4.4 MDA ... 68
4.4.5 SOD ... 70
4.4.6 Cu,Zn-SOD ginjal ... 71
4.4.7 Cu,Zn-SOD hati ... 74
4.4.8 Katalase ... 77
4.4.9 GSH-Px ... 78
4.5 Disfungsi sel endotelium ... 79
4.5.1 Vasorelaksasi ... 79
4.5.2 ED50 ... 81
4.5.3 Rasio sel endotelium ... 83
5 SIMPULAN DAN SARAN ... 86
5.1 Simpulan ... 86
5.2 Saran ... 86
DAFTAR PUSTAKA ………... 87
Halaman
1 Kriteria Diagnosis Diabetes Melitus ... 5
2 Klasifikasi Etiologis Diabetes Melitus ... 6
3 Kelarutan florotanin dalam berbagai pelarut ... 23
4 Harga Probit ... 31
Halaman
1 Efek kegemukan terhadap patogenesis diabetes melitus tipe 2 ... 8
2 Sistem detoksifikasi radikal bebas oleh enzim antioksidan ... 15
3 Mekanisme disfungsi sel endotelium akibat diabetes ... 21
4 Rumus bangun floroglusinol dan turunannya ... 23
5 Bagan alir ekstraksi dan pengujian ekstrak ... 28
6 Morfologi rumput laut coklat ... 47
7 Hubungan kadar florotanin dan aktivitas antioksidan ekstrak S. echinocarpum dalam berbagai pelarut ... 51
8 Partisi metanol ... 53
9 Eluat pertama dan kedua ... 53
10 Noktah floroglusinol, eluat pertama dan kedua ... 54
11 Spektra serapan ultra violet eluat kedua ekstrak metanol S. echinocarpum ... 54
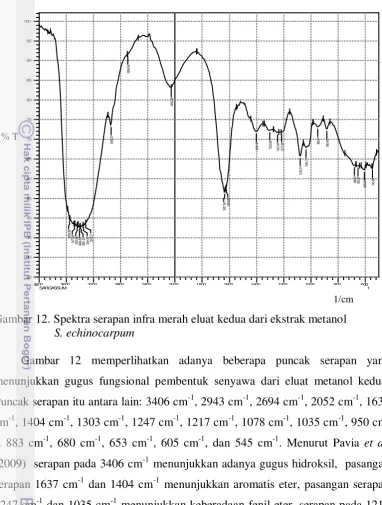
12 Spektra serapan infra merah eluat kedua ekstrak metanol S. echinocarpum ... 56
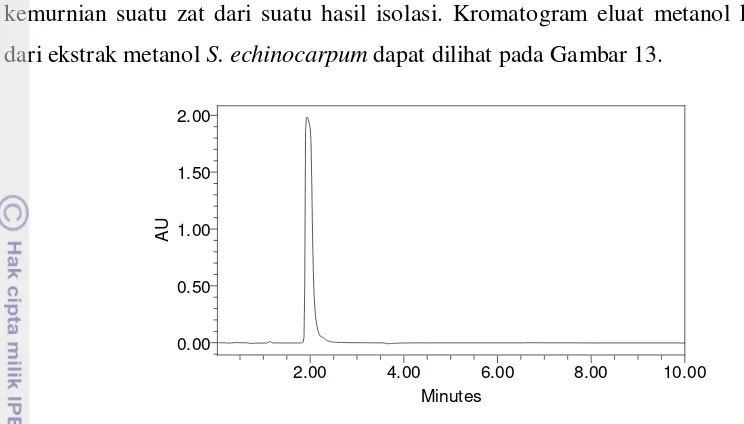
13 Kromatogram eluat kedua ekstrak metanol S. echinocarpum ... 57
14 Spektra masa eluat kedua ekstrak metanol S. echinocarpum ... 58
15 Struktur senyawa eluat kedua ... 58
16 Persentase kenaikan berat badan mencit akibat pemberian ekstrak metanol S. echinocarpum ... 59
17 Fotomikrograf hati mencit yang diberi ekstrak metanol S. echinocarpum dosis 0 mg/kg BB, 625 mg/kg BB, 1250 mg/kg BB, 2500 mg/kg BB, dan 5000 mg/kg BB ... 61
19 Fotomikrograf tubulus mencit yang diberi ekstrak metanol
S. echinocarpum dosis 0 mg/kg BB, 625 mg/kg BB,
1250 mg/kg BB, 2500 mg/kg BB, dan 5000 mg/kg BB ... 62 20 Skor histologis ginjal mencit yang diberi ekstrak metanol
S.echinocarpum ... 63
21 Kenaikan berat badan tikus normal dan tikus diabetes melitus yang diberi ekstrak metanol S. echinocarpum selama masa
penelitian ... 64 22 Kadar glukosa darah tikus normal dan tikus diabetes melitus
yang diberi ekstrak metanol S. echinocarpum selama masa
penelitian ... 66 23 Persentase HbA1c tikus normal dan tikus diabetes melitus
yang diberi ekstrak metanol S. echinocarpum ... 67
24 Kadar MDA serum tikus normal dan tikus diabetes melitus
yang diberi ekstrak metanol S. echinocarpum ... 69
25 Aktivitas SOD serum tikus normal dan tikus diabetes melitus
yang diberi ekstrak S. echinocarpum ... 70
26 Fotomikrograf profil Cu,Zn-SOD ginjal tikus normal (A), tikus diabetes melitus yang diberi ekstrak metanol
S. echinocarpum dosis 0 mg/kg (B), 150 mg/kg (C),
300 mg/kg (D), dan 450 mg/kg (E) ... 72
27 Persentase jumlah sel tubuli renalis pada berbagai tingkat kandungan Cu,Zn-SOD ginjal tikus normal dan tikus diabetes melitus yang diberi ekstrak metanol
S. echinocarpum ... 72
28 Fotomikrograf profil Cu,Zn-SOD hati tikus normal (A), tikus diabetes melitus yang diberi ekstrak metanol S. echinocarpum dosis 0 mg/kg (B), 150 mg/kg (C),
300 mg/kg (D), dan 450 mg/kg (E) ... 75
29 Persentase jumlah sel hati pada berbagai tingkat kandungan Cu,Zn-SOD hati tikus normal dan tikus diabetes melitus
yang diberi ekstrak metanol S. echinocarpum ... 75
30 Aktivitas katalase serum tikus normal dan tikus diabetes
melitus yang diberi ekstrak metanol S. echinocarpum ... 77
31 Aktivitas GSH-Px serum tikus normal dan tikus diabetes
32 Vasorelaksasi tikus normal dan tikus diabetes melitus
yang diberi ekstrak S.echinocarpum ... 79
33 ED50 tikus normal dan tikus diabetes melitus yang diberi
ekstrak metanol S.echinocarpum ... 81
34 Fotomikrograf sel endotelium aorta tikus normal (A), tikus diabetes melitus yang diberi ekstrak metanol S. echinocarpum dosis 0 mg kg-1 (B), 150 mg kg-1 (C),
300 mg kg-1 (D), dan 450 mg kg-1 (E). ... 83
35 Rasio sel endotelium aorta tikus normal dan tikus diabetes
Halaman
1 Sertifikat hasil identifikasi rumput laut coklat ... 107
2 Rendemen ekstraksi ... 108
3 Kadar florotanin ... 110
4 Aktivitas antioksidan ... 113
5 Keterangan kelaikan etik penelitian ... 116
6 Berat badan (uji toksisitas) ... 117
7 Histopatologis hati dan ginjal ... 119
8 Berat badan (uji diabetes) ... 123
9 Glukosa darah ... 126
10 Hemoglobin A1c ... 128
11 Malondialdehida (MDA) ... 130
12 Aktivitas Superoksida Dismutase (SOD) ... 132
13 Profil Cu,Zn-SOD ginjal ... 135
14 Profil Cu,Zn-SOD hati ... 140
15 Aktivitas katalase ... 145
16 Aktivitas glutation peroksidase ... 148
17 Vasorelaksasi ... 150
18 Dosis Efektif (ED50) ... 152
ACh Acethylcholine
AGEs Advances Glycation End Products
Akt protein kinase B
AMPK Adenosine Monophosphate Kinase
BH4
CaM Caldmodulin
Tetrahydrobiopterine
CAT Catalase
cGMP Cyclic-Guanylate Monophosphate
Cu,Zn-SOD Cuprum,Zinc-Superoxide Dismutase
DM Diabetes melitus
EDRF Endothelium Derived Relaxing Factors
EDCF Endothelium Derived Contracting Factors
EDHF Endothelium Derived Hyperpolarization Factors
eNOS endothelium Nitric Oxide Synthase
ecSOD extracellular Superoxide Dismutase
ERK extracellular signal regulated kinase
ESI Electrospray Ionization
ET-I Endothelin-I
GSH-Px Glutathione peroxidase
glu-ecSOD extracellular Superoxide Dismutase terglikasi
HbA1c Hemoglobin tipe A1c
NO Nitric Oxide
(hemoglobin terglikasi) NOS Nitric Oxide Synthase
PGI2
PGE
Prostasiklin
2
PI-3 Phosphatidilinositol-3
Prostaglandin
Stz Streptozotocin
SOD Superoxide Dismutase
1.1 Latar Belakang
Diabetes melitus (DM) adalah penyakit metabolik yang telah dikenal semenjak peradaban Mesir kuno. Jumlah penyandang DM di Indonesia pada tahun 2000 sebanyak 8,4 juta dan diperkirakan tahun 2030 akan mencapai 21,3 juta. Pemakaian insulin telah meningkatkan angka harapan hidup penyandang penyakit ini, namun seiring peningkatan tersebut penyakit penyulit (komplikasi) DM makin banyak ditemui (Kuller et al., 2000; Perkumpulan Endokrinologi Indonesia
(Perkeni), 2002; Wild et al., 2004).
Pengerasan dan penyempitan pembuluh arteri besar (aorta) atau aterosklerosis adalah salah satu manifestasi penyulit DM. Kejadian aterosklerosis pada penyandang diabetes umumnya lebih dini dan lebih sering dibandingkan bukan diabetes. Petanda awal terjadinya penyulit ini pada penyandang diabetes adalah didapatnya disfungsi sel endotelium. Kondisi ini salah satunya dicirikan oleh tingginya vasokonstriksi dibanding vasorelaksasi dan peningkatan stres oksidatif (Bierman, 1992; Gunawan, 2004; Kapuku et al., 2006; Bakker et al.,
2009).
Sel endotelium adalah sel epitel pembatas intima pembuluh darah dari komponen darah. Sel ini berperan penting dalam mengatur tonus otot polos pembuluh darah. Sel ini akan melepaskan faktor relaksasi (EDRF = Endothelium Derived Relaxing Factors) dan faktor kontraksi (EDCF = Endothelium Derived Contracting Factors) agar tekanan darah tetap berjalan normal. Sel ini juga
berperan pada migrasi dan pertumbuhan sel otot polos pembuluh darah, menghambat proses koagulasi darah dan merangsang disolusi bekuan darah yang telah terbentuk pada lumen pembuluh darah, dan mengatur adesi dan migrasi sel-sel radang pada dinding pembuluh darah. Tingginya kadar radikal bebas pada diabetes dapat mengakibatkan sel endotelium pembuluh darah menjadi lebih permeabel, adesif dan kontraktif (Haller, 1997; Hurst dan Lee, 2003).
Kerusakan sel endotelium pada keadaan hiperglikemik diawali oleh adanya peningkatan pembentukan anion superoksida (O2-). Radikal ini selanjutnya dapat
hidroksil bersifat sangat reaktif dan dapat bereaksi dengan komponen asam lemak tidak jenuh jamak (PUFA = Polyunsaturated Fatty Acids) yang banyak terdapat
pada membran sel. Reaksi ini dapat menimbulkan peroksidasi lemak dan membentuk radikal lipid peroksida. Radikal-radikal ini selanjutnya dapat merusak sel endotelium sehingga sel ini kehilangan fungsinya atau bahkan memberi efek sitotoksik (Flavahan, 1992; Baynes dan Thorpe, 1999; De Mattia et al., 2003).
Stres oksidatif adalah suatu keadaan adanya kadar dan aktivitas antioksidan enzim yang lebih rendah dibanding radikal bebas dalam tubuh. Saat hiperglikemik, radikal bebas dapat terbentuk secara tak terkendali melalui beberapa mekanisme, seperti: autooksidasi glukosa, aktivasi lintasan glikasi, dan pembentukan peroksinitrit. Secara selular, kegagalan respirasi pada mitokondria dan sintesis nitrit oksida (NO) menjadi penyebab pembentukan radikal bebas secara berlebih. Saat hiperglikemik kadar antioksidan dalam tubuh rendah karena banyak digunakan untuk menetralisasi radikal bebas dan antioksidan enzim tubuh menjadi terinaktivasi akibat mengalami glikasi (Bonnefont-Rousselot et al, 2000;
Evans et al. 2002; Maritim et al., 2003, Martin-Gallan et al, 2003).
Pencegahan disfungsi sel endotelium pada diabetes dapat dilakukan dengan mengonsumsi antioksidan seperti vitamin C, E, dan polifenol (Evans et al., 2002;
Hurst and Lee, 2003; Lee et al., 2004 dan Vita, 2005). Antioksidan ini dapat
mencegah disfungsi sel endotelium karena berkemampuan menurunkan radikal bebas dalam tubuh. Namun polifenol mempunyai konstanta kecepatan reaksi pendonoran hidrogen ke radikal bebas lebih besar dibanding vitamin E dan C (Rice-Evans et al., 1997) dan di samping itu bioaktif ini mampu menstimulasi
transkripsi dan memodulasi antioksidan enzim (Masella et al., 2005; Rodrigo et al., 2011).
Salah satu polifenol yang menunjukkan efektivitas pendonor hidrogen adalah florotanin (Shibata et al., 2008). Florotanin adalah polifenol yang
terkandung dalam rumput laut coklat (Quideau et al., 2011). Senyawa ini tersusun
Beberapa studi menunjukkan bahwa florotanin dapat menghambat pembentukan dan aktivitas radikal bebas. Wei et al. (2003) dan Mori et al. (2003)
melaporkan bahwa florotanin dari Sargassum kljelmanianum dan S. micracantum
dapat menghambat peroksidasi lemak. Kang et al. (2004) memperlihatkan bahwa
florotanin dari Ecklonia stolonifera mampu mencegah pembentukan spesies
oksigen reaktif. Okada et al. (2004) menunjukkan florotanin dari Eisenia bicyclis
mampu menghambat pembentukan anion superoksida dan advanced glycation end products (AGEs). Florotanin dari ekstrak S. ringgoldianum dapat berfungsi
sebagai pembersih (scavenger) radikal bebas (Nakai et al., 2006).
Sargassum tergolong rumput laut yang potensinya sangat besar dan
sebarannya hampir ada di seluruh pantai perairan Indonesia. Ada 11 jenis algae
Sargassum yang dikenal ada di perairan Indonesia, yaitu: S. echinocarpum, S. binderi, S. crassifolium, S. plagyophyllum, S. mollerii, S. siliquosum, S. hystrix, S.
gracilimum, S. duplicatum, S. cinereum, dan S. polycystum (Kadi 2005). Studi
pendahuluan mendapati S. polycystum, S. duplicatum dan S. cinereum terdapat di
perairan pantai Tanjung Benoa, Bali, sementara itu S. echinocarpum, S. polycystum, S. duplicatum dan S. filipendula terdapat di perairan pantai Pulau
Talango, Kabupaten Sumenep, Madura.
Florotanin rumput laut coklat telah menunjukkan berkemampuan sebagai penghambat aktivitas dan pembentukan radikal bebas serta peroksidasi lemak. Namun aktivitasnya sebagai komponen penurun stres oksidatif dan pencegah disfungsi sel endotelium akibat hiperglikemik serta penggunaan rumput laut coklat dari perairan Indonesia, seperti S. echinocarpum sebagai sumber komponen
aktif beraktivitas antioksidan belum pernah dilakukan. Oleh karena itu perlu diuji peran ekstrak rumput laut coklat (S. echinocarpum) sebagai komponen pencegah
stres oksidatif dan pencegah disfungsi sel endotelium pada tikus diabetes.
1.2 Tujuan Penelitian
Tujuan umum penelitian ini adalah untuk mengevaluasi aktivitas antioksidan ekstrak rumput laut coklat (S. echinocarpum) sebagai pencegah
- Mendapatkan rendemen, kadar florotanin, aktivitas antioksidan dan identitas bioaktif ekstrak rumput laut coklat (S. echinocarpum).
- Mendapatkan toksisitas akut ekstrak rumput laut coklat (S. echinocarpum).
- Mengevaluasi kemampuan ekstrak rumput laut coklat (S. echinocarpum)
dalam menurunkan kadar glukosa darah, HbA1c
- Mengevaluasi kemampuan ekstrak rumput laut coklat (S. echinocarpum)
dalam mempertahankan vasorelaksasi, sensitivitas reseptor dan rasio sel endotelium tikus diabetes melitus.
dan lipid peroksida serum dan mempertahankan aktivitas superoksida dismutase, katalase, dan glutation peroksidase serum serta profil Cu,Zn-SOD ginjal dan hati tikus diabetes melitus.
1.3 Hipotesis
Hipotesis penelitian ini adalah:
- Ekstrak rumput laut coklat (S. echinocarpum) mengandung florotanin,
beraktivitas antioksidan, dan senyawa aktifnya adalah turunan floroglusinol.
- Ekstrak rumput laut coklat (S. echinocarpum) tergolong tidak toksik.
- Ekstrak rumput laut coklat (S. echinocarpum) mampu menurunkan kadar
glukosa, HbA1c
- Ekstrak rumput laut coklat (S. echinocarpum) mampu mempertahankan
vasorelaksasi, sensitivitas reseptor dan rasio sel endotelium aorta tikus diabetes melitus.
2.1 Diabetes melitus
Diabetes melitus adalah penyakit gangguan metabolik yang telah lama dikenal dalam peradaban manusia. Georg Ebers seorang ahli Mesir kuno pada tahun 1862 mendapati suatu lembaran (papirus) dalam kuburan di Thebes tentang catatan adanya penyakit ini pada masyarakat Mesir kuno (± 1550 SM). Araeus pada abad pertengahan memberi gambaran bahwa tubuh penyandang penyakit ini menyusut menjadi air seni dan menyebutnya dengan istilah diabetes, sementara itu Willis dalam pengamatannya mendapati bahwa air seni penyandang penyakit ini terasa manis dan kondisi ini disebutnya dengan istilah melitus. Saat ini bila seseorang sering mengeluarkan air seni dan air seninya mengandung glukosa maka dapat dinyatakan orang tersebut menderita diabetes melitus (Pushparaj et al.
2001).
Diabetes melitus merupakan sindrom multifaktorial metabolik yang dicirikan oleh adanya hiperglikemia sebagai akibat gangguan sekresi insulin, kerja insulin atau kombinasinya. Gejala umumnya adalah poliuria (sering kencing), polidipsia (rasa haus yang terus-menerus), kehilangan berat badan dan kadang-kadang polifagia (perasaan lapar yang berlebih). Kriteria diagnosis diabetes melitus dapat dilihat pada Tabel 1.
Tabel 1. Kriteria Diagnosis Diabetes Melitus
1. HbA1C
2. Gula darah puasa ≥ 6,5 % ≥ 126 mg/dL
Puasa: tidak mengkonsumsi sumber kalori paling tidak selama 8 jam 3. Gula darah 2 jam ≥ 200 mg/dL pada uji toleransi gula secara oral
Tes toleransi glukosa oral (TTGO) yaitu pengujian 75 g glukosa anhidrat yang dilarutkan dalam air
4. Gejala diabetes dengan konsentrasi glukosa darah ≥ 200 mg/dL Gejala diabetes: poliuria, polidipsia, kehilangan berat badan
Sumber : American Diabetes Association (2011).
Tabel 2. Klasifikasi Etiologis Diabetes Melitus
I. Diabetes melitus tipe 1
(destruksi sel beta, umumnya mengarah pada defisiensi insulin absolut) A. melalui proses imun
B. Idiopatik
II. Diabetes melitus tipe 2
(bervariasi mulai yang predominan resistensi insulin disertai defisiensi insulin relatif sampai yang predominan gangguan sekresi insulin bersama resistensi insulin) III. Diabetes melitus tipe lain
A. Defek genetik fungsi sel beta:
Kromosom 12, HNF-1α (MODY3); Kromosom 7, glukokinase (MODY2); Kromosom 20, HNF-4α (MODY1); Kromosom 13, insulin promoter faktor-1 (IPF-faktor-1; MODY4); Kromosom faktor-17, HNF-faktor-1β (MODY5); Kromosom 2,
NeuroD1 (MODY6); DNA mitokondria; lainnya
B. Defek genetik kerja insulin
Insulin resisten tipe A, Leprechaunism, Sindrom Rabson-Mendenhall, Diabetes lipoatropik, lainnya
C. Penyakit eksokrin pankreas
Pankreatitis, Trauma/prankreatektomi, Neoplasia, Cystic fibrosis, Hemochromatosis, Pankreatopati fibrokalkulus, lainnya
D. Endokrinopati
Akromegali, Sindrom Cushing, Glukagonoma, Stomatostatinoma, Feokromositoma, Hipertiroidism, Aldosteronoma, lainnya
E. Karena obat/zat kimia: vacor, pentamidin, asam nikotinat, tiazid, glukokortikoid, hormon tiroid, interferon γ, dilantin, agonis β-adrenergik, diazosida, lainnya F. Infeksi : Rubella kongenital, cytomegalovirus, lainnya
G. Imunologi: sindrom “stiff-man” dan antibodi anti reseptor insulin
H. Sindrom genetik lain: sindrom Down, sindrom Klinefelter, sindrom Turner, sindrom Wolfram, atasia Friedreich, Huntington’s chorea, sindrom Lawrence-Moon-Btedl, Myotonic dystrophy, porphyria, sindrom Pruder-Willi, lainnya IV. Diabetes melitus Kehamilan
Sumber : American Diabetes Association (2011).
Individu yang menderita diabetes tipe 1 sangat membutuhkan insulin dari luar tubuh selama hidupnya guna menghindarkan ketoasidosis. Hal itu terjadi karena individu tersebut mengalami defisiensi insulin absolut yang diakibatkan oleh proses autoimun terhadap sel β pankreasnya. Kerusakan ini terjadi karena adanya suatu stimulus dari luar atau determinan genetik yang memungkinkan sel
β pankreas dikenali sebagai benda asing. Pulau Langerhans selanjutnya diinfiltrasi oleh sel T teraktivasi yang mengarah pada pengrusakan sel β pankreas dan akhirnya menjadi insulitis (Champe dan Harvey, 1994).
dan peningkatan sintesis 3-hidroksibutirat dan asetoasetat oleh hati. Perubahan metabolik ini terjadi sebagai akibat defisiensi insulin dan berlebihnya glukagon dalam darah (Brody, 1999).
Diabetes tipe 2 merupakan bentuk diabetes yang paling banyak diderita oleh masyarakat dunia, termasuk Indonesia. Empat ciri gangguan metabolik pada orang yang menderita diabetes melitus tipe 2 antara lain: kegemukan, kegagalan kerja insulin, disfungsi sekresi insulin dan peningkatan pembentukan glukosa endogen (Weyer et al, 1999).
Etiologi terjadinya diabetes melitus tipe 2 berawal dari faktor gen dan lingkungan (gaya hidup serta diet). Faktor tersebut dapat mengakibatkan adanya resistensi insulin. Dalam kondisi fungsi sel β pankreas normal, hiperinsulinemia akibat resistensi insulin dapat dikompensasi hingga kadar glukosa darah tetap normal. Tetapi bila fungsi sel β pankreas tidak normal, sekresi insulin akan menurun lalu terjadi hiperglikemia dan berlanjut menjadi diabetes melitus tipe 2 (Waspadji, 2003).
Penyandang diabetes melitus tipe 2 seringkali terkait dengan resistensi insulin, yaitu suatu keadaan di mana kandungan insulinnya cukup banyak namun efek fisiologisnya rendah (Lebovitz, 1999). Resistensi insulin dapat terjadi karena berbagai hal seperti kegagalan dalam transduksi signal, kisaran insulin yang abnormal, atau reseptor insulin yang gagal memberikan signal untuk mentranslokasikan transporter glukosa dari sitoplasma ke membran sel (Champe dan Harvey, 1994).
Penyandang diabetes melitus tipe 2 kebanyakan menderita kegemukan. Kegemukan dapat diakibatkan oleh dua faktor, yaitu faktor gen dan lingkungan. Faktor gen thrifty dapat mengakibatkan peningkatan penyimpanan lemak dan
menurunkan ambilan glukosa, sedangkan penyebab kegemukan akibat faktor lingkungan dapat berupa gaya hidup santai dan makan makanan dalam jumlah dan frekuensi yang banyak.
berlangsung lama, sel akan mengalami kekurangan glukosa sebagai sumber energinya. Guna memenuhi zat tersebut, sel melakukan glukoneogenesis. Glukoneogenesis pada jaringan lemak selain menghasilkan glukosa juga asam lemak. Bila keadaan ini terus berlanjut mengakibatkan kandungan asam lemak bebas dalam darah meningkat. Tingginya asam lemak bebas dalam darah dapat mengakibatkan penurunan ambilan glukosa oleh otot, meningkatkan produksi glukosa pada hati, dan dapat menghambat sekresi insulin oleh sel β pankreas. Penghambatan sekresi insulin oleh asam lemak bebas pada awalnya dapat dikompensasi, namun bila terus berlanjut sel β pankreas juga akan mengalami gangguan sekresi insulin. Sementara itu tingginya produksi glukosa hati dan rendahnya ambilan glukosa oleh otot akan mengakibatkan resistensi insulin. Keadaan ini bila berlangsung lama dan seiring rendahnya sekresi insulin menimbulkan kejadian hiperglikemik (Proietto et al., 1999; LeRoith dan Zick,
2001; Cefalu, 2001). Efek kegemukan terhadap patogenesis diabetes tipe 2 dapat dilihat pada Gambar 1.
Faktor genetik / lingkungan
Konsumsi kalori berlebih
Hiperinsulinemia
Peningkatan Asam Lemak Bebas & Trigliserida, serta penumpukan lemak
Sel Lemak Otot Hati Sel β
Resistensi insulin Kompensasi sel β
Resistensi insulin Perubahan ekspresi & makin meningkat pensignalan gen
Diabetes tipe 2 Penurunan sekresi insulin
2.2 Stres Oksidatif
Radikal bebas adalah atom atau molekul yang elektron terluarnya tidak berpasangan, hingga menjadikannya tidak stabil dan sangat reaktif. Pada kondisi tubuh normal, radikal bebas diproduksi untuk membunuh mikroorganisme dan menghancurkan sel kanker (Nadler and Natarajan, 2000). Dalam tubuh, radikal bebas dibentuk di membran sel dan organel seperti mitokondria, peroksisom, retikulum endoplasmik dan sitosol melalui reaksi-reaksi enzimatik fisiologis dalam proses metabolisme (Madhavi et al., 1996).
Radikal bebas dapat berkaitan pada atom oksigen dan nitrogen. Radikal bebas yang berkaitan dengan atom oksigen antara lain anion superoksida, hidrogen peroksida, hidroksil radikal, peroksil, dan alkoksil, sedang yang berkaitan dengan atom nitrogen adalah nitrit oksida, nitrogen dioksida, dan peroksinitrit (Fang et al., 2002).
Anion superoksida adalah bentuk tereduksi molekul oksigen yang dibentuk melalui penerimaan satu elektron. Anion superoksida adalah radikal bebas pertama kali yang dibentuk dari sistem pemindahan elektron pada mitokondria. Hidrogen peroksida dibentuk melalui reaksi dismutasi anion superoksida oleh superoksida dismutase. Radikal ini bersifat sangat mudah berdifusi, namun merupakan radikal bebas paling tidak reaktif. Hidroksil radikal adalah radikal bebas yang paling reaktif. Radikal ini dibentuk melalui pemecahan ikatan kovalen hidrogen dan oksigen saat radiasi sel, homolytic fission, reaksi
Fenton dan reaksi Haber Weiss. Radikal peroksil dan alkoksil dibentuk melalui dekomposisi alkil peroksida, iradiasi sinar ultra violet, dan homolisis peroksida.
Nitrit oksida adalah radikal bebas dengan elektron tidak berpasangan tunggal. Radikal ini dibentuk dari L-arginin oleh NO sintase dan bukan termasuk radikal bebas reaktif. Nitrogen dioksida dibentuk dari reaksi radikal peroksil dan nitrit oksida dan merupakan radikal bebas yang mampu memulai peroksidasi lemak. Peroksinitrit adalah radikal bebas yang dibentuk dari reaksi nitrit oksida dan anion superoksida. Radikal ini berkemampuan merusak jaringan dan mengoksidasi LDL (Lee et al., 2004).
sedang radikal nitrogen berperan dalam proses relaksasi dan proliferasi sel otot halus, adesi sel darah putih, agregasi platelet, angiogenesis, trombosis, tekanan pembuluh darah, hemodinamis, serta pada syaraf berperan sebagai neurotransmiter (Halliwell and Gutteridge, 1999, Droge, 2002).
Radikal bebas juga dapat bertindak sebagai oksidan terhadap enzim yang mengandung logam, radikal bebas, dan spesies reaktif lainnya yang dapat mengakibatkan oksidasi pada biomolekul hingga mengarah pada kerusakan sel, penyakit degeneratif, hingga kematian (Fridovich, 1999; McCord, 2000).
Stres oksidatif adalah suatu keadaan di mana kandungan oksidan atau radikal bebas dalam tubuh lebih banyak dibanding antioksidannya (Bonnefont-Rousselot et al, 2000). Menurut Jakus (2000), Bonnefont-Rousselot et al. (2000),
Maritim et al. (2003), Martin-Gallan et al, (2003), dan DeMattia et al. (2003) stres
oksidatif dapat dialami penyandang diabetes akibat autooksidasi glukosa, induksi dan aktivasi enzim-enzim lipoksigenase, aktivasi glikasi, dan penurunan aktivitas antioksidan enzim.
Saat hiperglikemik, glukosa yang berbentuk enediol mudah mengalami autooksidasi dalam suatu reaksi transisi yang diperantarai logam menjadi anion radikal enediol. Radikal ini selanjutnya diubah menjadi ketoaldehid yang dapat menghasilkan anion superoksida. Radikal ini selanjutnya berubah menjadi hidrogen peroksida dan akhirnya menjadi radikal hidroksil. Radikal hidroksil yang dihasilkan melalui autooksidasi glukosa secara spesifik menunjukkan pengrusakan terhadap protein.
Kondisi hiperglikemik dapat meningkatkan aktivitas dan ekspresi enzim-enzim lipoksigenase. Enzim lipoksigenase dapat mengoksidasi LDL dengan mekanisme pengoksidasian asam arakidonat dan linoleat. Produk peroksidasi asam arakidonat disebut isoprostan. Isoprostan jenis 8-epi-prostaglandin (PG) F2α
akan membentuk produk Amadori. Produk ini selanjutnya terdegradasi dengan kehilangan RNH2
Apoptosis adalah suatu proses kematian sel secara fisiologis yang dilakukan melalui suatu mekanisme rangkaian sel yang terprogram secara genetik. Apoptosis dapat dipicu oleh stres oksidatif, di samping karena adanya kerusakan DNA dan tiadanya faktor-faktor untuk kelangsungan hidup. Sel-sel yang mengalami apoptosis menunjukkan bahwa sitoplasmanya mengalami stres oksidatif. Sel yang mengalami apoptosis tidak menimbulkan kerusakan pada sel di
menjadi α-dikarbonil (deoksiglukoson). Senyawa ini lebih reaktif dibanding gula asal dalam kemampuannya bereaksi dengan asam amino dan interaksi ini membentuk produk akhir yang disebut advanced Maillard products atau advanced glycation end products (AGEs). AGEs ini dapat
membentuk anion superoksida (Yim et al., 1995; Meng et al., 1998).
Pembentukan radikal bebas tubuh yang berlebih saat diabetes melitus dapat memicu penurunan kadar antioksidan enzimatik tubuh serta mengubah keseimbangan oksidan dan antioksidan tubuh. Stres oksidatif pada diabetes melitus terjadi akibat pembentukan spesies oksigen reaktif secara berlebih dan penurunan antioksidan enzimatik secara drastis (Jakus, 2000, Evans et al. 2002).
Mekanisme pertahanan antioksidan enzim aktivitasnya akan mengalami penurunan saat tubuh mengalami hiperglikemia. Antioksidan ini tidak dapat bereaksi secara berantai saat hiperglikemia, hingga aktivitasnyapun menurun. Penurunan aktivitas antioksidan ini dapat juga diakibatkan oleh terhambatnya pertumbuhan sel endotelium oleh kondisi hiperglikemik. Sel endotelium yang rusak tidak dapat menghasilkan antioksidan enzim secara normal (Kashiwagi et al. 1999).
Stres oksidatif yang berlebihan akan menimbulkan kematian sel. Mekanismenya dapat melalui nekrosis atau apoptosis. Nekrosis adalah proses kematian sel atau jaringan yang terjadi secara tiba-tiba atau setelah terjadi pajanan radikal bebas pada sel atau jaringan tersebut. Nekrosis dapat mengakibatkan sel mengalami kehilangan intergritas mitokondria, membran plasma, membran peroksisom, dan lisosom, pembengkakan sel dan rupture, kerusakan dan lisis sel
sekitarnya, karena sel tersebut tidak melakukan proses penglepasan intraselular ke ekstraselular (Sesikeran et al., 2002).
2.2.1 Stres Oksidatif dan Kerusakan Sel
Stres oksidatif menurut Halliwell dan Gutteridge (1999) dapat merusak sel dengan cara mengganggu metabolisme kalsium, fragmentasi DNA, merusak bentuk dan keutuhan sel. Kandungan ion kalsium intraselular dipertahankan sangat rendah oleh enzim Ca2+ ATPase dan ion channel Ca2+. Kandungan ini akan meningkat secara perlahan untuk menghasilkan sinyal metabolik sebagai respons terhadap beberapa hormon. Radikal bebas dapat mengganggu atau merusak aktivitas enzim Ca2+ ATPase yang mengandung gugus sulfhidril (-SH). Enzim ini di dalam sel berfungsi sebagai pengendali konsentrasi ion Ca2+ dalam sel. Dengan adanya radikal bebas, aktivitas enzim ini menjadi inaktif hingga pengendalian konsentrasi Ca2+ terganggu. Ion Ca2+ yang berada di dalam mitokondria dan retikulum endoplasmik akan mengalami efluks ke dalam sitosol namun ion ini tidak bisa keluar dari sel. Akibatnya kadar Ca2+ sitosol meningkat dan dapat
meningkatkan aktivitas enzim-enzim yang tergantung pada konsentrasi Ca2+ yaitu:
protease, fosfolipase dan endonuklease.
Ion channel Ca2+ adalah suatu protein yang berfungsi menjaga kadar Ca2+
dalam intraselular tetap rendah. Saat sel mengalami stres oksidatif, komponen protein channel ini menjadi berubah atau rusak hingga kadar Ca2+ dalam sel dapat
menjadi meningkat. Tingginya kadar Ca2+ dalam sel dapat mengaktifkan enzim-enzim yang tergantung pada Ca2+
Kerusakan sel dapat juga diakibatkan oleh terfragmentasinya DNA sel. Hal ini terjadi karena rantai utama DNA dipotong endonuklease yang teraktivitasi oleh tingginya kadar Ca
seperti fosfolipase, endonuklease, dan protease. Teraktivasinya enzim tersebut dapat merusak integritas sitoplasma sehingga sel akan mengalami kematian.
2+. Fragmentasi juga dapat diakibatkan oleh reaksi antara
rusak, enzim ini akan menggunakan NAD+ sebagai substrat dalam jumlah besar sehingga metabolisme sel terganggu dan bahkan terhenti hingga sel menjadi mati.
Sitoskeleton, molekul adesi, dan penghubung antar sel merupakan komponen protein yang memiliki gugus thiol (-SH). Komponen ini dalam sel berfungsi sebagai penentu bentuk dan keutuhan sel. Apabila gugus ini teroksidasi oleh radikal bebas maka akan menimbulkan kerusakan dan lepasnya sel endotelium dari sel tetangganya atau dari matriks ekstraselular. Matriks ekstraselular sel terdiri dari komponen protein yang berupa serabut dan cairan
viscous. Komponen ini berinteraksi dengan sel endotelium untuk mengatur
perlekatan, migrasi, dan proliferasi sel endotelium. Apabila komponen ini teroksidasi oleh radikal bebas berakibat sel mengalami kerusakan dan terjadi penglepasan sel endotelium. Sedangkan membran basalis sebagai tempat perlekatan sel endotelium yang terdiri dari protein serabut kolagen dengan adanya radikal bebas akan mengalami kekakuan hingga sel endotelium pun terlepas.
Oksidasi radikal bebas terhadap gugus tiol juga dapat mengakibatkan
blebbing (pelepuhan) yaitu perubahan bentuk sel endotelium menjadi lebih bulat.
Jika proses pelepuhan lebih parah, tonjolan sel akan mengalami rupture dan
terbentuk lubang pada membran sehingga sel akan mati. Selain akibat oksidasi terhadap gugus tiol pelepuhan juga dapat diakibatkan oleh enzim protease yang teraktivasi oleh tingginya kadar Ca2+
Radikal bebas yang bersifat reaktif dapat menarik atom hidrogen asam lemak tidak jenuh rantai panjang dari membran sel. Senyawa karbon yang atom hidrogennya dikurangi memiliki elektron yang tidak berpasangan dan menjadi
dalam sitoplasma.
2.2.2 Stres Oksidatif dan Membran Sel
radikal bebas. Radikal hidroksil merupakan salah satu radikal bebas yang dapat mengawali peroksidasi lemak.
C-H + OH* C* + H2O
Ketika radikal C* terbentuk pada bagian hidrofob membran sel, akan memudahkan terjadi reaksi antara radikal ini dengan O2
Lipid* + O
yang terlarut dalam membran.
2 Lipid-O2
Lipid-O
* (radikal peroksil)
Radikal peroksil yang terbentuk ini bersifat sangat reaktif dan dapat mengoksidasi rantai asam lemak panjang tidak jenuh lainnya yang berdekatan.
2* + Lipid-H Lipid-O2
Mekanisme penghambatan aktivitas radikal bebas oleh antioksidan menurut Widodo et al. (1995) dapat berupa: 1. bekerja sebagai pengganti enzim
antioksidan yang telah habis, 2. bekerja dengan mengintervensi reaksi radikal bebas, dan 3. mematahkan reaksi rantai pada reaksi radikal bebas. Sementara itu
H + Lipid*
Reaksi-reaksi di atas akan berulang kembali dan keseluruhan proses tersebut bersambung dalam reaksi berantai radikal bebas hingga sel menjadi rusak dan mati.
Protein juga merupakan komponen penyusun membran sel. Komponen ini berada di bagian dalam dan luar membran. Radikal bebas dapat merusak komponen ini melalui reaksinya dengan asam amino penyusun protein yang mengandung gugus sulfhidril (-SH). Pembentukan ikatan disulfida akibat reaksi ini akan menimbulkan ikatan antar molekul protein secara silang hingga protein mengalami agregasi atau depolarisasi. Akibatnya protein menjadi kaku dan kehilangan fungsi sebagai kanal ion maupun pompa ion. Hilangnya fungsi ini memudahkan sel menjadi rusak dan mati.
2.2.3 Sistem Proteksi terhadap Radikal Bebas
Terbentuknya radikal bebas tidak akan menyebabkan kerusakan atau mengganggu proses fisiologis tubuh apabila sistem proteksi enzimatik dalam sel (Free radical scavenger) dan sistem proteksi non-enzimatik (antioksidan) cukup
Lee et al. (2004) menjelaskan bahwa antioksidan mampu mencegah kereaktifan
radikal bebas melalui tiga mekanisme: 1. pendonoran hidrogen, 2. pengkelasian logam, dan 3. quencher oksigen singlet. Antioksidan-antioksidan pendonor
hidrogen berperan menurunkan kereaktifan radikal bebas dengan memberikan hidrogennya pada radikal bebas. Antioksidan jenis pengkelat logam dapat mencegah pemercepatan tahap pemulaan oksidasi lemak, pembentukan oksigen tunggal, radikal alkoksil, radikal peroksil, dan radikal hidroksil dengan pembentukan ion kompleks dan senyawa koordinasi dengan logam. Antioksidan
quencher oksigen singlet berkemampuan mencegah pembentukan produk
teroksidasi melalui perubahan oksigen singlet menjadi oksigen triplet dengan pemindahan energi atau muatan tanpa pembentukan produk antara atau terlibat dalam pembentukan produk antara.
Enzim-enzim antioksidan seperti Superoxide dismutase (SOD), Glutathione peroxidase (GSH-Px) dan Catalase (CAT) berperan sebagai pertahanan antar sel dalam tubuh. Enzim-enzim ini mampu menurunkan kereaktifan radikal bebas melalui pendekomposisian spesies oksigen reaktif. SOD mengubah radikal superoksida menjadi hidrogen peroksida, GSH-Px mereduksi hidrogen peroksida menjadi air dan katalase mendetoksifikasi hidrogen peroksida menjadi air dan oksigen (Jakus, 2000, Nadler and Natarajan, 2000, Nedeljkovic et al., 2003). Secara ringkas sistem detoksifikasi radikal bebas oleh enzim
antioksidan dapat dilihat pada Gambar 2.
e- O
2- O2 O2- O2 H+
O2 O2- H2O2 .OH H2
SOD
H O
2O + O
2H
2 CAT
2
Gambar 2. Sistem detoksifikasi radikal bebas oleh enzim antioksidan O GSH-Px
sebagai antioksidan dengan cara menyumbangkan atom hidrogennya ke radikal bebas dan quenching oksigen singlet. Vitamin E dapat bertindak sebagai
antioksidan melalui pendonoran atom hidrogen dari gugus hidroksil pada cincin kromanol dan scavenger radikal bebas. Karotenoid bertindak sebagai antioksidan
karena kemampuannya sebagai quencher terhadap radikal bebas. Sementara itu
polifenol dapat bersifat sebagai antioksidan karena kemampuannya mendonorkan atom hidrogen, scavenger radikal bebas, dan pengkelat ion logam (Desphande et al., 1996; Lee et al., 2004).
Polifenol dapat bersifat sebagai antioksidan dijelaskan oleh Rice-Evans et al. (1997) karena senyawa ini mempunyai sifat pereduksi yakni agen pendonor
atau penyumbang hidrogen. Lebih lanjut ditegaskan bahwa aktivitas antioksidannya sangat ditentukan oleh: reaktivitasnya sebagai agen pendonor hidrogen (kaitannya dengan potensial reduksi), reaktivitasnya dengan antioksidan yang lain, potensial transisi pengkelat logam, dan kemampuannya untuk menstabilisasi dan mendelokalisasi elektron tak berpasangan.
Polifenol mempunyai struktur kimia yang ideal dalam kaitannya sebagai
scavenger radikal. Hal ini dapat dilihat dari uji interaksi, laju konstanta reaksi, dan
stabilitas radikal polifenol dengan radikal hidroksil (OH*), azida (N3*), anion
superoksida (*O2
-Sel endotelium adalah selapis sel epitel yang berbentuk poligonal dan berasal dari mesoderm. Sel ini terletak di bagian intima pembuluh darah dan melekat pada membran basalis. Sel endotelium mempunyai sebuah inti dengan panjang 5-25 µm dan tebal 3 µm. Sel ini memanjang seiring dengan aliran darah. Pada hubungan antar sel endotelium terdapat bagian yang overlapping untuk
membantu perlekatan pada pembuluh darah. Pada kondisi fisiologis sel ini merupakan pembatas intima dan media pembuluh darah dari pengaruh fisik ), dan lipid peroksil (LOO*) dibanding vitamin E dan C. Potensi polifenol sebagai antioksidan dapat juga diamati dari kecenderungan senyawa ini untuk mengkelat logam terutama besi dan tembaga, sehingga dapat menghambat pembentukan radikal bebas yang dikatalis oleh logam (Rice-Evans et al., 1997).
2.3 Sel Endotelium
komponen darah. Pada orang dewasa dengan berat badan 70 kg, sel ini meliputi area seluas 700 m2 dengan berat 11,5 kg (Haller, 1997).
Sel endotelium merupakan bagian yang sangat penting tidak saja sebagai sistem pembatas namun juga sebagai tempat beberapa reseptor dan angiogenesis. Sel ini juga berfungsi menghasilkan berbagai mediator yang berperan terhadap viskositas, kontraktilitas pembuluh darah, proliferasi sel otot polos dan interaksi elemen darah dengan dinding pembuluh darah. Sel endotelium yang sehat akan bersifat anti adesi dan anti trombosis serta berperan terhadap mengalirnya (fluiditas) aliran darah (Nystrom, 2005).
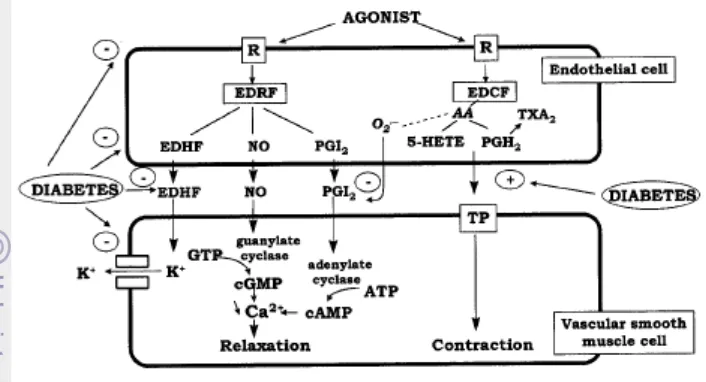
Fungsi sel endotelium tersebut dapat berlangsung karena adanya substansi yang disekresikan oleh sel endotelium ke darah dan otot halus. Substansi itu ada dua golongan besar yaitu Endothelium Derived Relaxing Factors (EDRF) yang
terdiri atas NO (nitrit oksida), prostasiklin (PGI2), dan faktor relaksasi
hiperpolarisasi, dan Endothelium Derived Contracting Factors (EDCF) yang
terdiri atas endotelin-I (ET-I), tromboksan A, prostaglandin, dan angiotensin II (Haller 1997).
Interaksi sel endotelium dan darah tidak hanya melibatkan interaksi komponen darah dengan sel, namun juga berperan terhadap aliran darah. Hal ini menjadikan sel endotelium bertanggung jawab terhadap pengaturan secara akut dan adaptasi secara kronis pembuluh darah. Pengaturan pembuluh darah secara akut sel endotelium ditandai dengan dihasilkannya faktor-faktor vasodilator, seperti nitrit oksida (NO), endothelium derived hyperpolarization factor (EDHF)
dan prostaglandin (PGI2/PGE2
NO dihasilkan melalui pengubahan L-arginin oleh NOS menjadi L-sitrulin dengan keberadaan oksigen dan kofaktor. Kofaktor yang terlibat dalam reaksi
). Sel ini juga mampu menginduksi vasokonstriktor yaitu endothelin-1 (ET-1) (deVriese et al. 2000).
tersebut antara lain: calmodulin, tetrahidrobiopterin (BH4), NADPH tereduksi, heme, FAD, dan FMN. Peningkatan konsentrasi Ca2+, fosforilasi Ser1177/1179 dari fosfatidilinositol-3 (PI-3) kinase dan Ser/Thr protein kinase Akt dapat mengaktifkan NOS. Produksi NO sangat tergantung pada ketersediaan L-arginin. L-Arginin di dalam sel dihasilkan dari L-sitrulin melalui aktivitas arginosuksinat sintase dan arginosuksinat liase (deVriese et al. 2000).
NO dilepaskan dari sel endotelium dalam merespons keberadaan senyawa kimia (asetilkolin [ACh], bradikinin [BK] atau ionofor Ca2+) dan tekanan fisik (tegangan geser, aliran darah) yang menghasilan vasodilatasi, menurunkan tekanan pembuluh darah, tekanan darah, menghambat agregasi dan adesi platelet, menghambat adesi dan migrasi leukosit, dan menurunkan proliferasi otot polos hingga pada akhirnya mencegah aterosklerosis. Fungsi NO ini dimediasi oleh
cyclic-Guanylate Monophosophate (cGMP) yang disintesis soluble guanylyl cyclase, yaitu suatu enzim yang mengandung heme (deVriese et al. 2000).
Beberapa penelitian menunjukkan bahwa polifenol pada anggur, teh hijau dan hitam, cokelat, kedelai, dan buah delima dapat meningkatkan modulasi eNOS untuk menghasilkan NO. Dalam penelitian tersebut diungkapkan bahwa polifenol dapat meningkatkan ketersediaan NO melalui peningkatan ekspresi eNOS, peningkatan ikatan CaM terhadap eNOS, peningkatan konsentrasi Ca2+ yang mendorong pada fosforilasi eNOS pada Ser1177
Disfungsi endotelium adalah suatu keadaan di mana produksi faktor vasodilator tidak seimbang dengan faktor vasokontriksi. Faktor vasodilator antara lain nitrit oksida, prostasiklin dan endothelial derived hyperpolarizing factor
melalui lintassan fosfatidilinositol-3-kinase (PI-3K)/Akt, dan menurunkan kadar anion superoksida.
2.3.2 Disfungsi Sel Endotelium
Sel endotelium pada pembuluh darah mudah mengalami stres hemodinamis yaitu tekanan akibat aliran, tekanan, dan viskositas darah. Apabila kondisi hemodinamis berubah, seperti hiperglikemia, maka fungsi sel endotelium sebagai pengendali perlintasan larutan, makromolekul ataupun sel darah ke dalam pembuluh darah akan berubah pula. Stres hemodinamis yang ringan dan sesaat masih memungkinkan sel endotelium tidak kehilangan fungsi utamanya melalui reendotelisasi, namun bila berlangsung lama dan berat sel endotelium akan kehilangan fungsinya (Drenckhahn and Ness, 1997).
Sel endotelium dapat menghasilkan granul membran protein 140 kDA (GMP-140) yang dapat mengikat neutrofil dan monosit, sehingga bila sel endotelium teraktivasi dengan cepat terjadi translokasi neutrofil dan monosit ke dalam membran. Setelah monosit menempel pada endotelium, segera monosit melanjutkan migrasinya ke lapisan intima. Proses migrasi monosit sangat berkaitan dengan adanya monocyte chemoatractant protein-1 (MCP-1) yang
(EDHF) dan faktor vasokontriksi adalah endotel-1, angiotensin II dan prostaglandin. Ketidak-seimbangan kedua faktor ini akan mengarah pada kondisi pro-aterogenesis (Eckel et al., 2002, Suryadipraja, 2003, Fonseca et al., 2004,
Nystrom, 2005).
Disfungsi sel endotelium dapat disebabkan oleh kerusakan morfologi atau struktur sebagai akibat terjadinya disintegrasi sel dan gangguan fungsi walau sel tidak mengalami disintegrasi. Kerusakan struktur dapat terjadi akibat aktivitas beberapa enzim proteolitik yang menyebabkan pecahnya matriks molekul adesi dan akibatnya sel endotelium menjadi terlepas. Terjadinya disfungsi endotelium merupakan awal pembentukan plak ateroma yang ditandai dengan meningkatnya adesi monosit pada endotelium arteri yang dipicu oleh intracellular adhesion molecul-1 (ICAM-1) yang juga akan menarik neutrofil dan monosit. Sel
endotelium saat kondisi hiperglikemia atau diabetes dapat rusak karena adanya faktor-faktor sirkulasi seperti tingginya asam lemak bebas, lipoprotein, turunan glikasi dan oksidasi (Eckel et al., 2002, Nystrom, 2005).
Sel endotelium pada kondisi hiperglikemik akan mengalami disfungsi akibat peningkatan permeabilitas dan penurunan vasorelaksan. Kondisi hiperglikemik dapat memicu pengaktifan protein kinase C (PKC) yang berada dalam sitoplasma sel endotelium. Teraktivasinya enzim ini akan mengaktifkan interaksi aktin dan miosin hingga terjadi permeabilitas atau kontraksi sel (Haller, 1997). Sowers and Lester (1999) menjelaskan bahwa saat hiperglikemia produksi vasorelaksan menurun hingga sel endotelium mengalami kontraksi.
Ada beberapa mekanisme penyebab disfungsi sel endotelium pada diabetes melitus, yaitu: penurunan produksi satu di antara EDRF, peningkatan inaktivasi EDRF, kegagalan difusi EDRF ke sel otot polos, penurunan respons otot polos terhadap EDRF dan peningkatan endhothelium-derived contracting factor (EDCF). Mekanisme disfungsi sel endotelium pada kondisi diabetes
Gambar 3. Mekanisme disfungsi sel endotelium akibat diabetes (deVriese et al. 2000)
Gambar 3 menunjukkan keadaan hiperglikemia dapat mengakibatkan keadaan disfungsi sel endotelium. Ada enam faktor yang menyebabkan kondisi ini muncul saat diabetes melitus. Pertama, hiperglikemik mengakibatkan perubahan konformasi membran sel endotelium dan reseptor, sehingga sel endotelium kurang sensitif terhadap keberadaan agonis. Kedua, hiperglikemik menurunkan produksi EDRF dan meningkatkan produksi EDCF yang berakibat pembuluh darah dominan mengalami kontraksi. Ketiga, hiperglikemik menghambat difusi EDHF sehingga efflux K+ terhambat dan konsentrasi Ca2+ menjadi tinggi yang berakibat
pembuluh darah menjadi tegang. Keempat, hiperglikemik menghambat kerja ion K+ channel yang mengakibatkan konsentrasi K+ dalam sitosol otot polos tetap tinggi dan berakibat otot polos tegang. Kelima, anion superoksida yang dihasilkan kondisi hiperglikemia berinteraksi dengan PGI2
Disfungsi sel endotelium dapat juga diakibatkan oleh degradasi NO dan stres oksidatif. NO yang disintesis melalui NOS atau dari donor NO secara non enzimatis akan diinaktivasi secara cepat melalui oksidasi menjadi nitrit atau nitrat. Ketidakcukupan BH
dan berakibat bioaktivitasnya untuk vasorelaksasi menurun. Keenam, hiperglikemik meningkatkan
vasoconstriction substances, seperti turunan prostaglandin yang berakibat
pembuluh darah mengalami konstraksi (deVriese et al. 2000).
4 juga berperan pada terjadinya disfungsi ini, karena aktivitas
menggantikan NO. Anion superoksida dapat bereaksi dengan NO membentuk peroksinitrit, yaitu salah satu spesies oksigen reaktif yang sangat toksik. Radikal ini dapat secara efektif dibersihkan oleh SOD dan katalase.
2.4 Florotanin
Polifenol adalah salah satu fitokimia yang banyak tersebar di alam ini dan ada sekitar 8000 strukturnya yang telah diketahui. Mulai dari yang berupa fenol sederhana hingga tanin yang berberat molekul lebih dari 30 kDa. Senyawa ini bila dikonsumsi dapat mencegah penyakit akibat radikal bebas. Senyawa ini mampu menurunkan kejadian penyakit tersebut karena sifatnya yang mudah mendonorkan atom hidrogennya pada radikal bebas (King, 1999; Chung et al., 1998; Lee et al.,
2004).
Tanin adalah salah satu polifenol yang banyak terkandung dalam tanaman. Senyawa ini terdiri dari tanin terkondensasi, tanin terhidrolisis dan florotanin. Tanin terkondensasi disusun oleh oligomer dan polimer flavonols, sedang tanin hidrolisat tersusun oleh asam galat dan elagat. Tanin terhidrolisis dalam pangan manusia hampir dapat dikatakan jarang dijumpai dibanding tanin terkondensasi (Cheynier, 1999; Chung et al. 1998, Quideau et al., 2011).
OH
Gambar 4. Rumus bangun floroglusinol dan turunannya
Florotanin dalam rumput laut coklat terdapat dalam organel sel yang disebut physodes. Organel ini terdapat dalam sitoplasma dan berbentuk lingkaran
atau elips. Organel ini mudah berpindah-pindah dan mampu memantulkan cahaya. Saat physodes bergerak dan berfusi ke dalam membran sel, florotanin akan
disekresikan ke membran sel. Florotanin yang tersekresi itu selanjutnya akan membentuk ikatan kompleks dengan asam alginat. Asam alginat adalah salah satu komponen utama membran sel rumput laut coklat selain alginat dan fukan (Ogino C, 1962; Shibata et al. 2004; Koivikko et al., 2005).
Florotanin merupakan senyawa yang berwarna coklat kekuning-kuningan. Senyawa ini akan terlihat coklat gelap bila ditempatkan di udara bebas dan memberikan warna merah bila direaksikan dengan asam klorida. Senyawa ini bersifat higroskopis, pereduksi kuat dan mudah larut dalam pelarut polar (Ogino C, 1962; Pavia et al., 2002). Kelarutan florotanin dalam berbagai pelarut dapat
dilihat pada Tabel 3.
Tabel 3. Kelarutan florotanin dalam berbagai pelarut
No. Pelarut Polaritas (P’) Kelarutan
Beberapa peneliti telah berhasil mengekstrak dan mengisolasi florotanin dari beberapa spesies rumput laut coklat. Nagayama et al. (2002) telah
mengekstraksi florotanin dari Eisenia bicyclis. Tepung rumput laut coklat ini
kecepatan 90 rpm selama 48 jam. Selanjutnya disaring dan dipekatkan dengan rotavapor. Ekstrak pekat selanjutnya ditambah metanol 240 mL, kloroform 480 mL, dan air bebas ion 180 mL. Setelah penambahan pelarut polar dan non-polar tersebut, lapisan bagian atasnya dipisahkan dan diekstrak dengan etil asetat 300 mL 2 kali. Ekstrak ini selanjutnya dipekatkan dengan rotavapor dan dinyatakan sebagai florotanin kasar.
Sementara itu Wei et al. (2003) telah mendapatkan florotanin berberat
molekul besar dari S. kjelmanianum. Rumput laut ini yang telah ditepungkan
diekstrak dengan etanol 85% (1:2; b/v) sambil digoyang selama semalam di dalam ruang gelap. Selanjutnya disaring untuk mendapatkan larutan coklat gelap. Larutan selanjutnya dipekatkan dengan rotavapor dan pekatan yang dihasilkan dicuci kloroform 3 kali dan dilanjutkan dengan eter 3 kali. Lapisan aqueous didialisis dengan akuabides selama semalam dalam ruang gelap bersuhu 4oC. Selama dialisis akuabides diganti 3 kali. Dialisat selanjutnya dipekatkan dengan rotavapor. Dialisat pekat selanjutnya dielusi dalam kromatografi kolom dengan toluen, aseton, dan aseton-metanol (1:2; v/v). Eluat aseton-metanol dinyatakan sebagai florotanin berberat molekul besar.
Okada et al. (2004) telah mengisolasi turunan florotanin yaitu eckol dan
dieckol dari Eiseniakurome. Rumput laut coklat ini sebelumnya direfluks metanol
(1:18;b/v) selama 3 jam dan 3 kali. Setelah metanol diuapkan, ekstrak selanjutnya difraksinasi dalam kolom kromatografi dengan eluen air, metanol, aseton, dan etil asetat. Fraksi metanol difraksinasi lagi dengan metanol aquoeus, metanol, dan aseton untuk menghasilkan fraksi 1-10. Fraksi 4 selanjutnya difraksinasi dengan metanol untuk mendapatkan fraksi 11-14. Fraksi 13 selanjutnya dielusi dengan n-butanol-n-propanol-H2
Berdasarkan pada strukturnya florotanin dapat berfungsi sebagai antioksidan. Beberapa penelitian menunjukkan senyawa ini mampu menunjukkan aktivitasnya sebagai antioksidan. Yan et al. (1998) mendapati bahwa aktivitas scavenger florotanin yang didapat dari S. kjelmanianum lebih kuat dibanding
vitamin E. Sementara itu Lim et al. (2002) mendapatkan bahwa florotanin yang
diperoleh dari S. siliquastrum mampu menghambat aktivitas anion superoksida