ANALISIS GENETIK DAN SELEKSI HIBRIDA JAGUNG PADA KONDISI OPTIMAL DAN SUBOPTIMAL
ANDI ADRIANI WAHDITIYA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK
CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul analisis genetik dan seleksi hibrida jagung pada kondisi optimal dan suboptimal adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Maret 2016
RINGKASAN
ANDI ADRIANI WAHDITIYA. Analisis Genetik dan Seleksi Hibrida Jagung pada Kondisi Optimal dan Suboptimal. Dibimbing oleh SURJONO HADI SUTJAHJO, WILLY BAYUARDI SUWARNO, dan MUHAMMAD AZRAI.
Perakitan jagung toleran kondisi suboptimal (cekaman kekeringan) sangat penting untuk perluasan jagung di lahan marjinal kering, terutama di luar Jawa. Penelitian bertujuan untuk mengevaluasi penampilan hibrida silang uji pada kondisi lingkungan pengairan optimal dan suboptimal (cekaman kekeringan), menganalisis varian genetik dan heritabilitas karakter hasil pada lingkungan pengairan optimal dan suboptimal (cekaman kekeringan), mengidentifikasi hibrida yang toleran lingkungan suboptimal (cekaman kekeringan).
Percobaan ini dilakukan di Kalimantan Selatan (lingkungan optimal) dan Sulawesi Selatan (lingkungan optimal dan suboptimal) pada bulan Mei 2014-April 2015, menggunakan Rancangan Kelompok Lengkap Teracak (RKLT) faktor tunggal dengan dua ulangan pada masing-masing lingkungan pengujian. Terdapat 14 parameter pengujian yakni tinggi tanaman, tinggi tongkol, rasio tinggi tongkol terhadap tinggi tanaman, umur berbunga jantan, umur berbunga betina, anthesis silking interval, plant aspect, husk cover, ear aspect, bobot tongkol kupasan, rendemen, kadar air, produksi dan bobot seribu biji. Analisis data meliputi sidik ragam per lingkungan dan gabungan antar lingkungan, analisis stress tolerance index, serta analisis nilai heritabilitas.
Hasil analisis gabungan antar lingkungan menunjukkan bahwa genotipe berpengaruh nyata terhadap parameter hasil. Berdasarkan analisis heritabilitas, diperoleh hasil bervariasi mulai dari rendah-tinggi, baik pada analisis tiap lingkungan maupun pada analisis lingkungan gabungan. Berdasarkan intensitas seleksi 15% diperoleh 12 genotipe terbaik, dan dengan intensitas seleksi 10% akhirnya diperoleh 5 genotipe terbaik (berdasarkan produksi tertinggi). Dari 230 genotipe yang diuji, terdapat 3 genotipe dengan kategori sangat toleran kekeringan, 111 genotipe dengan kategori toleran moderat kekeringan, dan 116 genotipe dengan kategori peka (berdasarkan nilai stress tolerance index yang diuji pada lingkungan yang berbeda). Genotipe pembentuk hibrida yang terpilih merupakan genotipe yang dinilai potensial digunakan untuk pengembangan jagung toleran cekaman kekeringan.
SUMMARY
ANDI ADRIANI WAHDITIYA. Genetic Analysis and Selection of Hybrid Maize in Optimum and Suboptimum Conditions. Supervised by SURJONO HADI SUTJAHJO, WILLY BAYUARDI SUWARNO, and MUHAMMAD AZRAI.
Development of maize varieties tolerant to drought is important for agricultural extensification to dry marginal lands, especially to those located outside the Java island. The research objectives were to evaluate agronomic performance, genetic variability and heritability of test crosses hybrids maize.
The experiments were conducted in South Kalimantan (optimum condition) and South Sulawesi (optimum and suboptimum condition) from May 2014- April 2015. There were 14 traits evaluated, namely plant height, ear height, plant height to ear height ratio, days to male flowering, days to females flowering, anthesis silking interval, plant aspect, husk cover, ear aspect, number of grain rows per ear, shelling percentage, ear weight, and grain yield. Data analyses include: analyze of variance (by environment and combined over environments), estimation of stress tolerance index, and analysis of heritability.
The results of combined analysis over environments showed that the genotype factor significantly affected all observed traits. Heritability estimates varied from low to high, both in the single and multiple environments analysis. Selection based on ear yield with an intensity of 15% resulted in best 12 genotypes, whereas with a selection intensity of 10% resulted in best 5 genotypes. From 230 genotypes tested, there are 3 highly drought-tolerant genotypes, 111 moderate drought tolerant genotypes and 116 drought sensitive genotypes, based on the stress tolerance index. The parent of selected genotypes are considered potential for developing new drought tolerant maize varieties.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
ANALISIS GENETIK DAN SELEKSI HIBRIDA JAGUNG PADA KONDISI OPTIMAL DAN SUBOPTIMAL
ANDI ADRIANI WAHDITIYA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PRAKATA
Alhamdulillah dengan izin Allah Subhanahu wa Ta‟ala, penulis akhirnya
dapat menyelesaikan tesis yang berjudul “Analisis Genetik Dan Seleksi Hibrida Jagung Pada Kondisi Optimal Dan Suboptimal”, sebagai salah satu persyaratan untuk menyelesaikan Program Strata-2 Program studi Pemuliaan dan Bioteknologi Tanaman Fakultas Pertanian Sekolah Pascasarjana Institut Pertanian Bogor.
Terima kasih tak terhingga penulis haturkan kepada komisi pembimbing Prof Dr Ir Surjono Hadi Sutjahjo MS selaku pembimbing I (ketua), Dr Willy Bayuardi Suwarmo SP MSi selaku pembimbing II dan Dr Muhammad Azrai SP MSi selaku pembimbing III, atas dukungan, arahan dan bimbingannya selama penyusunan dan penulisan tesis ini.
Hal yang sama dihaturkan kepada Dr Desta Wirnas SP MSi atas saran, kritikan dan koreksinya sebagai tim penguji, Dr Ir Yudiwanti Wahyu E.K. MSi selaku ketua Program Studi Pemuliaan dan Bioteknologi Tanaman atas setiap motivasi, nasihat dan kesempatan yang telah diberikan untuk melanjutkan yaitu Program Magister (S2) Pascasarjana Institut Pertanian Bogor.
Terima kasih banyak kepada Dr Roy Efendi SP MSi dan Dr Amin Nur SP Msi yang senantiasa meluangkan waktu untuk memberikan ilmu dan arahan kepada penulis.
Terima kasih yang tak terhingga serta khusus disampaikan kepada kedua orang tua yang terkasih, Ayahanda Dr H Muhammad Yasin MP dan Ibunda Ir Hj. Andi Rugaya MP atas setiap cinta yang terpancar dan doa restu yang teriring serta kasih sayang sepanjang masa dari kedua orang tua penulis.
Kepada teman-teman seperjuangan yang senantiasa memberi semangat,
dukungan moral Dhian Eka Wijaya, Mar‟atus Sholihah, Umi Salamah, Dia
Hasanuddin, Eni Togatorop, Rahmi Henda Yani, Desi Ratna Sari, Aqlima, Ratna Ningsih Tarrafanur, dan Yudia Azmi. Terima kasih yang terhingga atas bantuan dan dukungannya.
Terima kasih banyak kepada pemberi beasiswa BPP-DN (calon dosen) Dikti yang telah menjadi sponsor utama, atas semua pembiyaan selama hidup di negri perantauan Bogor. Terima kasih kepada Balai Penelitian Serealia Maros (BALITSEREAL) dan BPTP Kalimantan Selatan Selatan atas pendanaan penelitian yang diberikan sehingga penelitian tesis penulis dapat berjalan dengan lancar sesuai yang diharapkan.
Harapan penulis, semoga tesis ini dapat memberikan manfaat yang sebesar-besarnya bagi para penuntut ilmu, baik dalam bangku perkuliahan maupun bidang penelitian, guna membina generasi muda penerus bangsa yang lebih berkualitas dan berdaya saing.
Bogor, Maret 2016
DAFTAR ISI
RINGKASAN ii
SUMMARY iii
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiii
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis Penelitian 2
Manfaat Penelitian 2
2 TINJAUAN PUSTAKA 3
Pemuliaan Jagung 3
Top Cross (Silang Puncak) 3
Cekaman Kekeringan pada Jagung 4
Respon Tanaman Jagung terhadap Cekaman Kekeringan 5
Mekanisme Toleransi Kekeringan pada Tanaman Jagung 6
3 METODE PENELITIAN 8
Tempat dan Waktu Penelitian 8
Bahan Penelitian 8
Bahan Tetua 9
Metode Seleksi Bahan Tanam 9
Metode Penanaman dan Emaskulasi 10
Rancangan Percobaan 11
Pelaksanaan Penelitian 11
Analisis Data 13
4 HASIL DAN PEMBAHASAN 15
Evaluasi Toleransi Kekeringan Hibrida Silang Uji pada Tiga Lingkungan
yang Berbeda 15
Analisis Gabungan Antar Lingkungan 23
5 KESIMPULAN DAN SARAN 27
Saran 27
DAFTAR PUSTAKA 28
DAFTAR TABEL
analisis gabungan berbagai lingkungan 15
4 Nilai kuadrat tengah ulangan, genotipe, galat dan koefisien keragaman karakter vegetatif dan pembungaan tanaman jagung pada pengamatan
tiga lingkungan yang berbeda 15
5 Nilai kuadrat tengah ulangan, genotipe, galat dan koefisien keragaman karakter generatif dan produksi tanaman jagung pada pengamatan tiga
lingkungan yang berbeda 16
6 Nilai kuadrat tengah ulangan, genotipe jagung, galat dan koefisien keragaman karakter produksi tanaman jagung pada pengamatan tiga
lingkungan yang berbeda 17
7 Korelasi antar peubah tanaman jagung pada kondisi optimal Sulawesi
Selatan 19
8 Korelasi antar peubah tanaman jagung pada kondisi suboptimal
(cekaman kekeringan) Sulawesi Selatan 19
9 Korelasi antar peubah tanaman jagung pada kondisi optimal Kalimantan
Selatan Selatan 20
10 Nilai heritabilitas tiap lingkungan untuk karakter vegetatif dan pembungaan tanaman jagung pada pengamatan tiga lingkungan yang
berbeda 21
11 Nilai heritabilitas tiap lingkungan untuk karakter produksi tanaman jagung pada pengamatan tiga lingkungan yang berbeda 21 12 Genotipe jagung terbaik berdasarkan parameter produksi (ton/ha)
tertinggi pada setiap lingkungan dengan intensitas seleksi 15% 22 13 Nilai kuadrat tengah lingkungan, ulangan (lingkungan), genotipe,
genotipe x lingkungan untuk semua karakter pengamatan tanaman jagung
pada gabungan tiga lingkungan yang berbeda 23
14 Nilai heritabilitas untuk gabungan semua lingkungan pada pengamatan
semua karakter tanaman jagung 24
15 Rataan pada pengamatan parameter vegetatif dan pembungaan untuk
masing-masing genotipe tanaman jagung terbaik 25
14 Rataan pada pengamatan parameter produksi untuk masing-masing
genotipe tanaman jagung terbaik 26
15 Korelasi Spearman antar lingkungan untuk semua karakter pengamatan
DAFTAR GAMBAR
1 Skema persilangan tetua untuk menghasilkan populasi 8
2 Diagram metode seleksi ear to row tanaman Jagung 10
3 Posisi penanaman jantan dan betina pada persilangan Mr 14 dan S2 10
4 Pedoman skor penutupan klobot pada Jagung 12
5 Diagram batang jumlah genotipe jagung pada setiap kategori nilai stress
tolerance index 23
DAFTAR LAMPIRAN
1 Nama genotipe jagung, varietas pembanding, dan tetua yang digunakan
dalam percobaan 35
2 Lengas tanah pada pertanaman jagung keadaan optimal dan suboptimal 36
3 Gambar pertanaman jagung di lapangan 37
4 Gambar lima genotipe jagung terbaik 38
1 PENDAHULUAN
Latar Belakang
Konsep strategi pencapaian ketahanan pangan yang telah diluncurkan oleh pemerintah meliputi tiga target yakni: penguatan distribusi dan penguatan harga pangan, peningkatan ketersediaan dan penanganan rawan pangan serta percepatan penganekaragaman konsumsi dan keamanan pangan. Khusus untuk mendukung tercapainya target peningkatan ketersediaan dan penanganan rawan pangan maka perlu dirintis suatu teknologi yang dapat meningkatkan produksi tanaman pangan salah satunya adalah jagung.
Produksi jagung di Indonesia tahun 2014 sebanyak 19.03 juta ton pipilan kering atau mengalami kenaikan sebanyak 0.52 juta ton (2.81 persen) dibandingkan dengan tahun 2013 (Badan Pusat Statistik 2014). Sampai dengan tahun 2015 produksi jagung nasional belum mampu memenuhi kebutuhan di Indonesia baik itu sebagai pangan maupun sebagai pakan ternak. Hal ini bisa terjadi disebabkan oleh penerapan teknologi produksi jagung yang belum optimum, disamping itu juga karena adanya cekaman biotik dan abiotik (Zubactirodin et al. 2008).
Tantangan dalam pengembangan tanaman pangan ke depan, khususnya tanaman jagung adalah perubahan iklim yang mengakibatkan tanaman akan sangat terganggu terutama akibat kondisi cuacayang suboptimal (cekaman kekeringan). Kendala yang dihadapi pada cekaman kekeringan adalah tidak tercukupinya air dalam tanah untuk pertumbuhan dan perkembangan tanaman. Pertumbuhan pada fase tertentu sangat berpengaruh besar terhadap hasil (Mapegau 2001). Sehingga produktivitasnya rendah (Subandi 1988).
Kekeringan merupakan suatu keadaan dimana terjadi kekurangan air dalam tanah dan tanaman dalam periode pertumbuhan tanaman sehingga berpengaruh negatif terhadap pertumbuhan dan perkembangan tanaman. Kebutuhan air untuk pertumbuhan tanaman jagung bergantung pada keadaan iklim, metode pengairan yang digunakan, dan varietas jagung yang ditanam. Menurut Dahlan (2001), agar tanaman jagung dapat tumbuh baik, memerlukan curah hujan rata-rata 25 mm/minggu. Menurut Levit (1980) stress kekeringan pada tanaman disebabkan oleh dua hal: (1) kekurangan suplai air di daerah perakaran dan (2) laju evapotranspirasi melebihi laju absorbsi air oleh akar tanaman. Faktor yang pertama banyak dialami oleh tanaman yang ditanam pada lahan-lahan kering di daerah tropis.
Kondisi suboptimum lahan kering merupakan salah satu sumberdaya lahan potensial untuk pembangunan pertanian Indonesia, khususnya tanaman jagung. Wilayah dataran rendah yang tersebar terutama di Indonesia bagian timur, serta sebagian di Jawa, Kalimantan dan Sumatera.
2
selalu terancam oleh resiko kekurangan air yang mengakibatkan penurunan dan kehilangan hasil produksi (Ekmekei 2005).
Salah satu upaya untuk mengatasi permasalahan tersebut adalah perakitan varietas jagung hibrida unggul baru toleran cekaman kekeringan. Seleksi galur-galur generasi awal yang sesuai dengan lingkungan target pengembangan merupakan salah cara yang cukup efektif untuk memperbesar kemajuan seleksi dan mempercepat perolehan varietas yang diinginkan (Banziger et al. 1997).
Penggunaan varietas unggul yang dihasilkan melalui program pemuliaan atau seleksi sangat ditentukan oleh tersedianya keragaman genetik yang disebut plasma nutfah (Monneveux et al. 2006; Al Badeiry et al. 2014). Berkaitan dengan hal tersebut, pada penelitian ini akan dilakukan seleksi galur generasi awal yang toleran terhadap kekeringan berdasarkan evaluasi hibrida silang ujinya pada kondisi lingkungan pengairan normal di musim hujan dan evaluasi kekeringan di musim kemarau.
Tujuan Penelitian
Tujuan dari penelitian ini adalah:
1. Mengevaluasi penampilan hibrida silang uji pada kondisi lingkungan pengairan optimal dan suboptimal (cekaman kekeringan).
2. Menganalisis varian genetik dan heritabilitas karakter hasil pada lingkungan pengairan optimal dan suboptimal (cekaman kekeringan).
3. Mengidentifikasi galur-galur yang toleran lingkungan suboptimal (cekaman kekeringan).
Hipotesis Penelitian
Hipotesis yang diajukan dalam penelitian ini adalah terdapat satu atau lebih genotipe yang menunjukkan respon toleran terhadap cekaman kekeringan, sehingga memudahkan peneliti dalam menentukan tetua (yang berpotensi toleran cekaman kekeringan), untuk dilanjutkan ke penelitian berikutnya.
Manfaat Penelitian
3
2 TINJAUAN PUSTAKA
Pemuliaan Jagung
Strategi pemuliaan tanaman jagung untuk mendapatkan varietas unggul baru adalah dengan cara persilangan dan seleksi berulang sebagai usaha pemuliaan jangka panjang, introduksi dari luar negeri dan perbaikan populasi, serta seleksi untuk stabilitas hasil dilakukan pada berbagai sentra produksi jagung. Untuk mendukung terlaksananya kegiatan pemuliaan maka yang harus diperhatikan terlebih dahulu adalah ketersediaan dari bahan genetik atau sumber daya genetik yang beragam. Keanekaragaman plasma nutfah tanaman jagung digunakan sebagai sumber gen bagi para pemuliaan untuk merakit varietas jagung yang lebih unggul (Mejaya et al. 2003). Dalam upayah perbaikan genetik karakter yang diinginkan melalui program pemuliaan perlu penambahan plasma nuthfah baru guna meningkatkan keragaman dalam populasi (Surjono et al. 2005).
Peningkatan hasil tanaman dapat dicapai dengan penggunaan varietas hibrida. Varietas hibrida dapat diperoleh dengan metode pemuliaan, salah satunya dengan melakukan silang puncak dimana pola perkawinan silang puncak tersebut melibatkan sejumlah persilangan dari galur atau kultivar yang menggunakan tetua penguji. Tetua jantan digunakan sebagai penguji sedangkan tetua betina (galur, kultivar) yang diuji dapat berupa mandul jantan, self incompatible atau diemaskulasi sebelum kotak sari pecah (Stube 1980; Poespodarsono 1988).
Top Cross (Silang Puncak)
Silang puncak merupakan persilangan antara suatu galur/kultivar/varietas dengan penguji. Pola perkawinan silang puncak melibatkan sejumlah persilangan dari galur atau kultivar yang menggunakan tetua penguji. Silang puncak atau top cross, adalah progeni hibrida yang dihasilkan melalui penyerbukan suatu galur murni dengan suatu populasi yang menghasilkan pollen yang tercampur secara genetik (Willy 2008) Tetua jantan digunakan sebagai penguji sedangkan tetua betina (galur, kultivar) yang diuji dapat berupa mandul jantan, self incompatible atau diemaskulasi sebelum kotak sari pecah.
Tanaman yang mampu memberikan informasi jelas tentang sifat yang diinginkan bila dikombinasikan dengan tanaman teruji atau bila ditumbuhkan pada lingkungan berbeda (Stube 1980; Poespodarsono 1988).Menurut Hallauer (1975) silang puncak bertujuan untuk mengurangi jumlah galur yang terlalu besar yang akan diuji untuk membentuk varietas unggul baru hibrida maupun multini hasil tinggi. Silang puncak merupakan salah satu prosedur yang dapat digunakan untuk mengevaluasi galur-galur atau varietas yang potensial untuk dikembangkan lebih lanjut.
4
Cekaman Kekeringan pada Jagung
Sebagian besar tanaman jagung (Zea mays L.) di Indonesia (79 % dari areal pertanian jagung) ditanam pada lahan tegalan, sehingga ketersediaan air tergantung pada curah hujan (Dahlan 2001). Tanaman jagung yang mengalami cekaman air dapat mengalami kerusakan sel, kehilangan turgor, stomata tertutup, daun tanaman menggulung kemudian layu, pertukaran gas terganggu dan akhirnya tanaman tidak memberikan hasil pada kandungan lengas tanah yang sangat rendah (Rashidi dan Seyfi 2007). Tanaman jagung yang mengalami kekeringan akan berubah warna menjadi kuning pucat, daun tanaman menggulung seperti pensil dan tidak sinkron waktu pembentukan malai dan keluarnya rambut (Edmeades et al. 1992).
Fase perkecambahan dan fase vegetatif diduga merupakan fase yang sangat sensitif dalam kaitannya dengan ketersediaan air, sebab dapat menghambat proses pertumbuhan selanjutnya. Kekurangan air menghambat proses deferensiasi sel, pembelaan sel, dan pembentangan sel pada jaringan meristematik sehingga mengurangi jumlah biji. Jumlah biji yang terbentuk sangat ditentukan oleh jumlah fotosintat yang dapat ditranslokasikan untuk pembentukan biji tersebut (Sasmita 1996; Jatoi et al. 2014). Masa kritis tanaman jagung terhadap kekeringan adalah pada waktu tanaman berbunga, ketersediaan air pada waktu berbunga berhubungan dengan hasil biji. Kekurangan air pada waktu berbunga dapat mengurangi hasil sampai 22 % (Dahlan 2001). Cekaman air dapat mengurangi tinggi tanaman, luas daun, berat tajuk dan berat akar tanaman jagung.
Tanaman jagung yang mendapat cekaman air selama dua minggu sejak tanaman berumur tiga minggu segera memberikan tanggapan yang cepat sedangkan tanaman sorgum tanggapannya lebih lambat. Semakin tinggi cekaman air mengakibatkan makin rendahnya transpirasi dan meningkatnya ketahanan difusi daun (Sutoro et al. 1989).
Kekeringan, merupakan salah satu faktor eksternal yang mempengaruhi pertumbuhan tanaman (Farooq et al. 2009). Menurut (Levitt 1980; Bray 1997) kekeringan merupakan istilah untuk menyatakan bahwa tanaman mengalami kekurangan air akibat keterbatasan air dari lingkungan yaitu media tanam, sedangkan Mathius et al. (2001) menyatakan bahwa kekeringan disebabkan karena (1) kekurangan suplai air di daerah sistem perakaran dan (2) permintaan air yang berlebihan oleh daun karena laju evapotranspirasi lebih tinggi dibandingkan dengan laju absorpsi air oleh akar, meskipun keadaan air tanah tersedia cukup.
5
meliputi penurunan pertumbuhan batang dan daun, sedangkan perubahan biokimia dapat berupa akumulasi senyawa organik compatible yang berfungsi menjaga keseimbangan osmolit dalam tubuh tumbuhan. Salah satu senyawa organik kompatibel yang sering diamukulasi oleh tanaman ketika berada pada kondisi kekeringan yaitu prolin (Farooq et al. 2009). Akumulasi prolin terhadap cekaman kekeringan telah dilaporkan oleh banyak peneliti misalnya pada jagung Flirricks et al. (2012) menyatakan bahwa kekeringan akan menyebabkan penurunan pertumbuhan akar, penurunan panjang daun, indeks luas daun, dan keterlambatan memasuki fase reproduksi, serta penurunan hasil.
Respon Tanaman Jagung terhadap Cekaman Kekeringan
Kondisi pada tubuh tanaman terdapat hubungan yang erat antara absorbsi dengan perkembangan akar. Untuk tanaman yang akarnya berkembang kuat terjadi peningkatan absorbsi air dan relatif lebih toleran terhadap kekeringan. Banyak sifat-sifat tanaman baik morfologi maupun fisiologi yang dapat digunakan sebagai dasar penilaian sifat ketahanan terhadap kekeringan seperti pola kedalaman perakaran, jumlah stomata, lebar stomata, penyesuaian osmosis, peningkatan elastisitas dinding sel (Sammons et al. 1980; Kramer 1980).
Umumnya pengaruh fisiologi stres air pada tanaman yang paling menonjol dalam jaringan yang sedang tumbuh dengan cepat, yakni pada fase perkecambahan dan pertumbuhan awal vegetatif. Kemampuan benih berkecambah pada kandungan air tanah yang rendah tergantung kepada spesies. Setiap spesies memerlukan penyerapan air yang minimum untuk bisa berkecambah dan tampaknya mempunyai batas tegangan tersendiri. Nilai batas tersebut -1.25 MPa untuk jagung. Kondisi cekaman air, tanaman akan memperlihatkan berbagai respon sebagai mekanisme tanaman dalam usaha mengurangi cekaman yaitu: respon morfologi dengan mengurangi luas permukaan daun sehingga transpirasi menurun, mempercepat perkembangan perakaran terutama kearah bawah menyebabkan nisbah akar/pucuk meningkat sehingga tanaman lebih mampu mengabsorbsi air dari lapisan tanah yang lebih dalam sementara transpirasi dari bagian atas tanaman menurun (Herawati 2000), mengubah sudut daun pada posisi hampir sejajar dengan datangnya cahaya, agar suhu daun tidak segera meningkat sehingga transpirasi dapat ditekan, pembentukan lapisan kutikula pada permukaan daun dapat mengurangi penguapan. Selain itu lapisan lilin dapat meningkatkan pantulan cahaya, sehingga mengurangi suhu permukaan daun.
6
MPa sampai -0.25 MPa dan menurun 25% jika potensial air turun sampai -0,4 Mpa, serta mengulungan atau pelipatan daun. Tanaman kedelai berdaun lebar cendurung untuk mengulung daun keatas sehingga bulu-bulu (rambut) diatas permukaan bawah daun yang terbuka dapat merefleksikan lebih banyak cahaya.
Selain menunjukkan respon morfologi, ternyata tanaman yang mengalami kekeringan akan menunjukkan respon fisiologi pada kondisi kekeringan telah lama diketahui. Suatu hal yang cukup penting diantaranya adalah kemampuan tanaman mempertahankan tekanan turgor dengan menurunkan potensial osmotiknya (Jones et al. 1981).
Respon tanaman untuk mengatasi cekaman kekeringan adalah dengan pengaturan osmotik sel. Pada mekanisme ini terjadi sintesis dan akumulasi senyawa organik yang dapat menurunkan potensial osmotik sehingga menurunkan potensial air dalam sel tanpa membatasi fungsi enzim serta menjaga turgor sel. Beberapa senyawa yang berperan dalam penyesuaian osmotikal sel antara lain gula osmotik (Wang et al. 1995; Yakhushiji et al. 1998), prolin dan betain (Maestri et al. 1995), protein dehidrin (Close 1997) dan asam absisik (ABA) yang berperan dalam memacu akumulasi senyawa tersebut (Earl and Davis 2003). Menurut Fukai (1995) bahwa akumulasi hormon asam absisik (ABA) diperlukan untuk peningkatan proline pada kondisi potensial air rendah.
Hasil penelitian Sharp and Davies (1979); Westgate and Boyer (1985) menyatakan bahwa senyawa prolin berkontribusi lebih dari 50% terhadap osmotic adjustment pada akar jagung. Pembentukan senyawa osmoregulasi ini sebagai penanda biokimia untuk indikasi toleransi cekaman kekeringan. Banyak peneliti menyatakan bahwa prolin bebas banyak diakumulasi sebagai respon terhadap stress air yang dapat diamati pada daun-daun yang masih melekat maupun yang telah gugur pada banyak tanaman budidaya pada kondisi laboratorium (Abdelmoneim et al. 2014; Barnett and Beck et al. 1997; Routley and Singh 1966).
Akumulasi asam absisik (ABA) berkaitan juga dengan respon tanaman yang toleran cekaman kekeringan. Akar yang mengalami cekaman kekeringan, menurut Salisbury and Ross (1992) akan membentuk asam absisik lebih banyak dan diangkut melalui xilem menuju daun untuk menutup stomata. Selain itu kadar ABA endogen yang tinggi juga dapat diketahui dapat menginduksi peningkatan rasio pertumbuhan akar/tajuk (Biddington and Dearman 1982). Kenyataan ini menunjukkan respon yang berbeda dari akar dan tajuk terhadap ABA (Creelman et al. 1990). Pada tajuk, ABA menginduksi penghambatan sedangkan pada akar ABA mendorong pertumbuhan (Dallaire et al. 1994).
Mekanisme Toleransi Kekeringan pada Tanaman Jagung
7
yang terekspresi sebagai respons terhadap cekaman (Fukai and Cooper 1995). Menurut Blum (2002) bahwa kemampuan mempertahankan turgor atau status air sangat penting dalam toleransi kekeringan. Kemampuan tersebut secara kuantitatif lebih diperankan oleh karakter kuantitatif dibanding karakter adaptasi. Menurut Ahadiyat (2014) genotipe toleran cekaman kekeringan memiliki bobot kering akar yang besar dibanding genotipe peka, baik pada kondisi cekaman kekeringan maupun optimum. Sehingga karakter bobot akar dapat seleksi adalah karakter akar dapat diseleksi pada lingkungan optimum.
Berdasarkan kemampuan genetik tanaman, terdapat empat mekanisme adapatasi pada kondisi cekaman kekeringan yaitu drought escape, dehydration avoidance, dehydration tolerance dan drought recovery (Sopandie 2006). Menurut Sasmita (1996) tanaman seringkali menggunakan lebih dari satu mekanisme untuk beradaptasi pada kondisi cekaman kekeringan mekanisme tersebut adalah: melepaskan diri dari cekaman kekeringan yaitu kemampuan tanaman menyelesaikan siklus hidupnya sebelum mengalami defisit air yang parah, mekanisme ini ditunjukkan dengan perkembangan sistem pembungaan yang cepat dan perkembangan plastisitas jaringannya. Menurut Muhadjir (1988) mekanisme adaptasi berupa percepatan siklus hidup atau adanya genotipe genjah dengan umur pendek umumnya berdaya hasil rendah dibandingkan dengan yang berumur panjang.
Potensial air jaringan yang tinggi yang merupakan bentuk toleransi yaitu kemampuan tanaman yang tetap menjaga potensial jaringan dengan meningkatkan penyerapan air atau menekan kehilangan air (Banziger et al. 1997). Tanaman yang melakukan mekanisme toleransi mempunyai kemampuan untuk meningkatkan sistem perakaran dan konduktivitas hidrolitik atau kemampuan untuk menurunkan hantaran epidermis dengan regulasi stomata, pengurangan absorbsi radiasi dengan pembentukan lapisan lilin, bulu yang tebal dan penurunan permukaan evapotranspirasi melalui penyempitan daun serta pengguguran daun tua (Farooq et al. 2009). Oksidasi proline, setelah keadaan normal terjadi dengan cepat untuk menjaga kandungan proline yang rendah dalam tanaman. Beberapa penelitian menunjukkan bahwa tanaman yang toleran terhadap cekaman air memperlihatkan kemampuan mengakumulasi prolin (Sopandie 2006).
8
3 METODE PENELITIAN
Tempat dan Waktu Penelitian
Percobaan ini dilaksanakan di dua lokasi yaitu KP Tanah Laut, Kalimantan Selatan dan KP. Bilibili Sulawesi Selatan pada bulan Mei 2014-April 2015. Lokasi pengujian KP. Tanah Laut terletak pada ketinggian 100 m dpl, dan jenis tanah Inceptisol. Sedangkan lokasi pengujian KP. Bilibili Sulawesi Selatan terletak pada ketinggian 150 m dpl, dan jenis tanah Tropodult. Kedua tempat tersebut dipilih dengan alasan kedua lokasi tersebut merupakan representasi areal target pengembangan setelah varietas dirilis.
Bahan Penelitian
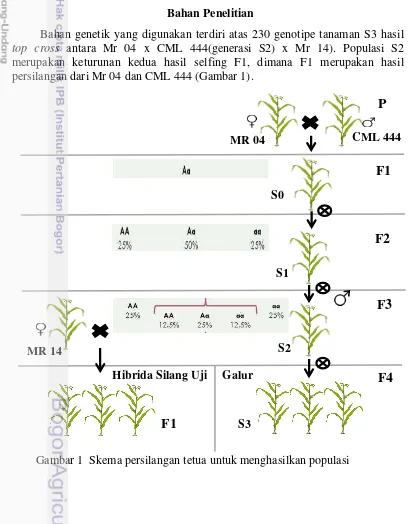
Bahan genetik yang digunakan terdiri atas 230 genotipe tanaman S3 hasil top cross antara Mr 04 x CML 444(generasi S2) x Mr 14). Populasi S2 merupakan keturunan kedua hasil selfing F1, dimana F1 merupakan hasil persilangan dari Mr 04 dan CML 444 (Gambar 1).
MR 04
P
F1
F2
F3
MR 14
Hibrida Silang Uji
F4
F1
CML 444
S0
S1
S2
S3
9
Bahan Tetua
Bahan tetua yang digunakan merupakan plasma nutfah dari Balitsereal dan CIMMYT (International Maize and Wheat Improvement Center), masing-masing tetua memiliki kelebihan dan kekurangan yang berbeda-beda (Tabel 1).
Tabel 1 Daftar genotipe, asal dan karakteristik tetua galur murni yang digunakan dalam pembentukan populasi percobaan
Tetua Genotipe Asal Karakteristik
P1 Mr 04 Balitsereal Biji berwarna kuning, peka terhadap kekeringan, sulit diperbanyak punya daya gabung khusus yang baik, daun terkulai, dan
Mr : Maros, CML: CIMMYT Maize Line
Tahap awal perakitan F1, yang digunakan sebagai tetua adalah Mr 04 (betina) dan CML 444 (jantan). Kemudian untuk hibrida silang uji, tetua yang digunakan adalah generasi S2 yang merupakan hasil selfing kedua dr F1 (jantan) dan disilangkan dengan Mr 14 (betina). Populasi S2 digunakan sebagai pejantan dengan tujuan agar tetap dapat dilakukan kegiatan selfing pada waktu yang bersamaan, sehingga dapat lebih mengefisienkan waktu.
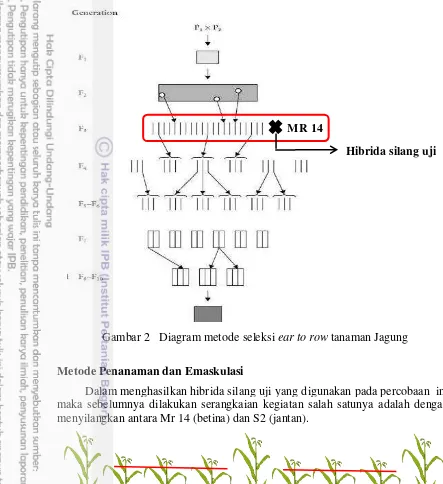
Metode Seleksi Bahan Tanam
Proses seleksi dan silsilah bahan tanaman tetua jagung yang digunakan dalam penelitian ini ditunjukkan pada Gambar 2. Untuk menghasilkan bahan tanam yang digunakan pada percobaan ini, F1 hasil persilangan awal telah mengalami serangkaian seleksi dan silang diri hingga mencapai keturunan generasi ke 3 atau S2. Adapun metode seleksi yang digunakan adalah metode seleksi tongkol ke baris (ear to row).
Pada musim pertama, tanam populasi dasar sekitar 3000-5000 tanaman (F1). Pilih 300-400 tanaman yang mempunyai karakter yang dikehendaki dan buat silangdiri untuk menghasilkan galur S1. Panen terpisah tanaman hasil silangdiri yang masih mempunyai karakter yang diinginkan.
Pada musim kedua, biji yang diperoleh pada musim 1 (S1) dari tiap tongkol ditanam satu baris sebanyak ±25 tanaman. Seleksi secara visual dilakukan antara famili dan dalam famili (baris) yang tanamannya tegap, tidak rebah, bebas hama penyakit, tongkol besar, produksi tinggi dan sebagainya, dan pilih 3-5 tanaman dari baris yang terpilih untuk silangdiri. Panen terpisah masing-masing tongkol, pilih 1-3 tongkol hasil silangdiri tiap baris terpilih dan diperoleh biji S2.
10
Gambar 2 Diagram metode seleksi ear to row tanaman Jagung
Metode Penanaman dan Emaskulasi
Dalam menghasilkan hibrida silang uji yang digunakan pada percobaan ini maka sebelumnya dilakukan serangkaian kegiatan salah satunya adalah dengan menyilangkan antara Mr 14 (betina) dan S2 (jantan).
Gambar 3 Posisi penanaman jantan dan betina pada persilangan Mr 14 dan S2 Garis merah menunjukkan perlakuan tanaman emaskulasi pada tetua betina
11
Rancangan Percobaan
Percobaan ini menggunakan Rancangan Kelompok Lengkap Teracak (RKLT) faktor tunggal dengan dua ulangan pada masing-masing lingkungan pengujian. Menurut Liu (1998) tiap lokasi yakni sebagai berikut :
Yij = µ + Gi + Kj + εij
Keterangan :
Yij = Nilai pengamatan pada genotipe ke-i, kelompok ke-j µ = Nilai tengah umum
Gi = Pengaruh genotipe ke-i, dimana i =1,2,3,...235 Kj = Pengaruh kelompok ke-j, dimana j=1,2
εij = Galat percobaan pada genotipe ke-i, kelompok ke-j
Model linier aditif untuk analisis gabungan antar lingkungan adalah sebagai berikut:
Yijk = µ + Li + K(L)ji + Gk + GLki + εijk
Keterangan :
Yijk = Nilai pengamatan pada lingkungan ke-i, kelompok ke-j, dan genotipe ke-k µ = Nilai tengah umum
Li = Pengaruh lingkungan ke-i, dimana i=1,2,3
K(L)ji = Pengaruh kelompok ke-j dalam lingkungan ke-i, dimana j=1,2 Gk = Pengaruh genotipe ke-k, dimana k =1,2,3,....235
GLki = Pengaruh interaksi antara genotipe ke-k dan lingkungan ke-i
εijk = Galat percobaan pada lingkungan ke-i, kelompok ke-j, dan genotipe ke-k
Pelaksanaan Penelitian
Tiap hibrida silang uji masing-masing ditanam satu baris per nomor dengan menggunakan plot baris sepanjang 5 m dengan jarak tanam 70 cm antar baris dan 20 cm dalam baris, ditumbuhkan 1 biji per lubang berdasarkan Rancangan Acak Kelompok dengan dua ulangan di tiap lingkungan. Pada umur 10 hst dilakukan pemupukan pertama dengan dengan dosis pupuk masing-masing 300 kg NPK Phonska/ha dan 100 kg urea/ha. Pemupukan kedua dilakukan pada umur 30 hari setelah tanam dengan dosis masing-masing 100 kg NPK Phonska/ha dan 250 kg urea/ha. Penyiangan dan pembumbunan dilakukan secara optimal, sedangkan aplikasi herbisida/insektisida menyesuaikan kebutuhan.
12
1. Tinggi tanaman (cm)
Pengamatan dilakukan setelah stadia pembungaan dengan cara mengukur tinggi tanaman dari permukaan tanah sampai dengan pangkal terakhir bunga jantan.
2. Tinggi letak tongkol (cm)
Diukur bersamaan dengan pengukuran tinggi tanaman. Pengukuran dilakukan dari permukaan tanah sampai dengan pangkal tongkol yang pertumbuhannya sempurna (umumnya pada tongkol bagian atas pada jagung prolifik).
3. Rasio tinggi tanaman dan tinggi tongkol
Dengan menghitung selisih antara tinggi tanaman dan tinggi tongkol 4. Umur 50% tanaman berbunga (hst) jantan
Pengamatan dilakukan setiap hari, sepanjang stadia pembungaan. Pencatatan umur berbunga jantan bukan ditandai setelah keluarnya bunga jantan (tassel), tetapi dihitung pada saat antesis atau ketika telah diproduksinya serbuk sari (pollen). Pollen berwarna kuning akan terlihat apabila tassel/malai digoyang. 5. Umur 50% tanaman berbunga (hst) betina
Pengamatan dilakukan setiap hari, sepanjang stadia pembungaan. Berbunga betina (keluar rambut) dicatat bila rambut telah keluar panjang >2 cm. 6. Anthesis silking interval
Dengan menghitung selisih waktu keluarnya bunga jantan dan bunga betina 7. Plant aspect/keragaan (penampilan) tanaman
Pengamatan dilakukan secara visual pada saat tanaman berumur 75 HST, yaitu pada waktu rambut tongkol telah mulai mengering tapi tanaman masih hijau. Penilaian menggunakan skor 1 (sangat baik), dan 5 (sangat buruk) terhadap petakan-petakan tanaman. Perhatian diarahkan pada keseragaman pertumbuhan tanaman, serangan hama dan penyakit secara umum, dan vigor. 8. Husk cover/penutupan klobot
Tingkat penutupan klobot diberi skor 1 (baik) sampai 5 (buruk), dengan kriteria sebagai berikut:
Skor 1 : Klobot menutup rapat dengan baik, sehingga beberapa tongkol dapat diikat menjadi satu pada ujung tongkol
Skor 2 : Klobot menutup ketat hanya sampai ujung tongkol saja Skor 3 : Klobot menutup agak longgar diujung tongkol
Skor 4 : Klobot menutup tongkol kurang baik, ujung tongkol terlihat
Skor 5 : Klobot menutup tongkol sangat jelek, sebagian biji nampak tidak dilindungi klobot.
13
9. Ear aspect /pengamatan keragaan (penampilan) tongkol
Dicatat waktu panen sebelum mengambil sampel untuk menentukan kadar air. Tongkol hasil panen disusun dan letakkan secara teratur kemudian diberi Skor 1 (terbaik) sampai 5 (terburuk) dengan mempertimbangkan kerusakan karena hama dan penyakit, ukuran, mengisinya biji dan keseragaman.
10.Bobot tongkol kupasan (kg)
Tongkol-tongkol yang dipanen adalah seluruh tongokol pada dua baris tengah per petak. Klobot tongkol dibuka kemudian ditimbang beratnya per petak. Data ini akan digunakan untuk menghitung hasil per petak.
11.Rendemen
Rendemen diukur dengan menimbang 10 tongkol kupasan basah yang diambil dari tongkol panen secara acak kemudian dipipil. Janggel tongkol ditimbang kembali sehingga rendemen dapat diketahui dengan rumus:
obot 10 tongkol kupasan basah- obot janggel obot 10 tongkol kupasan basah
12.Kadar air biji panen (%)
Setelah ditimbang bobot kupasan tongkol, diambil 5 tongkol sampel per petak kemudian setiap tongkol dipipil bijinya ±3-5 baris. Biji hasil pipilan dicampur kemudian diukur kadar airnya dengan menggunakan alat pengukur kadar air digital.
13.Hasil panen (ton/ha)
Dilakukan dengan cara mengkonversi hasil panen tongkol kupasan basah pada dua baris tengah tanaman per nomor dengan menggunakan rumus.
asil (tonha) 10000 100- A100-1 S
K.A = Kadar air biji waktu panen LP = Luas panen (m2).
B = Bobot tongkol kupasan (kg)
SP = Rata-rata „shelling percentage/rendemen‟
14.Bobot seribu biji
Bobot 1000 biji dalam kadar air 15 %. Untuk pengukuran ini tidak harus
„menunggu‟ kadar air 1 %. iji yang dipipil sejumlah ± 1000 butir dapat
langsung ditimbang dan diukur kadar air biji kemudian dikonversi pada kadar air 15%.
Analisis Data
Percobaan ini menggunakan analisis sidik ragam untuk tiap lingkungan dan gabungan berbagai lingkungan yakni sebagai berikut (Gomez and Gomez 1995):
Tabel 2 Tabel sidik ragam rancangan acak kelompok lengkap teracak untuk tiap lingkungan
Sumber Keragaman Derajat Bebas Jumlah Kuadrat Kuadrat Tengah E (KT)
Ulangan r-1 JK U KT Ulangan
Genotipe g-1 JK L KT Genotipe σ2e+ r σ2 g
14
Tabel 3 Tabel sidik ragam rancangan acak kelompok lengkap teracak untuk gabungan berbagai lingkungan
Sumber Keragaman Derajat Bebas Jumlah Kuadrat Kuadrat Tengah E (KT)
Lokasi (E) l-1 JK L KT Lokasi σ2e + g σ2 r / l + gr σ2 l
Ulangan/Lokasi l(r-1) JK U KT Ulangan σ2e + g σ2 r / l
Genotipe g-1 JK G KT Genotipe σ2e + r σ2 gl + rl σ2 g
G x E (g-1)(l-1) JK GxE KT GxE σ2e + r σ2gl
Galat l(r-1)(g-1) JK G KT Galat σ2e
Perhitungan stress tolerance index dilakukan untuk tiap genotipe berdasarkan rumus dari Henry (2013) yakni:
STI = Yd Yn / (MYn)2 Keterangan :
Yd : Hasil pada perlakuan stres kekeringan; Yn : Hasil pada perlakuan pengairan normal
MYn : Rata-rata hasil pada perlakuan pengairan normal
Tujuan perhitungan stress tolerance index adalah untuk melihat seberapa toleran genotipe yang diamati terhadap cekaman kekeringan yang diberikan yang dideskripsikan dengan nilai tertentu. Kategori nilai stress tolerance index dibagi menjadi tiga yakni : toleran (>1), moderat (0.5-1), peka (<0.5) (Fisher 1997).
Nilai heritabilitas arti luas (H) karakter hasil didefinisikan sebagai perbandingan antara ragam genetik (VG) dan ragam fenotip (VP). Mencari heritabilitas arti luas umtuk tiap lingkungan yakni sebagai berikut:
VG =
VP = Ragam fenotipe (entry mean basis) H = Heritabilitas dalam arti luas
15
VGxE = Ragam genotipe x lingkungan VP = Ragam fenotipe (entry mean basis) H = Heritabilitas dalam arti luas
Nilai heritabilitas menurut Kramer (1980) dibagi menjadi tiga yakni: tinggi (>70%), sedang (30%-70%), rendah (<30%).
Analisis data dilanjutkan dengan melakukan multiple seleksi pada lingkungan yang berbeda, dengan intensitas seleksi 15% kemudian dilanjutkan dengan intensitas seleksi 10%, dengan tujuan agar mempersempit jumlah genotipe yang diperoleh.
4 HASIL DAN PEMBAHASAN
Evaluasi Toleransi Kekeringan Hibrida Silang Uji pada Tiga Lingkungan yang Berbeda
Data hasil analisis ragam dari 235 genotipe (termasuk pembanding dan tetua) yang ditanam pada 3 lingkungan yang berbeda disajikan pada Tabel 4 sampai 6.
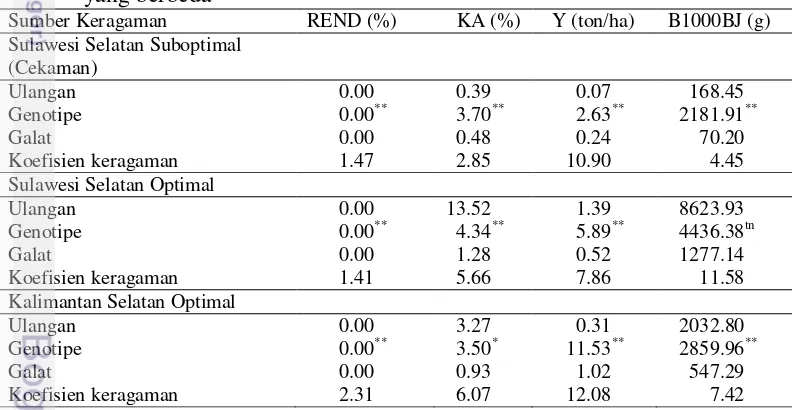
Tabel 4 Nilai kuadrat tengah ulangan, genotipe, galat dan koefisien keragaman karakter vegetatif dan pembungaan tanaman jagung pada pengamatan tiga lingkungan yang berbeda
16
Lingkungan Sulawesi Selatan suboptimal (cekaman), pengaruh genotipe sangat nyata terhadap tinggi tanaman, tinggi tongkol, rasio tinggi tongkol dan tinggi tanaman, umur berbunga jantan, dan umur berbunga betina. Hal ini berarti bahwa penampilan morfologi dipengaruhi oleh sifat genetik yang dibawah oleh tiap genotipe. Monneveux et al. (2006) menyatakan bahwa perbedaan sifat dari setiap tanaman dalam beberapa hal dapat dikendalikan oleh faktor genetik. Genotipe juga berpengaruh sangat nyata terhadap karakter-karakter tersebut pada lingkungan Sulawesi Selatan optimal, hal ini berarti genetik tanaman sangat mempengaruhi kenampakan morfologi tanaman. Pendapat ini didukung oleh Efendi (2014) yang menyatakan bahwa pada kondisi optimal, tanaman hampir tidak mengalami cekaman, baik itu abiotik maupun biotik sehingga faktor lingkungan berpengaruh relatif kecil terhadap keragaan morfologi tanaman, sehingga pengaruh genetik tanaman dapat terekspresi lebih baik.
Hal berbeda ditunjukkan pada Kalimantan Selatan optimal dimana untuk parameter tinggi tongkol dan rasio tinggi tanaman terhadap tinggi tongkol, pengaruh genotipe tidak nyata. Hal ini mengindikasikan bahwa pengaruh lingkungan terhadap kedua karakter cukup besar. Sesuai dengan pendapat Mundre (2002) yang menyatakan bahwa pertumbuhan tanaman dipengaruhi oleh banyak faktor baik internal maupun eksternal. Faktor eksternal yang mempengaruhi salah satunya adalah media tanam, dimana jika media tanam tidak dapat memenuhi nutrisi yang dibutuhkan oleh tanaman, maka tanaman tetap tidak dapat tumbuh dengan baik.
Tabel 5 Nilai kuadrat tengah ulangan, genotipe, galat dan koefisien keragaman karakter generatif dan produksi tanaman jagung pada pengamatan tiga lingkungan yang berbeda *= berpengaruh nyata pada taraf 5%, tn=tidak berpengaruh nyata; ASI=anthesis silking interval,
PA= plant aspect, HC= husk cover, EA= ear aspect, BTKP=bobot tongkol kupasan.
17
cover, ear aspect, dan bobot tongkol kupasan menunjukkan pengaruh genotipe sangat nyata. Sedangkan untuk peubah plant aspect, menunjukkan pengaruh genotipe nyata, dan untuk peubah anthesis silking interval menunjukkan pengaruh genotipe tidak nyata. Lingkungan Kalimantan Selatan optimal (Tabel 5) menunjukkan pengaruh genotipe sangat nyata terhadap semua peubah pengamatan kecuali pada peubah anthesis silking interval yang menunjukkan bahwa genotipe tidak berpengaruh nyata. Muhadjir (1988) berpendapat bahwa pertumbuhan dan perkembangan genotipe sangat dipengaruhi oleh lingkungan, proses fisiologis dipengaruhi oleh faktor lingkungan seperti iklim. Selain itu, kemampuan tanaman untuk tumbuh dan berkembang secara optimal saat pertumbuhan vegetatif menjadi salah satu indikator keberhasilan dalam menunjukkan potensi produksinya (Tabel 6) yang merupakan hasil interaksi antara faktor genetik dan faktor lingkungan.
Tabel 6 menunjukkan hasil analisis ragam pada tiga lingkungan untuk peubah rendemen, kadar air, hasil, dan bobot seribu biji. Pada ketiga lingkungan menunjukkan bahwa genotipe berpengaruh sangat nyata terhadap semua peubah yang diamati kecuali pada peubah bobot seribu biji pada lingkungan Sulawesi Selatan optimal, menunjukkan bahwa genotipe tidak berpengaruh nyata. Selain itu juga pada lingkungan Kalimantan Selatan optimal untuk peubah kadar air menunjukkan hasil yang berbeda, yakni pada peubah tersebut, genotipe menunjukkan hasil berpengaruh nyata. Menurut Makmur (1993) cekaman kekeringan tanaman jagung yang terjadi mulai pada fase pembungaan bunga sampai fase pematangan biji akan menurunkan pertumbuhan tinggi tanaman, rendemen, bobot seribu biji, serta hasil jagung per hektar. Pengaruh nyata atau sangat nyata dari genotipe ini diduga disebabkan adanya perbedaan atau keragaman secara genetik dari genotipe jagung yang diuji (Azizian 2014).
Tabel 6 Nilai kuadrat tengah ulangan, genotipe, galat dan koefisien keragaman karakter produksi tanaman jagung pada pengamatan tiga lingkungan yang berbeda
18
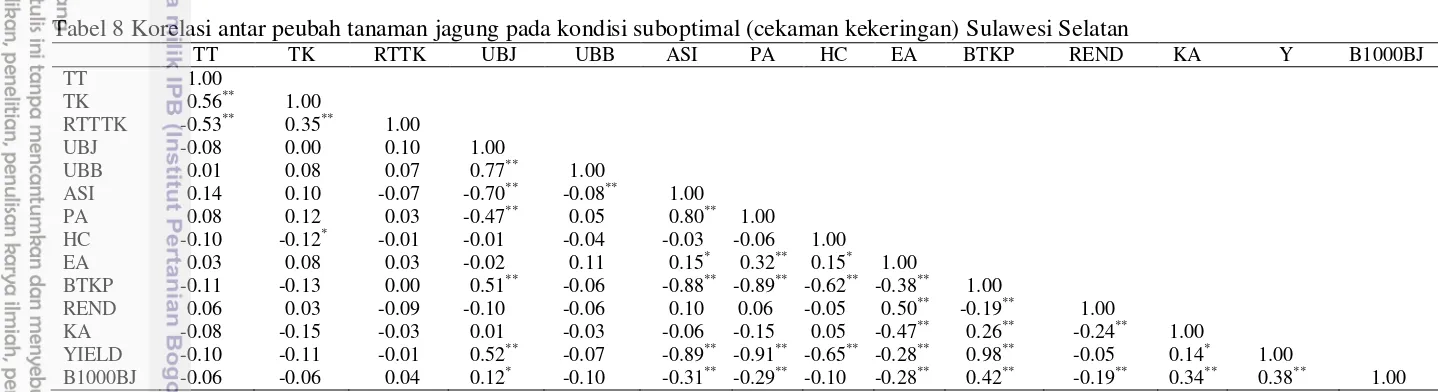
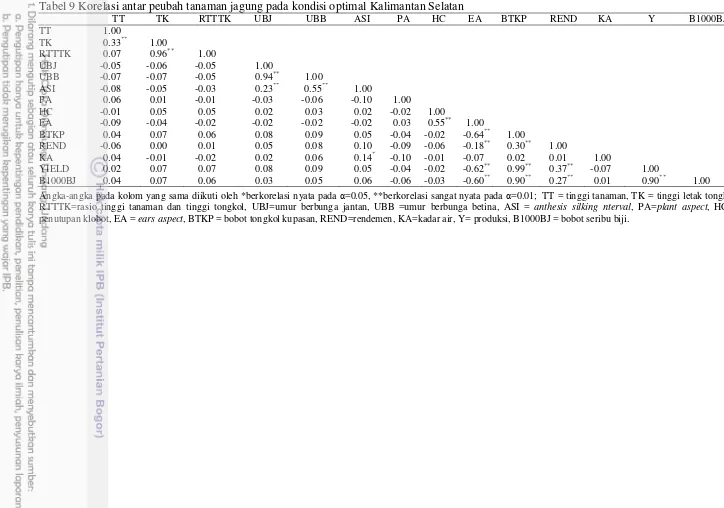
Berdasarkan hasil analisis korelasi pada masing-masing lingkungan maka diperoleh hasil yang disajikan pada Tabel 7-9. Tabel 7 menunjukkan hasil analisis korelasi antar peubah pada kondisi optimal (Sulawesi Selatan), dapat dilihat bahwa tinggi tanaman berkorelasi positif sangat nyata dengan tinggi tongkol, semakin tinggi tanaman maka semakin besar kemungkinan letak tongkol juga semakin tinggi, dan hal ini juga berpengaruh langsung pada parameter rasio tinggi tanaman dan tinggi letak tongkol. Pada parameter lain dapat dilihat bahwa umur berbunga jantan berkorelasi sangat nyata dengan parameter umur berbunga betina, dan anthesis silking interval berkorelasi sangat nyata dengan kedua parameter tersebut. Menurut Earl and Davis (2003) kondisi cekaman kekeringan menyebabkan pembentukan bunga jantan dan betina menjadi lebih lama, dan pemunculan bunga betina lebih lama dibanding bunga jantan.
Parameter keragaan tongkol berkorelasi nyata dengan parameter penutupan klobot, dimana diketahui bahwa semakin bagus penutupan klobot pada tongkol maka semakin sulit mikroorganisme dari luar merusak tongkol dan biji tanaman. Bobot tongkol kupasan menunjukkan hasil korelasi yang sangat nyata dengan penampakan tongkol (ear aspect), semakin bagus ear aspect dalam hal ini semakin besar dan semua biji utuh dan tidah terserang hama penyakit, maka semakin tinggi pula berat tongkol kupasannya.
Bobot tongkol kupasan pada tabel berkorelasi langsung dengan rendemen hasil. Produksi berkorelasi dengan keragaan tongkol, bobot tongkol kupasan dan kadar air tongkol oada kondisi optimal di Sulawesi Selatan. Tabel 8 menunjukkan analisis korelasi antar peubah pada lingkungan cekaman (Sulawesi), dapat dilihat bahawa hasil berkorelasi sangat nyata hampir dengan hampir semua parameter pengamatan. Umur berbunga jantan dan anthesis silking interval berkorelasi sangat nyata dengan hasil. Adanya cekaman kekeringan yang diberikan pada awal fase pembungaan akan sangat berpengaruh kepada umur munculnya bunga jantan dan betina serta interval muncul keduanya, karena pada waktu tersebut air sangat dibutuhkan untuk proses fotosintesis pembentukan bahan dan energi untuk memasuki fase reprroduksi pembentukan bunga dan pengisian biji. Pengaruh cekaman kekeringan terhadap tinggi tanaman.
Cekaman air atau cekaman kekeringan dapat mengurangi atau menekan pertumbuhan tinggi tanaman (Henry 2013; Jatoi et al. 2014). Pada kondisi cekaman kekeringan, tanaman jagung mengalami kelayuan atau penggulungan daun yang merupakan gejala kritis tanaman dan defisit air di manalaju kehilangan air melalui transpirasi lebih besar dibanding laju absorbsi air oleh akar (Banziger
19
Angka-angka pada kolom yang sama diikuti oleh *berkorelasi nyata pada α=0.05, **berkorelasi sangat nyata pada α=0.01, TT = tinggi tanaman, TK = tinggi letak tongkol,
RTTTK = rasio tinggi tanaman dan tinggi tongkol, UBJ = umur berbunga jantan, UBB = umur berbunga betina, ASI = anthesis silking nterval, PA = plant aspect, HC = penutupan klobot, EA= ears aspect, BTKP = bobot tongkol kupasan, REND = rendemen, KA = kadar air, Y= produksi, B1000BJ =bobot seribu biji.
Tabel 7 Korelasi antar peubah tanaman jagung pada kondisi optimal Sulawesi Selatan
TT TK RTTTK UBJ UBB ASI PA HC EA BTKP REND KA Y B1000BJ
Angka-angka pada kolom yang sama diikuti oleh *berkorelasi nyata pada α =0.05, **berkorelasi sangat nyata pada α=0.01; TT = tinggi tanaman, TK = tinggi letak tongkol, RTTTK = rasio tinggi tanaman dan tinggi tongkol, UBJ=umur berbunga jantan, UBB=bunga berbunga betina, ASI = anthesis silking nterval, PA=plant aspect, HC=penutupan
klobot, EA= ears aspect, BTKP = bobot tongkol kupasan, REND = rendemen, KA = kadar air, YIELD= produksi, B1000BJ=bobot seribu biji.
Tabel 8 Korelasi antar peubah tanaman jagung pada kondisi suboptimal (cekaman kekeringan) Sulawesi Selatan
20
Tabel 9 Korelasi antar peubah tanaman jagung pada kondisi optimal Kalimantan Selatan
TT TK RTTTK UBJ UBB ASI PA HC EA BTKP REND KA Y B1000BJ
TT -1.00
TK -0.33** -1.00
RTTTK -0.07 -0.96** -1.00
UBJ -0.05 -0.06 -0.05 -1.00
UBB -0.07 -0.07 -0.05 -0.94** -1.00
ASI -0.08 -0.05 -0.03 -0.23** 0.55** -1.00
PA -0.06 -0.01 -0.01 -0.03 -0.06 -0.10 -1.00
HC -0.01 -0.05 -0.05 -0.02 -0.03 -0.02 -0.02 -1.00
EA -0.09 -0.04 -0.02 -0.02 -0.02 -0.02 -0.03 -0.55** -1.00
BTKP -0.04 -0.07 -0.06 -0.08 -0.09 -0.05 -0.04 -0.02 -0.64** 1.00
REND -0.06 -0.00 -0.01 -0.05 -0.08 -0.10 -0.09 -0.06 -0.18** 0.30** 1.00
KA -0.04 -0.01 -0.02 -0.02 -0.06 -0.14* -0.10 -0.01 -0.07 0.02 0.01 -1.00
YIELD -0.02 -0.07 -0.07 -0.08 -0.09 -0.05 -0.04 -0.02 -0.62** 0.99** 0.37** -0.07 1.00
B1000BJ -0.04 -0.07 -0.06 -0.03 -0.05 -0.06 -0.06 -0.03 -0.60** 0.90** 0.27** -0.01 0.90** 1.00
21
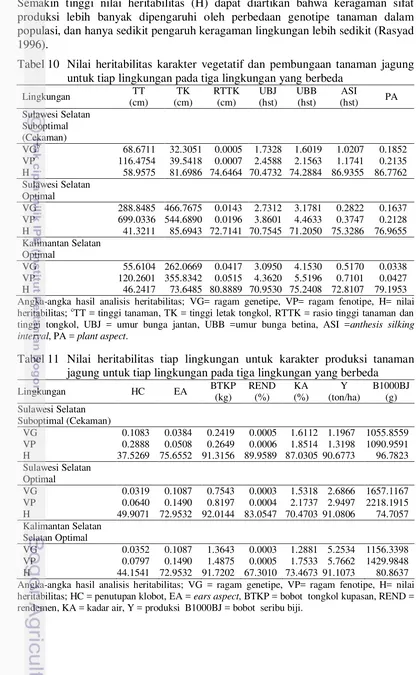
Heritabilitas dalam arti luas (H) merupakan dugaan yang bermanfaat untuk mengitung nilai estimasi kemajuan seleksi untuk suatu sifat di dalam populasi. Semakin tinggi nilai heritabilitas (H) dapat diartikan bahwa keragaman sifat produksi lebih banyak dipengaruhi oleh perbedaan genotipe tanaman dalam populasi, dan hanya sedikit pengaruh keragaman lingkungan lebih sedikit (Rasyad 1996).
Tabel 10 Nilai heritabilitas karakter vegetatif dan pembungaan tanaman jagung untuk tiap lingkungan pada tiga lingkungan yang berbeda
Lingkungan TT
Angka-angka hasil analisis heritabilitas; VG= ragam genetipe, VP= ragam fenotipe, H= nilai heritabilitas; cTT = tinggi tanaman, TK = tinggi letak tongkol, RTTK = rasio tinggi tanaman dan tinggi tongkol, UBJ = umur bunga jantan, UBB =umur bunga betina, ASI =anthesis silking
interval, PA = plant aspect.
Tabel 11 Nilai heritabilitas tiap lingkungan untuk karakter produksi tanaman jagung untuk tiap lingkungan pada tiga lingkungan yang berbeda
22
Karakter tinggi tanaman dan husk cover pada analisis heritabilitas antar lingkungan (Tabel 10-11) menunjukkan nilai heritabilitas sedang. Sedangkan untuk karakter ringgi tongkol, rasio tingi tanggi tongkol terhadap tinggi tanaman, umur berbunga jantan, umur berbunga betina, anthesis silking interval, plant aspect, ear aspect, bobot tongkol kupasan, rendemen, produksi dan, bobot seribu biji memiliki nilai heritabilitas tinggi. Nilai heritabilitas menurut Kramer (1980) dibagi menjadi tiga yakni: tinggi (>70%), sedang (30%-70%), rendah (<30%). Nilai heritabilitas yang tinggi mengindikasikan bahwa karakter tersebut lebih banyak dipengaruhi oleh genetiknya dibandingkan dengan lingkungan, sebaliknya jika nilai heritabilitas rendah berarti faktor lingkungan lebih mempengaruhi.
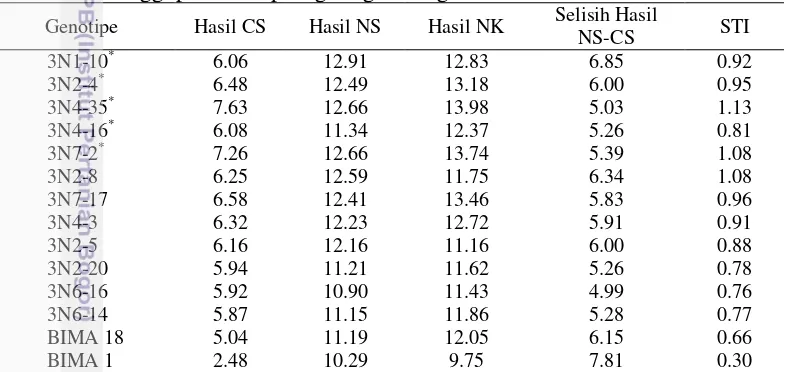
Setelah melakukan beberapa analisis, maka diperoleh dua belas genotipe dengan hasil tertinggi berdasarkan intensitas seleksi 15%, yang kemudian seleksi dipersempit ke taraf 10% sehingga diperoleh lima genotipe terbaik dengan hasil tertinggi pada ketiga lingkungan penanaman. Kelima genotipe tersebut dapat berproduksi dengan baik pada tiga lingkungan yang diberikan, bahkan kelimanya dapat berproduksi dengan baik melebihi varietas pembandingnya yakni Bima 1 dan Bima 18 (Tabel 10).
Tabel 12 Genotipe jagung terbaik berdasarkan parameter produksi (ton ha-1) tertinggi pada setiap lingkungan dengan intensitas seleksi 15%
Genotipe Hasil CS Hasil NS Hasil NK Selisih Hasil
Angka-angka hasil seleksi dari genotipe terbaik; CS=suboptimal (cekaman) Sulawesi, NS= optimal sulawesi Selatan, NK= optimal Kalimantan Selatan, STI= stress tolerance index, dan *= lima genotipe terbaik hasil seleksi dengan intensitas seleksi 10 %.
Hal ini berarti genotipe yang ditanam berproduksi dengan baik, bahkan lebih baik dari varietas pembandingnya, baik pada lingkungan optimal maupun lingkungan suboptimal (cekaman kekeringan). Hal ini sesuai dengan pendapat Widyasari et al. (2004) yang menyatakan bahwa suatu tanaman dikatakan toleran terhadap kondisi suboptimal (cekaman kekeringan) apabila tanaman tersebut tetap dapat berproduksi dengan baik walaupun ditumbuhkan pada lingkungan yang bercekaman.
23
111 genotipe moderat kekeringan, dan 116 genotipe peka kekeringan (Gambar 5), Berdasarkan pendapat Fisher (1997) yang menyatakan bahwa nilai STI dikategorikan menjadi 3 kategori yakni toleran (>1), moderat (0.5-1), peka (<0.5).
Gambar 5 Diagram batang jumlah genotipe jagung pada setiap kategori nilai stress tolerance index
Analisis Gabungan Antar Lingkungan
Pengujian dilakukan dengan menanam semua genotipe pada tiga lingkungan yang berbeda, kemudian datanya dianalisis dengan mengabungkan semuanya sehingga dapat memudahkan dalam melakukan evaluasi interaksi genotipe x lingkungan (GxE) pada populasi tersebut
Tabel 13 Nilai kuadrat tengah lingkungan, ulangan (lingkungan), genotipe, genotipe x lingkungan untuk semua karakter pengamatan tanaman jagung pada gabungan tiga lingkungan yang berbeda
Karakter Lingkungan Ulangan Genotipe G x E KK
B1000BJ 2390502.97** 3599.83** 3065.02** 1599.19** 9.280
Angka-angka pada kolom yang sama diikuti oleh **=berpengaruh sangat nyata pada taraf 1% *= berpengaruh nyata pada taraf 5% tn=tidak berpengaruh nyata; TT=tinggi tanaman, TK=tinggi tongkol, RTTTK=rasio tinggi tanaman dan tinggi tongkol, UBJ=umur berbunga jantan, UBB=umur berbunga betina, ASI=anthesis silking interval, PA= plant aspect, HC= husk cover,
EA= ears aspect, BTKP=bobot tongkol kupasan, REND=rendemen, KA=kadar air, Y=produksi,
24
Pada Tabel 13 hasil analisis ragam gabungan menunjukkan bahwa pengaruh genotipe dan interaksi GxE sangat nyata terhadap semua peubah yang diamati. Karena pengaruh interaksi GxE sangat nyata, maka seleksi genotipe terpilih dilakukan bukan berdasarkan rataan produksi gabungan, melainkan berdasarkan hasil rangking terpisah dari tiga lingkungan yang berbeda.
Tabel 14 Nilai heritabilitas untuk gabungan antar lingkungan pada pengamatan semua karakter tanaman jagung
B1000BJ 244.3048 484.4605 510.8371 47.82 Sedang
Angka-angka hasil analisis heritabilitas; VE =ragam lingkungan, VG =ragam genetik, VGxE = ragam interaksi genetik x lingkungan, VP = ragam fenotip, SD= standar deviasi, H = nilai heritabitas; TT=tinggi tanaman, TK=tinggi tongkol, RTTTK=rasio tinggi tanaman dan tinggi tongkol, UBJ=umur berbunga jantan, UBB=umur berbunga betina, ASI=anthesis silking interval,
PA= plant aspect, HC= husk cover, EA= ear aspect, BTKP=bobot tongkol kupasan,
REND=rendemen, KA=kadar air, Y=produksi , B1000BJ=Bobot 1000 biji
Heribilitas merupakan suatu tolak ukur yang digunakan dalam suatu seleksi, yaitu untuk mengetahui kemampuan tetua dalam menurunkan kesamaan sifat kepada keturunannya (Johnson et al. 2003). Lingkungan tumbuh tanaman tidak selalu merupakan lingkungan yang optimal bagi pertumbuhan sehingga seringkali tanaman tidak mampu mengekspresikan sifat-sifat genetiknya. Sifat genetik tanaman yang terekspresikan melalui fenotipiknya dipengaruhi oleh faktor genetik, lingkungan, dan interaksi antar faktor genetik dan lingkungan (Sutoro et al. 1989). Nilai heritabilitas harus diperhatikan dalam menyeleksi karakter tanaman, dimana sangat penting dalam menentukan metode seleksi dan pada generasi mana sebaiknya karakter yang diinginkan diseleksi (Azizian 2014).
25
pengaruh lingkungan kecil, sehingga faktor genetik lebih dominan dalam penampilan genetik tanaman. Nilai heritabilitas tinggi akan lebih meningkatkan keberhasilan seleksi (Azrai 2006).
Tabel 2 Rataan pada pengamatan parameter vegetatif dan pembungaan untuk masing-masing genotipe jagung terbaik
rataan umum+BNT 0.05, - =lebih kecil dari rataan umum-BNT 0.05
Pada Tabel 15-16 menyajikan data hasil perhitungan rataan genotipe-genotipe terbaik pada setiap peubah pengamatan. Pada peubah pengamatan tinggi tanaman, genotipe 3N6-16 memiliki nilai diataas rataan umum (rataan+BNT 0.05). Tinggi tongkol pada genotipe 3N2-4, 3N4-35, 3N4-16, dan 3N7-17 lebih besar dibandingkan dengan rataan umumnya. Pada peubah rasio tinggi tanaman tinggi tongkol, dari semua genotipe tidak ada yang menunjukkan nilai yang lebih atau kurang dari nilai rataan umum, hal yang sama juga pada umur berbunga jantan, umur berbunga betina, dan plant aspect. Pada peubah anthesis silking interval, genotipe 3N6-14 memiliki nilai lebih rendah dibandingkan dengan nilai rataan umum.
26
Banziger et al. (2000) kadar air yang rendah akan lebih baik dikarenakan dapat lebih tahan dengan penyimpanan. Hasil dan bobot seribu biji yang lebih tinggi dibandingkan nilai rataan umum (3N4-35 dan 3N7-2) merupakan salah satu tujuan yang ingin dicapai, sehingga dapat terlihat genotipe yang mana saja yang tetap dapat berproduksi dengan baik jika ditumbuhkan pada lingkungan yang optimum maupun lingkungan suboptimum
Tabel 16 Rataan pada pengamatan parameter produksi untuk masing-masing genotipe jagung terbaik
Parameter pengamatan meliputi HC=penutupan klobot, EA=ear aspect, BTKP=bobot tongkol kupasan, REND=rendemen, KA=kadar air, Y=produksi, B1000BJ=bobot seribu biji, *=lima genotipe terbaik, + =lebih besar dari hasil rataan umum+BNT 0.05, - =lebih kecil dari rataan umum-BNT 0.05
Tabel 17 Korelasi Spearman antar lingkungan untuk semua karakter pengamatan pada tanaman jagung
Karakter Pengamatan OS-OK SS-OS SS-OK
Tinggi tanaman (cm) Tinggi tongkol (cm)
Rasio tinggi tanaman dan tinggi tongkol (cm) Umur berbunga jantan (hst) Angka-angka pada kolom yang sama diikuti oleh *berkorelasi nyata pada α=0.05, **berkorelasi
sangat nyata pada α=0.01, SS= suboptimal (cekaman) Sulawesi Selatan, OS= optimal Sulawesi SelatanSulawesi, dan OK= optimal Kalimantan Selatan Selatan.
27
tinggi tongkol, bobot tongkol kupasan, rendemen, produksi dan nyata pada karakter tinggi tanaman dan ear aspect. Pada perbandingan lingkungan Sulawesi Selatansuboptimal dan Kalimantan Selatan optimal hanya bebrapa karakter yang menunjukkan hasil berpengaruh sangat nyata yakni karakter bobot tongkol kupasan, produksi, dan bobot seribu biji. Sedangkan yang berpengaruh nyata hanya satu karakter yakni ear aspect (kenampakan tongkol).
Korelasi positif dan nyata menandakan adanya kemiripan ranking genotipe diantara kedua lingkungan untuk karakter yang diamati. Sebaliknya korelasi yang negatif dan nyata mengindikasikan adanya hubungan ranking genotipe yang berlawanan. Korelasi yang positif dan nyata serta sangat nyata antar ketiga lingkungan seperti pada karakter produksi menandakan bahwa pengaruh interaksi GxE, meskipun nyata, diduga tidak menyebabkan perubahan ranking genotipe antar lingkungan secara dramatis.
5 KESIMPULAN DAN SARAN
Kesimpulan yang diperoleh dari hasil penelitian ini adalah:
a. Tingkat produktivitas hasil genotipe-genotipe yang diujikan berada pada kisaran 2-14 ton/ha.
b. Genotipe dengan tingkat produktivitas hasil terbaik pada lingkungan optimal Sulawesi Selatan yakni 3N1-10 sebesar 12.91 ton/ha, lingkungan suboptimal Sulawesi Selatan yakni 3N4-35 sebesar 7.63 ton/ha, lingkungan optimal Kalimantan Selatan yakni 3N4-35 sebesar 13.98 ton/ha.
c. Genotipe dengan produktivitas hasil terbaik pada analisis gabungan lingkungan yakni 3N4-35 sebesar 11.42 ton/ha.
a. Nilai heritabilitas menunjukkan hasil yang bervariasi mulai dari rendah-tinggi, baik pada analisis lingkungan yang berbeda (optimal dan suboptimal) maupun pada analisis lingkungan gabungan.
b. Hasil analisis gabungan menunjukkan adanya varian genetik yang luas, hal ini ditunjukkan dengan adanya interaksi antara genotipe dan lingkungan. 3. Berdasarkan nilai stress tolerance index hasil analisis lingkungan berbeda
dari 230 genotipe yang diujikan, terdapat 3 genotipe dengan kategori sangat toleran terhadap kekeringan yakni 3N4-35, 3N5-33, dan 3N7-2. 111 genotipe dengan kategori toleran moderat kekeringan, serta 116 genotipe dengan kategori peka
Saran
Perlu dilakukan penelitian lebih lanjut terhadap calon tetua dengan sifat– sifat yang terbaik dan terjamin kestabilannya, sehingga dapat digunakan sebagai tetua pada perakitan genotipe jagung terbaru yang toleran terhadap cekaman kekeringan.
1
28
DAFTAR PUSTAKA
Abdelmoneim TS, Mousa T, Abdelbagi I. 2014. Increasing plant tolerance to drought stress. Life Science J. 11(1): 10-17.
Al Badeiry NA, Al-Saadi AH, Merza TK. 2014. Analysis of genetic diversity in maize (Zea mays L.) varieties using simple sequence repeat (SSR) Markers. Journal of Pure and Applied Sciences. 6(22): 1768-1774.
Ahadiyat YR, Hidayat P, Susanto U. 2014. Drought tolerance, phosphorus efficiency and yield characters of upland rice lines. Food Agric. 26 (1): 25-34.
Anisa. 2011. Dampak cekaman kekeringan terhadap pertumbuhan, hasil, dan kandungan total prolina dan cabai. Jurnal Ilmiah Agrista 12: 19-27.
Arve HJ, Hanike LH, Abdullah. 2011. Eksplorasi plasma nutfah jagung di kawasan timur Indonesia. Mimbar Akademik, Universitas Haluoleo Kendari (ID) Edisi No. 8. Th VIII.
Azrai, M. 2006. Ragam interaksi genotip x lingkungan untuk infeksi penyakit bulai pada beberapa jagung koleksi Balitsereal. Agrivita. 28 (1): 45-53. Azizian A. 2014. Maize response to water levels. Int J of Plant Production 8(1):
131-162.
Badan Pusat Statistik Nasional. 2014. Produksi Pangan Tahun 2014. 21
Desember 2015; Bogor (ID). Tersedia pada:
http://www.bps.go.id/brs/view/id/1122.
Baddington, Dearman. 1982. Intergenotypic competition during evaluation of maize progenies under limited and adequate N supply. Field Crops Research. 44: 25-31.
Banziger M, Edmeades GO, Beck D, Bellon M. 1997. Breeding for drought and nitrogen stress tolerance in maize: From theory to practice. 28: 17-21. Bänziger M, Edmeades GO, Lafitte HR. 2000. Selection for drought tolerance
increases maize yields over a range of N levels. Crop Science 39:1035-1040.
Bardwaj, Yadav. 2012. Efficiency of high-nitrogen selection environments for improving maize for low-nitrogen target environments. Crop Science 37:1103-1109.
Blum A. 2002. Drought tolerance is it a complex trait. Field screening for drought tolerance in crop plants with emphasis on rice. Field screening for drought tolerance in crop plants with emphasis on rice. Int. Workshop on Field Screening for Drought Tolerance in Rice. 17–22. Patancheru, India (IND). 11–14 Dec. 2000.
Bray. 1997. Relationship between line and topcross performance under drought and non-stressed conditions in tropical maize. Developing Drought and Low N-Tolerant Maize. Proceedings of a Symposium. 383-386. CIMMYT, El Batán, Mexico (UMS), DF: CIMMYT. . March 25-29, 1997.
Close. 1997. Value of selection for osmotic potential in tropical maize. Agronomy Journal. 83:948-956.
29
Dahlan M. 2001. Pemuliaan tanaman untuk ketahanan terhadap kekeringan. Dalam Prosiding International Conference on Agricultural Development NTT. Timor Timur and Maluku Tenggara (ID). 11-15 Desember 2001. Dallaire SM, Houde Y, Gagne HS, Saini S, Boileau N, Chevrier, Sarhan F. 1994.
ABA and Low Temperature Induce Freezing Tolerance via Distinct Regulatory Patways in Wheat. Plant Cell Physiol. London (GB). Preager. 35 (1) : 1-9.
Edmeades GO, Bolanos J, Lafitte HR. 1992. Program in breeding for drought tolerance in maize. Proceeding of the 47th annual corn and sorghum industry. Research conference ASTA. 17 (1): 5-12.
Earl, Davis. 2003. Engineering Drought Tolerance in Plants : Discovering and Tailoring Genes to Unlock The Future. Cur. Op. in Biotechnol. 17: 113-122. Efendi. 2014. System analysis of plant traits to increase grain yield on limited
water supplies. Agron J . 93: 263-270.
Ekmekei. 2005. A breeding procedure designed to make maximum use of both general and specific combining ability. Agronomy Journal. 4: 360-367. Farooq TK, Alimuddin HN, Saipul FE. 2009. Selection improves drought
tolerance in tropical maize populations: II. Direct and correlated responses among secondary traits. Crop Science, 39(5): 1315-1324.
Fisher. 1997. Effective selection criteria for assesing stress tolerance. Proceeding of the international symposium on adaptation of vegetables and other food crops in temperature and water stress. AVRDC Publication. Tainan, Taiwan. Flirricks, Andonio, Selena. 2012. The influence of time lag between pollen
shedding and silking on the yield of maize. South African Journal of Agricultural Science. 10: 667-674.
Fukai S, Cooper M. 1995. Development of drought-resistant cultivars using physio-morphological traits in rice. Field Crops Research. 40: 67-86.
Gomez, K.A. dan Gomez A.A. (1995). Prosedur Statistik untuk Penelitian Pertanian.
hal :13 – 16, Edisi Kedua. Jakarta (ID): UI Pr.
Hallauer, A.R. 1975. Hybrid development and population improvement in maize by reciprocal full-sib selections. Egyptian Journal of Genetics and Cytology. 2: 84 101.
Henry A. 2013. Drought stress research in rice with emphasis on roots: accomplishments over the last 50 years. Plant Root. 7: 5-19.
Herawati T, Setiamihardja R. 2000. Pemuliaan Tanaman. Departemen Pertanian RI dengan Fakultas Pertanian Universitas Padjajaran, Jatinangor, Bandung (ID). Pajajaran Pr.
International Maize and Wheat Improvement Center (CIMMYT). 2000.
Managing trials and reporting data for CIMMYT‟s international maize testing program383-386. El Batán, Mexico (UMS), DF: CIMMYT.
Jatoi WA, Baloch M, Gul S. 2014. Heterosis for yield and traits in wheat under water stress conditions. J of An and Plant Sci. 24(1): 252-261.
Jones MM, Tumer NC and Osmond CB. 1981. Mechanism of Drought Resistance PP 15-53 in Paleg LG, and Aspinall (eds). The Physiology and Biochemistry of Drought Resistance in Plants. New York (US): Academic Pr.