KERAGAMAN GENETIK DAN MIKROEVOLUSI
Pyricularia grisea ASAL RUMPUT
SRI LISTIYOWATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi berjudul “Keragaman Genetik dan Mikroevolusi Pyricularia grisea Asal Rumput” adalah karya bersama saya dengan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini
Bogor, Februari 2012
ABSTRACT
SRI LISTIYOWATI. Genetic Diversity and Microevolution of Pyricularia grisea
from Grasses. Supervised by UTUT WIDYASTUTI, GAYUH RAHAYU, and ALEX HARTANA.
Pyricularia is the fungus caused rice and grasses blast disease. They are morphologically indistinguishable, therefore their specific epithet is often based on host. The concept of Pyricularia species using nucleotide sequences of current molecular approach is still on controversy, especially those infected weed grasses (Pyricularia grisea) and rice (Pyricularia oryzae). Yet genetic change caused by host alteration has not been explored. Therefore, this researchs were aimed to (i) study the populations diversity of Pyricularia from various grasses (Cynodon dactylon, Digitaria ciliaris, Eleusine indica, and Panicum repens) using five markers that consist of three kinds of sequence characterized amplified region (SCAR), i.e Cut1, PWL2 and Erg2 and two markers, magB and magC, repectively; (ii) to analyze genetic diversity of the basal samples of Pyricularia
from Digitaria ciliaris and its microevolution of Pyricularia d4 from D. ciliaris
following cross infection to rice and Panicum repens by adding markers of amplified fragment length polymorphism (AFLP), PCR of repetitive Pot2 (rep-Pot2), pathotype, and sequences of ITS and 5.8S rDNA nucleus; (iii) study phylogenetic relationship of Pyricularia all of from D. ciliaris (d4), P. repens
(pr10.a.S4), and from rice pathogen (ok6, ou6.S4) based sequences of ITS and 5.8S rDNA. This study used Pyricularia from grasses and rice from West Java that occurs in the same field as a model. The result showed DNA size of all markers of Cut1, PWL2 and Erg2, magB, and magC of Pyricularia from grasses were similar to that of rice origin. The frequency of those SCAR markers, i.e. Cut1, PWL2, and Erg2 were 78.1%, 54.1% and 95.1%, respectively. Based on present SCAR markers, they could be classified into five phenotypes (011, 101, 111, 001, 010) out of the eight possible groups. Cross infection of Pyricularia d4 from D. ciliaris grass to rice could induce genetic variation in their Cut1 and PWL2 markers, AFLP and rep-Pot2 phenotypes, as well as pathotype. On the other hand, no new variation revealed their ITS sequences. Moreover, the cross infection to another grass in different genus (P. repens) also caused genetic variation in AFLP and rep-Pot2 phenotypes, and ITS sequences. These results indicated that the cross infection might induce microevolution of Pyricularia d4. Sequences of ITS and 5.8S rDNA of Pyricularia all of from D. ciliaris (d4), and from rice pathogen (ok6, ou6.S4) were similar. Whereas those of Pyricularia from
P. repens (pr10.a.S4) had only one nucleotide different in their 5.8S rDNA sequences to those of Pyricularia d4. BLAST analysis of ITS and 5.8S rDNA sequences from five isolates showed 99% identical to Magnaporthe oryzae
pathogen of rice, cereals and cultivated grasses. The ITS sequences of the non-rice isolates were similar with isolates of the rice blast disease, so there might be gene flow among the pathogen on grasses and rice. Based on nomenclatural priority, the specific name for Pyricularia from D. ciliaris, P. repens, and rice were
Pyricularia grisea as the anamorphic of Magnaporthe grisea.
RINGKASAN
SRI LISTIYOWATI. Keragaman Genetik dan Mikroevolusi
Pyriculariagrisea Asal Rumput. Dibimbing oleh UTUT WIDYASTUTI, GAYUH
RAHAYU, dan ALEX HARTANA.
Rumput yang tumbuh di sekitar pertanaman padi telah dilaporkan sebagai inang Pyricularia. Pyricularia penginfeksi rumput memiliki keragaman genetik berbeda terhadap Pyricularia yang menginfeksi padi. Sampai saat ini analisis keragaman genetik Pyricularia dari rumput yang mengalami pergantian inang pada genus berbeda belum pernah dilaporkan. Pergantian inang atau perubahan patogenisitas diduga dapat menjadi faktor pendorong mikroevolusi pada
Pyricularia. Mikroevolusi merupakan perubahan yang dapat tampak dalam waktu relatif pendek (beberapa hari atau minggu) pada mikrob. Mikroevolusi
Pyricularia yang mengalami pergantian genus inang juga belum pernah diteliti. Spesies epitet grisea dari rumput dan oryzae dari padi ditetapkan berdasarkan inang. Kedua spesies tidak berbeda morfologinya. Berkembangnya berbagai pendekatan spesies, menyebabkan konsep spesies Pyricularia belum disetujui bersama. Couch dan Kohn pada tahun 2002 menempatkan Pyricularia yang patogen pada Digitaria sanguinalis sebagai Pyricularia grisea, yaitu sama seperti prinsip penamaan sebelumnya. Isolat-isolat Pyricularia dari rumput dan padi yang diperoleh dari ladang dan sawah di Jawa Barat menjadi model pada penelitian ini. Hasil penelitian ini diharapkan dapat berkontribusi dalam bentuk informasi pada manajemen penyakit blas pada padi di Jawa Barat.
Penelitian ini bertujuan menganalisis (i) keragaman populasi Pyricularia
dari rumput (Cydonon dactylon, Eleusine indica, Digitaria ciliaris, dan Panicum repens) berdasarkan lima penanda yang terdiri atas tiga penanda (Cut1, PWL2 dan Erg2) sequence characterized ampllified region (SCAR), dan dua penanda berupa magB, dan magC, (ii) keragaman genetik sampel basal Pyricularia dari rumput
Digitaria ciliaris dan mikroevolusi Pyricularia d4 yang berasal dari Digitaria ciliaris sebagai hasil induksi pergantian inang ke padi dan ke rumput Panicum repens berdasarkan lima penanda, yaitu SCAR, amplified fragment length polymorphism (AFLP), PCR pada repetitive Pot2 (rep-Pot2), dan hubungannya dengan ras fisiologi, serta sekuen ITS beserta 5.8S rDNA nukleus yang dihasilkan melalui amplifikasi dengan primer universal berupa its1 dan its4; (iii) hubungan filogenetik Pyricularia dari rumput dan padi berdasarkan sekuen ITS beserta 5.8S rDNA nukleus. Sekuen tersebut juga dihasilkan melalui amplifikasi dengan primer universal (its1 dan its4). Penelitian ini diharapkan dapat mengungkapkan peran pergantian inang pada perubahan genetik yang merupakan bagian dari mikroevolusi Pyricularia, dan hubungan filogenetik antara Pyricularia yang menginfeksi rumput dan padi.
rumput hanya memiliki lima fenotipe SCAR berdasarkan delapan kemungkinan fenotipe SCAR yang terdiri atas Cut1, PWL2 dan Erg2. Kelima fenotipe SCAR memiliki frekuensi yang berbeda, yaitu 19.5% berfenotipe 011, 41.5% berfenotipe 101, 31.7% berfenotipe 111, 2.5% berfenotipe 001, dan 4.9% berfenotipe 010. Populasi Pyricularia dari rumput memiliki 78.1% Cut1, 54.1% PWL2 dan 95.1% memiliki Erg2.
Riwayat perubahan genetik Pyricularia d4 yang mengalami pergantian inang digunakan sebagai model pada analisis mikroevolusinya. Sumber inokulum (suspensi konidium 104 - 105 mL-1) diinjeksikan ke pelepah daun inang pengganti tahap ke-1 (tiga varietas padi, rumput Cynodon dactylon, Digitaria sp., Ottochloa nodosa, dan Panicum repens). Pada inokulasi tahap ke-2, hanya turunan d4 hasil infeksi ke padi tahap ke-1 yang diinokulasikan silang ke varietas padi yang berbeda. Isolat d4 hanya mampu berganti inang ke padi var. Kencana bali, Cisokan, dan rumput P. repens. Isolat d4 yang berganti inang ke Kencana bali dan Cisokan mengalami perubahan fenotipe SCAR (Cut1 dan PWL2), AFLP dan rep-Pot2, serta ras fisiologinya, tetapi tidak mengalami perubahan sekuen ITS. Sebaliknya d4 berganti inang ke P. repens tidak mengalami perubahan Cut1, PWL2, dan ras fisiologinya, tetapi mengalami perubahan fenotipe AFLP dan rep-Pot2, serta mutasi transisi dua nukleotida ITS. Selain itu, d4 yang berganti inang ke P. repens mengalami tingkat perubahan fenotipe AFLP lebih besar dan perubahan fenotipe rep-Pot2 juga berbeda daripada d4 yang berganti inang ke Kencana bali dan Cisokan. Fenotipe rep-Pot2 dari d4 pada Kencana bali maupun Cisokan adalah sama. Sebaliknya, isolat d4 yang berganti inang ke Kencana bali memiliki tingkat perubahan fenotipe AFLP yang lebih tinggi, namun tingkat perubahan ras fisiologinya (023) dapat lebih rendah daripada d4 yang berganti inang ke Cisokan. Ras fisiologi d4 pada Cisokan dapat berubah menjadi 373, tetapi turunannya juga dapat tidak mengalami perubahan (000), masih seperti ras fisiologi inokulumnya (d4). Isolat d4 yang telah berganti inang ke padi tidak mengalami perubahan fenotipe SCAR, AFLP, rep-Pot2, dan sekuen ITS setelah berganti inang ke varietas padi yang sama. Sebaliknya, d4 yang telah berganti inang ke padi Cisokan mengalami perubahan fenotipe AFLP yang lebih tinggi ketika kemudian berganti inang ke padi Kencana bali.
Pyricularia dari D. ciliaris (d4), dan padi (ok6 dan ou6.S4) memiliki kesamaan sekuen ITS beserta 5.8S rDNA. Sedangkan Pyricularia dari P. repens
(pr10.a.S4) memiliki perbedaan satu nukleotida terhadap d4, ok6, atau ou6.S4. Hasil BLAST sekuen ITS beserta 5.8S rDNA dari d4 atau pr10.a.S4 menunjukkan sangat identik (99%) dengan Magnaporthe oryzae dari padi. Selain itu, sekuen ITS beserta 5.8S rDNA dari keempat isolat pada penelitian ini menunjukkan hubungan filogenetik sangat dekat dengan M. oryzae dari serealia lain. Sekuen ITS beserta 5.8S rDNA tidak mampu memisahkan isolat Pyricularia dari rumput dan padi. Walaupun demikian, berdasarkan azas prioritas maka Pyricularia dari rumput (d4 dan pr10.a.S4) lebih tepat dinamakan Pyricularia grisea dengan
Magnaporthegrisea sebagai teleomorfnya. Sedangkan Pyricularia dari padi (ok6 dan ou6.S4) berdasarkan prinsip prioritas juga dapat sebagai P. grisea.
© Hak Cipta milik Institut Pertanian Bogor, tahun 2012 Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumbernya.
a. pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah.
b. dan pengutipan tidak merugikan kepentingan yang wajar IPB.
KERAGAMAN GENETIK DAN MIKROEVOLUSI
Pyricularia grisea ASAL RUMPUT
SRI LISTIYOWATI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup : Dr. Iman Hidayat
Dr.Ir. Rika Raffiudin, MSi.
Judul Disertasi : Keragaman Genetik dan Mikroevolusi Pyricularia grisea Asal Rumput
Nama : Sri Listiyowati
NIM : G361030011
Disetujui Komisi Pembimbing
Dr.Ir. Utut Widyastuti, M.Si. Ketua
Dr.Ir. Gayuh Rahayu, M.S. Prof.Dr.Ir. Alex Hartana, M.Sc.
Anggota Anggota
Diketahui
Ketua Program Studi Biologi Dekan Sekolah Pascasarjana
Dr.Ir. Dedy Duryadi Solihin, DEA. Dr.Ir. Dahrul Syah, MSc.Agr
PRAKATA
Alhamdulillahirobbil alamin, penulis lafadkan atas segala rahmat dan karunia dari ALLAH SWT yang telah diberikan kepada penulis selama melakukan penelitian hingga penyelesaian penulisan disertasi ini. Disertasi yang berjudul Keragaman Genetik dan Mikroevolusi Pyricularia grisea Asal Rumput memuat hasil penelitian tentang keragaman cendawan blas pada beberapa spesies rumput yang tumbuh di sekitar pertanaman padi, dan perubahan genetik cendawan blas yang mengalami pergantian inang secara buatan. serta hubungan filogenetik antara cendawan blas pada rumput Digitaria ciliaris,
Panicum repens, dan padi. Hasil yang diperoleh diharapkan dapat berkontribusi informasi pada manajemen penyakit blas pada padi di Jawa Barat. Bagian dari disertasi ini telah disajikan pada Seminar Sains III di Bogor pada bulan November 2010 yang diselenggarakan oleh FMIPA IPB dengan MIPAnet, dengan judul Keragaman Genetik Hasil Pergantian Inang Pyricularia grisea Asal Rumput
Digitaria ciliaris. Selain itu sebuah artikel telah dipublikasikan di Jurnal Microbiology Indonesia 2011 berjudul Diversity of SCAR Markers of Pyricularia grisea Isolated from Digitaria ciliaris Following Cross Infection to Rice, 5(1):1-8. Melalui tulisan ini, penulis menyampaikan terima kasih dan penghargaan yang sebesar-besarnya kepada Dr.Ir. Utut Widyastuti, M.Si. selaku ketua komisi pembimbing, Dr.Ir. Gayuh Rahayu, MS., Prof.Dr.Ir. Alex Hartana, M.Sc. selaku anggota komisi pembimbing, atas segala curahan waktu, pikiran, nasihat dan arahan selama penelitian dan penulisan hasil disertasi. Terimakasih kepada bapak Dr.Ir. Muhammad Jusuf (Alm) atas bimbingan serta nasihat kepada penulis. Penulis juga menyampaikan terima kasih kepada seluruh jajaran pimpinan Institut Pertanian Bogor, yaitu Rektor yang telah mengizinkan penulis melanjutkan studi program doktor. Dekan Sekolah Pascasarjana IPB, Dr.Ir. Dedy Duryadi Solihin, DEA selaku Ketua Program Studi Biologi Sekolah Pascasarjana, Ketua Departemen Biologi FMIPA, dan Direktur Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) yang telah memfasilitasi studi S3 ini.
Penulis mengucapkan terima kasih kepada Direktorat Jenderal Pendidikan Tinggi (DIKTI) Kementrian Pendidikan Nasional Republik Indonesia yang telah membiayai pendidikan S3 melalui Beasiswa Pendidikan Pasca Sarjana (BPPS), Lembaga Penelitian dan Pengabdian kepada Masyarakat (LPPM) IPB yang telah memberikan dana penelitian dosen muda IPB pada tahun 2005 dengan judul, Cendawan Pyricularia grisea pada Padi dan Rumput di Sawah Sukabumi, dan pada tahun 2008 membiayai penelitian strategis dengan judul, Hubungan Kemampuan Pergantian Inang dengan Plastisitas Genetik pada Cendawan Blas Padi (SPK/Kontrak No 08/13.24.4/SPK/BG/2008). Penelitian ini juga dibiayai oleh Hibah Penelitian Mahasiswa Program Doktor IPB dari Direktorat Jenderal Pendidikan Tinggi (DIKTI) Kementrian Pendidikan Nasional Republik Indonesia tahun 2009, serta sebagian proyek IMHERE IPB atas nama Dr.Ir. Utut Widyastuti, M.Si.
sebagai penguji pada ujian terbuka pada 27 Januari 2012. Ucapan terima kasih juga penulis sampaikan kepada:
1. Dra. Anggiani Nasution (Kebun Percobaan Muara, Balai Besar Penelitian Tanaman Padi Bogor) yang telah menganalisis ras fisiologi isolat hasil pergantian inang
2. Dra. Alex Sumadijaya (Pusat Penelitian Biologi LIPI Bogor) yang telah membantu mengidentifikasi rumput
3. Syarifin Firdaus, M.Si; Muhammad Bahrelfi Belafif, M.Si.; Rida Oktorida M.Si.; Dr. Yaya Rukayadi M.Si; atas bantuan jurnal-jurnal yang diperlukan pada penelitian ini
4. Ika Atifah Zahroh S.Si; Ika Madona Pandia S.Si; Pepi Elvavina; Dwi Ambarwati S.Si dan Moch Rachmad Hidayat S.Si yang telah membantu pada penelitian ini; kepada Iqbal Kusnandarsyah S.Si yang membantu secara teknis pada kelengkapan penulisan disertasi
5. Rekan sejawat bagian Mikologi, yaitu Ir. Agustin W. Gunawan, MS.; Prof. Dr. Okky S. Dharmaputra, Dr.Ir. Lisdar M. Sudirman, Dr.Ir. Nampiah Sukarno yang telah memberikan dukungan dan perhatian kepada penulis selama masa studi
6. Rekan sejawat Dept. Biologi FMIPA IPB, yaitu Taruni Sri Prawasti, M.Si. atas dukungan moril dalam suka dan duka penulis, Dr.Ir. Miftahudin, Dr.Ir. Achmad Farajallah, M.Si., Dr.Ir. R.R Dyah Perwitasari, dan semua rekan staf pengajar Departemen Biologi FMIPA IPB atas segala bantuan, diskusi, dan dukungannya
7. Teman-teman seperjuangan: Dr Muzuni, MSi, Dr Dewi Indriyani MSi, Saleha Hannum, MSi, Yohana C Sulistianingsih, MSi; Ulung Anggito, M.Si, serta kepada para teknisi ibu Dewi, pak Edi Djaenudin, pak Mulya, pak Kusnadi, pak Adi, pak Yanto.
Penulis sampaikan terima kasih yang tulus ikhlas kepada suami atas segala pengertian, kesabaran, dan dukungannya sehingga penulis mampu melewati semuanya sampai selesai. Penulis menyampaikan permohonan maaf sebesar-besarnya kepada putriku dan suami atas segala waktu, tenaga dan pikiran yang banyak tersita untuk penelitian dan penyelesaian studi S3 ini. Kepada kedua orang tua dan adik-adik, penulis mengucapkan terima kasih atas do’a yang senantiasa dipanjatkan untuk kesehatan dan keberhasilan penulis.
Akhirnya penulis berharap tulisan ini dapat memberikan manfaat dan sumbangan bagi kemajuan ilmu pengetahuan biologi di Indonesia.
Bogor, Februari 2012
RIWAYAT HIDUP
Peneliti dilahirkan di Banyuwangi pada tanggal 14 Juli 1964 sebagai anak sulung dari pasangan Gilang Mahrodi dan Sriatun. Pendidikan sarjana ditempuh di Jurusan Biologi, Fakultas Ilmu Pengetahuan Alam (FMIPA) ITB dan meraih gelar sarjana pada tahun 1988. Penulis menjadi staf pengajar di Jurusan Biologi FMIPA IPB mulai tahun 1991, dan melanjutkan pendidikan Magister di Program Studi Biologi pada Program Pascasarjana IPB tahun 1994 dengan mendapat gelar Magister Sains tahun 1998. Kesempatan untuk melanjutkan ke program doktor pada program studi dan pada perguruan tinggi yang sama diperoleh pada tahun 2003 melalui Beasiswa Pendidikan Pascasarjana (BPPS) Ditjen Pendidikan Tinggi (DIKTI).
Selama mengikuti program S3, penulis menjadi anggota Himpunan Mikrobiologi Indonesia. Karya ilmiah berjudul ”Keragaman Genetik Hasil Pergantian Inang Pyricularia grisea Asal Rumput Digitaria ciliaris” telah disajikan pada Seminar Sains III di Bogor pada bulan November 2010 yang diselenggarakan oleh FMIPA IPB dengan MIPAnet. Sebuah artikel telah diterbitkan dengan judul ”Diversity of SCAR Markers of Pyricularia grisea
Isolated from Digitaria ciliaris Following Cross Infection to Rice” pada jurnal Microbiology Indonesia, 2011, 5(1):1-8. Karya-karya ilmiah tersebut merupakan bagian dari program S3 penulis.
Sampai saat ini penulis merupakan staf pengajar di Departemen Biologi, FMIPA IPB. Pada tahun 2006 turut serta dalam penulisan buku yang berjudul Cendawan dalam Praktik Laboratorium yang diterbitkan oleh Bagian Mikologi, Dept. Biologi FMIPA IPB, IPB Press.
DAFTAR ISI
Halaman
DAFTAR TABEL……… xiv
DAFTAR GAMBAR………... xv
DAFTAR LAMPIRAN……… xvii
I PENDAHULUAN Latar belakang……… 1
Tujuan……… 8
Hipotesis………. 8
Manfaat……….. 8
II RUANG LINGKUP PENELITIAN Waktu dan Tempat………. 11
Alur Penelitian ...………... 12
III KERAGAMAN Pyricularia ASAL RUMPUT BERDASARKAN PENANDA SCAR DAN MAG Abstrak………... 15
Pendahuluan………... 15
Bahan dan Metode …..………... 18
Hasil………... 23
Pembahasan………... 29
Simpulan………... 33
IV KERAGAMAN TURUNAN HASIL INFEKSI SILANG Pyricularia ASAL Digitaria ciliaris Abstrak………... 35
Pendahuluan………... 35
Bahan dan Metode ………... 39
Hasil………....……... 50
Pembahasan……… 65
Simpulan……… 78
V HUBUNGAN FILOGENETIK Pyricularia PADA RUMPUT DAN PADI Abstrak………... 81
Pendahuluan………... 81
Bahan dan Metode ……...………. 84
Hasil………... 86
Pembahasan……….... 90
Simpulan……….... 93
VI PEMBAHASAN UMUM………..….…... 95
VII SIMPULAN DAN SARAN Simpulan………..….…... 103
Saran…....………... 104
DAFTAR PUSTAKA……..……... 105
DAFTAR TABEL
Halaman 1 Isolat Pyricularia dari satu bercak blas pada beberapa rumput dari
tiga lokasi di Jawa Barat, dari padi di Lampung dan pada empat lokasi di Jawa Barat... 19 2 Ragam fenotipe berdasarkan tiga penanda SCAR... 22 3 Keragaman penanda SCAR dan mag pada Pyricularia dari beberapa
rumput dan padi…...………...….. 26 4 Skor dan gejala blas menurut IRRI (1996)... 48 5 Contoh pemberian nomor ras Pyricularia asal padi (Mogi et al.
1991)... 49 6 Hasil inokulasi Pyricularia d4 asal D. ciliaris dan o173 asal padi ke
padi var. Kencana bali, Cisokan, IR64, dan ke rumput P. repens, C. dactylon, Digitaria sp., O. nodosa; beserta penanda SCAR pada turunan hasil pergantian inang tahap ke-1... 52 7 Hasil inokulasi Pyricularia d~k dan d~c ke padi var. Kencana bali
dan Cisokan, beserta penanda SCAR pada turunan hasil pergantian
inang tahap ke-2... 53 8 Kultur yang dianalisis dengan metode AFLP dan disekuen ITS
beserta 5.8S rDNAnya ………... 54 9 Jarak genetik antara isolat klonal Pyricularia asal D. ciliaris dan
padi dari lokasi dan waktu yang sama, serta jarak genetiknya terhadap turunan Pyricularia d4 asal D. ciliaris dari hasil pergantian inang ke padi tahap ke-1 dan ke-2...
56 10 Jarak genetik antara Pyricularia o173 asal padi dan turunannya dari
hasil pergantian inang ke padi...……….. 56 11 Jarak genetik Pyricularia d4 asal D. ciliaris terhadap turunannya
dari hasil pergantian inang ke padi tahap ke-1 dan ke-2, serta ke rumput P. repens tahap ke-1.......... 57 12 Strain yang dianalisis melalui amplifikasi repetitive dari Pot2.…... 63 13 Reaksi Pyricularia d4 asal D. ciliaris dan turunannya dari hasil
pergantian inang ke padi tahap ke-1, dan ke rumput P. repens terhadap tujuh varietas diferensial Indonesia…...
64 14 Jarak genetik beserta penandanya pada penelusuran riwayat
Pyricularia d4 asal D. ciliaris dan o173 asal padi yang mengalami pergantian inang ... 69 15 Hasil BLAST sekuen ITS beserta 5.8S rDNA (hasil amplifikasi
primer universal its1 dan its4) Pyricularia d4 asal D. ciliaris... 88 16 Karakterisasi Pyricularia asal rumput (d4) dan padi (ok6 dan
DAFTAR GAMBAR
Halaman 1 Diagram alir penelitian………... 12 2 Fenotipe bercak blas pada daun rumput Digitaria ciliaris (a) dan
daun padi (b)... 20 3 Konidiofor cendawan blas pada rumput D. ciliaris...... 23 4 Susunan konidium pada konidiofor cendawan dari bercak blas pada
rumput D. ciliaris... 23 5 Konidium cendawan dari bercak blas pada daun rumput D. ciliaris.. 24 6 Tabung kecambah dan apresorium cendawan dari bercak blas pada
rumput D. ciliaris... 24 7 Penanda Cut1 pada Pyricularia dari rumput danpadi... 25 8 Penanda PWL2 pada Pyricularia dari rumput danpadi... 25 9 Penanda Erg2 pada Pyricularia dari rumput danpadi... 25 10 Penanda magB (a) dan magC (b) pada Pyricularia dari rumput dan
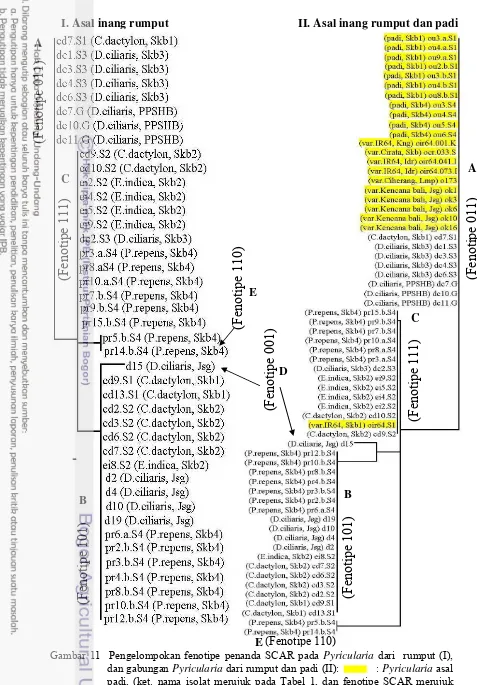
padi... 26 11 Pengelompokan fenotipe penanda SCAR pada Pyricularia dari
rumput (I), dan gabungan Pyricularia dari rumput dan padi (II)... 30 12 Daerah target amplifikasi DNA dengan primer universal its1 dan
its4... 49 13 Bercak blas hasil infeksi Pyricularia d4 asal D. ciliaris ke padi di
hari ke-9 setelah inokulasi, beserta bercak blas hasil infeksi o173 asal padi (kontrol) ke padi di hari ke-7 setelah inokulasi ... 50 14 Konidium Pyricularia d4 asal D. ciliaris dan turunannya dari hasil
pergantian inang ke padi tahap ke-1, beserta konidium o173 asal padi (kontrol)... 51 15 Bercak blas pada padi var. Kencana bali dan Cisokan hasil infeksi
Pyricularia d~k4.3 (a, b) dan d~c1.2 (c, d) di hari ke-7 setelah inokulasi... 51 16 Penanda SCAR Pyricularia d4 asal D. ciliaris dan turunannya dari
hasil pergantian inang ke padi pada tahap ke-1 dan ke-2, ke P. repens, beserta o173 dari padi (kontrol) dengan turunannya dari hasil pergantian inang ke padi...………... 53 17 Filogenetik berdasarkan 3 penanda AFLP………... 58 18 Pengelompokan melalui UPGMA berdasarkan 3 penanda AFLP... 59 19 Filogenetik dan pengelompokan melalui UPGMA berdasarkan 3
20 Fenotipe rep-Pot2 Pyricularia sampel basal asal D. ciliaris, turunan isolat basal (Pyricularia d4) dari hasil pergantian inang ke padi tahap ke-1 dan ke-2, ke P. repens; beserta o173 asal padi dengan turunannya dari hasil pergantian inang ke padi, dan Pyricularia asal padi dari lokasi dan waktu yang sama dengan Pyricularia d4 asal
D. ciliaris...... 63 21 Sekuen nukleotida bagian hilir 18S rDNA, daerah ITS beserta 5.8S
rDNA, dan bagian hulu 26S rDNA pada Pyricularia d4 dan turunannya dari pergantian inang ke P. repens (d~p1.1)…... 66 22 Penelusuran riwayat Pyricularia d4 asal D. ciliaris yang mengalami
pergantian inang ke padi tahap ke-1 dan ke-2, dan ke P. repens
berdasarkan 3 penanda AFLP, SCAR, rep-Pot2, dan ras fisiologi, serta sekuen ITS beserta 5.8S rDNA... 68 23 Filogenetik berdasarkan 3 penanda AFLP pada sampel basal
Pyricularia asal D. ciliaris dan turunan isolat basal (Pyricularia d4) dari hasil pergantian inang ke padi tahap ke-1 dan ke-2, ke P. repens; beserta Pyricularia o173 asal padi dengan turunannya dari hasil pergantian inang ke padi, dan Pyricularia asal padi dari lokasi dan waktu yang sama dengan Pyricularia d4 asal D. ciliaris... 77 24 Sekuen nukleotida bagian hilir 18S rDNA, daerah ITS beserta 5.8S
rDNA, dan bagian hulu 26S rDNA pada Pyricularia dari D. ciliaris
(d4), P. repens (pr10.a.S4) dan padi (ok6, ou6.S4)... ... 87 25 Filogenetik berdasarkan sekuen ITS beserta 5.8S rDNA pada
cendawan blas asal rumput dan padi yang tumbuh berdekatan pada dua lokasi di Jawa Barat terhadap data GeneBank …………... 90 26 Mikroevolusi Pyricularia d4 asal D. ciliaris pada pergantian inang
DAFTAR LAMPIRAN
Halaman 1 Spesies rumput yang memiliki bercak seperti penyakit blas pada
BAB I PENDAHULUAN Latar belakang
Bercak serupa penyakit blas padi dapat ditemukan pada beberapa spesies rumput yang tumbuh di areal penanaman padi di sawah Sukabumi. Areal persawahan ini termasuk wilayah endemik blas (Sobrizal et al. 2010). Sedangkan di Jasinga, Bogor, pada tahun 2005 beberapa galur padi percobaan di ladang terserang penyakit blas. Padi Kencana bali yang sedang diikutsertakan pada percobaan tersebut juga terserang penyakit blas. Areal tempat percobaan tersebut juga termasuk daerah endemik blas (Purwoko 2005), meskipun areal ini bukan lahan produksi padi dan tidak tampak areal persawahan di sekitarnya. Rumput
Digitaria ciliaris dan Panicum repens di rumah kaca Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) IPB juga memiliki bercak penyakit blas.
Morfologi Pyricularia yang diperoleh dari rumput dan padi ialah serupa (Rosman et al. 1990, Couch dan Kohn 2002). Nama Pyricularia grisea Sacc. muncul lebih awal sebagai spesies yang patogen pada Digitaria sanguinalis. Selanjutnya, Pyricularia oryzae Cav. muncul sebagai spesies patogen pada padi yang kemudian dikenal sebagai penyebab penyakit blas. Kedua spesies tersebut memiliki teleomorf yang sama pada awalnya, yaitu Magnaporthe grisea (Hebert) Barr (Ou 1985). Rosman et al. (1990) menempatkan P. oryzae sebagai sinonim dari P. grisea.
2
Pyricularia pada rumput di sekitar pertanaman padi di Filipina tidak menjadi sumber inokulum penyakit blas di padi (Borromeo et al. 1993), begitu pula di India, tidak ada aliran genetik Pyricularia patogen padi dan patogen selain padi (Rathour et al. 2006). Asosiasi spesifitas inang tampak dipertahankan melalui perbedaan pada patogenisitas di antara Pyricularia dari inang berbeda.
Pyricularia dari inang selain padi tidak patogen atau kurang patogen ke padi (Couch et al. 2005). Menurut Hamer et al (1989), seleksi inang untuk genotipe patogen yang spesifik terjadi selama pemuliaan dan budidaya padi.
Pyricularia dari Digitaria ciliaris dan Eragrostis sp. di Filipina memiliki karakteristik berbeda terhadap Pyricularia dari padi berdasarkan situs enzim restriksi DNA mitokondria (mtDNA) dan hasil hibridisasi dengan enam jenis probe (PGR613, PGR612, PGR46, PGR6G, PGR106, dan MGR586) dari elemen
repetitive (Borromeo et al. 1993). Pyricularia dari rumput juga membentuk kluster yang terpisah dari tanaman penting secara agronomi berdasarkan
restriction fragment length polymorphism (RFLP) dari rDNA dan sekuen ITS2 (Kusaba et al. 1999). Sampai saat ini analisis keragaman genetik Pyricularia yang mengalami pergantian inang dari genus yang berbeda belum pernah dilaporkan. Di Indonesia, sejak lama rumput yang tumbuh di sekitar pertanaman padi telah menjadi perhatian sehubungan dengan perkembangan penyakit blas, seperti dilaporkan oleh Rusli (1987), namun belum banyak informasi yang dapat diperoleh dari rumput-rumput di Indonesia.
Di Indonesia, rumput Echinochloa crusgalli (Cikampek), Leersia hexandra (Cilacap), Panicum maximum (Bali), dan Panicum repens (Subang) dilaporkan sebagai inang Pyricularia (DBPT Deptan 1992). Sedangkan rumput di sekitar pertanaman padi di Filipina sebagai inang Pyricularia ialah Brachiaria mutica, Brachiaria distachya, Dactyloctenium aegyptium, Digitaria ciliaris,
Echinochloa colona, Eleusine indica, L. hexandra, P. repens, Pennisetum
purpureum, dan Rottboellia exaltata (Mackill & Bonman 1986), Cenchrus echinatus, Cynodon dactylon, Cyperus brevifolius, Cyperus rotundus, Eragrostis
Menurut Couch dan Kohn (2002), Pyricularia pada rumput liar dan padi merupakan spesies terpisah. Pyricularia grisea merupakan spesies yang patogen pada Digitaria spp. (Digitaria horizontalis dari Brazil, Digitaria smutzi dari Jepang, dan Digitaria sp. dari USA dan Cina), dengan teleomorfnya ialah M. grisea, sedangkan P. oryzae merupakan spesies yang patogen pada padi dan berbagai anggota Graminae (serealia dan rumput) yang dibudidayakan. Teleomorf P. oryzae, yaitu Magnaporthe oryzae B. Couch. Magnaporthe oryzae
ditempatkan sebagai anggota M. grisea spesies kompleks. Serealia dan rumput budidaya tersebut adalah Eleusine coracana (finger millet), Eleusine indica,
Eragrostis curvula, Lolium perenne, Setaria sp. Selanjutnya, M. grisea kompleks digunakan sebagai nama cendawan penyebab blas pada padi dan beberapa anggota Graminae lain (Zellerhoff & Schaffrath 2006, Khang et al. 2008, Motallebi et al. 2009a).
Penyakit blas pada padi merupakan salah satu penyakit penting, karena menyebabkan banyak kehilangan hasil panen hingga 50% (Babujee & Gnanamanickam 2000). Pada tahun 1963, penyakit blas dilaporkan telah menyerang tanaman padi di 60 negara, dan pada tahun 1925 dilaporkan keberadaan penyakit blas pada padi di pulau Jawa (Parthasarathy & Ou 1963). Penyakit blas merupakan penyakit penting pada padi gogo (di lahan kering) di Indonesia. Penyakit tersebut telah menyebar luas, menyebabkan kerusakan padi sawah (Amir & Nasution 1993). Tingkat kerusakannya lebih besar pada padi di dataran tinggi. Pada tahun 1987-1988, frekuensi serangan penyakit blas di dataran rendah mencapai sekitar 56%, dan tahun 1990/1991 persawahan di Indramayu kehilangan produksi padi lebih dari 50% akibat serangan penyakit blas (DBPT Deptan 1992). Serangan penyakit tersebut cenderung meningkat setiap tahun. Pada tahun 2006, luas serangan penyakit blas paling tinggi terjadi di Lampung (1,923 ha) dan Jawa Barat (1853 ha). Sedangkan pada tahun 2007 dan 2008 berturut-turut luas serangannya paling tinggi di Jawa Barat (2864 ha dan 3205 ha) dan Sulawesi Selatan (2123 ha dan 2078 ha) (Deptan 2009).
4
2000). Padi var. Kencana bali merupakan salah satu anggota padi diferensial di Indonesia yang digunakan untuk uji patotipe (ras fisiologi). Padi tersebut rentan terhadap semua (27) ras Pyricularia patogen padi yang ada di Indonesia. Padi diferensial lainnya, di antaranya berupa padi var. Cisokan yang bersifat moderat resisten, yaitu resisten terhadap delapan ras atau dengan kata lain rentan terhadap serangan 21 ras Pyricularia patogen padi yang ada di Indonesia (DBPT Deptan 1992).
Isolat-isolat cendawan blas padi cenderung tidak stabil penampakan koloninya, fertilitas, dan patogenisitasnya selama disubkultur berulang-ulang di laboratorium (Valent & Chumley 1991). Pada penelitian pendahuluan juga tampak, umumnya cendawan blas yang berasal dari rumput memiliki warna koloni yang cepat berubah dari gelap menjadi putih selama disubkultur berulang-ulang di laboratorium. Selain itu, jumlah konidium cendawan blas yang berasal dari rumput sejak awal hasil isolasi lebih sedikit dibandingkan dengan cendawan blas yang diperoleh dari padi. Sporulasi Pyricularia dari rumput budidaya lebih baik daripada isolat dari rumput liar, kemungkinan karena perbedaan genetik sporulasinya (Rao et al. 1972). Pyricularia asal padi yang mengalami gangguan
magB menghasilkan mutan dengan efek pleotropi, yaitu mereduksi pertumbuhan somatik, konidiasi, pembentukan apresorium, dan patogenisitas.
Graminae lainnya (Ou 1985), dan Pyricularia dari berbagai inang bervariasi fertilitasnya (Zeigler 1998). Oleh karena itu, keragaman cendawan blas dari rumput dapat berdasarkan penanda molekuler pada lokus magB dan magC dari cendawan blas yang patogen pada padi.
Banyak penanda molekuler lain yang juga dapat digunakan untuk menganalisis keragaman genetik. Sebagai contoh, sebanyak 14 pasang primer sequence characterized amplified region marker (SCAR) telah digunakan untuk memonitor rekombinasi dan migrasi populasi Pyricularia patogen padi dari benua Afrika, Amerika Utara, Amerika Selatan, Asia, dan Eropa (Soubabere et al. 2000). Begitu juga di Indonesia, sebanyak tiga pasang primer SCAR telah digunakan untuk menunjukkan keragaman genetik Pyricularia patogen padi dari beberapa daerah endemik blas (Reflinur et al. 2005). Sampai saat ini belum diperoleh informasi keragaman penanda SCAR pada Pyricularia dari rumput di sekitar pertanaman padi, khususnya di Indonesia. Oleh karena itu pada penelitian ini akan dilakukan analisis keragaman Pyricularia dari beberapa spesies rumput menggunakan tiga penanda SCAR, yaitu Cut1, PWL2, dan Erg2.
Pada daur penyakit, hubungan inang utama dan inang alternatif untuk mempertahankan propagul yang dapat menginfeksi sangat penting. Beberapa
Pyricularia dari rumput hanya dapat menginfeksi padi yang rentan (Mackill & Bonman 1986; Singh & Singh 1988; DBPT Deptan 1992), dan beberapa rumput lain (DBPT Deptan 1992). Hasil uji di rumah kaca menunjukkan, Pyricularia
yang bukan berasal dari padi umumnya avirulen atau lemah virulensinya terhadap padi (Couch et al. 2005). Kemampuan infeksi silang Pyricularia yang dilakukan oleh para peneliti tersebut di atas tidak didasarkan pada riwayat inokulan spora tunggal. Oleh sebab itu, analisis genetik sampel basal (isolat-isolat satu klonal dengan inokulan) sangat diperlukan untuk menduga tingkat mikroevolusi yang muncul akibat pergantian inang.
6
berulang (repeatable) (Mettler et al. 1988). Mutasi titik, penyusunan kembali gen (genetic rearrangements), dan transfer gen merupakan proses yang menyumbang terjadi evolusi pada organisme, termasuk evolusi mikroba. Contoh mikroevolusi
Cochliobolus carbonatum pada jagung di alam yang mengalami insersi
transposon pada gen resistensinya (Multani et al. 1998). Contoh lainnya ialah keragaman Fusarium oxysporum f. sp. albedinis penyebab layu pada palm berdasarkan keserasian vegetatif (vegetative compatibility=VCG), RFLP mtDNA, dan random amplified polymorphic DNA (RAPD) (Fernandez et al. 1997). Proses mikroevolusi pada jangka waktu lama menghasilkan perkembangan spesies baru atau subspesies dan disebut dengan istilah makroevolusi (Morschhäuser et al. 2000).
Sampai saat ini belum diperoleh laporan tentang mikroevolusi Pyricularia
yang mengalami pergantian genus inang. Hal yang menarik ialah injeksi konidium patogen blas padi ke kultivar padi yang panikelnya resisten penyakit blas menghasilkan isolat turunan (asal bercak blas pada spikeletnya) dengan sifat-sifat patogenisitas dan genetik yang bervariasi (Namai & Iwade 2002, Namai 2011). Fenomena yang sama mungkin dapat terjadi juga pada cendawan blas asal rumput, pergantian inang kemungkinan dapat menimbulkan variasi patogenisitas dan genetiknya. Perubahan genetik ini diduga dapat bersifat stabil dan perubahan ini dapat diamati dengan menggunakan penanda molekuler.
Penanda molekuler lainnya yang dapat digunakan untuk mengamati keragaman genetik di antaranya metode amplified fragment lenght polymorphism
(AFLP). AFLP telah digunakan untuk membedakan Pyricularia asal padi indica
dan japonica (Thuan et al. 2000). AFLP juga dipakai oleh Tredway et al. (2005) untuk menunjukkan keragaman genetika Pyricularia asal rumput industri, padi, dan gandum. AFLP tidak digunakan untuk analisis filogenetik pada tingkat di atas spesies (Robinson & Harris 1999). AFLP memiliki potensi sebagai sumber informasi filogenetik sistematika molekuler pada takson yang berkerabat sangat dekat, sangat baik untuk mempelajari hubungan filogenetik ketika sekuen internal transcribed spacer (ITS) bersifat terkonservasi (Koopman 2005).
juga dapat digunakan untuk menganalisis keragaman isolat Pyricularia patogen padi. Daerah ITS rDNA nukleus paling cepat mengalami variasi diantara populasi (White et al. 1990). Sekuen ITS yang banyak bervariasi dapat disejajarkan (aligned) dengan tingkat kepercayaan hanya antara taksa yang sangat berkerabat dekat (Guarro et al. 1999). Oleh sebab itu identitas, keragaman genetik sampel basal, dan stabilitas genetik cendawan blas asal rumput yang mengalami pergantian inang dipelajari secara molekuler dengan bantuan teknik SCAR, AFLP, rep-Pot2 dan hubungannya dengan ras fisiologi, serta sekuen ITS.
Di seluruh dunia, cendawan blas yang dominan di lapang ialah anamorfnya (Kato et al. 2000). Teleomorf tidak pernah ditemukan di lapang, meskipun terdapat indikasi keberadaan siklus seksual di lapang berdasarkan penanda molekuler repeat-induced point mutation (RIP) (Ikeda et al. 2002). Teleomorf hanya dihasilkan di laboratorium dari penyilangan dua isolat yang membawa jenis mating type berbeda, dan salah satu isolat tersebut bersifat hermaprodit (Zeigler 1998). Oleh karena itu spesies epitet seringkali berdasarkan inang. Perkembangan pendekatan spesies mempengaruhi identitas Pyricularia, sehingga koreksi penamaan masih berlangsung. Lokus actin, β-tubulin, calmodulin, dan ITS rDNA telah menjadi objek penelitian untuk memecahkan masalah penamaan dan hubungan filogenetik antara isolat. Lokus tersebut juga digunakan untuk menganalisis variasi genetik populasi Pyricularia. Oleh sebab itu karakterisasi cendawan blas pada rumput dan padi sangat diperlukan untuk menetapkan nama yang tepat terutama kaitannya dengan mikroevolusinya akibat pergantian inang.
Daerah ITS dapat digunakan untuk membedakan spesies Trichophyton
8 Tujuan
Penelitian ini bertujuan untuk
(i) Menganalisis keragaman populasi Pyricularia dari beberapa rumput yang tumbuh berdekatan dengan padi berdasarkan penanda SCAR yang terdiri atas Cut1, PWL2 dan Erg2; serta penanda magB, dan magC
(ii) Menganalisis keragaman genetik sampel basal Pyricularia dari rumput D.
ciliaris dan mikroevolusi hasil induksi pergantian genus inangnya
berdasarkan lima penanda, yaitu penanda SCAR, AFLP, PCR repetitive Pot2 (rep-Pot2), dan hubungannya dengan ras fisiologi (patotipe), serta sekuen ITS beserta 5.8S rDNA nukleus
(iii) Menganalisis hubungan filogenetik berdasarkan sekuen ITS beserta 5.8S rDNA nukleus antara Pyricularia dari rumput dan padi yang diperoleh dari lokasi dan waktu yang sama.
Hipotesis :
1) Lokus SCAR yaitu Cut1, PWL2, Erg2, magB, dan magC dapat digunakan sebagai dasar analisis tingkat keragaman Pyricularia patogen rumput
2) Isolat-isolat Pyricularia hasil pertumbuhan konidium tunggal dari satu bercak blas membentuk karakter sampel basal dengan heterogenisitas terbatas dan perubahan genotipe inang memicu keragaman genetik dari sampel basal
Pyricularia
3) Terdapat hubungan filogenetik yang sangat dekat antara Pyricularia pada rumput yang tumbuh di sekitar padidengan Pyricularia yang berada pada padi, sehingga keduanya merupakan spesies yang sama, yaitu sebagai Pyricularia grisea
Manfaat
Keragaman penanda SCAR (Cut1, PWL2, Erg2), magB dan magC pada
Pyricularia dari beberapa spesies rumput yang tumbuh berdekatan dengan padi di sawah Sukabumi dan ladang Jasinga, Bogor untuk menduga tingkat variasi
BAB II
RUANG LINGKUP PENELITIAN
Waktu dan Tempat
Penelitian dilaksanakan mulai Oktober 2005 – Oktober 2007 dan September 2008 - Januari 2010, bertempat di Laboratorium Genetika Cendawan, Laboratorium Biotechnology Research Indonesia –Netherland (BIORIN), dan Rumah Kaca Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) IPB.
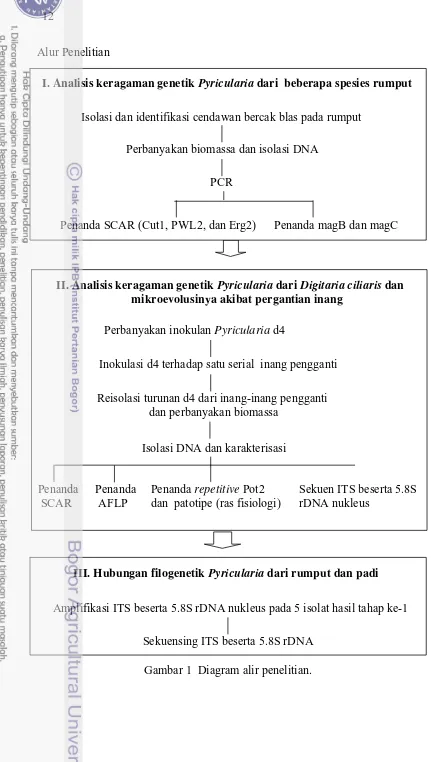
Alur Penelitian
12
Alur Penelitian
III. Hubungan filogenetik Pyricularia dari rumput dan padi
Amplifikasi ITS beserta 5.8S rDNA nukleus pada 5 isolat hasil tahap ke-1 Sekuensing ITS beserta 5.8S rDNA
I. Analisis keragaman genetik Pyricularia dari beberapa spesies rumput
Isolasi dan identifikasi cendawan bercak blas pada rumput
PCR
[image:38.595.43.474.41.803.2]Penanda SCAR (Cut1, PWL2, dan Erg2) Penanda magB dan magC Perbanyakan biomassa dan isolasi DNA
Gambar 1 Diagram alir penelitian. Isolasi DNA dan karakterisasi Perbanyakan inokulan Pyricularia d4
Inokulasi d4 terhadap satu serial inang pengganti Reisolasi turunan d4 dari inang-inang pengganti
dan perbanyakan biomassa
Penanda AFLP
Penanda repetitive Pot2 dan patotipe (ras fisiologi) Penanda
SCAR
Sekuen ITS beserta 5.8S rDNA nukleus
Program yang digunakan untuk menganalisis data hasil penelitian ini adalah Phylogenetic Analysis Using Parsimony (PAUP*) version 4.0b10 for 32-bit Microsoft Windows (Swofford 2002) dan NTSYS Spc version 2 (Rohlf 1998). PAUP* digunakan untuk menganalisis pengelompokan keragaman penanda SCAR dari beberapa spesies rumput melalui distance dengan UPGMA. Program ini juga digunakan untuk menganalisis data AFLP dari hasil pergantian inang, yaitu mengkonstruksi filogram melalui distance (Robinson & Harris 1999) dengan heuristic. Pada analisis tersebut, tingkat perubahan pola AFLP dari
BAB III
KERAGAMAN Pyricularia ASAL RUMPUT BERDASARKAN PENANDA SCAR DAN MAG
Abstrak
Karakterisasi penanda sequence characterized amplified region (SCAR), magB, dan magC pada populasi Pyricularia dari rumput yang tumbuh liar disekitar tanaman padi belum dilaporkan. Oleh karena itu dilakukan karakterisasi tiga penanda SCAR (Cut1, PWL2, Erg2), dan dua penanda lainnya berupa magB dan magC pada Pyricularia dari beberapa spesies rumput yang tumbuh berdekatan dengan padi untuk menduga tingkat variasi Pyricularia di Jawa Barat. Sebanyak 41 isolat Pyricularia dari Cydonon dactylon, Eleusine indica, Digitaria ciliaris, dan Panicum repens menunjukkan semuanya memiliki kesamaan ukuran fragmen DNA hasil amplifikasi kelima penanda. Ukuran fragmen Cut1, PWL2, dan Erg2 berturut-turut ± 1700 pb, 900 pb dan 1400 pb. Sedangkan ukuran fragmen magB dan magC masing-masing ± 1330 pb dan 1550 pb. Ukuran fragmen Cut1, PWL2, Erg2, magB, dan magC tersebut juga sama dengan ukuran fragmen 22 isolat Pyricularia dari padi. Sampel Pyricularia dari empat spesies rumput memiliki lima fenotipe SCAR yang dikonstruksi berdasarkan hasil amplifikasi dengan urutan Cut1, PWL2, dan Erg2. Kelima pola fenotipe ini menunjukkan frekuensi yang berbeda, yaitu 19.5% berfenotipe 011 (A), 41.5% berfenotipe 101 (B), 31.7% berfenotipe 111 (C), 2.5% berfenotipe 001 (D), dan 4.9% berfenotipe 010 (E). Fenotipe SCAR Pyricularia dari satu bercak blas rumput kebanyakan terdiri atas dua fenotipe SCAR, dengan variasi fenotipe terutama pada Cut1 dan PWL2. Frekuensi gen penanda SCAR juga bervariasi, yaitu sebanyak 78.1% Pyricularia dari rumput memiliki Cut1, hanya 54.1% yang memiliki PWL2, dan 95.1% memiliki Erg2. Hanya satu isolat Pyricularia
yang tidak menghasilkan amplikon magC, sehingga magB dan magC tidak dapat menunjukkan keragaman genetik Pyricularia darirumput.
Kata kunci: Pyricularia, Cydonon dactylon, Eleusine indica, Digitaria ciliaris, Panicum repens, padi, SCAR, magB, magC
Pendahuluan
Beberapa spesies rumput yang tumbuh di sekitar pertanaman padi telah dilaporkan sebagai inang Pyricularia. Di Indonesia, rumput seperti Echinochloa crusgalli (Cikampek), Leersia hexandra (Cilacap), Panicum repens (Subang), dan
Panicum maximum (Bali) sebagai inang cendawan blas (Deptan DBPT 1992).
Leersia hexandra dan P. repens juga menjadi inang cendawan blas di Filipina, selain itu bercak blas juga ditemukan pada Brachiaria distachya, Brachiaria mutica, Dactyloctenium aegyptium, Digitaria ciliaris, Echinochloa colona,
16
Cyperus iria, dan Cyperus rotundus (Singh & Singh 1988), serta Pennisetum purpureum di Ghana (Nutsugah et al. 2008).
Pada penelitian pendahuluan tampak bahwa cendawan blas yang berasal dari rumput memiliki warna koloni yang cepat berubah dari gelap menjadi putih selama disubkultur berulang-ulang di laboratorium. Selain itu, jumlah konidium cendawan blas yang berasal dari rumput sejak awal hasil isolasi lebih sedikit dibandingkan dengan cendawan blas yang diperoleh dari padi. Sporulasi
Pyricularia dari rumput budidaya lebih baik daripada isolat dari rumput liar, kemungkinan karena perbedaan genetik sporulasinya (Rao et al. 1972).
Pyricularia dari padi yang mengalami gangguan magB menghasilkan mutan dengan efek pleotropi, yaitu mereduksi pertumbuhan somatik, konidiasi, pembentukan apresorium, dan patogenisitas. Isolat-isolat cendawan blas padi dilaporkan cenderung tidak stabil dalam penampakan koloni, fertilitas, dan patogenisitasnya selama disubkultur di laboratorium (Valent & Chumley 1991).
Pembentukan apresorium pada mutan nul magB dapat dipulihkan melalui penambahan cAMP (Liu & Dean 1997). Mutasi dominan magB menyebabkan autolisis koloni yang telah tua, pembentukan melanin tertunda, reduksi reproduksi seksual dan aseksual. Selanjutnya, mutan dominan magB mampu menghasilkan apresorium pada permukaan hidrofobik dan hidrofilik, meskipun perkembangan pada permukaan hidrofilik tertunda. Mutan dari tipe mutasi magB lainnya adalah tidak menyebabkan perubahan fenotipe yang drastis, hanya meningkatkan sensitivitas terhadap penghambatan konidiasi melalui tekanan osmotik (Fang & Dean 2000). Sedangkan delesi pada magC menyebabkan mutannya mengalami reduksi konidiasi, tetapi tidak mempunyai efek pada pertumbuhan miselium atau pembentukan apresorium. Sebaliknya delesi magA tidak memiliki efek pada pertumbuhan somatik, konidiasi, ataupun pembentukan apresorium (Liu & Dean 1997). Gen magB dan magC beserta magA berada pada kromosom yang terpisah dan hanya satu salinan (copy) dalam genom
Pyricularia dari padi(Dean 1997).
1998). Sebagai contoh, 78 sampel dari populasi Pyricularia dari Stenotaphrum secundatum didominasi oleh gen kawin tipe Mat1-1 dan bersifat steril, hanya satu sampel yang jantan fertil dengan gen kawin tipe Mat1-2, tidak ditemukan betina yang fertil, meskipun ditemukan dua macam gen tipe kawin. Contoh lainnya 87 sampel populasi Pyricularia dari inang Festuca arundinaceae juga memiliki gen kawin tipe Mat1-1, dengan betina fertilnya berjumlah 47. Sebanyak 47 sampel betina fertil tersebut yang meghasilkan peritesium kosong ialah 19 sampel. Frekuensi gen tipe kawin lawan jenisnya dari populasi Pyricularia pada kedua inang sangat rendah (0-5.7%) (Tredway et al. 2003). Hal tersebut di atas mungkin berhubungan dengan subunit α dari protein G yang disandikan oleh tiga gen, yaitu
magA, magB, dan magC (Dean 1997). Gen subunit α dari protein G mengontrol pertumbuhan, perkembangan, patogenisitas, dan diperlukan untuk perkawinan
Pyricularia dari padi (Liu & Dean 1997). Oleh karena itu penanda magB dan magC dapat digunakan sebagai dasar keragaman pada Pyricularia untuk mendapatkan informasi keberadaan kedua penanda tersebut sehubungan dengan pemisahan spesies dan aliran genetik antara Pyricularia pada inang rumput dan padi.
Isolat yang tidak berasal dari padi umumnya tidak patogen terhadap padi, ataupun hanya memberikan reaksi patogen lemah (Singh dan Singh 1988, Couch
et al. 2005). Demikian halnya di Indonesia, semua isolat Pyricularia dari Leersia hexandra, Panicum maximum, Panicum repens, dan Echinochloa crusgalli hanya patogen terhadap padi var. Kencana bali. Sedangkan isolat dari L. hexandra hanya patogen terhadap P. repens, tidak terjadi saling infeksi silang dari isolat lainnya (DBPT Deptan 1992). Sebaliknya, Pyricularia NBG-A8401 dari padi mampu menginfeksi lima spesies gulma yang berupa rumput (Brachiaria distachya,
Echinochloa colona, L. hexandra, Leptochloa chinensis, dan Rottboellia exaltata), dan dua strain lainnya dari Pyricularia asal padi (2017 dan 43) hanya mampu menginfeksi L. chinensis (Mackill & Bonman 1986).
18
amplified region marker (SCAR). Haplotipe Pyricularia padi dari Asia memiliki keragaman tinggi, sehingga berada di semua grup. Sebaliknya haplotipe
Pyricularia padi dari Eropa tidak menyebar, berada dalam satu grup dengan
Pyricularia padi dari Amerika Utara, Amerika Selatan, dan Asia (Soubabere et al. 2000). Sebanyak 16 jenis primer penanda SCAR yang dikembangkan oleh Soubabere et al. (2001) untuk memonitor rekombinasi dan migrasi populasi
Pyricularia padi. Sebanyak tiga jenis (Cut1, PWL2, dan Erg2) penanda SCAR dari ke 16 jenis tersebut dapat menunjukkan keragaman genetik (haplotipe)
Pyricularia dari padi daerah endemik blas, yaitu Sumatra Utara, Sumatra Barat, Lampung, Sukabumi, dan Bogor (Reflinur et al. 2005).
Sampai saat ini belum diperoleh informasi keragaman penanda SCAR dan mag pada Pyricularia penginfeksi rumput yang tumbuh liar di sekitar tanaman padi. Oleh sebab itu penelitian ini bertujuan menganalisis keragaman
Pyricularia dari beberapa spesies rumput yang tumbuh di sekitar tanaman padi berdasarkan lima macam penanda molekuler, yaitu tiga penanda SCAR (Cut1, PWL2, Erg2), dan dua penanda lainnya berupa magB, dan magC. Hasil penelitian ini diharapkan dapat menduga tingkat variasi Pyricularia di Jawa Barat.
Bahan dan Metode
Tempat. Penelitian dilaksanakan di Laboratorium Genetika Cendawan dan Biorin di Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) IPB.
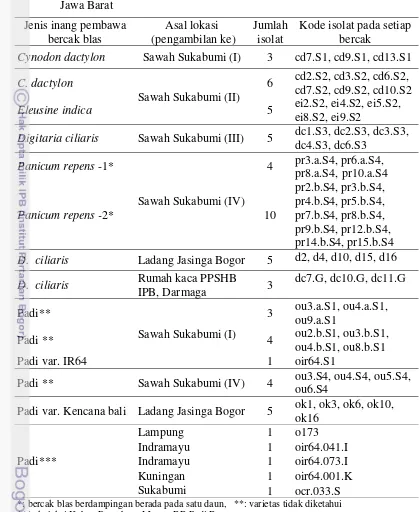
Bahan. Bercak blas pada helaian daun spesies rumput dari tiga lokasi berbeda, yaitu dari empat spesies rumput di sekitar pertanaman padi pada sawah di Sukabumi dengan waktu pengambilan berbeda, satu spesies rumput di sekitar pertanaman padi pada ladang di Jasinga Bogor, dan satu spesies rumput di rumah kaca PPSHB IPB, Kampus Dermaga (Tabel 1). Sebagai pembanding, bercak blas dari helaian daun padi diambil dari lokasi dan waktu yang sama dengan pengambilan sampel dari rumput. Selain itu, sebagai kontrol digunakan
Tabel 1 Isolat Pyricularia dari satu bercak blas pada beberapa rumput dari tiga lokasi di Jawa Barat, dan dari padi di Lampung dan pada empat lokasi di Jawa Barat
Jenis inang pembawa bercak blas
Asal lokasi (pengambilan ke)
Jumlah isolat
Kode isolat pada setiap bercak
Cynodon dactylon Sawah Sukabumi (I) 3 cd7.S1, cd9.S1, cd13.S1
C. dactylon
Sawah Sukabumi (II)
6 cd2.S2, cd3.S2, cd6.S2, cd7.S2, cd9.S2, cd10.S2
Eleusine indica 5 ei2.S2, ei4.S2, ei5.S2,
ei8.S2, ei9.S2
Digitaria ciliaris Sawah Sukabumi (III) 5 dc1.S3, dc2.S3, dc3.S3,
dc4.S3, dc6.S3
Panicum repens -1*
Sawah Sukabumi (IV)
4 pr3.a.S4, pr6.a.S4, pr8.a.S4, pr10.a.S4
Panicum repens-2* 10
pr2.b.S4, pr3.b.S4, pr4.b.S4, pr5.b.S4, pr7.b.S4, pr8.b.S4, pr9.b.S4, pr12.b.S4, pr14.b.S4, pr15.b.S4
D. ciliaris Ladang Jasinga Bogor 5 d2, d4, d10, d15, d16
D. ciliaris Rumah kaca PPSHB
IPB, Darmaga 3
dc7.G, dc10.G, dc11.G
Padi**
Sawah Sukabumi (I)
3 ou3.a.S1, ou4.a.S1, ou9.a.S1
Padi ** 4 ou2.b.S1, ou3.b.S1,
ou4.b.S1, ou8.b.S1
Padi var. IR64 1 oir64.S1
Padi ** Sawah Sukabumi (IV) 4 ou3.S4, ou4.S4, ou5.S4, ou6.S4
Padi var. Kencana bali Ladang Jasinga Bogor 5 ok1, ok3, ok6, ok10, ok16
Padi***
Lampung 1 o173
Indramayu 1 oir64.041.I Indramayu 1 oir64.073.I
Kuningan 1 oir64.001.K
Sukabumi 1 ocr.033.S
*: bercak blas berdampingan berada pada satu daun, **: varietas tidak diketahui ***: koleksi Kebun Percobaan Muara, BB Padi Bogor
Isolasi Konidium Tunggal dan Identifikasi Cendawan. Satu bercak blas (Gambar 2) dicuci dengan air mengalir dan dilembapkan semalam, kemudian
sejumlah konidium tunggal diisolasi dengan bantuan mikroskop (modifikasi dari
Bonman et al. 1987). Konidium tunggal dikecambahkan pada medium agar-agar
20
Isolat hasil perkecambahan ditumbuhkan pada medium cawan Potato Dextrose Agar (PDA, Difco) dan disimpan pada agar-agar miring PDA.
Kultur cendawan dari konidium tunggal yang diperoleh dari hasil isolasi diidentifikasi dengan bantuan mikroskop dan menggunakan buku acuan Ou (1985). Kultur cendawan terlebih dahulu ditumbuhkan pada medium sporulasi, yaitu medium oatmeal (30 g oatmeal L-1, 20 g agar-agar L-1, 5 g sukrosa L-1, modifikasi Tsurushima et al. 2005). Hifa aerial kultur cendawan berumur 7-8 hari dihilangkan secara aseptik dengan bantuan kaca objek dan akuades steril. Selanjutnya kultur ditutup dengan plastik transparan dan diberi lubang untuk aerasi serta disinari n-UV terus menerus selama 4-5 hari untuk menginduksi pembentukan konidium.
[image:46.595.72.479.81.475.2]Isolasi dan Amplifikasi DNA. DNA genom cendawan diisolasi dari miselium yang ditumbuhkan pada 25 mL medium cair (5 g L-1 sukrosa, 2 g L-1 ekstrak khamir, dan 2 g L-1 pepton, modifikasi Crawford et al. 1986) selama enam hari pada mesin pengocok. Miselium dipanen dan diisolasi genomnya menurut prosedur yang dijelaskan oleh Raeder dan Broda (1985) dengan volume lebih besar dan sedikit modifikasi. Miselium digerus dalam mortar steril sampai terbentuk pasta. Pasta disuspensikan dalam 4 mL larutan penyangga ekstrak (200 mM Tris HCl pH 8.5; 250 mM NaCl; 25 mM EDTA; 0.5% SDS). Sebanyak 2.8 mL fenol dan 1.2 mL campuran kloroform dan isoamil alkohol (CIA=24:1) ditambahkan ke dalam suspensi dan suspensi dibolak-balik secara perlahan. Suspensi disentrifugasi selama 30 menit pada 4,000 rpm dengan suhu 6 °C. Fase cairan bagian atas segera dipindahkan ke tabung baru dan dipresipitasi dengan menambahkan 1x volume isopropanol dingin. Hasil presipitasi disentrifugasi
Gambar 2 Fenotipe bercak blas pada daun rumput dan padi: a. Digitaria ciliaris, dan b. padi.
a
b
selama 20 menit pada 4,000 rpm dengan suhu 6 °C. Endapan dibilas dengan etanol dingin konsentrasi 70% dan disentrifugasi kembali selama 10 menit. Endapan dikeringkan dengan pompa vakum selama 15 menit, dan dilarutkan dalam 100 uL TE 1x (10 mm Tris HCl pH 8, 1 mmol EDTA), serta ditambahkan 0.2x volume RNAse 20 mg mL-1. Larutan diinkubasi semalam pada 37 °C, kemudian ditambahkan 900 uL TE 1x dan larutan diekstrak kembali dengan menambahkan 1xvolume CIA, dan disentrifugasi selama 10 menit pada 4,000 rpm dengan suhu 6 °C. Cairan bagian atas dipresipitasikan kembali dengan menambahkan isopropanol dingin seperti tahapan sebelumnya sampai diperoleh endapan yang dilarutkan dalam 100 uL TE 1x. DNA yang dianalisis memiliki tingkat kemurnian (A260/A280) 1.6-1.9.
DNA cendawan hasil isolasi diamplifikasi melalui PCR dengan menggunakan lima penanda molekuler. Kelima penanda tersebut terdiri atas tiga jenis penanda SCAR (Cut1, PWL2, Erg2) yang merupakan bagian dari 16 penanda SCAR yang dihasilkan oleh Soubabere et al. (2001). Sedangkan dua penanda lainnya, yaitu magB, dan magC masing-masing dikonstruksi berdasarkan sekuen nomor akses AF011341 dan AF011342 (Liu & Dean 1997). Primer
forward dan reverse magB masing-masing terletak pada nukleotida nomor 841-861 dan 2154- 2174. Sedangkan primer forward dan reverse magC masing-masing terletak pada nukleotida nomor 808-828 dan 2334-2354. Susunan nukleotida (primer) kelima penanda sebagai berikut:
22
Total reaksi PCR sebanyak 20 μL, mengandung sekitar 100 ng DNA genom cetakan; 10 μL 2x PCR master mix (0.05 unit μL-1 Taq DNA polymerase, 4 mM MgCl, 0.4 mM masing-masing dNTP), dan 0.6 ρmol masing-masing primer. Program PCR meliputi pradenaturasi pada suhu 95 °C selama 15 menit, dilanjutkan 35 siklus pada suhu 94 °C selama 1 menit, suhu pelekatan (annealing) primer Cut1, PWL2, dan Erg2 adalah 60 °C selama 45 detik, sedangkan suhu pelekatan primer magB dan magC adalah 56 °C, juga selama 45 detik. Program selanjutnya adalah pemanjangan (elongation) pada suhu 72 °C selama 2 menit. Tahap akhir proses PCR pada suhu 72 °C selama 15 menit. Produk PCR di visualisasi melalui elektroforesis pada gel agarosa 1% (w/v) dalam TAE 1x (0.04 M Tris-asetat dan 0.001 M EDTA), dan perendaman gel dalam 0.5 μg mL-1 etidium bromida. Pengamatan dilakukan terhadap keberadaan pita DNA pada gel agarosa yang diletakkan pada UV transluminator (Labquip) yang dilengkapi kamera digital (Olymphus). Data molekuler diperoleh melalui dua kali ulangan.
Analisis data. Amplikon pada isolat dari rumput dinyatakan ada (=1) bila terdapat fragmen DNA hasil amplifikasi. Sebaliknya jika tidak tampak fragmen DNA hasil amplifikasi dari suatu penanda diberi nilai 0. Analisis terhadap keragaman fenotipe penanda SCAR, magB, dan magC hanya dilakukan pada isolat-isolat dalam satu klon. Fenotipe SCAR berdasarkan kombinasi keberadaan Cut1, PWL2, dan Erg2 seperti yang dilakukan oleh Reflinur et al. (2005), tetapi dimodifikasi urutannya (Tabel 2). Fenotipe SCAR Pyricularia dianalisis lebih lanjut untuk mengetahui tipe pengelompokannya. Pada analisis ini juga dilakukan penggabungan fenotipe SCAR Pyricularia dari rumput dan padi Dendogram dikonstruksi melalui distance dengan UPGMA pada program Phylogenetic Analysis Using Parsimony (PAUP), version 4.0b10 for 32-bit Microsoft Windows (Swofford 2002).
Tabel 2 Ragam fenotipe berdasarkan tiga penanda SCAR
Penanda SCAR
Fenotipe Cut1 Pwl2 Erg2
A 0 1 1
B 1 0 1
C 1 1 1
D 0 0 1
Hasil
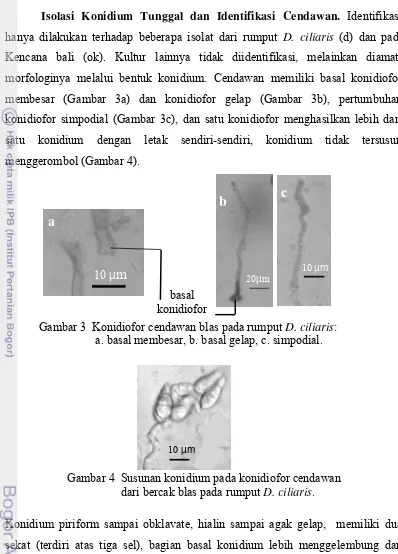
Isolasi Konidium Tunggal dan Identifikasi Cendawan. Identifikasi hanya dilakukan terhadap beberapa isolat dari rumput D. ciliaris (d) dan padi Kencana bali (ok). Kultur lainnya tidak diidentifikasi, melainkan diamati morfologinya melalui bentuk konidium. Cendawan memiliki basal konidiofor membesar (Gambar 3a) dan konidiofor gelap (Gambar 3b), pertumbuhan konidiofor simpodial (Gambar 3c), dan satu konidiofor menghasilkan lebih dari satu konidium dengan letak sendiri-sendiri, konidium tidak tersusun menggerombol (Gambar 4).
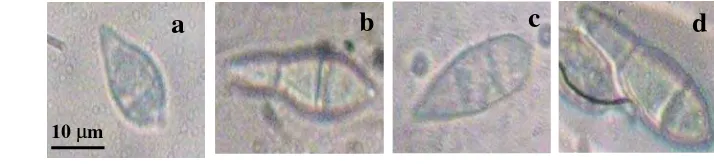
Konidium piriform sampai obklavate, hialin sampai agak gelap, memiliki dua sekat (terdiri atas tiga sel), bagian basal konidium lebih menggelembung dan terdapat embelan kecil, dan bagian atas konidium agak mengecil dan sekat atasnya sering menggenting (Gambar 5). Ukuran konidium-konidium dari bercak blas daun pada rumput D. ciliaris ialah (19.3-25.9) μm x (8.4-9.6) μm, sedangkan dari daun padi (15.1-23.7) μm x (8.1-9.6) μm, serta rataan ukuran konidium
[image:49.595.108.506.96.650.2]Pyricularia asal padi ras fisiologi 173 ialah 36.7 μm x 8.9 μm (Pandia 2008). Gambar 3 Konidiofor cendawan blas pada rumput D. ciliaris:
a. basal membesar, b. basal gelap, c. simpodial.
b
20µm basal
konidiofor
a
10 µ
m
c
10 µm
Gambar 4 Susunan konidium pada konidiofor cendawan dari bercak blas pada rumput D. ciliaris.
24
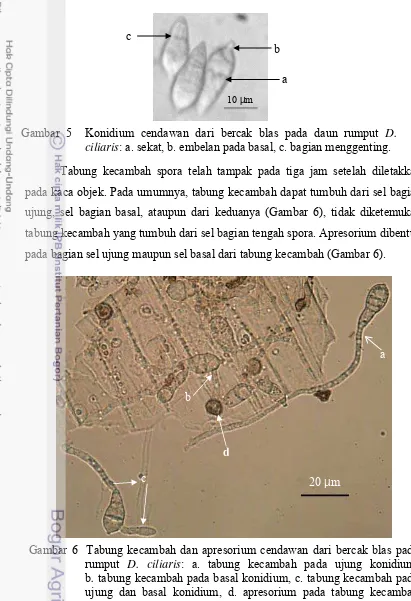
Tabung kecambah spora telah tampak pada tiga jam setelah diletakkan pada kaca objek. Pada umumnya, tabung kecambah dapat tumbuh dari sel bagian ujung, sel bagian basal, ataupun dari keduanya (Gambar 6), tidak diketemukan tabung kecambah yang tumbuh dari sel bagian tengah spora. Apresorium dibentuk pada bagian sel ujung maupun sel basal dari tabung kecambah (Gambar 6).
[image:50.595.60.471.72.673.2]Pada penelitian ini, isolat yang dianalisis adalah Pyricularia yang diperoleh dari bercak blas pada rumput yang tumbuh liar di sekitar pertanaman padi. Isolat dari padi hanya digunakan sebagai pembanding. Oleh karena itu Gambar 5 Konidium cendawan dari bercak blas pada daun rumput D.
ciliaris: a. sekat, b. embelan pada basal, c. bagian menggenting. b
a c
10 µm
a
b c 20 µm
Gambar 6 Tabung kecambah dan apresorium cendawan dari bercak blas pada rumput D. ciliaris: a. tabung kecambah pada ujung konidium, b. tabung kecambah pada basal konidium, c. tabung kecambah pada ujung dan basal konidium, d. apresorium pada tabung kecambah basal konidium.
c 20 µm
d
a
jumlah isolat dari padi yang dianalisis lebih sedikit dari jumlah isolat dari bercak blas pada rumput. Sebagai kontrol digunakan Pyricularia dari padi yang telah diketahui tipe ras fisiologinya (patotipe), koleksi BB Padi Bogor.
Amplikon Penanda Molekuler. Ukuran amplikon ketiga penanda SCAR pada 41 Pyricularia dari rumput sama dengan amplikon 22 Pyricularia dari padi. Ukuran amplikon ketiga penanda SCAR tersebut, yaitu Cut1, PWL2, dan Erg2 berturut-turut ± 1700 pb (Gambar 7), ± 900 pb (Gambar 8), dan ± 1400 pb (Gambar 9). Keberadaan amplikon ini sangat bervariasi bergantung asal inang dan lokasi asal bercak (Tabel 1). Ukuran amplikon penanda magB dan magC pada
Pyricularia dari rumput sama seperti amplikon Pyricularia dari padi, yaitu berurut-turut ± 1330 pb (Gambar 10a) dan 1550 bp (Gambar 10b).
Gambar 9 Penanda Erg2 pada Pyricularia dari rumput dan padi: M. DNA standar, 1. cd9.S2, 2. dc2.S3, 3. d2, 4. ei2.S2, k. kontrol negatif, 5. pr3.a.S4, 6. pr2.b.S4, 7. ou3.a.S1, 8. ouir64.S1, 9. ok1, 10. ocr.033; (ket. nama isolat mengacu pada Tabel 1).
2000 pb 1650 pb
1000 pb 850 pb
[image:51.595.105.505.91.820.2]±1400 pb M 1 2 3 4 k 5 6 7 8 9 10
Gambar 7 Penanda Cut1 pada Pyricularia dari rumput dan padi: M. DNA standar, 1. cd9.S2, 2. dc2.S3, 3. d4, k. kontrol negatif, 4. ei2.S2, 5. pr3.a.S4, 6. ouir64.S1; (ket. nama isolat mengacu pada Tabel 1).
M 1 2 3 k 4 5 6
±1700 pb 2000 pb
1000 pb
Gambar 8 Penanda PWL2 pada Pyricularia dari rumput dan padi: M. DNA standar, 1. cd9.S2, k. kontrol negatif, 2. dc2.S3, 3. ei2.S2, 4. pr3.a.S4, 5. ou3.a.S1, 6. ouir64.S1; (ket. nama isolat mengacu pada Tabel 1).
M 1 k 2 3 4 5 6
± 900 pb 650 pb
26
Keberadaan penanda Cut1, PWL2, dan Erg2 sangat bervariasi. Berdasarkan keragaman tersebut, Pyricularia dari rumput terdiri atas lima fenotipe SCAR, yaitu A, B, C, D, dan E (Tabel 3). Pada satu bercak Pyricularia
umumnya ditemukan dua fenotipe SCAR, kecuali Pyricularia dari P. repens
memiliki tiga fenotipe SCAR dan Pyricularia pada D. ciliaris dari rumah kaca PPSHB IPB hanya memiliki satu fenotipe SCAR, yaitu A (Tabel 3), sebaliknya, fenotipe SCAR Pyricularia dari padi lebih homogen. Hampir semua Pyricularia
dari padi ber fenotipe SCAR tipe A, kecuali Pyricularia dari padi IR64 berfenotipe berbeda (C).
Tabel 3 Keragaman penanda SCAR dan mag pada Pyricularia
dari beberapa rumput dan padi
Kode Isolat Cut1 PWL2 Erg2 Fenotipe B C SCAR Mag Rumput
cd7.S1 0 1 1 A 1 1
cd9.S1 1 0 1 B 1 1
cd13.S1 1 0 1 B 1 1
cd2.S2 1 0 1 B 1 1
cd3.S2 1 0 1 B 1 1
cd6. S2 1 0 1 B 1 1
cd7.S2 1 0 1 B 1 1
cd9.S2 1 1 1 C 1 1
cd10.S2 1 1 1 C 1 1
dc1.S3 0 1 1 A 1 1
dc2.S3 1 1 1 C 1 1
dc3.S3 0 1 1 A 1 1
Gambar 10 Penanda magB (a) dan magC (b) pada Pyricularia dari rumput dan padi: M. DNA standar, 1. cd9.S2, 2. dc2.S3, 3. d2, 4. ei2.S2, 5. pr3.a.S4, 6. pr2.b.S4, 7. ou3.a.S1, 8. ouir64.S1, 9. ok1, 10. ocr.033; k. kontrol negatif (ket. nama isolat mengacu pada Tabel 1).
b
k 1 2 3 4 5 6 7 8 9 10 M
2000 pb 1650 pb
1000 pb ±1550 pb
a
M 1 2 3 4 5 6 7 8 9 k ±1330 bp2000 bp 1650 bp
Tabel 3 Keragaman penanda SCAR dan mag pada Pyricularia dari beberapa rumput dan padi (Lanjutan)
Kode Isolat Cut1 PWL2 Erg2 Fenotipe B C SCAR Mag
dc4.S3 0 1 1 A 1 1
dc6.S3 0 1 1 A 1 1
d2 1 0 1 B 1 1
d4 1 0 1 B 1 1
d10 1 0 1 B 1 1
d15 0 0 1 D 1 0
d16 1 0 1 B 1 1
dc7.G 0 1 1 A 1 1
dc10.G 0 1 1 A 1 1
dc11.G 0 1 1 A 1 1
ei2.S2 1 1 1 C 1 1
ei4.S2 1 1 1 C 1 1
ei5.S2 1 1 1 C 1 1
ei8.S2 1 0 1 B 1 1
ei9.S2 1 1 1 C 1 1
pr3.a.S4 1 1 1 C tk tk
pr6.a.S4 1 0 1 B tk tk
pr8.a.S4 1 1 1 C tk tk
pr10.a.S4 1 1 1 C tk tk
pr2.b.S4 1 0 1 B tk tk
pr3.b.S4 1 0 1 B tk tk
pr4.b.S4 1 0 1 B tk tk
pr5.b.S4 1 1 0 E tk tk
pr7.b.S4 1 1 1 C tk tk
pr8.b.S4 1 0 1 B tk tk
pr9.b.S4 1 1 1 C tk tk
pr12.b.S4 1 0 1 B tk tk
pr14.b.S4 1 1 0 E tk tk
pr15.b.S4 1 1 1 C tk tk
Padi
ou3.a.S1 0 1 1 A 1 1
ou4.a.S1 0 1 1 A 1 1
ou9.a.S1 0 1 1 A 1 1
ou2.b.S1 0 1 1 A 1 1
ou3.b.S1 0 1 1 A 1 1
ou4.b.S1 0 1 1 A 1 1
ou8.b.S1 0 1 1 A 1 1
oir64.S1 1 1 1 C 1 1
ou3.S4 0 1 1 A tk tk
ou4.S4 0 1 1 A tk tk
28
Tabel 3 Keragaman penanda SCAR dan mag pada Pyricularia dari beberapa rumput dan padi (Lanjutan)
Kode Isolat SCAR Mag
Cut1 PWL2 Erg2 Fenotipe B C
ou6.S4 0 1 1 A tk tk
ok1 0 1 1 A 1 1
ok3 0 1 1 A 1 1
ok6 0 1 1 A 1 0
ok10 0 1 1 A 1 0
ok16 0 1 1 A 1 1
oir64.001.K 0 1 1 A 1 1
ocr.033.S 0 1 1 A 1 1
oir64.041.I 0 1 1 A 1 1
oir64.073.I 0 1 1 A 1 1
o173 0 1 1 A 1 1
1= ada, 0= tidak ada, tk = tidak dikarakterisasi; (ket. nama isolat merujuk pada Tabel 1)
Jenis rumput menentukan variasi fenotipe SCAR. Pada bercak-bercak blas di C. dactylon terdapat tiga fenotipe yaitu A, B, dan C, dengan B sebagai fenotipe dominan. Sedangkan pada D. ciliaris terdapat empat fenotipe yaitu A, B, C, dan D dengan fenotipe A yang dominan. Pada E. indica terdapat dua fenotipe yaitu C dan B, C sebagai fenotipe dominan. Pada P. repens terdapat tiga fenotipe, yaitu C, B, dan E, dengan fenotipe E paling jarang ditemukan.
Pada lokasi berbeda, fenotipe SCAR Pyricularia rumput juga berbeda, tetapi tidak demikian dengan fenotipe SCAR Pyricularia padi. Pyricularia pada
D. ciliaris dari Sukabumi memiliki fenotipe SCAR A dan C, sedangkan
Pyricularia pada D. ciliaris dari Jasinga memiliki fenotipe SCAR B dan D, serta fenotipe SCAR Pyricularia pada D. ciliaris dari rumah kaca hanya memiliki tipe A. Fenotipe SCAR Pyricularia dari Sukabumi didominasi oleh fenotipe B dan C, keduanya bernilai sama. Hanya fenotipe SCAR B yang dominan di Jasinga, dan di Jasinga tidak memiliki fenotipe SCAR C. Sebaliknya, fenotipe SCAR
Pyricularia padi tidak beragam, semua berfenotipe SCAR A, kecuali Pyricularia
Frekuensi penanda SCAR juga bervariasi. Sebanyak 78.1% Pyricularia
dari rumput memiliki Cut1, hanya 54.1% yang memiliki PWL2 dan 95.1% memiliki Erg2 (hanya 4.9% yang tidak memiliki Erg2). Sebaliknya, hanya satu
Pyricularia dari padi (IR64 dari Sukabumi) yang memiliki Cut1, dan 100% sampel Pyricularia dari padi memiliki PWL2, lebih tinggi daripada Pyricularia
dari rumput. Semua Pyricularia dari padi menunjukkan hasil amplifikasi penanda SCAR Erg2. Begitu juga semua Pyricularia dari rumput dan padi memiliki penanda magB. Hanya satu Pyricularia dari jenis rumput D. ciliaris (d15), dan dua Pyricularia dari padi var. Kencana bali (ok6 dan ok10) yang tidak menunjukkan amplikon magC. Ketiga Pyricularia ini diperoleh dari bercak blas di ladang Jasinga Bogor (Tabel 1). Penanda mag tidak dapat menunjukkan keragaman Pyricularia dari rumput dan padi.