PERBAND
AKTIVITAS
PENDEK
FAKULTAS MATE
IN
DINGAN METODE HIDROLISIS DA
S PEMBENTUKAN ASAM LEMAK
EK PADA PATI TAKTERCERNA TIP
DESI AWALINA
DEPARTEMEN KIMIA
ATEMATIKA DAN ILMU PENGETAH
INSTITUT PERTANIAN BOGOR
BOGOR
2012
DALAM
K RANTAI
TIPE 3
ABSTRAK
DESI AWALINA. Perbandingan Metode Hidrolisis dalam Aktivitas Pembentukan
Asam Lemak Rantai Pendek pada Pati Taktercerna Tipe 3. Dibimbing oleh
SUMINAR S ACHMADI.
Pati taktercerna ialah fraksi pati yang tidak terserap oleh sistem pencernaan
pada individu yang sehat, salah satunya ialah RS3, yang merupakan pati
retrogradasi dan memiliki kemiripan proses dengan daya cerna pati dalam tubuh
manusia. Penelitian ini bertujuan mengevaluasi kemampuan kelompok bakteri
Eubacterium rectale
dan
Clostridium butyricum
dalam memproduksi asam lemak
rantai pendek (SCFA:
short chain fatty acid
), yaitu asam asetat, propionat, dan
butirat dari RS3 yang dihidrolisis menggunakan asam klorida 2 N,
α
-amilase, dan
pululanase. Rendemen pati taktercerna yang didapat setelah proses hidrolisis
menggunakan HCl 2 N,
α
-amilase, dan pululanase berturut-turut adalah 6.6%,
2.3%, dan 48.1%, dengan derajat polimerisasi pati berturut-turut 15, 18, dan 25
residu, yang berarti telah memenuhi ketentuan sebagai RS3. Penelitian ini
menghasilkan nisbah butirat:propionat:asetat rerata sebesar 130:60:50, yang
mendekati metabolisme ideal SCFA. Jenis SCFA menggunakan media RCM
(
reinforced clostridial medium
) mirip dengan yang diproduksi menggunakan
media PYG (
pepton yeast glucose
) pada setiap perlakuan hidrolisis, walaupun
dengan ragam yang sangat berbeda.
Kata kunci: asam lemak rantai pendek, pati taktercerna, RS3, SCFA
ABSTRACT
DESI AWALINA. Comparison of Hydrolysis Methods and Their Effect on
Activity in Formation of Short Chain Fatty Acids of Resistant Starch Type 3.
Supervised by SUMINAR S ACHMADI.
Resistant starch is a fraction of starch not absorbed by the digestive system
in healthy individuals, one of which is RS3, a retrograded starch and has
similarities with digestibility process of starch in human body. This study aimed
to evaluate the ability of
Eubacterium rectale
and
Clostrydium butyricum
consortium in producing short chain fatty acids (SCFA), namely acetic, propionic,
and butyric acids from RS3 under 3 different hydrolysis methods. Yield of
resistant starch from hydrolysis using 2 N HCl ,
α
-amylase, and pullulanase were
6.6%, 2.3%, and 48.1%, respectively, with the degree of polymerization were 15,
18, and 25 residues, respectively, meaning that they meet the requirement as RS3.
This experiment resulted an average of butyrate:propionate:acetate which was
130:60:50, which was close to an ideal SCFA metabolism. Fatty acids produced
using RCM (
reinforced clostridial medium
) medium was similar to that of using
PYG (
pepton yeast glucose
) medium, although the levels were significantly
different among the hydrolysis methods used.
PERBANDINGAN METODE HIDROLISIS DALAM
AKTIVITAS PEMBENTUKAN ASAM LEMAK RANTAI
PENDEK PADA PATI TAKTERCERNA TIPE 3
DESI AWALINA
Skipsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Kimia
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi
: Perbandingan Metode Hidrolisis dalam Aktivitas Pembentukan
Asam Lemak Rantai Pendek pada Pati Taktercerna Tipe 3
Nama
: Desi Awalina
NIM
: G44086035
Disetujui
Pembimbing,
Prof Dr Suminar S Achmadi
NIP 19480427 197412 2 001
Diketahui
Ketua Departemen Kimia,
Prof Dr Ir Tun Tedja Irawadi, MS
NIP 19501227 197603 2 002
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala
karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Judul yang dipilih dalam
penelitian yang dilaksanakan sejak bulan Agustus 2010 ini ialah Perbandingan
Metode Hidrolisis dalam Aktivitas Pembentukan Asam Lemak Rantai Pendek
pada Pati Taktercerna Tipe 3.
Ucapan terima kasih kepada Ibu Prof Dr Suminar S Achmadi yang telah
membantu dan membimbing dalam penelitian ini. Terima kasih pula kepada Ibu
Prof Maggy T Suhartono, Ibu Ir Endang Yuli Purwani, MSi, dan Ibu Winda
Haliza beserta segenap karyawan Laboratorium Mikrobiologi dan Laboratorium
Kimia Balai Besar Penelitian Pascapanen, Bogor. Penelitian ini dapat
terselesaikan atas dorongan semangat dan bantuan dari sahabat-sahabatku Melly,
Kirty, Lia, Rania, dan Martha.
Bogor, Maret 2012
RIWAYAT HIDUP
Penulis dilahirkan di Bukittinggi pada tanggal 17 Desember 1983 dari ayah
Safri dan ibu Nurhayati. Penulis merupakan putri kedua dari tiga bersaudara.
Tahun 2002 penulis lulus dari SMUN 2 Bogor dan pada tahun yang sama
lulus seleksi masuk IPB pada program studi D3 Analisis Kimia, Fakultas
Matematika dan Ilmu Pengetahuan Alam. Tahun 2005 penulis lulus dari program
studi D3 Analisis Kimia, dan pada tahun yang sama penulis diterima bekerja
sebagai staf
Method and Development
di PT Eisai Indonesia.
DAFTAR ISI
Halaman
DAFTAR TABEL ... vii
DAFTAR GAMBAR ... vii
DAFTAR LAMPIRAN ... vii
PENDAHULUAN
Latar Belakang ... 1
Tujuan ... 1
METODE PENELITIAN
Alat dan Bahan ... 1
Lingkup Penelitian ... 2
Pencirian Pati Ubi Jalar ... 2
Retrogradasi Pati ... 2
Hidrolisis Pati dan Produksi RS3 ... 2
Pencirian dan Penentuan Derajat Polimerisasi RS3 ... 2
Fermentasi (
in vitro
) ... 4
Analisis Aktivitas SCFA ... 4
Analisis Statistika ... 4
HASIL DAN PEMBAHASAN
Ciri Kimia dan Visual Pati Ubi Jalar ... 4
Pati Retrogradasi ... 5
Hidrolisat Pati dan Produk RS3 ... 5
Ciri Kimia dan Derajat Polimerisasi RS3 ... 6
Produk Fermentasi (
in vitro
) dan Aktivitasnya ... 7
SIMPULAN ... 9
DAFTAR PUSTAKA ... ... 10
DAFTAR TABEL
Halaman
1 Ciri kimia sampel ubi jalar varietas cangkuang dibandingkan
dengan 2 varietas lainnya ... 4
2 Rendemen RS3 hasil hidrolisis menggunakan HCl 2 N,
α
-amilase, dan pululanase ... 5
3 Derajat polimerisasi produk hidrolisis dengan menggunakan
HCl 2 N,
α
-amilase, dan pululanase ... 7
4 Profil SCFA hasil fermentasi
in vitro
RS3 dengan
masing-masing 3 kali ulangan dalam satuan mol/L ... 8
5 SCFA temuan beberapa peneliti lainnya ... 8
6 Analisis statistik SCFA dari
C. butyricum
pada perlakuan
hidrolisis yang berbeda ... 9
7 Analisis statistik SCFA dari
E. rectale
pada perlakuan hidrolisis
yang berbeda ... 9
DAFTAR GAMBAR
Halaman
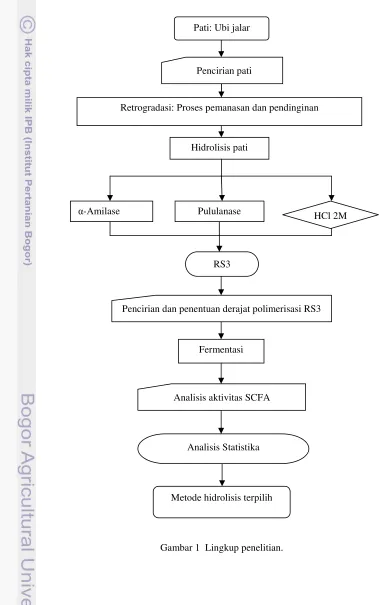
1 Lingkup penelitian ... 3
2 Mikroskop polarisasi pati ubi jalar varietas cangkuang dengan perbesaran
200
X
... 5
3 Skema hidrolisis pati menggunakan enzim ... 6
4 Kurva fraksionasi RS3 hasil hidrolisis untuk penentuan DP ... 7
DAFTAR LAMPIRAN
Halaman
1 Komposisi medium RCM dan PYG ... 11
PENDAHULUAN
Latar Belakang
Pentingnya kesehatan sistem pencernaan makin disadari seiring dengan perubahan pola konsumsi pangan dan gaya hidup yang penuh dengan tekanan (stres). Keadaan ini mendorong dilakukannya riset tentang kontribusi ingredien pangan dalam bentuk pati taktercerna tipe 3 (resistant starch type 3; selanjutnya disebut RS3) dalam mencegah kanker kolon (colorectal cancer; selanjutnya disebut CRC). RS didefinisikan sebagai fraksi pati yang tidak terserap oleh sistem pencernaan pada individu yang sehat (EURESTA 1993). Ada 4 kelompok RS, yaitu RS1, RS2, RS3, dan RS4 (Sajilata et al. 2006). RS1 adalah pati yang secara fisik terperangkap di dalam matriks sehingga tidak dapat diakses oleh enzim. RS2 adalah granul pati mentah yang didominasi oleh struktur kristalin sehingga sulit dihidrolisis oleh enzim. RS3 adalah pati retrogradasi dan RS4 adalah pati modifikasi secara kimiawi.
RS3 dipilih karena memiliki kemiripan proses dengan daya cerna pati dalam tubuh manusia. RS3 lolos dari sistem pencernaan yang sehat dan langsung memasuki usus besar selanjutnya difermentasi oleh mikroflora yang ada di dalamnya. Fermentasi dilakukan oleh konsorsium bakteri antara lain Eubacterium rectale dan Clostridium butyricum. Kelompok bakteri tersebut diketahui sangat menguntungkan; mereka memproduksi asam lemak rantai pendek (SCFA: short chain fatty acid), yaitu asam asetat, propionat, dan butirat (Bird et al. 2000, Ramsay et al. 2006). SCFA adalah sumber energi utama bagi sel-sel kolon yang mampu mengurangi risiko terjadinya CRC (Augenlicht et al. 2002). Dari uraian ini, sifat molekul RS mungkin saja memengaruhi produksi SCFA khususnya butirat oleh bakteri penghuni kolon. Jika SCFA meningkat, maka proliferasi sel CRC dapat dihambat. Oleh karena itu, peningkatan asupan RS dapat dijadikan dasar untuk pertumbuhan bakteri penghasil butirat. Kemampuan bakteri memfermentasi RS secara in vitro diharapkan dapat menggambarkan kemampuannya secara in vivo.
Berdasarkan percobaan Purwani dan Suhartono (2009), pati asal beras dan sagu memiliki potensi sebagai RS3 yang dapat memproduksi butirat secara enzimatis, dan secara in vitro dapat menghambat proliferasi sel CRC. Dengan asumsi tersebut, ubi jalar (Ipomoea batatas) yang juga merupakan
sumber karbohidarat akan berpotensi sebagai penghasil RS3 yang bersifat apoptosis terhadap sel CRC.
Modifikasi pati menjadi RS3 memerlukan tahap hidrolisis. Hidrolisis secara enzimatis memiliki perbedaan mendasar dengan hidrolisis secara asam. Hidrolisis secara asam memutus rantai pati secara acak, sedangkan hidrolisis secara enzimatis memutus rantai pati secara spesifik pada percabangan tertentu. Hidrolisis secara enzimatis lebih menguntungkan dibandingkan dengan hidrolisis asam karena prosesnya lebih spesifik, kondisi prosesnya dapat dikendalikan, biaya pemurnian lebih murah, dan kerusakan warna dapat diminimumkan.
Kuantitas butirat yang dihasilkan dari hidrolisis pati ubi jalar belum dikaji secara detail. Perbandingan hidrolisis dengan menggunakan α-amilase, pululanase, dan asam klorida 2 N dapat digunakan dalam produksi RS3 skala industri, dengan memperhatikan biaya dalam proses hidrolisisnya.
Informasi mengenai sifat fisiko-kimia ubi jalar juga masih sangat terbatas. Konsekuensi lebih lanjut adalah perlunya dilakukan uji sifat fungsional ubi jalar sebagai penghasil RS3 yang digunakan oleh bakteri untuk menghasilkan SCFA.
Tujuan
Penelitian ini bertujuan:
a. menghasilkan RS3 berbasis pati ubi jalar dengan 3 perlakuan hidrolisis yang dapat dimanfaatkan dengan baik oleh bakteri penghuni kolon (in vitro) serta mengevaluasi ciri molekulnya;
b. membandingkan profil SCFA dan enzim pendegradasi pati yang dihasilkan oleh bakteri penghasil butirat yang ditumbuhkan pada medium berisi RS3; dan
c. mengidentifikasi RS3 yang dapat digunakan oleh bakteri untuk menghasilkan SCFA secara optimum.
METODE PENELITIAN
Alat dan Bahan
19091-136 (60 m × 0.250 mm). Helium digunakan sebagai gas pembawa dengan laju 1.8 mL/menit. Nisbah split 40:1. Suhu oven dijaga pada 90 °C selama 0.5 menit, kemudian dinaikkan menjadi 110 °C dengan laju 10 °C/menit, dinaikkan lagi menjadi 170 °C dengan laju 5 °C/menit dan akhirnya dinaikkan menjadi 210 °C dengan laju 20 °C/menit. Suhu injektor dan detektor adalah 275 °C. Bahan-bahan yang digunakan adalah ubi jalar dari Balai Penelitian Tanaman Kacang-kacangan dan Umbi-umbian, Malang. Galur bakteri diperoleh dari koleksi kultur mikroorganisme Balai Besar Veteriner Bogor. Enzim yang digunakan adalah enzim α -amilase dan pululanase, enzim ini diperoleh dari NOVO melalui distributornya di Jakarta.
Lingkup Penelitian
Pati ubi jalar diperoleh melalui ekstraksi ubi jalar mentah menggunakan air bersih. Pencirian pati dilakukan terhadap 3 varietas pati berbeda, meliputi kadar air, abu, lemak, protein, serat kasar, dan amilosa.
RS3 dibuat dari pati retrogradasi yang diberi perlakuan panas dan dihidrolisis dengan enzim dan asam. RS3 kemudian ditetapkan ciri kimianya. Proses fermentasi diterapkan untuk mengevaluasi sifat fungsional RS3. Dua jenis bakteri penghasil butirat dilibatkan dalam fermentasi RS3, yaitu C. butyricum dan E. rectale; keduanya adalah bakteri penghasil butirat yang secara normal ditemukan di dalam kolon. Analisis aktivitas SCFA dilakukan untuk menentukan metode hidrolisis terbaik dalam produksi SCFA yang dilakukan. Skema penelitian dicantumkan dalam Gambar 1.
Pencirian Pati Ubi Jalar
Pencirian pati dilakukan terhadap 3 varietas pati berbeda, meliputi pencirian kadar air, abu, lemak, protein, serat kasar, amilosa, dan mikroskop polarisasi pati. Kadar air, abu, lemak, protein, serat kasar, dan amilosa dianalisis dengan metode standar AOAC (2000).
Retrogradasi Pati
Retrogradasi berupa proses pemanasan dan pendinginan pada pati. Sebanyak 50 g pati dilarutkan ke dalam 250 mL akuades kemudian dipanaskan hingga homogen dan mengental berbentuk gel. Gel dikemas dalam plastik tahan panas, udara di dalam plastik
dihilangkan dengan vakum, kemudian gel diautoklaf selama 30 menit pada suhu 121 °C. Gel didinginkan pada suhu ruang selama 1 jam, kemudian disimpan pada 4 °C selama 24 jam.
Hidrolisis Pati dan Produksi RS3
Hidrolisis untuk memotong polimer pati menggunakan enzim dan asam dilakukan pada produksi RS3. Setiap sampel hasil retrogradasi ditambah 1250 mL akuades dan dihaluskan dengan blender. Untuk hidrolisis asam, sampel dipanaskan pada suhu 90 °C sambil diaduk, lalu ditambahkan HCl 2 N hingga pH 1 pada suhu 90 °C dan diaduk selama 30 menit. Untuk hidrolisis enzimatik, enzim pululanase ditambahkan sebanyak 1.0 mL pada suhu 60 °C dan diaduk selama 3 jam, sedangkan enzim α-amilase ditambahkan sebanyak 0.1 mL pada suhu 45 °C dan diaduk selama 3 jam. Setiap sampel didinginkan pada suhu ruang selama 1 jam, kemudian disimpan pada 4 °C selama 24 jam. Sampel dicuci dan disaring, kemudian dibilas dengan 1000 mL akuades, dan disimpan kembali pada 4 °C selama 24 jam. Setelah dicuci dan disaring kembali, sampel dikeringkan menggunakan drum pengering.
Pencirian dan Penentuan Derajat Polimerisasi RS3
Kadar RS3 dianalisis menurut metode Goni et al. (1996). Analisis dilakukan secara triplo. Tahapan untuk analisis RS ialah sebagai berikut. Sampel (100 mg) ditambah dengan 10 mL KCl-HCl pH 1.5. Sebanyak 0.2 mL larutan pepsin (1 g pepsin/10 mL bufer KCl-HCl) ditambahkan ke dalamnya, dan diinkubasi 37 °C selama 1 jam dengan pengadukan konstan. Sampel ditambah dengan 9 mL bufer tris-maleat pH 6.9 (pH disesuaikan dengan HCl 2 M atau NaOH 0.5 M). Larutan enzim α-amilase (40 mg α -amilase/mL bufer tris-maleat) dan pululanase (40 mg pululanase/mL bufer tris-maleat) masing-masing ditambahkan ke dalamnya, diinkubasi pada 37 °C, 16 jam, dengan pengocokan konstan. Campuran disentrifus (3000 rpm, 15 menit), supernatan dibuang.
ditambahkan, diinkubasi selama 45 menit pada suhu 60 °C dengan pengadukan konstan. Campuran disentrifus (15 menit, 3000 rpm) dan supernatannya dikumpulkan di dalam labu takar. Residu dibilas dengan 10 mL akuades, bilasan disatukan dengan supernatan, kemudian ditera hingga volume 25‒1000 mL.
Bergantung pada kadar RS, sentrifus dapat diganti dengan filtrasi. Kadar glukosa total di dalam supernatan dihitung dengan metode fenol sulfat. Kadar RS dihitung sebagai total glukosa × 0.9.
Gambar 1 Lingkup penelitian. Metode hidrolisis terpilih
Hidrolisis pati
Retrogradasi: Proses pemanasan dan pendinginan
α-Amilase Pululanase HCl 2M
Pencirian dan penentuan derajat polimerisasi RS3 RS3
Pati: Ubi jalar
Pencirian pati
Fermentasi
Analisis aktivitas SCFA
Derajat polimerisasi (DP) pada setiap fraksi ditentukan dengan filtrasi gel pada kolom Sephadex G-25 (Lu et al. 1996). Total karbohidrat di setiap fraksi ditentukan dengan metode fenol-asam sulfat dan konsentrasinya dipantau pada panjang gelombang 490 nm (AOAC 2005). Nilai gula pereduksi ditentukan dengan metode dinitrosalisilat dan konsentrasinya dipantau pada panjang gelombang 540 nm (AOAC 2000). DP merupakan nisbah antara total karbohidrat dan nilai gula pereduksi.
Fermentasi (in vitro)
Dua galur bakteri yang digunakan untuk penelitian ialah C. butyricum dan E. rectale. Galur bakteri (dalam keadaan terliofilisasi) diaktifkan kemudian dipelihara di dalam medium sesuai dengan rekomendasi dari kolektornya. C. butyricum di dalam medium RCM (reinforced clostridial medium) dan E. rectale dalam medium PYG (pepton yeast glucose) yang dimodifikasi. Komposisi kedua jenis medium dicantumkan dalam Lampiran 1. Kultur disegarkan secara berkala. Kultur stok disiapkan untuk setiap galur.
Kultur stok diinokulasi ke dalam tabung yang berisi 20 mL medium RS3. Nilai pH medium diatur pada pH 7, sesuai dengan pH kolon. Tabung diinkubasi secara anaerob pada suhu 37 °C selama 48 jam. Setiap galur ditumbuhkan dalam medium basal yang mengandung 10 g RS3. Sel bakteri dipisahkan dengan cara sentrifugasi.
Analisis Aktivitas SCFA
SCFA diukur menggunakan GC dengan 3 kali ulangan. Standar yang digunakan dalam analisis adalah asetat, propionat, dan butirat. Kalibrasi standar dicantumkan dalam Lampiran 2.
Analisis Statistika
Analisis statistik yang diterapkan adalah ANOVA dua-arah yang dilanjutkan dengan uji pembeda Tukey. Perangkat lunak SPSS 15.0 digunakan untuk keperluan tersebut.
Korelasi antara aktivitas enzim dan produksi SCFA dengan perbandingan antara aktivitas enzim dan HCl yang digunakan juga dianalisis.
HASIL DAN PEMBAHASAN
Ciri Kimia dan Visual Pati Ubi Jalar
Sebelum digunakan sebagai bahan dasar pembuatan RS3, pati ubi jalar varietas cangkuang dianalisis ciri kimianya dan dibandingkan dengan 2 varietas pati ubi jalar lainnya. Berdasarkan pengamatan (Tabel 1), sampel yang digunakan telah memenuhi persyaratan dalam pembentukan RS3, yaitu kadar amilosa lebih besar dari 30.00% dengan kadar abu, lemak, protein, dan serat kasar yang sangat rendah. Banyak faktor yang memengaruhi terbentuknya RS3, termasuk pati yang tidak dapat diakses secara fisik. Struktur granular pati, dan derajat retrogradasi dapat membatasi kecernaan pati (Topping & Clifton 2001). Dua varietas lainnya, yaitu salosa dan ace tidak digunakan untuk analisis lebih lanjut karena kadar amilosa berada di bawah 30.00%. Retrogradasi pati akan menghasilkan polimer linear berantai pendek yang tidak larut dan bersifat resisten terhadap enzim pencernaan. Oleh karena itu, sumber pati beramilosa tinggi akan memiliki kadar RS yang tinggi pula.
Tabel 1 Ciri kimia sampel ubi jalar varietas cangkuang dibandingkan dengan 2 varietas lainnya
Komponen (%)
Varietas
Cangkuang Salosa Ace Air 19.10 10.53 10.30
Abu 0.90 0.55 0.21
Lemak 1.88 1.99 0.46
Gambar 2 Mikroskop polarisasi pati ubi jalar varietas cangkuang dengan perbesaran 200X.
Kisaran diameter pati varietas cangkuang yang dilihat dari mikroskop polarisasi dengan perbesaran 200X berada antara 8.0 µm dan 70.6 µm. Kisaran rerata dari 5 titik pengamatan varietas cangkuang sebesar 34.88 µ m. Kisaran diameter berpengaruh pada laju hidrolisis, yaitu laju hidrolisis akan semakin cepat bila ukuran granul pati semakin kecil. Secara visual, pati ubi jalar berwarna putih gading dan warna antar varietas tidak dapat dibedakan.
Pati Retrogradasi
Pati retrogradasi berbentuk gel bening dengan warna abu-abu. Proses pembuatan pati retrogradasi melalui 2 tahap, yakni (1) gelatinisasi yang menyebabkan struktur granul mengembang akibat penyerapan air yang berlebih selama pemanasan, (2) retrogradasi pati yang mengakibatkan rekristalisasi rantai
amilosa dan amilopektin. Proses gelatinisasi menyebabkan pati lebih mudah tercerna dibandingkan dalam bentuk mentahnya, tetapi gel pati tersebut tidak stabil dan membentuk kristal saat pendinginan. Peristiwa tersebut dinamakan retrogradasi. Menurut Wunderlich (1976), mekanisme retrogradasi melalui 3 tahapan, yakni nukleasi, pertumbuhan kristal, dan tahap penyempurnaan.
Hidrolisat Pati dan Produk RS3
Hidrolisat pati berupa cairan kuning kecokelatan, yakni gula sederhana yang kemudian dipisahkan dari fraksi pati yang tidak dapat terhidrolisis. Fraksi pati yang tidak dapat terhidrolisis ini yang kemudian disebut pati taktercerna (RS3). Rendemen (Tabel 2) paling tinggi didapat dari hidrolisis menggunakan enzim pululanase, yakni hampir 50%.
Tabel 2 Rendemen RS3 hasil hidrolisis menggunakan HCl 2 N, α-amilase, danpululanase
Bobot HCl 2 N α-Amilase Pululanase
Sebelum (g) 50.00 50.00 50.00
Sesudah (g) 3.31 1.14 24.05
Gambar 3 Skema hidrolisis pati menggunakan enzim (Mathewson 1998).
Enzim-enzim pencerna pati meliputi α- dan β amilase, glukoamilase, dan isoamilase atau pululanase (Mathewson 1998). Enzim α -amilase (EC 3.2.1.1) merupakan enzim penghidrolisis yang memotong ikatan glikosidik α-(1,4) secara acak dari bagian dalam molekul pati; enzim ini dikenal dengan endoamilase. Enzim β-amilase (EC 3.2.1.2) menghidrolisis ikatan β-(1,4) dari ujung nonpereduksi dan menghasilkan maltosa. Glukoamilase (EC 3.2.1.3) mengonversi maltosa dan fragmen besar pati (hasil hidrolisis α- dan β-amilase) menjadi glukosa. Isoamilase memotong ikatan α-(1,6). Skema hidrolisis pati menggunakan enzim terlihat pada Gambar 3.
Hidrolisis secara enzimatis memiliki perbedaan mendasar dengan hidrolisis secara asam. Hidrolisis secara enzimatis memutus rantai pati secara spesifik pada ikatan glikosidik tertentu, sedangkan hidrolisis secara asam memutus rantai pati secara acak. Pada hidrolisis dengan asam, parameter suhu dan konsentrasi asam saling berinteraksi. Semakin tinggi konsentrasi asam, semakin rendah suhu proses, dan sebaliknya. Suhu dan konsentrasi asam terbaik untuk proses likuifikasi pati ubi jalar jenis unggul adalah 95 °C dan pH 1 (Ega 2002).
Ciri Kimia dan Derajat Polimerisasi RS3
Kadar pati taktercerna yang didapat setelah proses hidrolisis berturut-turut menggunakan HCl 2 N, α-amilase, dan
pululanase adalah 15%, 20%, dan 12%. Beberapa faktor yang dapat memengaruhi terjadinya perbedaan kadar pati taktercerna, di antaranya pemotongan rantai pada saat hidrolisis, kisaran diameter pati, dan faktor penghambat hidrolisis seperti adanya lipid dan protein. Adanya kompleks rantai dengan lipid dapat menghambat pembentukan RS (Mangala et al. 1999).
DP merupakan jumlah struktur berulang dalam rantai polimer. Polimer dengan DP besar (>104) disebut polimer tinggi, sedangkan polimer dengan DP rendah (<104) disebut oligomer. Derajat polimerisasi RS3 hasil hidrolisis dengan HCl 2 N, α-amilase, dan pululanase secara berturut-turut sebagai berikut: 15, 18, dan 25 residu. Setelah hidrolisis, rantai glukosa RS3 yang dibentuk lebih pendek dibandingkan dengan rantai glukosa pati. Salah satu syarat pembentukan RS adalah rantai glukosa pati dengan panjang rantai 30‒40 residu (Mangala et al. 1999). Hidrolisis rantai amilosa oleh enzim berlangsung secara terus-menerus hingga tersisa rantai glukosa 10‒20 unit. Pada rantai amilopektin, enzim α-(1,4) bekerja dengan cara menyisakan rantai glukosa 10-20 unit dan menyisakan fragmen percabangan yang mengandung ikatan glikosidik α-(1,4) (Mathewson 1998). Penentuan DP berdasarkan nisbah antara nilai total karbohidrat dan nilai gula pereduksi pada hasil hidrolisis setiap sampel. Derajat polimerisasi RS3 terlihat pada Tabel-3. Ujung pati mereduksi
α
- amilase
α
- amilase
pululanase
Tabel 3 Derajat polimerisasi produk hidrolisis dengan menggunakan HCl 2 N, α-amilase, dan pululanase
Gula pereduksi (mg/mL) Total karbohidrat (mg/mL) DP
HCl 2 N 4.00 58.05 14.5
α-Amilase 3.00 52.48 17.5
Pululanase 2.32 57.08 24.6
Kurva fraksionasi menggambarkan besarnya DP berbanding lurus dengan area kurva yang terbentuk (Gambar 4). Semua hasil hidrolisis pati yang membentuk RS3 pada penelitian ini memiliki DP < 104, sehingga dapat digolongkan ke dalam oligomer. Data ini sejalan dengan perhitungan DP melalui nisbah total karbohidrat:gula pereduksi pada Tabel 3, yaitu tidak lebih dari 25 residu.
Produk Fermentasi (in vitro) dan Aktivitasnya
Fermentasi in vitro RS3 menghasilkan asam lemak rantai pendek, yakni asetat, propionat, dan butirat. Profil SCFA yang dihasilkan dengan menggunakan 3 macam perlakuan hidrolisis dan 2 macam bakteri penghasil butirat tercantum dalam Tabel 4. Hasil fermentasi selain SCFA adalah gas CO2, H2, dan CH4 serta produk lain seperti asam laktat, etanol, dan suksinat yang dibentuk oleh
tipe bakteri yang berbeda. Secara khusus, SCFA terutama butirat adalah sumber energi utama bagi sel-sel kolon. Hampir 70‒90% SCFA dimetabolisme oleh sel kolon. Metabolisme ideal SCFA terjadi pada nisbah butirat:propionat:asetat sebesar 90:30:50. Sekitar 70% konsumsi oksigen sel kolon diperoleh dari oksidasi SCFA terutama butirat (Cook & Sellin 1998). Penelitian ini menghasilkan nisbah butirat:propionat:asetat rerata sebesar 130:60:50. Hasil tersebut mendekati metabolisme ideal SCFA.
Produksi SCFA menggunakan media RCM jumlahnya mirip dengan produksi SCFA menggunakan media PYG pada setiap perlakuan hidrolisis, walaupun terdapat ragam kadar yang sangat nyata akibat perbedaan perlakuan hidrolisis yang digunakan. SCFA yang diperoleh dalam penelitian ini cocok dengan SCFA yang telah ditemukan beberapa peneliti lainnya yang dicantumkan dalam Tabel-5.
Tabel 4 Profil SCFA hasil fermentasi in vitro RS3 dengan masing-masing 3 kali ulangan dalam satuan mol/L
Media Hidrolisis Statistika Asetat Propionat Butirat
RCM
HCl 2 N Rerata 0.04 0.06 0.13
SD 0.01 0.02 0.03
α-Amilase Rerata 0.07 0.05 0.12
SD 0.02 0.02 0.04
Pululanase Rerata 0.06 0.07 0.13
SD 0.01 0.01 0.02
PYG
HCl 2 N Rerata 0.04 0.09 0.12
SD 0.02 0.03 0.02
α-Amilase Rerata 0.07 0.06 0.14
SD 0.01 0.02 0.02
Pululanase Rerata 0.03 0.07 0.12
SD 0.02 0.02 0.03
Tabel 5 SCFA temuan beberapa peneliti lainnya
Penelitian fermentasi Kondisi Medium Fermentasi SCFA (mmol/L) Asetat Butirat Reid et al.
1996
39 oC, 48 jam, pH 6,
50% CO2, 50% N2
Nutrient broth (Unipath, UK) + 0.5% pati kentang retrogradasi
4.6 23.57
Nutrient broth (Unipath, UK) + 1.0% pati kentang retrogradasi
8.43 36.17
Nutrient broth (Unipath, UK) + 0.5% pati kentang retrogradasi + 0.5 mg/mL
pankreatin
14.72 49.72
Nutrient broth (Unipath, UK) + 1.0% pati kentang retrogradasi + 0.5 mg/mL
pankreatin
33.77 76.51
Vandak et al. 1995
37 oC, pH 6.5, agitasi 150 rev/min,
N2
(g/L) : KH2PO4, 2; (NH4)2SO4, 2; MgSO4·7H2O, 0.6; FeSO4·7H2O, 0.03;
MnSO4·7H2O, 0.01; dadih, 30
33.31 112.36
(g/L) : KH2PO4, 2; (NH4)2SO4, 2; MgSO4·7H2O, 0.6; FeSO4·7H2O, 0.03; MnSO4·7H2O, 0.01; dadih, 30; ekstrak
ragi, 5
79.9 211.1
(g/L) : KH2PO4, 2; (NH4)2SO4, 2; MgSO4·7H2O, 0.6; FeSO4·7H2O, 0.03; MnSO4·7H2O, 0.01; dadih, 30 + biotin 50
µg/L
76.6 210
Purwani & Suhartono
2009
37 oC, botol tertutup, 48
jam
(g/L): ekstrak ragi, 3; bubuk daging, 10; pepton, 10; larutan kanji, 1; natrium
klorida, 5; natrium asetat, 3; sistein hidroklorida, 0.5; RS3 pati sagu dengan perlakuan pululanase, 2; Medium pH 6.8.
61.77 6.15
(g/L): ekstrak ragi, 3; bubuk daging, 10; pepton 10; larutan kanji, 1; natrium klorida, 5; natrium asetat, 3; sistein hidroklorida, 0.5; RS3 pati beras dengan perlakuan pululanase, 2; Medium pH 6.8.
Produksi SCFA tidak dipengaruhi oleh kadar RS3 substratnya, tidak juga dipengaruhi oleh DP dan rendemen yang dihasilkan setelah proses hidrolisis dengan 3 perlakuan yang berbeda. Hasil tersebut dikuatkan dengan analisis statistika lanjut (Tabel 6 dan 7).
Perlakuan hidrolisis pada media RCM memberikan hasil di bawah 0.05 pada tingkat kepercayaan 95%. Hal tersebut menjelaskan bahwa perlakuan hidrolisis tidak mempengaruhi produksi SCFA pada media RCM. Kejadian yang sama juga dapat dilihat pada media PYG pada Tabel 7.
SIMPULAN
Ubi jalar varietas cangkuang mampu memproduksi RS3 dengan kemampuan pembentukan SCFA. Dibandingkan dengan HCl 2N dan α-amilase, pululanase merupakan katalis terbaik dalam pembentukan RS3. Penggunaan medium RCM dan PYG tidak menghasilkan SCFA yang berbeda nyata pada 3 macam perlakuan hidrolisis yang digunakan yaitu HCl 2N, α-amilase, dan pululanase. Tidak ada pengaruh kadar RS3 dan derajat polimerisasi dalam produksi SCFA.
Produksi SCFA menggunakan katalis pululanase dalam hidrolisis pembentukan RS3 disarankan karena menghasilkan rendemen 7 kali lebih banyak dibandingkan katalis HCl 2N dan 21 kali lebih banyak dibandingkan katalis α-amilase, untuk digunakan dalam produksi SCFA.
Tabel 6 Analisis statistik SCFA dari C. butyricum pada perlakuan hidrolisis yang berbeda
Perlakuan (i) Perlakuan (j) Perbedaan
rerata (i-j) Galat baku Sig Tingkat kepercayaan 95%
HCl 2 N α-Amilase -0.0056 0.00912 0.817 0.0177
Pululanase -0.0100 0.00912 0.528 0.0133
α-Amilase Pululanase -0.0044 0.00912 0.878 0.0188
HCl 2 N 0.0056 0.00912 0.817 0.0288
Pululanase α-Amilase 0.0044 0.00912 0.878 0.0277
HCl 2 N 0.0100 0.00912 0.528 0.0333
Tabel 7 Analisis statistik SCFA dari E. rectale pada perlakuan hidrolisis yang berbeda
Perlakuan (i) Perlakuan (j) Perbedaan
rerata (i-j) Galat baku Sig Tingkat kepercayaan 95%
HCl 2 N α-Amilase -0.0078 0.01058 0.746 0.0192
Pululanase 0.0111 0.01058 0.556 0.0381
α-Amilase Pululanase 0.0189 0.01058 0.203 0.0259
HCl 2 N 0.0078 0.01058 0.746 0.0348
Pululanase α-Amilase -0.0189 0.01058 0.203 0.0081
DAFTAR PUSTAKA
[AOAC] Association of Official Analytical Chemists. 2005. Official Methods of Analysis. Ed ke-18. Maryland: AOAC. Augenlicht et al. 2002. Short chain fatty acid
and colon cancer. Am Soc Nutr Sci Supl 3804-3808.
Bird A, Brown IL, Topping DL. 2000. Starches, resistant starches, the gut microflora and human health. Curr Issues Intest Microbiol 1:25-37.
Cook SI, Sellin JH. 1998. Review article: Short chain fatty acids in health and desease. Aliment Pharmacol Ther 12:499-507.
[EURESTA] European Flair Concerted Action on Resistant Starch. 1993. Wageningen: Wageningen University. Ega L. 2002. Kajian sifat fisik dan kimia serta
pola hidrolisis pati ubi jalar jenis unggul secara enzimatis dan asam [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Goni I, Garcia-Diz L, Manas E, Saura-Calixto F. 1996. Analysis of resistant starch: A method for foods and food products. Food Chem 56:445-449.
Lu S, Ching-Yung C, Cheng-Yi L. 1996. Gel chromatography fractionation and thermal characterization of rice starch affected by hydrothermal treatment. Cereal Chem 73:5-11.
Mangala SL, Udayasankar K, Tharanathan RN. 1999. Resistant starch from processed cereals: The influence of amylopectin and
non-carbohydrate constituents in its formation. Food Chem 64:391-396. Mathewson RP. 1998. Common enzyme
reaction. Cereal Food World 47:798-807. Purwani EY, Suhartono MT. 2009. Study on
resistant starch type 3 derived from rice and sago as functional food ingredient with capability to prevent colorectal cancer (CRC) disease [laporan akhir]. Jakarta: Indonesian Danone Institute Foundation, (tidak dipublikasikan). Ramsay AG, Scott KP, Martin JC, Rincon
MT, Flint HJ. 2006. Cell-associated α -amylases of butyrate-producing Firmicute bacteria from the human colon. Microbiol 152:3281-3290.
Reid CA, Hillman K, Henderson C, Glass H. 1996. Fermentation of native and processed starches by the porcine fecal anaerobe CIostridium butyricum (NCIMB 7423). J Appl Bacteriol 80:191-198. Sajilata MG, Singhai RS, Kulkarni PR. 2006.
Resistant starch-a review. Comp Rev Food Sci Food Safety 5:1-17.
Topping DL, Clifton PM. 2001. Short-shain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol Rev 81:1031-1064.
Vandak D, Tomaska M. Zigova J, Sturdik E. 1995. Short note: Effect of growth supplement and whey pretreatment on butyric acid production by Clostridium butyricum. World J Microbiol Biotechnol 11:363.
Lampiran 1 Komposisi medium RCM dan PYG
Medium pertumbuhan
C. butyricum
yang digunakan adalah RCM
(
reinforced clostridial medium
), selanjutnya disebut sebagai medium basal.
Komposisi RCM
Komponen
Jumlah (g/liter)
Ekstrak ragi
3.0
Serbuk `Lab-Lemco’ (ekstrak kaldu sapi)
10.0
Pepton
10.0
Glukosa
5.0
Larutan kanji
1.0
Natrium klorida
5.0
Natrium asetat
3.0
Sistein hidroklorida
0.5
Seluruh bahan tersebut, kecuali sistein hidroklorida dicampur, kemudian pH
medium diatur menjadi 6.8 dan ditambahkan indikator potensial redoks resazurin
sebesar 0.1 mL/100 mL. Penambahan resazurin akan menyebabkan warna
medium menjadi merah muda saat dipanaskan. Campuran medium tersebut
dididihkan hingga indikator resazurin berubah warna dari merah muda menjadi
tidak berwarna, kemudian didinginkan dengan diembusi gas CO
2.Setelah dingin,
sistein hidroklorida ditambahkan dan diaduk rata. Medium dimasukkan ke dalam
tabung reaksi bertutup, kemudian disterilisasi pada 121 °C selama 20 menit.
Media dapat disimpan pada suhu beku sebelum digunakan.
Medium pertumbuhan
E. rectale
yang digunakan adalah PYG (
pepton yeast
glucose
) yang dimodifikasi.
Komposisi medium PYG
Komponen
Jumlah
Unit
Tripticase pepton
5.00
g
Pepton
5.00
g
Ekstrak ragi
10.00
g
Ekstrak daging
5.00
g
Glukosa
5.00
g
K
2HPO
42.00
g
Tween 80
1.00
mL
Resazurin
1.00
mg
Larutan garam (lihat bawah)
40.00
mL
Akua distilata
950.00
mL
Larutan haemin (lihat bawah)
10.00
mL
Larutan Vitamin K1 (lihat bawah)
0.20
mL
Larutan campuran mineral
Mineral
Jumlah
Unit
CaCl
2·2 H
2O
0.25
g
MgSO4·7 H2O
0.50
g
K2HPO4
1.00
g
KH
2PO
41.00
g
NaHCO
310.00
g
NaCl
2.00
g
Akuades
1000.00
mL
Larutan Haemin:
Sebanyak 50 mg haemin dilarutkan dalam 1 mL NaOH 1 N, dijadikan 100
mL dengan akuades, dan disimpan dalam lemari pendingin.
Vitamin K1
Sebanyak 0.1 mL vitamin K1 dilarutkan dalam 20.00 mL etanol 95%, dan
disaring-steril. Setelah itu, disimpan di dalam botol cokelat.
Lampiran 2 Kalibrasi standar SCFA
Kurva standar
Std (µL/mL)
Asetat
Propionat
Butirat
10
2.62544
7.26075
9.53571
20
9.08551
13.71954
17.43600
30
17.41830
22.54874
26.73470
50
28.46358
40.34441
43.52949
Kurva standar asetat
Kurva standar propionat
y= 0.651x- 3.512
R² = 0.993
0 5 10 15 20 25 30 35
0 10 20 30 40 50 60
A
re
a
µL/mL
y= 0.839x- 2.105
R² = 0.996
0 5 10 15 20 25 30 35 40 45
0 10 20 30 40 50 60
A
re
a
Kurva standar butirat
Analisis sampel
Media
Perlakuan
Hidrolisis
Ulangan
Konsentrasi (mol/L)
Asetat
Propionat
Butirat
RCM
α
-Amilase
1
0.09
0.04
0.15
2
0.06
0.05
0.08
3
0.07
0.07
0.12
pululanase
1
0.05
0.06
0.14
2
0.07
0.08
0.14
3
0.06
0.06
0.11
HCl 2 N
1
0.04
0.06
0.12
2
0.04
0.08
0.16
3
0.03
0.04
0.11
PYG
α
-Amilase
1
0.06
0.04
0.14
2
0.06
0.07
0.12
3
0.08
0.08
0.16
pululanase
1
0.05
0.06
0.09
2
0.02
0.05
0.15
3
0.01
0.09
0.12
HCl 2 N
1
0.02
0.05
0.11
2
0.04
0.11
0.11
3
0.05
0.10
0.15
y= 0.855x+ 0.781
R² = 0.999
0 5 10 15 20 25 30 35 40 45 50
0 10 20 30 40 50 60
A
re
a