DETEKSI KERUSAKAN AKROSOM SPERMATOZOA
DOMBA DENGAN TEKNIK HISTOKIMIA LEKTIN
SELAMA PROSES PEMBEKUAN
LISA DWI FANNESSIA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Deteksi Kerusakan Akrosom Spermatozoa Domba dengan Teknik Histokimia Lektin selama Proses Pembekuan adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
LISA DWI FANNESSIA. Deteksi Kerusakan Akrosom Spermatozoa Domba dengan Teknik Histokimia Lektin selama Proses Pembekuan. Dibimbing oleh MOHAMAD AGUS SETIADI, NI WAYAN KURNIANI KARJA dan I KETUT MUDITE ADNYANE.
Proses pembekuan dapat menyebabkan kerusakan pada membran plasma dan akrosom spermatozoa sehingga dapat menurunkan fertilitas spermatozoa. Penelitian bertujuan untuk mengevaluasi kerusakan akrosom dengan menggunakan teknik histokimia lektin selama proses pembekuan. Semen dikoleksi dari domba berumur 1-2 tahun dua kali dalam satu minggu menggunakan vagina buatan. Segera setelah ditampung, semen dievaluasi karakteristiknya kemudian diencerkan dengan medium Niwa dan Sasaki Freezing (NSF).
Semen dikemas di dalam straw mini (0,25 ml) dan diekuilibrasi pada suhu 4oC selama dua jam. Straw kemudian dibekukan serta disimpan dalam tabung N2 cair. Evaluasi karakteristik spermatozoa (motilitas, viabilitas, dan membran plasma utuh) dan status akrosom dilakukan selama proses pembekuan. Deteksi status akrosom spermatozoa diamati dengan menggunakan metode pewarnaan histokimia lektin yaitu metode Fluorescens isothiocyanate (FITC) dan Avidin-Biotin-Complex (ABC). Data karakteristik spermatozoa dan status akrosom spermatozoa dianalisis dengan analisis sidik ragam (ANOVA).
Persentase motilitas, viabilitas dan membran plasma utuh spermatozoa sebelum pembekuan (83 ± 2,7%; 88,8 ± 2,6%; 88,2 ± 3,7%) mengalami penurunan (P<0,05) setelah ekuilibrasi (71 ± 4,2%; 84,2 ± 5,0%; 76,2 ± 1,3%) dan setelah thawing (40 ± 3,5%; 61,08 ± 3,3%; 51,2 ± 10,4%). Persentase akrosom spermatozoa intak dengan metode FITC dan ABC selama proses pembekuan masing-masing adalah 93,63 ± 2,73%; 88,04 ± 3,2% dan 81,73 ± 4,77% VS 94,54 ± 0,26%; 88,17 ± 0,38% dan 79,38 ± 2,06%. Dapat disimpulkan bahwa hasil penelitian menunjukkan terjadi penurunan kualitas spermatozoa selama proses pembekuan. Lebih lanjut, status akrosom spermatozoa dapat dideteksi dengan baik menggunakan kedua metode pewarnaan histokimia lektin.
SUMMARY
LISA DWI FANNESSIA. Detection of Acrosomal Damage of Ram Spermatozoa using Lectin Histhochemical Technique during Freezing Process. Supervised by MOHAMAD AGUS SETIADI, NI WAYAN KURNIANI KARJA and I KETUT MUDITE ADNYANE.
Freezing process caused damage of spermatozoa plasma membrane and acrosome that leading to the decrease of spermatozoa fertility. Research was conducted to evaluate acrosomal damage during freezing process by using lectin histhochemical technique. Semen was collected twice a week using artificial vagina from 1-2 years old ram. Immediately after collection, characteristic of semen quality was evaluated then diluted with Niwa and Sasaki Freezing (NSF) medium. Semen was loaded into 0.25 ml mini straws and equilibrated at 4oC for two hours. Straws were then frozen and stored in liquid nitrogen. Evaluation of sperm characteristic (motility, viability and plasma membrane integrity) and acrosomal damage were done during freezing process.
Detection of acrosomal integrity was observed using Fluorescens isothiocyanate (FITC) and Avidin-Biotin-Complex (ABC) staining methods. Data of characteristic spermatozoa and acrosomal integrity were analyzed using ANOVA. Result of the experiments showed that the percentage of motility, viability and plasma membrane integrity of spermatozoa before freezing (83 ± 2.7%; 88.8 ± 2.6%; 88.2 ± 3.7%) were significantly decreased (P<0.05) after equilibration (71 ± 4.2%; 84.2 ± 5.0%; 76.2 ± 1.3%) and after thawing (40 ± 3.5%; 61.08 ± 3.3%; 51.2 ± 10.4%).
The percentage of acrosomal intact spermatozoa using FITC and ABC methods during freezing process were 93.63 ± 2.73%; 88.04 ± 3.2% and 81.73 ± 4.77% VS 94.54 ± 0.26%; 88.17 ± 0.38% and 79.38 ± 2.06% respectively. In conclusion, the characteristic of spermatozoa were significantly decrease (P<0.05) during freezing process. Furthermore, the integrity of acrosome spermatozoa during freezing process can be well analyzed using lectin histhochemical staining methods.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Reproduksi
DETEKSI KERUSAKAN AKROSOM SPERMATOZOA
DOMBA DENGAN TEKNIK HISTOKIMIA LEKTIN
SELAMA PROSES PEMBEKUAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
Judul Tesis : Deteksi Kerusakan Akrosom Spermatozoa Domba dengan Teknik Histokimia Lektin selama Proses Pembekuan
Nama : Lisa Dwi Fannessia NIM : B352120021
Disetujui oleh Komisi Pembimbing
Prof Dr Drh Mohamad Agus Setiadi Ketua
Drh Ni Wayan Kurniani Karja, MP., PhD Anggota
DDrh I Ketut Mudite Adnyane, MSi., PhD., PAVet Anggota
Diketahui oleh
Ketua Program Studi Biologi Reproduksi
Prof Dr Drh Mohamad Agus Setiadi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan November 2013 ini ialah Status Akrosom, dengan judul Deteksi Kerusakan Akrosom Spermatozoa Domba dengan Teknik Histokimia Lektin selama Proses Pembekuan.
Terima kasih penulis ucapkan kepada Prof Dr Drh Mohamad Agus Setiadi sebagai ketua komisi pembimbing, Drh Ni Wayan Kurniani Karja, MP., PhD dan Drh I Ketut Mudite Adnyane, MSi., PhD., PAVet selaku anggota komisi pembimbing atas bimbingan, juga perhatian dan nasehatnya.
Penghargaan dan ucapan terima kasih penulis sampaikan kepada semua staf pengajar dan karyawan Program Studi Biologi Reproduksi Sekolah Pascasarjana Institut Pertanian Bogor yang telah membantu dan memberi dukungan kepada penulis sampai selesainya penyusunan tesis ini. Selanjutnya, ucapan terima kasih penulis sampaikan kepada rekan-rekan seperjuangan pada Program Studi BRP dan IBH 2012.
Rasa hormat dan terima kasih penulis persembahkan kepada orang tua dan seluruh keluarga atas doa dan kasih sayangnya. Demikian pula kepada pihak yang tidak bisa disebutkan satu persatu yang telah memberikan perhatian, saran serta kritik yang membangun penulis mengucapkan terima kasih dan penghargaan yang setinggi-tingginya.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL x
DAFTAR GAMBAR x
DAFTAR LAMPIRAN x
1 PENDAHULUAN 1
Latar Belakang 1
Kerangka Pemikiran 2
Tujuan Penelitian 3
Manfaat Penelitian 3
2 TINJAUAN PUSTAKA 3
Spermatozoa 3
Akrosom 5
Karbohidrat, Glikokonjugat dan Lektin 7
Kriopreservasi Spermatozoa 10
3 METODE 11 Waktu dan Tempat Penelitian 11 Koleksi Semen 11 Kriopreservasi Spermatozoa 11 Evaluasi Karakteristik dan Deteksi Status Akrosom 11
Analisis Data 15
4 HASIL DAN PEMBAHASAN 15
Karakteristik Spermatozoa selama Proses Pembekuan 15
Deteksi Kerusakan Akrosom Spermatozoa 18
5 SIMPULAN 20
DAFTAR PUSTAKA 20
LAMPIRAN 25
RIWAYAT HIDUP 29
DAFTAR TABEL
1 Jenis lektin, sumber dan afinitasnya terhadap karbohidrat 9
DAFTAR GAMBAR
1 Spermatozoa dengan bagian-bagiannya 5
2 Skematik daerah kepala spermatozoa mamalia 6
3 Tahap perubahan spermatid 7
4 Spermatozoa dengan pewarnaan metode eosin-nigrosin 13 5 Gambaran integritas membran spermatozoa domba 14 6 Karakteristik spermatozoa domba selama proses pembekuan 16 7 Status akrosom spermatozoa domba dengan pewarnaan histokimia
lektin 18
8 Status akrosom spermatozoa rusak selama proses pembekuan 19
DAFTAR LAMPIRAN
1 Komposisi medium Niwa dan Sasaki Freezing untuk kriopreservasi
spermatozoa 26
2 Prosedur pewarnaan histokimia lektin metode ABC (Avidin Biotin
Complex) 27
3 Prosedur pewarnaan histokimia lektin metode FITC (Fluorescens
1
PENDAHULUAN
Spermatozoa/semen biasanya disimpan dalam bentuk beku sehingga dapat digunakan dalam jangka waktu yang tidak terbatas. Namun demikian, spermatozoa mamalia sangat rentan mengalami kerusakan sel selama proses pembekuan (Partyka et al. 2011). Pada umumnya masalah selama proses pembekuan berkisar pada dua kejadian utama yaitu cold shock dan perubahan-perubahan intraseluler akibat pengeluaran air yang berhubungan dengan pembentukan kristal es. Kerusakan sel spermatozoa akibat adanya kejadian tersebut diantaranya kerusakan akrosom, kerusakan organ intraseluler, perubahan permeabilitas membran serta penurunan motilitas dan viabilitas (Gazali dan Tambing 2002).
Partyka et al. (2010) dan Hernandez et al. (2012) melaporkan bahwa terjadi kenaikan kerusakan akrosom spermatozoa selama proses pembekuan. Kerusakan akrosom dapat digunakan sebagai indikator salah satu penyebab berkurangnya fungsi spermatozoa (Esteves dan Verza Jr 2011). Spermatozoa harus mempunyai akrosom dalam kondisi utuh guna memicu proses fertilisasi, karena akrosom spermatozoa yang telah mengalami kerusakan tidak dapat membuahi sel telur (Esteves et al. 2007). Oleh karena itu, diperlukan teknik untuk mendeteksi kerusakan akrosom yang efektif dan akurat.
Akrosom merupakan derivat dari apparatus Golgi yang terbentuk pada tahap awal spermiogenesis, utamanya terjadi pada fase spermatid dan terletak pada bagian anterior dari kepala spermatozoa serta terdiri dari enzim-enzim protease, diantaranya proakrosin, hyaluronidase dan phospholipase (Toshimori dan Ito 2003; Florman et al. 2008; Esteves dan Verza Jr 2011). Akrosom memegang peranan penting dalam proses fertilisasi. Inisiasi ikatan spermatozoa dengan zona pelusida akan memicu terjadinya reaksi akrosom dan menyebabkan pelepasan serta aktivasi dari enzim akrosom, sehingga spermatozoa mampu melakukan penetrasi zona pelusida (Inoue et al. 2005; Miranda et al. 2009).
Spermatozoa dari kepala sampai ekor dilapisi oleh membran dengan struktur yang sangat kompleks dalam susunan mosaik yang teratur dan memiliki peran biologik spesifik pada permukaannya. Membran plasma spermatozoa diperkirakan terdiri dari 300 protein yang berbeda dan sekitar 92% protein membran ekstraseluler pada semua sel eukariotik berupa glikokonjugat (Schroter et al. 1999), termasuk yang terdapat pada membran akrosom luar spermatozoa. Akrosom mengandung glikoprotein yang disekresikan oleh retikulum endoplasma dan apparatus Golgi, demikian pula dengan enzim-enzim yang digunakan pada saat penetrasi spermatozoa, seperti akrosin dan hyaluronidase (Curry dan Watson 1995).
2
Teknik histokimia lektin telah banyak digunakan untuk mendeteksi status akrosom spermatozoa pada banyak spesies seperti babi (Siciliano et al. 2008), tikus (Lybaert et al. 2009), ayam (Partyka et al. 2010), kuda (Cocchia et al. 2011), sapi (Odhiambo et al. 2011), kambing (Batista et al. 2011) dan Canada goose (Partyka et al. 2011). Pada penelitian ini dilakukan analisa kerusakan akrosom spermatozoa domba selama proses pembekuan menggunakan dua metode pewarnaan histokimia lektin yaitu metode Fluorescens isothiocyanate (FITC) dan metode Avidin-Biotin-Complex (ABC). Metode FITC merupakan metode pewarnaan single-step dimana lektin yang telah dilabel fluoresens akan berikatan langsung dengan karbohidrat spesifik yang terdapat pada permukaan sel dan diamati dengan menggunakan mikroskop fluoresens, sementara pada metode ABC melibatkan afinitas terhadap molekul avidin-biotin membentuk ikatan kompleks permanen untuk selanjutnya digunakan kromogen diamino benzidine (DAB) sebagai substansi penanda membentuk kompleks dengan enzim peroksidase, dan kompleks yang terbentuk dalam kromogen DAB menghasilkan warna coklat yang dapat diamati menggunakan mikroskop cahaya (Hsu et al. 1981; Maji et al. 2010).
Kerangka Pemikiran
Pembekuan spermatozoa adalah suatu proses penghentian sementara kegiatan hidup dari sel tanpa mematikan fungsi sel, reaksi metaboliknya berhenti mendekati total. Dalam proses pembekuan semen, akan terbentuk kristal-kristal es, terjadi penumpukan elektrolit dan bahan terlarut lainnya di dalam larutan atau di dalam sel. Kristal es intraseluler dapat merusak spermatozoa secara mekanik, sementara konsentrasi elektrolit yang berlebihan akan melarutkan selubung lipoprotein dinding sel spermatozoa pada waktu thawing sehingga permeabilitas membran sel spermatozoa akan berubah yang memicu terjadinya kerusakan, termasuk mengakibatkan kerusakan akrosom spermatozoa. Lebih lanjut, kerusakan akrosom spermatozoa menyebabkan kegagalan kemampuan spermatozoa untuk membuahi ovum.
3 cukup kuat dan reversible pada permukaan sel tanpa masuk ke dalam sel, maka lektin semakin diketahui mempunyai kemampuan untuk dapat mengevaluasi struktur membran maupun fungsi glikokonjugat.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengevaluasi kerusakan akrosom spermatozoa dengan menggunakan teknik histokimia lektin selama proses pembekuan.
Manfaat Penelitian
Hasil penelitian diharapkan dapat memberikan manfaat antara lain:
1. Memberikan informasi metode terbaik untuk deteksi kerusakan akrosom spermatozoa
2. Memberikan gambaran kualitas spermatozoa yang lebih spesifik dan akurat selama proses pembekuan
2
TINJAUAN PUSTAKA
Spermatozoa
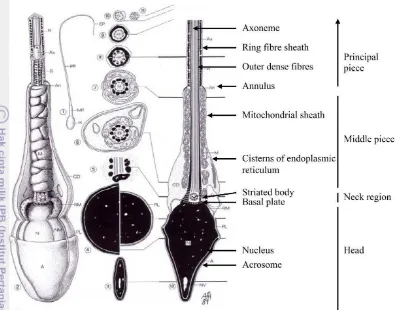
Spermatozoa merupakan gamet jantan yang diproduksi oleh tubuli seminiferi testis melalui proses yang disebut spermatogenesis. Struktur spermatozoa terdiri atas tiga bagian yaitu kepala, bagian tengah dan ekor (Gambar 1). Kepala spermatozoa sebagai pembawa materi inti berperan dalam menerjemahkan informasi genetik. Bagian tengah ekor terdapat sejumlah mitokondria yang dipercaya dapat memberikan energi untuk daya gerak. Bagian ini mengandung sebagian besar mekanisme daya gerak spermatozoa dan memiliki peranan yang sangat penting terhadap motilitas. Spermatozoa sebagai hasil akhir proses spermatogenesis merupakan sel yang berbentuk memanjang dengan bagian kepala sedikit pipih dan ekor yang panjang (Garner dan Hafez 2000). Lebih lanjut untuk proses fertilisasi, spermatozoa harus mempunyai cukup energi untuk pergerakan, protein dan senyawa lain yang penting selama berada dalam saluran reproduksi betina sehingga dapat melakukan fertilisasi.
4
dalam penembusan corona radiata dan zona pellucida pada proses fertilisasi (Toshimori dan Ito 2003; Bearden et al. 2004).
Ekor spermatozoa terdiri atas bagian leher (neck), tengah (middle), principal dan ujung (end). Fungsi dari ekor spermatozoa adalah membuat spermatozoa dapat bergerak di saluran reproduksi betina dan untuk penetrasi zona pelusida. Komponen penyusun flagella adalah aksonema dan serabut pekat. Aksonema spermatozoa mamalia dikelilingi sembilan serabut pekat dari akhir bagian leher sampai ekor bagian utama. Fungsi aksonema belum diketahui dengan pasti karena serabut ini tidak mempunyai protein kontraktil namun mempunyai ATPase dan kalsium yang mungkin berperan dalam kontrol motilitas (Curry dan Watson 1995).
Bagian leher merupakan bagian yang paling pendek dan yang menghubungkan antara kepala dengan ekor. Ekor spermatozoa mengandung serabut-serabut fibril (axial filament) yang tersusun secara radial. Axial filament ini tersusun mulai dari sentriol atas dan berjalan sampai dengan ujung ekor. Susunannya dari luar ke tengah adalah 9 filamen besar, 9 pasang filamen kecil dan 2 filamen kecil di pusat. Serabut-serabut ini bertanggung jawab terhadap pergerakan spermatozoa (Garner dan Hafez 2000). Pada middle piece, bagian badan spermatozoa mamalia dari ujung bawah bagian leher sampai annulus (struktur pita yang melingkar antara bagian badan dan ekor utama). Karakteristik utama dari bagian ini adalah dengan adanya mitokondria yang tersusun secara heliks mengelilingi sumbu memanjang. Mitokondria merupakan tempat metabolisme yang menghasilkan energi untuk pergerakan spermatozoa. Membran mitokondria sangat stabil dan tahan terhadap tekanan selama pergerakan flagella. Membran dalam mitokondria adalah bagian yang memproduksi energi untuk spermatozoa dan posisi mitokondria mengelilingi flagella, yang siap untuk mensuplai energi (ATP) ke aksonema untuk motilitas (Curry dan Watson 1995). Pada principal piece, bagian terpanjang dari flagella, mulai dari annulus sampai ujung atas dari bagian ujung ekor. Bagian ini ditandai dengan keberadaan selubung serabut yang terdapat hanya 2 filamen pusat dikelilingi 9 pasang filamen kecil. Sementara itu, pada end piece yang merupakan bagian akhir dari ekor hanya mengandung 2 filamen pusat yang diselubungi membran (Curry dan Watson 1995; Bearden et al. 2004).
5
Gambar 1 Spermatozoa dengan bagian-bagiannya (Holstein et al. 2003)
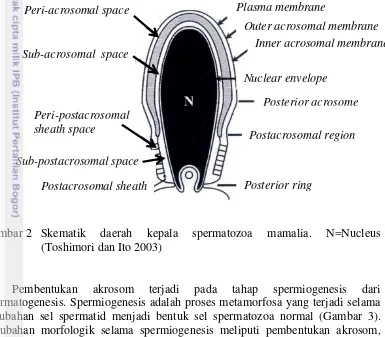
Akrosom
Struktur dasar kepala spermatozoa biasanya mirip antar spesies mamalia tetapi ukuran dan bentuk dari nukleus dan akrosom bervariasi serta bersifat spesifik untuk masing-masing spesies (Esteves dan Verza Jr 2011). Pada manusia, akrosom relatif kecil, menutupi dua pertiga kepala tetapi tidak memanjang di tepi anterior. Kepala spermatozoa terbagi menjadi dua daerah bagian yaitu daerah akrosom dan daerah post akrosom (Gambar 2). Akrosom berada pada setengah bagian dari kepala spermatozoa dan dilapisi oleh membran akrosom dalam dan membran akrosom luar. Membran akrosom bagian luar berada langsung dekat dengan membran plasma. Membran akrosom melingkupi bagian kecil yang berisi matriks akrosom. Matriks akrosom terdiri dari sejumlah enzim hidrolitik, utamanya yaitu hyaluronidase dan acrosin serta beberapa enzim lainnya seperti asam fosfatase, fosfolipase, N-asetilglukosaminidase dan kolagenase, termasuk didalamnya juga terdapat banyak karbohidrat. Enzim akan dilepaskan saat terjadi reaksi akrosom (Curry dan Watson 1995).
6
utuh ketika reaksi akrosom terjadi. Membran plasma yang menutupi bagian akrosom anterior berperan dalam dua proses utama yaitu sebagai sisi dari “ sperm-egg recognition” atau pengenalan spermatozoa dengan oosit dan penetrasi zona pelusida serta melakukan fusi dengan membran akrosom luar saat terjadi reaksi akrosom. Inisiasi ikatan zona pelusida diperantarai oleh keberadaan glikokonjugat yang terdapat pada membran anterior akrosom dan hal ini ditunjukkan dengan adanya ikatan dengan lektin. Sementara itu, pada daerah post akrosom terdapat selubung post akrosom (postacrosomal sheath) yang terletak diantara membran plasma dan selaput inti (Curry dan Watson 1995).
Gambar 2 Skematik daerah kepala spermatozoa mamalia. N=Nucleus (Toshimori dan Ito 2003)
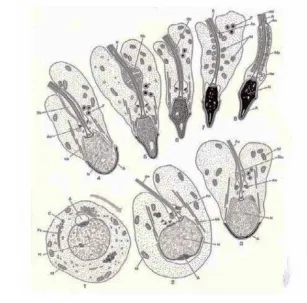
Pembentukan akrosom terjadi pada tahap spermiogenesis dari spermatogenesis. Spermiogenesis adalah proses metamorfosa yang terjadi selama perubahan sel spermatid menjadi bentuk sel spermatozoa normal (Gambar 3). Perubahan morfologik selama spermiogenesis meliputi pembentukan akrosom, kepala, leher dan ekor spermatozoa. Spermiogenesis dibagi dalam tahap Golgi, tahap tudung (capping), tahap akrosom dan tahap pematangan (maturation) (Hess dan Franca 2005).
Golgi
Tahap Golgi merupakan tahap pertama dari pembentukan akrosom. Pada tahap ini terbentuk granula proakrosomal di gelembung Golgi yang selanjutnya bergabung membentuk butir akrosom tunggal dalam gelembung akrosomal. Sentriol proksimal akan bergerak dari sitoplasma ke dasar nukleus yang nantinya akan menjadi leher antara kepala dan ekor, sedangkan sentriol distal akan berkembang menjadi aksonema (flagella di ekor).
7
Tudung
Butir akrosom pada tahap ini akan bergerak ke arah anterior inti. Butir-butir akrosom akan memipih yang disebut akrosom dan terdapat membran. Ekor akan terbentuk dari sentriol distal. Pada fase ini, spermatid akan bergerak ke arah lumen tubuli seminiferi.
Akrosom
Akrosom akan berkembang dan menutup hampir 2/3 area kepala. Kepala dan sitoplasma akan memanjang dan inti mengalami kondensasi.
Pematangan
Spermatid akan memanjang dan akan dilepaskan ke lumen sedangkan sisa sitoplasma akan bergerak ke arah posterior. Mitokondria akan mengelilingi flagella dari dasar inti sampai dengan 1/3 dari ekor.
Gambar 3 Tahap perubahan spermatid (Holstein et al. 2003)
Karbohidrat, Glikokonjugat dan Lektin
8
(terdapat pada membran akrosom luar) serta fertilisasi. Pada umumnya dapat dijumpai sekitar tujuh karbohidrat yang ada di mamalia, seperti mannosyl (Man), glucosyl (Glc), galactosyl (Gal), Fucosyl (Fuc), acetylgalactosyl (GalNAc), acetylglucosaminyl (GlcNAc) dan sialic acid (Desantis et al. 2006).
Glikokonjugat merupakan karbohidrat yang berikatan secara kovalen pada protein atau lipid. Glikoprotein adalah kompleks antara karbohidrat dengan protein, sementara glikolipid adalah kompleks karbohidrat dengan lipid. Glikoprotein adalah mukopolisakarida yang lebih banyak dan bervariasi dibandingkan polisakarida dan proteoglikan. Perubahan atau modifikasi glikokaliks spermatozoa seiring dengan penghilangan ataupun adanya protein spesifik atau glikoprotein yang terdapat pada saluran reproduksi betina dapat mengakibatkan terjadinya kapasitasi dan reaksi akrosom. Selanjutnya kapasitasi maupun reaksi akrosom yang terjadi pada beberapa spesies mamalia diketahui berperan dalam pengenalan gamet maupun fertilisasi (Purohit et al. 2008).
Glikokaliks terdapat di bagian antara permukaan sel spermatozoa dengan lingkungan ekstraseluler. Begitu pun pada oosit juga mencakup matriks glikokaliks, zona pelusida (ZP). Namun, tidak seperti zona pelusida pada oosit, glikokaliks yang ada di spermatozoa diperkirakan terdapat 50 sampai 150 glikokonjugat yang berbeda-beda. Selanjutnya, tidak seperti zona pelusida, beberapa penelitian menunjukkan bahwa permukaan spermatozoa tidaklah sama tetapi terdiri dari area yang memiliki fungsi yang berbeda, dan hal ini menggambarkan distribusi glikokonjugat pada area tertentu dari sel dapat memberi dugaan yang berkaitan dengan kemungkinan fungsinya di lokasi tersebut. Sejak adanya molekul yang terdapat pada permukaan sel spermatozoa ketika spermatozoa memperoleh kemampuan untuk membuahi, maka hal ini
dikenal dengan “major maturation antigen”. Komposisi dari glikokaliks utamanya berasal dari lektin spesifik, berikatan dengan rantai gula dengan tipe yang berbeda, kemudian bersatu menyerupai gugus gula membentuk permukaan spermatozoa (Schroter et al. 1999).
Lektin merupakan molekul protein yang dihasilkan dari tanaman serta hewan yang termasuk dalam golongan nonenzimatik tetapi tidak masuk dalam golongan immunoglobulin. Lektin dapat ditemukan pada jaringan tubuh hewan dan tumbuhan, terutama biji dari kacang-kacangan. Lektin termasuk protein besar (BM 20.000-300.000) sehingga dimungkinkan untuk mengikat molekul kovalen untuk beberapa grup asam amino bebas tanpa mengganggu ikatan karbohidrat tersebut. Karena afinitas lektin terhadap residu monosakarida sangat spesifik, maka lektin dapat digunakan untuk mendeteksi keberadaan maupun distribusi glikokonjugat (Kiernan 1990).
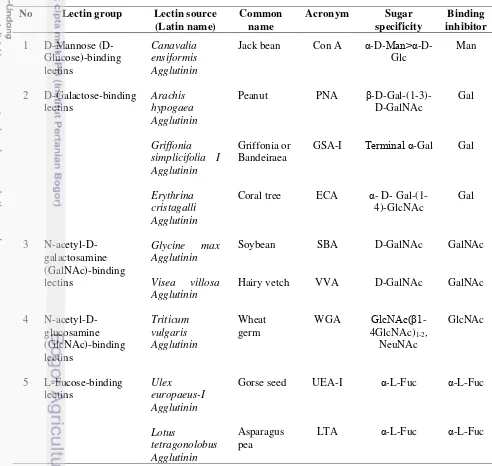
9 serta uronic acid. Glikokonjugat yang mengandung residu glukosa dan manosa berperan dalam transport ion, sedangkan yang mengandung residu gula fukosa terlibat dalam perlekatan antar sel dan pengaturan difusi substrat antar sel. Glikokonjugat dengan residu gula N-asetil-D-glukosamin berperan dalam transport cairan dan ion, sementara glikokonjugat dengan residu gula galaktosa terlibat dalam perlekatan antar sel dan penanda dalam diferensiasi sel (Spicer dan Schulte 1992). Jenis lektin, sumber dan afinitasnya terhadap karbohidrat dirangkum pada Tabel 1.
Tabel 1 Jenis lektin, sumber dan afinitasnya terhadap karbohidrat (Aly dan Salem-Bekhit 2012)
Man, mannose; Glc, glucose; GalNAc, acetylgalactosamine; GlcNAc. acetylglucosamine; NeuNAc, N-acetylneuraminic acid (sialic acid); α-L-Fuc, α-L-Fucose.
No Lectin group Lectin source
10
Kriopreservasi Spermatozoa
Fenomena utama selama proses kriopreservasi yang dapat menurunkan viabilitas sel spermatozoa yaitu kejutan dingin (cold shock) dan perubahan intraseluler akibat pengeluaran air yang berkaitan dengan pembentukan kristal es. Selain itu ada faktor tambahan yakni peroksidasi lipid. Kerusakan umum pada sel spermatozoa selama proses kriopreservasi akibat adanya fenomena tersebut adalah kerusakan mekanik yang ditandai dengan kerusakan organel sitoplasma atau pecah karena ekspansi es, konsentrasi larutan menjadi toksik dan tebal akibat adanya dehidrasi dari suspensi media baik intra maupun ekstraseluler dan perubahan fisik serta kimiawi diantaranya presipitasi, denaturasi, koagulasi dari protein, disosiasi ion dan kehilangan sifat-sifat absorpsi atau sifat-sifat pengikat air (Hu et al. 2006; Camara et al. 2011).
Kejutan dingin terjadi karena adanya penurunan suhu secara mendadak di bawah suhu 0°C. Hal ini berkaitan dengan fase transisi dari membran lipid yang menyebabkan terjadinya fase pemisahan dan penurunan sifat-sifat permeabilitas selektif dan membran biologik sel hidup. Efek kejutan dingin pada spermatozoa adalah penurunan aktivitas flagella, kerusakan organ intraseluler dan kerusakan membran sel. Pengaruh utama kejutan dingin terhadap sel spermatozoa adalah penurunan motilitas dan viabilitas, perubahan permeabilitas dan perubahan komponen lipid pada membran. Jumlah spermatozoa motil mengalami penurunan disertai pelepasan enzim, perpindahan ion melewati membran dan penurunan kandungan lipid seperti fosfolipid dan kolesterol yang sangat berperan dalam mempertahankan integritas membran plasma (Holt 2000).
Pembentukan kristal-kristal es selama proses kriopreservasi menyebabkan terjadi penumpukan elektrolit di dalam sel yang mengakibatkan terjadinya kerusakan sel secara mekanik, dimana elektrolit yang menumpuk akan merusak dinding sel sehingga pada waktu thawing, permeabilitas membran plasma akan berubah dan sel akan mati. Pembentukan kristal-kristal es kemungkinan berkaitan dengan perubahan tekanan osmotik dalam fraksi yang tidak mengalami pembekuan (Watson 2000).
11 rantai oksidasi, sementara apabila lisosom pecah akan mengeluarkan asam hidrolase sehingga akan mencerna bagian sel yang lain.
Radikal bebas yang merupakan senyawa oksigen reaktif atau Reactive Oxygen Species (ROS) adalah molekul atau oksidan yang sangat reaktif walaupun derajat kekuatannya berbeda-beda karena memiliki satu atau lebih elektron yang tidak berpasangan dalam orbital luarnya, sehingga dapat bereaksi dengan molekul sel dengan cara mengikat elektron dari molekul sel tersebut yang mengakibatkan reaksi berantai yang dapat menghasilkan radikal bebas baru, seperti superoxide dismutase (SOD) dan nitric oxide (NO). Dengan demikian, radikal bebas dapat mengganggu integritas sel dan dapat bereaksi dengan komponen-komponen sel, baik komponen struktural seperti molekul-molekul penyusun membran maupun komponen fungsional, seperti enzim-enzim dan DNA. Tingginya komposisi spermatozoa dengan asam lemak tidak jenuh memiliki konsekuensi yang tidak menguntungkan karena menjadi rentan terhadap peroksidasi lipid (Meggiolaro et al. 2003). Peroksidasi lipid menyebabkan kerusakan matrik struktur lipid yang menyebabkan instabilitas membran karena terputusnya rantai asam lemak menjadi senyawa yang bersifat toksik terhadap sel, seperti malonaldehide (MDA). Pada sel spermatozoa efek toksik yang disebabkan oleh peroksidasi lipid mengakibatkan rendahnya motilitas spermatozoa, kerusakan enzim intraseluler serta kerusakan membran plasma spermatozoa terutama pada bagian akrosom sehingga juga menurunkan fertilitasnya (Sarlos et al. 2002).
3
METODE
Waktu dan Tempat Penelitian
12
Koleksi Semen
Semen segar dikoleksi dari domba lokal jantan yang sudah mencapai sexual maturity berumur antara 1-2 tahun dengan bobot badan berkisar antara 25-30 kg. Domba dipelihara secara intensif dalam kandang individual yang berada di Unit Rehabilitasi Reproduksi, Fakultas Kedokteran Hewan, Institut Pertanian Bogor. Pakan hijauan dan konsentrat sebagai sumber makanan pokok bagi domba, sedangkan air minum diberikan secara ad libitum. Penampungan semen dilakukan sebanyak dua kali seminggu dengan menggunakan vagina buatan.
Kriopreservasi Spermatozoa
Semen segar yang sudah dievaluasi karakteristiknya diencerkan dengan medium Niwa dan Sasaki Freezing (NSF) untuk kemudian dibekukan (Kikuchi et al. 1999; Karja et al. 2006). Hanya semen yang mempunyai karakteristik baik dengan persentase motilitas minimal 70% digunakan proses pembekuan. Dalam proses pembekuan, penambahan bahan pengencer dilakukan dengan metode two step freezing yaitu dimulai dengan penambahan medium NSF I kemudian dilakukan ekuilibrasi pada suhu 4°C selama 2 jam sebelum ditambahkan dengan medium NSF II dan diekuilibrasi kembali pada suhu yang sama selama 5 menit. Komposisi medium freezing I (NSF I) terdiri dari 20% (v:v) kuning telur, 8,8% (w/v) laktosa (Merck, Germany) dan 20 mg/ml ampicillin, sedangkan medium freezing II (NSF II) terdiri dari 92,52% (v:v) medium freezing I, 1,48% (v:v) orvus ES paste dan 6% (v:v) gliserol (Lampiran 1). Perbandingan medium NSF I dan NSF II adalah sebesar 1:1. Semendikemas ke dalam straw berukuran 0,25 ml (I.V.M., France) kemudian diletakkan pada Styrofoam plate dalam uap nitrogen cair berjarak sekitar 4 cm dari permukaan nitrogen cair selama 20 menit kemudian segera dimasukkan dalam kontainer nitrogen cair untuk penyimpanan.
Evaluasi Karakteristik dan Deteksi Status Akrosom
Evaluasi semen selama proses pembekuan (semen segar, setelah ekuilibrasi dan setelah thawing) dilakukan terhadap karakteristik spermatozoa yang meliputi persentase motilitas, viabilitas dan membran plasma utuh (MPU) spermatozoa serta deteksi status akrosom dengan pewarnaan histokimia lektin. Thawing semen beku dilakukan pada water bath dengan temperatur 32°C selama 30 detik.
1) Penilaian persentase motilitas
13 2) Persentase spermatozoa hidup
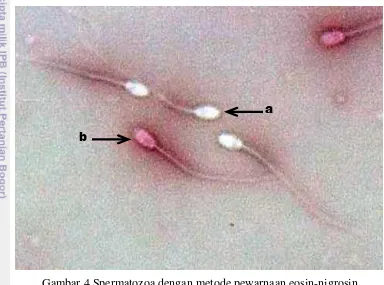
Penentuan persentase viabilitas dari spermatozoa dilakukan dengan menggunakan metode pewarnaan eosin-nigrosin (Cocchia et al. 2011). Sebanyak 10 µl sampel semen dan ditambah 40 µl eosin-nigrosin kemudian dicampur diatas gelas objek dan dibuat preparat ulas. Setelah itu, dikeringkan menggunakan heating table selama 15 detik sebelum dilakukan pengamatan dan sebanyak 100 spermatozoa diamati dengan mikroskop cahaya pembesaran 400x. Spermatozoa yang dikategorikan hidup adalah spermatozoa yang tidak menyerap zat warna sehingga pada bagian kepala spermatozoa tidak terwarnai (putih), sedangkan spermatozoa yang dikategorikan mati adalah spermatozoa yang menyerap zat warna sehingga pada bagian kepalanya akan berwarna merah (Gambar 4).
Gambar 4 Spermatozoa dengan metode pewarnaan eosin-nigrosin. Spermatozoa hidup (a) dan spermatozoa mati (b).
3) Penilaian keutuhan membran plasma
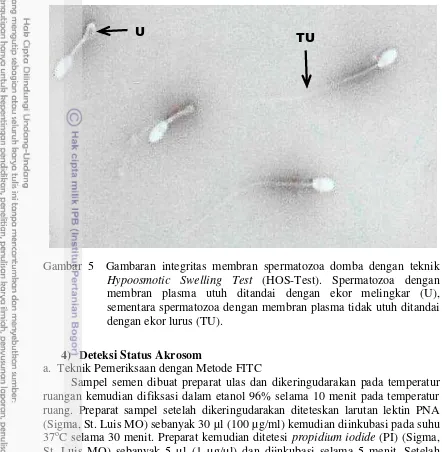
Penilaian persentase keutuhan membran plasma spermatozoa diperiksa menggunakan teknik Hypoosmotic Swelling Test (HOS-Test) dengan komposisi 0,135 g fruktosa (Merck, Germany) dan 0,0737 g trisodium citrate 2H2O dalam 10 ml air mili-Q. Sampel semen sebanyak 20 μl diencerkan dengan 80 μl larutan HOS dan dibiarkan selama 10 menit dalam water bath (37oC). Untuk keperluan
pengamatan, diteteskan 10 μl sampel semen pada gelas objek yang ditutup dengan gelas penutup dan evaluasi dilakukan dengan mikroskop cahaya pembesaran 400x terhadap 100 spermatozoa. Spermatozoa yang memiliki membran plasma utuh ditandai oleh ekor yang melingkar atau menggelembung, sedangkan spermatozoa yang memiliki membran plasma tidak utuh ditandai oleh ekor yang lurus (Gambar 5) (Perez-Llano et al. 2006).
14
Gambar 5 Gambaran integritas membran spermatozoa domba dengan teknik Hypoosmotic Swelling Test (HOS-Test). Spermatozoa dengan membran plasma utuh ditandai dengan ekor melingkar (U), sementara spermatozoa dengan membran plasma tidak utuh ditandai dengan ekor lurus (TU).
4) Deteksi Status Akrosom
a. Teknik Pemeriksaan dengan Metode FITC
Sampel semen dibuat preparat ulas dan dikeringudarakan pada temperatur ruangan kemudian difiksasi dalam etanol 96% selama 10 menit pada temperatur ruang. Preparat sampel setelah dikeringudarakan diteteskan larutan lektin PNA (Sigma, St. Luis MO) sebanyak 30 µl (100 µg/ml) kemudian diinkubasi pada suhu 37oC selama 30 menit. Preparat kemudian ditetesi propidium iodide (PI) (Sigma, St. Luis MO) sebanyak 5 µl (1 µg/µl) dan diinkubasi selama 5 menit. Setelah inkubasi, preparat kemudian dicuci dengan PBS (phosphate-buffered saline) sebanyak tiga kali untuk membersihkan sisa pereaksi yang tidak berikatan, kemudian ditutup menggunakan cover glass. Pemeriksaan status akrosom dilakukan menggunakan mikroskop fluoresens (Nikkon, Eclipse E600, Japan) pada panjang gelombang 380-420 nm. Jumlah spermatozoa yang diamati pada setiap perlakuan adalah 200 spermatozoa. Hasil pemeriksaan dengan metode FITC dibedakan ke dalam dua kategori, yaitu spermatozoa dengan akrosom berwarna hijau dikategorikan sebagai akrosom intak, sementara spermatozoa tidak berwarna hijau dikategorikan sebagai akrosom rusak sesuai metode Cocchia et al. (2011). Semua proses pewarnaan dilakukan dalam ruang gelap. Prosedur pewarnaan histokimia lektin metode FITC secara garis besar tercantum pada Lampiran 3.
U
15 b. Teknik Pemeriksaan dengan Metode ABC
Teknik ABC yang dipakai dalam penelitian ini sesuai dengan metode Hsu et al. (1981) dengan beberapa modifikasi. Sampel semen dibuat preparat ulas kemudian dikeringudarakan untuk selanjutnya difiksasi dalam larutan glutaraldehyde 4% dan didiamkan pada suhu 4oC selama 3 hari. Preparat yang sudah difiksasi kemudian dicuci dengan PBS selama 15 menit serta direndam dengan larutan 0,3% H2O2 dalam metanol selama 30 menit untuk menghilangkan peroksidase endogen dan kemudian dicuci kembali dengan PBS.
Preparat diinkubasi dengan lektin PNA (biotinylated PNA) (Vector Lab, Inc., USA) selama satu malam. Setelah itu, preparat dicuci dengan PBS. Selanjutnya, pada masing-masing preparat diteteskan ABC kit (Avidin-Biotin Peroxidase Complex) (Vector Lab, Inc., USA) dan diinkubasi pada suhu 37oC selama 30 menit untuk kemudian dicuci dengan PBS. Untuk visualisasi, preparat ditetesi dengan larutan diamino benzidine (DAB) (Vector Lab, Inc., USA) dan didiamkan dalam waktu 15 menit. Terakhir dilakukan pencucian preparat dengan akuades, kemudian preparat didehidrasi, dilakukan clearing dan mounting. Reaksi yang terjadi ditunjukkan dengan warna coklat pada bagian akrosom yang mengindikasikan akrosom intak, sementara itu spermatozoa tidak berwarna dikategorikan sebagai akrosom rusak. Jumlah spermatozoa yang diamati pada setiap perlakuan adalah 200 spermatozoa. Prosedur pewarnaan histokimia lektin metode ABC secara garis besar tercantum pada Lampiran 2.
Analisis Data
Data karakteristik spermatozoa serta evaluasi pewarnaan histokimia lektin disajikan dalam bentuk persentase dan dianalisis dengan analisis sidik ragam (ANOVA) dalam bentuk rancangan acak lengkap dengan lima kali ulangan. Apabila terdapat perbedaan dilanjutkan dengan uji beda nyata terkecil (Steel dan Torrie 1993).
4
HASIL DAN PEMBAHASAN
Karakteristik Spermatozoa Domba Selama Proses Pembekuan
16
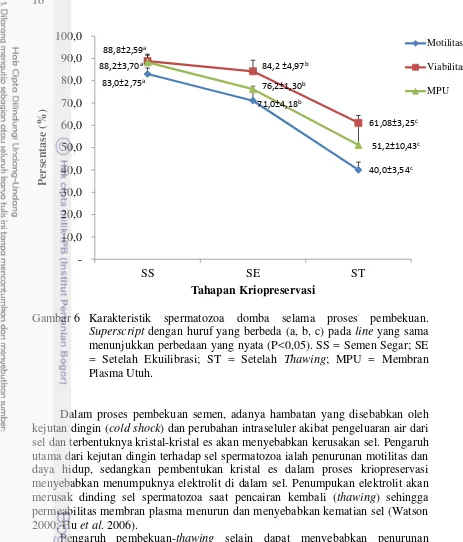
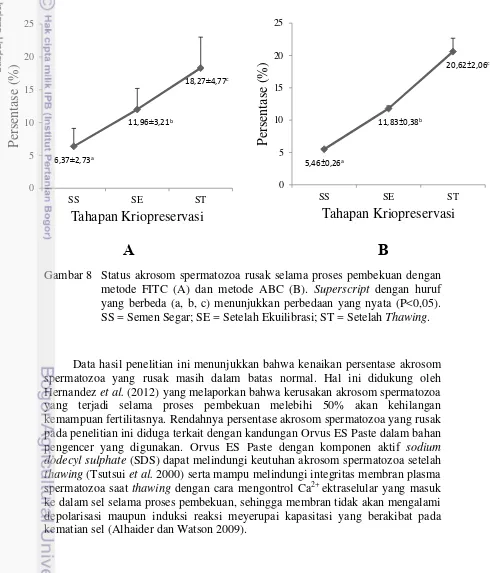
Gambar 6 Karakteristik spermatozoa domba selama proses pembekuan. Superscript dengan huruf yang berbeda (a, b, c) pada line yang sama menunjukkan perbedaan yang nyata (P<0,05). SS = Semen Segar; SE = Setelah Ekuilibrasi; ST = Setelah Thawing; MPU = Membran Plasma Utuh.
Dalam proses pembekuan semen, adanya hambatan yang disebabkan oleh kejutan dingin (cold shock) dan perubahan intraseluler akibat pengeluaran air dari sel dan terbentuknya kristal-kristal es akan menyebabkan kerusakan sel. Pengaruh utama dari kejutan dingin terhadap sel spermatozoa ialah penurunan motilitas dan daya hidup, sedangkan pembentukan kristal es dalam proses kriopreservasi menyebabkan menumpuknya elektrolit di dalam sel. Penumpukan elektrolit akan merusak dinding sel spermatozoa saat pencairan kembali (thawing) sehingga permeabilitas membran plasma menurun dan menyebabkan kematian sel (Watson 2000; Hu et al. 2006).
Pengaruh pembekuan-thawing selain dapat menyebabkan penurunan motilitas juga dilaporkan dapat menyebabkan kerusakan pada organel-organel sel seperti mitokondria spermatozoa (Kasai et al. 2002; Partyka et al. 2010). Energi berupa Adenosine Tri Phosphate (ATP) yang merupakan hasil metabolisme di dalam membran mitokondria diduga berperan untuk menggertak mikrotubul yang menyebabkan terjadinya pergesekan diantara mikrotubul, sehingga spermatozoa mampu bergerak secara bebas (Silva dan Gadella 2006). Lebih lanjut, Gadea et al. (2007) menambahkan bahwa motilitas spermatozoa sangat bergantung pada suplai energi berupa Adenosine Tri Phosphate (ATP) hasil metabolisme yang akan berlangsung dengan baik apabila membran plasma sel ada dalam keadaan yang utuh, sehingga mampu dengan baik mengatur lalu lintas keluar masuk sel substrat
17 dan elektrolit-elektrolit yang dibutuhkan dalam proses metabolisme. Pada membran plasma sel terdapat banyak makromolekul seperti protein, lipoprotein, glikoprotein, dan lain-lain yang dapat berfungsi sebagai enzim, reseptor, saluran atau pembawa (carrier). Makromolekul-makromolekul inilah yang berfungsi memfasilitasi lalu lintas keluar masuk seluruh substrat dan elektrolit pada sel. Substrat dan elektrolit harus difasilitasi karena tidak dapat menembus secara difusi bebas membran plasma sel spermatozoa yang bersifat semipermeabel.
Penurunan viabilitas spermatozoa juga terlihat setelah spermatozoa mengalami proses pembekuan yaitu sebesar 27,72%, meskipun demikian persentase viabilitas spermatozoa setelah thawing pada penelitian ini masih memenuhi standar (61,08%), karena menurut Hernandez et al. (2012) selama proses pembekuan-thawing hampir 50% spermatozoa mati. Tingginya persentase viabilitas spermatozoa pada penelitian ini diduga terkait dengan penggunaan medium NSF I dan NSF II yang mengandung Orvus ES Paste. Komponen aktif pada Orvus ES Paste yaitu sodium dodecyl sulphate (SDS) dalam bahan pengencer dapat meningkatkan viabilitas dan fluiditas membran plasma spermatozoa, sehingga dapat mempertahankan viabilitas spermatozoa setelah thawing seperti yang dilaporkan Alhaider dan Watson (2009). Lebih lanjut dilaporkan bahwa Orvus ES Paste mampu meningkatkan daya hidup spermatozoa setelah pencairan kembali (thawing) dengan bertindak sebagai surfaktan untuk menstabilkan membran sel dan untuk melindungi spermatozoa dari efek toksik gliserol selama proses pembekuan (Ponglowhapan dan Chatdarong 2008).
Integritas membran plasma harus tetap terjaga agar kelangsungan hidup spermatozoa, motilitas dan kemampuan fertilisasi dapat dipertahankan. Hal tersebut dikarenakan selain berfungsi sebagai pelindung secara fisik organel-organel sel spermatozoa termasuk akrosom, membran plasma juga berfungsi dalam pengaturan keluar-masuknya zat-zat nutrisi dan ion-ion. Selain itu, membran plasma juga berfungsi sebagai penjaga keseimbangan elektrolit intra dan ekstraseluler. Rusaknya membran plasma akan mengakibatkan terganggunya proses metabolisme dan proses fisiologis yang akan mengakibatkan kematian spermatozoa (Zhu dan Liu 2000). Hasil penelitian menunjukkan bahwa terjadi penurunan persentase membran plasma utuh spermatozoa setelah proses pembekuan sebesar 37%, sehingga hanya tersisa 51,2% masih memiliki membran plasma utuh.
18
Deteksi Kerusakan Akrosom Spermatozoa Domba
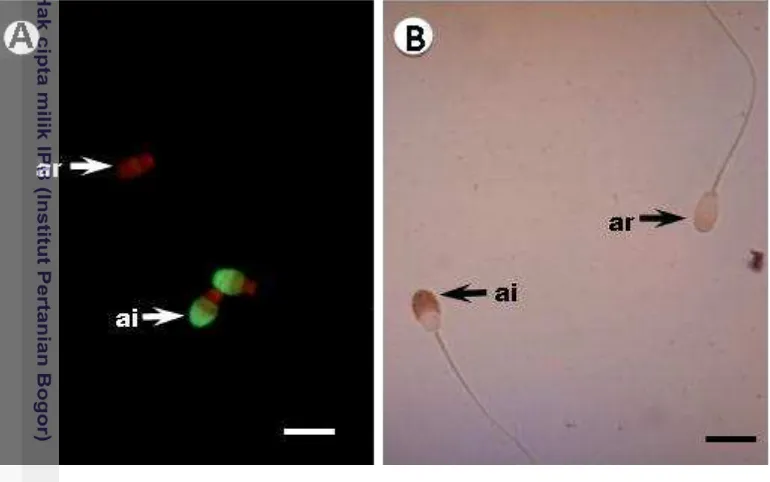
Keutuhan akrosom merupakan kunci keberhasilan terjadinya proses fertilisasi, karena hanya spermatozoa dengan akrosom utuh yang mampu melakukan penetrasi zona pelusida dan melakukan fusi dengan membran plasma oosit (Celeghini et al. 2010). Status akrosom spermatozoa domba selama proses pembekuan pada penelitian ini dideteksi dengan menggunakan dua teknik pewarnaan histokimia lektin yaitu FITC dan ABC. Hasil yang diperoleh seperti terlihat pada Gambar 7.
Gambar 7 Status akrosom spermatozoa domba dengan pewarnaan histokimia lektin metode FITC (A): Spermatozoa dengan akrosom berwarna hijau dikategorikan sebagai akrosom intak (ai), sedangkan spermatozoa tidak berwarna hijau dikategorikan sebagai akrosom rusak (ar). Status akrosom spermatozoa domba dengan pewarnaan histokimia lektin metode ABC (B): Spermatozoa dengan akrosom berwarna coklat dikategorikan sebagai akrosom intak (ai), sedangkan spermatozoa yang tidak berwarna dikategorikan sebagai akrosom rusak (ar). Skala = 10 µm.
19 secara efektif untuk mengevaluasi status akrosom spermatozoa sehingga dapat memberikan gambaran fertilitas dari spermatozoa.
Deteksi status akrosom spermatozoa dengan dua metode yang digunakan pada penelitian ini (metode FITC dan ABC) menunjukkan hasil yang sama yaitu terjadi peningkatan persentase akrosom spermatozoa rusak selama proses pembekuan (P<0,05) (Gambar 8). Kenaikan akrosom spermatozoa rusak setelah thawing terdeteksi dengan menggunakan metode FITC sebesar 11,9%, sementara dengan metode ABC sebesar 15,16%.
Gambar 8 Status akrosom spermatozoa rusak selama proses pembekuan dengan metode FITC (A) dan metode ABC (B). Superscript dengan huruf yang berbeda (a, b, c) menunjukkan perbedaan yang nyata (P<0,05). SS = Semen Segar; SE = Setelah Ekuilibrasi; ST = Setelah Thawing.
Data hasil penelitian ini menunjukkan bahwa kenaikan persentase akrosom spermatozoa yang rusak masih dalam batas normal. Hal ini didukung oleh Hernandez et al. (2012) yang melaporkan bahwa kerusakan akrosom spermatozoa yang terjadi selama proses pembekuan melebihi 50% akan kehilangan kemampuan fertilitasnya. Rendahnya persentase akrosom spermatozoa yang rusak pada penelitian ini diduga terkait dengan kandungan Orvus ES Paste dalam bahan pengencer yang digunakan. Orvus ES Paste dengan komponen aktif sodium dodecyl sulphate (SDS) dapat melindungi keutuhan akrosom spermatozoa setelah thawing (Tsutsui et al. 2000) serta mampu melindungi integritas membran plasma spermatozoa saat thawing dengan cara mengontrol Ca2+ ektraselular yang masuk ke dalam sel selama proses pembekuan, sehingga membran tidak akan mengalami depolarisasi maupun induksi reaksi meyerupai kapasitasi yang berakibat pada kematian sel (Alhaider dan Watson 2009).
20
Menurut Mizutani et al. (2010) menyatakan bahwa sodium dodecyl sulphate (SDS) merupakan cairan deterjen anionik terlarut yang berperan mempertahankan dan melarutkan membran protein pada konsentrasi tinggi, sehingga dapat melindungi spermatozoa melawan cold shock serta meningkatkan kemampuan bertahan spermatozoa selama proses pembekuan. Mekanisme kerja dari Orvus ES Paste dengan komponen aktif SDS dalam melindungi tudung akrosom spermatozoa belum diketahui secara spesifik, namun demikian beberapa penelitian telah membuktikan bahwa penambahan Orvus ES Paste ke dalam bahan pengencer mampu melindungi akrosom spermatozoa setelah thawing. Hasil penelitian Mizutani et al. (2010) menunjukkan bahwa persentase status akrosom spermatozoa kucing dengan akrosom spermatozoa intak setelah thawing yang diberi penambahan Orvus ES Paste lebih tinggi (20,6±1,2%) dibandingkan dengan tanpa penambahan Orvus ES Paste (12,2±1,7%). Hasil serupa juga dilaporkan Tsutsui et al. (2000) yang menyatakan bahwa penambahan Orvus ES Paste dalam metode pembekuan pada babi dan anjing menunjukkan bahwa Orvus ES Paste mampu melindungi tudung akrosom spermatozoa.
Menurut Morton (2010), SDS tidak bekerja secara langsung pada membran plasma spermatozoa, melainkan akan memodifikasi partikel kuning telur dengan melarutkan komponen lipid dari kuning telur yang dapat melindungi membran plasma. Tsutsui et al. (2000) juga menambahkan bahwa efek proteksi SDS sebagai cairan deterjen anionik terlarut adalah untuk melarutkan dan menambah dispersi dari lipoprotein kuning telur serta memperkuat ikatan antara kuning telur dan membran plasma spermatozoa, sehingga kuning telur yang awalnya tidak larut secara penuh dapat terdispersi secara lebih baik dan mampu melindungi membran plasma spermatozoa, termasuk melindungi keutuhan akrosom spermatozoa.
5
SIMPULAN
Status akrosom spermatozoa dapat dideteksi menggunakan lektin PNA metode FITC dan metode ABC dengan efektifitas yang sama sehingga dapat digunakan untuk penilaian terhadap kualitas dan fertilitas spermatozoa.
DAFTAR PUSTAKA
Agungpriyono S, Kurohmaru M, Kimura J, Wahid AH, Sasaki M, Kitamura N, Yamada J, Fukuta K, Zuki AB. 2009. Distribution of lectin-bindings in the testis of the Lesser Mouse Deer, Tragulus javanicus. Anat Histol Embryol 38: 208-213.
21 Aly K, Salem-Bekhit MM. 2012. Histochemical mapping of glycoconjugates in the eyeball of the one Humped Camel (Camellus dromedarius). J Pharm Biomed Sci 2(4): 33-46.
Bag S, Joshi A, Naqvi SMK, Rawat PS, Mittal JP. 2002. Effect of freezing temperature, at which straws were plunged into liquid nitrogen, on the post-thaw motility and acrosomal status of ram spermatozoa. Anim Reprod Sci 72: 175-183.
Baker SS, Thomas M, Thaler CD. 2004. Sperm membrane dynamics assessed by changes in lectin fluorescence before and after capacitation. J Androl 25(5): 744-751.
Batista AM, Silva SV, Soares AT, Monteiro Jr PLJ, Wischral A, Guerra MMP. 2011. Comparison of capripure and percoll density gradients for sperm separation of frozen thawed goat spermatozoa. Anim Reprod 8: 81-84. Bearden HJ, Fuquay JW, Willard ST. 2004. Applied Animal Reproduction. 6th Ed.
New Jersey: Pearson education Inc.
Blesbois E, Grasseau I, Seigneurin F. 2005. Membrane fluidity and the ability of domestic bird spermatozoa to survive cryopreservation. Reproduction 129: 371-378.
Camara DR, Silva SV, Almeida FC. Nunes JF, Guerra MMP. 2011. Effects of antioxidants and duration of pre-freezing equilibration on frozen-thawed ram semen. Theriogenology 76: 342-350.
Celeghini ECC, de Andrade AFC, Raphael CF, Nascimento J, Ticianelli JS, de Arruda RP. 2010. Damage assessment of the equine sperm membranes by fluorimetric technique. Braz Arch Biol Technol 53(6): 1285-1292.
Cocchia N, Pasolini MP, Mancini R, Petrazzuolo O, Cristofaro I, Rosapane I, Sica A, Tortora G, Lorizio R, Paraggio G, Mancini A. 2011. Effect of SOD (superoxide dismutase) protein supplementation in semen extenders on motility, viability, acrosome status and ERK (extracelluler signal-regulated kinase) protein phosphorylation of chilled stallion spermatozoa. Theriogenology 75: 1201-1210.
Curry MR dan Watson PF. 1995. Sperm structure and function. Di dalam: Grudzinkas JG dan Yovich JL, Editor. Gametes-The Spermatozoon. Cambridge: Cambridge University Press.
Desantis S, Ventriglia G, Zubani D, Deflorio M, Megalofonou P, Acone F, Zarrili A, Palmieri G, De Metrio G. 2006. Histochemical analysis of glycoconjugates in the domestic cat testis. Histol Histopathol 21: 11-22. Esteves SC, Sharma RK, Thomas AJ Jr, Agarwal A. 2000. Improvement in
22
Esteves SC, Sharma RK, Thomas Jr AJ, Agarwal A. 2007. Evaluation of acrosomal status and sperm viability in fresh and cryopreserved specimens by the use of fluorescent peanut agglutinin lectin in conjuction with hypo-osmotic sweeling test. Int Braz J Urol 33(3): 364-376.
Esteves SC, Verza Jr S. 2011. Relationship of in vitro acrosome reaction to sperm function: An Update. The open Reproductive Science Journal 3: 72-84. Florman HM, Jungnickel MK, Sutton KA. 2008. Regulating the acrosome
reaction. Int J Dev Biol 52(5-6): 503-510.
Gadea J, Gumbao D, Novas SC, Zquez FAZ, Grullo LA. 2007. Supplementation of the dilution medium after thawing with reduced glutathione improves function and the in vitro fertilizing ability of frozen-thawed bull spermatozoa. Andrology 7: 1-10.
Gadella BM. 2008. Sperm membrane physiology and relevance for fertilization. Anim Reprod Sci 107: 229-236.
Gazali M, Tambing SN. 2002. Kriopreservasi sel spermatozoa. Hayati 9(1): 27-32. Garner DL, Hafez ESE. 2000. Spermatozoa and seminal plasma. In: Hafez ESE,
Hafez B, 7th Ed. Reproduction in farm animals. USA: Lippincott Williams & Wilkins.
Hernandez PJE, Fernandez RF, Rodriguez SJL, Soto MYG, Verona JEH, Garcia RAD. 2012. Post-thaw acrosomal viability and reaction in sperm obtained from equine epididymis tail. Rev Salud Anim 34(2): 84-88.
Hess R, Franca LR. 2005. History of the sertoli cell discovery. In: Griswold M, Skinner M, Ed. Sertoli cell biology. New York: Academic Press.
Holstein AF, Schulze W, Davidoff M. 2003. Understanding spermatogenesis is a prerequisite for treatment. Reprod Biol Endocrinol 1(107): 1-16.
Hsu S, Raine L, Fanger, H. 1981. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: a comparison between ABC and unlabeled antibody (PAP) procedures. J Histochem Cytochem 29(4): 577-580.
Hu Jian-hong, Li Qing-Wang, Gang-Li, Chen Xiao-Yu, Hai-Yang, Zhang Shu-Shan, Wang Li-Qiang. 2006. The cryoprotective effect on frozen-thawed boar semen of egg yolk low density lipoproteins. J Anim Sci 19(4): 486-494. Holt WV. 2000. Basic aspects of frozen storage of semen. Anim Reprod Sci 62:
3-22.
23 Kasai T, Ogawa K, Mizuno K, Nagai S, Uchida Y, Ohta S, Fujie M, Suzuki K, Hirata S, Hoshi K. 2002. Relationship between sperm mitochondrial membrane potential, sperm motility and fertility potential. Asian J Androl 4: 97-103.
Kiernan JA. 1990. Histological and Histochemical Methods: Theory & Practice 2nd Ed. New York: Pergamon Press. methodology for the detection and quantification of the acrosome reaction in mouse spermatozoa. Histol Histopathol 24: 999-1007.
Maji S, Datta U, Hembram ML. 2010. Cell surface changes associated with in vitro capacitation and acrosome reaction of goat epididymal sperm by a marine bio-active compound from the snail Telescopium telescopium. Vet Arhiv 80(5): 561-570.
Meggiolaro D, Porcelli F, Consiglio AL, Carnevali A, Crepaldi P, Molteni L, Ferrandi B. 2003. Effect of lipid peroxidation on the immunocythochemical detection of a leukocyte antigenic determinant in fresh and cryopreserved bovine spermatozoa. J Anim Sci 2: 255-263.
Miranda PV, Allaire A, Sosnik J, Visconti PE. 2009. Localization of low-density detergent-resistant membrane proteins in intact and acrosome-reacted mouse sperm. Biol Reprod 80: 897-904.
Mizutani T, Sumigama S, Nagakubo K, Shimizu N, Oba H, Hori, Tsutsui T. 2010. Usefulness of addition of orvus ES paste and sodium lauryl sulfate to frozen feline semen. Theriogenology 72(1): 23-27.
Morton KM, Evans G, Maxwell WMC. 2010. Effect of glycerol concentration, Equex-STM supplementation and liquid storage prior to freezing on the motility and acrosome integrity of frozen-thawed epididymal alpaca (Vicugna pacos) sperm. Theriogenology 74: 311-316.
Nur Z, Zik B, Ustuner B, Tutuncu S, Sagirkaya H, Ozguden CG, Gunay U, Dogan I. 2011. Effect of freezing rate on acrosome and chromatin integrity in ram semen. Ankara Univ Vet Fak Derg 58: 267-272.
Odhiambo JF, Sutovsky M, DeJarnette JM, Marshall C, Sutovsky P. 2011. Adaptation of ubiquitin-PNA based sperm quality assay for semen evaluation by a conventional flow cytometer and a dedicated platform for flow cytometric semen analysis. Theriogenology 76: 1168-1176.
Partyka A, Ni-zanski W, Lukaszewicz E. 2010. Evaluation of fresh and frozen-thawed fowl semen by flow cytometry. Theriogenology 74: 1017-1027. Partyka A, Lukaszewicz E, Ni-zanski W, Twardon J. 2011. Detection of lipid
24
Perez-Llano B, Enciso M, Garcia-Casado P, Sala R, Gosalvez J. 2006. Sperm DNA fragmentation in boars is delayed or abolished by using sperm extenders. Theriogenology 66: 2137-2143.
Ponglowhapan S, Chatdarong K. 2008. Effects of Equex STM Paste on the quality of frozen-thawed epididymal dog spermatozoa. Theriogenology 69: 666-672.
Purohit S, Laloraya M, Kumar PG. 2008. Distribution of N- and O-linked oligosaccharides on surface of spermatozoa from normal and infertile subjects. Andrologia 40(1): 7-12.
Sarlos P, Molnar A, Kokai M, Gabor Gy, Ratky J. 2002. Comparative evaluation of the effect of antioxidants in the conservation of ram semen. Acta Vet Hung 50(2): 235-245.
Schroter S, Osterhoff C, McArdle W, Ivell R. 1999. The glycocalyx of the sperm surface. Hum Reprod 5(4): 302-313.
Siciliano L, Maeciano V, Carpino A. 2008. Proteosome-like vesicles stimulate acrosome reaction of pig spermatozoa. Reprod Biol Endocrinol 6(5): 1-7. Silva PFN, Gadella BM. 2006. Detection of damage in mammalian sperm cells.
Theriogenology 65: 958-978.
Spicer SS, Schulte BA. 1992. Diversity of cell glycoconjugates shown histochemically: a perspective. J Hictochem Cytochem 40(1): 1-38.
Steel RGD, Torrie JH. 1993. Prinsip dan Prosedur Statistika. Jakarta: Gramedia Pustaka Utama.
Talaei T, Esmaeelpour T, Aekiyash F, Bahmanpour S. 2010. Effects of cryopreservation on plasma membrane glycoconjugates of human spermatozoa. Iran J Reprod Med 8(3): 119-124.
Toshimori K, Ito C. 2003. Formation and organization of the mammalian sperm head. Arch Histol Cytol 66(5): 383-396.
Tsutsui T, Hase M, Hori T, Ito T, Kawakami E. 2000. Effects of orvus ES paste on canine spermatozoal longevity after freezing and thawing. J Vet Med Sci 62(5): 533–535.
Watson PF. 2000. The causes of reduced fertility with cryopreserved semen. Anim Reprod Sci 60-61: 481-492.
25
26
Lampiran 1 Komposisi medium Niwa dan Sasaki Freezing (NSF) untuk kriopreservasi spermatozoa
1. Medium Freezing NSF I
Komponen Jumlah
Laktosa 8,8%
Kuning Telur 20%
Distilled Water 80%
Ampicilin 20 mg/ml
2. Medium Freezing NSF II
Komponen Jumlah
Medium Freezing NSF I 92,52%
Orvus ES Paste 1,48%
27 Lampiran 2 Prosedur pewarnaan histokimia lektin metode ABC (Avidin Biotin
Complex)
Pewarnaan Histokimia Lektin untuk Akrosom Spermatozoa
Metode ABC (
Avidin Biotin Complex
)
1. Sediaan spermatozoa dibuat preparat ulas
2. Fiksasi glutaraldehyde 4% 3 hari (refrigerator 4oC)
3. PBS 0,01 M (3x) @ 5 menit
4. Blocking „endogenous peroxidase‟
Larutan 0,3% H2O2 dalam metanol 30 menit
5. PBS 0,01 M (3x) @ 5 menit
6. Lektin PNA (biotinylated PNA) @ 30 µl 1 malam (refrigerator 4oC)
7. PBS 0,01 M (3x) @ 5 menit
8. ABC Kit @ 60 µl 30 menit (inkubator 37oC)
9. PBS 0,01 M (3x) @ 5 menit
10. DAB (diaminobenzidine) 1-15 menit*
11. Cuci dengan akuades
12. Dehidrasi, clearing, mounting
Keterangan:
28
Lampiran 3 Prosedur pewarnaan histokimia lektin metode FITC (Fluorescens isothiocyanate)
Pewarnaan Histokimia Lektin untuk Akrosom Spermatozoa
Metode FITC (
Fluorescens isothiocyanate
)
1. Sediaan spermatozoa dibuat preparat ulas
2. Fiksasi etanol 96% 10 menit
3. Lektin PNA @ 30 µl 30 menit (inkubator 37oC) 4. PI (Propidium Iodide) @ 5 µl 5 menit
5. PBS 0,01 M (3x) @ 5 menit
29
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 26 Agustus 1989 sebagai anak kedua dari pasangan bapak Khairuman Esa Wijaya (Alm) dan ibu Lili Muslikhah. Pendidikan sarjana ditempuh di Fakultas Biologi Universitas Jenderal Soedirman, Purwokerto dan lulus pada tahun 2012. Pada tahun yang sama penulis diterima di Program Studi Biologi Reproduksi Fakultas Kedokteran Hewan pada Program Pascasarjana IPB. Beasiswa Program Pascasarjana diperoleh dari Beasiswa Unggulan DIKTI 2012.
Karya ilmiah dengan judul “Deteksi Kerusakan Akrosom Spermatozoa