EFEKTIVITAS PROMOTER KERATIN IKAN
FLOUNDER
JEPANG Paralichthys olivaceus DAN PROMOTER
HEATSHOCK IKAN RAINBOW TROUT Oncorhynchus mykiss
PADA IKAN NILA Oreochromis niloticus
ARIEF EKO PRASETIYO
SKRIPSI
PROGRAM STUDI TEKNOLOGI DAN MANAJEMEN AKUAKULTUR FAKULTAS PERIKANAN DAN ILMU KELAUTAN
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER INFORMASI
Dengan ini saya menyampaikan bahwa Skripsi yang berjudul:
EFEKTIVITAS PROMOTER KERATIN IKAN FLOUNDER JEPANG Paralichthys olivaceus DAN PROMOTER HEATSHOCK IKAN RAINBOW TROUT Oncorhynchus mykiss PADA IKAN NILA Oreochromis niloticus
adalah benar merupakan hasil karya yang belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Bogor, Januari 2009
RINGKASAN
ARIEF EKO PRASETIYO. Efektivitas Promoter Keratin Ikan Flounder Jepang Paralichthys olivaceus dan Promoter Heatshock Ikan Rainbow Trout Oncorhynchus mykiss pada Ikan Nila Oreochromis niloticus. Dibimbing oleh KOMAR SUMANTADINATA dan ALIMUDDIN.
Promoter (elemen regulator) adalah bagian dari DNA yang merupakan tempat menempelnya RNA polimerase dan mengarahkannya agar proses transkripsi terjadi sehingga efektivitas promoter suatu konstruksi gen berperan penting dalam menentukan keberhasilan transgenik. Pada penelitian ini dilakukan pengujian efektivitas 2 jenis promoter dalam rangka pembuatan ikan transgenik pada spesies ikan budidaya di Indonesia. Kedua jenis promoter tersebut adalah promoter keratin dari ikan flounder Jepang Paralichthys olivaceus dan promoter heatshock dari ikan rainbow trout Oncorhynchus mykiss yang diujikan pada ikan nila Oreochromis niloticus. Guna mengamati efektivitas kedua promoter, maka disambungkan dengan gen Green Fluorescent Protein (GFP).
Induk ikan nila dipijahkan secara semi buatan, kemudian telur yang sudah terbuahi disuntik dengan konstruksi gen dengan konsentrasi 50 ng/µ l. Telur yang sudah disuntik kemudian diinkubasi ke dalam akuarium dengan suhu air 30oC. Parameter yang diamati antara lain adalah derajat kelangsungan hidup embrio, derajat penetasan, persentase embrio mengekspresikan transgen, dan tingkat ekspresi gen GFP. Pada parameter tingkat ekspresi gen GFP, embrio ikan nila yang mengekspresikan GFP dikelompokkan menjadi 3 kelas yaitu: pendar hijau kurang terang, pendar hijau terang, dan pendar hijau sangat terang.
EFEKTIVITAS PROMOTER KERATIN IKAN
FLOUNDER
JEPANG Paralichthys olivaceus DAN PROMOTER
HEATSHOCK IKAN RAINBOW TROUT Oncorhynchus mykiss
PADA IKAN NILA Oreochromis niloticus
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Perikanan pada Fakultas Perikanan dan Ilmu Kelautan
Institut Pertanian Bogor
Oleh:
Arief Eko Prasetiyo C14104041
PROGRAM STUDI TEKNOLOGI DAN MANAJEMEN AKUAKULTUR FAKULTAS PERIKANAN DAN ILMU KELAUTAN
Judul : Efektivitas Promoter Keratin Ikan Flounder Jepang Paralichthys olivaceus dan Promoter Heatshock Ikan Rainbow Trout Oncorhynchus mykiss pada Ikan Nila Oreochromis niloticus
Nama : Arief Eko Prasetiyo
NRP : C14104041
Disetujui,
Pembimbing I Pembimbing II
Prof. Dr. Komar Sumantadinata NIP. 130 345 017
Dr. Alimuddin NIP. 132 133 953
Diketahui,
Dekan Fakultas Perikanan dan Ilmu Kelautan
Prof. Dr. Ir. Indra Jaya, M.Sc NIP. 131 578 799
DAFTAR RIWAYAT HIDUP
Penulis dilahirkan di Nganjuk pada tanggal 15 April 1986 dari rahim seorang ibu Titik Pangastuti dan bapak Suharto. Pendidikan formal yang telah dilalui adalah SMUN I Puri Mojokerto dan lulus pada tahun 2004. Pada tahun yang sama penulis melanjutkan pendidikan tinggi di IPB melalui jalur Undangan Seleksi Masuk Institut Pertanian Bogor dan memilih program studi Teknologi dan Manajemen Akuakultur, Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan.
Selama mengikuti perkuliahan, penulis pernah melaksanakan kegiatan praktek kerja lapang (PKL) di Balai Besar Pengembangan Budidaya Air Payau (BBPBAP) Jepara. Penulis juga pernah menjadi asisten mata kuliah Pendidikan Agama Islam semester ganjil 2005/2006, asisten mata kuliah Ikhtiologi periode 2006/2007 hingga 2007/2008, asisten mata kuliah Dasar-dasar Genetika Ikan periode 2007/2008, dan asisten mata kuliah Dasar-dasar Akuakultur periode 2006/2007 hingga 2008/2009.
Selain itu penulis juga aktif dalam kegiatan keorganisasian mahasiswa dan tercatat sebagai pengurus Badan Eksekutif Mahasiswa Tingkat Persiapan Bersama (BEM-TPB) selaku Ketua Departemen Politik dan Organisasi 2004/2005, pengurus Badan Eksekutif Mahasiswa Fakultas Perikanan dan Ilmu Kelautan (BEM FPIK) selaku Sekretaris Departemen Pengembangan Sumber Daya Manusia 2005/2006, dan pada periode 2006/2007 penulis mendapatkan amanah sebagai Ketua BEM FPIK.
KATA PENGANTAR
Puji syukur tiada hentinya penulis panjatkan kepada Allah SWT atas segala limpahan karuniaNya sehingga karya ilmiah ini berhasil terselesaikan. Tema yang dipilih pada penelitian ini adalah genetika ikan, dengan judul
”Efektivitas Promoter Keratin Ikan Flounder Jepang Paralichthys olivaceus dan Ikan Rainbow Trout Onchorhyncus mykiss pada Ikan Nila Oreochromis niloticus”.
Keberhasilan penulis dalam menyelesaikan penelitian ini tidak semata didapatkan sendiri, melainkan dengan bantuan orang-orang sekitar. Untuk itu penulis mengucapkan terima kasih kepada:
1. Prof. Dr. Komar Sumantadinata selaku Pembimbing I dan Pembimbing Akademik yang telah membimbing penulis sejak awal perkuliahan sampai dengan penyusunan karya ilmiah ini.
2. Dr. Alimuddin selaku Pembimbing II yang telah membimbing dan mengarahkan penulis selama melakukan penelitian sampai dengan penyusunan karya ilmiah ini.
3. Ibu dan Bapak yang telah memberikan doa restu dan dukungan moril dan materil.
4. Bpk. Adi Sucipto, Bpk. Dian Hardiantho, Bpk. Ayi Santika dan Bpk. Maskur atas bantuan dan ijinnya sehingga penelitian ini dapat terlaksana dengan baik di BBPBAT Sukabumi.
5. Saudari Lina Mulyati, Anna Octavera, Prihanik Marlina, dan Dwi Hany Yanti yang telah membantu persiapan dan pelaksanaan penelitian.
6. Radi Ihlas Albani, Mauluddin, Firman Nazrasul Hakim, dan Lazuardi Yudha dan semua orang yang tidak bisa disebutkan satu persatu.
Penulis telah berusaha semaksimal mungkin dalam penulisan ini. Akhirnya, diantara kelebihan dan kekurangannya, penulis berharap skripsi ini dapat bermanfaat bagi penulis dan para pembaca pada umumnya.
Bogor, Januari 2009
Arief Eko Prasetiyo
DAFTAR ISI
Halaman
KATA PENGANTAR ... i
DAFTAR GAMBAR ... iv
DAFTAR LAMPIRAN ... v
I. PENDAHULUAN ... 1
1.1 Latar Belakang ... 1
1.2 Tujuan ... 3
II. TINJAUAN PUSTAKA ... 4
2.1 Ikan Nila Oreochromis niloticus ... 4
2.2 Teknologi Transgenesis dengan Mikroinjeksi ... 5
2.3 Promoter ... 6
2.4 Promoter Keratin ... 8
2.5 Promoter Heatshock ... 9
2.6 Gen Green Fluorescent Protein (GFP) ...10
2.7 Ekspresi Gen Sementara (Transient Gene Expression) ...10
III. BAHAN DAN METODE ...12
3.1 Waktu dan Tempat ...12
3.2 Prosedur Kerja ...12
3.2.1 Pemeliharaan Induk, Pemijahan dan Pengambilan Telur ...12
3.2.2 Persiapan Mikroinjeksi dan Loading DNA ...13
3.2.3 Pelaksanaan Mikroinjeksi ...15
3.2.3 Pengamatan GFP ...15
3.3 Parameter yang Diamati dan Analisis Data ...16
3.3.1 Derajat Kelangsungan Hidup Embrio (DKHE) ...16
3.3.2 Derajat Penetasan (DP) ...17
3.3.3 Persentase Embrio Mengekspresikan Transgen (PEMT) ...17
3.3.4 Tingkat Ekspresi Gen GFP ...17
IV. HASIL DAN PEMBAHASAN ...19
4.1 Hasil...19
4.1.1 Derajat Kelangsungan Hidup Embrio (DKHE) dan Derajat Penetasan (DP) ...19
4.1.2 Pola Ekspresi Gen GFP dengan Promoter Keratin dan Promoter Heatshock ...19
4.1.3 Persentase Embrio Mengekspresikan Transgen (PEMT) dan Tingkat Ekspresi Gen GFP ...20
4.2 Pembahasan ...22
V. KESIMPULAN dan SARAN ...25
5.1 Kesimpulan ... 25
5.2 Saran ... 25
DAFTAR PUSTAKA ...26
LAMPIRAN ...33
DAFTAR GAMBAR
Halaman
1. Ikan nila Oreochromis niloticus ... 4
2. Sekuens promoter keratin (Yazawa et al., 2005) ... 8
3. Telur ikan nila O. niloticus yang diletakkan pada cekungan gel agarose ...13
4. Seperangkat peralatan mikroinjeksi ...14
5. Telur ikan nila O. niloticus yang sedang diinjeksi gen GFP ...15
6. Satu set peralatan untuk pengamatan GFP ...16
7. Pola ekspresi Gen GFP dengan promoter keratin dan promoter heatshock pada embrio ikan nila O.niloticus ...20
8. Tiga kelas ekspresi gen GFP pada embrio ikan nila O.niloticus ...20
9. Ekspresi gen GFP dengan promoter keratin dan promoter heatshock pada embrio ikan nila O. niloticus ...21
EFEKTIVITAS PROMOTER KERATIN IKAN
FLOUNDER
JEPANG Paralichthys olivaceus DAN PROMOTER
HEATSHOCK IKAN RAINBOW TROUT Oncorhynchus mykiss
PADA IKAN NILA Oreochromis niloticus
ARIEF EKO PRASETIYO
SKRIPSI
PROGRAM STUDI TEKNOLOGI DAN MANAJEMEN AKUAKULTUR FAKULTAS PERIKANAN DAN ILMU KELAUTAN
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER INFORMASI
Dengan ini saya menyampaikan bahwa Skripsi yang berjudul:
EFEKTIVITAS PROMOTER KERATIN IKAN FLOUNDER JEPANG Paralichthys olivaceus DAN PROMOTER HEATSHOCK IKAN RAINBOW TROUT Oncorhynchus mykiss PADA IKAN NILA Oreochromis niloticus
adalah benar merupakan hasil karya yang belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Bogor, Januari 2009
RINGKASAN
ARIEF EKO PRASETIYO. Efektivitas Promoter Keratin Ikan Flounder Jepang Paralichthys olivaceus dan Promoter Heatshock Ikan Rainbow Trout Oncorhynchus mykiss pada Ikan Nila Oreochromis niloticus. Dibimbing oleh KOMAR SUMANTADINATA dan ALIMUDDIN.
Promoter (elemen regulator) adalah bagian dari DNA yang merupakan tempat menempelnya RNA polimerase dan mengarahkannya agar proses transkripsi terjadi sehingga efektivitas promoter suatu konstruksi gen berperan penting dalam menentukan keberhasilan transgenik. Pada penelitian ini dilakukan pengujian efektivitas 2 jenis promoter dalam rangka pembuatan ikan transgenik pada spesies ikan budidaya di Indonesia. Kedua jenis promoter tersebut adalah promoter keratin dari ikan flounder Jepang Paralichthys olivaceus dan promoter heatshock dari ikan rainbow trout Oncorhynchus mykiss yang diujikan pada ikan nila Oreochromis niloticus. Guna mengamati efektivitas kedua promoter, maka disambungkan dengan gen Green Fluorescent Protein (GFP).
Induk ikan nila dipijahkan secara semi buatan, kemudian telur yang sudah terbuahi disuntik dengan konstruksi gen dengan konsentrasi 50 ng/µ l. Telur yang sudah disuntik kemudian diinkubasi ke dalam akuarium dengan suhu air 30oC. Parameter yang diamati antara lain adalah derajat kelangsungan hidup embrio, derajat penetasan, persentase embrio mengekspresikan transgen, dan tingkat ekspresi gen GFP. Pada parameter tingkat ekspresi gen GFP, embrio ikan nila yang mengekspresikan GFP dikelompokkan menjadi 3 kelas yaitu: pendar hijau kurang terang, pendar hijau terang, dan pendar hijau sangat terang.
EFEKTIVITAS PROMOTER KERATIN IKAN
FLOUNDER
JEPANG Paralichthys olivaceus DAN PROMOTER
HEATSHOCK IKAN RAINBOW TROUT Oncorhynchus mykiss
PADA IKAN NILA Oreochromis niloticus
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Perikanan pada Fakultas Perikanan dan Ilmu Kelautan
Institut Pertanian Bogor
Oleh:
Arief Eko Prasetiyo C14104041
PROGRAM STUDI TEKNOLOGI DAN MANAJEMEN AKUAKULTUR FAKULTAS PERIKANAN DAN ILMU KELAUTAN
Judul : Efektivitas Promoter Keratin Ikan Flounder Jepang Paralichthys olivaceus dan Promoter Heatshock Ikan Rainbow Trout Oncorhynchus mykiss pada Ikan Nila Oreochromis niloticus
Nama : Arief Eko Prasetiyo
NRP : C14104041
Disetujui,
Pembimbing I Pembimbing II
Prof. Dr. Komar Sumantadinata NIP. 130 345 017
Dr. Alimuddin NIP. 132 133 953
Diketahui,
Dekan Fakultas Perikanan dan Ilmu Kelautan
Prof. Dr. Ir. Indra Jaya, M.Sc NIP. 131 578 799
DAFTAR RIWAYAT HIDUP
Penulis dilahirkan di Nganjuk pada tanggal 15 April 1986 dari rahim seorang ibu Titik Pangastuti dan bapak Suharto. Pendidikan formal yang telah dilalui adalah SMUN I Puri Mojokerto dan lulus pada tahun 2004. Pada tahun yang sama penulis melanjutkan pendidikan tinggi di IPB melalui jalur Undangan Seleksi Masuk Institut Pertanian Bogor dan memilih program studi Teknologi dan Manajemen Akuakultur, Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan.
Selama mengikuti perkuliahan, penulis pernah melaksanakan kegiatan praktek kerja lapang (PKL) di Balai Besar Pengembangan Budidaya Air Payau (BBPBAP) Jepara. Penulis juga pernah menjadi asisten mata kuliah Pendidikan Agama Islam semester ganjil 2005/2006, asisten mata kuliah Ikhtiologi periode 2006/2007 hingga 2007/2008, asisten mata kuliah Dasar-dasar Genetika Ikan periode 2007/2008, dan asisten mata kuliah Dasar-dasar Akuakultur periode 2006/2007 hingga 2008/2009.
Selain itu penulis juga aktif dalam kegiatan keorganisasian mahasiswa dan tercatat sebagai pengurus Badan Eksekutif Mahasiswa Tingkat Persiapan Bersama (BEM-TPB) selaku Ketua Departemen Politik dan Organisasi 2004/2005, pengurus Badan Eksekutif Mahasiswa Fakultas Perikanan dan Ilmu Kelautan (BEM FPIK) selaku Sekretaris Departemen Pengembangan Sumber Daya Manusia 2005/2006, dan pada periode 2006/2007 penulis mendapatkan amanah sebagai Ketua BEM FPIK.
KATA PENGANTAR
Puji syukur tiada hentinya penulis panjatkan kepada Allah SWT atas segala limpahan karuniaNya sehingga karya ilmiah ini berhasil terselesaikan. Tema yang dipilih pada penelitian ini adalah genetika ikan, dengan judul
”Efektivitas Promoter Keratin Ikan Flounder Jepang Paralichthys olivaceus dan Ikan Rainbow Trout Onchorhyncus mykiss pada Ikan Nila Oreochromis niloticus”.
Keberhasilan penulis dalam menyelesaikan penelitian ini tidak semata didapatkan sendiri, melainkan dengan bantuan orang-orang sekitar. Untuk itu penulis mengucapkan terima kasih kepada:
1. Prof. Dr. Komar Sumantadinata selaku Pembimbing I dan Pembimbing Akademik yang telah membimbing penulis sejak awal perkuliahan sampai dengan penyusunan karya ilmiah ini.
2. Dr. Alimuddin selaku Pembimbing II yang telah membimbing dan mengarahkan penulis selama melakukan penelitian sampai dengan penyusunan karya ilmiah ini.
3. Ibu dan Bapak yang telah memberikan doa restu dan dukungan moril dan materil.
4. Bpk. Adi Sucipto, Bpk. Dian Hardiantho, Bpk. Ayi Santika dan Bpk. Maskur atas bantuan dan ijinnya sehingga penelitian ini dapat terlaksana dengan baik di BBPBAT Sukabumi.
5. Saudari Lina Mulyati, Anna Octavera, Prihanik Marlina, dan Dwi Hany Yanti yang telah membantu persiapan dan pelaksanaan penelitian.
6. Radi Ihlas Albani, Mauluddin, Firman Nazrasul Hakim, dan Lazuardi Yudha dan semua orang yang tidak bisa disebutkan satu persatu.
Penulis telah berusaha semaksimal mungkin dalam penulisan ini. Akhirnya, diantara kelebihan dan kekurangannya, penulis berharap skripsi ini dapat bermanfaat bagi penulis dan para pembaca pada umumnya.
Bogor, Januari 2009
Arief Eko Prasetiyo
DAFTAR ISI
Halaman
KATA PENGANTAR ... i
DAFTAR GAMBAR ... iv
DAFTAR LAMPIRAN ... v
I. PENDAHULUAN ... 1
1.1 Latar Belakang ... 1
1.2 Tujuan ... 3
II. TINJAUAN PUSTAKA ... 4
2.1 Ikan Nila Oreochromis niloticus ... 4
2.2 Teknologi Transgenesis dengan Mikroinjeksi ... 5
2.3 Promoter ... 6
2.4 Promoter Keratin ... 8
2.5 Promoter Heatshock ... 9
2.6 Gen Green Fluorescent Protein (GFP) ...10
2.7 Ekspresi Gen Sementara (Transient Gene Expression) ...10
III. BAHAN DAN METODE ...12
3.1 Waktu dan Tempat ...12
3.2 Prosedur Kerja ...12
3.2.1 Pemeliharaan Induk, Pemijahan dan Pengambilan Telur ...12
3.2.2 Persiapan Mikroinjeksi dan Loading DNA ...13
3.2.3 Pelaksanaan Mikroinjeksi ...15
3.2.3 Pengamatan GFP ...15
3.3 Parameter yang Diamati dan Analisis Data ...16
3.3.1 Derajat Kelangsungan Hidup Embrio (DKHE) ...16
3.3.2 Derajat Penetasan (DP) ...17
3.3.3 Persentase Embrio Mengekspresikan Transgen (PEMT) ...17
3.3.4 Tingkat Ekspresi Gen GFP ...17
IV. HASIL DAN PEMBAHASAN ...19
4.1 Hasil...19
4.1.1 Derajat Kelangsungan Hidup Embrio (DKHE) dan Derajat Penetasan (DP) ...19
4.1.2 Pola Ekspresi Gen GFP dengan Promoter Keratin dan Promoter Heatshock ...19
4.1.3 Persentase Embrio Mengekspresikan Transgen (PEMT) dan Tingkat Ekspresi Gen GFP ...20
4.2 Pembahasan ...22
V. KESIMPULAN dan SARAN ...25
5.1 Kesimpulan ... 25
5.2 Saran ... 25
DAFTAR PUSTAKA ...26
LAMPIRAN ...33
DAFTAR GAMBAR
Halaman
1. Ikan nila Oreochromis niloticus ... 4
2. Sekuens promoter keratin (Yazawa et al., 2005) ... 8
3. Telur ikan nila O. niloticus yang diletakkan pada cekungan gel agarose ...13
4. Seperangkat peralatan mikroinjeksi ...14
5. Telur ikan nila O. niloticus yang sedang diinjeksi gen GFP ...15
6. Satu set peralatan untuk pengamatan GFP ...16
7. Pola ekspresi Gen GFP dengan promoter keratin dan promoter heatshock pada embrio ikan nila O.niloticus ...20
8. Tiga kelas ekspresi gen GFP pada embrio ikan nila O.niloticus ...20
9. Ekspresi gen GFP dengan promoter keratin dan promoter heatshock pada embrio ikan nila O. niloticus ...21
DAFTAR LAMPIRAN
Halaman
1. Embriogenesis ikan nila O. niloticus (Fujimura & Okada, 2007) ...34
2. Pemijahan ikan nila O. niloticus dengan sistem semi buatan ...35
3. Persiapan alat mikroinjeksi...36
4. Konstruksi DNA (Keratin-GFP) dan (Heatshock-GFP) ...37
5. Persiapan kontruksi DNA berupa perbanyakan dan isolasi plasmid ...38
6. Proses memasukkan DNA dalam jarum mikroinjeksi (Loading DNA) ...39
7. Derajat Kelangsungan Hidup Embrio (DKHE) dan Derajat Penetasan (DP), pada ikan nila O. niloticus ...40
8. Persentase Embrio Mengekspresikan Transgen (PEMT) dan tingkat ekspresi gen GFP pada ikan nila O. niloticus ...41
I. PENDAHULUAN
1.1 Latar Belakang
Teknologi transgenesis adalah teknologi rekayasa melalui introduksi DNA asing (eksogenus DNA) ke dalam hewan uji guna mengubah dan memanipulasikan struktur genetiknya (Glick & Pasternak, 2003). Dalam usaha budidaya perikanan teknologi transgenesis ini dapat bermanfaat untuk meningkatkan laju pertumbuhan ikan dan krustase (Devlin et al., 1995), meningkatkan daya tahan terhadap penyakit (Dunham, 2004), mengurangi laju konsumsi oksigen pada ikan (Cook et al., 2000), serta untuk memproduksi protein farmaseutika/nutraseutika yang berguna bagi akuakultur dan kesehatan manusia (Kinoshita & Ozato, 1995; Fletcher & Davies, 1991; Collas et al., 2000).
Hackett (1993) menyebutkan bahwa, ada tiga tahapan utama untuk menghasilkan ikan transgenik, yaitu (1) mempersiapkan konstruksi gen yang tersusun atas gen penyandi protein tertentu dan promoter (regulator element); (2) mengintroduksi konstruksi gen ke dalam inti sel pada embrio yang sedang berkembang supaya bisa didistribusikan ke semua jaringan ikan; (3) mengidentifikasi individu ikan yang mengekspresikan gen asing atau transgen. Promoter adalah bagian dari DNA yang merupakan tempat menempelnya RNA polimerase dan mengarahkannya agar proses transkripsi terjadi (Glick & Pasternak, 2003). Efektivitas promoter suatu konstruksi gen berperan penting dalam menentukan keberhasilan teknologi transgenesis (Alimuddin et al., 2008; Nugrahani, 2008).
Promoter keratin merupakan promoter yang diisolasi dari ikan flounder Jepang Paralichthys olivaceus (Hirono et al., 2003). Pada awalnya promoter keratin digunakan dalam pembuatan ikan transgenik yang ditujukan untuk mengontrol ekspresi gen-gen yang berhubungan dengan imunitas ikan (Yazawa et al., 2005) dan pembuatan ikan hias warna-warni karena efektivitasnya yang tinggi
ikan zebra (Yazawa et al., 2005). Promoter keratin ikan flounder Jepang diketahui memiliki sifat dapat aktif dimana-mana (ubiquitous) dan aktif kapan saja diperlukan (house keeping) (Yazawa et al., 2005).
Promoter heatshock adalah suatu promoter yang aktivasi dan efektivitas optimalnya bergantung pada pengaruh rangsangan yang berasal dari lingkungan (Ojima, 2007). Sehingga promoter heatshock tergolong sebagai promoter yang dapat diinduksi (inducible promoter). Kemampuan tersebut sering dimanfaatkan oleh para peneliti untuk mendeteksi (biodetector) suatu kondisi lingkungan yang menyimpang serta proses studi biologi suatu organisme (Milioni et al., 2001; Wu et al., 2007). Sementara itu, sifat promoter heatshock yang diisolasi dari ikan rainbow trout (Kawamura & Yoshizaki, tidak dipublikasikan) belum diverifikasi. Namun demikian, ada beberapa promoter sejenis yang memiliki sifat serupa dengan promoter heatshock telah dilaporkan tergolong sebagaiinducible promoter yaitu promoter hspb1/hsp27 (Ojima, 2007). Promoter lain yang serupa dengan heatshock yaitu hsp27, dapat pula digunakan untuk mengembangkan organisme
transgenik sebagai biodetector kualitas lingkungan dari pencemaran arsenik (Wu et al., 2007).
Kedua promoter tersebut diintroduksi ke Indonesia oleh Dr. Alimuddin pada tahun 2008, sehingga perlu dilakukan pengujian terkait efektivitasnya pada ikan-ikan budidaya di Indonesia. Pada percobaan ini dilakukan pengujian efektivitas promoter keratin dan heatshock pada ikan nila Oreochromis niloticus. Masing-masing promoter tersebut disambungkan dengan gen penyandi protein berpendar hijau (green fluorescent protein, GFP) sehingga terbentuk konstruksi keratin-GFP (k-GFP) dan konstruksi heatshock-GFP (hsc-GFP). Efektivitas
LacZ yang membutuhkan X-gal untuk visualisasi ekspresinya (Iyengar et al.,
1996).
1.2 Tujuan
Penelitian ini bertujuan untuk mengetahui efektifitas promoter keratin ikan flounder Jepang dan promoter heatshock ikan rainbow trout pada ikan nila dengan
II. TINJAUAN PUSTAKA
2.1 Ikan Nila Oreochromis niloticus
Ikan nila (Gambar 1) adalah ikan yang tergolong ke dalam famili Cichlidae genus Oreochromis dan memiliki nama latin spesies Oreochromis niloticus (Trewavas, 1983). Ikan nila merupakan salah satu jenis ikan yang
diintroduksikan dari luar negeri. Ikan tersebut berasal dari Afrika bagian timur di Sungai Nil, Danau Tangayika, Chad, Nigeria dan Kenya, lalu dibawa oleh orang ke Eropa, Amerika, negara-negara Timur Tengah dan Asia. Di Indonesia benih ikan nila secara resmi didatangkan dari Taiwan oleh Balai Penelitian Perikanan Air Tawar tahun 1969 (Suyanto, 1994 dalam Saputra, 2007).
Gambar 1. Ikan nila Oreochromis niloticus
Ikan nila memiliki karakteristik sebagai ikan parental care yang merawat anaknya dengan menggunakan mulut (mouth breeder) (Effendie, 1997). Ikan ini dicirikan dengan garis vertikal yang berwarna gelap pada sirip ekornya sebanyak 6 buah. Selain pada sirip ekor, garis tersebut juga terdapat pada sirip punggung dan sirip anal (Suyanto, 1994 dalam Saputra, 2007).
Keunikan lain dari ikan nila ditunjukkan dari bentuk telurnya yang lonjong serta perkembangan embriogenesisnya (Lampiran 1) yang mencapai 90-110 jam pasca pembuahan, seperti yang dilaporkan oleh Fujimura & Okada (2007).
(Majumdar & McAndrew, 1986; Kocher et al., 1998; Oliveira & Wright, 1998; McConnell et al., 2000; Maclean et al., 2002; Lee et al., 2003; Katagiri et al. 2005; Lee et al., 2005; Santini & Bernardi, 2005; Fujimura & Okada, 2007; Kobayashi et al., 2007). Ikan nila merupakan salah satu ikan konsumsi yang menjadi target dari pengembangan rekayasa transgenik selain ikan mas, salmon dan channel catfish (Alestrom, 2007).
2.2 Teknologi Transgenesis dengan Mikroinjeksi
Teknologi transgenesis atau teknologi transfer gen ini merupakan suatu proses mengintroduksi DNA eksogenus atau DNA asing ke embrio dengan tujuan untuk memanipulasi struktur genetiknya (Glick & Pasternak, 2003). Dalam usaha budidaya perikanan teknologi transgenik ini berguna untuk meningkatkan laju pertumbuhan ikan dan krustase (Devlin et al., 1995), meningkatkan daya tahan terhadap penyakit (Dunham, 2004), mengurangi laju konsumsi oksigen pada ikan (Cook et al., 2000), serta untuk memproduksi protein farmaseutika/nutraseutika yang berguna bagi akuakultur dan kesehatan manusia (Kinoshita & Ozato, 1995; Fletcher & Davies, 1991; Collas et al., 2000). Sebagai contoh aplikasi teknologi transgenesis yang telah berhasil diantaranya adalah peningkatan daya tahan terhadap bakteri patogen dengan mengintroduksikan peptida cecropin-B pada channel catfish transgenik (Dunham, et al., 2000 dalam Dunham, 2004); peningkatan laju pertumbuhan pada ikan salmon Pasifik (Devlin et al., 1995), ikan mud loach (Nam et al., 2001), dan ikan nila (Kobayashi et al., 2007).
Kinoshita dan Ozato (1995) menjelaskan bahwa teknologi transgenesis tidak hanya berperan dalam kegiatan produksi, melainkan juga memiliki peranan dalam studi terkait fungsi, proses dan pola ekspresi dari suatu konstruksi gen. Garcia et al. (1998) dalam Ath-thar (2007) menjelaskan bahwa transfer DNA eksogenus ke dalam embrio ikan maupun hewan lainnya memiliki dua tujuan utama yaitu terkait studi tentang fungsi pengaturan gen selama perkembangan dan manipulasi genetika dari ikan komersial untuk memproduksi galur transgenik yang memiliki nilai komersial yang lebih tinggi.
(1996) menyebutkan bahwa pada saat ini, mikroinjeksi DNA asing ke dalam sitoplasma adalah metode yang paling banyak digunakan untuk kepentingan komersial. Hackett (1993) menambahkan bahwa mikroinjeksi adalah teknik yang paling sering digunakan pada ikan.
Pada teknik mikroinjeksi guna memastikan material genetik masuk ke pronuklei, konsentrasi yang tinggi dari konstruksi DNA (104-107 kopi) biasanya diinjeksikan ke dalam sitoplasma dari telur yang telah dibuahi (Ath-thar, 2007). Injeksi dengan jumlah kopi DNA yang tinggi meningkatkan integrasi transgen (DNA yang ditransfer), tetapi hal itu meningkatkan resiko kematian pada embrio (Ath-thar, 2007; Purwanti, 2007). Integrasi transgen pada DNA inang umumnya tidak terjadi pada fase satu sel, sehingga tidak semua sel ikan membawa transgen. Kejadian tersebut dikenal dengan istilah mozaik (Zbikowska, 2003).
Keterbatasan metode ini antara lain telur harus diperlakukan satu persatu dan injeksi secara langsung ke nukleus tidak mungkin dilakukan pada ikan, tidak seperti pada mamalia. Mikroinjeksi ke sitoplasma memerlukan jumlah kopi gen yang banyak. Hal ini yang menyebabkan tingginya kejadian mozaik (Ath-thar, 2007). Mikroinjeksi juga memiliki resiko tinggi terhadap kelangsungan hidup embrio (Zbikowska, 2003; Ath-thar, 2007). Namun metode mikroinjeksi memiliki kelebihan yaitu efisiensi penggunaan DNA dan adanya kemungkinan untuk dapat mengontrol letak injeksi pada telur. Untuk telur dari jenis ikan yang sulit untuk didekorionasi mikroinjeksi dilakukan melalui lubang mikrofil. Pada metode ini secara umum telur langsung diinjeksi pada blastodisk. Tempat injeksi yang acak ini sangat berpengaruh terhadap gerakan transfer DNA dari yolk atau sitoplasma ke nukleus. Transfer DNA lebih cepat ketika telur diinjeksi pada blastodisk yang berlawanan arah dengan vegetal-pole (Collas et al., 2000).
2.3 Promoter
penting dalam menentukan keberhasilan transgenesis (Alimuddin et al., 2008; Nugrahani, 2008).
Promoter merupakan sekuen DNA yang terletak di hulu (upstream, terminal 5) dari lokasi dimulainya transkripsi (Hackett, 1993). Daerah promoter adalah cis-acting (faktor transkripsi); promoter mempengaruhi transkripsi dalam segmen DNA yang sama dimana promoter tersebut berada. Sekuen ini dikenali oleh RNA polimerase yang kemudian menempel dan mengendalikan proses transkripsi (Hacket, 1993; Glick & Pasternak, 2003). Promoter merupakan salah satu penentu/pengatur spatial-temporal ekspresi gen, sehingga promoter bisa dianalogikan sebagai switch suatu gen.
Beberapa promoter telah diisolasi dan dilakukan proses pengujian oleh beberapa peneliti, antara lain cytomegalovirus (CMV) dari virus manusia (Dunham, 2004); actin dari ikan mas (Liu et al., 1990), ikan medaka (Takagi et al., 1994), ikan zebra (Higashijima et al., 1997), ikan mud loach (Noh et al., 2003), ikan kerapu bebek (Nugrahani, 2008) dan ikan nila (Alimuddin et al.,2008).
Efektivitas suatu promoter dipengaruhi oleh kesesuaian cis-acting promoter dan trans-acting inang (Iyengar et al., 1996; Alimuddin, 2003) sehingga dalam penerapannya sering kali digunakan promoter yang homolog dengan hewan uji (Kawamoto et al., 1989). Keberadaan sekuen pemicu aktivitas (enhancer) pada konstruksi gen dapat pula memberikan pengaruh positif terhadap efektivitas suatu promoter (Iyengar et al., 1996). Penggunaan promoter yang tidak sesuai dapat berakibat terhambatnya proses ekspresi gen, sebaliknya apabila promoter yang digunakan sesuai maka dapat meningkatkan derajat integrasi dan tingkat ekspresi gen tersebut pada sel inang (Dunham, 2004).
2.4 Promoter Keratin
Promoter keratin merupakan promoter yang diisolasi dari ikan flounder Jepang Paralichthys olivaceus (Hirono et al., 2003). Pada awalnya promoter keratin merupakan promoter yang digunakan pada teknologi transgenesis yang terkait dengan sistem imun, karena efektivitasnya yang tinggi pada jaringan kulit (Gong et al., 2002). Namun Giordano et al. (1990) menyebutkan bahwa efektivitas promoter keratin tidak hanya terbatas pada jaringan kulit dan epitel, tapi juga terdapat pada sel yang sedang berkembang dan sel saraf tertentu. Sedangkan Yazawa et al. (2005) menjelaskan bahwa promoter keratin yang diujikan pada ikan zebra mampu bersifat dapat aktif dimana-mana atau tidak spesifik jaringan tertentu (ubiquitous) dan dapat aktif kapan saja diperlukan (house keeping).
Para peneliti telah melakukan karakterisasi promoter keratin pada berbagai jenis ikan, antara lain pada ikan rainbow trout (Markl & Franke, 1988; Markl et al., 1989), ikan koki (Giordano et al., 1990; Druger et al.,1992), catfish (Arenas et
al., 1995), common carp (Groff et al., 1997), ikan hiu (Schaffeld et al., 1998), serta ikan zebra (Conrad et al., 1998; Yazawa et al.,2005).
Gambar 2. Sekuens promoter keratin (Yazawa et al., 2005)
coding region dan asam amino ditunjukkan dengan huruf yang dicetak tebal,
bagian awal transkripsi ditunjukkan dengan tanda bintang (*) dan bagian predictive transcriptional factor binding ditunjukkan dengan garis bawah.
2.5 Promoter Heatshock
Promoter heatshock merupakan promoter yang diisolasi dari ikan rainbow trout (Kawamura & Yoshizaki, tidak dipublikasikan) dan dikonstruksi oleh Yamamoto dan Yoshizaki (tidak dipublikasikan), diintroduksikan ke Indonesia pada tahun 2008 oleh Dr. Alimuddin. Promoter heatshock memiliki kesamaan karakter dengan beberapa promoter, antara lain promoter hsp90α & hsp90β (Krone & Sass, 1994; Krone et al., 1997; Milioni et al., 2001), hsp70 (Yeh & Hsu, 2002), hsp16.6 (Feng Fang, 2003), hsf (Ian Birch-Martin et al., 2005), hsf1 (Bajac & Shaham, 2007), hsp27 (Wu et al., 2007).
Promoter ini tergolong sebagai inducible promoter, yaitu suatu promoter yang aktivasi dan efektivitas optimalnya bergantung pada pengaruh rangsangan yang berasal dari lingkungan (Ojima, 2007). Kemampuan tersebut sering dimanfaatkan oleh para peneliti untuk mendeteksi suatu kondisi lingkungan yang menyimpang (biodetector) serta proses studi biologi suatu organisme (Milioni et al., 2001;Wu et al., 2007).
Aktivitas promoter ini pernah diuji pada Cyanobacterium (Feng Fang, 2003), Caenorhabditis elegans (Bacaj & Shaham, 2007), Drosophila (Ian Birch-Martin et al., 2005) dan ikan zebra (Krone & Sass, 1994; Krone et al., 1997; Lele et al., 1997: Yeh & Hsu, 2002; Wu et al., 2007).
2.6 Gen Green Fluorescent Protein (GFP)
Gen GFP (Green Fluorescent Protein) adalah gen berpendar hijau yang diisolasi dari Aequorea victoria (Felts et al., 2001), gen ini memiliki kandungan protein yang dapat berpendar dan divisualisasikan ekspresinya pada sel dengan menggunakan bantuan sinar UV atau mikroskop fluoresen (Chalfie, 1994 dalam Iyengar et al., 1996). GFP memiliki kelebihan yaitu untuk mendeteksi ekspresinya tidak memerlukan substrat tambahan, sehingga dapat digunakan sebagai marker (penanda) dalam pengujian efektivitas suatu promoter (Chou et al., 2001; Gong et al., 2002; Yazawa et al., 2005), dan dapat pula berperan sebagai gen target dalam
pembuatan ikan hias berpendar yang berwarna-warni (Gong et al., 2002).
Pengujian promoter dengan menggunakan GFP sudah banyak dilaporkan berhasil pada beberapa jenis promoter dan spesies yang berbeda antara lain pengujian promoter β-actin pada ikan rainbow trout (Boonanuntanasarn et al., 2002; Yoshizaki, 2001), ikan nila (Kobayashi, 2007), ikan zebra (Williams et al., 1996; Alimuddin et al., 2005), ikan lele (Ath-thar, 2007) dan ikan mas (Purwanti, 2007); pengujian promoter hsp70 (Seok et al., 2007), hsp27 (Wu et al., 2007), dan promoter TNF (Tumor Necrosis Factor), C3, gelatinaseB, keratin (Yazawa et al., 2005) pada ikan zebra.
2.8 Ekspresi Gen Sementara (Transient Gene Expression)
Ekspresi yang dihasilkan oleh suatu gen berhubungan dengan keberadaan dari sekuen intron (Betancourt et al., 1993 dalam Hackett, 1993). Lebih lanjut Betancourt et al. (1993) dalam Hackett (1993) menjelaskan bahwa penggunaan kombinasi intron atau promoter yang tidak sesuai dapat mengakibatkan pemotongan RNA yang salah, sehingga berpengaruh terhadap penghambatan transkripsi dan instabilitas pada mRNA.
penelitian transgenik, diantaranya pada ikan kakap Sparus auratus (Garcia-Pozo et al., 1998), ikan zebra (Higashijima et al., 1997; Meng et al., 1999), ikan medaka (Winkler et al., 1991; Hamada et al., 1998; Chou et al., 2001), ikan lele (Volckaert et al., 1997; Ath-thar, 2007), dan ikan mas (Purwanti, 2007).
Ath-thar (2007) menyebutkan bahwa ekspresi transgen mulai muncul pada saat embrio memasuki fase gastrula awal atau fase permulaan adanya epiboly. Sedangkan Iyengar et al., (1996) menyebutkan bahwa ekspresi tertinggi pada perkembangan embrio, umumnya terjadi pada fase gastrula. Kondisi ini diakibatkan akumulasi DNA yang diinjeksikan yang berlanjut pada peningkatan replikasi pada fase pembelahan (cleavage) dan akumulasi dari enzim RNA polymerase II yang menyebabkan dimulainya transkripsi pada fase MBT (Mid Blastula Transition) (Iyengar et al., 1996; Ath-thar, 2007). Pada pengujian yang dilakukan pada ikan medaka (Hamada et al.,1998) disebutkan bahwa ekspresi gen wtGFP (wild type GFP) dan mtGFP (mutant GFP) dimulai pada fase midblastula
III. BAHAN DAN METODE
3.1 Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan April hingga September 2008. Empat bulan pertama (April hingga Juli) merupakan penelitian pendahuluan berupa persiapan dan perbanyakan konstruksi gen dilaksanakan di Laboratorium Pengembangbiakan dan Genetika Ikan, Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Sementara itu, penelitian utama dilaksanakan pada bulan Agustus hingga September bertempat di Hatchery Nila dan Laboratorium Patologi Balai Besar Pengembangan Budidaya Air Tawar (BBPBAT) Sukabumi.
3.2 Prosedur Kerja
3.2.1 Pemeliharaan Induk, Pemijahan dan Pengambilan Telur
Induk dipelihara dalam 40 buah akuarium berukuran 60x40x40 cm dengan sistem resirkulasi. Masing-masing akuarium diisikan satu ekor induk ikan nila, dengan susunan berurutan jantan-betina. Induk tersebut dipelihara pada kisaran suhu 30oC. Pakan yang digunakan untuk induk, berupa pellet udang P5 yang diberikan dari pukul 07.00 hingga 16.00 secara satiasi.
Pemijahan dilakukan dengan sistem semi buatan (Lampiran 2). Induk jantan dan induk betina yang telah matang, dicampurkan menjadi satu dengan cara memindahkan induk jantan ke dalam akuarium induk betina. Matangnya suatu induk ikan nila, dicirikan dengan adanya tonjolan genital pore. Setelah induk ikan nila betina mengeluarkan sebagian sedikit telurnya, maka induk jantan segera dipindahkan. Induk betina kemudian diambil secara hati-hati dengan menggunakan 2 buah serok secara rangkap (dengan susunan yang memiliki ukuran mata jaring lebih besar di atas), lalu distriping pada wadah mangkuk yang telah disediakan. Setelah telur terkumpul di dalam mangkuk, maka induk jantan distriping pada wadah yang sama untuk mengeluarkan spermanya.
dapat disimpan untuk selanjutnya dilakukan perlakuan mikroinjeksi. Pemijahan dengan sistem semi buatan ini dilakukan guna mendapatkan telur fase satu sel pada saat yang sama dalam jumlah relatif banyak.
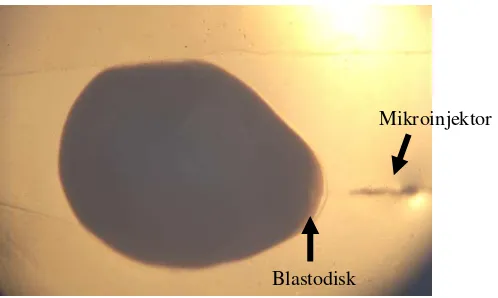
Telur yang sudah dibuahi kemudian disusun diatas cekungan gel agarose 2% dengan menggunakan pipet. Sekitar 8 butir telur dapat diletakkan pada cekungan gel agarose tersebut (Gambar 3). Telur tersebut disusun dengan posisi blastodisk terletak pada bagian cekungan gel yang lebih lebar, sehingga nantinya blastodisk berhadapan langsung dengan arah datangnya jarum mikroinjeksi.
Gambar 3. Telur ikan nila O. niloticus yang diletakkan pada cekungan gel agarose
3.2.2 Persiapan Mikroinjeksi dan Loading DNA
Persiapan alat mikroinjeksi (Lampiran 3) dilakukan dengan cara micromanipulator ditempelkan pada magnetic stand yang telah melekat pada
[image:34.595.191.436.271.433.2]pengamatan serta mengetahui bahwa jarum telah menembus blastodisk telur, digunakan mikroskop (Zeiss Stemi DV4). Rangkaian satu set alat mikroinjeksi ditampilkan pada Gambar 4.
Gambar 4. Seperangkat peralatan mikroinjeksi
Konstruksi DNA (Lampiran 4) yang digunakan berbentuk plasmid yang tersusun dari promoter, gen GFP dan poly A. Pada konstruksi promoter keratin-GFP diligasikan dengan vektor pEkeratin-GFP-NI, sedangkan pada konstruksi heatshock-GFP diligasikan dengan vektor pGEM-T easy. Persiapan kontruksi berupa perbanyakan dan isolasi plasmid disajikan pada Lampiran 5.
memberi sedikit rongga antara larutan DNA dengan minyak mineral. Langkah terakhir adalah capillary glass dilepas dari needle holder dan diganti dengan jarum mikroinjeksi yang telah berisi larutan DNA dengan minyak mineral.
3.2.3 Pelaksanaan Mikroinjeksi
[image:36.595.205.454.391.540.2]Pelaksanaan mikroinjeksi dilakukan segera setelah telur nila dibuahi dan disusun pada cekungan gel agarose. Telur ikan nila yang sudah disusun pada gel plat diletakkan berhadapan dengan jarum mikroinjeksi yang telah diatur posisinya dengan bantuan micromanipulator. Kemudian cairan DNA gen GFP diinjeksikan dengan perlahan pada blastodisk (Gambar 5), masing-masing dilakukan pada 30 butir telur sebanyak 2 kali ulangan. Guna menahan agar telur tidak berputar ketika proses injeksi, maka digunakan tusuk gigi yang dihimpitkan pada bagian belakang telur. Cairan DNA yang diinjeksikan mencapai kira-kira seperlima dari volume blastodisk yang diinjeksi (Ath-thar, 2007).
Gambar 5. Telur ikan nila O. niloticus yang sedang diinjeksi gen GFP
Telur ikan nila yang sudah diinjeksi gen GFP lalu dipindahkan dengan menggunakan mangkuk dan diinkubasi ke dalam akuarium pada suhu 30oC, kemudian dilakukan pengamatan ekspresi GFP.
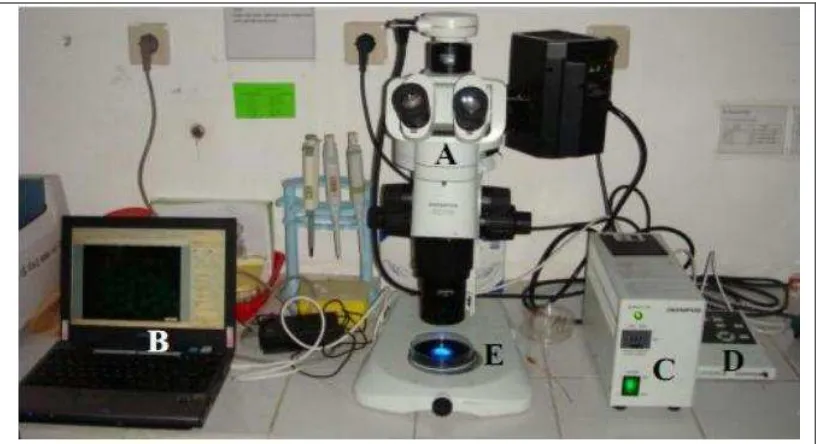
3.2.4 Pengamatan GFP
Pengamatan GFP menggunakan 1 set peralatan untuk pengamatan GFP (Gambar 6) yang terdiri dari mikroskop stereo zoom (Olympus SZX16) yang dilengkapi filter GFP (Olympus SZX2-FGFPHQ), high speed compact color
Blastodisk
digital camera Olympus D20, burner Olympus, remote controller Olympus DP-20
serta laptop yang memiliki software Olympus DH2-BW.
[image:37.595.108.516.332.554.2]Pengamatan GFP dilakukan dengan cara membagi telur menjadi 2 kelompok, satu kelompok digunakan untuk pengamatan pada setiap jam (10 butir) sehingga dapat diketahui waktu ekspresi awal pendaran GFP, puncak ekspresi dan waktu hilangnya ekspresi pendaran GFP. Kelompok lainnya berupa kelompok perlakuan (30 butir) yang hanya dilakukan pengamatan pada saat titik puncak ekspresi pendaran GFP dan pada saat pendaran GFP tidak terlihat lagi. Pada kelompok perlakuan, kegiatan tersebut dilanjutkan hingga telur menetas. Metode seperti ini dilakukan guna menghindari pengaruh penggunaan mikroskop fluoresen pada telur.
Gambar 6. Satu set peralatan untuk pengamatan GFP
3.3 Parameter yang Diamati dan Analisis Data 3.3.1 Derajat Kelangsungan Hidup Embrio (DKHE)
embrio ini dilakukan sebelum telur menetas, menggunakan rumus dari Effendi (1979) sebagai berikut:
� = ℎ ℎ
ℎ � × 100%
3.3.2 Derajat Penetasan (DP)
Derajat penetasan (DP) adalah nilai yang menunjukkan persentase dari jumlah telur ikan nila yang menetas. Perhitungan dilakukan setelah telur menetas secara keseluruhan, menggunakan rumus dari Effendi (1979) sebagai berikut:
�= ℎ
ℎ � × 100%
3.3.3 Persentase Embrio Mengekspresikan Transgen (PEMT)
Persentase embrio mengekspesikan transgen (PEMT) ini menentukan persentase keberhasilan dari injeksi DNA ke dalam telur. Persentase ini didapatkan dari perbandingan jumlah telur yang di dalamnya terdapat ekspresi gen yang telah diinjeksikan dibandingkan dengan jumlah total telur yang telah diinjeksi. Perhitungan derajat ekspresi gen ini dapat dihitung dengan rumus dari Ath-thar (2007) sebagai berikut:
� ��= ℎ � �
ℎ × 100%
3.3.4 Tingkat Ekspresi Gen GFP
3.3.5 Analisis Data
IV. HASIL DAN PEMBAHASAN 4.1 Hasil
4.1.1 Derajat Kelangsungan Hidup Embrio (DKHE) dan Derajat Penetasan (DP)
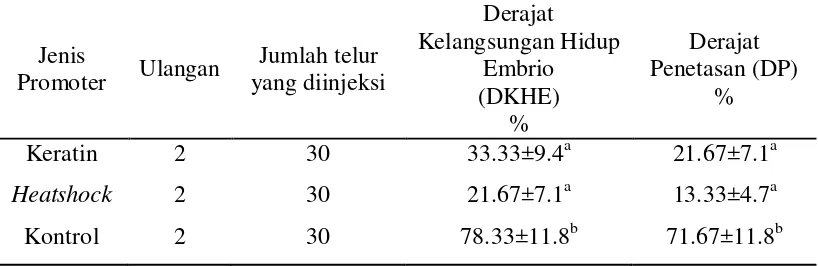
[image:40.595.116.525.362.495.2]Seperti yang ditunjukkan pada Tabel 1, diketahui bahwa parameter DKHE dan DP larva pada kontrol memiliki kisaran nilai yang lebih tinggi (71.67±11.8%), dibandingkan dengan kisaran nilai DKHE dan DP yang diinjeksikan GFP dengan menggunakan promoter keratin (21.67±7.1%) dan promoter heatshock (13.33±4.7%). Sedangkan promoter keratin dan heatshock pada paramater DKHE dan DP menunjukkan nilai yang relatif sama. Data keseluruhan ditunjukkan pada Lampiran 7.
Tabel 1. Derajat Kelangsungan Hidup Embrio dan Derajat Penetasan pada Ikan Nila (O.niloticus)
Jenis
Promoter Ulangan
Jumlah telur yang diinjeksi Derajat Kelangsungan Hidup Embrio (DKHE) % Derajat Penetasan (DP) %
Keratin 2 30 33.33±9.4a 21.67±7.1a
Heatshock 2 30 21.67±7.1a 13.33±4.7a
Kontrol 2 30 78.33±11.8b 71.67±11.8b
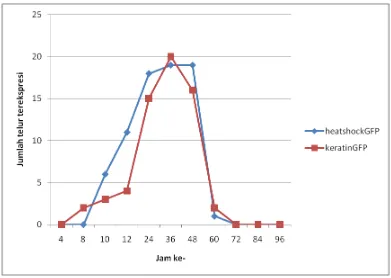
4.1.2 Pola Ekspresi Gen GFP dengan Promoter Keratin dan Promoter Heatshock
Gambar 7. Pola ekspresi gen GFP dengan promoter keratin dan promoter heatshock pada embrio ikan nila O.niloticus
4.1.3 Persentase Embrio Mengekspresikan Transgen (PEMT) dan Tingkat Ekspresi Gen GFP
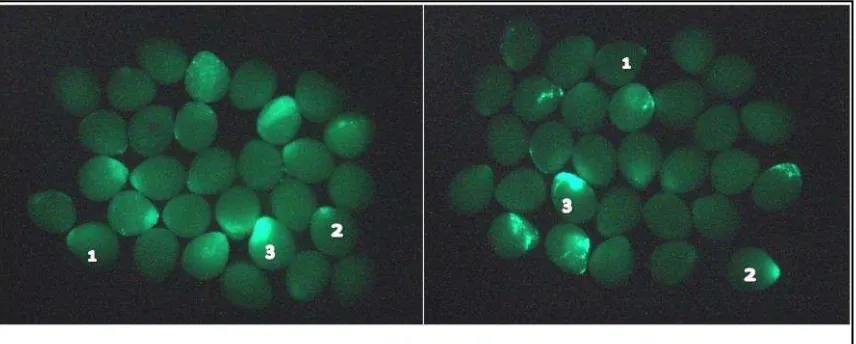
[image:41.595.117.508.82.360.2]Tingkat ekspresi gen GFP ditunjukkan dengan 3 kelas ekspresi gen GFP yang ditampilkan pada Gambar 8.
[image:41.595.173.488.531.728.2]Pada Tabel 2 dan Gambar 9 diketahui bahwa promoter keratin dan heatshock pada parameter persentase embrio mengekspresikan transgen (PEMT) relatif sama, namun promoter keratin mampu mengontrol ekspresi GFP lebih baik dibandingkan dengan promoter heatshock. Hal ini dapat diketahui dari rataan jumlah telur yang mengekspresikan GFP pada tingkat 3 (pendar hijau sangat terang) oleh promoter keratin lebih banyak (8±1.4) dibandingkan dengan promoter heatshock (5±0.0). Jumlah embrio mengekspresikan transgen menunjukkan nilai yang sama; 14±1.4% pada promoter keratin, 15±1.4% pada promoter heatshock. Data keseluruhan ditunjukkan pada Lampiran 8.
Tabel 2. Persentase Embrio Mengekspresikan Transgen (PEMT) dan Tingkat Ekspresi GFP pada Embrio Ikan Nila O.niloticus
Jenis
Promoter Ulangan
Jumlah telur yang diinjeksi
Persentase Embrio Mengekspresikan
Transgen (PEMT)
%
Tingkat Ekspresi (Jumlah Telur)
(1) (2) (3)
Keratin 2 30 43.34±4.7a 1.5±0.7a 3±1.4a 8±1.4a
Heatshock 2 30 50.00±4.7a 5.5±0.7b 4.5±0.7a 5±0.0b
(1): ekspresi pendar hijau kurang terang; (2): ekspresi pendar hijau terang, dan (3): ekspresi pendar hijau sangat terang.
[image:42.595.114.562.319.445.2] [image:42.595.117.545.502.674.2]4.2 Pembahasan
Telur kontrol memiliki nilai derajat kelangsungan hidup embrio (DKHE) dan derajat penetasan (DP) yang cukup tinggi menunjukkan bahwa kualitas telur ikan nila hasil pembuahan semi buatan memiliki kualitas yang baik.
Nilai DKHE dan DP pada telur kontrol relatif lebih tinggi dibandingkan dengan telur yang diberi perlakuan mikroinjeksi gen GFP dengan promoter keratin dan promoter heatshock. Rendahnya derajat kelangsungan hidup embrio pada telur yang dimikroinjeksi merupakan suatu hal yang umumnya terjadi, dikarenakan tingginya resiko perlakuan mikroinjeksi terhadap kelangsungan hidup embrio (Zbikowska, 2003; Dunham et al., 1987 dalam Ath-thar, 2007). Pada umumnya rendahnya kelangsungan hidup embrio diakibatkan oleh tidak tepatnya jarum mikroinjeksi masuk ke dalam blastodisk, sehingga jarum mikroinjeksi tersebut mengenai yolk sac dan terjadi kerusakan pada embrio. Kerusakan pada embrio dapat mengakibatkan infeksi pada perkembangan organogenesis embrio dan menghambat pertumbuhan embrio serta abnormalitas sehingga embrio tidak mampu menetas.
Pada persentase embrio mengekspresikan transgen (PEMT), kedua promoter menunjukkan nilai yang relatif sama. Hal ini membuktikan bahwa promoter keratin yang diisolasi dari ikan flounder Jepang maupun heatshock diisolasi dari ikan rainbow trout dapat aktif pada ikan nila. Sehingga kedua jenis promoter tidak bersifat spesifik pada suatu jenis ikan tertentu.
Puncak ekspresi gen GFP yang diaktivasi oleh kedua promoter terjadi pada jam-36 setelah injeksi, atau ketika telur ikan nila memasuki akhir pembentukan epiboly. Kondisi ini serupa dengan yang disampaikan Iyengar et al. (1996) bahwa kondisi seperti terjadi karena adanya akumulasi dari DNA eksogenus (DNA yang diinjeksikan) tereplikasi dari fase pembelahan (cleavage) dan akumulasi dari hasil transkripsi pada fase MBT.
Hilangnya ekspresi gen GFP pada jam ke-72 pasca mikroinjeksi, menunjukkan bahwa ekspresi gen GFP yang dipicu oleh promoter keratin maupun heatshock bersifat transient (sementara). Ekspresi sementara ini terkait dengan ketahanan dari DNA yang diinjeksikan ke dalam telur. Ekspresi sementara ini sering terjadi pada beberapa penelitian transgenik, diantaranya pada ikan medaka Oryzias latipes (Winkler et al., 1991; Hamada et al., 1998; Chou et al., 2001), ikan zebra Danio rerio (Higashijima et al., 1997; Meng et al., 1999), ikan lele Clarias gariepinus (Volckaert et al., 1997; Ath-thar, 2007), kakap Sparus auratus
(Garcia-Pozo et al., 1998), dan ikan mas Cyprinus carpio (Purwanti, 2007). Promoter keratin menunjukkan aktivitas yang berbeda dengan yang dilaporkan oleh Yazawa et al. (2005) dalam pengujian pada ikan zebra yang menyebutkan bahwa, aktivitas promoter keratin mampu mendorong GFP terekspresi dari embrio hingga ikan zebra tumbuh dewasa. Hal ini dikarenakan pada penelitian Yazawa et al. (2005) pada ikan zebra yang diteliti sudah F2, sedangkan pada penelitian ikan nila ini masih F0. Aktivitas yang sama dengan ikan zebra mungkin sama terjadi bila ikan nila sudah F2.
cis-acting yang dimiliki oleh promoter keratin lebih dikenali oleh elemen
trans-acting ikan nila, dibandingkan dengan elemen cis-acting yang dimiliki oleh
promoter heatshock. Kurang sesuainya elemen cis-acting pada promoter heatshock dengan trans-acting pada ikan nila, dapat berakibat pada terhambatnya
mekanisme transkripsi sehingga tidak menghasilkan ekspresi yang optimal. Hal tersebut senada dengan penjelasan Iyengar et al. (1996) dan Alimuddin (2003) yang menyebutkan bahwa efektivitas suatu promoter dalam mengendalikan gen untuk terekspresi sangat terkait erat dengan kesesuaian antara elemen cis-acting pada promoter dengan elemen trans-acting pada inang target.
V. KESIMPULAN DAN SARAN 5.1 Kesimpulan
Efektivitas promoter keratin lebih baik dibandingkan dengan promoter heatshock pada ikan nila Oreochromis niloticus. Promoter keratin memiliki pola
aktivitas sementara relatif sama dengan promoter heatshock, namun pada promoter keratin awal aktivitasnya terjadi lebih cepat.
5.2 Saran
DAFTAR PUSTAKA
Alestrom P .2001. Genetically Modified Fish in Future Aquaculture: Technical, Environmental, and Management Considerations. Biotechnology Seminar Paper. International Service National Agricultural Research Netherland.
Alimuddin, Octavera A, Arifin OZ, Sumantadinata K .2008. Karakterisasi
promoter β-actin ikan nila Oreochromis niloticus. Jurnal Akuakultur Indonesia 7(2):115-127.
Alimuddin, Yoshizaki G, Kiron V, Satoh S and Takeuchi T. 2005. Enhancement of EPA and DHA biosynthesis by over-expression of Masu Salmon ∆6 -Desaturase-Like gene in Zebrafish. Transgenic Research, 14 : 159 – 165. Alimuddin. 2003. Introduction and expression of foreign Δ6 desaturase-Like gene
in a Teleostean fish. Thesis. Graduate School of Fisheries Science. Tokyo University of Fisheries.
Arenas MI, Fraile B, De Miguel M, and Paniagua R. 1995. Intermediate filaments in the testis of the teleost mosquito fish (Gambusia affini holbrooki): a light and electron microscope immunocytochemical study and Western blotting analysis. Histochem J 27: 329–337.
Ath-thar MHF. 2007. Efektivitas promoter β-actin ikan medaka Oryzias latipes dengan penanda gen hrGFP (Humanized Renilla reniformis Green Fluorescent protein) pada ikan lele Clarias sp. Keturunan F0. Skripsi. Departemen Budidaya Perairan. Perikanan dan Ilmu Kelautan. Institut Pertanian Bogor.
Bacaj T, and Shaham S. 2007. Temporal control of cell specific transgene expression in C.elegans. Genetics: Published Articles Ahead of Print: 107.074369.
Boonanuntanasarn S, Yoshizaki G, Takeuchi Y, Morita T and Takeuchi T. 2002. Gene knock-down in Rainbow trout embryo using antisense morpholino phosphorodiamidate oligonucleotides. Marine Biotechnology 4:248-257. Chou CY, Horng LS and Tsai HJ. 2001. Uniform GFP-expression in transgenic
Medaka Oryzias latipes at the F0 generation. Transgenic Research 10:303-315.
Conrad M, Lemb K, Schubert T, and Markl J. 1998. Biochemical identification and tissue-specific expression patterns of keratins in the zebrafish (Danio rerio). Cell Tissue Res 293: 195–205.
Cook JT, McNiven MA and Sutterlin AM. 2000. Metabolic rate of Pre-smolt Growth-enhanced transgenic Atlantic Salmon Salmo salar. Aquaculture 188: 33-45.
Devlin RH, Yesaki TY, Donaldson EM, Du SJ and Hew CL. 1995. Production of germline transgenic Pacific Salmonids with dramatically increased growth performance. Canadian Journal of Fisheries Aquatic Sciences, 52 : 1376 – 1384.
Druger RK, Levine EM, Glasgow E, Jones PS, and Schchter N. 1992. Cloning of a type I Keratin from Goldfish optic nerve: Differential Expression of Keratin During Regeneration. Differentiation 52: 33-43.
Dunham RA. 2004. Aquaculture and Fisheries Biotechnology : Genetic Approaches. CABI Publishing. Cambridge, MA, USA.
Effendi MI. 1979. Metode Biologi Perikanan. Yayasan Dewi Sri. Bogor. Effendie I. 1997. Biologi Perikanan. Yayasan Pustaka Nusatama.Yogyakarta. Felts K, Rogers B, Chen K, Ji H, Sorge J and Vaillancourt P. 2001. Recombinant
Renilla reniformis GFP displays low toxicity. Stratagene 13:85-87.
Feng Fang. 2003. Expression of heat shock genes HSP16.6 and HTPG in the cyanobacterium Synechocystis. Disertation. The Graduated School. Miamy University.
Fletcher GL. and Davies PL. 1991. Transgenic fish for aquaculture. Genetic Engineering 13:331-371.
Food and Agriculture Organization (FAO). 2004. The State of World Fisheries and Aquaculture. http://www.fao.org/sof/sofia/index_en.html. Diakses 10
November 2008.
Fujimura K and Okada N. 2007. Development of embryo, larva and early juvenile of Nile Tilapia Oreochromis niloticus (pisces: chiclidae) developmental staging system. Develop Growth Differ 49 : 301-324.
Giordano S, Hall C, Quitschke W, Glasgow E, Schechter N. 1990. Keratin 8 of simple ephitelia is expressed in glia of the Goldfish nervous system. Differentiation 44: 163-172.
Glick BR and Pasternak JJ. 2003. Molecular Biotechnology : Principles and Applications of Recombinant DNA. Third ed. ASM Press. Washington DC.
Gong Z, Ju B, Wang X, He J, Wan H, Sudha PM, and Yan T. 2002. Green fluorescent protein expression in germ line transmitted transgenic zebrafish under a stratified ephitelial promoter from keratin8. Developmental Dynamics 223: 204-215.
Groff JM, Naydan DK, Higgins RJ, Zinkl JG. 1997. Cytokeratin filament expression in epithelial and non-epithelial tissues of the Common Carp (Cyprinus carpio). Cell Tissue Res 287 : 375–384.
Hackett PB. 1993. The Molecular Biology of Transgenic Fish. In: Hocachka and Mommesen (Eds.). Biochemistry and Molecular Biology of Fishes Vol. 2 (pp.218-221). Elsevier Science Publishers BV.
Hamada K, Tamaki K, Sasado T, Watai Y, Kani S, Wakamtsu Y, Ozato K, Kinoshita M, Kohno R, Takagi S and Kimura M. 1998. Usefulness of the
Medaka β-actin promoter investigated using a mutant GFP reporter gene in transgenic Medaka Oryzias latipes. Molecular Marine Biology and Biotechnology 7:173-180.
Higashijima S, Okamoto H, Ueno N, Hotta Y and Eguchi G. 1997. High-frequency generation of transgenic Zebrafish which reliably express GFP in whole muscles or the whole body by using promoter of Zebrafish origin. Developmental Biology 192:289-299.
Hirono I, Aoki T, Shimizu N, and Takashima F.2003. Immunorelated-genes of the Japanese Flounder Paralichthys olivaceus. Aquatic genomics in Hirono I, Aoki T, Shimizu N, and Takashima F (eds) pp : 286-300.
Ian Birch-Martin, Shan G, Huen D, McGirr R, White RAH, and Russel S. 2005. Genomic analysis of Heat-shock factor target in Drosophila. Genome Biology 6: R63.
Iyengar A, Muller F and Maclean N. 1996. Regulation and expression of transgenes fish – A Review. Transgenic Research 5:147-166.
Katagiri T, Kidd C, and Tomasino E. 2005. A BAC-based physical map of Nile Tilapia genome. BMC Genom 6: 89.
Kato K, Takagi M, Tamaru Y, Akiyama S, Konishi T, Murata O, and Kumai H.
region for gene transfer by microinjection in Red Sea Bream Pagrus major. Fisheries Science 73: 440-445.
Kawamoto T, Makino K, Orita S, Nakata A,and Kakunaga T.1989. DNA bending
and binding factors of the human β-actin promoter. Nucleic Acids Research 17(2): 523-537.
Kinoshita M and Ozato K. 1995. Cytoplasmic microinjection of DNA into fertilized Medaka Oryzias latipes eggs. The Fish Biology Journal MEDAKA 7:59-64.
Kobayashi SI, Alimuddin, Morita T, Miwa M, Lu J, Endo M, Takeuchi T and Yoshizaki G. 2007. Transgenic Nile Tilapia Oreochromis niloticus over-expressing growth hormone show reduced ammonia excretion. Aquaculture 270: 427-435.
Kocher TD, Lee WJ, Sobowleska H, Penman D, and McAndrew B.1998. A genetic linkage map of a cychlid fish, the Tilapia (Oreochromis niloticus). Genetics 148 :1225-1232.
Krone PH, and Sass JB. 1994. Hsp90α and hsp90β genes are present in the Zebrafish and are differentially regulated in developing embryos. Biochem. Biophys. Res. Commun 204 : 746–752.
Krone PH, Sass JB, Lele Z. 1997. Heat shock protein gene expression during embryonic development of the zebrafish. Cell Mol Life Sci. 53 :122–129. Lee BY, Penman DJ, and Kocher TD. 2003. Identification of sex determining
region in the Nile Tilapia (Oreochromis niloticus) using bulket segregant analysis. Animal Genetic 34: 379-383.
Lele Z, Engel S, and Krone PH. 1997. Hsp47 and hsp70 genes expression is differentially regulated in a stress and tissue specific manner in Zebrafish embryos. Dev. Genet. 21: 123–133.
Liu Z, Moav B, Faras AJ, Guise KS, Kapuscinski AR and Hackett PB. 1990.
Functional analysis of elements affecting expression of the β-actin gene of Carp. Molecular Cell Biology 10:3432-3440.
Majumdar KC, and McAndrew BC. 1986. Relative DNA content of somatic nuclei and chromosomal studies in Three Generation, Tilapia, Sharoterodon, and Oreochromis of the tribe Tilapiini (Pisces, Cichlidae). Genetica 68: 175-188.
Markl J, Winter S, and Franke WW. 1989. The catalog and the expression complexity of cytokeratins in lower vertebrate: Biochemical identification of cytokeratins in a teleost fish, Rainbow trout. Eur J Cell Biol 50: 1-16. McClean N, Rahman MA, Sohm F.2002. Transgenic Tilapia and the Tilapia
genome. Gene 295: 265-277.
McConnell SK, Beynon C, Leamon L, and Skibinski DO. 2000. Microsatellite marker based genetic linkage of Oreochromis aureus and O. niloticus (Cichlidae): Extensive linkage segment homologies revealed. Animal Genetic 331: 214-218.
Melamed P, Rosenfelt , Elizur A, and Yaron Z. 1998. Endocrin regulation of gonadotrophin and growth hormone gene transcription in fish. Comp Bochem Physiol C Pharmacol Toxicol Endocrinol 119: 325-338.
Meng A, Jessen JR and Lin S. 1999. Transgenesis. Methods in Cell Biology In : Detrich HW III, Westerfield M and Zon LI (eds) Vol. 60 (pp.133-148). Milioni D, Franz G, Sung R, and Hatzopoulos P. 2001.Gene expression during
heat-shock in embryogenic carrot cell lines. Plant Cell, Tissue and Organ Culture 65: 221–228.
Nam YK, Noh JK, Cho YS, Cho HJ, Cho KN, Kim CG and Kim DS. 2001. Dramatically accelerated growth and extraordinary gigantism of transgenic Mud Loach Misgurnus mizolepis. Transgenic Research 10:353-362. Noh JK, Cho KN, Han EH, Kim A, Lee JS, Kim DS, and Kim CG. 2003.
Genomic cloning of Mud Loach Misgurnus mizolepis (Cypriniformes, Cobitidae) Beta-actin gene and usefulness of its promoter region for fish transgenesis. Marine Biotechnology 5 (3): 244-252.
Nugrahani W. 2008. Isolasi Promoter β-Actin Ikan Kerapu Bebek Cromileptes altivelis dengan Metode Degenerate PCR. Skripsi. Departemen Budidaya Perairan. Fakultas Perikanan dan Ilmu Kelautan. Institut Pertanian Bogor. Ojima N. 2007. Rainbow trout hspb1(hsp27): Identification of two mRNA splice
variants that show predominant expression in muscle tissues. Comparative Biochemistry and Physiology part B 148: 277-285.
Oliviera C, and Wright JM. 1998. Molecular cytogenetic analysis of heterochromatin in the chromosome of Tilapia Oreocromis niloticus (Teleostei : Cichlidae). Chromosome Res 6: 205-211.
Santini S and Benardi G. 2005. Organization and base composition of Tilapia Hox genes: Implications for the Evolution of Hox cluster in Fish. Gene 346: 51-61.
Saputra A. 2007. Pertumbuhan Benih Ikan Nila Hasil Sex Reversal, Benih Genetically Male Tilapia dan benih ikan nila YY. Skripsi. Departemen Budidaya Perairan. Perikanan dan Ilmu Kelautan. Institut Pertanian Bogor. Schaffeld M, Lobbecke A, Lieb B, Markl J. 1998. Tracing keratin evolution: catalog, expression patterns and primary structure of Shark (Scyliorhinus stellaris) keratins. Eur J Cell Biol 77: 69 – 80.
Seok SH, Baek MW, Lee HY, Kim DJ, Na YR, Noh KJ, Park SH, Lee HK, Lee BH, Ryu DY, and Park JH. 2007. Quantitative GFP fluorescence as an indicator of arsenite developmental toxicity in mosaic heat shock protein 70 transgenic Zebrafish. Toxicology and Pharmacology 225: 154-161. Takagi S, Sasado G, Tamiya G, Ozato K, Wakamatsu Y, Takeshita A and Kimura
M. 1994. An efficient expression vector for transgenic Medaka construction. Molecular Marine Biology and Biotechnology 3:192-199 Trewavas E. 1983.Tilapiine fishes of the genera Sarotherodon, Oreochromis and
Danakilia. British Mus. Nat. Hist. 583 p.
Volckaert FA, Hellemans BA, Galbusera P and Ollevier F. 1994. Replication, expression and fate of foreign DNA during embryonic and larval development of the African Catfish Clarias gariepinus. Molecular Marine Biology and Biotechnology 3(2):57-69.
Williams DW, Muller F, Lavender FL, Orban L, and Maclean N. 1996. High transgene activity in the yolk syucitial layer affects quantitative transient expression assay in Zebrafish Danio rerio embryos. Transgenic Research 5:433-442.
Winkler, C, Vielkind JR and Schartl M. 1990. Transient expression of foreign DNA during embryonic and larval development of the Medaka fish Oryzias latipes. Mol. Gen. Genet. 226:129 – 140.
Wright PA, and Land MD. 1998. Urea production and transport in teleost fishes. Comp Biochem Physiol. Physiol A: Mol Integr Physiol 119: 47-54.
Wu YL, Pan X, Sudha PM, Wang H, Kee PW, and Gong Z. 2007. Developmental of a heatshock inducible GFP transgenic Zebrafish line by using the Zebrafish hsp27 promoter. Gene 408: 85-94.
Yeh F, and Hsu T. 2002. Differential regulation of spontaneous and heat-induced HSP 70 expression in developing Zebrafish (Danio rerio). J. Exp. Zool. 293 : 349–359.
Yoshizaki, G. 2001. Gene transfer in Salmonidae: Applications to aquaculture. Suisanzoshoku. 49(2):137-142.
Lampiran 1. Embriogenesis ikan nila O.niloticus (Fujimura & Okada, 2007)
Lampiran 2. Pemijahan ikan nila dengan sistem semi buatan
1
2
3
4
1. Identifikasi induk jantan dan betina yang siap memijah (ditandai dengan adanya tonjolan pada daerah lubang urogenitalnya
2. Kedua induk kemudian dicampurkan dengan cara memindahkan induk jantan pada akuarium induk betina
3. Setelah induk terjadi pemijahan secara alami, kemudian dilakukan stripping pada induk betina dan induk jantan
Lampiran 3. Persiapan alat mikroinjeksi
Keterangan :
A. 1 set alat mikroinjeksi B. Injektor
C. Gel plat mikroinjeksi dan telur yang telah disusun guna diinjeksi D. Micromanipulator
A B
Lampiran 4. Konstruksi DNA (Keratin-GFP) dan (Heatshock-GFP)
a. Konstruksi DNA Keratin-GFP (Yazawa et al.,2005)
Lampiran 6. Proses memasukkan DNA kedalam jarum mikroinjeksi (Loading DNA)
1. Larutan DNA diambil dengan menggunakan mikrotip sebanyak ± 5µ l
2. Selanjutnya dengan menggunakan mikrotip, larutan DNA tersebut dimasukkan ke dalam jarum mikroinjeksi.
3. Capillary glass ditempelkan pada needle holder kemudian dimasukkan minyak mineral hingga penuh
4. Dengan bantuan capillary glass, minyak mineral dimasukkan ke dalam jarum mikroinjeksi hingga penuh, dengan memberi sedikit rongga antara larutan DNA dengan minyak mineral.
Lampiran7. Derajat Kelangsungan Hidup Embrio (DKHE) dan Derajat Penetasan (DP) pada ikan nila O.niloticus
1
30
26,67
16,67
2
30
40
26,67
rataan
30
33,33
21,67
simpangan baku
0
9,4
7,1
1
30
16,67
10
2
30
26,67
16,67
rataan
30
21,67
13,33
simpangan baku
0
7,1
4,7
1
30
70
63,33
2
30
86,67
80
rataan
30
78,33
71,67
simpangan baku
0
11,8
11,8
Jumlah telur yang diinjeksi
Derajat Kelangsungan Hidup Embrio
Derajat Penetasan
Keratin
HSC
Kontrol
Lampiran 8. Persentase embrio mengekspresikan transgen (PEMT) dan tingkat ekspresi Gen GFP pada ikan nila O.niloticus
1
30
46,67
2 butir
14,29%
4 butir
28,57%
7 butir
50%
2
30
40,00
1 butir
8,33%
2 butir
16,67%
9 butir
75,00%
rataan
30
43,34
1,5
11,31%
3
22,62%
8
62,50%
simpangan baku
0
4,72
0,71
4,21
1,41
8,42
1,41
17,68
1
30
46,67
5 butir
35,71%
4 butir
28,57%
5 butir
35,71%
2
30
53,33
6 butir
35,50%
5 butir
31,25%
5 butir
31,25%
rataan
30
50,00
5,5
36,62%
4,5
29,92%
5
33,48%
simpangan baku
0
4,71
0,7
1,3
0,7
1,9
0,0
3,2
(1): ekspresi pendar hijau kurang terang; (2): ekspresi pendar hijau terang, dan (3): ekspresi pendar hijau sangat terang.
Tingkat Ekspresi
1
2
3
Jenis Promoter
Ulangan
Keratin
HSC