PENGARUH LAMA KETIADAAN INANG
Spodoptera litura
F.
(LEPIDOPTERA: NOCTUIDAE) TERHADAP POLA
REPRODUKSI PARASITOID
Snellenius manilae
ASHMEAD
(HYMENOPTERA: BRACONIDAE)
MOHAMAD ELDIARY AKBAR
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
PENGARUH KETIADAAN INANG
Spodoptera litura
F.
(LEPIDOPTERA: NOCTUIDAE) TERHADAP POLA
REPRODUKSI PARASITOID
Snellenius manilae
Ashmead
(HYMENOPTERA: BRACONIDAE)
MOHAMAD ELDIARY AKBAR
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Skripsi : Pengaruh Lama Ketiadaan Inang Spodoptera litura F. (Lepidoptera: Noctuidae) Terhadap Pola Reproduksi Parasitoid Snellenius manilae Asmead (Hymenoptera: Braconidae).
Nama Mahasiswa : Mohamad Eldiary Akbar
NRP : A34062988
Disetujui Dosen Pembimbing
Dr. Ir. Damayanti Buchori, M.Sc NIP 19601028 198603 2 002
Diketahui
Ketua Departemen Proteksi Tanaman
Prof. Dr. Ir. Dadang, M.Sc NIP 19640204 199002 1 002
ABSTRAK
MOHAMAD ELDIARY AKBAR. Pengaruh Lama Ketiadaan Inang Spodoptera litura F. (Lepidoptera: Noctuidae) Terhadap Pola Reproduksi Parasitoid
Snellenius manilae Ashmead (Hymenoptera: Braconidae). Dibimbing oleh DAMAYANTI BUCHORI.
Pengendalian hayati merupakan salah satu solusi dalam masalah hama pertanian yang seharusnya lebih banyak dipraktekkan di lapangan karena lebih ramah lingkungan dan mengurangi dampak buruk dari penggunaan pestisida. Salah satu agens hayati yang dapat digunakan dalam pengendalian hayati adalah parasitoid. Imago betina parasitoid berperan penting dalam pencarian dan pemilihan inang yang sesuai untuk melakukan oviposisi sehingga perilaku reproduksi parasitoid merupakan salah satu tumpuan keberhasilan dalam penggunaan teknik pengendalian hayati. Tujuan penelitian ini adalah mengetahui pengaruh lama ketiadaan inang S. litura terhadap perilaku reproduksi parasitoid
S. manilae. Penelitian dilakukan di Laboratorium Bioekologi Predator dan Parasitoid, Departemen Proteksi Tanaman, Fakultas Pertanian IPB. Penelitian ini terdiri atas 14 perlakuan dengan variasi waktu ketiadaan inang selama 1, 2, 3, 4, 5, 6, dan 7 hari. Ketiadaan inang mempengaruhi perilaku reproduksi parasitoid S. manilae. Ketiadaan inang cenderung meningkatkan parasitisasi dan juga lama hidup imago parasitoid. Ketiadaan inang juga cenderung menyebabkan penurunan jumlah telur yang diproduksi parasitoid. Parasitoid yang diberi inang pada awal kemunculan menjadi imago cenderung memproduksi lebih banyak telur daripada yang tidak diberi inang pada awal kemunculan menjadi imago. Parasitoid dapat langsung melakukan oviposisi walaupun tidak diberi inang selama 7 hari berturut-turut.
Penulis dilahirkan di Bandung pada tanggal 18 Oktober 1988 sebagai putra kedua dari empat bersaudara pasangan Ir. Akhmad Subandi dan Triarena Jayanti. Pada tahun 1994 penulis memulai sekolah di SD Negeri Mohamad Toha II dan lulus pada tahun 2000. Penulis melanjutkan ke pendidikan menengah di SLTP Negeri 2 Bandung dan lulus pada tahun 2003, kemudian melanjutkan pendidikan di SMA Negeri 4 Bandung dan lulus tahun 2006. Pada tahun 2006 juga, penulis diterima di IPB melalui jalur Seleksi Penerimaan Mahasiswa Baru (SPMB) dan tahun 2007 diterima di Departemen Proteksi Tanaman, Institut Pertanian Bogor.
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT karena berkat kehadirat-Nya penulis dapat menyelesaikan skripsi yang berjudul ”Pengaruh Lama Ketiadaan Inang Spodoptera litura F (Lepidoptera: Noctuidae) Terhadap Pola Reproduksi Parasitoid Snellenius manilae Ashmead (Hymenoptera: Braconidae)” yang bertujuan memperoleh gelar Sarjana Pertanian pada Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Penulis menyampaikan terima kasih kepada Dr. Ir. Damayanti Buchori, M.Sc yang telah memberikan pengetahuan, pengarahan, dukungan, dan bimbingan sejak awal hingga akhir penelitian. Ucapan terima kasih juga penulis sampaikan kepada Dr. Ir. Kikin H Mutaqin, M.Si selaku dosen penguji tamu yang telah memberikan dukungan atas penulisan skripsi ini. Ucapan terima kasih juga penulis sampaikan kepada Ir. Titiek S. Yuliani, SU selaku dosen pembimbing akademik yang telah memberikan bimbingan, pengetahuan, dan dukungan sejak awal penulis masuk Departemen Proteksi Tanaman IPB. Tak lupa penulis mengucapkan terima kasih kepada seluruh staf pengajar dan laboran Departemen Proteksi Tanaman.
Terima kasih untuk Ibunda Triarena Jayanti dan Ayahanda Ir. Akhmad Subandi tercinta serta ketiga saudara yang penulis sayangi, Kory Pranita A, S.Si Dona Astari N, dan Ahmad Riky A. Ucapan terima kasih juga ditujukan kepada rekan-rekan di Laboratorium Bioekologi Parasitoid dan Predator dan teman-teman baik di dalam maupun di luar Departemen Proteksi Tanaman (Adiyat Yori Rambe, Agus F Tambun, Agus Setiawan, Ahmad Faishol, Aliet Seignoritha, Asmawati Ahmad, Ari Widiastuti, Atrie Yuni Sonia, Dede Tiara, Dedek Kusvianti, Dolpina A Ratissa, Edwina Triwibowo, Ellyta Sariani, Fitra Murgianto, Gamatriani M, Iin Maena, Indri Ahdiaty, Isnendi A, Lia Nazirah, Lutfi Afifah, Meike Isna Rahmawati, M. Julyanda, Sarah Nila, Sari Nurulita, Radhian Ardi Prabowo, Rio Ardian, Sarifa Yahtum Latuconsina, Sofyan Zuhri, Sunarti Tambunan, Tuti Legiastuti, Vani Nur Oktaviani S, Veronica T Simanjuntak, Yulius Dika C, Zalzilatul Hikmia) yang telah memberikan persahabatan dan dukungan selama ini.
Penulis menyadari banyak kekurangan dalam penulisan skripsi ini. Oleh karena itu, saran dan kritik yang membangun sangat diharapkan demi penulisan yang lebih baik. Penulis berharap skripsi ini dapat bermanfaat dan memberikan pengetahuan bagi siapa saja yang membacanya.
Bogor, Desember 2011
DAFTAR ISI
Halaman
DAFTAR TABEL ... ix
DAFTAR GAMBAR ... x
DAFTAR LAMPIRAN ... xi
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 3
Manfaat... 3
TINJAUAN PUSTAKA... 4
Pengendalian Hayati ... 4
Parasitoid ... 6
Perilaku Reproduksi Parasitoid... 7
Ketiadaan Inang ... 8
Superparasitisme ... 10
Spodoptera litura (Fabricius) ... 11
Snellenius manilae ... 12
BAHAN DAN METODE ... 15
Waktu dan Tempat ... 15
Metode Penelitian ... 15
Perbanyakan S. litura ... 15
Perbanyakan Parasitoid S. manilae ... 16
Pengamatan ketiadaan inang S. litura terhadap produksi telur parasitoid S. manilae. ... 17
Analisis Data ... 20
HASIL DAN PEMBAHASAN ... 21
Hasil Penelitian ... 21
Pengaruh Ketiadaan Inang Terhadap Oviposisi di Hari Pertama Setelah Perlakuan ... 21
Pengaruh Ketiadaan Inang Terhadap Kemampuan Parasitisasi S. manilae di hari ke-8 ... 22
Pengaruh Ketiadaan Inang Terhadap Produksi Telur S. manilae ... 23
Pengaruh Ketiadaan Inang Terhadap Sisa Telur dalam Ovari S. manilae .... 24
Pengaruh Ketiadaan Inang Terhadap Lama Hidup Imago S. manilae ... 26
Pengaruh Ketiadaan Inang Terhadap Superparasitisasi S. manilae ... 27
Pembahasan ... 30
Pengaruh Ketiadaan Inang Terhadap Perilaku Oviposisi S. manilae ... 30
Pengaruh Ketiadaan Inang Terhadap Produksi Telur dan Lama Hidup S. manilae ... 32
Pengaruh Ketiadaan Inang Terhadap Superparasitisasi S. manilae ... 35
KESIMPULAN DAN SARAN ... 38
Kesimpulan ... 38
Saran ... 38
DAFTAR PUSTAKA ... 39
DAFTAR TABEL
DAFTAR GAMBAR
Halaman
1. Waktu perlakuan ketiadaan inang. ... 18
2. Jumlah telur yang diletakkan oleh parasitoid S. manilae pada hari petama setelah ketiadaan inang S. litura... 21
3. Tingkat parasitisasi S. manilae pada hari ke-8. ... 23
4. Jumlah telur parasitoid S. manilae pada hari ke-8 dan hingga hari ke-8. ... 24
5. Rata-rata sisa telur S. manilae di dalam ovari. ... 25
6. Rata-rata jumlah total telur parasitoid S. manilae. ... 26
DAFTAR LAMPIRAN
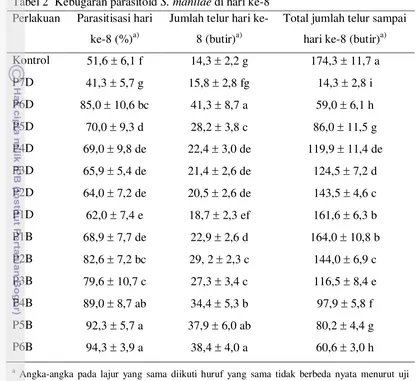
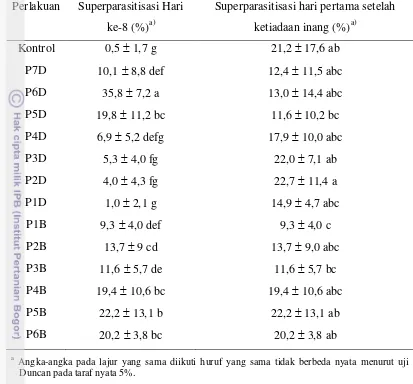
Halaman 1. Kebugaran parasitoid S. manilae di hari ke-8... 42 2. Total kebugaran parasitoid S. manilae. ... 43 3. Tingkat superparasitisasi S. manilae di hari ke-8 dan hari pertama setelah
PENDAHULUAN
Latar Belakang
Parasitoid merupakan salah satu jenis musuh alami yang dapat digunakan untuk pengendalian hayati. Menurut Godfray (1994), larva parasitoid makan di dalam tubuh arthropoda lain yang menjadi inang sehingga menyebabkan kematian inangnya. Parasitoid hanya memerlukan satu inang untuk menyelesaikan perkembangannnya dan sering kali sejumlah parasitoid berkembang secara berkelompok dalam satu inang yang sama. Elzinga (2004) menyebutkan beberapa kelebihan penggunaan parasitoid, yaitu agens hayati ini biasanya sangat selektif, resistensi serangga lebih sedikit terjadi dibandingkan pada penggunaan pestisida, pengaruh terhadap ekosistem lebih sedikit dan parasitoid lebih tidak berbahaya pada manusia dibandingkan penggunaan pestisida.
Parasitoid Snellenius (=Microplitis) manilae Ashmead (Hymenoptera: Braconidae) merupakan salah satu endoparasitoid larva S. litura (Xu & Yang 1983 dalam Waterhouse & Norris 1987). Parasitoid S. manilae ditemukan memarasit larva S. litura pada larva instar-instar awal sehingga kematian larva S. litura terjadi lebih dini. Hal tersebut menguntungkan karena dapat mencegah terjadinya kerugian yang lebih besar. Prabowo (1994) menyebutkan S. manilae
banyak ditemukan pada populasi S. litura yang menyerang tanaman talas di daerah Tajur dan Sindangbarang Bogor. Ulat grayak (S. litura Fabr.) merupakan hama penting yang sering dijumpai pada berbagai tanaman budidaya di Indonesia. Hama ini bersifat polifag yang inangnya tidak terbatas hanya pada tanaman budidaya saja, tetapi juga pada tanaman liar. S. litura menyerang bagian daun pada tanaman sehingga dapat mengganggu proses fotosintesis pada tanaman yang berakibat pada penurunan hasil produksi. Kalshoven (1981) menyebutkan tanaman inang S. litura di antaranya tembakau, kedelai, jagung, ubi jalar, kakao, kacang tanah, jarak, pisang, bayam, gandum, kangkung, bawang merah, babadotan, dan tanaman hias. Status sebagai hama utama pada beberapa tanaman budidaya menjadikan pengendalian hama ini sangat penting. Hal tersebut dilakukan untuk menghindari kerusakan tanaman demi tercapainya peningkatan hasil produksi pertanian.
3
(Hidayatullah 2000) dan efektifitas parasitisasi (Ratna 2008). Penelitian mengenai aspek biologi reproduksi dari parasitoid betina penting karena parasitoid betina berperan penting dalam kegiatan parasitisasi inang. Salah satu tumpuan keberhasilan pengendalian hayati menggunakan parasitoid sangat tergantung pada perilaku reproduksi imago betina parasitoid. Hal ini disebabkan imago parasitoid betina memliki peran yang penting dalam mencari dan memilih inang yang sesuai baginya untuk melakukan oviposisi. Selain itu imago parasitoid betina juga berperan dalam keberlangsungan populasi parasitoid karena imago betina yang menentukan inang yang sesuai dan baik bagi keturunannya untuk tumbuh dan berkembang dengan baik. Hal ini menunjukkan bahwa berbagai aspek yang dapat mempengaruhi biologi dan ekologi parasitoid perlu dipelajari dengan lebih seksama.
Terkadang selama beberapa waktu inang tidak tersedia bagi parasitoid. Berdasarkan beberapa studi awal, ketidaktersediaan inang tersebut dapat mempengaruhi perilaku peletakan telur parasitoid. Heriyano (2000) dalam penelitiannya menyebutkan bahwa pemuasaan parasitoid Eriborus argenteopilsus
(Hymenoptera: Ichneumonidae) tidak mempengaruhi kemampuan untuk memarasit, namun lama waktu parasitoid tidak mendapatkan inang ternyata dapat mempengaruhi kemampuan reproduksi parasitoid. Hal tersebut dapat berpengaruh terhadap keberhasilan pengendalian hayati ketika diterapkan dilapangan.
Tujuan
Penelitian ini bertujuan untuk mengetahui pengaruh ketiadaan inang S. litura terhadap tanggap reproduksi S. manilae.
Manfaat
TINJAUAN PUSTAKA
Pengendalian Hayati
Menurut Huffaker & Messenger (1976), istilah pengendalian hayati pertama kali digunakan oleh Smith (1919) untuk menunjukkan pentingnya penggunaan musuh-musuh alami untuk mengontrol hama serangga. Pengendalian hayati melibatkan penggunaan populasi musuh alami untuk menekan populasi hama hingga kepadatan yang lebih rendah, baik secara permanen maupun secara sementara (Driesche et al. 2008). Sedangkan Norris et al. (2003) mendefinisikan pengendalian hayati sebagai penggunaan parasitoid, predator, pathogen, antagonis atau populasi kompetitor untuk menekan populasi hama, membuat hama menjadi lebih sedikit kelimpahannya dan lebih sedikit merusak daripada seharusnya bila agens hayati tidak ada.
Ruang lingkup pengendalian hayati telah meluas dari pemanfaatan serangga entomophagus untuk mengendalikan hama-hama serangga ke arah pemanfaatan keseluruhan rangkaian organism untuk menendalikan serangga-serangga, tungau, keong, vertebrata, dan tanaman. Organisme-organisme yang telah digunakan untuk pengendalian hayati meliputi virus, bakteri, fungi, nematoda, keong, serangga, tungau dan berbagai jenis vertebrata (Huffaker & Messenger 1976).
5
yang lainnya. Kelebihan keenam adalah teknik pengendalian hayati secara tradisional telah diakui tidak mengganggu ekosistem. Pada beberapa hal, bagaimanapun pelepasan musuh alami di daerah yang baru mungkin terdapat pengaruh-pengaruh yang tak diharapkan pada non-target.
Menurut Waterhouse (1993), terdapat empat cara utama bagaimana pengendalian hayati diterapkan, yaitu pertama adalah memasukkan musuh alami dari luar ke negara yang baru, kedua adalah memelihara secara massal musuh alami dan melepaskannya, merupakan pendekatan yang disebut sebagai metode inundasi. Metode ketiga adalah memelihara secara masal musuh alami yang telah ada tetapi tidak efektif pada saat tersebut karena jumlahnya yang rendah, pendekatan ini disebut juga metode augmentasi. Keempat adalah metode manajemen.
Studi dan penelitian mengenai pengendalian hayati di Indonesia telah banyak dilakukan oleh para ahli terutama di tingkat laboratorium dan rumah kaca. Pada tingkat penerapan di lapangan, baru sebagian kecil saja yang telah dimanfaatkan. Terdapat beberapa hal yang sering menjadi kendala dalam penerapan teknik pengendalian hayati, antara lain adalah perlunya berbagai penelitian mendasar untuk mengetahui peranan agens hayati secara pasti, diperlukan fasilitas-fasilitas untuk mendukung penelitian mulai dari eksplorasi hingga sumberdaya manusia, serta petani sudah terbiasa dengan cara pengendalian yang memberikan hasil cepat sehingga tidak tertarik dengan cara pengendalian hayati yang baru memberikan hasil dalam jangka waktu yang relatif lama (Abdurrahman 2011).
Parasitoid
Menurut Godfray (1994), larva parasitoid makan di dalam tubuh arthropoda lain yang menjadi inang sehingga menyebabkan kematian inangnya. Parasitoid hanya memerlukan satu inang untuk menyelesaikan perkembangannnya dan sering kali sejumlah parasitoid berkembang secara berkelompok dalam satu inang yang sama. Semua stadia hidup serangga dapat diparasitisasi (Drieasche et al. 2008). Menurut Godfray (1994), kelebihan parasitoid dibanding musuh alami yang lain adalah parasitoid mampu mengendalikan hama secara spesifik, populasinya di lapang relatif cukup tinggi dan mampu menekan populasi serangga hama secara signifikan. Elzinga (2004) juga menyebutkan beberapa kelebihan penggunaan parasitoid, yaitu agens hayati ini biasanya sangat selektif, resistensi serangga lebih sedikit terjadi dibandingkan pada penggunaan pestisida, pengaruh terhadap ekosistem lebih sedikit dan parasitoid lebih tidak berbahaya pada manusia dibandingkan penggunaan pestisida.
Berdasarkan tempat perkembangan larvanya, parasitoid dibagi menjadi endoparasitoid dan ektoparasitoid. Driesche et al. (2008) menjelaskan bahwa parasitoid yang larvanya berkembang di dalam inangnya disebut endoparasitoid, sedangkan yang berkembang di luar tubuh inangnya disebut ektoparasitoid. Parasitoid terbagi menjadi dua kelompok, yaitu koinobiont dan idiobiont. Parasitoid yang inangnya tidak berkembang lebih jauh setelah terparasit termasuk ke dalam kelompok idiobiont, sedangkan parasitoid yang inangnya tetap melanjutkan perkembangan paling tidak selama beberapa waktu setelah terparasit termasuk kelompok koinobiont. S. manilae termasuk ke dalam kelompok koinobiont karena larva inang tetap melanjutkan perkembangannya selama beberapa waktu setelah terparasit hingga larva parasitoid keluar untuk berpupa. Kelompok koinobiont biasanya menyerang larva, lebih sering instar-instar awal (Quicke 1997).
7
Berdasarkan proses pematangan telur, parasitoid terbagi menjadi dua kelompok yaitu synovigenic dan pro-ovigenic. Menurut Quicke (1997), serangga
synovigenic melakukan pematangan telur selama sebagian atau seluruh masa hidup dewasanya, sedangkan serangga pro-ovigenic muncul menjadi dewasa dengan telur yang sudah matang. Driesche et al. (2008) juga menjelaskan bahwa spesies yang muncul menjadi imago dengan suplai telur seumur hidup tersedia, memungkinkan serangan yang cepat pada banyak inang. Sebaliknya, telur spesies
synovigenic berkembang secara teratur selama masa hidup imago betina. Parasitoid-parasitoid synovigenic membutuhkan protein untuk mematangkan telur. Beberapa parasitoid synovigenic memakan nectar atau embun madu, tetapi ada beberapa yang memakan haemolimph inang. Namun Quicke (1997) juga menyatakan diantara kedua kelompok tersebut terdapat kelompok yang disebut
pro-synovigenic yaitu serangga yang muncul menjadi dewasa dengan sejumlah telur yang sudah matang tetapi dapat melakukan pematangan telur lebih banyak lagi saat pasokan asli telur mereka habis.
Perilaku Reproduksi Parasitoid
Prabowo (1996) menyatakan masa pra oviposisi imago betina S. manilae
cukup singkat, yaitu kurang dari satu hari. Masa pra oviposisi adalah waktu sejak imago keluar hingga saat pertama kali meletakkan telur. Imago betina dapat meletakkan telur dan mempunyai keturunan sejak hari pertama pemunculannya. Selain itu imago jantan dan betina yang keluar pada waktu yang bersamaan dapat segera melakukan kopulasi.
Kebanyakan parasitoid melakukan pematangan telur selama masa hidup dewasanya atau disebut synovigenic (Godfray 1994). Flanders (1950 dalam
Quicke 1997) menyebutkan bahwa ada hubungan yang kuat antara menjadi
synovigenic dan oosorption. Oosorption adalah penyerapan kembali telur yang telah matang. Hal ini diketahui terjadi paling tidak pada beberapa anggota dari tujuh ordo, yaitu Thysanura, Dermaptera, Orthoptera, Heteroptera, Diptera, Coleoptera, dan Hymenoptera (Bell & Bohm 1975 dalam Quicke 1997).
telur akan meningkat pada permulaan kondisi ketiadaan inang kemudian menurun setelahnya akibat terjadinya penyerapan kembali telur (Drost & Carde 1992).
Ketiadaan Inang
Populasi inang di alam tidak menentu, banyak faktor yang mempengaruhi sehingga populasinya berfluktuasi. Ketersediaan inang dapat mempengaruhi produksi telur parasitoid. Penyerapan kembali telur adalah mekanisme yang memungkinkan imago betina untuk mendaur ulang nutrisi yang tersimpan di dalam telur-telur mereka untuk pemeliharaan somatik pada saat terjadi stress ketika makanan atau inang berkurang di alam (Jervis & Kidd 1986 dalam
Hougardy et al. 2005) atau untuk memelihara pasokan telur matang yang baru secara konstan hingga imago betina dapat melanjutkan oviposisi ketika inang menjadi sedikit (Rivero-Lynch & Godfray 1997 dalam Hougardy et al. 2005). Menurut Hougardy et al. (2005), pemberian inang pada imago betina sebelum pelepasan di lapangan dalam beberapa kasus memberi dampak positif terhadap potensi reproduksi di masa depan karena dapat merangsang oogenesis dan kemampuan menemukan inang. Namun pemberian inang segera setelah imago muncul, walaupun hanya untuk beberapa hari, dapat menurunkan kapasitas reproduksi yang tersisa secara dramatis. Dalam kondisi lapangan, Mastrus ridibundus (Hymenoptera: Ichneumonidae) mungkin secara teratur memiliki pengalaman-pengalaman bahwa ketiadaan inang sementara memberikan imago betina waktu untuk mengisi kembali ovariolnya dengan telur-telur matang yang baru dan hal ini dapat menjelaskan terjadinya tingkat serangan terhadap inang yang rendah. Namun Hougardy et al. (2005) juga dalam penelitiannya menyatakan bahwa tidak ditemukan adanya penyerapan kembali telur pada imago betina M. ridibundus yang tidak diberi inang.
9
relatif lama. Selain itu Hougardy et al. (2005) dalam penelitiannya menyebutkan bahwa imago betina M. ridibundus yang tidak diberi inang Cydia pomonella L. (Lepidoptera: Tortricidae) memiliki sediaan telur yang yang lebih banyak dibandingkan imago betina yang terus menerus diberi inang. Imago betina yang tidak diberi inang dapat segera menggunakan telur-telur yang ada di dalam ovariolnya, bahkan setelah 6 hari tidak diberi inang. Oviposisi mencapai puncaknya pada hari pertama inang tersedia, dengan jumlah telur yang diletakkan meningkat seiring dengan tingkat ketiadaan inang. Juga diluar hari pertama reproduksi, tingkat oviposisi harian dan serangan terhadap inang menurun sebagai akibat dari ketiadaan inang dan secara keseluruhan, imago betina tersebut memiliki keperidian seumur hidup yang lebih rendah dan menyerang lebih sedikit inang daripada imago betina yang secara berkelanjutan diberi inang. Peningkatan lama ketiadaan inang tidak mempengaruhi keberhasilan M. ridibundus dalam pencarian inang walaupun tidak mendapat inang hingga 9 hari (Hougardy & Mills 2007).
Hegazi et al. (2007) dalam penelitiannya pada Microplitis rufiventris
(Hymenoptera: Braconidae) menyebutkan bahwa pola reproduksi telur pada M. rufiventris bergantung pada ketersediaan inang untuk melakukan parasitisasi. Telur-telur yang matang akan diakumulasi dalam oviduct ketika inang tidak tersedia, pematangan telur berlanjut hingga daya dukung dukung oviduct tercapai. Ketiadaan inang juga mengubah fisiologi internal imago betina dan menyebabkan terjadinya modifikasi produksi telur dan lama hidup imago.Salah satu akibat dari ketiadaan inang adalah penipisan jumlah telur dalam oviduct yang sering diinterpretasikan sebagai bukti terjadinya oosorption (Labeyrie 1959a, 1960; King 1963 dalam Hegazi et al. 2007).
Studi mengenai hubungan antara ketiadaan inang dengan penyerapan kembali telur juga telah dilakukan oleh Carbone et al. (2008) pada parasitoid telur
yang diberi inang. Lama hidup A. nitens meningkat secara signifikan pada perlakuan ketiadaan inang dengan pemberian makanan, hal ini dapat disebabkan tidak adanya energi yang digunakan untuk reproduksi, ketersediaan makanan dan adanya penyerapan kembali telur. Hal yang sebaliknya terjadi pada parasitoid
Venturia canescens (Hymenoptera: Ichneumonidae) dalam penelitian yang dilakukan oleh Eliopoulos et al. (2005). Imago yang diberi makanan namun tidak diberi inang tidak secara signifikan hidup lebih lama dibandingkan imago yang diberi makanan dan inang.
Superparasitisme
Godfray (1994) menjelaskan bahwa superparasitisme adalah peletakkan telur pada inang yang telah terparasit oleh spesies parasitoid yang sama. Collins & Dixon (1986 dalam Hougardy 2005) juga menyebutkan bahwa ketiadaan inang berkepanjangan dapat meningkatkan superparasitisme. Ratna (2008) menyatakan bahwa efisiensi parasitisasi bagi parasitoid soliter seperti S. manilae dapat ditunjukkan melalui data tingkat parasitisasi optimal yaitu keberadaan larva terparasit oleh hanya satu telur. Sebaliknya superparasitisme merupakan pengurangan tingkat efisiensi pemarasitan. Kejadian superparasitisasi ini mengakibatkan ketidakefektifan parasitisasi karena dapat mengurangi tingkat keberhasilan hidup parasitoid yang berkembang dalam tubuh inang. Di dalam tubuh inang akan terjadi kompetisi nutrisi antar larva parasitoid yang berhasil menetas dari telur, menyebabkan kedua larva mati atau satu individu dapat berkembang dengan pertumbuhan tidak optimal sehingga terjadi penurunan kebugaran parasitoid yang berhasil menjadi imago (Godfray 1994).
Studi mengenai superparasitisme telah banyak dilakukan. Fenlon et al. (2009) dalam penelitiannya terhadap parasitoid Aphidius colemani (Hymenoptera: Braconidae) menyebutkan bahwa tingkat superparasitisme menurun seiring dengan meningkatnya kepadatan populasi inang. Hal penting yang menentukan terjadinya superparasitisme adalah kemampuan parasitoid untuk mendiskriminasi inang. Mehrnejad & Copland (2007) dalam studinya mengenai endoparasitoid
11
mungkin disebabkan oleh pengaruh penanda internal maupun eksternal yang ditinggalkan oleh imago betina yang melakukan oviposisi sebelumnya. Selain itu juga sekresi dari kelenjar Dufour berperan penting sebagai bahan kimia penanda inang yang telah diparasit. Superparasitisme juga menyebabkan peningkatan kematian inang oleh karena itu dalam pengendalian hayati, diharapkan parasitoid yang efektif memiliki kemampuan untuk menemukan inang dengan cepat dan mampu mendiskriminasi inang (Mehrnejad & Copland 2007).
Spodoptera litura (Fabricius)
Spodoptera litura merupakan serangga hama tanaman yang bersifat polifag. Kisaran inangnya sangat luas mencakup tanaman budidaya maupun tanaman non-budidaya. Menurut Kalshoven (1981), tanaman inang S. litura di Indonesia diantaranya tembakau, kentang, kedelai, jagung, ubi jalar, kakao, kacang tanah, jarak, pisang, bayam, gandum, kangkung, bawang merah, babadotan, dan tanaman hias. Hill (1983 dalam Waterhouse & Norris 1987) menyebutkan S. litura merupakan hama utama pada tanaman kapas, padi, tomat, dan tembakau. Selain itu juga menyerang jeruk, kakao, karet, legume, sorgum, jagung, dan jarak.
S. litura sering juga disebut dengan nama ulat grayak. Serangga ini termasuk ke dalam Ordo Lepidoptera, Famili Noctuidae. S. litura menjadi hama pada saat memasuki stadia larva. S.litura mungkin merupakan hewan asli India dan Asia Tenggara (Waterhouse & Norris 1987). Serangga ini tersebar di daerah Asia dan Kepulauan Pasifik. Waterhouse & Norris (1987) menyebutkan S. litura
tersebar di Korea, Jepang, Filipina, Indonesia, Australia, Papua Nugini, Selandia Baru, Hawaii serta negara-negara kepulauan pasifik seperti Fiji, Vanuatu, Toga, Guam, Kepulauan Solomon, dan Kaledonia Baru.
malam hari dalam kelompok yang terdiri dari 200 sampai 300 butir (Waterhouse & Norris 1987). Lama stadium telur adalah tiga hari (Tampenawas 1981). Menurut Kalshoven (1981) total telur yang dapat diproduksi oleh imago S. litura
adalah 2000-3000 telur.
Kalshoven (1981) menyebutkan larva menetas setelah tiga sampai lima hari dan hidup secara gregarius. Setelah beberapa hari larva-larva tersebut akan menyebar. Menurut Tampenawas (1981), stadia larva terdiri dari enam instar. Larva instar satu mempunyai kepala hitam dan tubuh berwarna hijau kekuningan. Larva instar dua mempunyai kepala berwarna cokelat muda, warna tubuh mula-mula hijau kekuningan kemudian menjadi hijau cokelat. Larva instar tiga variasi warnanya lebih jelas. Warna dasar hijau cokelat dengan garis-garis putih dan cokelat sepanjang tubunya. Larva instar empat mempunyai warna dasar abu-abu dan pada bagian dorsal terdapat tiga garis kning memanjang. Larva instar lima berwarna hitam, garis kuning pada bagian dorsal berubah menjadi agak jingga. Larva instar enam mempunyai bentuk yang sama dengan instar lima hanya ukuran tubuh yang menjadi lebih besar. Pada saat stadia larva, S. litura memakan bagian daun tumbuhan inangnya dan merupakan hama utama beberapa tanaman budidaya.
Kalshoven (1981) menyebutkan stadia pupa terjadi di dalam tanah pada sel sederhana yang terbuat dari tanah. Menurut Tampenawas (1981), larva yang akan memasuki masa pra pupa ukuran tubuh mengecil dan berusaha mencari tempat persembunyian. Pupa berwarna cokelat. Stadia pupa terjadi selama 8 sampai 9 hari. Imago S. litura adalah ngengat yang termasuk serangga nokturnal, yaitu hanya aktif pada malam hari sedangkan pada siang hari bersembunyi. Menurut Tampenawas (1981), tubuh imago berwarna cokelat muda dengan panjang tubuh berkisar antara 15-20 mm. Lama siklus hidup S. litura adalah 27 hari. Imago betina kawin beberapa kali (Kalshoven 1981).
Snellenius manilae
13
entomophagus, baik dalam jumlah spesies yang memiliki perilaku entomophagus, juga dalam frekuensi dan efektivitas dimana mereka menyerang serangga hama tanaman pertanian. Quicke (1997) menyebutkan bahwa delapan puluh persen spesies parasitoid adalah anggota ordo Hymenoptera. Famili yang paling penting dan paling sering ditemui memiliki perilaku sebagai parasit adalah Ichneumonidae dan Braconidae, yang menyerang berbagai macam ulat, larva dan imago kumbang, dan lain-lain (Clausen 1997).
Beberapa spesies dari famili Braconidae memiliki nilai yang cukup besar dalam pengendalian hayati serangga-serangga hama. Driesche et al. (2008) menyebutkan bahwa family Braconidae telah digunakan secara luas dalam pengendalian hayati, terutama untuk melawan aphid, Lepidoptera, Coleoptera dan Diptera. Menurut Clausen (1940), perilaku hiperparasitisme lebih sedikit berkembang pada famili Braconidae daripada famili Ichneumonidae dan hampir seluruh famili Braconidae menguntungkan. Parasitisme oleh famili Braconidae dapat terjadi secara internal ataupun eksternal dan modifkasi perilaku berhubungan dengan perilaku stadia inang yang menjadi sasaran serangan. Sejumlah besar spesies dari famili Braconidae dapat melakukan oviposisi langsung pada hari kemunculan imago betina dari pupa.
Menurut Clausen (1940), pemilihan inang subfamili Microgasterinae terbatas terutama pada larva ordo Lepidoptera. Genus yang dominan adalah
Apanteles, Microgaster dan Microplitis. Genus-genus tersebut sangat umum dijumpai. Menurut Prabowo (1994), parasitoid ini merupakan endoparasitoid larva
Spodoptera litura yang bersifat koinobiont. Koinobiont adalah parasitoid yang inangnya tetap hidup setelah terparasitisasi dan selama perkembangan parasitoid (Quicke 1997).
Telur dan larva S. manilae berkembang di dalam tubuh larva S. litura
selama dua hari. Inang yang terparasit akan mati satu sampai dua hari setelah parasitoid keluar dari inang (Prabowo 1996).
Stadia larva terjadi di dalam tubuh inang hingga inang siap untuk berpupa. Larva berkembang di dalam haemosel inang. Menurut Prabowo (1996), stadia larva terdiri dari empat instar. Larva instar I ditemukan pada 48 jam setelah peletakkan telur, sedangkan larva instar IV ditemukan pada 168 jam setelah peletakkan telur. Stadia pupa terjadi di luar tubuh inang. Larva parasitoid akan keluar dari tubuh inang ketika telah siap untuk berpupa. Stadia pra pupa parasitoid
S. manilae terdiri dari dua tahap, yaitu eonymph dan pronymph. Pupa parasitoid bertipe eksarata dengan embelan tubuh bebas dan tampak jelas. Pupa terbentuk dalam kokon yang berwarna cokelat (Prabowo 1996).
Lama hidup imago S. manilae yang diberi makan madu mampu hidup selama 9 hari, sedang yang diberi air hanya mampu hidup selama 4 sampai 5 hari. Imago S. manilae sulit dibedakan karena sangat mirip. Imago jantan dan betina dapat dibedakan secara langsung dengan melihat bagian metasoma dan panjang antena. Metasoma imago jantan dari arah ventral terlihat lebih langsing daripada imago betina dan pangkal metasoma betina relatif lebih terang daripada imago jantan. Pada imago jantan antena terlihat lebih panjang daripada panjang badannya daripada imago betina. Rata-rata panjang ovipositor pada imago betina adalah 0,15 mm (Prabowo 1996).
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilakukan di Laboratorium Bioekologi Parasitoid dan Musuh Alami, Departemen Proteksi Tanaman, Fakultas Pertanian IPB. Pelaksanaan penelitian dimulai pada bulan Januari – Juni 2011.
Bahan dan Alat
Bahan yang digunakan pada penelitian ini adalah serangga inang
Spodoptera litura, parasitoid larva Snellenius manilae, daun talas yang digunakan untuk pakan larva S. litura, madu 20% untuk pakan imago S. litura dan S. manilae, dan daun kedelai yang digunakan untuk meletakkan larva S. litura saat perlakuan.
Alat-alat yang digunakan pada penelitian ini adalah kotak plastik berukuran 35 cm x 5 cm x 7 cm yang tutupnya telah dilubangi dan ditutup kain kasa agar sirkulasi udara lancar, tabung reaksi, cawan petri, kapas, pinset, serbuk gergaji, kertas tisu, kertas buram, dan wadah plastik berbentuk silinder sebagai tempat pemeliharaan imago S. litura. Untuk pembedahan digunakan mikroskop stereo dan jarum bertangkai.
Metode Penelitian
Perbanyakan S. litura
Larva S. litura diambil dari lahan pertanaman talas di daerah Situ Gede dan Kemang, Bogor. Larva yang diperoleh selanjutnya dipelihara untuk perbanyakan di laboratorium. Larva S. litura dipelihara dalam kotak plastik berukuran 35 cm x 25 cm x 7 cm. Selama kegiatan perbanyakan, larva S. litura
dikumpulkan ke dalam wadah plastik dengan jumlah pupa dibatasi 20 pupa per wadah.
Imago S. litura yang telah keluar dari pupa dipindahkan ke dalam wadah plastik silinder. Bagian dalam wadah plastik tersebut diberi lapisan kertas buram sebagai tempat untuk meletakkan telur bagi imago S. litura sehingga telur yang telah diletakkan lebih mudah dikumpulkan. Imago S. litura diberi makanan berupa larutan madu 20% yang diresapkan pada bantalan kapas dan diletakkan pada dasar wadah.
Telur S. litura dikumpulkan kemudian diletakkan pada gelas plastik bertutup dengan jumlah kelompok telur tidak lebih dari 3 kelompok pada setiap gelas plastik. Larva yang menetas dari telur kemudian diberi pakan berupa daun talas yang masih muda.
Perbanyakan Parasitoid S. manilae
Larva S. litura instar II dan III diambil dari lahan pertanaman talas dengan harapan telah terparasit oleh S. manillae secara alamiah. Larva yang diperoleh kemudian dipelihara di laboratorium. Larva dimasukkan ke dalam kotak plastik berukuran 35 cm x 25 cm x 7 cm, lalu ditunggu hingga larva parasitoid S. manilae
keluar dari tubuh larva S. litura untuk berpupa. Larva S. litura yang tidak terparasit di pisahkan untuk kemudian dipelihara. Pupa S. manilae dikumpulkan lalu dimasukkan ke dalam tabung reaksi yang telah diolesi madu yang diencerkan sampai 20% sebagai pakan lalu tabung ditutup dengan tisu. Setiap tabung reaksi berisi lima pupa S. manilae. Hal ini bertujuan untuk mempermudah pengamatan jenis kelamin imago S. manilae.
Perbanyakan S. manilae selanjutnya dilakukan dengan menggunakan larva
S. litura yang dihasilkan dari perbanyakan di laboratorium. Larva S. litura instar II diletakkan pada potongan daun talas kemudian dimasukkan ke dalam tabung reaksi. Setiap tabung berisi 30 ekor larva S. litura. Seekor imago betina S. manilae
dimasukkan ke dalam tabung yang berisi larva instar II S. litura. Larva S. litura
17
yang tidak terparasit dipisahkan untuk kemudian dimanfaatkan untuk perbanyakan inang S. litura.
Pengamatan ketiadaan inang S. litura terhadap produksi telur parasitoid S. manilae.
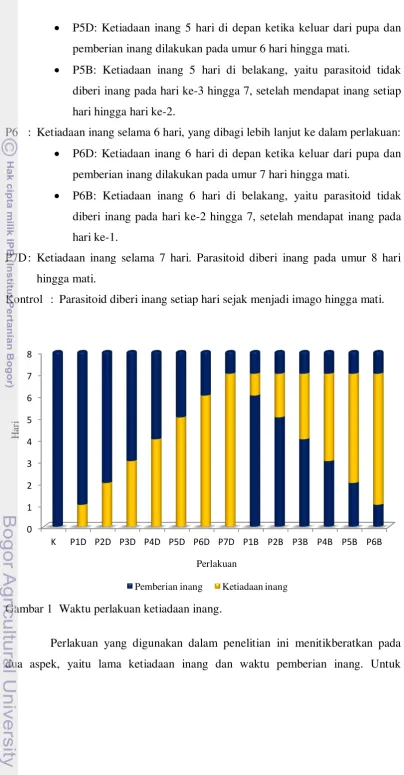
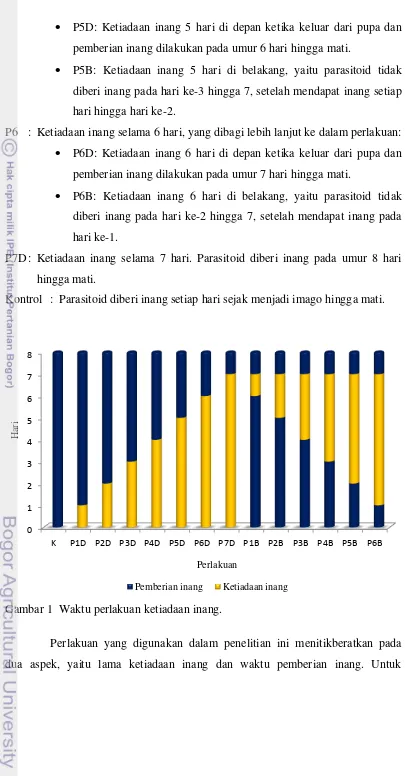
Penelitian ini disusun dengan Rancangan Acak Lengkap. Berikut adalah tingkat perlakuan yang digunakan, yaitu:
P1 : Ketiadaan inang selama 1 hari, yang dibagi lebih lanjut ke dalam perlakuan: P1D: Ketiadaan inang 1 hari di depan ketika keluar dari pupa dan
pemberian inang dilakukan pada umur 2 hari hingga mati.
P1B: Ketiadaan inang 1 hari di belakang, yaitu parasitoid tidak diberi inang pada hari ke-7 setelah mendapat inang setiap hari hingga hari ke-6.
P2 : Ketiadaan inang selama 2 hari, yang dibagi lebih lanjut ke dalam perlakuan: P2D: Ketiadaan inang 2 hari di depan ketika keluar dari pupa dan
pemberian inang dilakukan pada umur 3 hari hingga mati.
P2B: Ketiadaan inang 2 hari di belakang, yaitu parasitoid tidak diberi inang pada hari ke-6 dan 7, setelah mendapat inang setiap hari hingga hari ke-5.
P3 : Ketiadaan inang selama 3 hari, yang dibagi lebih lanjut ke dalam perlakuan: P3D: Ketiadaan inang 3 hari di depan ketika keluar dari pupa dan
pemberian inang dilakukan pada umur 4 hari hingga mati.
P3B: Ketiadaan inang 3 hari di belakang, yaitu parasitoid tidak diberi inang pada hari ke-5 hingga 7, setelah mendapat inang setiap hari hingga hari ke-4.
P4 : Ketiadaan inang selama 4 hari, yang dibagi lebih lanjut ke dalam perlakuan: P4D: Ketiadaan inang 4 hari di depan ketika keluar dari pupa dan
pemberian inang dilakukan pada umur 5 hari hingga mati.
P4B: Ketiadaan inang 4 hari di belakang, yaitu parasitoid tidak diberi inang pada hari ke-4 hingga 7, setelah mendapat inang setiap hari hingga hari ke-3.
P5D: Ketiadaan inang 5 hari di depan ketika keluar dari pupa dan pemberian inang dilakukan pada umur 6 hari hingga mati.
P5B: Ketiadaan inang 5 hari di belakang, yaitu parasitoid tidak diberi inang pada hari ke-3 hingga 7, setelah mendapat inang setiap hari hingga hari ke-2.
P6 : Ketiadaan inang selama 6 hari, yang dibagi lebih lanjut ke dalam perlakuan: P6D: Ketiadaan inang 6 hari di depan ketika keluar dari pupa dan
pemberian inang dilakukan pada umur 7 hari hingga mati.
P6B: Ketiadaan inang 6 hari di belakang, yaitu parasitoid tidak diberi inang pada hari ke-2 hingga 7, setelah mendapat inang pada hari ke-1.
P7D : Ketiadaan inang selama 7 hari. Parasitoid diberi inang pada umur 8 hari hingga mati.
Kontrol : Parasitoid diberi inang setiap hari sejak menjadi imago hingga mati.
Gambar 1 Waktu perlakuan ketiadaan inang.
Perlakuan yang digunakan dalam penelitian ini menitikberatkan pada dua aspek, yaitu lama ketiadaan inang dan waktu pemberian inang. Untuk
0 1 2 3 4 5 6 7 8
K P1D P2D P3D P4D P5D P6D P7D P1B P2B P3B P4B P5B P6B
H
a
ri
Perlakuan
19
mempermudah dalam pemberian kode pada 13 tingkat perlakuan yang digunakan maka digunakan kode P1D, P2D, P3D, P4D, P5D, P6D, P7D, P1B, P2B, P3B, P4B, P5B, dan P6B. Huruf ‘P’ merupakan kependekan dari ‘perlakuan’. Angka setelah huruf ‘P’ menunjukkan lama hari ketiadaan inang. Huruf ‘D’ setelah angka menunjukkan bahwa perlakuan ketiadaan inang diberikan di depan, yaitu perlakuan ketiadaan inang diberikan sejak awal S. manilae menjadi imago, sedangkan huruf ‘B’ menunjukkan bahwa perlakuan ketiadaan inang diberikan di belakang, yaitu perlakuan ketiadaan inang mulai diberikan setelah imago S. manilae mendapatkan inang pada awal kemunculannya menjadi imago.
Untuk setiap tingkat perlakuan dilakukan 10 kali ulangan. Larva inang dipaparkan selama 24 jam. Setiap 24 jam larva inang diganti dengan yang baru. Jumlah inang yang diberikan setiap 24 jam pada masing-masing unit perlakuan adalah 30 larva S. litura instar II akhir. Parasitoid yang digunakan adalah imago betina yang telah berkopulasi.
Parameter yang diamati/dihitung adalah: 1. Tingkat parasitisasi di hari ke-8
Jumlah inang yang terparasit dihitung dengan cara tubuh inang dibedah terlebih dahulu setelah 24 jam pemaparan. Pembedahan dilakukan dengan cara ujung posterior larva inangg ditarik dengan pinset halus sehingga bagian kutikula robek dan hemolimf serta telur parasitoid keluar. Persentase parasitisasi dihitung dengan rumus:
2. Jumlah telur yang diletakkan pada hari ke-8
Jumlah telur dihitung dengan cara tubuh inang dibedah terlebih dahulu. Jumlah telur yang terdapat pada tubuh larva inang dihitung.
3. Jumlah telur yang diletakkan pada hari pertama setelah pemuasaan.
Jumlah telur dihitung dengan cara tubuh inang dibedah terlebih dahulu kemudian jumlah telur yang terdapat dalam tubuh inang dihitung.
Jumlah telur yang tersisa dalam ovari S. manilae dihitung dengan cara tubuh parasitoid dibedah ketika parasitoid mati. Jumlah telur yang terdapat di dalam ovari dicatat.
5. Total produksi telur
Jumlah telur yang diletakkan sejak hari pertama parasitoid menjadi imago hingga parasitoid mati.
6. Lama hidup imago betina S. manilae
Jumlah hari sejak S. manilae menjadi imago hingga mati. 7. Tingkat superparasitisasi pada hari ke-8
Superparasitisasi yang terjadi dihitung dengan menggunakan rumus:
8. Tingkat superparasitisasi pada hari pertama setelah pemuasaan
Superparasitisasi yang terjadi dihitung dengan menggunakan rumus yang sama dengan tingkat superparasitisasi hari ke-8.
Pada parameter pertama (tingkat parasitisasi hari ke-8), kedua (jumlah telur yang diletakkan pada hari ke-8), dan ketujuh (tingkat superparasitisasi hari ke-8) waktu pengamatan yang digunakan adalah pada saat imago parasitoid berumur 8 hari. Pemilihan hari ke-8 sebagai waktu pengamatan parameter tersebut didasarkan pada uji pendahuluan yang telah dilakukan pada seratus individu imago betina S. manilae. Berdasarkan uji pendahuluan tersebut diketahui bahwa imago betina S. manilae yang terus-menerus diberi inang sejak kemunculannya menjadi imago dengan pemberian pakan berupa madu 20% rata-rata lama hidupnya hanya 8 hari, sehingga hari ke-8 digunakan sebagai waktu pengamatan parameter-parameter tersebut.
Analisis Data
HASIL DAN PEMBAHASAN
Hasil Penelitian
Pengaruh Ketiadaan Inang Terhadap Oviposisi di Hari Pertama Setelah Perlakuan
Hasil penelitian menunjukkan bahwa semakin lama S. manilae tidak mendapatkan inang maka oviposisi pada hari pertama mendapatkan inang semakin meningkat. Namun peningkatan ini hanya berlaku pada S. manilae yang diberi perlakuan ketiadaan inang di belakang (P1B hingga P6B). Pada perlakuan ketiadaan inang di depan (P1D hingga P7D), peningkatan oviposisi pada hari pertama mendapatkan inang hanya terjadi pada S. manilae yang tidak mendapatkan inang 1 hingga 3 hari (P1D, P2D, dan P3D). Bila S. manilae tidak mendapatkan inang lebih dari 3 hari berturut-turut (P4D, P5D, P6D dan P7D) ternyata menurunkan oviposisi pada hari pertama mendapatkan inang.
Gambar 2 Jumlah telur yang diletakkan oleh parasitoid S. manilae pada hari petama setelah ketiadaan inang S. litura.
*Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji Duncan pada taraf nyata 5%. K: Kontrol; D: ketiadaan inang di awal; B: ketiadaan inang di belakang; Angka: jumlah hari ketiadaan inang.
d* d
cd bc
cd
e e
e d
c cd
ab
a a
0 5 10 15 20 25 30 35 40 45
K P1D P2D P3D P4D P5D P6D P7D P1B P2B P3B P4B P5B P6B
Hasil tersebut menunjukkan bahwa saat imago tidak menemukan inang dan lama ketiadaan inang memengaruhi pola reproduksi. Parasitod S. manila
ternyata dapat langsung meletakkan telur ketika mendapatkan inang walaupun sebelumnya tidak mendapatkan inang hingga 7 hari berturut-turut (P7D), walaupun jumlahnya terendah dibandingkan dengan perlakuan lain. Rata-rata jumlah telur pada perlakuan P7D adalah 14,3 butir telur, tidak berbeda nyata terhadap P6D dan P5D (Gambar 2). Perbedaan yang mencolok terlihat antara perlakuan P6D dengan P6B. Walaupun keduanya sama-sama tidak diberi inang selama 6 hari, namun oviposisi pada hari pertama mendapat inang ternyata sangat jauh berbeda. Jumlah rata-rata telur yang diletakkan pada perlakuan P6B adalah 38,4 butir sedangkan P6D hanya 17,7 butir (Gambar 2).
Pengaruh Ketiadaan Inang Terhadap Kemampuan Parasitisasi S. manilae di
hari ke-8
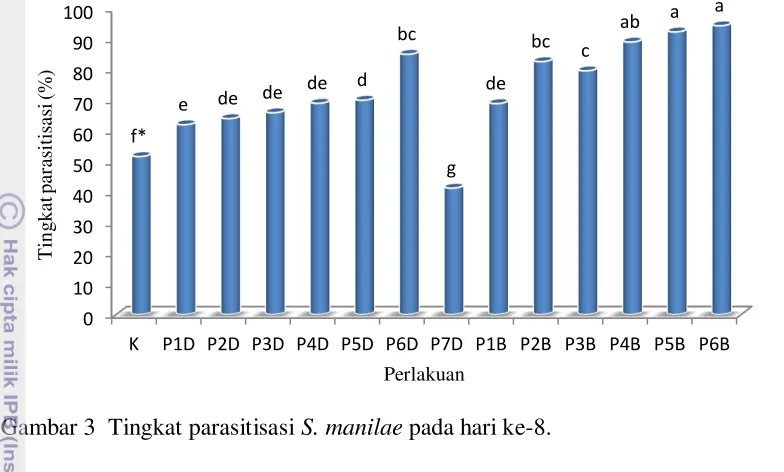
Hasil dari perlakuan ketiadaan inang dapat dilihat pada kemampuan parasitisasi hari ke-8. Hasil yang diperoleh dalam penelitian ini menunjukkan bahwa lama ketiadaan inang ternyata memengaruhi tingkat parasitisasi pada hari ke-8. Semakin lama S. manilae tidak mendapatkan inang maka parasitisasi pada hari ke-8 cenderung meningkat. Hal tersebut terjadi baik pada perlakuan dengan ketiadaan inang di depan (P1D hingga P7D) maupun perlakuan dengan ketiadaan inang di belakang (P1B hingga P6B).
23
Gambar 3 Tingkat parasitisasi S. manilae pada hari ke-8.
*Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji Duncan pada taraf nyata 5%. K: Kontrol; D: ketiadaan inang di awal; B: ketiadaan inang di belakang; Angka: jumlah hari ketiadaan inang.
Pengaruh Ketiadaan Inang Terhadap Produksi Telur S. manilae
Semakin lama S. manilae tidak mendapatkan inang maka jumlah telur yang diletakkan pada hari ke-8 semakin meningkat. Hal ini terjadi baik pada perlakuan-perlakuan dengan ketiadaan inang di depan (P1D hingga P7D) maupun perlakuan-perlakuan ketiadaan inang di belakang (P1B hingga P6B). Pada perlakuan ketiadaan inang di depan, lama ketiadaan inang hingga 7 hari (P7D) ternyata dapat menurunkan jumlah telur yang diletakkan pada hari ke-8. Jumlah telur tertinggi yang diletakkan oleh parasitoid pada hari ke-8 terdapat pada perlakuan P6D, yaitu sebesar 41,3 butir telur, tidak berbeda nyata terhadap P5B dan P6B (Gambar 4).
Ketiadaan inang ternyata memberikan pengaruh yang sebaliknya pada jumlah telur yang diletakkan hingga hari ke-8. Semakin lama S. manilae tidak mendapatkan inang maka jumlah telur yang diproduksi hingga hari ke-8 semakin menurun. Kecenderungan penurunan jumlah telur ini terjadi baik pada perlakuan dengan ketiadaan inang di depan (P1D hingga P7D) maupun perlakuan-perlakuan ketiadaan inang di belakang (P1B hingga P6B). Pada perlakuan-perlakuan dengan
0 10 20 30 40 50 60 70 80 90 100
K P1D P2D P3D P4D P5D P6D P7D P1B P2B P3B P4B P5B P6B
f*
e de de
de d
bc
g de
bc c
ab a a
0 20 40 60 80 100 120 140 160 180 200
K P1D P2D P3D P4D P5D P6D P7D P1B P2B P3B P4B P5B P6B
Jum
la
h
te
lur
(b
ut
ir
)
Perlakuan
Jumlah telur hari ke-8 Jumlah telur sampai hari ke-8
ketiadaan inang di depan, lama ketiadaan inang lebih dari 6 hari (P7D) menyebabkan jumlah produksi telur turun secara drastis. Rata-rata jumlah telur tertinggi terdapat pada kontrol (174,3 butir telur), sedangkan yang terendah terdapat pada perlakuan P7D (14,3 butir telur) yang berbeda nyata terhadap perlakuan lainnya (Gambar 4).
Gambar 4 Jumlah telur parasitoid S. manilae pada hari ke-8 dan hingga hari ke-8. Pengaruh Ketiadaan Inang Terhadap Sisa Telur dalam Ovari S. manilae
25
Gambar 5 Rata-rata sisa telur S. manilae di dalam ovari.
*Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji Duncan pada taraf nyata 5%. K: Kontrol; D: ketiadaan inang di awal; B: ketiadaan inang di belakang; Angka: jumlah hari ketiadaan inang.
Jumlah telur yang tersisa dalam ovari tertinggi terdapat pada perlakuan P6B (99,0 butir telur) berbeda nyata dengan semua perlakuan lainnya (Gambar 5). Perbedaan yang nyata terlihat antara perlakuan P6D dan P6B. Walaupun keduanya tidak mendapatkan inang selama 6 hari berturut-turut, namun sisa telur di dalam ovari parasitoid pada perlakuan P6D hanya sekitar setengah dari jumlah telur yang tersisa dalam ovari parasitoid pada perlakuan P6B.
Pengaruh Ketiadaan Inang Terhadap Jumlah Total Produksi Telur S. manilae
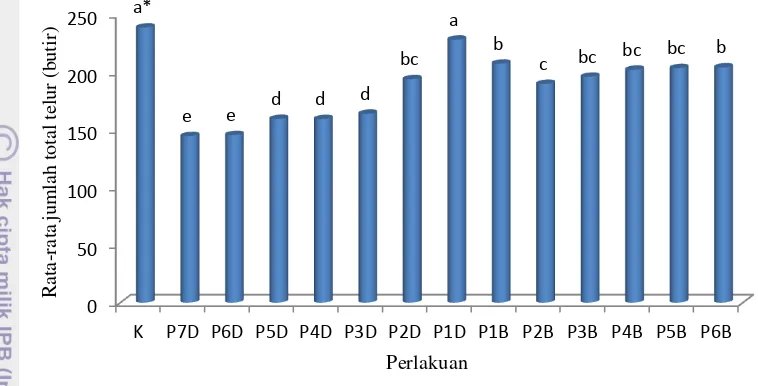
Pada perlakuan ketiadaan inang di depan semakin lama S. manilae tidak mendapatkan inang maka jumlah total produksi telurnya semakin menurun. Dengan kata lain, bila S. manile tidak mendapatkan inang sejak awal menjadi imago maka ketiadaan inang cenderung menyebabkan penurunan jumlah total produksi telurnya. Hal yang berbeda terjadi pada perlakuan-perlakuan dengan ketiadaan inang di belakang (P1B hingga P6B). Pada perlakuan-perlakuan tersebut lama ketiadaan inang cenderung tidak memengaruhi jumlah total telur
0 10 20 30 40 50 60 70 80 90 100
K P1D P2D P3D P4D P5D P6D P7D P1B P2B P3B P4B P5B P6B
d* d
ef
g f
ef e
cd
f ef
yang diproduksi. Jumlah total telur yang diproduksi cenderung tetap, walaupun jumlahnya lebih rendah daripada kontrol.
Gambar 6 Rata-rata jumlah total telur parasitoid S. manilae.
*Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji Duncan pada taraf nyata 5%. K: Kontrol; D: ketiadaan inang di awal; B: ketiadaan inang di belakang; Angka: jumlah hari ketiadaan inang.
Rata-rata jumlah telur tertinggi terlihat pada kontrol (238,7 butir telur) tidak berbeda nyata terhadap P1D namun berbeda nyata terhadap perlakuan lainnya (Gambar 6). Pada kontrol, imago betina S. manilae selalu diberi inang sejak kemunculannya menjadi imago hingga hari kematiannya sehingga dapat terus meletakkan telur.
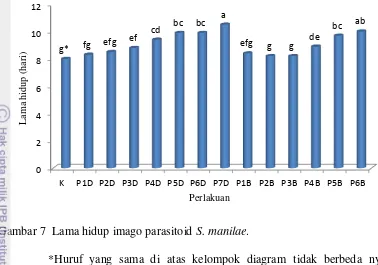
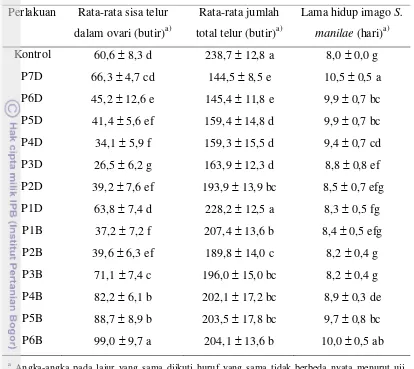
Pengaruh Ketiadaan Inang Terhadap Lama Hidup Imago S. manilae
Lama hidup imago betina S. manilae yang diberi perlakuan ketiadaan inang dalam jangka waktu tertentu ditunjukkan pada Gambar 5. Berdasarkan hasil penelitian diketahui bahwa lama ketiadaan inang ternyata memengaruhi lama hidup imago S. manilae. Semakin lama imago S. manilae tidak mendapatkan inang maka lama hidupnya cenderung semakin meningkat. Peningkatan lama hidup ini terjadi baik pada perlakuan-perlakuan dengan ketiadaan inang di depan (P1D hingga P7D) maupun perlakuan-perlakuan dengan ketiadaan inang di belakang (P1B hingga P6B).
0 50 100 150 200 250
K P7D P6D P5D P4D P3D P2D P1D P1B P2B P3B P4B P5B P6B
a*
e e d d
d bc
a b
c bc bc bc b
27
Gambar 7 Lama hidup imago parasitoid S. manilae.
*Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji Duncan pada taraf nyata 5%. K: Kontrol; D: ketiadaan inang di awal; B: ketiadaan inang di belakang; Angka: jumlah hari ketiadaan inang.
Menurut Prabowo (1996), imago betina S. manilae yang diberi makan madu dapat hidup hingga 9 hari, sedangkan bila hanya diberi air hanya mampu bertahan hidup hingga 5 hari. Rata-rata lama hidup tertinggi terjadi pada parasitoid dengan perlakuan P7D (10,5 hari) tidak berbeda nyata terhadap P6B namun berbeda nyata terhadap perlakuan lainnya (Gambar 7). Pada perlakuan P7D, parasitoid tidak diberikan inang sejak awal menjadi imago hingga 7 hari berturut-turut. Lama hidup terendah terjadi pada imago betina S. manilae pada kontrol (8 hari). Pada kontrol, imago betina S. manilae terus menerus diberi inang sejak kemunculannya sebagai imago hingga mati. Hal tersebut sesuai dengan hasil penelitian Ratna (2008) bahwa S. manilae yang diberi inang terus-menerus lama hidupnya berkisar antara 5 hingga 8 hari.
Pengaruh Ketiadaan Inang Terhadap Superparasitisasi S. manilae
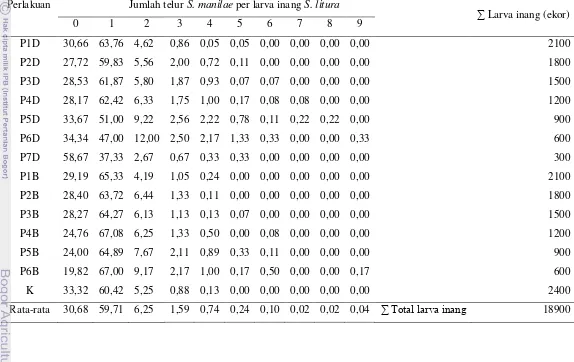
Superparasitisme merupakan kejadian dimana imago parasitoid meletakkan telur lebih dari satu pada setiap larva inang. Tabel 1 menunjukkan persentase kejadian superparasitisasi dengan jumlah telur yang diletakkan pada
0 2 4 6 8 10 12
K P1D P2D P3D P4D P5D P6D P7D P1B P2B P3B P4B P5B P6B
g* fg efg
ef cd
bc bc a
efg g g de
bc ab
L
am
a
h
idup
(h
ar
i)
satu larva mulai dari satu hingga sembilan butir telur. Secara umum superparasitisasi yang terjadi sangatlah kecil persentasenya. Pada kejadian superparasitisasi ternyata diketahui bahwa kebanyakan yang terjadi adalah peletakkan telur sebanyak dua butir telur per larva inang. Superparasitisasi yang terjadi persentasenya masih lebih kecil dibandingkan dengan parasitisasi normal (jumlah telur satu butir per larva inang). Secara umum persentase parasitisasi normal nilainya diatas 50%, kecuali pada perlakuan P6D (47,00%) dan P7D (37,33%).
29
Tabel 1 Persentase superparasitisasi S. manilae.
Perlakuan Jumlah telur S. manilae per larva inang S. litura
∑ Larva inang (ekor) 0 1 2 3 4 5 6 7 8 9
Pembahasan
Pengaruh Ketiadaan Inang Terhadap Perilaku Oviposisi S. manilae
Waktu ketiadaan inang ternyata memengaruhi kemampuan parasitoid S. manilae dalam meletakkan telurnya. Secara umum ketiadaan inang di awal kehidupan imago S. manilae menyebabkan penurunan kegiatan oviposisi ketika pertama kali mendapatkan inang. Hal yang sebaliknya terjadi bila ketiadaan inang terjadi setelah imago S. manilae bertemu inang. Pada parasitoid yang memiliki pengalaman bertemu inang (perlakuan ketiadaan inang di belakang), lama ketiadaan inang ternyata menyebabkan peningkatan oviposisi pada hari pertama bertemu inang. Hal tersebut sesuai dengan yang diutarakan oleh Vinson (1985
dalam Ratna 2008) bahwa imago betina parasitoid cenderung meningkatkan oviposisi ketika jumlah inang terbatas dalam jangka waktu yang relatif lama. Hal yang sebaliknya terjadi pada parasitoid yang tidak memiliki pengalaman bertemu inang (perlakuan ketiadaan inang di awal).
Hasil penelitian menunjukkan bahwa ketiadaan inang selama 6 hari akan menyebabkan oviposisi menurun bila perlakuan diberikan di awal kehidupan imago, namun bila perlakuan baru diberikan di belakang oviposisi cenderung meningkat. Hal tersebut menunjukkan bahwa pengalaman pertama mendapatkan inang ternyata dapat menyebabkan parasitoid meningkatan perilaku oviposisi walaupun tidak mendapatkan inang selama 6 hari berturut-turut. Hal ini sesuai dengan pernyataan Drost & Carde (1992) yang menyebutkan bahwa paparan awal inang pada parasitoid dapat mendorong perilaku reproduksi parasitoid menjadi lebih aktif. Ketiadaan inang hingga 5 hari atau lebih yang diikuti dengan tidak adanya pengalaman bertemu inang ternyata dapat menurunkan jumlah telur yang diletakkan pada hari pertama bertemu inang. Penyebab lain yang mungkin adalah terhentinya produksi telur akibat ketiadaan inang dalam waktu yang relatif lama.
Drost & Carde (1992) menyebutkan bahwa ketiadaan inang pada spesies
31
dan synovigenic. Golongan pro-ovigenic adalah parasitoid betina yang muncul menjadi imago dengan sejumlah telur yang telah matang, sedangkan golongan
synovigenic muncul menjadi imago tanpa atau dengan sedikit telur yang matang dan kemudian akan memproduksi telur sepanjang hidupnya (Godfray 1994). Namun Quicke (1997) juga menyatakan diantara kedua kelompok tersebut terdapat kelompok yang disebut pro-synovigenic yaitu serangga yang muncul menjadi dewasa dengan sejumlah telur yang sudah matang tetapi dapat melakukan pematangan telur lebih banyak lagi saat pasokan asli telur mereka habis.
Perilaku oviposisi cenderung meningkat seiring meningkatnya lama ketiadaan inang. Hal tersebut menunjukkan bahwa pengalaman bertemu inang ternyata memengaruhi perilaku oviposisi S. manilae. Akan tetapi lama ketiadaan inang ternyata tidak memengaruhi kemampuan parasitoid dalam melakukan oviposisi. Parasitod S. manilae tetap mampu langsung meletakkan telur ketika mendapatkan inang walaupun sebelumnya tidak mendapatkan inang hingga 7 hari berturut-turut walaupun jumlah telurnya rendah. Kemampuan meletakkan telur langsung walaupun dalam jangka waktu yang relatif lama tidak mendapatkan inang merupakan hal yang penting yang dibutuhkan dari suatu musuh alami. Hal yang sama terjadi pada penelitian yang dilakukan oleh Hougardy et al. (2005) terhadap M. ridibundus yang dapat langsung menggunakan telur-telur dalam ovariolnya bahkan setelah 6 hari tidak mendapatkan inang.
sebelumnya sehingga tidak memiliki pengalaman dalam pemarasitan. Drost & Carde (1992) menyebutkan bahwa ketiadaan inang parasitoid dapat menyebabkan rendahnya tingkat parasitisasi karena belum adanya pengalaman oviposisi.
Pengaruh Ketiadaan Inang Terhadap Produksi Telur dan Lama Hidup S. manilae
Waktu pemberian inang ternyata tidak memengaruhi jumlah telur yang diletakkan. Walaupun demikian, lama ketiadaan inang ternyata dapat memengaruhi produksi telur S. manilae. Semakin lama S. manilae tidak mendapatkan inang maka jumlah telur yang diletakkan oleh S. manilae baik pada imago yang mendapat perlakuan di awal kehidupan maupun di belakang sama-sama cenderung menurun. Hal tersebut dapat disebabkan semakin lama ketiadaan inang maka semakin sedikit inang yang dapat digunakan untuk meletakkan telur-telurnya. Jumlah keturunan yang dihasilkan oleh parasitoid pun akan semakin menurun akibat tidak tersedianya inang sebagai tempat bagi telur-telur parasitoid untuk tumbuh dan berkembang. Vinson (1985 dalam Ratna 2008) menyebutkan bahwa kecenderungan imago betina parasitoid meningkatkan oviposisi terjadi bila sediaan jumlah inang terbatas dalam jangka waktu yang relatif lama. Namun dalam penelitian ini peningkatan hanya terjadi pada hari pertama parasitoid bertemu inang.
33
beberapa kasus memberi dampak positif terhadap potensi reproduksi di masa depan karena dapat merangsang oogenesis dan kemampuan menemukan inang.
Secara umum lama ketiadaan inang memengaruhi jumlah telur yang tersisa di ovari dan juga menyebabkan peningkatan sisa telur dalam ovari parasitoid. Hal ini dapat menjadi indikasi bahwa ketika parasitoid tidak mendapatkan inang, maka telur yang ada akan disimpan hingga saatnya bertemu inang. Namun peningkatan yang terjadi berbeda antara parasitoid yang diberi inang sejak awal kemunculan menjadi imago dengan yang sama sekali tidak diberi inang pada awal kemunculan menjadi imago. Ketiadaan inang selama 6 hari di awal kehidupan imago S. manilae ternyata menyebabkan sisa telur dalam ovari hanya setengahnya dari imago yang mengalami ketiadaan inang di belakang. Hal tersebut menunjukkan bahwa pemberian inang diawal kemunculan imago parasitoid dapat menstimulasi parasitoid untuk meningkatkan produksi telurnya walaupun tidak mendapatkan inang hingga 6 hari berturut-turut.
menghemat energi ketika dalam kondisi stress seperti suhu rendah dan kekurangan makanan.
Hal yang sebaliknya terjadi pada pemberian perlakuan di belakang. Rata-rata jumlah total telur cenderung meningkat seiring dengan meningkatnya waktu ketiadaan inang. Peningkatan rata-rata jumlah total telur ini dapat dipicu oleh ketersediaan inang di awal kemunculan parasitoid menjadi imago yang kemudian menstimulasi produksi telur walaupun pada beberapa hari berikutnya tidak tersedia inang. Pemberian inang secara berkelanjutan seperti pada kontrol memungkinkan imago betina S. manilae untuk terus melakukan oviposisi sepanjang hidupnya sehingga telur yang diletakkan lebih banyak dibandingkan imago betina S. manilae pada perlakuan lainnya. Selain itu juga ketersediaan inang yang berkelanjutan dapat mendorong produksi telur terjadi sepanjang hidupnya. Drost & Carde (1992) menyebutkan dalam penelitiannya mengenai pengaruh ketiadaan inang pada Brachymeria intermedia bahwa total produksi telur bergantung pada jumlah inang yang ditemuinya, hal tersebut menunjukkan bahwa parasitoid mengatur produksi telur pada inang yang tersedia.
Hasil penelitian memperlihatkan secara umum ketiadaan inang memengaruhi lama hidup imago betina S. manilae. Kecenderungan yang terlihat adalah semakin lama imago betina S. manilae tidak mendapatkan inang, maka lama hidupnya semakin meningkat. Ketiadaan inang dalam jangka waktu tersebut menyebabkan inang menyimpan energi yang seharusnya digunakan untuk oviposisi sehingga umur imago betina menjadi lebih panjang. Hal serupa terjadi pada parasitoid A. nitens. Lama hidup A. nitens meningkat secara signifikan pada perlakuan ketiadaan inang dengan pemberian makanan, yang dapat disebabkan tidak adanya energi yang digunakan untuk reproduksi, ketersediaan makanan dan adanya penyerapan kembali telur (Carbone et al. 2008). Namun hal yang sebaliknya terjadi pada parasitoid Venturia canescens (Hymenoptera: Ichneumonidae) dalam penelitian yang dilakukan oleh Eliopoulos et al. (2005). Imago yang diberi makanan namun tidak diberi inang tidak secara signifikan hidup lebih lama dibandingkan imago yang diberi makanan dan inang.
35
matang (oosorption) sebagai respon terhadap ketiadaan inang. Penyerapan kembali telur adalah mekanisme yang memungkinkan imago betina untuk mendaur ulang nutrisi yang tersimpan di dalam telur-telur mereka untuk pemeliharaan somatik pada saat terjadi stress ketika makanan atau inang berkurang di alam (Jervis & Kidd 1986 dalam Hougardy et al. 2005) atau untuk memelihara pasokan telur matang yang baru secara konstan hingga imago betina dapat melanjutkan oviposisi ketika inang menjadi sedikit (Rivero-Lynch & Godfray 1997 dalam Hougardy et al. 2005). Selain itu Quicke (1997) juga menyebutkan oosorption terjadi dalam beberapa hari setelah pematangan telur bila tidak tersedia inang. Nutrisi yang dihasilkan dari proses penyerapan kembali telur digunakan sebagai sumber energi untuk hidup parasitoid. Pemberian inang secara terus menerus menjadikan imago betina S. manilae terus melakukan oviposisi dan mendorong produksi telur sepanjang hidupnya. Oviposisi dan produksi telur membutuhkan energi sehingga dapat memperpendek lama hidup imago.
Pengaruh Ketiadaan Inang Terhadap Superparasitisasi S. manilae
Superparasitisme menurut Quicke (1997) adalah peletakkan telur lebih dari yang normal pada individu inang yang dilakukan oleh suatu parasitoid. Lebih jauh lagi Godfray (1994) menyebutkan mengenai self-superparasitism, yaitu superparasitisme yang dilakukan oleh individu parasitoid yang sama. Prabowo (1996) menyebutkan bahwa S. manilae merupakan parasitoid soliter, yaitu hanya meletakkan satu telur pada satu individu inang. Namun berdasarkan hasil penelitian ditemukan adanya kejadian superparasitisme.
Secara umum imago S. manilae tidak melakukan superparasitisasi karena
secara terus-menerus persentase parasitisasi optimal didapatkan sebesar 44,7% dan kejadian superparasitisasi 19,6%.
Superparasitisasi yang terjadi jumlahnya jauh lebih rendah daripada parasitisasi normal. Selain itu juga perilaku superparasitisme dihindari oleh parasitoid karena menurut Ratna (2008) superparasitisasi dapat menurunkan tingkat keberhasilan hidup parasitoid dalam tubuh inang. Godfray (1994) juga menyebutkan bahwa superparasitisme menyebabkan terjadinya kompetisi nutrisi antar larva parasitoid yang berhasil menetas dari telur, sehingga menyebabkan kedua larva mati atau satu individu dapat berkembang dengan pertumbuhan tidak optimal sehingga terjadi penurunan kebugaran parasitoid yang berhasil menjadi imago.
Superparasitisasi yang terjadi didominasi oleh peletakan 2 butir telur per larva inang dan menurun seiring bertambahnya jumlah telur yang diletakkan per larva inang. Hal tersebut dapat disebabkan parasitoid sebisa mungkin menghindari peletakan lebih dari 2 butir telur per larva inang. Semakin banyak jumlah telur yang diletakkan per larva inang makan tingkat keberhasilan hidup parasitoid dalam tubuh inang semakin kecil akibat adanya kompetisi yang terjadi. Selain itu juga superparasitisme dapat menyebabkan peningkatan kematian inang (Mehrnejad & Copland 2007).
37
dan belum belajar mengenai kelimpahan mutlak inang maupun berapa proporsi inang yang telah diparasitisasi (Godfray 1994).
KESIMPULAN DAN SARAN
Kesimpulan
Ketiadaan inang ternyata mempengaruhi pola reproduksi imago S. manilae. Secara umum ketiadaan inang di awal kehidupan imago S. manilae
menyebabkan penurunan perilaku oviposisi ketika pertama kali mendapatkan inang. Hal yang sebaliknya terjadi bila ketiadaan inang terjadi setelah imago S. manilae bertemu inang. Pengalaman pertama mendapatkan inang ternyata dapat menyebabkan parasitoid meningkatan perilaku oviposisi. Lama ketiadaan inang ternyata tidak memengaruhi kemampuan parasitoid dalam melakukan oviposisi. Ketiadaan inang selama 6 hari akan menyebabkan parasitisasi menurun jika perlakuan diberikan di awal kehidupan imago S. manilae, namun jika perlakuan diberikan di belakang setelah bertemu inang maka parasitisasi cenderung lebih tinggi. Waktu pemberian inang ternyata tidak memengaruhi jumlah telur yang diletakkan, namun semakin lama S. manilae tidak mendapatkan inang maka jumlah telur yang diletakkan oleh S. manilae cenderung menurun. Ketiadaan inang di awal kehidupan imago menyebabkan jumlah telur yang tersisa dalam ovari S. manilae lebih rendah daripada ketiadaan inang di belakang. Jumlah total telur yang diproduksi cenderung menurun seiring meningkatnya lama ketiadaan inang bila perlakuan diberikan di awal kehidupan imago S. manilae. Ketiadaan inang juga cenderung menyebabkan lama hidup imago S. manilae meningkat.
Superparasitisme ditemukan terjadi pada S. manilae. Kejadian superparasitisasi pada penelitian ini pada umumnya terjadi akibat jumlah telur yang diletakkan melebihi jumlah inang yang dipaparkan, namun superparasitisme juga ditemukan terjadi akibat parasitoid tidak pernah bertemu inang sebelumnya sehingga belum memiliki pengalaman dalam oviposisi dan diskriminasi inang.
Saran
DAFTAR PUSTAKA
Abdurrahman. 2011. Pengendalian Penyakit Tumbuhan Secara Hayati yang Ekologis dan Berkelanjutan. http://epetani.deptan.go.id/blog/pengendalian- penyakit-tumbuhan-secara-hayati-yang-ekologis-dan-berkelanjutan-bag-7-1972. [2 Oktober 2011].
Carbone SS, Nieto MP, Rivera AC. 2008. Egg resorption behavior by the solitary egg parasitoid Anaphes nitens under natural condition. Entomologia Experimentalis et Applicata 127: 191-198.
Darwati R. 1999. Pengaruh umur parasitoid terhadap persentase dan keberhasilan hidup Snellenius (=Microplitis) manilae Ashmead (Hymenoptera: Braconidae) pada inang Spodoptera litura Fabricius (Lepidoptera: Noctuidae) [skripsi]. Bogor: Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Institut Pertanian Bogor.
Driesche RV, Mark H, Ted C. 2008. Control of Pest and Weeds by Natural Enemies: An Introduction to Biological Control. Oxford: Blackwell Publishing.
Drost YC, Carde RT. 1992. Influence of host deprivation of egg load and oviposition behavior of Brachymeria intermedia, a parasitoid of gypsy moth. Physiol. Entomol. 17 (3): 230-234.
Eliopoulos PA, Stathas GJ, Bouras SL. 2005. Effects and interactions of temperature, host deprivation and adult feeding on the longevity of the parasitoid Venturia canescens (Hymenoptera: Ichneumonidae). Europea