PEMANFAATAN TUNGAU PREDATOR EKSOTIS DAN
POTENSI TUNGAU PREDATOR LOKAL SEBAGAI AGENS
PENGENDALI HAYATI TUNGAU HAMA PADA TANAMAN
STROBERI
EDWIN ISWELLA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Pemanfaatan Tungau Predator Eksotis dan Potensi Tungau Predator Lokal sebagai Agens Pengendali Hayati Tungau Hama pada Tanaman Stroberi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2015
Edwin Iswella
RINGKASAN
EDWIN ISWELLA. Pemanfaatan Tungau Predator Eksotis dan Potensi Tungau Predator Lokal sebagai Agens Pengendali Hayati Tungau Hama pada Tanaman Stroberi. Dibimbing oleh PUDJIANTO dan SUGENG SANTOSO.
Tungau laba-laba merupakan hama penting dan merusak pada tanaman stroberi di Indonesia. Pengendalian tungau laba-laba menggunakan tungau predator famili Phytoseiidae telah banyak dilakukan di luar negeri. Sejak tahun 2009, satu spesies tungau predator eksotis, Phytoseiulus persimilisAthias Henriot
(Acari: Phytoseiidae) telah digunakan untuk mengendalikan T. urticae pada
tanaman stroberi di rumah kaca. Penelitian ini bertujuan untuk mengetahui kemampuan menetap dan perkembangan populasi P. persimilis serta potensi
tungau predator lokal N. longispinosus pada tanaman stroberi.
Penelitian dilaksanakan di rumah kaca PT Strawberindo Lestari, Cianjur, Jawa Barat serta Laboratorium Bionomi dan Ekologi Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor dari bulan September 2014 sampai dengan Februari 2015. Penelitian yang dilakukan adalah pengamatan populasi tungau hama, tungau predator eksotis, dan tungau predator lokal di rumah kaca; pengamatan keberadaan tungau predator eksotis di luar rumah kaca dan uji kemampuan memangsa serta potensi kanibalisme tungau predator lokal di laboratorium.
Hasil penelitian menunjukkan bahwa tungau predator P. persimilis tidak
ditemukan baik di dalam maupun di luar rumah kaca. Tungau predator yang ditemukan adalah N. longispinosus dengan populasi yang cenderung konstan
selama 8 minggu pengamatan. Populasi T. urticae berfluktuasi dan semakin
menurun di semua lokasi selama 8 minggu pengamatan. Pada uji kemampuan memangsa, rata-rata telur T. urticae dan T. kanzawai yang dimangsa imago
betina predator adalah 14.15 1.87 dan 15.15 1.20 butir. Jumlah ini secara
nyata lebih tinggi (P<0.05) dibandingkan telur yang dimangsa oleh deutonimfa predator. Rata-rata imago betina T. urticae dan T. kanzawai yang dimangsa
deutonimfa predator adalah 1.05 0.32 dan 1.05 0.41 ekor. Hasil ini tidak
berbeda nyata (P=0.486) dengan jumlah yang dimangsa oleh imago betina predator (1.95 0.32; 2.15 0.57 ekor). Rata-rata jumlah telur yang diletakkan
imago betina N. longispinosusyang diberi mangsa berupa telur (2.00 0.39; 1.80 0.73 butir) secara nyata lebih tinggi (P<0.05) dibanding dengan yang diberi
mangsa imago betina T. urticaedan T. kanzawai(0.90 0.13; 0.70 0.20 butir).
Deutonimfa maupun imago betina N. longispinosusmemiliki sifat kanibal.
Rata-rata predator yang dimangsa konspesifik baik pada perlakuan komposisi fase yang sama maupun komposisi fase campuran semakin meningkat dengan meningkatnya kerapatan predator. Pada saat kondisi tidak ada mangsa, imago betina N. longispinosus masih dapat menghasilkan telur. Rata-rata jumlah telur yang
diletakkan imago betina N. longispinosus semakin meningkat dengan
meningkatnya kerapatan predator.
SUMMARY
EDWIN ISWELLA. The Use of Exotic Predatory Mite and the Potency of Native Predatory Mite as Biological Control Agent of Phytophagous Mite on Strawberry. Supervised by PUDJIANTO and SUGENG SANTOSO.
The two-spotted spider mite (TSSM) is one of the most important pests on strawberry grown in fields and greenhouses in Indonesia. The control of the pest with predatory mites has been carried out in many countries for several decades. Since 2009, a species of exotic predatory mite, Phytoseiulus persimilis Athias
Henriot (Acari: Phytoseiidae), has been introduced to Indonesia for the control of TSSM on strawberry grown in greenhouses. The objectives of this study were to investigate the establishment of the exotic predatory mites, and the potency of native predatory mites as biological control agent of TSSM. The research was conducted by observing the population of the exotic and native predatory mites inside and outside greenhouses. The predation rate and cannibalistic behavior of native predatory mite were also studied in laboratory. The results indicated that the exotic predatory mite,P. persimilis,was not found neither inside nor outside
the greenhouses. A native species of predatory mite,Neoseiulus longispinosus
Evans (Acari: Phytoseiidae) was found. Predation rate of deutonymphs and adult of N. longispinosus on egg was higher than on adult female of T. urticaeand T. kanzawai. Fecundity of N. longispinosuswhen fed with eggs of T. urticae andT. kanzawai was higher than with the female adults. Cannibalism was encountered in
both the deutonymphs and the female adults of N. longispinosuswhen the prey
was not available.
€ Hak Cipta Milik IPB, Tahun 2015 Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
PEMANFAATAN TUNGAU PREDATOR EKSOTIS DAN
POTENSI TUNGAU PREDATOR LOKAL SEBAGAI AGENS
PENGENDALI HAYATI TUNGAU HAMA PADA TANAMAN
STROBERI
EDWIN ISWELLA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Entomologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Pemanfaatan Tungau Predator Eksotis dan Potensi Tungau Predator Lokal sebagai Agens Pengendali Hayati Tungau Hama pada Tanaman Stroberi
Nama : Edwin Iswella NIM : A 351130394
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karuniaNya sehingga penulis bisa menyelesaikan tesis ini. Tema penelitian ini adalah pengendalian hayati, dengan judul Pemanfaatan Tungau Predator Eksotis dan Potensi Tungau Predator Lokal sebagai Agens Pengendali Hayati Tungau Hama pada Tanaman Stroberi.
Penulis mengucapkan terima kasih dan penghargaan yang sebesar-besarnya kepada Dr Ir Pudjianto, MSi dan Dr Ir Sugeng Santoso, MAgr selaku komisi pembimbing yang telah banyak memberi ilmu, saran, masukan dan bimbingannya selama proses penelitian dan penyusunan tesis ini. Terima kasih penulis sampaikan kepada Dr Ir Retno Dyah Puspitarini, MS sebagai penguji luar komisi atas saran dan masukannya. Terima kasih kepada Prof Dr Ir Sri Hendrastuti Hidayat, MSc selaku Ketua Program Studi Fitopatologi atas arahan serta bimbingan selama perkuliahan. Ucapan terimakasih juga disampaikan kepada segenap Pimpinan Pusat Badan Karantina Pertanian yang telah memberikan beasiswa pendidikan pada Sekolah Pascasarjana IPB dan PT Strawberindo Lestari yang telah mengizinkan dan menyediakan bahan serta tempat penelitian.
Penghargaan penulis kepada Dr Ir Tri Asmira Damayanti, MAgr atas rekomendasinya saat penulis mendaftar ke Sekolah Pascasarjana IPB. Terima kasih penulis sampaikan kepada Dra Latifatul Ainy, MSi atas rekomendasi, dukungan dan nasehatnya. Penulis juga mengucapkan terima kasih kepada seluruh rekan mahasiswa S2 Entomologi dan Fitopatologi Badan Karantina Pertanian angkatan tahun 2013 atas persahabatan, dukungan dan bantuannya selama melaksanakan studi, teman-teman di Laboratorium Bionomi dan Ekologi Serangga khususnya Pak Wawan Yuandi atas bantuannya selama penelitian. Ucapan terima kasih penulis sampaikan kepada Bapak Sumartono dan Bapak Asep di PT Strawberindo Lestari atas bantuannya. Terima kasih yang terdalam teruntuk istriku tercinta Kartika Candra Sari dan putriku tersayang Haura Sahda Nadhifa atas doa, dorongan dan dukungannya hingga penulis menyelesaikan studi.
Akhir kata semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2015
DAFTAR ISI
Tungau Hama pada Tanaman Stroberi (Fragaria spp.) 3
Tetranychus urticaeKoch (Acari: Tetranychidae) 3
Taksonomi dan Morfologi 3
Siklus Hidup 4
Penyebaran 4
Pengendalian 4
Phytoseiulus persimilisAthias Henriot (Acari: Phytoseiidae) 5
Taksonomi dan Morfologi 5
Populasi Tungau Hama dan Tungau Predator di Dalam Rumah
Kaca 10
Keberadaan Tungau Predator Eksotis di Luar Rumah Kaca 11 Tingkat Pemangsaan N. longispinosus 12
Pemeliharaan N. longispinosus, T. urticae, dan T. kanzawai 12
Kemampuan MemangsaN. longispinosusterhadap T. urticae,
dan T. Kanzawai 13
Pengaruh Fase Mangsa terhadap Tingkat Oviposisi N.
longispinosus 13
Potensi KanibalismeN. longispinosus 13
Analisis Data 14
HASIL DAN PEMBAHASAN 15
Keberadaan Tungau Predator Eksotis di dalam dan di luar Rumah
Kaca 15
Perkembangan Populasi Tungau Predator Lokal dan Tungau Hama di
Kemampuan Memangsa N. longispinosus 21
Pengaruh Fase Mangsa terhadap Tingkat Oviposisi N. longispinosus 23
Potensi KanibalismeN. longispinosus 24
Tingkat Oviposisi N. longispinosuspada Uji Kanibalisme 26
SIMPULAN DAN SARAN 27
DAFTAR PUSTAKA 28
LAMPIRAN 31
DAFTAR TABEL
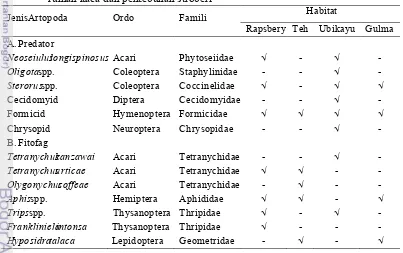
1 Jenis artopod fitofag dan predator yang ditemukan pada habitat
di luar rumah kaca dan perkebunan stroberi 16 2 Rata-rata telur dan imago betina T. urticaedan T. kanzawai
yang dimangsa deutonimfa dan imago betina N. longispinosus 21
3 Tingkat kanibalisme N. longispinosus pada komposisi fase
sama dan campuran pada beberapa tingkat kerapatan 25
DAFTAR GAMBAR
1 Imago T. urticaedengan dua spot berwarna gelap pada bagian
dorsal (Zhang 2003) 4
2 Bagian dorsal P. persimilisyang memiliki 23 pasang seta
dengan seta median j4–j6 dan s4 berukuran panjang (Gerson et
al. 2003) 5
3 Siklus hidup tungau predator P. persimilis(CABI 2007) 6
4 Bagian dorsal N. longispinosusdengan 17 pasang seta, sebagian
seta dorsal berukuran panjang kecuali seta j1 dan s5 (Zhang
2003) 7
5 Blok pelepasan predator eksotis di dalam rumah kaca PT
Strawberindo Lestari 10
6 Titik pengambilan sampel di sekitar rumah kaca 11 7 Titik pengambilan sampel di sekitar perkebunan stroberi 12 8 Arena pemeliharaan N. longispinosus, T. urticae dan T.
kanzawai pada cawan petri (A) dan pada tanaman ubi kayu (B) 12
9 Arena percobaan uji kemampuan memangsa N. longispinosus 13
10 Rata-rata populasi N. longispinosusdan T. urticaepada (A)
blok pelepasan predator eksotis dan (B) blok tanpa pelepasan
selama 8 minggu pengamatan 18
11 Rata-rata (SD) telur yang diletakkan oleh imago betina N.
longispinosuspada saat diberi mangsa dengan fase berbeda 23
12 Rata-rata (SE) telur yang diletakkan oleh satu imago betina
DAFTAR LAMPIRAN
1 Hasil analisis ragam (ANOVA) yang dilanjutkan dengan uji Tukey pada taraf nyata 95% terhadap kemampuan memangsa
N. longispinosus 31
2 Hasil analisis ragam (ANOVA) yang dilanjutkan dengan uji Tukey pada taraf nyata 95% pengaruh fase mangsa terhadap
peletakan telur N. Longispinosus 33
3 Hasil analisis ragam (ANOVA) yang dilanjutkan dengan uji Tukey pada taraf nyata 95% terhadap potensi kanibalisme N.
longispinosus pada komposisi fase sama 34
4 Hasil analisis ragam (ANOVA) yang dilanjutkan dengan uji Tukey pada taraf nyata 95% terhadap potensi kanibalisme N.
longispinosuspada komposisi fase campuran 36
5 Fase tungau predator N. longispinosus: A. Telur, B. Larva, C.
Protonimfa, D. Deutonimfa, E. Imago betina, F. Imago jantan 37 6 Beberapa perilaku tungau predator N. longispinosus: A.
Predasi, B. Kanibalisme, C. Kopulasi 38 7 Tungau fitofag yang ditemukan : A. Tetranychus kanzawai, B.
Tetranychus urticae 38
8 Predator tungau hama lain yang ditemukan : a. Oligotaspp.
pada ubi kayu, b. Larva Cecidomyiidae pada ubi kayu dan c.
Larva Chrysopidae pada ubi kayu 39
9 Artopoda fitofag lain yang ditemukan : A. Nimfa Empoasca
spp. pada teh, B. Hyposidra talacapada teh, C. Aphisspp. pada
gulma D. Tripsspp. pada stroberi, E. Paracoccus marginatus
PENDAHULUAN
Latar Belakang
Tungau laba-laba, Tetranychus urticae Koch (Acari: Tetranychidae),
merupakan hama penting pada tanaman stroberi yang dibudidayakan di dalam rumah kaca dan di lapangan. Di Florida Amerika Serikat, kehilangan hasil akibat serangan tungau laba-laba pada tanaman stroberi dapat mencapai 50% (Strand 1994). Pengendalian tungau laba-laba tersebut umumnya dilakukan secara kimiawi dengan menggunakan akarisida. Saat ini telah banyak dikembangkan teknik pengendalian hayati untuk mengendalikan hama tersebut, salah satunya dengan menggunakan agens hayati tungau predator famili Phytoseiidae.
Tungau predator dari Famili Phytoseiidae telah diketahui sebagai musuh alami yang sangat penting bagi tungau laba-laba (Gotoh et al.2003). Phytoseiidae
umumnya hidup pada tanaman dan di permukaan tanah. Tungau predator tersebut umumnya memangsa tungau hama, serangga kecil, nematoda, cendawan, dan dapat juga memakan nektar serta eksudat tanaman (Zhang 2003). Tungau predator Famili Phytoseiidae telah dikenal sebagai tungau predator yang efektif dan paling banyak diteliti terkait dengan keefektifannya dalam mengendalikan tungau laba-laba, tungau hama lain dan trips (Gerson et al. 2003). Tungau phytoseiid dapat
bereproduksi sepanjang tahun pada daerah tropis dan subtropis serta di dalam rumah kaca pada daerah temperate (Zhang 2003). Famili Phytoseiidae terbagi dalam tiga subfamili yaitu Amblyseiinae, Typhlodrominae dan Phytoseiinae (Gerson et al. 2003). Spesies komersial yang banyak digunakan di rumah kaca
adalah genus Neoseiulus dan Phytoseiulus yang termasuk dalam subfamili Amblyseiinae dan Phytoseiinae (Zhang 2003).
Phytoseiulus persimilis Athias Henriot (Acari: Phytoseiidae) merupakan
tungau predator yang pertama kali digunakan dalam pengendalian hayati di rumah kaca secara komersial (CABI 2007). P. persimilispertama kali diidentifikasi pada
tanaman mawar yang dibudidayakan pada rumah kaca di Algeria pada tahun 1957 dan umumnya hidup pada iklim Mediterania (Zhang 2003 ; CABI 2007). Sejak itu, P. persimilistelah menyebar ke seluruh dunia dan telah sukses dikembangkan
untuk pengendalian T. urticae pada tanaman mentimun, lada, tomat, stroberi dan
bunga potong di dalam rumah kaca atau di lapangan (Zhang 2003). P. persimilis
sangat aktif bergerak dan memangsa seluruh fase dari T. urticae, terutama fase
telur serta sangat efektif dalam menekan populasi hama tersebut (White at al.
2012). P. persimilis termasuk dalam phytoseiid tipe I yang bersifat spesialis
terhadap mangsa tungau laba-laba khususnya Tetranychus spp. (Gerson et al.
2003).
Program pengendalian T. urticae menggunakan P. persimilisefektif selama
beberapa dekade tanpa ditemukan keberadaan tungau predator di luar rumah kaca dan dampak yang merugikan bagi fauna asli pada iklim dingin di Eropa Barat (Van Lenteran et al. 2006). Hal yang sama juga dinyatakan oleh Gerson et al.
(2003), bahwa P. persimilis dapat mengendalikan tungau laba-laba pada berbagai
seiring dengan menurunnya populasi mangsa sehingga sangat disarankan untuk digunakan dalam program pengendalian augmentatif di rumah kaca.
Sejak tahun 2009, satu spesies tungau predator eksotis, P. persimilis telah
diintroduksi ke Indonesia dari Belanda untuk mengendalikan T. urticae pada
tanaman stroberi di rumah kaca (Santoso 2014, komunikasi pribadi). Tungau predator tersebut telah dilepaskan pada pertanaman stroberi di dalam rumah kaca selama beberapa tahun terakhir hingga saat ini. Pengamatan terhadap kemampuan menetap dan perkembangan populasi tungau predator eksotis perlu dilakukan sebagai salah satu cara untuk mengevaluasi program pengendalian hama.
Sebagai salah satu upaya untuk mengatasi ketergantungan terhadap agens hayati eksotis, perlu dicari alternatif pengendalian dengan memanfaatkan agens hayati lokal. Pada beberapa tanaman budidaya, termasuk stroberi, terdapat beberapa musuh alami atau predator lokal. Salah satunya adalah tungau predator
Neoseiulus longispinosus Evans (Acari: Phytoseiidae). Di Indonesia N. longispinosus pertama kali dikenal dengan nama Thyplodromus longispinosus
(Evans 1952). Di beberapa negara Neoseiulus(Amblyseius) spp. telah digunakan
untuk pengendalian hayati T. urticaepada stroberi dan tanaman budidaya lainnya
(Cross et al. 2001). Untuk itulah perlu dilakukan penelitian mengenai potensi
tungau predator lokal tersebut sebagai agens hayati tungau laba-laba pada tanaman stroberi.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui kemampuan menetap tungau predator eksotis P. persimilis dan potensi tungau predator lokal N. longispinosus
sebagai agens pengendali hayati T. urticaepada tanaman stroberi.
Manfaat Penelitian
Manfaat penelitian ini adalah untuk memberikan informasi mengenai keamanan tungau predator eksotis P. persimilis yang diintroduksi untuk
mengendalikan T. urticae serta potensi pemanfaatan tungau predator lokal N. longispinosus dalam mengendalikan tungau laba-laba pada tanaman stroberi.
TINJAUAN PUSTAKA
Tungau Hama pada Tanaman Stroberi
Stroberi merupakan tanaman buah berupa herba yang ditemukan pertama kali di Chile, Amerika Selatan. Salah satu spesies tanaman stroberi yaitu Fragaria chiloensis menyebar ke berbagai negara di benua Amerika, Eropa dan Asia.
Spesies lain, yaitu F. vesca lebih menyebar luas dibandingkan spesies lainnya.
Jenis stroberi ini pula yang pertama kali masuk dan dikembangkan di Indonesia (Gunawan 1996).
Stroberi memiliki cukup banyak hama dari kelompok Arthopoda, Molusca dan Nematoda (Cross et al. 2001). Terdapat kurang lebih 90 spesies hama pada
stroberi dan 10 spesies hama diantaranya sangat penting secara ekonomi. Hama tersebut diantaranya capsid bugs (Lygus rugulipennis Poppius (Hemiptera:
Miridae), aphids, strawberry blossom weevil (Anthonomus rubi Herbst
(Coleoptera: Curculionidae), vine weevil (Otiorhynchus sulcatus Fabricius
(Coleoptera: Curculionidae), thrips, two-spotted spider mite (Tetranychus urticae
Koch (Acari: Tetranychidae), strawberry mite (Tarsonemus pallidusZimmerman
(Acari: Tarsonematidae) dan slugs (Cross et al. 2001). Sebagian hama bersifat
polifag dan keberadaannya pada tanaman inang biasanya disertai dengan musuh alaminya.
Tetranychus urticae merupakan tungau hama yang sangat penting dan
menyebabkan kerugian cukup besar pada stroberi, seperti yang terjadi pada daerah dimana stroberi dibudidayakan dengan sistem produksi tahunan di California USA (Strand 1994). Menurut Zhang (2003), tungau laba-laba umum ditemui pada tanaman yang dibudidayakan di dalam rumah kaca. Populasi T. urticae yang
tinggi pada daun stroberi dapat mengurangi kapasitas fotosintesis dan transpirasi sehingga menyebabkan menurunnya hasil panen buah. Kehilangan hasil dapat mencapai 25% dan dapat lebih tinggi jika serangan tungau hama dalam populasi tinggi terjadi pada awal musim tanam (Walsh et al. 1998).
Serangan tungau hama ini pada suatu musim juga dapat menurunkan hasil panen pada musim tanam berikutnya. Tungau laba-laba hidup dan makan pada permukaan bawah daun. Pada saat musim panas aktivitas makan dan reproduksinya meningkat secara signifikan. Tungau laba-laba menjadi dorman saat cuaca dingin, dan imago betina berkembang menjadi berwarna kemerahan. Tungau lain yang menyerang stroberi adalah tungau T. pallidusyang biasa disebut strawberry miteatau cyclamen mite(Zhang 2003).
Tetranychus urticaeKoch (Acari: Tetranychidae)
Taksonomi dan Morfologi
Tetranychus urticae termasuk dalam filum Arthropoda, kelas Arachnida,
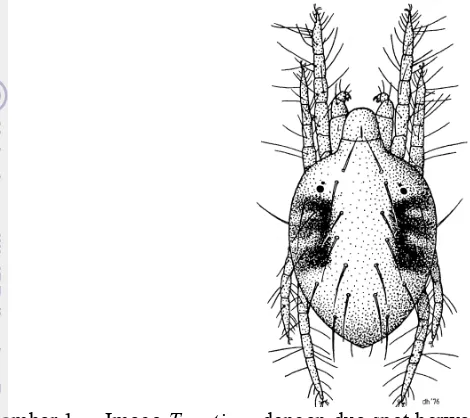
kekuningan dengan bercak berwarna hitam. Imago betina berukuran 400-500 ‚m dan jantan berukuran lebih kecil (Zhang 2003). Tungau betina memiliki bercak berwarna hitam pada bagian dorsal (Gambar 1). Tungkai berwana kekuningan. Tungau jantan berukuran lebih kecil dibanding tungau betina dan berbentuk menyerupai segitiga (Gerson et al. 2003).
Gambar 1 Imago T. urticaedengan dua spot berwarna gelap pada bagian dorsal
idiosoma (Zhang 2003)
Siklus Hidup
Perkembangan hidup terdiri dari telur, larva, protonimfa, deutonimfa dan imago. Lama waktu perkembangan dari telur hingga dewasa semakin menurun dengan meningkatnya suhu dan perkembangan dapat berlangsung kurang dari satu minggu pada saat suhu optimal (30-32ƒC). Pada suhu 15-8 ƒC, perkembangan berlangsung selama 16 hari. Perkembangan tungau jantan lebih cepat dibandingkan tungau betina (Zhang 2003).
Penyebaran
Tungau T. urticaebersifat kosmopolit dan umum berada pada tanaman yang
dibudidayakan di rumah kaca di seluruh dunia. T. urticae merupakan spesies
tungau laba-laba yang bersifat polifag dan dilaporkan telah menyerang lebih dari 150 tanaman inang yang memiliki nilai ekonomi yang tinggi. Tungau ini menyerang lebih dari 300 spesies tanaman di dalam rumah kaca (Zhang 2003).
Pengendalian
Pengendalian hayati T. urticae oleh tungau phytoseiid banyak dilakukan
pada industri rumah kaca di Eropa, sebagian Asia dan Afrika, Australia dan Amerika Utara (Zhang 2003). Spesies yang umum digunakan yaitu P. persimilis, P. micropilis, Neoseiulus californicus, N. fallacis, N. longispinosus dan Galendromus occidentalis (Zhang 2003). Pengendalian kimia tungau laba-laba
pada beberapa negara yang berbeda. Pengendalian secara kultur teknis dan fisik juga dapat digunakan untuk mengatasi tungau ini (Zhang 2003).
Pengendalian kimia tungau laba-laba perlu untuk dipertimbangkan akibat perkembangan resistensi yang cepat dan penurunan jumlah akarisida yang terdaftar. Bahan kimia dan metode pengendaliannya sangat tergantung dari tingkat resistensi tungau, kondisi tanaman dan lingkungan, serta ketersediaan akarisida di negara yang berbeda (Zhang 2003).
Phytoseiulus persimilisAthias Henriot (Acari: Phytoseiidae)
Taksonomi dan Morfologi
Phytoseiulus persimilistermasuk dalam filum Arthropoda, kelas Arachnida,
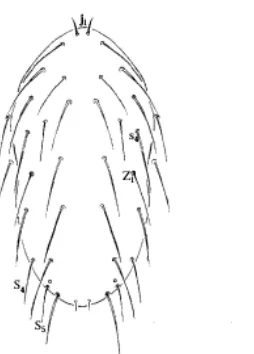
ordo Mesostigmata, dan famili Phytoseiidae. Imago berwarna orange cerah dengan bentuk seperti buah pir dan memiliki tungkai panjang. Panjang imago 0.5 mm. Telur berbentuk oval dengan panjang 0.3 mm. Betina dapat memproduksi telur sebanyak 5 telur per hari. Imago pada bagian dorsal memiliki 23 pasang seta dengan seta median j4–j6 dan s4 berukuran panjang (Gerson et al. 2003) (Gambar
2). Stigmata terletak diantara koksa ke III dan ke IV (Gerson et al. 2003).
Pada kondisi suhu, kelembapan, dan ketersediaan makanan yang optimal P. persimilis dapat menghasilkan 60 telur selama hidupnya (CABI 2007). Apabila
tingkat ketersediaan mangsa tercukupi, predator dapat menghasilkan 78 telur selama 15 hari pada suhu 26 ƒC dan kelembapan nisbi 80% (Gerson et al. 2003).
Gambar 2 Bagian dorsal P. persimilis yang memiliki 23 pasang seta dengan
seta median j4–j6 dan s4 berukuran panjang (Gerson et al. 2003)
Siklus Hidup
Siklus hidup P. persimilisterdiri dari telur, larva, nimfa dan imago (Gambar
2007). Zhang (2003) menyatakan bahwa perkembangan P. persimilis dari telur
hingga dewasa selama 3.6 hari untuk jantan dan 4.1 hari untuk betina pada suhu 26 ⁰C. Pada rumah kaca dengan suhu berfluktuasi diantara 11.5 dan 23.5 ⁰C dengan kelembapan nisbi berkisar 36-95%, perkembangan dari telur hingga dewasa berlangsung selama 11 hari, sementara masa praoviposisi betina selama 2.6 hari (Zhang 2003).
Gambar 3 Siklus hidup tungau predator P. persimilis(CABI 2007)
Penyebaran
Phytoseiulus persimilis merupakan tungau predator dengan kemampuan
memangsa yang tinggi, hidup terutama pada daerah Mediterania (Zhang 2003). P. persimilis pertama kali masuk secara tidak sengaja ke Jerman dari Cile melalui
importasi tanaman anggrek pada tahun 1958. Sejak itu tungau predator tersebut telah menyebar ke seluruh dunia dan telah sukses dikembangkan untuk pengendalian tungau laba-laba pada berbagai tanaman budidaya baik di lapangan dan di rumah kaca (Gerson et al. 2003; Zhang 2003; CABI 2007).
Keefektifan Pemangsaan
Phytoseiulus persimilis termasuk dalam tungau phytoseiid tipe I yang
bersifat spesialis terhadap mangsa tungau laba-laba khususnya Tetranychus spp.
(Gerson et al. 2003). Fase larva P. persimilis tidak makan, fase nimfa
mengkonsumsi telur dan protonimfa tungau laba-laba sedangkan fase imago mengkonsumsi seluruh fase dari mangsa. Perkembangan yang cepat dari P. persimilismerupakan hal yang sangat efektif dalam pemangsaan tungau laba-laba.
Tungau predator tersebut memiliki reproduksi yang lebih cepat dibandingkan dengan tungau hama. Imago berpindah dari satu tanaman ke tanaman lainnya melalui kontak fisik antara tanaman dan di sepanjang jaring tungau laba. Imago betina dapat mengkonsumsi 5 ekor imago tungau laba-laba atau 20 ekor larva dan telur dalam sehari. Pada kondisi suhu ideal (15-20 ⁰C) dan kelembapan nisbi (60-70%), pemangsaan berlangsung optimal (CABI 2007).
Telur Larva
Protonimfa
Deutonimfa
Neoseiulus longispinosusEvans (Acari: Phytoseiidae)
Taksonomi dan Morfologi
Tungau predator N. longispinosus termasuk dalam famili Phytoseiidae, ordo
Mesostigmata. N. longispinosus sangat berhubungan dekat dengan N. womersleyi secara biosistematika (Zhang 2003; Gerson et al. 2003). Telur
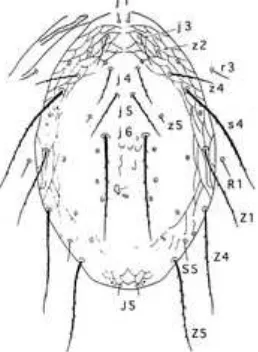
berbentuk oval dan transparan. Pradewasa transparan sementara tungau dewasa berwarna kuning. Imago betina berukuran 350 ‚m. Bagian dorsal memiliki 17 pasang seta. Kecuali seta j1 dan s5, sebagian seta dorsal berukuran panjang dan berakhir pada seta selanjutnya (Zhang 2003) (Gambar 4).
Gambar 4 Bagian dorsal N. longispinosusdengan 17 pasang seta, sebagian seta
dorsal berukuran panjang kecuali seta j1 dan s5 (Zhang 2003)
Siklus Hidup
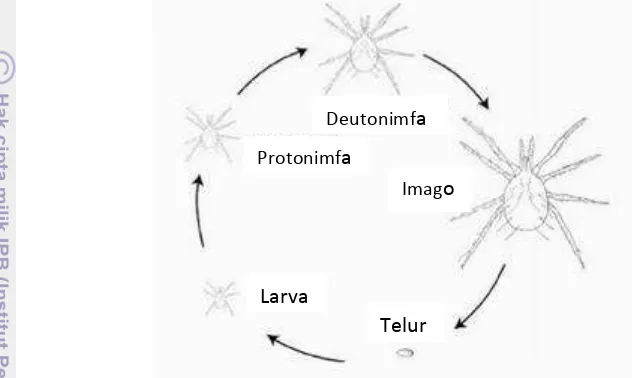
Siklus hidup N. longispinosus terdiri dari telur, larva, protonimfa,
deutonimfa dan imago. Perkembangan fase pradewasa berlangsung selama 5 hari pada suhu 28 ƒC (Zhang 2003). Fase telur berlangsung selama 1-2 hari (Puspitarini 2005). Masa fase larva pada umumnya relatif singkat dan biasanya hanya dalam hitungan jam. Lama fase nimfa biasanya berlangsung selama satu hari. Lama hidup tungau jantan lebih pendek dibandingkan tungau betina (Zhang 2003).
Penyebaran
Tungau predator N. longispinosus berasal dari Rusia, China dan Jepang
menyebar ke Asia Tenggara menuju Australia dan Hawaii (Zhang 2003). Gerson
et al. (2003) menyatakan bahwa N. longispinosuslebih banyak ditemukan ke arah
selatan yaitu di India, Cina bagian selatan, Taiwan, Filiphina, Malaysia, Indonesia dan New Zealand.
Kefektifan Pemangsaan
Imago N. longispinosus dapat memangsa 26.33 telur per hari dan
meletakkan 2.92 telur per hari. Imago betina dapat meletakan telur sebanyak 27.5 telur selama 16.1 hari masa oviposisi, memangsa 11.7 larva, 9.3 nimfa atau 5.1 imago broad miteper hari. Fase larva N. longispinosusmemangsa 3.8 larva dan
1.4 nimfa. Sementara protonimfa memangsa 9.2 larva, 7.9 nimfa dan 3.2 imago (Zhang 2003). N. longispinosus berasosiasi dan memangsa tungau yang
menginfestasi bambu di China dan efisien dalam mengendalikan tungau hama tersebut pada suhu tinggi (30-35 ÄC) (Zhang et al. 1999).
Pengendalian Hayati
Agens hayati adalah setiap organisme yang meliputi: spesies, sub spesies, varietas semua jenis serangga, nematoda, protozoa, cendawan, bakteri, virus,
mikoplasma serta organisme lainnya dalam semua tahap perkembangannya yang dapat dipergunakan untuk keperluan pengendalian hama dan penyakit/organisme pengganggu, proses produksi, pengolahan hasil pertanian dan berbagai keperluan lainnya. (Kepmentan Nomor 411 Tahun 1995). Agens hayati memiliki beberapa keunggulan dibandingkan bila dibandingkan dengan sistem pengendalian lainnya, yaitu: selektivitas tinggi, ramah lingkungan, tersedia di alam, mampu berkembang serta menyebar sendiri atau dengan bantuan perantara, dan tidak menyebabkan resistensi hama (Sosromarsono & Untung 2000). Pada dasarnya agens hayati di bagi ke dalam 4 kelompok besar yaitu agens hayati predator, agens hayati parasitoid, patogen serangga (entomopatogen) dan agens hayati antagonis.
Huffaker et al. (1971) menyebutkan bahwa empat karakteristik utama yang
berkaitan dengan efesiensi dari parasit, predator, dan agens yang memiliki keunggulan tergantung pada a) kemampuan adaptasi terhadap berbagai kondisi lingkungan fisik, b) kapasitas pencarian mangsa dan pergerakannya, c) kemampuan peningkatan relatif dengan mangsa dan kemampuan memangsa, d) faktor dalam lainnya seperti, kesesuaian inang, spesifikasi inang, kemampuan membedakan, kemampuan untuk bertahan pada inang alternatif dan kebiasaan khusus yang membentuk hubungan antara kerapatan mangsa dengan persebaran pada inang dan populasinya sendiri.
Agens hayati untuk berbagai keperluan tersebut seringkali harus diintroduksi dari luar negeri karena agen hayati yang diperlukan belum tersedia di Indonesia atau belum tersedia dalam jumlah yang mencukupi keperluan. Teknik pengendalian hayati dengan menggunakan parasitoid dan predator dapat dikelompokkan dalam 3 kategori yaitu, konservasi, introduksi, dan augmentasi. Meskipun ketiga teknik pengendalian hayati tersebut berbeda tetapi dalam pelaksanaannya sering digunakan secara bersamaan (Huffaker et al.1971).
dengan otorisasi pemasukan agens hayati, baik dalam bentuk yang telah diformulasi maupun yang belum diformulasi, ke dalam wilayah Indonesia akan efektif, tidak berdampak negatif terhadap kesehatan manusia dan hewan, dan aman terhadap lingkungan. Izin pemasukan agens hayati dapat diberikan dalam bentuk keputusan Menteri Pertanian, sedangkan penolakan permohonan izin pemasukan agen hayati diberikan dalam bentuk surat penolakan. Izin hanya berlaku untuk jangka waktu enam bulan sejak tanggal penerbitannya dan dapat dipergunakan dalam satu kali pemasukan (Kepmentan No 411 Tahun 1995).
BAHAN DAN METODE
Tempat dan Waktu
Penelitian dilaksanakan di rumah kaca PT Strawberindo Lestari, Cianjur, Jawa Barat dan Laboratorium Bionomi dan Ekologi Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor dari bulan September 2014 sampai dengan Februari 2015.
Bahan dan Alat
Bahan dan alat yang digunakan antara lain sampel daun stroberi, sampel tanaman lain (rapsberi, teh, ubi kayu dan gulma), kapas, busa, polybag ukuran 30 cm x 30 cm, cawan petri plastik, kotak plastik berukuran 35 cm x 27.5 cm x 6 cm, mikroskop stereo, mikroskop kompon, objek glass, lampu penerang, kantung plastik, kompas, kuas halus, perangkap (sticky trap), kaca pembesar, alat
penghitung (counter) dan alat bantu lainnya.
Metode Penelitian
Populasi Tungau Hama dan Tungau Predator di Dalam Rumah Kaca
Pelepasan tungau predator eksotis P. persimilisdilakukan terakhir kali pada
bulan April 2014. Pelepasan dilakukan pada beberapa blok di dalam rumah kaca (Gambar 5). Pengamatan populasi P. persimilis dan T. urticae dilakukan pada
blok tanaman stroberi di dalam rumah kaca yang dilepaskan dengan P. persimilis
dan blok tanpa pelepasan. Pada setiap blok pengamatan ditetapkan sebanyak 30 tanaman contoh secara acak sistematis. Pada setiap tanaman contoh diambil sebanyak 3 daun. Pengambilan sampel daun dilakukan setiap 1 minggu selama 8 minggu. Sampel dibawa ke laboratorium untuk diamati dan dihitung populasi tungau hama T. urticae, tungau predator eksotis P. persimilisdan tungau predator
lokal.
Keberadaan Tungau Predator Eksotis di Luar Rumah Kaca
Pengamatan dilakukan untuk mengetahui keberadaan, kemampuan menetap dan peluang penyebaran tungau predator eksotis di luar lingkungan rumah kaca. Agens hayati diketahui dapat menyebar secara aktif dan pasif. Penyebaran secara aktif dengan berjalan melewati jaring kasa rumah kaca. Penyebaran secara pasif agens hayati dapat melalui lalu lintas pekerja, alat, produk panen buah stroberi atau bagian tanaman lainnya pada saat pemeliharaan dan proses pemanenan di dalam rumah kaca tersebut.
Pengambilan sampel tanaman dilakukan di dua tempat. Pengambilan sampel pertama dilakukan terhadap tanaman yang berada di sekitar rumah kaca. Pengambilan sampel dilakukan secara purposive sampling. Tanaman yang
diambil sampelnya dapat berupa tanaman rapsberi, gulma atau tanaman lainnya. Pengambilan sampel tanaman di sekitar rumah kaca dilakukan mengikuti empat arah mata angin dengan jarak 5, 15 dan 30 meter dari rumah kaca (Gambar 6). Pada setiap arah mata angin diambil sebanyak tiga sampel tanaman. Pengambilan sampel dilakukan sebanyak tiga kali pada setiap lokasi. Sampel tanaman selanjutnya dibawa ke laboratorium untuk diamati, dihitung dan diidentifikasi terhadap artopod yang ditemukan.
Gambar 6 Titik pengambilan sampel di sekitar rumah kaca
Pengambilan sampel selanjutnya dilakukan terhadap tanaman di sekitar perkebunan PT Strawberindo Lestari dilakukan pada jarak kurang lebih 200, 500, 1000 dan 2000 meter dari rumah kaca mengikuti arah jalan (Gambar 7). Pengambilan sampel dilakukan secara purposive sampling terhadap tanaman yang
berada di sekitar perkebunan stroberi. Sampel dapat berupa tanaman teh, ubi kayu, gulma dan tanaman lainnya. Pada setiap titik diambil sebanyak tiga sampel tanaman. Sampel tanaman selanjutnya dibawa ke laboratorium untuk diamati, dihitung dan diidentifikasi terhadap arthopod yang ditemukan. Pengambilan sampel tanaman dilakukan sebanyak tiga kali pada setiap lokasi.
Gambar 7 Titik pengambilan sampel di sekitar perkebunan stroberi
Tingkat Pemangsaan N. longispinosus
Pengujian bertujuan untuk mengetahui potensi pemangsaan tungau predator lokal N. longispinosus terhadap beberapa tungau hama. Dalam pengujian ini,
digunakan tungau hama T. urticae dan T. kanzawai Kishida (Acari:
Tetranychidae) sebagai mangsa. Rata-rata suhu harian dan kelembapan nisbi di dalam laboratorium adalah 29 †C (27-31 †C) dan 59.9% (43-79%).
Pemeliharaan N. longispinosus, T. urticae danT. kanzawai. Tungau hama
diambil dari pertanaman stroberi dan ubi kayu yang berada di sekitar perkebunan. Pemeliharaan dilakukan pada arena pemeliharaan cawan petri plastik (d = 6 cm) yang didalamnya diletakkan busa (d = 5.5 cm) (Gambar 8A). Di atas busa diletakkan kapas berukuran 5 cm x 4 cm. Busa dan kapas dijaga agar selalu basah dengan diberi air setiap hari. Di atas kapas diletakkan daun ubi kayu atau daun stroberi dengan luasan yang tidak melebihi luasan kapas dan dilakukan penggantian daun setiap tiga hari. Pemeliharaan T. urticae dan T. kanzawai juga
dilakukan pada tanaman ubi kayu pada pot plastik yang diletakkan di dalam kotak kaca (Gambar 8B).
Gambar 8 Arena pemeliharaan N. longispinosus, T. urticae dan T. kanzawai
pada cawan petri (A) dan pada tanaman ubi kayu (B)
A B
Tungau predator N. longispinosus didapatkan dari dalam rumah kaca.
Tungau predator N. longispinosusdipelihara dengan mangsa T. kanzawai dan T. urticaepada daun ubi kayu atau stroberi. Tungau hama T.kanzawai dan T. urticae
didapatkan dari pembiakan masal di laboratorium. Pada tiap cawan petri ditempatkan selembar daun ubi kayu atau stroberi. Tungau predator diinfestasikan pada daun yang telah berisi mangsa T. kanzawai dan T. urticae tersebut.
Penggantian daun dilakukan setiap tiga hari.
Kemampuan memangsa N. longispinosus terhadap T. urticae dan T. kanzawai. Predator yang diuji adalah fase deutonimfa dan imago betina N. longispinosus. Mangsa yang digunakan adalah fase telur dan imago betina T. urticaedan T.kanzawai. Pada arena percobaan diletakkan masing-masing 30 butir
telur atau 10 ekor imago betina T. urticae dan T. kanzawai. Penyediaan mangsa
telur dilakukan dengan cara meletakkan 10-15 imago betina T. urticae atau T. kanzawai pada daun stroberi berukuran 2 cm x 2 cm dalam cawan petri dan
dibiarkan untuk meletakkan telur selama 24 jam, kemudian imago betina tungau disingkirkan. Pada arena percobaan berukuran 2 cm x 2 cm diletakkan masing-masing seekor deutonimfa atau imago betina N. longispinosus yang telah
dilaparkan selama 6 jam (Gambar 8). Setelah 24 jam predator disingkirkan dan banyaknya telur dan imago betina T. urticae atau T. kanzawai yang dimangsa
dihitung. Banyaknya imago betina yang dimangsa didasarkan dari jumlah bangkai yang dijumpai, sedangkan banyaknya telur yang dimangsa dihitung dari jumlah telur awal dikurangi dengan jumlah telur yang menetas menjadi larva. Pengujian diulang sebanyak lima kali.
Pengaruh Fase Mangsa terhadap Tingkat Oviposisi N. longispinosus.
Pengamatan dilakukan pada percobaan kemampuan pemangsaan imago betina N. longispinosus terhadap fase telur dan imago betina T. urticae dan T. kanzawai.
Jumlah telur N. longispinosusyang diletakkan dihitung setelah 24 jam.
Gambar 9 Arena percobaan uji kemampuan memangsa N. longispinosus
Potensi Kanibalisme N. longispinosus
Pengujian potensi kanibalisme terdiri dari dua tahap. Pengujian pertama dilakukan untuk mengetahui kanibalisme tungau predator pada komposisi mangsa fase yang sama. Fase predator yang digunakan adalah deutonimfa dan imago betina N. longispinosusdengan kerapatan 2, 4, 6, dan 8 ekor predator. Deutonimfa
jam setelah perlakuan kemudian dengan mencatat jumlah predator yang dimangsa konspesifik. Jumlah telur predator yang diletakkan dihitung 24 jam setelah perlakuan. Pengujian diulang sebanyak lima kali.
Pengujian selanjutnya dilakukan untuk mengetahui tingkat kanibalisme tungau predator pada komposisi fase campuran. Kerapatan predator yang diuji yaitu 2, 4, 6, dan 8 ekor predator. Deutonimfa dan imago betina dengan perbandingan 1:1 diletakkan pada arena yang sama dengan pengujian kemampuan memangsa. Predator dilaparkan terlebih dahulu selama 6 jam sebelum perlakuan. Pengamatan dilakukan 24 jam setelah perlakuan dengan mencatat jumlah predator yang dimangsa konspesifik. Seperti halnya pada pengujian komposisi fase yang sama, jumlah telur predator yang diletakkan juga dihitung setelah 24 jam. Pengujian diulang sebanyak lima kali.
Analisis Data
Data pengamatan populasi tungau predator dan tungau hama di dalam dan di luar rumah kaca diolah dengan program Ms Excel 2007. Pengujian kemampuan memangsa menggunakan rancangan acak lengkap (RAL) faktorial dengan tiga faktor yaitu fase tungau predator, spesies tungau hama dan fase tungau hama, Pengujian potensi kanibalisme menggunakan rancangan acak lengkap (RAL). Data hasil pengamatan diolah menggunakan ANOVA dengan program Minitab 16 dilanjutkan dengan Uji Tukey α = 0.05.
HASIL DAN PEMBAHASAN
Keberadaan Tungau Predator Eksotis di dalam dan di luar Rumah Kaca
Hasil pengamatan menunjukkan tungau predator eksotis P. persimilis tidak
ditemukan di dalam rumah kaca. Berdasarkan informasi yang diperoleh pelepasan tungau predator P. persimilis di dalam rumah kaca, dilakukan terakhir kali pada
bulan April tahun 2014. Rentang waktu yang cukup lama antara waktu pelepasan dengan waktu pengamatan ( 7 bulan), menyebabkan populasi P. persimilis
menurun secara drastis sehingga tidak dapat bertahan. Biologi dan karakteristik P. persimilis merupakan faktor penting yang juga berpengaruh terhadap
perkembangan populasinya.
P. persimilis termasuk dalam phytoseiid tipe I yang bersifat spesialis
terhadap mangsa tungau laba-laba khususnya Tetranychus spp. (Gerson et al.
2003; Zhang 2003). Tungau predator spesialis ini tidak mengkonsumsi makanan tambahan lain seperti nektar, tepung sari atau eksudat tanaman seperti halnya tungau predator generalis. Hal ini menyebabkan perkembangan populasi P. persimilis sangat bergantung pada perkembangan populasi mangsanya, sehingga
saat populasi mangsa menurun maka populasi P. persimilisjuga menurun.
Hal tersebut merupakan salah satu keuntungan dari karakteristik agens hayati yang digunakan untuk pengendalian secara augmentatif. Dengan karakteristik tersebut, tungau predator eksotis tidak berpotensi untuk menjadi spesies invasif. Gerson et al. (2003) menyatakan bahwa, populasi P. persimilis
akan menurun seiring dengan menurunnya jumlah populasi mangsa sehingga sangat disarankan untuk digunakan dalam program pengendalian secara augmentatif di dalam rumah kaca.
Tungau predator P. persimilis merupakan tungau predator yang telah
dikenal luas dan umum digunakan untuk pengendalian hayati di rumah kaca karena karakteristiknya. Tungau predator tersebut memiliki kemampuan untuk menekan populasi tungau hama dengan cepat terutama apabila dilepas saat populasi hama tinggi, namun populasinya akan cepat menurun saat populasi hama rendah (Cross et al. 2001). Penggunaan tungau predator P. persimilis terlihat
seperti pestisida sehingga sering disebut dengan living pesticideatau biopesticide
karena dapat memberikan pengaruh yang sangat cepat tetapi hanya memiliki efek jangka pendek atau short term effect (Gerson et al. 2003). Menurut Cross et al.
(2003), P. persimilis tidak dapat bertahan pada musim dingin sehingga harus
dilepaskan pada setiap musim tanam. Untuk itulah pelepasannya harus dilakukan secara periodik untuk menjamin keberhasilan program pengendalian hayati tungau laba-laba di dalam rumah kaca.
Menurut petunjuk penggunaan dari produsen agens hayati tersebut, pengaplikasian atau pelepasan P. persimilis harus dilakukan secara berkala atau
periodik. Hal tersebut menjadi salah satu kelemahan karena program pengendalian hama menjadi kurang efisien ditinjau dari segi teknis dan ekonomis. Kemungkinan faktor lain yang menyebabkan tidak ditemukannya P. persimilis
yaitu, kemungkinan adanya kompetisi interspesies dengan predator lokal salah satunya N. longispinosus atau artopod predator lainnya yang turut mempengaruhi
Pengamatan terhadap keberadaan tungau predator eksotis P. persimilis di
luar rumah kaca dilakukan pada lingkungan di sekitar rumah kaca dan perkebunan stroberi. Pengamatan dilakukan pada tanaman rapsberi, teh, ubi kayu dan berbagai jenis gulma seperti Ageratum conyzoides (L.) (Asterales: Asteraceae), Chromolaena odorata (L.) (Asterales: Asteraceae), Axonopus compressus (Sw.)
(Poales: Poaceae), Boreria alata (Aubl.) (Gentianales: Rubiaceae), Digitaria adsendence (L.) (Poales: Poaceae), dan Paspalum conjugatum (L.) (Poales:
Poaceae).
Kelompok Artopoda fitofag yang ditemukan yaitu Aphis spp. (Hemiptera:
Aphididae), Thrips spp. (Thysanoptera: Thripidae), Frankliniela intonsa
(Thysanoptera: Thripidae), Hyposidra talaca (Lepidoptera: Geometridae), Olygonychus coffeae, T. kanzawai, T. urticae (Acari: Tetranychidae). Kelompok
Artopoda predator yang ditemukan yaitu N. longispinosus(Acari: Phytoseiidae), Sterorus spp. (Coccinelidae: Coleoptera), larva Cecidomyiid (Diptera:
Cecidomyidae), larva Chrysopid (Neuroptera: Chrysopidae), Formicid (Hymenoptera: Formicidae), dan Oligotaspp. (Coleoptera: Staphylinidae) (Tabel
1). Beberapa diantara artopod predator tersebut, juga berperan sebagai predator tungau laba-laba yaitu N. longispinosus, larva Cecidomyiid, larva Chrysopid, dan Oligotaspp.
Neoseiulus longispinosusAcari Phytoseiidae √ - √
-Oligotaspp. Coleoptera Staphylinidae - - √
-Sterorusspp. Coleoptera Coccinelidae √ - √ √
Cecidomyid Diptera Cecidomyidae - - √ -Formicid Hymenoptera Formicidae √ √ √ √ Chrysopid Neuroptera Chrysopidae - - √ -B. Fitofag
Tetranychus kanzawai Acari Tetranychidae - - √
-Tetranychus urticae Acari Tetranychidae √ √ -
-Olygonychus coffeae Acari Tetranychidae - √ -
-Aphis spp. Hemiptera Aphididae √ √ - √
Trips spp. Thysanoptera Thripidae √ - √
-Frankliniela intonsa Thysanoptera Thripidae √ - -
-Hyposidra talaca Lepidoptera Geometridae - √ - √
Tungau predator yang ditemukan pada sampel tanaman merupakan tungau predator lokal yaitu N. longispinosus. Tungau predator ini banyak ditemukan pada
tanaman ubi kayu dan rapsberi di sekitar perkebunan stroberi. Pada tanaman ubi kayu tungau ditemukan berasosiasi dengan T. kanzawaisedangkan pada tanaman
rapsberi ditemukan berasosiasi dengan T. urticae. Tungau hama T. kanzawai
sangat intens ditemukan pada tanaman ubi kayu dengan populasi yang cukup tinggi. Sementara T. urticae ditemukan pada stroberi dan rapsberi. Menurut
Santoso (2015, komunikasi pribadi) tungau hama T. urticae umumnya berada
pada tanaman yang dibudidayakan di dataran tinggi seperti krisan, mawar, dan stroberi, sementara T. kanzawai cenderung ditemukan pada tanaman budidaya
dataran rendah seperti ubi kayu. N. longispinosus banyak ditemukan berasosiasi
dengan T. kanzawaipada tanaman ubi kayu, hal ini dapat dijadikan pertimbangan
untuk melakukan pembiakan masal atau rearing N. longispinosus menggunakan T. kanzawaisebagai alternatif mangsa selain T. urticae.
Agens hayati memiliki peluang untuk keluar dari lingkungan pengujian. Salah satunya karena faktor kemampuan pemencaran dari agens hayati tersebut. Pemencaran agens hayati dapat terjadi secara aktif dan pasif. Menurut Jung (2005) tungau predator phytoseiid memiliki 2 mode penyebaran yaitu ambulatory dispersaldan aerial dispersal. Hasil penelitian Jung (2005) menunjukkan bahwa,
tungau predator spesialis P. persimilis memiliki kemampuan bergerak (walking activity) dan aerial dispersal yang lebih tinggi dibandingkan tungau predator
generalis Neoseiulus fallacis dan N. californicus. Menurut Hoy (2011) tungau
predator P. persimilis memiliki tungkai yang panjang dan sangat aktif bergerak
dan diketahui dapat bergerak (walking) dari daun ke daun, tanaman ke tanaman,
dan dapat bergerak beberapa meter diatas tanah dari tanaman budidaya ke gulma atau sebaliknya. Beberapa tungau phytoseiid dan tetranychid juga memiliki kemampuan aerial dispersal. Hasil penelitian Hoy (2011) menunjukkan bahwa, Metaseilus occidentalis (Acari: Phytoseiidae) dan tungau laba-laba ditemukan
terperangkap pada perangkap lekat (stick trap) yang berjarak 200 meter dari
pertanaman anggrek. Untuk alasan inilah keamanan dari rumah kaca sangat diperlukan terutama jika digunakan untuk lingkungan pengujian agens hayati eksotis.
Keamanan lingkungan pengujian merupakan hal penting dalam pengendalian hayati secara augmentatif di dalam rumah kaca. Hal ini bertujuan untuk mencegah agens hayati eksotis keluar dari lingkungan pengujian menjadi spesies invasif. Tidak ditemukannya P. persimilis di luar rumah kaca
menunjukkan bahwa rumah kaca tersebut memiliki tingkat keamanan yang cukup baik dalam mencegah keluarnya tungau predator dari lingkungan pengujian. Menurut Van Lenteran et al. (2006), program pengendalian T. urticae
menggunakan P. persimilis efektif selama beberapa dekade tanpa ditemukan
keberadaan tungau predator tersebut di luar rumah kaca dan dampak yang merugikan bagi fauna asli pada iklim dingin di Eropa Barat. Hal yang sama juga diungkapkan oleh Gerson et al. (2003), bahwa tungau predator P. persimilisdapat
mengendalikan tungau laba-laba pada berbagai tanaman pada daerah temperate dan subtropis, serta tidak dapat menetap pada flora asli.
Di beberapa negara terdapat beberapa kasus yang dilaporkan tentang kemampuan menetap tungau predator eksotik, namun hal tersebut sangat jarang terjadi. Walter et al. (2006) melaporkan bahwa, P. persimilis, A. californicusdan M. occidentalis dapat menetap pada beberapa lokasi setelah pelepasan predator
secara augmentatif. Namun demikian, walaupun P. persimilis dapat menetap di
Australia, tetapi predator tersebut tidak dapat masuk lebih jauh ke dalam hutan hujan (Walter et al. 2006).
Perkembangan Populasi Tungau Predator Lokal dan Tungau Hama di dalam Rumah Kaca
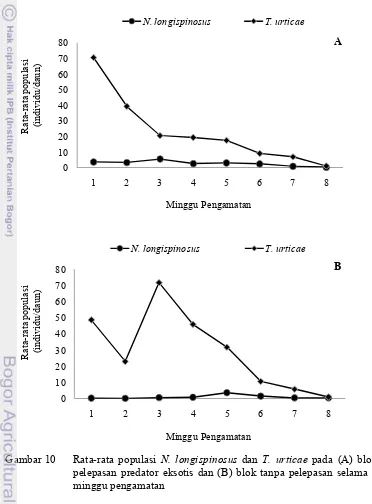
Hasil pengamatan menunjukkan tidak ditemukan tungau predator eksotis P. persimilis di dalam rumah kaca dan hanya ditemukan tungau predator lokal N. longispinosus. Gambar 10 menunjukkan perkembangan populasi tungau predator
lokal N. longispinosusdan tungau hama T. urticae pada blok pelepasan tungau
predator eksotis dan blok tanpa pelepasan selama 8 minggu pengamatan.
Gambar 10 Rata-rata populasi N. longispinosus dan T. urticae pada (A) blok
Pada blok pelepasan predator eksotis, populasi N. longispinosus cenderung
semakin menurun hingga pengamatan minggu terakhir (Gambar 10A). Rata-rata populasi N. longispinosus tertinggi pada minggu ke-3 yaitu sebanyak 5.47
individu per daun dan terendah pada minggu ke-8 sebanyak 0.23 individu per daun (Gambar 10A). Rata-rata N. longispinosus semakin meningkat hingga
minggu ke-5 dan merupakan populasi tertinggi yaitu sebanyak 3.65 individu per daun. Populasi N. longispinosus terendah pada minggu ke-2 sebanyak 0.10
individu per daun (Gambar 10B). Populasi N. longispinosus kemudian menurun
hingga minggu terakhir pengamatan (Gambar 10B).
Tungau predator N. longispinosus termasuk dalam phytoseiid tipe II yang
memangsa tidak hanya tungau laba-laba Tetranychusspp. tetapi juga spesies lain
yang menghasilkan sedikit jaring dan mengkonsumsi nektar serta eksudat tanaman (Gerson et al. 2003). Hal ini diduga sebagai penyebab N. longispinosus
dapat bertahan dengan baik karena mampu bertahan pada rentang makanan yang bervariasi. Sebagaimana yang dinyatakan oleh Cross et al. (2001) bahwa Neoseiulusspp. memiliki kemampuan untuk bertahan pada rentang makanan yang
bervariasi sehingga mampu menjaga keseimbangan antara predator dan mangsanya dalam jangka waktu yang cukup lama dan beberapa spesies juga dapat mengendalikan tungau tarsonemid. N. longispinosus juga merupakan spesies asli
atau lokal sehingga telah beradaptasi dengan lingkungannya.
Strand (1994) menyatakan bahwa Neoseiulus californicusMcGregor (Acari:
Phytoseiidae) dapat bertahan dengan memakan nektar tetapi tidak dapat berkembang biak jika tidak ada tungau laba-laba sehingga jumlahnya tidak menurun drastis seperti P. persimilis ketika populasi tungau hama berkurang.
Menurut Cross et al. (2001) tungau predator P. persimilis tetap menjadi predator
yang efektif untuk mengendalikan T. urticae khususnya untuk stroberi yang
dibudidayakan di dalam naungan yang terlindungi. Sehingga untuk pengendalian secara augmentatif agens hayati yang bersifat spesialis akan lebih cocok dibandingkan predator yang bersifat generalis. Sementara phytoseiid lainnya seperti Neoseiulus spp. menjadi alternatif terbaik untuk mengendalikan tungau
laba-laba pada beberapa kondisi tertentu.
N. longispinosus dianggap cukup berpotensi untuk dikembangkan sebagai
agens pengendali hayati T. urticae. Salah satu caranya dengan melakukan teknik
konservasi terhadap tungau predator lokal tersebut. Teknik konservasi merupakan
salah satu teknik pengendalian hayati dengan memanfaatkan keberadaan agens hayati lokal. Teknik konservasi dapat dilakukan dengan cara memodifikasi lingkungan yang sesuai untuk perkembangan tungau predator lokal N. longispinousus. Salah satunya dengan pemeliharaan jenis gulma yang berbunga
sebagai pakan tambahan untuk N. longispinosus. Pengembangan tungau predator
lokal N. longispinosus memiliki keuntungan yaitu lebih efisien dan ekonomis
karena tidak perlu melakukan importasi. Pemeliharaan atau rearing untuk
mencapai kuantitas produksi yang cukup digunakan untuk pengendalian hayati tungau laba-laba menjadi tantangan tersendiri.
Menurut Alatawi et al. (2011), dalam program pengendalian hayati secara
mangsa sebelum predator eksotis dilepaskan menjadi sangat penting untuk keberhasilan program pengendalian hama secara augmentatif.
Efektifitas dari tungau predator dalam menekan populasi tungau hama diduga dipengaruhi oleh musim. Hasil penelitian Mori & Saito (1979), menyatakan bahwa terdapat perbedaan tingkat penekanan terhadap populasi T. urticae oleh P. persimilis dan Amblyseius longispinosuspada musim panas dan
musim gugur di dalam rumah kaca. Tingkat penekanan populasi T. urticaepada
musim gugur oleh 2 spesies phytoseiid tersebut lebih rendah dibandingkan pada musim panas dan diketahui bahwa P. persimilisberperan lebih efektif dibanding A. longispinosuspada musim gugur (Mori & Saito 1979). Hasil penelitian Mori &
Saito (1979) juga menunjukkan bahwa kepadatan populasi T. urticae tertekan
pada tingkat yang rendah oleh P. persimilis dan A. longispinosus yang
diintroduksikan secara berkala. Hal tersebut diduga disebabkan oleh pengaruh musim yang berimplikasi terhadap dinamika populasi tungau hama dan musuh alaminya. Dengan adanya perkembangan populasi tungau hama akibat pengaruh musim menyebabkan populasi tungau predator ikut terpengaruh.
Pada blok pelepasan predator eksotis, rata-rata populasi T. urticae
cenderung semakin menurun hingga pengamatan minggu terakhir (Gambar 10A). Populasi T. urticaetertinggi pada minggu ke-1 sebanyak 70.53 individu per daun,
sementara populasi terendah pada minggu ke-8 sebanyak 6.83 individu per daun. Menurunnya populasi T. urticaediduga dipengaruhi oleh semakin meningkatnya
umur tanaman dan musim. Pada blok yang dilepaskan dengan tungau predator eksotis P. persimilis umur tanaman stroberi mencapai ‰ 10 bulan. Umumnya
pergantian tanaman stroberi dilakukan pada saat umur tanaman mencapai 1 tahun atau lebih. Populasi T. urticaeakan menurun seiring dengan meningkatnya umur
tanaman. Faktor lain yang mempengaruhi adalah waktu pengambilan sampel tanaman. Pengambilan sampel dilakukan menjelang musim hujan (November-Desember 2014). Pada musim hujan populasi tungau hama umumnya akan semakin menurun. Hal ini sesuai dengan yang dikemukakan oleh Zhang (2003), bahwa pada saat musim kemarau aktivitas makan dan reproduksi T. urticae
meningkat secara signifikan dan sebaliknya pada musim hujan aktivitas makan dan reproduksi akan menurun.
Pada blok tanpa pelepasan, secara umum populasi T. urticae cenderung
berfluktuasi dan semakin menurun hingga pengamatan minggu terakhir (Gambar 10B). Populasi T. urticae menurun pada minggu ke-2 kemudian meningkat
kembali pada minggu ke-3 yang merupakan populasi tertinggi sebanyak 71.70 individu per daun. Populasi T. urticae kemudian semakin menurun hingga
pengamatan minggu terakhir (Gambar 10B). Populasi T. urticaeterendah adalah
pada minggu ke-8 sebanyak 0.94 individu per daun. Pada saat populasi T. urticae
menurun secara drastis pada minggu ke-6 populasi N. longispinousus mencapai
populasi tertinggi pada minggu ke-5 dan ke-6. Menurunnya populasi T. urticae
diduga dipengaruhi dengan semakin meningkatnya umur tanaman dan musim saat pengambilan sampel. Pada blok tanpa pelepasan, umur tanaman stroberi mencapai ‰ 7 bulan atau lebih muda dibandingkan umur tanaman stroberi pada blok pelepasan. Sehingga bila dilihat populasi T. urticae pada blok tanpa pelepasan
mempengaruhi, diduga karena pada blok pelepasan predator eksotis P. persimilis,
populasi T. urticaetelah mengalami penekanan oleh predator eksotis tersebut.
Praktek pengendalian kimiawi dan teknik budidaya yang dilakukan, diduga juga mempengaruhi perkembangan populasi tungau predator dan tungau hama di dalam rumah kaca. Selain pengendalian hayati, aplikasi akarisida juga masih dilakukan di dalam rumah kaca. Sehingga berpengaruh terhadap perkembangan populasi tungau hama T. urticae tersebut. Pemeliharaan dengan teknik
pemangkasan (pruning) yaitu dengan menyingkirkan daun stroberi yang sudah tua
atau kering, dapat mempengaruhi perkembangan populasi tungau hama dan tungau predator apabila daun yang disingkirkan tersebut terinfestasi oleh tungau hama.
Kemampuan Memangsa N. longispinosus
Rata-rata pemangsaan tungau predator N. longispinosus terhadap tungau
hama T. urticae tidak berbeda nyata dengan mangsa T. kanzawai (P=0.486),
sementara rata-rata pemangsaan antara fase deutonimfa dan imago betina predator berbeda nyata (P<0.05) dan rata-rata pemangsaan terhadap fase telur dan imago betina hama berbeda nyata (P<0.05) (Lampiran 1). Rata-rata telur T. urticaedan T. kanzawai yang dimangsa imago betina predator sebanyak 14.15 1.87 butir
dan 15.15 1.20 butir secara nyata (P<0.05) lebih tinggi dibandingkan telur yang
dimangsa oleh deutonimfa predator (9.00 2.03 butir dan 9.15 1.14 butir telur)
(Tabel 2). Rata-rata imago betina T. urticae dan T. kanzawai yang dimangsa
deutonimfa predator (1.05 0.32; 1.05 0.41 ekor) tidak berbeda nyata dengan
imago betina predator (1.95 0.32; 2.15 0.57 ekor) (Tabel 2). Tingkat
pemangsaan terendah adalah pada fase deutonimfa predator yang memangsa 1.05
0.32 ekor dan 1.05 0.41 ekor imago betina T. urticaedan T. kanzawai.
Tabel 2 Rata-rata telur dan imago betina T. urticae dan T. kanzawai yang
dimangsa deutonimfa dan imago betina N. longispinosus
Fase
N.longispinosus
Jumlah fase yang dimangsa SDa
T. urticae T. kanzawai
Telur Imago betina Telur Imago betina Deutonimfa 9.00 2.03 b 1.05 0.32 c 9.15 1.14 b 1.05 0.41 c
Imago betina 14.15 1.87 a 1.95 0.32 c 15.15 1.20 a 2.15 0.57 c a Rata-ratayang diikuti oleh huruf yang sama tidak berbeda nyata pada uji Tukey α = 0.05
Tingkat pemangsaan N. longispinosus baik fase deutonimfa maupun fase
imago betina terhadap mangsa berupa telur lebih tinggi dibandingkan mangsa berupa imago T. urticae dan T. kanzawai. Hal ini sesuai dengan hasil penelitian
Puspitarini (2005) yang menyatakan bahwa rata-rata telur Panonychus citri
McGregor (Acari: Tetranychidae) yang dimangsa deutonimfa dan imago betina A. longispinosus secara nyata lebih tinggi dibandingkan nimfa P. citri. Fase imago
betina A. longispinosus dapat memangsa 9.8 ‰ 2.50 sementara fase deutonimfa
menunjukkan bahwa sebagian besar imago N. longispinosus lebih banyak
dijumpai pada daun yang berisi telur T. kanzawai yang menunjukkan bahwa
predator memiliki ketertarikan yang cukup tinggi terhadap mangsa dalam fase telur.
Tungau predator lebih banyak memangsa fase telur karena lebih mudah untuk memangsa telur dibandingkan fase imago yang aktif bergerak dan berkaitan dengan kandungan nutrisi fase mangsa. Mangsa pada fase nimfa dan imago aktif bergerak dan sering memberikan perlawanan sehingga untuk menangkap dan memangsanya diperlukan waktu dan energi yang lebih banyak (Huffacker et al.
1970). Mangsa yang berbeda menyediakan kandungan nutrisi yang berbeda pula (Hoy 2011). Fase telur diketahui memiliki kandungan protein yang tinggi (Sabelis 1985). Kandungan protein telur yang tinggi dibutuhkan oleh imago betina predator untuk pembentukan telur. Pemangsaan predator terhadap telur atau fase pradewasa dari mangsa sangat membantu dalam menekan perkembangan populasi hama.
Tingkat pemangsaan juga ditentukan oleh faktor suhu (Rahman et al. 2012).
Tingkat pemangsaaan A. longispinosus terhadap Aponycus corpuzae Rimando
(Acari : Tetranychidae) dan Schizotetranychus nanjingensis Ma & Yuan (Acari :
Tetranychidae) ditentukan oleh suhu (Zhang et al. 1998; Zhang et al. 1999).
Tungau predator A. longispinosus efektif mengendalikan A. corpuzae pada
rentang suhu 15 ƒC sampai 35 ƒC dan paling efektif pada suhu 25 ƒC (Zhang et al.
1998). Hasil penelitian Zhang et al. (1999) menunjukkan bahwa A. longispinosus
efektif mengendalikan S. nanjingensis pada suhu 30-35 ƒC dan tetapi kurang
efektif pada suhu 10-15 ƒC. Dapat disimpulkan bahwa tingkat pemangsaan N. longispinosus lebih efektif pada suhu tinggi dibandingkan suhu rendah. Pada
pengujian ini suhu rata-rata laboratorium adalah 29 ƒC dan dapat dianggap merupakan suhu yang optimal bagi N. longispinosusuntuk memangsa T. urticae
dan T. kanzawai.
Efesiensi pemangsaan diduga juga terkait dengan efisiensi pencarian mangsa oleh predator. Menurut Huffaker (1971) predator melakukan pencarian mangsa secara acak sampai terjadi kontak dengan mangsa. Hal yang sama juga diungkapkan oleh Krips et al. (1999), bahwa Phytoseiulus persimilis melakukan
pencarian mangsa secara acak. Sebagian besar tingkah laku pemangsa cenderung mencari mangsa yang mempunyai sifat berkelompok daripada mangsa yang sifatnya terpencar (Marchal et al. 1996). Pencarian mangsa oleh predator juga
dipengaruhi oleh keadaan permukaan daun. Hasil penelitian Krips et al. (1999)
menunjukkan bahwa efisiensi pencarian P. persimilis terhadap mangsanya pada Gerbera sp. dipengaruhi oleh kepadatan trikoma pada permukaan daun.
Permukaan arena percobaan berupa daun stroberi diketahui memiliki trikoma yang cukup banyak, sehingga cukup mempengaruhi efisiensi pencarian mangsa. Luas dari arena percobaan dan kepadatan mangsa juga merupakan faktor yang mempengaruhi efisiensi pencarian mangsa oleh predator. Arena percobaan daun stroberi berukuran 2 cm x 2 cm. Pada arena percobaan yang berbeda luasan maka efesiensi predator dalam mencari mangsa akan berbeda juga.
McMurty & Rodriguez (1987) menyatakan bahwa, tingkat kelaparan mempengaruhi rasio pertemuan dan penangkapan mangsa oleh predator. Krips et al. (1999) juga menyatakan bahwa rata-rata pemangsaan tergantung pada rata-rata
memangsa mangsa yang ditemuinya. Pada percobaan, tungau predator N. longispinosus telah dilaparkan selama 6 jam sebelum perlakuan. Perlakuan
tersebut diduga dapat mempengaruhi tingkat pemangsaan tungau predator.
Berdasarkan hasil pengujian, N. longispinosusmemiliki tingkat pemangsaan
yang sama atau tidak berbeda nyata (P=0.486) terhadap T. urticae dan T. kanzawai (Lampiran 1). Kedua mangsa tersebut masih tergolong dalam satu
famili Tetranychidae sehingga tingkat pemangsaan N. longispinosus tidak
berbeda. Berdasarkan hasil tersebut tungau hama T. kanzawai berpotensi
digunakan untuk pembiakan atau rearing N. longispinosussebagai mangsa selain T. urticae. Hal ini karena T. kanzawai cukup mudah ditemui dan
dikembangbiakan pada tanaman ubi kayu sehingga lebih efisien dan ekonomis dibandingkan dengan pengembangbiakan N. longispinosus dengan mangsa T. urticaepada tanaman stroberi.
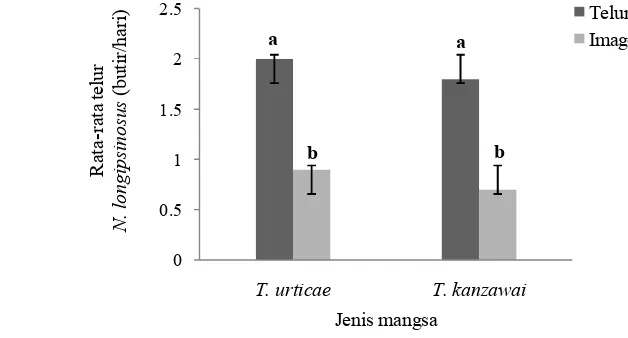
Pengaruh Fase Mangsa terhadap Tingkat Oviposisi N. longispinosus
Gambar 11 menunjukkan rata-rata telur yang diletakkan imago betina N. longispinosuspada saat diberi mangsa dengan fase yang berbeda. Rata-rata telur
yang diletakkan imago betina predator N. longispinosus pada saat diberikan
mangsa telur secara nyata lebih tinggi (P<0.05) dibanding dengan saat diberi
Umumnya phytoseiid dapat menghasilkan 2-4 telur perhari dengan total 20-50 telur per imago betina selama hidupnya (Hoy 2011).
Gambar 11 Rata-rata ( SD) telur yang diletakkan oleh imago betina N. longispinosuspada saat diberi mangsa dengan fase berbeda
Hasil pengujian ini sesuai dengan hasil penelitian Oliviera et al. (2007) yang
menunjukkan rata-rata oviposisi P. macropilis Banks (Acari: Phytoseidae)