1
EKSPLORASI PROTEIN TOKSIN Bacillus thuringiensis DARI
TANAH DI KABUPATEN TANGERANG
MOHAMMAD HERO SHIDDIQI
PROGRAM STUDI KIMIA
FAKULTAS SAINS DAN TEKNOLOGI
UNIVERSITAS ISLAM NEGERI SYARIF HIDAYATULLAH
JAKARTA
2
EKSPLORASI PROTEIN TOKSIN Bacillus thuringiensis DARI
TANAH DI KABUPATEN TANGERANG
Skripsi
Sebagai Salah Satu Syarat Untuk Memperoleh Gelar Sarjana Sains Program Studi Kimia
Fakultas Sains dan Teknologi
Universitas Islam Negeri Syarif Hidayatullah Jakarta
Oleh :
MOHAMMAD HERO SHIDDIQI
106096003240PROGRAM STUDI KIMIA
FAKULTAS SAINS DAN TEKNOLOGI
UNIVERSITAS ISLAM NEGERI SYARIF HIDAYATULLAH
JAKARTA
3
EKSPLORASI PROTEIN TOKSIN Bacillus thuringiensis DARI
TANAH DI KABUPATEN TANGERANG
Skripsi
Sebagai Salah Satu Syarat Untuk Memperoleh Gelar Sarjana Sains Program Studi Kimia
Fakultas Sains dan Teknologi
Universitas Islam Negeri Syarif Hidayatullah Jakarta
Oleh :
MOHAMMAD HERO SHIDDIQI
106096003240Menyetujui,
Pembimbing I Pembimbing II
Drs. Eddy Jusuf, DES NIP. 19500915 197803 1 001
S. Hermanto, M.Si NIP. 19750810 200501 1 005
Mengetahui,
Ketua Program Studi Kimia
4 PENGESAHAN UJIAN
Skripsi berjudul “Eksplorasi Protein Toksin Bacillus thuringiensis dari Tanah di Kabupaten Tangerang” yang ditulis oleh Mohammad Hero Shiddiqi, NIM 106096003240 telah diuji dan dinyatakan “Lulus” dalam sidang Munaqosyah Fakultas Sains dan Teknologi, Universitas Islam Negeri Syarif Hidayatullah Jakarta pada tanggal 1 Desember 2011. Skripsi ini telah diterima sebagai salah
Pembimbing I Pembimbing II
Drs. Eddy Jusuf, DES NIP. 19500915 197803 1 001
S. Hermanto, M.Si NIP. 19750810 200501 1 005
Mengetahui,
Dekan Fakultas Sains dan Teknologi Ketua Program Studi Kimia
DR. Syopiansyah Jaya Putra, M.Sis NIP. 19680117 200112 1 001
5 PERNYATAAN
DENGAN INI SAYA MENYATAKAN BAHWA SKRIPSI INI ADALAH
HASIL KARYA SENDIRI YANG BELUM PERNAH DIAJUKAN SEBAGAI
SKRIPSI ATAU KARYA ILMIAH PADA PERGURUAN TINGGI ATAU
LEMBAGA MANAPUN.
Jakarta, Desember 2011
6 Mohammad Hero Shiddiqi, Eksplorasi Protein Toksin Bacillus thuringiensis
dari Tanah di Kabupaten Tangerang. Dibawah bimbingan Eddy Jusuf dan S. Hermanto.
ABSTRAK
Bacillus thuringiensis (Bt) merupakan bakteri gram positif berspora penghasil protein toksin yang dapat bersifat sebagai insektisidal dan sitosidal. Penelitian ini dilakukan dengan tujuan mendapatkan isolat lokal B.thuringiensis dari tanah di Kabupaten Tangerang dan memprediksi bobot molekulnya. Sampel tanah yang di uji antara lain : Ciherang, Ciputat, Pasar Kemis, Kosambi, Parung Secab, Puspiptek, Pamulang, dan Cileduk. Isolasi B.thuringiensis dilakukan dengan menggunakan metode Travera et al. (1987). Protein toksin disolasi dari bakteri menggunakan metode Bel et al. (1997) dan dianalisis kandungan proteinnya dengan metode Lowry. Profil protein dianalisis menggunakan SDS-PAGE dengan konsentrasi resolving gel 12 % dan stacking gel 4 %. Hasil penelitian menunjukkan terdapat 6 sampel tanah yang positif terdapat
B.thuringiensis, yaitu Ciherang, Pasar Kemis, Kosambi, Parung Secab, Pamulang dan Cileduk. Dari profil protein hasil SDS-PAGE terdapat 5 jenis protein toksin dari 10 sampel protein yang diprediksi bersifat insektisidal atau ICP, yaitu Cry 15, Cry 23, Cry 25, Cry 30, dan Cry 35, serta terdapat 3 jenis protein toksin dari 5 sampel protein yang diprediksi bersifat sitosidal atau parasporin, yaitu PS-2 (Cry 46), PS-3 (Cry41), dan PS-4 (Cry 45).
7 Mohammad Hero Shiddiqi, Exploration of Toxin Protein Bacillus thuringiensis from Soil in Kabupaten Tangerang. Supervised by. Eddy Jusuf and S. Hermanto.
ABSTRACT
Bacillus thuringiensis (Bt) is a gram positive sporulated bacterium and toxin protein producer that can act as insecticidal and sitocidal agents. This research was conducted with a purpose to get local isolate of B.thuringiensis from the soil of Tangerang District and determine protein molecular weight. Samples of soil were taken from Ciherang, Ciputat, Pasar Kemis, Kosambi, Parung Secab, Puspiptek, Pamulang, and Cileduk Villages. Isolation of B. thuringiensis was conducted by using Travera et al. method (1987), throught the isolated toxin protein from bacterium by using Bel et al. method (1997) and protein analysed with Lowry method. Protein profile was analysed by mean of SDS-PAGE with concentration resolving gel 12% and stacking gel 4%. The result showed that 6 samples were positive of B. thuringiensis, ie from Ciherang, Pasar Kemis, Kosambi, Parung Secab, Pamulang and Cileduk. Profile of SDS-PAGE result showed 5 type of toxin proteins from 10 samples predicted as an insecticidal agents or ICP, i.e Cry 15, Cry 23, Cry 25, Cry 30, and Cry 35, and 3 type of toxin proteins from 5 samples predicted as an sitocidal agents or parasporine, i.e PS-2 (Cry 46), PS-3 (Cry41), and PS-4 (Cry 45).
viii
KATA PENGANTAR
Assalamu ’alaikum wr. wb.
Segala puji serta syukur penulis panjatkan kepada Allah SWT atas rahmat,
hidayah, dan inayah-Nya, sehingga penulis dapat menyelesaikan skripsi ini
dengan judul “Ekplorasi Protein Toksin Bakteri Bacillus thuringiensis dari Tanah di Kabupaten Tangerang”.
Skripsi ini merupakan tahapan akhir dan tugas utama dari serangkaian kuliah
yang harus diselesaikan dalam menempuh pendidikan strata satu pada Program
Studi Kimia, UIN Syarif Hidayatullah Jakarta.
Terselesaikannya skripsi ini tidak terlepas dari peranan berbagai pihak yang
telah ikut secara langsung maupun tidak langsung. Maka dalam kesempatan ini,
penulis mengucapkan banyak terima kasih kepada :
1. Bapak Drs. Eddy Jusuf, DES, selaku Pembimbing I yang dengan sabar telah banyak memberikan pengarahan, masukan dan motivasi dalam penelitian
dan penyusunan tugas akhir ini.
2. Bapak S. Hermanto, M.Si, selaku Pembimbing II sekaligus Pembimbing Akademik yang telah banyak memberikan arahan dan motivasi dalam
penyusunan tugas akhir dan menyelesaikan kuliah ini dengan baik.
3. Bapak DR. Syopiansyah Jaya Putra, M.Sis, selaku Dekan Fakultas Sains dan
Teknologi, Universitas Islam Negeri Syarif Hidayatullah Jakarta.
4. Bapak Drs. Dede Sukandar, M.Si, selaku Ketua Program Studi Kimia
Fakultas Sains dan Teknologi, Universitas Islam Negeri Syarif Hidayatullah
ix 5. Ayahanda Amat Rusdi, Ibunda Nahidhotul Millah, serta adik-adik tercinta
atas dorongan dan do’anya yang senantiasa mengiringi gerak langkah
penulis.
6. Mba Pipit dan Mba Prita yang telah banyak memberikan bantuan selama
proses penelitian berlangsung di Pusat Laboratorium Terpadu UIN.
7. Dini Novalia Pratiwi yang telah banyak memberikan masukan dan
semangatnya baik moril maupun spiritual hingga tahap penyusunan tugas
akhir ini.
8. Teman-teman seperjuangan yang juga telah memberikan semangat dan
sarannya sehingga tugas akhir ini dapat terselesaikan.
Penulis menyadari bahwa masih banyak kekurangan dalam penulisan skripsi
ini. Segala kritik dan saran yang membangun diharapkan dari pembaca.
Akhir kata, penulis ucapkan terima kasih dan berdo’a semoga semua amal
baik yang terlah diberikan tersebut mendapat balasan dari Allah SWT, dan skripsi
ini dapat bermanfaat bagi pembaca untuk perkembangan ilmu pengetahuan dan
teknologi serta semoga Allah swt selalu melimpahkan taufik dan hidayah-Nya
kepada kita semua. Amiin.
Wassalamu ‘alaikum wr. wb.
Jakarta, Desember 2011
x
DAFTAR ISI
Halaman Judul ... Halaman
KATA PENGANTAR ... viii
DAFTAR ISI ... x
DAFTAR TABEL ... xiii
DAFTAR GAMBAR ... xiv
DAFTAR LAMPIRAN ... xvi
BAB I PENDAHULUAN ... 1
1.1. Latar Belakang ... 1
1.2. Perumusan Masalah ... 3
1.3. Hipotesis ... 3
1.4. Tujuan Penelitian ... 4
1.5. Manfaat Penelitian ... 4
BAB II TINJAUAN PUSTAKA ... 5
2.1. Bacillus thuringiensis ( Bt ) ... 5
2.1.1. Klasifikasi ... 5
2.1.2. Morfologi dan Fisiologi ... 5
2.1.3. Identifikasi dan Screening ... 7
2.2. Protein ... 8
2.2.1. Struktur Protein ... 8
2.2.2. Klasifikasi protein ... 12
2.2.3. Metode Identifikasi Protein ... 13
2.3. Protein Toksin pada Bacillus thuringiensis ... 14
xi
2.3.2. Parasporin ... 17
2.4. SDS PAGE ... 21
2.4.1. Prinsip Dasar ... 21
2.4.2. Aplikasi (Keunggulan dan Kelemahannya) ... 22
BAB III METODE PENELITIAN ... 24
3.1. Waktu dan Tempat Penelitian ... 24
3.2. Alat dan Bahan ... 24
3.3. Cara Kerja ... 25
3.3.1. Isolasi bakteri dari tanah ... 25
3.3.2. Isolasi protein ... 26
3.3.3. Kuantitifikasi protein dengan Lowry ... 27
3.3.4. SDS-PAGE (Laemli, 1970) ... 27
3.4. Desain Penelitian ... 30
3.4.1. Isolasi B. thurigiensis ... 30
3.4.2. Isolasi protein toksin ... 31
BAB IV HASIL DAN PEMBAHASAN ... 32
4.1. Isolasi B.thuringiensis dari tanah di Kabupaten Tangerang ... 32
4.2. Isolasi protein toksin ... 34
4.3. Kuantifikasi protein dengan Lowry ... 36
4.4. Profil protein hasil SDS-PAGE ... 39
BAB V KESIMPULAN DAN SARAN ... 47
5.1. Kesimpulan ... 47
5.2. Saran ... 47
xiii
DAFTAR TABEL
Halaman
Tabel 2.1. Jenis-jenis protein Cry dari Bacillus thuringiensis berdasarkan ukuran dan serangga sasarannya
………... 16
Tabel 2.2. Spektrum sitotoksisitas empat macam parasporin terhadap berbagai tipe sel kanker dan normal ... 19
Tabel 4.1. Isolat dari sampel tanah yang menghasilkan protein kristal
……… 34
Tabel 4.2. Nilai absorbansi dan konsentrasi sampel protein kristal ... 38
xiv
Gambar 2.5. Beberapa jenis ikatan yang terdapat pada polipeptida ... 11
Gambar 2.6. Struktur kuartener protein ... 11
Gambar 2.7. Struktur tiga dimensi protein Cry 2Ab10 ... 15
Gambar 4.1. Isolasi B. thuringiensis dari tanah : (a) Isolat B. thuringiensis pembanding dan (b) Isolat Pasar Kemis……….. 32 yang menghasilkan protein kristal. (b). Isolat B.thuringiensis pembanding ( Isolat Cibinong, 5 k). Perbesaran 50 x 10 .... ... 33
Gambar 4.4. Peremajaan bakteri berkristal di medium Luria Bertani (LB) ……… 35
Gambar 4.5. Isolasi bakteri berkristal di medium 2xSG ………..
xv Gambar 4.9. Hasil analisa SDS PAGE pada sampel protein Cdk 3,
Kem 22, Ch 7, Cdk 4, Ch 3 Pm 14, Cdk 14, Kem 3, dan
Cdk 9………... 40
Gambar 4.10. Hasil analisa SDS PAGE pada sampel protein Pm 6, Kos 15, Kem 24, Kem 7, Par 16, dan Kem 5 ……… 41
Gambar 4.11. Hasil analisa SDS PAGE pada sampel protein Pm 23, Kem 10, Cdk 9 dan Kem 18 ...
41
Gambar 4.12. Mekanisme kerja ICP………. 44
xvi
Lampiran 7. Standar protein marker catalog #161-0318
(BIO-RAD)... 59
Lampiran 11. Kurva standar protein marker pada analisa sampel protein Pm 6, Kos 15, Kem 24, Kem7, Par 16, dan Kem
5……….... 63
Lampiran 12. Kurva standar protein marker pada analisa sampel protein Pm 23, Cdk 9, Kem 10, dan Kem 18………... 64
Lampiran 13. Hasil uji mikroskop perbesaran 50 x 10………... 65
Lampiran 14 Isolasi 24 koloni terpilih………... 66
1
BAB I
PENDAHULUAN
1.1. Latar Belakang
Bacillus thuringiensis merupakan suatu jenis bakteri gram positif yang
terdiri dari sejumlah besar subspesies atau varietas dan galur-galur (strains) yang
ditemukan hampir di semua habitat. Bakteri ini pertama kali ditemukan tahun
1901 oleh Ishiwata, yaitu peneliti Jepang pada ulat sutra (Bombyx mori) yang
diketahui patogen terhadap serangga (Herlambang, 2007).
Dilihat dari sifat morfologi maupun fisiologinya bakteri ini memiliki
persamaan dengan Bacillus cereus, yang membedakan dengan B.thuringiensis
adalah adanya kristal protein yang bersifat toksin terhadap serangga (Benhard et
al., 1993). Protein toksin ini pertama kali dikenal sebagai parasporal crystalline
inclusion selanjutnya disebut sebagai δ-endotoksin atau Insecticidal Crystal
Protein (ICP) yang dibagi dalam dua kategori protein, yaitu: protein Cry (dari
kata Crystal) dan protein Cyt (dari kata Cytolytic) (Jusuf, 2009).
Sejak tahun 1930, ICP telah digunakan sebagai pestisida hayati
(bioinsektisida) untuk menanggulangi serangga yang menjadi hama tanaman
pertanian. B. thuringiensis dikenal sebagai agensia bahan baku pestisida yang baik
dalam pertanian dan aman terhadap kesehatan serta ramah lingkungan. Sifat
ramah lingkungan tersebut dikarenakan protein kristal yang diisolasi dari B.
thuringiensis mempunyai target yang spesifik sehingga tidak mematikan serangga
yang bukan sasaran dan mudah terurai, serta tidak menumpuk dan mencemari
2 Pada awal abad 21, telah diketahui juga bahwa beberapa tipe protein yang
disintesis oleh B. thuringiensis memiliki kemampuan menghambat tumbuh sel-sel
kanker pada manusia. Yokohama et al. (1988) pertama kali melaporkan bahwa
protein 25 kDa yang diisolasi dari B. thuringiensis subs. israelensis ONR-60A
dapat menghambat tumbuh kultur sel leukemia tikus. Protein Cry anti kanker
manusia dari galur B. thuringiensis disebut sebagai parasporin, diperkenalkan
pertama kali oleh Mizuki et al. (2000). Istilah parasporin didefinisikan sebagai
protein-protein δ-endotoksin yang non-hemolitik tetapi memiliki kemampuan
preferensial membunuh sel kanker. Penemuan tersebut menjadi alternatif dalam
menangani penyakit kanker. Pada dasarnya parasporin merupakan protein Cry
juga seperti contoh protein PS-1 adalah Cry 31, PS-2 adalah Cry 46, PS-3 adalah
Cry 41, dan PS-4 adalah Cry 45 (Ohba et al., 2009).
Penyebaran bakteri ini sangat luas, didapatkan di tanah (Martin dan
Travers, 1989), pada makanan ternak (Meadow et al., 1992), pada batang dan
daun pepohonan (Smith dan Couche, 1991) dan lingkungan perairan (Thanabalu
et al., 1992). Bangkai serangga merupakan sumber nutrisi yang memenuhi dalam
kelangsungan hidup bakteri ini, dimana pada saat nutrisi akan habis maka B.
thuringiensis akan membentuk spora dan mensintesis protein kristal toksik
terhadap jenis serangga tertentu.
Umumnya bangkai serangga terdapat pada tanah sehingga pada penelitian
ini isolasi B. thuringiensis dilakukan pada tanah yang berasal dari Kabupaten
Tangerang. Isolasi dimaksudkan untuk melengkapi informasi mengenai sebaran B.
thuringiensis pada daerah tersebut. Penelitian ini dilakukan untuk mengeksplorasi
3 anti serangga dan anti kanker, kemudian mengidentifikasi tipe protein yang
didapatkan berdasarkan bobot molekulnya. Identifikasi dilakukan dengan
SDS-PAGE (Sodium Dodecyl Sulfate Polyacrilamid Gel Electroforesis), di mana dari
keseluruhan isolat yang diperoleh diharapkan mampu menghasilkan protein toksin
sesuai dengan karakteristik bobot molekulnya.
1.2. Perumusan Masalah
Berdasarkan latar belakang penelitian, maka rumusan masalah yang
diajukan adalah sebagai berikut :
1) Apakah isolasi bakteri dari tanah di Kabupaten Tangerang mampu
menghasilkan B.thuringiensis ?
2) Bagaimana profil protein toksin hasil isolasi dari isolat lokal B.thuringiensis
berdasarkan karakteristik bobot molekulnya ?
1.3. Hipotesis
Isolat lokal B.thuringiensis mampu menghasilkan protein toksin (Cry atau
Cyt) dengan karakteristik bobot molekul yang sama atau berbeda dengan isolat
4 1.4. Tujuan Penelitian
Penelitian ini dilakukan untuk :
1) Mendapatkan isolat B.thuringiensis dari tanah di Kabupaten Tangerang.
2) Mengetahui tipe dan karakteristik protein toksin yang didapat, dengan
berdasarkan bobot molekulnya dan dibandingkan dengan data base dari hasil
penelitian sebelumnya.
1.5. Manfaat Penelitian
Manfaat penelitian adalah sebagai berikut :
1. Melengkapi informasi mengenai sebaran isolat lokal B.thuringiensis yang ada
di wilayah JABODETABEK.
2. Memberikan kontribusi bagi pencarian sumber-sumber baru penghasil
bioinsektisida alami yang ramah lingkungan.
3. Studi awal pemanfaatan protein toksin sebagai alternatif pengobatan penyakit
5 BAB II
TINJAUAN PUSTAKA
2.1. Bacillus thuringiensis ( Bt ) 2.1.1. Klasifikasi
Divisio : Schizophyta
Kelas : Schizomycetes
Ordo : Eubacteriales
Familia : Bacillaceae
Genus : Bacillus
Spesies : thuringiensis Perbesaran 100 x 10. Gambar 2.1.Bacillus thuringiensis.
(Herlambang, 2007)
2.1.2. Morfologi dan Fisiologi
Bacillus thuringiensis (Bt) merupakan bakteri gram positif dengan bentuk
batang, membentuk spora dengan panjang 3 – 5 m dan lebar 1 – 1,2 m,
bergerak aktif (motil) dengan flagella peritrich (di seluruh dinding sel) , bersifat
fakultatif aerob dan bisa didapatkan di seluruh benua dan kepulauan dari
ketinggian 0 sampai 2000 meter di atas permukaan laut dan zona tropis hingga
artik. Bakteri ini bisa didapatkan di habitat alam seperti pada tanah, pepohonan,
debu penyimpanan biji-bijian serealia, pakan ternak, dan serangga yang mati.
Termasuk bakteri mesofil dengan kisaran suhu pertumbuhan 15 – 45 oC, dengan
suhu optimum antara 26-37 oC , dan kisaran pH pertumbuhan antara 5,5 sampai
6 hijau kebiruan, berukuran 1,0 – 1,3 m dengan posisi terminal, sedangkan protein
kristal berukuran 0,6 sampai 2,0 m bergantung dari tipe-nya masing-masing
(Zeigler, 1999).
Gambar 2.2. B. thuringiensis isolat 84-HS-1-11, Ket : S = Spora, P = Protein Cry ( Parasporal Inclusion) ( Mizuki et. al., 2000 ).
Secara alami B. thuringiensis adalah bakteri entemopatogen dan sampai
saat ini telah diidentifikasi sebanyak 70 subspesies atau varietas yang berbeda,
yang dibedakan oleh perbedaan serulogi dari antigen flagella-nya, dan
menghasilkan lebih dari 300 protein Cry dari 1000 galur dan hanya beberapa galur
yang telah dimanfaatkan.
Protein Cry merupakan bentuk protoksin yang disintesis oleh B.
thuringiensis bersamaan dengan pembentukan spora yang berada di dalam sel
sampai sel mengalami lisis sesudah sporulasi sempurna. Spora pada bakteri
tersebut merupakan suatu usaha perlindungan diri dari pengaruh lingkungan luar
yang buruk, hal ini terjadi karena dinding bakteri yang bersifat impermeabel.
Protein Cry yang dihasilkan merupakan bagian dari 25 % berat kering
bakteri yang terdiri dari suatu molekul glikoprotein dengan massa molekular 30
7 tidak larut dalam air ataupun pelarut organik, tetapi larut dalam larutan alkali dan
terdenaturasi oleh panas, asam lambung dan enzim protease lambung sehingga
terlarut dalam air dan membentuk toksin aktif yang akan tetap aktif meskipun
dipanaskan hingga suhu 80˚C selama 20 menit (Dini, 2005).
2.1.3. Identifikasi dan Screening
B. thuringiensis dapat diisolasi dari berbagai sumber, antara lain mudah
ditemukan di tanah dengan populasi yang cukup tinggi, bagian tumbuhan, kotoran
hewan,dan dari serangga yang terinfeksi di lapangan. Ohba dan Aizawa (1979)
telah berhasil memperoleh 105 spora bakteri B. thuringiensis untuk 1 gram tanah
dan selanjutnya Ohba dan Aizawa (1985) telah berhasil mengisolasi 189 kultur
bakteri B. thuringiensis dari 139 sampel tanah di Jepang. Identifikasi
B.thuringiensis dapat dilakukan dengan berbagai cara, beberapa diantaranya
antara lain metode Aizawa et al. (1975), Metode Johson dan Bishop (1996), dan
metode Travera et al, (1987). Pada peneitian ini metode yang digunakan adalah
metode Travera.
Metode Travera et al. (1987) menggunakan medium T-3 sebagai medium
untuk pertumbuhan dan sporulasi B. thuringiensis. Dimana sampel tanah yang
akan diidentifikasi terlebih dahulu dihomogenkan dengan buffer fosfat dan
dipanaskan dengan suhu 70° C selama 30 menit. Pemanasan dilakukan untuk
membunuh sel-sel bakteri atau mikroorganisme yang sedang tumbuh termasuk
spora bakteri lain yang sedang tumbuh. Kemudian diratakan di medium T-3 dan
8 dengan B. thuringiensis. Protein Cry terbentuk setelah proses sporulasi pada
waktu inkubasi hari ke tiga.
Screening B. thuringiensis pada medium dengan koloni yang terbentuk
dapat diamati dari permukaan yang kasar, licin agak mengkilat, warna koloni
putih kekuningan. Pengamatan dengan mikroskop dapat dilihat dari bentuk sel
vegetatifnya yang berbentuk batang dan memiliki spora dan kristal di dalamnya.
Bakteri berspora namun tidak memiliki kristal bukanlah B. thuringiensis namun
diduga merupakan B. cereus yang memiliki banyak kesamaan namun tidak
ditemukan protein kristal pada bakteri tersebut (Salaki et al., 2009).
2.2. Protein
Protein berasal dari kata Proteos (utama atau pertama) merupakan senyawa
makromolekul yang memiliki peranan penting pada setiap makhluk hidup
(Lehninger, 1982). Protein adalah suatu polipeptida dengan bobot molekul yang
sangat bervariasi, dari 5000 hingga satu juta. Disamping berat molekul yang
berbeda-beda, protein memiliki sifat yang berbeda-beda pula (Poedjiadi, 1994),
dengan fungsi yang spesifik ditentukan gen yang sesuai (Lehninger, 1982).
2.2.1. Struktur Protein
Protein tanpa memandang fungsi dan aktifitas biologisnya dibangun oleh
susunan dasar yang sama yaitu 20 asam amino yang molekulnya sendiri tidak
memiliki aktivitas biologis. Masing-masing asam amino dalam suatu protein
terintegrasi melalui ikatan peptida yang tersusun secara kovalen membentuk
struktur yang beragam bergantung dari proses pembentukan dan fungsi dari
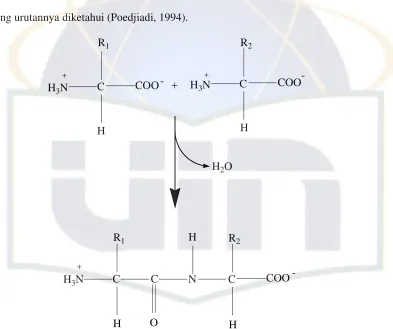
9 Struktur protein terdiri dari struktur primer, sekunder, tersier, dan
kuartener. Struktur primer menunjukkan jumlah, jenis, dan urutan asam amino
dalam molekul protein. Struktur primer protein juga menunjukkan ikatan peptida
yang urutannya diketahui (Poedjiadi, 1994).
R1 R2
Gambar 2.3. Struktur primer protein (Poedjiadi, 1994).
Struktur sekunder protein adalah struktur protein yang dihasilkan oleh
adanya interaksi ikatan hidrogen. Struktur sekunder terdiri dari α- heliks (spiral)
dan β- sheets (lembaran berlipat). Terdapat dua bentuk struktur β- sheets, yaitu
paralel dan anti paralel. Bentuk paralel terjadi apabila rantai polipeptida yang
berikatan melalui ikatan hidrogen itu sejajar dan searah, sedangkan bentuk anti
paralel terjadi apabila rantai polipeptida berikatan dalam posisi sejajar tapi
10 α-heliks
Β-sheets (paralel) β-sheets (anti paralel)
Gambar 2.4. Struktur sekunder protein (Poedjiadi, 1994)
Struktur tersier menunjukkan kecendrungan polipeptida membentuk
lipatan atau gulungan, sehingga membentuk struktur yang lebih kompleks.
Struktur ini dimantapkan oleh adanya beberapa ikatan antara gugus R pada
molekul asam amino yang membentuk protein. Beberapa jenis ikatan tersebut
11 rantai samping non polar, (d) interaksi dipol-dipol, dan (e) ikatan disulfida yaitu
suatu ikatan kovalen antara residu sistein (Poedjiadi, 1994).
Gambar 2.5. Beberapa jenis ikatan yang terdapat pada polipeptida (Poedjiadi, 1994)
Struktur kuartener menunjukkan adanya interaksi intermolekuler antar
unit-unit protein. Sebagaian besar protein globular terdiri atas beberapa rantai
polipeptida yang terpisah. Rantai polipeptida ini saling berinteraksi membentuk
persekutuan (Poerdjiadi, 1994). Gambar 2.6 menunjukkan suatu model struktur
kuartener.
12 2.2.2. Klasifikasi protein
Golongan protein berdasarkan fungsi biologisnya, antara lain :
a. Biokatalisator/ enzim dari hampir semua reaksi yang terjadi pada makhluk
hidup.
b. Protein pengangkut yaitu hemoglobin (Hb) yang mengikat dan membawa
oksigen ke dalam jaringan peri-peri dan lipoprotein yang membawa lipid
dari hati ke organ lain.
c. Antibodi yaitu immunoglobin yang berperan dalam sistem kekebalan
tubuh.
d. Protein pengatur yaitu hormon insulin yang berperan mengatur
keseimbangan kadar glukosa dalam darah.
e. Protein Struktural (keratin dan kolagen).
f. Protein kontraktil (myosin).
g. Protein nutrient yaitu ovalbumin dan kasein.
h. Protein Toksin, pada penelitian ini protein Cry termasuk protein toksin
yang dihasilkan oleh B.thuringiensis untuk mempertahankan hidupnya.
Golongan protein berdasarkan struktur, antara lain :
a. Protein fiber/ serat merupakan protein yang tidak larut dalam air, fleksibel
dan lentur.
b. Protein globular merupakan protein yang larut dalam air dan tidak stabil
13 2.2.3. Metode Identifikasi Protein
Identifikasi protein dapat dilakukan secara kualitatif maupun kuantitatif.
Secara kualitatif, yaitu melalui reaksi xantoprotein, Hopkins-Cole, reaksi Millon,
reaksi natriunnitroprusida, dan reaksi Sakaguchi. Sedangkan secara kuantitatif,
yaitu metode Kjeldahl, metode titrasi formol, metode Lowry, metode biuret dan
metode spetofotometer UV (Apriyanto, 1989). Pada penelitian ini metode yang
digunakan dalam mengidentifikasi protein adalah metode Lowry.
Metode lowry dikembangkan pada tahun 1951 dengan menggunakan
reagen pendetektor Folin-Ciocalteu. Reagen ini biasa digunakan untuk mendeteksi
gugus-gugus fenolik. Dalam analisa protein dengan menggunakan reagen
Folin-Ciocalteu dapat mendeteksi residu tirosin yang mengandung gugus fenolik
melalui reaksi reduksi oksidasi dimana gugus fenolik tirosin akan mereduksi
gugus fosfotungstat dan fosfomolibdat yang terdapat pada reagen tersebut menjadi
tungsten dan molibden yang berwarna biru. Intensitas warna kompleks sebanding
dengan kandungan protein dalam sampel yang dianalisa (Apriyanto et al., 1989).
Hasil reduksi ini dapat dianalisa lebih lanjut dengan melihat puncak absorbsi yang
lebar pada daerah panjang gelombang sinar tampak (600-800 nm).
Sensitifitas metode ini mengalami perbaikan yang cukup signifikan
apabila digabung dengan metode biuret atau penambahan ion Cu, dimana
kompleks Cu-protein yang dihasilkan reagen biuret akan menyebabkan reduksi
fosfotungstat dan fosfomolibdat dalam reagen Folin-Ciocalteu, sementara
residu-residu tirosin dan triptofan mereduksi sisanya.
Dalam analisa kadar protein dengan metode Lowry, diperlukan protein
14 rentang konsentrasi tertentu dimana konsentrasi sampel protein berada didalam
rentang tersebut (Hermanto, 2008).
2.3. Protein Toksin pada Bacillus thuringiensis 2.3.1. ICP (Insecticidal Crystal Protein)
ICP (Insecticidal Crystal Protein) merupakan protein Cry (δ-endotoksin)
yang dihasilkan Bacillus thuringiensis yang bersifat anti serangga. Tiap-tiap
protein Cry memiliki toksisitas yang spesifik dengan sasaran serangga yang
spesifik dan dapat juga memiliki beberapa serangga sasaran (Zeigler, 1999).
Mekanisme kerja dari ICP adalah dengan termakan langsung oleh larva
pada saat memakan daun atau bagian tumbuhan lain yang mengandung protein
kristal. Kemudian protoksin tersebut masuk ke dalam usus dan terlarut oleh asam
serta enzim protease sehingga menjadi toksin. Toksin ini secara spesifik akan
menempel pada membran-membran sel di dalam usus serangga tersebut. Dalam
waktu 1-3 jam sel-sel akan mengalami gangguan osmotik dikarenakan toksin
tersebut meningkatkan derajat permeabilitas dinding sel, sehingga sel akan
menggembung lalu membentuk lubang-lubang pada membran dan pecah (Dini,
2005).
Perubahan biokimia yang tidak stabil tersebut akhirnya menyebabkan
serangga lemas, aktivitas makannya menurun dan tidak merespon untuk
pertumbuhan selanjutnya hingga terjadi perubahan secara fisik pada warna
tubuhnya dimulai warna kecoklatan pada bagian anterior sampai pada bagian
15 Struktur protein δ-endotoksin memiliki tiga daerah (domain) aktif rantai
polipeptida berdasarkan perbedaan struktur dan fungsi yang mengakibatkan
kematian serangga (gambar 7 ), yaitu domain I merupakan 7 gabungan rantai helix
(disimpulkan α = alpa) dapat menyebabkan pembentukan lubang-lubang pada
usus serangga dengan membentuk “ion channel”, domain II membentuk untaian
polipeptida yang panjang (disimpulkan β = beta) dan berikatan dengan ujung α-7
(domain I) berfungsi sebagai pengenal reseptor sel-sel epitel serangga. Domain III
banyak mengandung arginin dan diperkirakan berperan dalam menstabilkan “ion
channel”.
Gambar 2.7. Struktur tiga dimensi protein Cry 2Ab10 (Lin et al., 2007)
Umumnya protein Cry dengan bentuk kubus toksik terhadap jenis
serangga ordo Lepidoptera dan Diptera. Sedangkan protein Cry dengan bentuk
bipiramidal toksik terhadap jenis serangga ordo Lepidoptera dan yang berbentuk Domain III
Domain I
16 oval hanya toksik pada jenis serangga ordo Diptera (Dini, 2005). Bobot dari
protein Cry juga menentukan sifat toksik pada serangga tertentu seperti terlihat
pada Tabel 2.1.
Tabel 2.1.Beberapa jenis-jenis protein Cry dari Bacillus thuringiensis berdasarkan ukuran dan serangga sasarannya.
17 2.3.2. Parasporin
Parasporin adalah protein Cry yang memiliki kemampuan sitosidal
terhadap sel kanker. Berdasarkan hasil penelitian hingga tahun 2008, telah
diketahui ada empat jenis parasporin, yaitu PS-1, PS-2, PS-3 dan PS-4. Pada tahun
2005, Hiromi juga menemukan protein kristal 29 kDa yang disebut protein p-29.
Aktivitas sitosidal parasporin terhadap sel kanker hanya terjadi bila protein
tersebut didegradasi oleh enzim-enzim protease menjadi satu molekul protein
kecil (Jusuf, 2010).
PS-1 merupakan protein Cry Aa/b/c berbentuk spherical dengan struktur 3
domain, bukan toksin pembentukan pori-pori membran. Agriculture and
Agri-food Canada (AAFC) telah mengisolasi dan karakterisasi satu protein Cry31Aa2
dari galur B. thuringiensis M15 yang menunjukkan aktivitas sitosidal in vitro
khususnya terhadap sel HepG2 (human hepatocyte cancer cells) dan sel-sel jurkat
(leukemic T cells). Mizuki et. al. (2000) mengisolasi protein dari B.thuringiensis
nomor isolat 84-HS-1-11 asal Hiroshima berukuran ± 81.045 kDa yang tersusun
dari 723 asam amino, disandi oleh satu gen berukuran 2169 bp. Protein ini dikenal
sebagai Cry31Aa2. Sekuen asam amino tediri dari 5 conserved block seperti
biasanya yang terdapat protein Cry pada umumnya, tetapi homolginya dengan
protein Cry maupun Cyt sangat rendah (< 25%).
Aktivitas sitosidal terjadi apabila protein telah didegradasi oleh protease
menjadi molekul yang kecil berukuran 40 sampai 60 kDa. Tripsin dan proteinase
K dapat mengaktifkan parasporin, tetapi chymotrypsin tidak dapat mengaktifkan,
Aktivitas sitosidal sangat kuat terhadap MOLT-4 (human leukemic T cells) dan
18 thuringiensis var. dakota A1547 yang diisolasi oleh Yamagiwa et al. (2002) juga
gen dari protein yang telah dimurnikan diklon pada sel lain dan menghasilkan
protein rekombinan yang memiliki aktivitas sitosidal yang kuat terhadap sel-sel
kanker hati dan usus tanpa efek terhadap sel normalnya.
Mekanisme sitosidal dari protein ini adalah meningkatkan dengan cepat
kepekatan ion bebas Ca2+ intraseluler dengan tanpa perubahan permeabilitas
membran plasma dan sel-sel kanker dibunuh melalui apoptosis. pada sel HeLa
yang ditreatment dengan parasporin-1, dapat diobservasi adanya degradasi
pro-caspase-3 dan poly (ADP-ribose) polymerase. Hal ini ditunjukkan oleh adanya
penurunan sintesis protein selular dan DNA pada sel HeLa. Tingkat kepekatan ion
bebas Ca2+ naik tajam 1-3 menit setelah pemberian PS-1, dan hasil uji
menunjukkan bahwa derajat sensitivitas sel berbanding lurus dengan besarnya
peningkatan kepekatan Ca2+ intraseluler. Jadi, PS-1 mengaktifkan apoptotic
signaling pada sel-sel kanker yang ditreatment sebagai akibat meningkatnya level
Ca2+ dan influx Ca 2+ ini merupakan langkah awal dalam jalur proses toksisistas
PS-1. Bentuk toksin hasil pemecahan oleh protease adalah 15 dan 56 kDa,
sementara bentuk reseptor pada sel sasaran belum diketahui (Kitada et al., 2005).
PS-2 adalah protein Cry46Aa atau disebut juga sebagai Mtx-like protein
dengan bentuk tidak beraturan yang ditunjukkan oleh protein PS2Aa1. Proses
sitosidal terjadi dengan meningkatkan permeabilitas sel kanker. Tahap awalnya,
peningkatan pada reseptor putative (GPI-anchored proteins) yang berada pada
lipid rafts (detergents resistans membrane) yang diikuti dengan oligomerisasi dan
pembentukan pori pada membran plasma dengan efek sitolisis (Kitada et al.,
19 Protein kristal yang tergolong PS-2 diperoleh dari galur B. thuringiensis
TK-E6 yang dikenal sebagai PS2Ab adalah polipeptida dengan 304 asam amino
dan berat molekul diprediksi sekitar 33.107 kDa (Hayakawa et al., 2007). Sekuen
asam amino PS2Ab menunjukkan homologi signifikan (84% identitas). PS2Aa
yang sebelumnya ditemukan terdapat dalam galur B. thuringiensis var. dakota
A1547. Protein Kristal PS2Ab diproses dengan proteinase K menghasilkan
protein aktif berukuran 29 kDa yang memiliki sitotoksisistas kuat terhadap sel-sel
MOLT-4 dan Jurkat dengan nilai EC50 masing-masing 0,545 dan 0,745 ng/mL.
Toksisitas PS2Ab ternyata jauh lebih tinggi dibandingkan PS2Aa setelah beberapa
kali pengujian (Jusuf, 2010). Spektrum sitotoksisitas dari parasporin terhadap
beberapa tipe kanker dapat dilihat pada Tabel 2.2.
Tabel 2.2. Spektrum sitotoksisitas empat macam parasporin terhadap berbagai tipe sel kanker dan normal ( Ohba et al., 2009 ).
20 Protein kristal PS-3 atau Cry41Aa/b berbentuk bipiramidal berukuran 88
kDa dengan struktur tiga domain. setelah pemecahan protease menjadi aktif pada
toksik dengan ukuran 64 kDa. Mekanisme penghambatan tumbuh sel kanker
belum diungkap dan reseptor yang mengenali protein ini pada membran sel-sel
kanker belum diketahui (Kitada et al., 2005).
PS-4 digolongkan dalam dua macam, yaitu Cry45Aa yang disebut epsilon
toxin like dan Cry42Aa yang merupakan struktur tiga domain, berukuran 31 kDa
dan bentuk aktif pada ukuran 27 kDa, dimana bentuk protein reseptor pada sel
target belum diketahui (Kitada et al., 2005). Didapatkan pada galur B.
thuringiensis var. shandongiensis strain 89-T-34-22 yang dikenal sebagai
Cry45Aa (Okumura et al., 2004).
Sekuen asam amino PS-4 yang ditelusurkan berdasarkan sekuen gen
penyandinya memiliki homologi sangat rendah dibandingkan dengan umumnya
protein Cry maupun dengan ketiga jenis parasporin tersebut di atas. Pelarutan
dengan alkali dan proteinase K menghasilkan protein dengan aktivitas sitotoksik
kuat terhadap MOLT-4 dan sitotoksik lemah terhadap sel-sel T normal, tetapi efek
terhadap sel-sel HeLa tidak terlihat. Uemori et al. (2008) mengidentifikasikan dua
gen protein parasporin masing-masing gen ps1Aa3 dengan panjang 2.619 bp
menyandi protein 81 kDa (PS1Aa3) dan gen ps1Ab1 dengan panjang 2.178 bp
menyandi protein 82 kDa (PS1Ab1) dari B. thuringiensis strain B0195. Sekuen
asam amino PS1Aa3 ternyata 100% identik dengan protein referensi PS1Aa1,
sedang PS1Ab1 adalah 86,4% identik dengan PS1Aa1. Protein rekombinan
21 proteolitik mampu menginduksi sitolisis sel HeLa, tetapi tidak berpengaruh
terhadap sel non-kanker UtSMC (human uterine smoot muxcle cells)
2.4. SDS PAGE (Sodium Dodecyl Sulfate Polyacrilamid Gel Electroforesis) 2.4.1. Prinsip Dasar
Elektroforesis merupakan proses bergeraknya molekul bermuatan pada
suatu medan listrik. Kecepatan molekul-molekul yang bergerak dalam medan
listrik bergantung pada muatan, bentuk dan ukuran. Dengan demikian
elektroforesis dapat digunakan untuk separasi makromolekuler (seperti protein
dan asam nukleat). Posisi molekul yang tersparasi pada gel dapat dideteksi dengan
pewarnaan autoradiografi, ataupun dilakukan dengan densitometer (Ikmalia,
2008).
Salah satu metode elektroforesis yang umumnya digunakan untuk analisa
campuran protein secara kualitatif adalah SDS‐PAGE (Sodium Dodecyl Sulfate
Polyacrilamid Gel Electroforesis). Prinsip penggunaan metode ini adalah
pembentukan polimer dari komponen akrilamida dengan ikatan silamg
N.N` bisakrilamida. Kisi-kisi tersebut berfungsi sebagai saringan
molekul sehingga konsentrasi atau rasio akrilamida dengan bisakrilamida dapat
diatur untuk mengoptimalkan migrasi komponen protein (Wilson dan Walker,
2000).
Elektroforesis gel SDS dilakukan pada pH netral dengan adanya SDS dan
β-merkaptoetanol. Dengan adanya SDS, protein rantai ganda akan terdisosiasi
menjadi rantai-rantai individual dan terdenaturasi oleh detergen ini, sehingga
22 membentuk konfigurasi random coil. Peristiwa ini dibantu dengan adanya
β-merkaptoetanol yang memutus ikatan disulfida antar ataupun dalam rantai
(mereduksi ikatan disulfida menjadi gugus sulfihidril). Di samping itu, SDS
berikatan dengan protein membentuk kompleks SDS-Protein yang tidak memiliki
struktur sekunder dan membawa muatan negatif yang diakibatkan oleh adanya
gugus-gugus anion dari SDS (Nur et al., 1992).
2.4.2. Aplikasi SDS-PAGE
SDS PAGE dapat diaplikasikan berdasarkan kegunaannya antara lain
(Ikmalia, 2008) :
1. Menentukan berat molekul (estimasi). Penetapan BM secara lebih teliti dapat
dilakukan dengan ultrasentrifuge, meskipun dengan elektroforesis sudah
memenuhi syarat.
2. Dapat mendeteksi terjadinya pemalsuan bahan.
3. Dapat mendeteksi terjadinya kerusakan bahan seperti protein dalam
pengolahan dan penyimpanan.
4. Untuk memisahkan spesies molekul yang berbeda secara kualitatif maupun
kuantitatif, yang selanjutnya masing-masing spesies dapat dianalisis.
5. Menetapkan titik isoelektrik protein.
Keunggulan dari SDS PAGE adalah pita protein yang terbentuk menjadi
lebih stabil dikarenakan adanya medium penyangga yaitu gel poliakrilamida,
bersifat transparan sehingga dapat diamati dengan mudah, dapat memperoleh
resolusi yang lebih baik dan ukuran pori dapat diatur sehingga pemisahan
23 sering digunakan dalam elektroforesis gel untuk analisis protein, tapi tidak dapat
digunakan untuk menganalisa sejumlah protein kompleks yang diperlukan
aktifitas biologi dari protein untuk pengujian protein selanjutnya. Hal ini
disebabkan karena protein yang telah di elektroforesis telah mengalami denaturasi
24
BAB III
METODE PENELITIAN
3.1. Waktu dan Tempat Penelitian
Penelitian ini berlangsung selama 8 bulan dari bulan Agustus 2010 sampai
Maret 2011. Tempat penelitian dilakukan di Pusat Penelitian Bioteknologi,
Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong dan Pusat Laboratorium
Terpadu (PLT), UIN Syarif Hidayatullah Jakarta.
3.2. Bahan dan Alat
Bahan yang digunakan adalah beberapa sampel tanah dari Kabupaten
Tangerang (Ciherang, Ciputat, Pasar Kemis, Kosambi, Parung Secab, Puspiptek,
Pamulang, dan Cileduk), galur B. thuringiensis pembanding ( Isolat Cibinong, 5
k), triptosa, tripton, yeast extract, Na3PO4, dan MnCl2, NaCl, HCl, EDTA,
sukrosa, bromophenol blue, SDS, beta-merkaptoetanol, dithiothreitol, Acryl/bis
(sigma), ammonium persulfat (APS), coomassie brilliant blue, glisin, resolving
gel buffer (1,5M Tris-HCl pH 8,8), stacking gel buffer (0,5 M Tris-HCl pH 6,8),
metanol, H2SO4, NH4OH, Na2CO3, CuSO4.5H2O, Na.K.tartrate, reagen folin,
N,N,N’,N’,tetramethylethylenediamine (TEMED), BSA, standar protein marker
catalog #161-0318 (Bio-Rad), aquabides, dan aquades.
Alat yang digunakan pada penelitian ini adalah erlenmeyer, vortex,
timbangan digital, cawan petri, labu ukur, mikroskop, tabung reaksi, pH meter,
25 flow, inkubator, spektrofotometer UV-VIS Shimadzu, dan Mini-Protean Gel
Elektrophoresis 3 cell BIO-RAD.
3.3. Cara Kerja
3.3.1. Isolasi bakteri dari tanah
Teknik isolasi yang dilakukan menggunakan metode Travera et al. (1987).
Sampel tanah ditimbang sebanyak 1 gram. Kemudian dimasukkan ke dalam
tabung 20 mL yang telah berisi 10 mL buffer fosfat 0,05 M dengan pH 6,8,
dikocok dengan kuat selama 15 menit. Setelah itu, dipanaskan pada suhu 70˚C
selama 30 menit dalam waterbath, lalu dikocok lagi dengan kuat untuk meratakan
penyebaran spora selama 15 menit. Sebanyak 5 dan 10 l suspensi disebarkan
pada cawan petri yang berisi agar T-3 (per-liter mengandung 3 g tripton, 2 g
triptosa, 1,5 g yeast extract, 0,05 M Na3PO4, 0,005 M MnCl2 dalam 12 g agar ).
Pengerjaan dilakukan secara duplo. Kemudian diinkubasi pada suhu 28˚C selama
kurang lebih 48 jam, hingga muncul koloni yang memiliki kesamaan morfologi,
warna, aroma yang sama dengan B. thuringiensis pembanding.
Dari setiap sampel dipilih 24 koloni yang betul-betul mirip dengan B.
thuringiensis pembanding, lalu ditransfer ke agar T-3 yang baru di cawan petri
dan diinkubasi pada suhu 28˚C sampai terjadi sporulasi selama kurang lebih
selama 72 jam. Penapisan isolat dari 24 koloni terpilih setiap sampelnya
dilakukan dengan observasi mikroskop menggunakan mikroskop fase kontras atau
cahaya biasa dengan memastikan adanya bentuk protein kristal. Pada observasi
mikroskop, koloni dihomogenkan dengan menggunakan aquades steril pada kaca
26 Koloni yang dipastikan membentuk protein kristal dipanen dan dikoleksi sebagai
isolat baru serta disimpan pada agar Luria-Bertani miring (per liter mengandung
10 g tripton, 5 g yeast extract, 10 g NaCl dalam 15 g agar) dan diberi nomor isolat
sesuai dengan asal sampel.
3.3.2. Isolasi protein
Nomor-nomor isolat bakteri berkristal ditumbuhkan pada agar
Luria-Bertani dalam cawan petri sebanyak 2 kali reinokulasi, kemudian diinkubasi pada
suhu 28˚C selama 24 jam. Setelah itu koloni bakteri yang tumbuh diinokulasikan
dalam plat agar 2xSG ( per-liter mengandung 16 g nutrient broth, 2 g KCl, 0,5 g
MgSO4.7H2O, 2 mL glukosa 50 %, 1 mL Ca(NO3) 1 M, 1 mL MnCl2 0,1 M, 1 mL
FeSO4 1 mM dalam 17 g agar ), lalu diinkubasi hingga bersporulasi selama kurang
lebih 48 sampai 72 jam. Koloni yang telah bersporulasi dipanen dan dimasukkan
ke dalam microtube 1,5 mL yang berisi 1 mL NaCl 0,5 M dingin direndam dalam
es dan dikocok hingga menjadi suspensi homogen. Suspensi disentrifugasi pada
13000x g selama 10 menit, supernatan dibuang dan hanya disisakan pellet pada
microtube. Pellet ditambahkan lagi 1 mL NaCl dan disentrifugasi kembali.
Kemudian pellet yang diperoleh diresuspensi dengan 140 l campuran 1 %
SDS-0,01 % β-merkaptoetanol dan direbus selama 10 menit.
Suspensi kemudian disentriugasi kembali pada 10000x g selama 10 menit
dan supernatan yang terjadi diambil sebagai sampel untuk dianalisis secara
27 3.3.3. Kuantitifikasi protein dengan Lowry
a. Lowry Concentrate 2x :
- Reagen Cooper (20 g Na2CO3 dalam 260 mL H2O, 0,4 CuSO4.5H2O
dalam 20 mL H2O dan 0,2 g Na.K.tartrate dalam 20 mL H2O.
- 10 g SDS dalam 100 mL H2O
- 4 g NaOH dalam 100 mL H2O
Pada waktu digunakan, 3 bagian reagen cooper ditambahkan 1 bagian SDS
dan 1 bagian NaOH.
b. 0,2 N Folin reagent : campur 10 mL 2 N folin dengan 90 mL H2O.
Terhadap 400 ul sampel ditambahkan 400 l Lowry Concentrate 2x,
inkubasi pada suhu ruang, ± 10 menit. Selanjutnya ditambahkan 200 l 0,2 N
Folin reagent, lalu dihomogenkan dengan vortex setiap kali penambahan.
Campuran diinkubasi 30 menit pada suhu ruang, dan dibaca dengan
spektrofotometer pada panjang gelombang 750 nm dengan menggunakan
BSA sebagai pembanding.
3.3.4. SDS-PAGE (Laemli, 1970)
Karakterisasi protein dilakukan dengan elektroforesis SDS-PAGE, dengan
menggunakan 12 % separating gel dan 4% stacking gel. Pewarnaan dengan 0,1 %
coomassie brilliant blue, 50 % metanol dan 10 % asam asetat .
a. Preparasi sampel
Supernatan dari hasil isolasi protein diambil sebanyak 30 l
dimasukkan ke dalam microtube 100 l, kemudian ditambahkan 20 l buffer
28 bromophenol blue, 2,5% SDS, 7,4 mM dithiothreitol). Campuran
dihomogenkan dan dipanaskan selama 10 menit pada suhu 100 °C.
b. Preparasi gel elektroforesis
- Resolving gel (12%)
Aquabides sebanyak 3,4 mL ditambahkan 30% degassed
Acrylamide/bis sebanyak 4 mL, lalu dihomogenkan. Campuran tersebut
ditambahkan resolving gel buffer (1,5M Tris-HCl pH 8,8) sebanyak 2,5
mL, lalu dihomogenkan dan ditambahkan SDS 10 % (w/v) sebanyak 0,1
mL. Setelah itu, ditambahkan ammonium persulfat (APS) 10 % sebanyak
50 l, lalu homogenkan. Kemudian ditambahkan dengan TEMED
sebanyak 5 l dan dihomogenkan kembali.
- Stacking gel (4%)
Aquabides sebanyak 6,1 mL ditambahkan 30% degassed
Acrylamide/bis sebanyak 1,3 mL, lalu dihomogenkan. Campuran tersebut
ditambahkan stacking gel buffer (0,5 M Tris-HCl pH 6,8) sebanyak 2,5
mL, lalu dihomogenkan dan ditambahkan SDS 10 % (w/v) sebanyak 0,1
mL. Setelah itu, ditambahkan ammonium persulfat (APS) 10 % sebanyak
50 l, lalu homogenkan. Kemudian ditambahkan dengan TEMED
sebanyak 10 l dan dihomogenkan kembali.
c. Pembuatan kolom gel
Resolving gel dimasukkan sedikit demi sedikit dengan menggunakan
mikropipet ke dalam alat elektroforesis hingga batas untuk stacking gel.
Kemudian ditambahkan aquabides, untuk meratakan resolving gel. Setelah
29 dipasangkan sisir pembentuk sumur atau kolom, dan dibiarkan hingga
membeku lalu sisir dapat diangkat. Hasil gel tersebut kemudian dipasang
pada perangkat elektroforesis.
d. Loading sampel
Larutan buffer dimasukkan ke dalam tangki elektroforesis. Kemudian
sampel sebanyak 10 l dimasukkan ke dalam kolom gel dengan hati-hati lalu
di elektroforesis selama ± 100 menit dengan tegangan elektrik 120 volt.
e. Pewarnaan dan pencucian gel
Gel diangkat lalu diwarnai dengan coomassie brilliant blue stainning
gel selama semalaman. Kemudian gel diangkat dan dimasukkan ke dalam
30
Bt. dibandingkan Morfologi Isolat
Kandidat yang memiliki
31 3.4.2. Isolasi protein toksin
Inokulasi media LB (duplo) Inkubasi ± 24 jam, 28 °C
Isolat Hasil medium LB
Inokulasi media 2xSG Inkubasi ± 48 – 72 jam, 28 °C
Isolat Hasil medium 2xSG
+ 1 mL NaCl 0,5 M Resuspensi
Pellet Supenatan
+ 1 mL NaCl 0,5 M Resuspensi
Pellet Supernatan
+ 140 l 1 % SDS- 0,01 β-mercaptoetanol Dipanaskan, diresuspensi
Pellet Supernatan
H
ngiensis dari Tanah di Kabupaten Tangerang
asil isolasi yang dilakukan terhadap beberapa samp
ang diperoleh 6 sampel yang diduga memiliki
huringiensis pembanding, yaitu Ciherang, Pasa
ab, Pamulang dan Cileduk. Kemiripan tersebut te
emiliki permukaan yang kasar, agak mengkilat,
an seperti yang terlihat pada gambar 4.1.
asi B. thuringiensis dari tanah : (a) Isolat B. thuring mbanding dan (b) Isolat Pasar Kemis.
Ciputat dan Puspiptek tidak ditemukan adan
ngkinan disebabkan pada sampel tanah yang dia
berulang kali. B. thuringiensis merupakan
ga pada suatu saat tertentu dipengaruhi oleh beber
i, epizootik dan endozootik (Salaki et al., 2009).
Koloni-koloni da
ding, diseleksi kembali dengan pemilihan 24 kolo
mbar 4.2.
(a) (b)
lasi 24 koloni terpilih: (a) Isolat Ciherang dan (b) I mulang.
koloni tersebut dimaksudkan untuk menyeleks
miliki kemiripan dengan B. thuringiensis pemba
ngujian menggunakan mikroskop. Dari pemiliha
dari koloni yang dipilih terdapat protein kri
B. thuringiensis. Berikut merupakan hasil
erbesaran 50 x 10 pada salah satu koloni sa
nding seperti yang terlihat pada gambar 4.3.
(a) (b)
uji mikroskop : (a). Isolat Kem 7 salah satu is asilkan protein kristal. (b). Isolat B.thu nding (Isolat Cibinong, 5 k). Keterangan: S = spor
34 Berdasarkan Gambar 4.3, pada hasil uji mikroskop terlihat bahwa isolat
Kem 7 memiliki kemiripan dengan isolat B.thuringiensis pembanding (Isolat
Cibinong, 5 k). Hal ini dapat diamati dari adanya spora yang berbentuk bulat dan
protein kristal berbentuk lancip dan berwarna kehitaman seperti pada isolat
B.thuringiensis pembanding.
Hasil penapisan 24 koloni terpilih dengan menggunakan mikroskop
didapatkan 30 nomor isolat yang menghasilkan protein kristal. Isolat-isolat dari
tanah yang dipastikan membentuk protein kristal antara lain, terlihat pada tabel
4.1.
Tabel 4.1. Isolat dari sampel tanah yang menghasilkan protein kristal
Sampel Tanah Isolat Penghasil Protein Kristal Ciherang (Ch) Ch-3 dan Ch 7
Parung (Par) Par 8, Par 9, Par 11, Par 16, Par 17, dan Par 18 Pamulang (Pm) Pm 6, Pm 10, Pm14, Pm 19, dan Pm 23
Cileduk (Cdk) Cdk 3, Cdk 4, Cdk 9, dan Cdk 14 Kosambi (Kos) Kos 6, Kos 7, Kos 9, Kos 15 dan Kos 16
Pasar Kemis (Kem) Kem 3, Kem 5, Kem 6, Kem 7, Kem10, Kem 22, Kem 23, dan Kem 24
Tabel 4.1 menunjukkan bahwa, sampel tanah dari Pasar Kemis memiliki
isolat terbanyak dan Ciherang memiliki isolat terkecil sebagai penghasil protein
berkristal. Hal ini menandakan pada sampel tanah yang diambil dari Pasar Kemis
memiliki sumber air, mineral, karbon, dan nitrogen yang cukup banyak bagi B.
thuringiensis serta kondisi lingkungan seperti pH, kelarutan oksigen dan
35 4.2. Isolasi protein toksin
Isolat dari sampel tanah yang menghasilkan protein kristal diremajakan
pada medium Luria Bertani (LB). Hal ini dikarenakan medium LB merupakan
medium dengan kandungan jumlah karbon, nirogen dan mineral yang cukup
banyak bagi kelangsungan hidup B. thutingiensis. Sedangkan medium T-3 dan
2xSG memiliki kandungan karbon, nitrogen dan mineral yang sedikit sehingga
memungkinkan B. thutingiensis untuk bersporulasi. Peremajaan pada medium LB
seperti yang terlihat pada gambar 4.4.
Gambar 4.4. Peremajaan bakteri berkristal di medium Luria Bertani (LB)
Gambar 4.4 menunjukkan adanya koloni yang tumbuh dari hasil
peremajaan di medium LB. Koloni tersebut berwarna putih kekuningan dengan
jumlah yang cukup banyak pada setiap isolat setelah diinkubasi 24 jam, hal ini
menandakan bahwa bakteri dapat hidup dengan baik pada medium LB.
Peremajaan tersebut dimaksudkan agar bakteri dapat lebih produktif dalam
menghasilkan protein kristal saat di medium sporulasi.
Koloni hasil dari peremajaan dipindahkan pada medium 2xSG agar bakteri
bersporulasi sehingga dihasilkan protein kristal. Isolasi bakteri pada medium
36 Gambar 4.5. Isolasi bakteri berkristal di medium 2xSG
Gambar 4.5 menunjukkan bahwa adanya koloni yang tumbuh pada
medium 2xSG, dimana koloni yang tumbuh merupakan koloni yang telah
bersporulasi. Proses sporulasi tersebut merupakan usaha dari bakteri berkristal
dalam mempertahankan diri dengan lingkungan yang membahayakan
kehidupannya (Dini, 2005). Terjadinya proses sporulasi diamati dengan
menggunakan mikroskop, yaitu pada saat sudah terbentuknya spora dan protein
kristal pada isolat yang diamati.
Hasil dari inokulasi pada medium 2xSG yang sudah mengalami sporulasi,
dipanen dan dibersihkan dengan NaCl untuk menghilangkan medium yang
terbawa pada spora dan kristal saat proses pemanenan. Pemisahan spora dan
protein kristal dilakukan dengan menambahkan campuran 1% SDS - 0,01%
β-merkaptoetanol, dimana protein kristal yang bercampur dengan spora akan
terpisahkan karena adanya β-merkaptoetanol kemudian protein kristal yang telah
terpisah akan terlarut karena diikat oleh SDS.
4.3. Kuantifikasi protein dengan Lowry
Kuantifikasi dengan Lowry dilakukan untuk mengetahui kadar protein
37 kristal, yaitu Ch-3, Ch 7, Par 8, Par 9, Par 11, Par 16, Par 17, Par 18, Pm 6, Pm
10, Pm 14, Pm 19, Pm 23, Cdk 3, Cdk 4, Cdk 9, Cdk 14, Kos 6, Kos 7, Kos 9,
Kos 15, Kos 16, Kem 3, Kem 5, Kem 6, Kem 7, Kem10, Kem 22, Kem 23, dan
Kem 24. Sedangkan standar pembanding yang digunakan adalah BSA (Bovine
Serum Albumin) dengan konsentrasi 1 ppm, 5 ppm, 10 ppm, 50 ppm, dan 100
ppm.
Pada pengujian kadar protein dengan menggunakan metode Lowry, semua
sampel dan standar pembanding membentuk kompleks biru keunguan. Hal ini
disebabkan adanya ion Cu+2 yang terdapat dalam kompleks Cu, akan memutus
ikatan peptida serta bersama-sama dengan adanya residu tirosin dan triptofan
menyebabkan reduksi fosfotungstat dan fosfomolibdat yang terdapat dalam reagen
menjadi tungsten dan molibden yang berwarna biru keunguan. Reaksi yang terjadi
pada metode Lowry seperti yang terlihat pada gambar 4.6.
Gambar 4.6. Reaksi pada metode Lowry (Dini, 2005)
Intensitas warna kompleks sebanding dengan kandungan protein dalam
38 warna kompleks yang dihasilkan sampel protein kristal diperoleh konsentrasi
protein sampel sebagai berikut :
Tabel 4.2. Nilai absorbansi dan konsentrasi sampel protein kristal
Sampel Absorbansi Konsentrasi Protein (ppm)
Ch 3 0,402 56,714
Dari hasil pengujian kadar protein, diperoleh konsentrasi terbesar pada
sampel Cdk 3 yaitu 99 ppm, dengan absorbansi sebesar 0,698. Sedangkan
39 absorbansi sebesar 0,112. Perbedaaan kadar protein pada masing-masing sampel
dapat disebabkan dari berbedanya jumlah biomassa isolat yang didapat dan
dipengaruhi oleh proses sporulasi dari isolat pada masing-masing sampel dalam
menghasilkan protein toksin.
4.4. Profil Protein Hasil SDS-PAGE
Analisa SDS-PAGE dilakukan untuk mengestimasi bobot molekul protein
sampel yang didapat dari hasil isolasi B. thuringiensis pada tanah di kabupaten
Tangerang. Kemudian mencocokkan hasil bobot molekul yang didapat dengan
sumber referensi untuk memprediksi jenis protein toksin yang didapat.
Adapun hasil-hasil analisa SDS-PAGE yang dilakukan pada setiap sampel
disajikan pada gambar berikut :
M 1 2 3 4 5 6 7
40 M 1 2 3 4 5 6 7
Gambar 4.8. Hasil analisa SDS PAGE pada sampel protein 1 (Kem 6), 2 (Par 18), 3 (Par 11), 4 (Kos 16), 5 (Par 9), 6 (Pm 19), dan 7 (Pm 10).
M 1 2 3 4 5 6 7 8 9
41 M 1 2 3 4 5 6
Gambar 4.10. Hasil analisa SDS PAGE pada sampel protein 1 (Pm 6), 2 (Kos 15), 3 (Kem 24), 4 (Kem7), 5 (Par 16), dan 6 (Kem 5).
M 1 2 3 4
Gambar 4.11. Hasil analisa SDS PAGE pada sampel protein 1 (Pm 23), 2 (Kem 10), 3 (Cdk 9), dan 4 (Par 18).
Hasil SDS PAGE untuk ke 30 isolat, diinterpretasikan melalui persamaan
42 hingga 12. Hasil prediksi protein toksin dari masing-masing protein sampel
berdasarkan referensi seperti terlihat pada tabel 4.3.
Tabel 4.3. Bobot molekul protein sampel dan prediksi protein toksinnya.
Sampel Bobot Molekul (kDa)
Prediksi protein toksin Sumber
Cdk 3 29 Cry 23 Arrieta et al., 2004
Pada tabel 4.3 dapat dilihat bahwa terdapat 20 sampel protein toksin yang
43 sampel protein lainnya belum dapat diprediksi jenis protein toksinnya. Protein Cyt
yang diprediksi didapat hanya 1 tipe yang terdiri dari 5 sampel protein yaitu Cyt
A. Terdapat 5 tipe protein toksin yang diprediksi bersifat insektisidal atau lebih
dikenal dengan ICP (Insecticidal Crystal Protein) yang terdiri dari 10 sampel
protein, antara lain Cry 15, Cry 23, Cry 25, Cry 30, dan Cry 35. Hal ini
didasarkan pada penelitian sebelumnya dimana, protein Cry 15 memiliki sifat
toksin terhadap serangga ordo lepidoptera (Brown dan Whiteley, 1992), protein
Cry 23 memiliki sifat toksin terhadap serangga ordo diptera (Arrieta et al., 2004),
protein Cry 25 memiliki sifat toksin terhadap serangga ordo coleoptera, protein
Cry 30 memiliki sifat toksin terhadap serangga ordo dipteral (Zeigler,1999), dan
protein Cry 35 memiliki sifat toksin terhadap serangga ordo diptera (Ernest et al.,
2005).
Mekanisme dari protein Cry yang bersifat insektisidal, yaitu dengan
termakan langsung oleh serangga. Protein Cry tersebut menjadi toksin setelah
mengalami proteolisis, kemudian menyebabkan gangguan osmotik sehingga sel
membengkak dan pecah lalu menyebabkan kematian pada serangga (Hofte dan
44 Gambar 4.12. Mekanisme kerja ICP (Jusuf, 2009)
Parasporin adalah protein Cry yang memiliki kemampuan sitosidal
terhadap sel kanker. Berdasarkan hasil penelitian hingga tahun 2008, telah
diketahui ada empat jenis parasporin, yaitu PS-1 (Cry 31), PS-2 (Cry 46), PS-3
(Cry41) dan PS-4 (Cry 45) (Jusuf, 2010). Berdasarkan tabel 4.3, juga terdapat 3
tipe protein toksin yang diprediksi bersifat sitosidal atau disebut parasporin (PS)
dari 5 sampel protein, antara lain PS-2 (Cry 46), PS-3 (Cry41), dan PS-4 (Cry 45).
Mekanisme dari PS-1 yaitu dengan meningkatkan dengan cepat kepekatan
ion bebas Ca2+ intraseluler dengan tanpa perubahan permeabilitas membran
plasma dan sel-sel kanker dibunuh melalui apoptosis (Kitada et al., 2005). Pada
PS-2 proses sitosidal terjadi dengan meningkatkan permeabilitas sel kanker
(Kitada et al., 2005). Sedangkan untuk PS-3 dan PS-4 hingga saat ini belum
diketahui mekanisme kerjanya terhadap sel kanker. Berikut merupakan
45 Gambar 4.13. Mekanisme kerja PS-2 (Jusuf, 2010)
Protein Cyt A yang dihasilkan oleh B.thuringiensis memiliki bobot
molekul 27 kDa (Balaraman et al., 2005) dan 28 kDa (Hofte dan Whiteley,
1989). Sampel protein Ch 3 dan Ch 7 memiliki bobot molekul sebesar 27 kDa,
serta sampel protein toksin Par 17, Kos 16, dan Cdk 14 memiliki bobot molekul
sebesar 28 kDa. Sehingga diprediksi sampel protein Ch 3, Ch 7, Par 17, Kos 16
dan Cdk 14 merupakan protein Cyt A.
Pada penelitian yang dilakukan Arrieta et al. (2004), protein toksin yang
dihasilkan oleh B.thuringiensis dengan bobot molekul 29 kDa digolongkan
sebagai protein Cry 23. Sampel protein Kos 7, Cdk 3, Cdk, 4, dan Cdk 9 memiliki
bobot molekul sebesar 29 kDa, sehingga diprediksi merupakan protein Cry 23.
Sampel protein Kos 9 dengan bobot molekul sebesar 34 kDa diprediksi
sebagai protein Cry 15, hal ini didasarkan pada penelitian yang dilakukan Brown
dan Whiteley (1992) pada B.thuringiensis subs thompsoni bahwa protein Cry
dengan bobot molekul 34 kDa digolongkan sebagai protein Cry 15.
Zeigler (1999) menyatakan dalam Bacillus Genetic Stock Center of
46
bobot molekul 35 kDa digolongkan sebagai protein Cry 30 dan protein Cry
dengan bobot molekul 100 kDa digolongkan sebagai Cry 25. Protein sampel Par
16 dan Kem 5 memiliki bobot molekul sebesar 35 kDa sehingga diprediksi
merupakan protein Cry 30, sedangkan sampel protein Kem 23 memiliki bobot
molekul 100 kDa diprediksi merupakan protein Cry 25. Ernest et al. (2005),
melakukan karakterisasi terhadap protein Cry 35 dan didapat bobot molekul
sebesar 44 kDa. Sampel protein Par 18 dan Kos 6 memiliki bobot molekul sebesar
44 kDa sehingga diprediksi merupakan protein Cry 35.
Hayakawa et al. (2007) menggolongkan protein kristal dengan bobot
molekul ± 33,107 kDa merupakan PS 2 (Cry 46) yang diperoleh dari galur
B.thuringiensis TK-E6. Sampel protein Par 11 memiliki bobot molekul 33 kDa,
sehingga diprediksi sebagai PS 2 (Cry 46). Sampel protein Kem 22 dengan bobot
molekul 88 kDa diprediksi sebagai PS 3 (Cry 41), sedangkan Pm 23, Par 9 dan
Kem 7 dengan bobot molekul 31 kDa diprediksi sebagai PS 4 (Cry 45). Hal ini
didasarkan pada penelitian Kitada et al. (2005) yang mengidentifikasi parasporin
dan sel kanker sasarannya, parasporin dengan bobot molekul 88 kDa digolongkan
sebagai PS 3 (Cry 41) dan parasporin dengan bobot molekul 31 kDa digolongkan
sebagai PS 4 (Cry45).
Berdasarkan hasil-hasil SDS PAGE, terlihat bahwa dari beberapa isolat
yang menghasilkan protein toksin, memiliki karakteristik bobot molekul yang
berbeda-beda. Hal ini disebabkan karena B.thuringiensis dapat memproduksi satu
atau lebih protein toksin selama proses sporulasi (Hofte dan Whiteley, 1989).
Namun untuk mengkonfirmasi hal tersebut perlu dilakukan pengujian in vitro (bio
47
BAB V
KESIMPULAN DAN SARAN
5.1. Kesimpulan
Dari hasil penelitian ini dapat disimpulkan beberapa hal, antara lain:
1. Terdapat 30 isolat B.thuringiensis yang ditemukan dari 8 sampel tanah di
Kabupaten Tangerang.
2. Ke 30 isolat B.thuringiensis tersebut menghasilkan 8 tipe protein Cry dan 1
protein Cyt dengan bobot molekul 27 dan 28 kDa (Cyt A), 29 kDa (Cry 23),
31 kDa (Cry 45/ PS-4), 33 kDa (Cry 46/ PS-2), 34 kDa (Cry 15), 35 kDa (Cry
30), 44 kDa (Cry 35), 88 kDa (Cry 41/ PS-3), dan 100 kDa (Cry 25).
5.2. Saran
Berdasarkan hasil penelitian ini, maka disarankan agar dilakukan
perlakuan lebih lanjut, antara lain:
1. Perlu dilakukan isolasi protein toksin dengan jumlah biomassa yang lebih
banyak dan dilakukan pemurnian protein toksin.
2. Perlu adanya pengujian secara in vitro (bio assay) untuk mengidentifikasi
aktivitas sebagai anti kanker dan anti serangga dari protein toksin yang