SELEKSI DAN STUDI PEWARISAN SERTA
PENGEMBANGAN MARKA SSR PENANDA KETAHANAN
TERHADAP PENYAKIT LAYU BAKTERI (
Ralstonia
solanacearum
) PADA TOMAT
EKA JAN VIRGIN HAQUARSUM
A253130091
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis Seleksi dan Studi Pewarisan serta Pengembangan Marka SSR Penanda Ketahanan terhadap Penyakit Layu Bakteri (Ralstonia solanacearum) pada Tomat adalah karya saya sendiri dan belum
diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, November 2016
RINGKASAN
EKA JAN VIRGIN HAQUARSUM. Seleksi dan Studi Pewarisan Serta Pengembangan Marka SSR Penanda Ketahanan terhadap Penyakit Layu Bakteri
(Ralstonia solanacearum) pada Tomat. Dibimbing oleh SURJONO HADI
SUTJAHJO, KIKIN HAMZAH MUTAQIN dan CATUR HERISON.
Tomat yang dimanfaatkan dalam bentuk buah segar ataupun olahan merupakan tanaman sayuran buah penting, baik di Indonesia maupun di dunia. Di Indonesia, kebutuhan tomat masih belum terpenuhi oleh produksi dalam negeri sehingga berpengaruh pada harga yang sering berfluktuasi. Beberapa masalah yang mempengaruhi produksi tomat telah diketahui, salah satunya serangan penyakit layu bakteri. Penyakit yang tersebar luas di area tropis dan subtropis ini disebabkan oleh bakteri Ralstonia solanacearum. Bakteri ini bersifat gram negatif
dan merupakan bakteri tular tanah.
Penelitian ini terdiri dari empat percobaan, yang meliputi : (1) seleksi tiga puluh genotipe tomat lokal koleksi terhadap karakter ketahanan penyakit layu bakteri (Ralstonia solanacearum) yang bertujuan untuk menyeleksi ketahanan
genotipe tomat lokal koleksi terhadap penyakit layu bakteri, (2) studi pewarisan karakter ketahanan terhadap penyakit layu bakteri pada tomat yang bertujuan untuk mempelajari pola pewarisan sifat ketahanan terhadap penyakit layu bakteri pada tomat, dan (3) identifikasi marka SSR (Simple Sequence Repeats) penanda
ketahanan terhadap penyakit layu bakteri pada tomat yang bertujuan untuk mendapatkan marka SSR penanda ketahanan terhadap penyakit layu bakteri pada tomat.
Percobaan pertama dan kedua dilakukan dengan menggunakan isolat bakteri yang berasal dari tanaman sakit di lapang. Tanaman diinokulasi pada saat minggu keempat setelah semai dengan cara pelukaan ujung akar dan direndam pada 20 mL suspensi bakteri selama 30 menit. Kemudian tanaman disiram oleh suspensi bakteri tersebut.
Percobaan ketiga dilakukan melalui tiga tahapan, yaitu: seleksi primer dengan menggunakan dua genotipe tahan dan rentan (hasil dari percobaan pertama), BSA pada populasi F2, dan identifikasi ketahanan tomat koleksi dengan menggunakan primer terpilih.
Setiap genotipe tomat lokal koleksi menunjukkan ketahanan yang berbeda. Dari tiga puluh genotipe tomat lokal koleksi terdapat 1 genotipe sangat tahan (Kudamati 1), 8 genotipe tahan (Gondol Lonjong, Khemir, Kudamati 3, Lombok 3, Makasar 3, Situbondo Bulat Kecil, Situbondo Gelombang, dan Tanah Datar), 3 genotipe agak tahan (Aceh 1, Aceh 5, dan Bajawa), 8 agak rentan (Cherry NTT, Kali Acai, Kefamenanu 3, Kefamenanu 6, Kefamenanu7, Makasar 1, Makasar 2, dan Makasar 4), 1 rentan (Meranti 1) dan 9 genotipe sangat rentan (Aceh 2, Aceh 3, Kefamenanu 9, Kefamenanu 12, Kefamenanu 14, Lombok 1, Lombok 2, Lombok 4, dan Meranti 2). Genotipe Kudamati 1 dan Lombok 4 digunakan untuk merakit populasi F1, F1R, dan F2 pada Percobaan Kedua.
yang dikendalikan oleh dua pasang gen mayor dengan aksi gen duplikat resesif epistasis.
Tiga tahapan yang dilakukan pada percobaan tiga menunjukkan bahwa primer TOM-144 diduga bersifat polimorfik sebagai penanda ketahanan terhadap penyakit layu bakteri. Hasil pita amplifikasi menunjukkan bahwa genotipe tahan memiliki ukuran yang berkisar pada 144 bp. Primer TOM-144 mampu mengidentifikasi ketahanan tomat terhadap penyakit layu bakteri.
Hasil penelitian ini dapat dimanfaatkan oleh para pemulia untuk dapat mengembangkan varietas tahan berupa galur murni ataupun hibrida. Marka SSR yang didapat juga dapat mengidentifikasi secara cepat ketahanan terhadap penyakit layu bakteri pada genotipe atau varietas lainnya.
SUMMARY
EKA JAN VIRGIN HAQUARSUM. Selection, Inheritance and Development of SSR Marker to Bacterial Wilt Disease (Ralstonia solanacearum) Resistance on
Tomato. Supervised by SURJONO HADI SUTJAHJO, KIKIN HAMZAH MUTAQIN and CATUR HERISON.
Tomato is an important vegetable which consumed as a fresh or processed product. In Indonesia, the domestic production is still insufficient resulted in prices fluctuation. One of problems which affect the production of tomato is bacterial wilt disease that caused by Ralstonia solanacearum. This disease is
widespread in tropical and subtropical areas. R. solanacearum is gram-negative
and soil borne bacterium.
There were four experiments, i.e. (1) selection of thirty local tomato genotypes to bacterial wilt (R. solanacearum) disease resistance which aimed to
select the bacterial wilt disease resistance on tomato, (2) inheritance of bacterial wilt disease resistance trait on tomato which aimed to study of inheritance to bacterial wilt disease resistance on tomato, and (3) identification of SSR (Simple Sequence Repeats) marker to bacterial wilt disease resistance in tomato which
aimed to identify by SSR marker to bacterial wilt disease resistance on tomato. The first and second experiments were done using field isolated inoculum were collected from the infected plants in the field. Plants were inoculated by wounding the root using scissor at four weeks after seeding. Plants were immersed in 20 mL of bacterial suspension for thirty minutes, planted in soil medium in pot, and then watered by the bacterial suspension.
The third experiment was done by three steps, i.e (1) primer screening used two resistant and susceptible genotypes (from the result of the first experiment), confirm (2) BSA on F2 population, and (3) identification the bacterial wilt disease resistance by selected primer.
Each of local genotypes of tomato showed different resistances. There were 1 highly resistant (Kudamati 1), 8 resistants (Gondol Lonjong, Khemir, Kudamati 3, Lombok 3, Makasar 3, Situbondo Bulat Kecil, Situbondo Gelombang, and Tanah Datar), 3 moderately resistants (Aceh 1, Aceh 5, and Bajawa), 8 moderately susceptibles (Cherry NTT, Kali Acai, Kefamenanu 3, Kefamenanu 6, Kefamenanu7, Makasar 1, Makasar 2, and Makasar 4), 1 susceptible (Meranti 1), and 9 very susceptibles (Aceh 2, Aceh 3, Kefamenanu 9, Kefamenanu 12, Kefamenanu 14, Lombok 1, Lombok 2, Lombok 4, and Meranti 2). Kudamati 1 and Lombok 4 were used for generate F1, F1R, and F2 population.
Testing on F1 population showed that the inheritance of resistance to bacterial wilt disease on tomato is dominant without maternal effect with the degree of dominance is uncomplete. χ2 testing on F2 population had a ratio of 9:7 (resistant : susceptible) which indicated that the character is a qualitative character controlled by two pairs of major genes with duplicate recessive epistasis.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Departemen Agronomi dan Hortikultura
SELEKSI DAN STUDI PEWARISAN SERTA
PENGEMBANGAN MARKA SSR PENANDA KETAHANAN
TERHADAP PENYAKIT LAYU BAKTERI (
Ralstonia
solanacearum
) PADA TOMAT
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas segala karunia dan kasih sayang-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih pada penelitian ini adalah Seleksi dan Studi Pewarisan serta Pengembangan Marka SSR Penanda Ketahanan terhadap Penyakit Layu Bakteri (Ralstonia solanacearum) pada Tomat.
Terima kasih penulis sampaikan kepada:
1. Prof Dr Ir Surjono Hadi Sutjahjo, MS, Dr Ir Kikin Mutaqin, MSi, dan Dr Ir Catur Herison, MSc, selaku komisi pembimbing yang telah banyak memberikan arahan, bimbingan, saran, dan motivasi sejak perencanaan penelitian sampai penyusunan tesis ini selesai.
2. Dr Desta Wirnas, SP, MS selaku dosen penguji luar komisi yang telah banyak memberi masukan.
3. DIKTI, yang telah memberikan beasiswa BPPDN.
4. Tim Program Penelitian KKP3N yang diketuai oleh Prof Dr Ir Surjono Hadi Sutjahjo, MS, yang telah memberi dana untuk penelitian ini.
5. Teman-teman seangkatan PBT ’13 dan teman-teman di Laboratorium Plant and Molecular Biology 2.
6. Mama Eliya Wati dan Bapak Abdul Kadir Jailani untuk doa, motivasi, dan omelan yang selalu mengiringi setiap langkahku.
7. Ibu dan Om Ujang yang sudah banyak membantu.
8. Saudara-saudariku: Dwi Lika Shauma Nur Hanifah Haquarsum, Elin Nur Afifah Amatullah Haquarsum, Ahmad Marzuki Haquarsum, dan Tika Hafizhah Haquarsum, untuk warna yang berbeda di tiap harinya.
9. Semua pihak yang telah membantu selama penelitian.
Semoga karya ilmiah ini bermanfaat bagi pengembangan ilmu pengetahuan.
Bogor, November 2016
DAFTAR ISI
1.3 Hipotesis Penelitian 2
1.4 Ruang Lingkup Penelitian 2
2. TINJAUAN PUSTAKA 2.1 Tomat
2.2 Koleksi Plasma Nutfah Tomat
4 4
2.3 Penyakit Layu Bakteri 5
2.4 Pemuliaan Tanaman 6
2.5 Penanda Molekuler 7
3. SELEKSI TIGA PULUH GENOTIPE TOMAT LOKAL TERHADAP KARAKTER KETAHANAN PENYAKIT LAYU BAKTERI
(Ralstonia solanacearum)
3.1 Pendahuluan 3.2 Bahan dan Metode 3.3 Hasil dan Pembahasan 3.4 Kesimpulan 4. STUDI PEWARISAN KARAKTER KETAHANAN TERHADAP
PENYAKIT LAYU BAKTERI (Ralstonia solanacearum) PADA
TOMAT
5. IDENTIFIKASI MARKA SSR PENANDA KETAHANAN TERHADAP PENYAKIT LAYU BAKTERI (Ralstonia
solanacearum) PADA TOMAT
5.1 Pendahuluan 5.2 Bahan dan Metode 5.3 Hasil dan Pembahasan 5.4 Kesimpulan
5.5 Daftar Pustaka
34 35 38 46 46
6. PEMBAHASAN UMUM 48
7. KESIMPULAN DAN SARAN 7.1 0Kesimpulan
7.2 0Saran
50 50 50
DAFTAR PUSTAKA 51
LAMPIRAN 57
DAFTAR TABEL
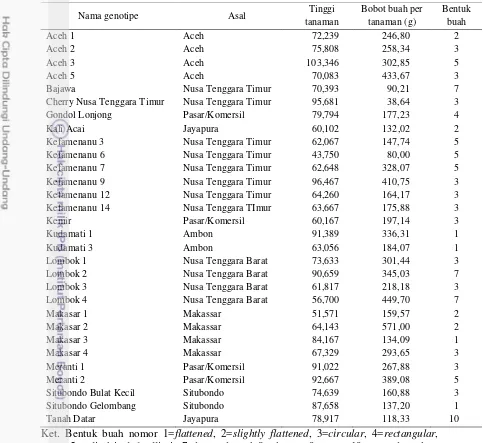
1 Deskripsi tiga puluh genotipe tomat lokal koleksi Laboratorium
Genetik dan Pemuliaan Tanaman IPB 5
2 Respon ketahanan tomat terhadap penyakit layu bakteri (R.
solanacearum) berdasarkan kejadian penyakit 12
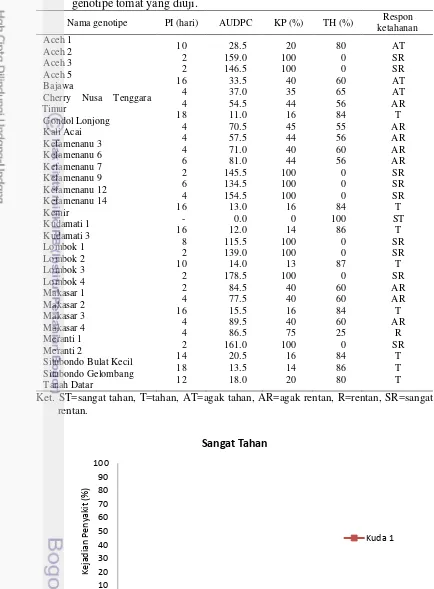
3 Periode inkubasi (PI), Area Under the Disease Progress Curve
(AUDPC), kejadian penyakit (KP), tanaman hidup (TH), dan
respon ketahanan genotipe tomat yang diuji 13
4 Respon ketahanan tomat terhadap penyakit layu bakteri (R.
solanacearum) berdasarkan kejadian penyakit. 24
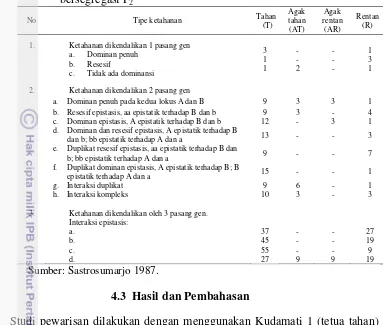
5 Klasifikasi derajat dominansi berdasarkan nilai potensi rasio (hp) 25 6 Nisbah fenotipik frekuensi karakter resistensi tanaman terhadap
penyakit yang dikendalikan oleh gen mayor dalam populasi
bersegregasi F2. 26
7 Periode Inkubasi (PI), Area Under the Disease Progress Curve
(AUDPC), Kejadian Penyakit (KP), Tanaman Hidup (TH), dan
respon ketahanan populasi tomat yang diuji. 27 8 Hasil uji χ2 pada populasi F2 berdasarkan kejadian penyakit
dengan beberapa nisbah Mendel 30
9 Jenis primer yang digunakan dan urutan basanya 36
DAFTAR RUMUS
1 Kejadian Penyakit 11
2 Area Under the Disease Progress Curve 12
3 Jumlah Minimum Tanaman F2 23
4 Kejadian Penyakit 24
5 Area Under the Disease Progress Curve 24
6 Derajat Dominansi 25
DAFTAR GAMBAR
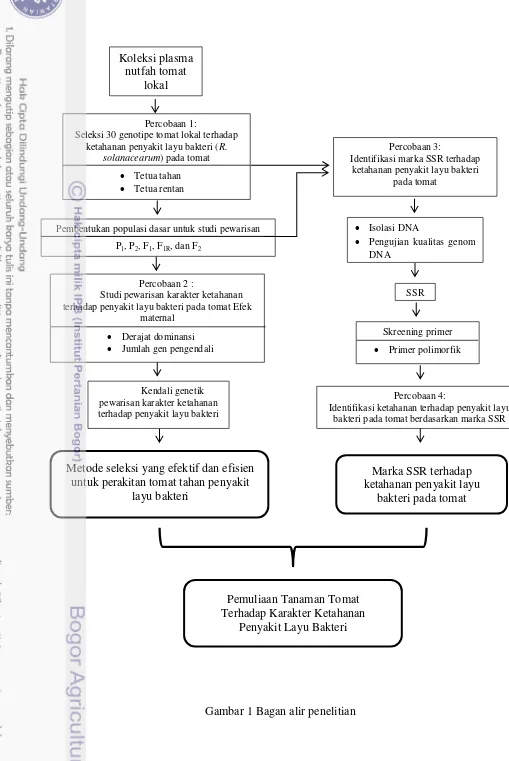
1 Bagan Alir Penelitian 3
2 Grafik kejadian penyakit kelas sangat tahan 13
3 Grafik kejadian penyakit kelas tahan 14
4 Grafik kejadian penyakit kelas agak tahan 14
5 Grafik kejadian penyakit kelas agak rentan 14
6 Grafik kejadian penyakit kelas rentan 15
7 Grafik kejadian penyakit kelas sangat rentan 15 8 A.Respon ketahanan tanaman muda terhadap pemberian
inokulasi R. solanacearum (4 hari setelah inokulasi) B. Gejala
layunya daun muda pada tanaman yang sudah besar (10 hari setelah inokulasi). C. Pembentukan akar adventif pada tanaman dewasa yang terserang (30 hsi). D. Profil browning pada
batang dengan potongan vertikal. E. Ooz bakteri yang keluar dari batang tanaman sakit. F. Koloni R. solanacearum pada
media TTC 16
9 Histogram sebaran frekuensi tanaman hidup pada populasi P1,
P2, F1, dan F2 27
10 Respon ketahanan tanaman pada hari ke-2 setelah inokulasi 29 11 Respon ketahanan tanaman pada hari ke-10 setelah inokulasi 29 12 Respon ketahanan tanaman pada hari ke-20 setelah inokulasi 29 13 Respon ketahanan tanaman pada hari ke-30 setelah inokulasi 29 14 Hasil elektroforesis 10 primer dengan menggunakan agarose. 39
15 Hasil elektroforesis 5 primer dengan menggunakan PAGE. 40 16 Hasil elektroforesis 5 primer dengan menggunakan PAGE. 40 17 Hasil elektroforesis 10 primer dengan menggunakan agarose. 41
18 Hasil elektroforesis 10 primer dengan menggunakan PAGE. 41 19 Hasil elektroforesis 5 primer dengan menggunakan agarose. 42
20 Hasil elektroforesis 5 primer dengan menggunakan PAGE. 43
DAFTAR LAMPIRAN
1 Pembuatan Media TTC 58
2 Pembuatan Larutan Stok Ekstraksi DNA 58
3 Pembuatan Larutan Penyangga 58
4 Ethidium bromide 59
5 Pembuatan Larutan Stok Bahan Pembuatan Gel Polyakrilamid 59
1 PENDAHULUAN
1.1 Latar Belakang
Tomat adalah salah satu jenis tanaman sayuran penting di Indonesia selain kentang, cabe, dan bawang. Kebutuhan masyarakat akan tomat tergolong besar, terlihat pada data bahwa hingga Januari 2015 Indonesia masih melakukan impor sebesar 1.117, 348 ton untuk dapat memenuhi kebutuhan tersebut (BPS 2015). Tingkat produksi dalam negeri yang masih rendah disebabkan oleh beberapa faktor, di antaranya faktor genetik varietas yang ditanam masih belum optimal dengan cekaman lingkungan yang ada. Menurut Purwati (2008) budidaya memerlukan varietas unggul yang cocok dengan lokasi penanaman, teknik budidaya yang tepat, pemupukan yang berimbang, dan pengendalian hama dan penyakit yang efektif sehingga hasil produksi yang tinggi dapat dicapai.
Salah satu organisme pengganggu tanaman tomat yang sangat merugikan adalah bakteri Ralstonia solanacearum yang menyebabkan penyakit layu bakteri
dan merupakan salah satu penyakit tanaman paling berbahaya yang tersebar luas di daerah tropis dan subtropis (Hayward 1964). Patogen ini memiliki kemampuan bertahan hidup dalam waktu yang lama di dalam tanah sehingga patogen sulit dikendalikan (Saddler 2005) dan dapat menurunkan hasil sekitar 15-35% (Hayward 1964), serta menyerang tanaman pertanian lainnya, seperti kacang tanah, pisang, kentang, tembakau, dan suku Solanaceae lainnya (Persley et al.
1985).
Kegiatan pemuliaan tanaman diharapkan dapat menghasilkan varietas tahan untuk menanggulangi serangan penyakit layu bakteri. Kegiatan pemuliaan tanaman akan efektif bila memperhatikan pewarisan karakter. Beberapa hasil penelitian mengenai pewarisan karakter ketahanan terhadap penyakit layu bakteri hingga saat ini masih belum konsisten. Menurut Monma dan Sakata (1993) karakter ketahanan tomat terhadap layu bakteri dikendalikan oleh aksi gen resesif dengan karakter kualitatif maupun kuantitatif. Osiru et al. (2001) menyatakan
ketahanan tomat terhadap penyakit layu bakteri dikendalikan oleh dua gen, dan Lafortune et al. (2005) menyatakan karakter ketahanan cabai terhadap layu bakteri
dikendalikan oleh sedikit gen dan dipengaruhi oleh efek aditif. Berbagai hasil penelitian tersebut membuat studi pewarisan karakter ketahanan tomat terhadap layu bakteri masih perlu dilakukan.
Menurut Ganapathy et al. (2012), memahami perbedaan genetik antar
individu merupakan hal yang penting dalam pemuliaan. Dalam mempelajari perbedaan genetik Prabakaran et al. (2010) menyatakan bahwa hal tersebut dapat
dievaluasi secara penampakan morfologi, protein pada biji, isozim dan penanda molekuler. Penanda molekuler sangat penting dalam meningkatkan efisiensi pemuliaan tanaman (Moon 2006). Penanda molekuler dapat memberikan karakterisasi komprehensif sumber daya genetik (Sehgal et al. 2012). Berbeda
dengan sifat-sifat morfologi, penanda molekuler dapat memperlihatkan perbedaan antar berbagai genotipe pada tingkat DNA, efisien untuk karakterisasi plasma nutfah, konservasi, manajemen dan tidak dipengaruhi oleh faktor lingkungan (Prabakaran et al. 2010; Tommasini et al. 2003). Penanda molekuler sekarang
Dalam aplikasinya, penanda molekuler semakin efektif dengan adanya teknik Bulk segregant analysis (BSA). BSA adalah langkah acak untuk
mengidentifikasi penanda pada daerah spesifik. Setiap bulk terdiri dari individu
yang identik untuk setiap karakter yang berbeda (misal: tahan dan rentan) (Michelmone et al. 1991). BSA telah digunakan untuk mempermudah kegiatan
molekuler berbasis PCR, di antaranya pada penanda RAPD (Michelmone et al.
1991) dan RAFL (Hämäläinen et al. 1997).
1.2 Tujuan Penelitian
1. Menyeleksi 30 genotipe tomat lokal terhadap ketahanan penyakit layu bakteri (R. solanacearum).
2. Mempelajari pola pewarisan karakter ketahanan tomat terhadap penyakit layu bakteri (R. solanacearum).
3. Memperoleh marka SSR yang polimorfik pada metode BSA.
4. Mengidentifikasi ketahanan tomat terhadap penyakit layu bakteri (R.
solanacearum) berdasarkan marka SSR
1.3 Hipotesis Penelitian
1. Terdapat keragaman ketahanan terhadap penyakit layu bakteri (R. solanacearum) pada genotipe tomat lokal.
2. Terdapat lebih dari satu gen yang berperan dalam pewarisan karakter ketahanan penyakit layu bakteri (R. solanacearum).
3. Terdapat marka SSR yang spesifik pada metode BSA.
4. Terdapat keragaman ketahanan tomat terhadap penyakit layu bakteri (R.
solanacearum) berdasarkan marka SSR.
1.4 Ruang Lingkup Penelitian
Penelitian ini memiliki ruang lingkup yang saling berkaitan menjadi satu kesatuan dalam mencapai tujuan penelitian yang meliputi: (1) seleksi karakter ketahanan terhadap penyakit layu bakteri (R. solanacearum) pada tomat, (2) studi
pewarisan karakter ketahanan terhadap penyakit layu bakteri pada tomat, (3) identifikasi marka SSR (Simple Sequence Repeats) penanda ketahanan terhadap
Koleksi plasma nutfah tomat
lokal
Percobaan 1:
Seleksi 30 genotipe tomat lokal terhadap ketahanan penyakit layu bakteri (R.
solanacearum) pada tomat
Tetua tahan Tetua rentan
Pembentukan populasi dasar untuk studi pewarisan P1, P2, F1, F1R, dan F2
Percobaan 2 :
Studi pewarisan karakter ketahanan terhadap penyakit layu bakteri pada tomat Efek
maternal
Pengujian kualitas genom DNA
Kendali genetik pewarisan karakter ketahanan terhadap penyakit layu bakteri
Metode seleksi yang efektif dan efisien untuk perakitan tomat tahan penyakit
layu bakteri
Percobaan 4:
Identifikasi ketahanan terhadap penyakit layu bakteri pada tomat berdasarkan marka SSR
Marka SSR terhadap ketahanan penyakit layu
bakteri pada tomat
Gambar 1 Bagan alir penelitian Pemuliaan Tanaman Tomat Terhadap Karakter Ketahanan
2 TINJAUAN PUSTAKA
2.1 Tomat
Tomat (Lycopersicon esculentum Mill) merupakan tanaman budidaya
penting di seluruh dunia. Menurut Jagatheeswari (2014) tomat memiliki jenis tomat liar dan budidaya yang mana dapat tumbuh dengan baik di daerah mesofit. Tomat memiliki batang yang tidak berkayu dengan cabang utama yang ditutupi oleh rambut epidermis. Daun tomat memiliki kedudukan berselang-seling. Pembungaan bersifat majemuk tak berbatas (inflorescentia racemosa) dengan
bunga yang memiliki ukuran kecil, bewarna kuning, dan bersifat hermafrodit. Tomat mampu tumbuh di berbagai habitat, dari daerah di sekitar permukaan laut hingga 3.300 m di atas permukaan laut (Peralta et al. 2006). Sejak abad
ke-20, telah dilakukan pengelompokkan berbagai kultivar tomat berdasarkan morfologi melalui pemuliaan tanaman. Melalui domestikasi, kegiatan penelitian dan pemuliaan yang dilakukan oleh para ilmuwan dan petani di seluruh dunia, varietas tomat (sebagian besar hibrida) telah dikembangkan dengan berbagai bentuk, warna, dan ukuran (Bai & Lindhout 2007).
Laary dan Joanne (2007) melaporkan ada dugaan bahwa daerah asal tomat adalah Peru atau Meksiko. Meskipun belum ada bukti yang pasti mengenai waktu dan daerah asalnya, Meksiko diduga kuat merupakan daerah asal tomat, sedangkan Peru sebagai pusat keanekaragaman untuk kerabat liar.
2.2 Koleksi Plasma Nutfah Tomat
Syukur et al. (2012) menyatakan bahwa koleksi plasma nutfah merupakan
sumber kekayaan keragaman genetik bagi kegiatan pemuliaan tanaman dimana memiliki tujuan untuk mempelajari tingkat keragaman yang ada dan untuk menyelamatkan keragaman genetik. Menurut Sutjahjo (2013) keragaman yang tinggi pada peubah pertumbuhan vegetatif dan generatif merupakan potensi yang sangat baik sebagai material genetik dalam perakitan tomat unggul.
Koleksi plasma nutfah adalah hasil eksplorasi dari tempat dimana terdapat keragaman genetik yang tinggi (center of origin) atau dari tempat dimana tanaman
itu secara intensif dibudidayakan sejak lama (center of diversity) (Syukur et al.
Tabel 1 Deskripsi tiga puluh genotipe tomat lokal koleksi Laboratorium Genetik dan Pemuliaan Tanaman IPB
Nama genotipe Asal Tinggi
tanaman
Cherry Nusa Tenggara Timur Nusa Tenggara Timur 95,681 38,64 3
Gondol Lonjong Pasar/Komersil 79,794 177,23 4
Kali Acai Jayapura 60,102 132,02 2
Kefamenanu 3 Nusa Tenggara Timur 62,067 147,74 5 Kefamenanu 6 Nusa Tenggara Timur 43,750 80,00 5 Kefamenanu 7 Nusa Tenggara Timur 62,648 328,07 5 Kefamenanu 9 Nusa Tenggara Timur 96,467 410,75 3 Kefamenanu 12 Nusa Tenggara Timur 64,260 164,17 3 Kefamenanu 14 Nusa Tenggara TImur 63,667 175,88 3
Kemir Pasar/Komersil 60,167 197,14 3
Situbondo Bulat Kecil Situbondo 74,639 160,88 3
Situbondo Gelombang Situbondo 87,658 137,20 1
Tanah Datar Jayapura 78,917 118,33 10
Ket. Bentuk buah nomor 1=flattened, 2=slightly flattened, 3=circular, 4=rectangular,
5=cylindrical, 6=elliptic, 7=heart-shaped, 8=obovate, 9=ovate, 10=pear-shaped.
2.3 Penyakit Layu Bakteri
Penyakit layu bakteri disebabkan oleh bakteri tular tanah dan air yang bersifat nonfluoresens dari family Ralstonia (Setyari et al. 2013) dan merupakan
penyakit yang sangat merugikan (Hayward 1991; Fujiwara et al. 2008). Bakteri
ini semula termasuk ke dalam genus Pseudomonas, namun Yabuuchi et al. (1992)
menyatakan berdasarkan sebuah penelitian taksonomi spesies nonfluoresens
tertentu dari genus Pseudomonas bahwa genus Burkholderia diusulkan untuk terpisah dan nama Burkholderia solanacearum diusulkan. Penelitian selanjutnya
mengungkapkan bahwa R. solanacearum cukup berbeda dari anggota lain dari
genus tersebut sehingga genus baru diusulkan dengan nama Ralstonia (Yabuuchi
Ralstonia solanacearum biasanya menyerang area tropis dan subtropis
(Kelman 1998), serta memiliki inang berupa tanaman penting seperti tomat, terong, kentang, tembakau, cabai, pisang, kacang, dan jahe. Inang patogen ini diyakini berkisar 450 spesies (Swanson et al. 2005).
R. solanacearum merupakan patogen dengan kisaran inang yang sangat
luas, tetapi memiliki ras yang beragam yang hanya menyerang inang tertentu. Spesies yang tumbuh di daerah tropis, subtropis, dan daerah dengan temperatur tinggi memiliki kemungkinan rentan terhadap salah satu ras tersebut (James et al.
2003; Chandrashekara & Prasannakumar 2010).
Terserangnya tanaman di areal pertanaman terjadi secara acak tergantung kondisi lingkungan yang menguntungkan bagi patogen (Nguyen & Ranamukhaarachchi 2010). Infeksi R. solanacearum terjadi melalui luka pada
perakaran tanaman yang disebabkan kesalahan saat pindah tanam, nematoda, atau organisme lainnya (Yulianah 2007). Bakteri masuk ke jaringan tanaman bersamaan dengan terjadinya proses penyerapan hara dan air secara difusi oleh jaringan xylem. Selanjutnya bakteri akan menetap di pembuluh xylem dan merusak sel yang ditempatinya sehingga menyebabkan proses pengangkutan hara dan air terganggu oleh massa bakteri (Agrios 2005). R. solanacearum juga
memiliki kemampuan untuk menghancurkan sel-sel pembuluh xylem dengan mengeluarkan enzim penghancur yang mengandung selulosa dan pectin yang
dikenal dengan nama enzim selulase dan pektinase. Terganggunya proses angkut hara dan air akan menyebabkan tanaman layu dan mati (Agrios 2005).
Penyakit ini memiliki gejala serangan pertama berupa layunya daun termuda dan kelihatan lemah, terutama pada siang hari (Vanitha et al. 2009). Gejala
selanjutnya yaitu berupa perubahan warna batang menjadi coklat yang disebabkan terganggunya system jaringan angkut tanaman. Tanaman yang telah terserang akan mengeluarkan ooz bakteri apabila potongan batang direndam pada air (Nguyen & Ranamukhaarachchi 2010). Tanaman yang terserang juga cenderung lebih banyak membentuk akar adventif sampai setinggi bunga (Semangun 2007). Gejala umum serangan R. solanacearum adalah tanaman seperti kekurangan air,
daun muda pada pucuk tanaman menjadi layu, dan daun-daun tua menguning (Cavalcante et al. 1995). Secara keseluruhan gejala yang ditimbulkan merupakan
akibat dari terserangnya jaringan pembuluh xylem (Agrios 2005). 2.4 Pemuliaan Tanaman Tomat
Pemuliaan tanaman adalah perpaduan antara seni dan ilmu dalam merakit keragaman genetik suatu populasi tanaman tertentu menjadi lebih baik atau unggul dari sebelumnya. Pada mulanya pemuliaan tanaman dititikberatkan pada pemilihan atau seleksi karena yang memegang peranan adalah kemampuan pemulia tanaman untuk menilai atau meramalkan tanaman yang dapat menjadi varietas lebih unggul (Syukur et al. 2012). Tujuan pemuliaan tanaman secara lebih
Dalam mengoptimalkan kegiatan pemuliaan, ada beberapa cabang ilmu yang berkaitan, di antaranya: genetika, botani, fisiologi tanaman, agronomi, entomologi, patologi, biokimia, serta ilmu statistik dan komputer (Chahal & Gosal 2003). Kegiatan pemuliaan tanaman pada dasarnya diawali dengan kegiatan koleksi berbagai genotipe. Setelah dilakukan koleksi, tanaman-tanaman tersebut diseleksi sesuai dengan karakter-karakter yang diinginkan. Demi meningkatkan efektifitas seleksi kegiatan perluasan keragaman genetik perlu dilakukan, yang meliputi hibridisasi, mutasi, ataupun rekayasa genetika. Setelah perluasan genetik dilakukan maka langkah selanjutnya adalah seleksi. Metode seleksi yang dipilih tergantung dari tipe penyerbukan. Hasil seleksi kemudian akan dievaluasi untuk mengetahui kestabilan dari calon varietas tersebut (Syukur et al. 2012).
Tujuan dari program pemuliaan tomat bervariasi tergantung pada lokasi, kebutuhan, dan sumber daya. Secara umum, tujuan pemuliaan dalam tomat telah melalui empat tahap, yaitu pemuliaan untuk peningkatan hasil pada tahun 1970-an, masa simpan produk pada tahun 1980-1970-an, kualitas rasa tahun 1990-1970-an, dan kualitas gizi pada saat ini. Dalam rangka mencapai keuntungan yang memuaskan, petani diharuskan memproduksi buah dalam skala tinggi dengan biaya produksi serendah mungkin sehingga mayoritas tujuan pemuliaan fokus pada karakteristik yang mengurangi biaya produksi atau memastikan produksi dengan hasil dan kualitas yang tinggi (Bai & Lindhout 2007).
Hingga saat ini salah satu bidang pemuliaan tanaman yang masih terus diteliti adalah peningkatan resistensi, baik terhadap cekaman biotik ataupun abiotik. Menurut Bai dan Lindhout (2007) salah satu masalah yang paling menonjol dalam pemuliaan tomat adalah resistensi terhadap hama penyakit dimana tomat memiliki lebih dari 200 spesies hama patogen yang dapat menyebabkan kerugian ekonomi yang signifikan.
2.5 Penanda Molekuler
Kegiatan pemuliaan tanaman tergantung pada identifikasi dan pemanfaatan
variasi genetik. Penggunaan penanda molekuler untuk mengetahui ‘track loci’ dan
daerah genom tanaman sekarang merupakan kegiatan rutinitas pada kegiatan pemuliaan. Susunan penanda genetik seperti penanda berbasis morfologi, biokimia dan DNA telah digunakan di berbagai bidang, termasuk di bagian pengembangan hasil tanaman. Perbaikan genetik pada tanaman memerlukan informasi sifat ekonomi tanaman tersebut, seperti hasil produksi, ketahanan hama penyakit atau cekaman abiotik untuk menjadi target kegiatan pemuliaan tanaman (Langridge & Chalmers 2004; Jehan & Lakhanpaul 2006). Identifikasi genetik dengan pendekatan molekuler untuk mendapatkan informasi genetik terkait karakter yang diinginkan sangat dibutuhkan dalam kegiatan pemuliaan agar mendapat hasil yang tepat (Nuraida 2012). Selain itu menurut Prabakaran et al.
(2010) penanda molekuler merupakan cara tepat untuk mengetahui variasi genetik dan identitas kultivar. Sehgal et al. (2012) menyatakan bahwa penanda molekuler
Penanda molekuler memiliki peran penting dalam bidang bioteknologi dan studi genetik. Ada berbagai tipe penanda molekuler, yaitu yang tidak berbasis PCR (RFLP) dan yang berbasis PCR (RAPD, AFLP, SSR, SNP, dan lain sebagainya) (Kumar et al. 2009). Berbeda dengan pernyataan Jehan dan
Lakhanpaul (2006) yang mengemukakan penanda molekuler dapat diklasifikasi menjadi 3 kelas, yaitu: penanda berbasis hibridisasi (RFLP), penanda berbasis PCR (RAPD dan modifikasinya), dan penanda yang menggabungkan prinsip hibridisasi dan PCR (SSR, AFLP, dan modifikasi lainnya).
Dewasa ini penanda molekuler menjadi pilihan terdepan untuk studi keragaman genetik tanaman. Penanda molekuler hingga kini merupakan teknik yang menjanjikan, cepat, dan tergolong murah. Penanda molekuler yang ideal harus mampu memenuhi beberapa kriteria, yaitu: tingkat polimorfik yang tinggi, bersifat kodominan, selektif, reprodusibilitas tinggi, murah,dan mudah dilakukan (Kumar et al. 2009).
PCR (Polymerase Chain Reaction) sebagai teknik dasar dalam kegiatan
molekuler sangat membantu penggunaan penanda molekuler. PCR adalah teknik untuk memperbanyak untai DNA secara banyak dengan waktu yang relatif cepat (Innis et al. 1990). Teknik identifikasi berbasis PCR dapat digunakan untuk
mendeteksi keragaman pada tanaman (Jones et al. 1997).
Salah satu teknik penanda molekuler yang berbasis PCR adalah Simple Sequence Repeats (SSR). Menurut Powell et al. (1996) SSR adalah kelompok
urutan DNA berulang yang merupakan bagian yang signifikan dari genom eukariot. Bagian tersebut dapat menyajikan informasi penanda genetik yang sangat penting. SSR telah menjadi penanda yang sangat berguna dalam berbagai aspek studi genetika molekuler dalam dekade terakhir, di antaranya pada bidang: studi keanekaragaman genetik (Ashley et al. 2003), fingerprinting (Rongwen et al. 1995), studi ekologi-genetik (Li et al. 2000), dan marker-assisted selection
(Fazio et al. 2003). Penggunaan SSR semakin meluas dikarenakan penanda ini
bersifat kodominan, reproducible, banyak terdapat pada individu eukariot dan
3 SELEKSI TIGA PULUH GENOTIPE TOMAT LOKAL
TERHADAP KARAKTER KETAHANAN PENYAKIT
LAYU BAKTERI (
Ralstonia solanacearum
)
Selection of thirty local tomato genotypes to bacterial wilt (R. solanacearum) disease resistance
Abstrak
Penyakit layu bakteri yang disebabkan oleh bakteri Ralstonia solanacearum
merupakan penyakit penting pada tomat yang dapat menurunkan hasil hingga 35%. Budidaya menggunakan varietas tahan adalah langkah awal yang penting dalam pengendalian penyakit ini. Tujuan penelitian adalah untuk menyeleksi ketahanan genotipe tomat lokal koleksi terhadap penyakit layu bakteri. Bahan tanam yang digunakan meliputi tiga puluh genotipe tomat lokal yang dikoleksi dari berbagai daerah di Indonesia. Tanaman diinokulasi dengan menggunakan isolat lapang pada saat bibit berusia empat minggu setelah semai. Pengamatan selama tiga puluh hari pada karakter periode inkubasi, kejadian penyakit, area under the disease curve, dan persentase tanaman hidup menunjukkan bahwa
setiap genotipe memiliki respon ketahanan yang berbeda. Genotipe yang bersifat sangat tahan adalah Kudamati 1 dan yang bersifat sangat rentan adalah Lombok 4. Kata kunci: tomat, Ralstonia solanacearum, inokulasi
Abstract
Bacterial wilt disease caused by Ralstonia solanacearum is an important disease on tomato that could reduce productivity by up to 35%. Cultivate the resistant variety is an important early step in controlling this disease. This research aimed to select collected local tomato genotypes resistance to bacterial wilt. The materials were thirty local genotipes that collected from various regions in Indonesia. Plants were inoculated by field isolated inoculum at four weeks after seeding. A thirty days observation on the characteristics of incubation period, disease incident, area under the disease curve, and living plants percentage revealed that each genotype has different resistance responses. The results showed that the highly resistant and susceptible were Kudamati 1 and Lombok 4, respectively.
3.1 Pendahuluan
Budidaya tomat hingga saat ini masih memiliki banyak kendala di lapangan, baik gangguan biotik maupun abiotik. Terutama penanaman tomat di dataran rendah yang memiliki permasalahan berupa serangan penyakit layu bakteri yang dapat menyebabkan kerugian yang besar hingga gagal panen (Álvarez et al.
2010). Penyakit ini disebabkan oleh Ralstonia solanacearum. Menurut Agrios
(2005) dan Yamada et al. (2007), R. solanacearum menyerang areal pertanaman
yang memiliki suhu tinggi dan keberadaannya terbatas di daerah yang berhawa panas, sehingga menjadi patogen penting di daerah tropis dan subtropis. Patogen mudah menyebar melalui irigasi dan peralatan pertanian yang telah terkontaminasi (Yamada et al. 2007).
Tingginya tingkat kerusakan akibat patogen ini menjadikannya sebagai patogen paling berbahaya nomor dua di dunia (Mansfield et al. 2012). Patogen
2013) dan suku Solanaceae lainnya.
Menurut Ayana et al. (2011) dan Xue et al. (2011), sejauh ini tidak ada
pengendalian penyakit yang efektif secara umum. Beberapa pendekatan untuk mengendalikan serangan R. solanacearum telah dilakukan, seperti penggunaan
agens hayati (Xue et al. 2009) dan bakterisida. Penggunaan agen hayati yang
dapat mengendalikan jumlah populasi patogen ternyata belum mampu bekerja secara optimal menekan populasi patogen hingga akhir masa pertumbuhan (Maharina et al. 2014). Pengendalian dengan menggunakan bakterisida dalam
jangka waktu panjang akan menimbulkan dampak yang tidak diinginkan, seperti timbulnya resistensi patogen. Selain itu, pengendalian secara kimiawi pun juga sebenarnya belum efektif dalam menanggulangi serangan R. solanacearum
ataupun menyembuhkan tanaman yang telah terserang (Young et al. 2012).
Sejauh ini resistensi secara genetik sangat diharapkan demi pengendalian penyakit yang efektif dan ramah lingkungan dibandingkan penggunaan bahan kimia (Mejri
et al. 2012).
Tujuan penelitian ini adalah untuk menyeleksi genotipe tomat lokal yang tahan terhadap penyakit layu bakteri.
3.2 Bahan dan Metode
3.2.1 Waktu dan Tempat
Penelitian bertempat di Kebun Percobaan Leuwikopo pada Februari-April 2015. Pembibitan, penanaman, dan pemberian inokulum dilakukan di rumah plastik yang berada di ketinggian 250 m dpl.
3.2.2 Bahan Tanam
3, Lombok 4, Makasar 1, Makasar 2, Makasar 3, Makasar 4, Meranti 1, Meranti 2, Situbondo Bulat Kecil, Situbondo Gelombang, dan Tanah Datar). Benih disemai sebanyak lima belas benih dari masing-masing genotipe dalam tray persemaian
yang berisi media campuran tanah dan pupuk kandang dengan perbandingan 2:1 (v/v).
3.2.3 Persiapan Isolat R. solanacearum
Bakteri R. solanacearum yang digunakan merupakan isolat yang diambil
dari tanaman sakit di lapang. Pangkal batang tanaman Solanaceae yang terinfeksi dengan kemiringan 450 dan direndam dengan menggunakan aquades steril selama
24 jam. Tanaman yang mengeluarkan ooz bakteri digunakan sebagai sumber inokulum.
3.2.4 Uji Isolat R. solanacearum
Isolat R. solanacearum diuji dengan menggunakan media TTC (Triphenyl Tetrazolium Chloride). Koloni bakteri R. solanacearum dicirikan berwarna putih
susu dengan bagian tengah yang berwarna merah.
3.2.5 Inokulasi Bakteri R. solanacearum
Tanaman diinokulasi pada saat pindah tanam di usia 4 minggu. Tanaman dilukai dengan cara menggunting ujung akarnya, kemudian direndam dengan suspensi bakteri sebanyak 20 mL selama 30 menit. Selanjutnya tanaman ditanam di polybag berukuran 30 cm. Suspensi bakteri kemudian disiram ke tanaman. 3.2.6 Pengamatan
Pengamatan dilakukan 2 kali seminggu sejak 2 hari setelah inokulasi (hsi) selama 30 hari. Peubah yang diamati adalah :
1. Periode inkubasi
Yaitu waktu yang diperlukan bakteri untuk dapat menimbulkan gejala gangguan terhadap tanaman. Masa inkubasi diamati 2 hari sekali setelah inokulasi.
2. Kejadian penyakit
Yaitu pengamatan kejadian penyakit yang diamati mulai umur 2 HSI sampai usia 30 HSI. Kejadian penyakit diukur dengan menggunakan rumus:
Kejadian Penyakit = n x 100% ... (1) N
Keterangan : n
3. Area under the disease progress curve (AUDPC)
Yaitu suatu perhitungan kuantitatif untuk mempermudah mengambil kesimpulan mengenai intensitas serangan suatu penyakit (kejadian atau keparahan penyakit) yang berkembang dalam suatu periode waktu.
AUDPC = ∑ ( )
Nilai kejadian atau keparahan penyakit pada waktu ke-i Waktu ke-i
4. Tanaman Hidup
Persentase tanaman hidup per genotipe dihitung pada hari ketigapuluh setelah inokulasi.
5. Respon Ketahanan
Respon ketahanan setiap genotipe ditunjukkan berdasarkan kejadian penyakit menggunakan metode Peter et al. (1993) yang dimodifikasi
(Tabel 2).
Tabel 2 Respon ketahanan tomat terhadap penyakit layu bakteri (R. solanacearum) berdasarkan kejadian penyakit
Kejadian penyakit (%) Respon ketahanan 0 ≤ X < 5
3.3 Hasil dan Pembahasan
Pengamatan pada peubah menunjukkan bahwa genotipe yang diuji memberikan respon yang berbeda. Hasil pengamatan selama 30 hari menunjukkan bahwa dari 30 genotipe tomat lokal koleksi terdapat 1 genotipe sangat tahan (Kudamati 1), 8 genotipe tahan (Gondol Lonjong, Kemir, Kudamati 3, Lombok 3, Makasar 3, Situbondo Bulat Kecil, Situbondo Gelombang, dan Tanah Datar), dan 9 genotipe sangat rentan (Aceh 2, Aceh 3, Kefamenanu 9, Kefamenanu 12, Kefamenanu 14, Lombok 1, Lombok 2, Lombok 4, dan Meranti 2). Genotipe tahan dicirikan dengan tingkat kejadian penyakit yang rendah dan persentase tanaman hidup yang tinggi.
Tabel 3 Periode inkubasi (PI), Area Under the Disease Progress Curve (AUDPC),
Cherry Nusa Tenggara Timur
Ket. ST=sangat tahan, T=tahan, AT=agak tahan, AR=agak rentan, R=rentan, SR=sangat rentan.
Gambar 2 Grafik kejadian penyakit kelas sangat tahan tomat
Gambar 3 Grafik kejadian penyakit kelas tahan tomat
Gambar 4 Grafik kejadian penyakit kelas agak tahan tomat
Gambar 5 Grafik kejadian penyakit kelas agak rentan tomat
Gambar 6 Grafik kejadian penyakit kelas rentan tomat
Gambar 7 Grafik kejadian penyakit kelas sangat rentan tomat
Genotipe yang sangat rentan, seperti genotipe Lombok 4 dan Meranti 2 (Tabel 3), kematian dapat terjadi pada hari kedua setelah inokulasi tanpa menunjukkan gejala terlebih dahulu (Gambar 8.A1), sedangkan genotipe sangat tahan (Kudamati 1) tidak menunjukkan gejala apa pun (Gambar 8.A2). Genotipe yang tergolong rentan, agak rentan, dan agak tahan akan menampakkan gejala terhadap infeksi patogen (Gambar 8.B, 8.C, dan 8.D). Gejala awal serangan berupa layunya daun pertama tanaman (Gambar 8.B). Tanaman menunjukkan gejala ini umumnya ketika memasuki hari keempat setelah inokulasi. Layunya daun disebabkan tidak adanya pasokan air yang cukup ke bagian tanaman. Berkurangnya pasokan air terjadi karena patogen telah merusak dan menghambat kerja xylem (Agrios 2005). R. solanacearum secara agresif menyerang pembuluh
xylem. Gejala ini akan semakin terlihat pada saat tengah hari dimana suhu sedang mengalami kenaikan. Layunya daun seringkali dianggap hanya karena faktor kekurangan air, namun gejala tersebut akan dilanjutkan dengan gejala lainnya berupa munculnya akar adventif di sepanjang batang tanaman (Gambar 8.C). Akar adventif yang muncul menandakan bahwa jaringan xylem telah mengalami
kerusakan yang berat. Xylem yang telah dirusak oleh patogen akan mengalami
browning (Gambar 8.D). Pada saat gejala telah mencapai tahap browning maka
tanaman telah mengalami serangan yang parah. Tanaman akan mengeluarkan ooz bakteri apabila potongan batang dan akar direndam di air (Gambar 8.E). Ooz bakteri dipastikan merupakan bakteri R. solanacearum dengan menggunakan
media TTC. Menurut Chaudhry dan Rasshid (2011) bakteri R. solanacearum pada
media TTC dicirikan dengan warna pink yang dikelilingi oleh selaput berwarna putih susu (Gambar 8.F).
Gambar 8 A.Respon ketahanan tanaman muda terhadap pemberian inokulasi R.
solanacearum (4 hari setelah inokulasi) B. Gejala layunya daun muda pada
tanaman yang sudah besar (10 hari setelah inokulasi). C. Pembentukan akar
adventif pada tanaman dewasa yang terserang (30 hsi). D. Profil browning pada
batang dengan potongan vertikal. E. Ooz bakteri yang keluar dari batang
tanaman sakit. F. Koloni R. solanacearum pada media TTC
Patogen mulai menyerang tanaman melalui tahapan awal berupa kolonisasi pada perakaran dimana patogen masuk melalui organ perakaran tanaman yang terbuka. Menurut Agrios (2005) kesalahan pindah tanam dan nematoda merupakan hal yang dapat menyebabkan luka pada perakaran tanaman sehingga infeksi R. solanacearum pertama kali akan terjadi pada area tersebut (Zuluaga et al. 2015). Pada Percobaan ini tanaman sampel diberi pelukaan di ujung akar
dengan menggunakan gunting. Pelukaan dilakukan untuk memastikan bahwa patogen dapat menginfeksi tanaman sampel. Berdasarkan fakta bahwa patogen memanfaatkan sistem vascular untuk menyebar ke seluruh bagian tanaman (Agrios 2005) maka tanaman sampel direndam beberapa saat di dalam suspensi bakteri. Proses angkut air dan hara oleh jaringan xylem memungkinkan patogen terdistribusi ke dalam tubuh tanaman.
pembuluh xylem menyebabkan terputusnya distribusi air dan hara yang dibutuhkan tanaman sehingga mengakibatkan kelayuan.
Cepatnya proses infeksi dan kematian yang ditimbulkan menjadikan patogen ini sebagai patogen penting dunia. Menurut Champoiseau dan Momol (2009) serangan R. solanacearum memang sulit untuk dikendalikan dan belum
ada satu metode tunggal yang menunjukkan tingkat efisiensi 100% dalam menekan serangan patogen ini. Berbagai cara pengendalian telah diusahakan untuk dapat menekan serangannya, namun belum memberikan hasil yang memuaskan. Beberapa metode pengendalian yang penting sejauh ini meliputi teknik kontrol biologi, pengendalian kimiawi, kultur teknis, dan penanaman varietas tahan (Tahat & Sijam 2010).
Teknik kontrol biologi menggunakan bakteri endofit Staphylococcus epidermidis dan rizobacteria Pseudomonas fluorescens dilaporkan dapat menekan
serangan penyakit layu bakteri pada tomat (Nawangsih & Wardani 2014). Penggunaan kontrol biologi memang belum mampu menanggulangi penyakit ini secara menyeluruh, namun teknik ini juga merupakan kunci penting dalam pertanian berkelanjutan.
Pengendalian kimiawi tidak dapat memberikan hasil yang memuaskan dalam menekan serangan penyakit layu bakteri karena area serangan patogen yang terletak di dalam jaringan xylem. Patogen juga semakin sulit dikendalikan secara kimiawi mengingat patogen mampu bertahan hidup di tanah (Tahat & Sijam 2010). Pada pengendalian secara kultur teknis juga menghadapi kendala berupa luasnya inang dari patogen ini sehingga cara ini sulit untuk dilakukan.
Pengendalian dengan menggunakan varietas tahan sejauh ini memberikan harapan yang paling menjanjikan. Menurut Tahat dan Sijam (2010) tomat yang memiliki ketahanan mampu menekan perbanyakan patogen dan membatasi penyebaran patogen di dalam jaringan xylem sehingga patogen tidak mampu berpindah dari protoxylem ke bagian xylem primer atau bagian xylem lainnya. Berdasarkan fenomena tersebut maka penting untuk dilakukannya kegiatan seleksi demi mendapatkan varietas yang secara genetik tahan.
Percobaan seleksi dilakukan dengan memanfaatkan genotipe tomat lokal koleksi yang diperoleh dari berbagai daerah di Indonesia. Syukur et al. (2012)
dibudidayakan. Penggunaan genotipe tahan diharapkan dapat meminimalisir kerusakan dan kerugian yang ditimbulkan patogen.
3.4 Kesimpulan
Genotipe tomat lokal koleksi memiliki respon ketahanan yang beragam terhadap penyakit layu bakteri. Berdasarkan masa inkubasi dan nilai keparahan penyakit diketahui bahwa genotipe Kudamati 1 merupakan genotipe yang sangat tahan; Gondol Lonjong, Kemir, Kudamati 3, Lombok 3, Makasar 3, Situbondo Bulat Kecil, Situbondo Gelombang, dan Tanah Datar merupakan genotipe tahan; sedangkan genotipe Aceh 2, Aceh 3, Kefamenanu 9, Kefamenanu 12, Kefamenanu 14, Lombok 1, Lombok 2, Lombok 4, dan Meranti 2 merupakan genotipe yang sangat rentan.
3.5 Daftar Pustaka
Agrios GN. 2005. Plant Pathology. Ed ke-5. San Diego (US): Academic Press.
Álvarez B, Biosca EG, López MM. 2010. On the life of Ralstonia solanacearum,
a destructive bacterial plant pathogen. A. Méndez-Vilas (Ed.). pp. 267-279.
Aliye N, Fininsa C, Hiskias Y. 2008. Evaluation of rhizosphere bacterial antagonistsvfor their potential to bioprotect potato (Solanum tuberosum)
against bacterial wilt (Ralstonia solanacearum). Biological Control.
47:282-288.
Ayana G, Fininsa C, Ahmed S, Wydra K. 2011. Effects of soil amendment on bacterial wilt caused by Ralstonia solanacearum and tomato yields in
Ethiopia. Journal of Plant Protection Research. 51(1):74-76.
Bi-hao C, Jian-ju L, Yong W, Guo-jo C. 2009. Inheritance and identification of SCAR marker linked to bacterial wilt-resistance in eggplant. African Journal of Biotechnology. 8(20):5201-5207.
Champoiseau PG, Momol TM. 2009. Bacterial wilt of tomato [educational
modules]. Florida (US): University of Florida.
Chaudhry Z, Rashid H. 2011. Isolation and characterization of Ralstonia solanacearum from infected tomato plants of soan skesar valley of Punjab. Pak. J. Bot. 43(6):2979-2985.
Crus APZ, Ferreira V, Pianzzola MJ, Siri MI, Coll NS, Valss M. 2014. A novel, sensitive method to evaluate potato germplasm for bacterial wilt resistance using a luminescent Ralstonia solanacearum reporter strain. Molecular Plant-Microba Interactions. 27(2):277-285.
Grover A, Chakrabarti SK, Azmi W, Khurana AMP. 2012. Rapid method for isolation of PCR amplifiable genomic DNA of Ralstonia solanacearum
infested in potato tubers. Scientific Research. 2:441-446.
Jones JDG, Dangl JL. 2006. The plant immune system. Nature Publishing Group.
444:323-329.
Kumar R, Barman A, Jha G, Ray SK. 2013. Identification and establisment of genomic identify of Ralstonia solanacearum isolated from a wilted chili
plant at Tezpur, North East India. Current Science. 105(11):1571-1578.
Pada Budidaya Tanaman Tomat. Jurnal Produksi Tanaman.
1(6):506-513.
Mansfield J, Genin S, Magori S, Citovsky V, Sriariyanum M, Ronald P, Dow M, Verdier V, Beer SV, Machado MA, Toth I, Salmond G, Foster GD. 2012. Top 10 plant pathogenic bacteria in molecular plant pathology. Molecular Plant Pathology. 13(6):614-629.
Mejri S, Mabrouk Y, Voisin M, Delavault P, Simier P, Saidi M, Belhadj O. 2012. Variation in quantitative characters of faba bean after seed irradiation and associated molecular changes. African Journal of Biotechnology.
11(33):8383-8390.
Nawangsih AA, Wardani FF. 2014. Interaksi antara bakteri endofit dan bakteri perakaran pemacu pertumbuhan tanaman dalam menekan penyakit layu bakteri pada tomat. Jurnal Fitopatologi Indonesia. 10(5):145-152.
Ozaki K, Watabe H. 2009. Bacterial wilt of geranium and portulaca caused by
Ralstonia solanacearum in Japan. Bull. Minamikyushu Univ. 39A:67-71.
Peter RA, Gopalakrishnan TR, Rajan S, Kumar SPG. 1993. Breeding for Resistance to Bacterial Wilt in Tomato, Eggplant and Pepper. Di dalam: Hartman GL, Hayward AC, editor. Bacterial Wilt. Proceeding of an International Conference Held at Kaoshiung, 28-31 Okt 1992. AVRDC, ACIAR ICRISAT CIP and Rothamsted Experimental Station. hlm 183-190.
Rodrigues LMR, Destéfano SAL, Diniz MCT, Comparoni R, Neto JR. 2011. Pathogenicity of Brazilian strains of Ralstonia solanacearum in Strelitzia reginae seedlings. Tropical Plant Pathology. 36(6):409-413.
Siri MI, Sanabria A, Pianzzola MJ. 2011. Genetic diversity and aggressiveness of
Ralstonia solanacearum strains causing bacterial wilt of potato in
Uruguay. Plant Disease. 95(10):1292-1301.
Syukur M, Sujiprihati S, Yunianti R. 2012. Teknik Pemuliaan Tanaman. Bogor
(ID): Penebar Swadaya.
Tahat MM, Sijam K. 2010. Ralstonia solanacearum: the bacterial wilt caused
agent. Asian Journal of Plant Science. 9(7):385-393.
Xue QY, Chen Y, Li SM, Chen LF, Ding GC, Guo DW, Guo JH. 2009. Evaluation of the strains of Acinetobacter and Enterobacter as potential
biocontrol agents against Ralstonia wilt of tomato. Biol. Control.
48:252-258.
Xue QY, Yin YN, Yang W, Heuer H, Prior P, Guo JH, Smalla K. 2011. Genetic diversity of Ralstonia solanacearum strains from China assessed by
PCR-based fingerprints to unravel host plant-and-site-dependent distribution patterns. FEMS. Microbiol. Ecol. 75:507-519.
Yamada T, Kawasaki T, Nagata S, Fujiwara A, Usami S, Fujie M. 2007. New bacteriophages that infect the phytopathogen Ralstonia solanacearum. Mikrobiologi. 153:2630-2639.
Young BJ, Wu J, Lee HJ, Jo EJ, Murugaiyan SK, Chung E, Lee SW. 2012. Biocontrol potential of a lytic bacteriophage PE204 against bacterial wilt of tomato. J. Microbiol. Biotechnol. 22(12):1613-1620.
4 STUDI PEWARISAN KARAKTER KETAHANAN
TERHADAP PENYAKIT LAYU BAKTERI
(
Ralstonia solanacearum
) PADA TOMAT
Inheritance of Bacterial Wilt (Ralstonia solanacearum) Disease Resistance Trait on Tomat
Abstrak
Ralstonia solanacearum penyebab penyakit layu bakteri memiliki kisaran
inang yang luas dan susah dikendalikan. Penggunaan varietas tahan merupakan upaya pengendalian penyakit yang efektif, efisien, ekonomis, dan ramah lingkungan. Tujuan penelitian ini adalah untuk mempelajari pola pewarisan sifat ketahanan terhadap penyakit layu bakteri pada tomat. Percobaan menggunakan Kudamati 1 (genotipe tahan) dan Lombok 4 (genotipe rentan) untuk membentuk populasi F1, F1R, dan F2. Pengamatan pada populasi F1 dan F1R menunjukkan bahwa karakter ini bersifat dominan tanpa dipengaruhi efek maternal dengan derajat dominansi tidak sempurna. Berdasarkan uji χ2 pada populasi F2, segregasi memiliki rasio 9:7 (tahan : rentan) yang menandakan bahwa karakter merupakan karakter kualitatif yang dikendalikan oleh dua pasang gen mayor dengan aksi gen duplikat resesif epistasis.
Kata kunci: tomato, Ralstonia solanacearum, pola pewarisan, dominan Abstract
Ralstonia solanacearum causes of bacterial wilt disease has a wide host range and difficultly controlling. Cultivate the resistant variety is an effective, efficient, economical, and environmentally friendly controlling. The objective this research was to study of inheritance to bacterial wilt disease resistance on tomato. This experiment used Kudamati 1 (resistant genotype) and Lombok 4 (susceptible genotype) for generate F1, F1R, and F2 population. Observation on F1 dan F1R population showed that the character is dominant without maternal effect with the degree of dominance is uncomplete. Based on χ2 testing in F2 population, the rasio segregation was 9:7 (resistant : susceptible) which indicated the character is qualitative character controlled by two genes with duplicate recessive epistasis.
4.1 Pendahuluan
Penyakit layu bakteri yang disebabkan oleh bakteri Ralstonia solanacearum merupakan salah satu penyakit penting pada tanaman tomat. Patogen tersebar luas di daerah tropis dan subtropis, serta memiliki kisaran inang yang luas sehingga sulit untuk dikendalikan. Budidaya menggunakan varietas tahan merupakan langkah awal yang sangat menentukan keberhasilan budidaya sehingga sangat penting untuk mengetahui informasi pewarisan mengenai karakter ketahanan dalam tujuan mempermudah perakitan varietas tahan tersebut. Mejri et al. (2012)
menyatakan bahwa pengendalian penyakit yang dimulai dari resistensi genetik merupakan pilihan yang tepat. Dengan kata lain budidaya varietas tahan merupakan pengendalian yang efektif.
Varietas tahan dirakit melalui kegiatan pemuliaan tanaman yang mana akan efektif apabila informasi pewarisan karakter telah diketahui. Berbagai penelitian sejauh ini memberikan informasi jumlah gen pengendali yang berbeda. Grimault
et al. (1995) melaporkan bahwa karakter ketahanan tomat terhadap penyakit layu
bakteri dikendalikan oleh satu gen. Berbeda dengan hasil penelitian tersebut, Osiru et al. (2001) menyatakan dikendalikan 2 gen dan Yue et al. (1995) tiga gen.
Selain itu Shou et al. (2006) menyatakan bahwa karakter ketahanan penyakit layu
bakteri pada tomat juga dipengaruhi oleh efek maternal.
Sejauh ini beberapa penelitian mengenai informasi pewarisan karakter ketahanan tomat terhadap penyakit layu bakteri telah dilakukan, namun belum memberikan informasi yang konsisten, sehingga masih perlu dilakukan konfirmasi demi mendapatkan informasi yang sesuai dengan varietas tomat yang dibudidayakan.
Tujuan penelitian ini adalah untuk mempelajari pola pewarisan karakter ketahanan terhadap penyakit layu bakteri pada tomat.
4.2 Bahan dan Metode
4.2.1 Waktu dan Tempat
Penelitian dilaksanakan di Kebun Percobaan Leuwikopo dari bulan Mei-Desember 2015. Pembibitan, penanaman, dan pemberian inokulum dilakukan di rumah plastik yang berada di ketinggian 250 m dpl
4.2.2 Pembentukan Populasi Dasar
Pembentukan populasi dasar meliputi F1, F1R, dan F2 dengan memanfaatkan genotipe Kudamati 1 sebagai tetua tahan dan genotipe Lombok 4 sebagai tetua rentan. Benih tetua disemai dalam tray persemaian yang berisi media campuran
tanah dan pupuk kandang dengan perbandingan 2:1 (v/v). Bibit dipelihara hingga berumur 3 minggu dan siap dipindahkan di polybag berukuran 30 x 30 cm.
Pemeliharaan yang dilakukan meliputi pemupukan, penyiraman, pengendalian organisme pengganggu tanaman, dan pemberian ajir. Tanaman diajir dan diikat untuk mencegah kemungkinan roboh.
4.2.3 Studi Pewarisan Sifat Ketahanan Terhadap Penyakit Layu Bakteri Bahan percobaan yang digunakan adalah populasi dasar yang terdiri dari P1, P2, F1, F1R dan F2. P1, P2, F1, F1R ditanam sebanyak 15 tanaman. Jumlah minimum tanaman F2 ditentukan berdasarkan perhitungan populasi minimum dengan menggunakan rumus:
Peluang kegagalan mendapatkan genotipe yang diinginkan
Berdasarkan rumus di atas dengan asumsi bahwa ketahanan terhadap penyakit layu bakteri dikendalikan oleh tiga gen, maka jumlah tanaman minimum pada populasi F2 adalah sebagai berikut:
n = (log F) (log q) = log (63/64) log 0.05 = -1.30103 -0.00684 = 191
Populasi F2 yang digunakan dalam percobaan sebanyak 215 tanaman.
Benih populasi P1, P2, F1, F1R dan F2 disemai dalam tray persemaian yang
berisi media campuran tanah dan pupuk kandang dengan perbandingan 2:1 (v/v). Bibit dipelihara hingga berumur 4 minggu dan dipersiapkan untuk pindah tanam sekaligus inokulasi. Inokulum R. solanacearum yang digunakan merupakan isolat
yang diambil dari tanaman sakit di lapang. Tanaman Solanaceae yang sakit dipotong pangkal batangnya dengan kemiringan 450 dan direndam dengan menggunakan aquades steril selama 24 jam. Tanaman yang mengeluarkan ooz bakteri digunakan sebagai sumber inokulum. Isolat R. solanacearum diuji dengan
menggunakan media TTC (Triphenyl Tetrazolium Chloride). Koloni bakteri R. solanacearum dicirikan berwarna putih susu dengan bagian yang berwarna merah
di tengahnya.
Tanaman yang akan diinokulasi dilukai dengan cara menggunting ujung akarnya, kemudian direndam dengan suspensi bakteri sebanyak 20 mL selama 30 menit. Selanjutnya tanaman ditanam di polybag berukuran 30 cm. Suspensi bakteri kemudian disiram ke tanaman. Pengamatan dilakukan 2 kali seminggu sejak 2 hari setelah inokulasi selama 30 hari. Peubah yang diamati adalah :
1. Periode inkubasi
Yaitu waktu yang diperlukan bakteri untuk dapat menimbulkan gejala gangguan terhadap tanaman. Masa inkubasi diamati 2 hari sekali setelah inokulasi.
2. Kejadian penyakit
Yaitu pengamatan kejadian penyakit yang diamati mulai umur 2 HSI sampai usia 30 HSI. Kejadian penyakit diukur dengan menggunakan rumus:
Kejadian Penyakit = n x 100% ... (4) N
Keterangan : n
3. Area under the disease progress curve (AUDPC)
Yaitu suatu perhitungan kuantitatif untuk mempermudah mengambbil kesimpulan mengenai intensitas serangan suatu penyakit (kejadian atau keparahan penyakit) yang berkembang dalam suatu periode waktu.
= Nilai kejadian atau keparahan penyakit pada waktu ke-i Waktu ke-i 4. Tanaman Hidup
Persentase tanaman hidup per genotipe dihitung pada hari ketigapuluh setelah inokulasi.
5. Respon Ketahanan
Respon ketahanan setiap genotipe ditunjukkan berdasarkan kejadian penyakit menggunakan metode Peter et al. (1993) yang dimodifikasi
(Tabel 4).
Tabel 4 Respon ketahanan tomat terhadap penyakit layu bakteri (R. solanacearum) berdasarkan kejadian penyakit
Kejadian penyakit (%) Respon ketahanan 0 ≤ X < 5
Data peubah hasil pengamatan akan dianalisa, yang meliputi: 1. Efek maternal
Pengaruh tetua betina pada pewarisan karakter ketahanan terhadap penyakit layu bakteri pada tomat dianalisis dengan cara membandingkan tanaman F1 dan F1R yang telah diinokulasi. Peubah yang digunakan meliputi periode inkubasi, kejadian penyakit, AUDPC, dan respon ketahanan.
2. Derajat dominansi
Pendugaan aksi gen pengendali karakter ketahanan tomat terhadap penyakit layu bakteri dilakukan dengan penghitungan derajat dominansi menggunakan rumus pendugaan potensi rasio (hp) (Petr 1959). Peubah yang digunakan adalah persentase tanaman hidup.
hp = ̅̅̅̅ - ̅̅̅̅ ... (6)
Berdasarkan nilai potensi rasio maka aksi gen diklasifikasikan seperti pada Tabel 5.
Tabel 5 Klasifikasi derajat dominansi berdasarkan nilai potensi rasio (hp) Kisaran nilai hp Derajat dominansi
hp = 0
hp = 1 atau hp = -1 0 < hp < 1
-1 < hp < 0 hp > 1 atau hp < -1
Tidak ada dominansi
Dominan atau resesif sempurna Dominan tidak sempurna
Resesif dengan aksi gen tidak penuh Overdominan
3. Estimasi jumlah gen pengendali
Sebaran frekuensi populasi F2 diuji untuk melihat sebaran yang dimiliki normal atau tidak dengan peubah respon ketahanan. Pendugaan jumlah gen pengendali berdasarkan sebaran frekuensi populasi F2 dibagi menjadi 2 kelompok, yaitu:
1) Sifat kuantitatif, apabila data menyebar secara kontinyu.
2) Sifat kualitatif, apabila data menyebar secara diskontinyu sehingga pendugaan jumlah gen dilakukan dengan menggunakan uji Chi
Kuadrat (χ2) untuk menguji kesesuaian nilai pengamatan dengan nilai
harapan. Jika nilai χ2 hitung lebih kecil dari χ2 tabel, maka sebaran populasi F2 mengikuti nisbah fenotipik yang diharapkan. Penghitungan dilakukan dengan menggunakan Microsoft Excel 2010.
χ2 = ∑
... (7)
Keterangan : Oi
Ei
=
= Nilai pengamatan Nilai harapan
Tabel 6 Nisbah fenotipik frekuensi karakter resistensi tanaman terhadap penyakit yang dikendalikan oleh gen mayor dalam populasi bersegregasi F2
1. Ketahanan dikendalikan 1 pasang gen a. Dominan penuh
2. Ketahanan dikendalikan 2 pasang gen
a. Dominan penuh pada kedua lokus A dan B 9 3 3 1 b. Resesif epistasis, aa epistatik terhadap B dan b 9 3 - 4 c. Dominan epistasis, A epistatik terhadap B dan b 12 - 3 1 d. Dominan dan resesif epistasis, A epistatik terhadap B
dan b; bb epistatik terhadap A dan a 13 - - 3 e. Duplikat resesif epistasis, aa epistatik terhadap B dan
b; bb epistatik terhadap A dan a 9 - - 7 f. Duplikat dominan epistasis, A epistatik terhadap B; B
epistatik terhadap A dan a 15 - - 1
g. Interaksi duplikat 9 6 - 1
h. Interaksi kompleks 10 3 - 3
3. Ketahanan dikendalikan oleh 3 pasang gen. Interaksi epistasis:
Studi pewarisan dilakukan dengan menggunakan Kudamati 1 (tetua tahan) dan Lombok 4 (tetua rentan). Populasi F1 dan F1R diuji untuk mendapatkan informasi mengenai efek tetua betina dan derajat dominansi sedangkan populasi F2 untuk mengetahui jumlah gen pengendali karakter tersebut.
Tabel 7 menunjukkan populasi F1 dan F1R memiliki respon ketahanan yang sama, yaitu tahan. Periode inkubasi yang lama juga memperkuat dugaan bahwa kedua populasi termasuk ke dalam kategori tahan.
Hasil pengamatan pada populasi F1 dan F1R menunjukkan tidak ada perbedaan yang dapat membuktikan bahwa tidak adanya pengaruh tetua betina dalam pewarisan karakter ketahanan terhadap penyakit layu bakteri. Analisis efek tetua betina dilakukan dengan membandingkan hasil pengamatan antara populasi F1 dan F1R tanpa menggunakan uji T dikarenakan data pengamatan tidak memungkinkan untuk dilakukan analisis uji T berdasarkan fakta bahwa tanaman tomat yang terserang penyakit layu bakteri tidak dapat dilakukan pengkelasan (scoring) berdasarkan keparahan penyakit. Ralstonia solanacearum yang mampu
Tabel 7 Periode Inkubasi (PI), Area Under the Disease Progress Curve
(AUDPC), Kejadian Penyakit (KP), Tanaman Hidup (TH), dan respon ketahanan populasi tomat yang diuji
Nama genotipe PI (hari) AUDPC KP (%) TH (%) Respon ketahanan
Ket. ST=sangat tahan, T=tahan, AT=agak tahan, AR=agak rentan, R=rentan, SR=sangat rentan.
Dari pengujian populasi F1 dan F1R juga dapat diduga bahwa karakter ketahanan bersifat dominan. Pendekatan untuk menduga derajat dominansi dilakukan dengan menggunakan hasil pengamatan persentase tanaman hidup, yang meliputi: nilai tengah kedua tetua, rata-rata nilai tetua tertinggi (tetua tahan), dan rata-rata nilai F1. Hasil perhitungan dengan menggunakan rumus Petr (1959) menunjukkan nilai hp= 0.6. Nilai potensi rasio tersebut menggambarkan rata-rata persentase tanaman hidup pada populasi F1 berada pada posisi tetua tahan. Hal ini menunjukkan bahwa karakter ketahanan tomat terhadap penyakit layu bakteri dikendalikan oleh dominansi tidak sempurna.
Frekuensi tanaman hidup yang menunjukkan respon ketahanan pada kedua tetua memiliki sebaran nilai yang besar. Sebaran F1 dan F2 mengarah pada tetua tahan (Gambar 9, 10, 11, 12, dan 13). Hasil pengamatan pada populasi F2 yang berjumlah 215 tanaman menunjukkan dua respon ketahanan, yaitu tahan (hidup) sebanyak 121 tanaman dan rentan (mati) sebanyak 94 tanaman. Sebaran data yang diperolah menunjukkan bahwa karakter ketahanan terhadap penyakit layu bakteri bersifat diskontinyu, yang menandakan bahwa karakter ini merupakan sifat kualitatif.
Pendugaan jumlah gen pengendali dilakukan menggunakan analisis Mendel melalui uji Chi Square mengingat karakter bersifat kualitatif.
Gambar 9 Histogram sebaran frekuensi tanaman hidup pada populasi P1, P2, F1, dan F2.
Pengelompokan berdasarkan hidup mati sampel pada akhir pengamatan pada hari ke-30 setelah inokulasi
0
15
0 15
tahan rentan
Populasi P2
12
3
0 15
tahan rentan
Populasi F1
121
94
0 215
tahan rentan
Gambar 10 Respon ketahanan tanaman pada hari ke-2 setelah inokulasi
Gambar 11 Respon ketahanan tanaman pada hari ke-10 setelah inokulasi
Gambar 12 Respon ketahanan tanaman pada hari ke-20 setelah inokulasi