IDENTIFIKASI SECARA MOLEKULAR KANDIDAT MUTAN
Gα
DARI
KEDELAI KULTIVAR SLAMET BERDASARKAN mRNA
HAYATUL FAJRI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
HAYATUL FAJRI. Identifikasi Secara Molekular Kandidat Mutan Gen Gα dari Kedelai Kultivar Slamet berdasarkan mRNA. Dibimbing oleh UTUT WIDYASTUTI SUHARSONO dan ARIS TJAHJOLEKSONO.
Protein heterotrimerik G terdiri atas tiga subunit yaitu α, , dan (Gα, G dan G ) yang terletak di membran internal sel. Protein Gα teraktivasi ketika adanya cekaman biotik atau abiotik dari luar sel. Protein Gα diduga berperan terhadap ketahanan cekaman Aluminium (Al). Kedelai kultivar Slamet merupakan kedelai toleran Al yang memiliki satu kopi gen Gα. Irradiasi sinar gamma pada biji kedelai kultivar Slamet menyebabkan mutasi random pada kedelai kultivar Slamet generasi M2. Kandidat mutan Gα M2 diseleksi berdasarkan keadaan stomata menutup pada siang hari dan tinggi tanaman. Mutan Gα M4 nomor 338 dan 103 mengalami kerusakan jaringan sel akar lebih besar dibandingkan tanaman non mutan. Tanaman nomor 338/3S1.174/12/14 (M4) dan 103/3S3.103/8/5/1/4 (M5) mengalami mutasi gen Gα pada ruas U3-L4 (ekson 5 – ekson 11). Sedangkan tanaman nomor 344/3S1.196/14/1/8 (M4) diduga mutan gen Gα. Ketiga nomor tersebut ditanam kembali sampai generasi ke 6 dan diseleksi kandidat mutan Gα berdasarkan fenotipe lebih pendek dari pada Slamet non mutan dan stomata menutup >50%. Dipilih empat nomor dari M5 dan tiga nomor dari M6 untuk diidentifikasi mutan Gα bedasarkan mRNA.
Tanaman M5 nomor 344/3S1.196/14/1/8/7 mengalami penurunan ekspresi gen Gα pada ruas L5-L4 (ekson 7- ekson 11). Generasi selanjutnya (M6) yaitu nomor 344/3S1.196/14/1/8/7/5 mengalami mutasi gen Gα pada ruas L1 sampai L3 (ekson 2 – ekson 6). Penurunan ekspresi gen
Gα dapat terjadi karena adanya mutasi pada promotor gen Gα. Tiga tanaman M5 lain diduga mengalami mutasi gen Gα. Dua tanaman M6 yaitu nomor 103/3S3.103/8/5/1/4/8 mengalami mutasi pada ruas L3-L2 (ekson 6 – ekson 15) dan nomor 103/3S3.103/8/5/1/4/9 pada ruas U3-L4 (ekson 5 – ekson 11). Mutasi yang terjadi pada dua nomor ini adalah penurunan ekspresi gen yang ditandai dengan lebih tipisnya pita gen Gα dibandingan tanaman Slamet non mutan.
ABSTRACT
HAYATUL FAJRI. Molecular Identification of Mutant Candidate of Gα Gene from Soybean cv. Slamet Based on mRNA. Supervised by UTUT WIDYASTUTI SUHARSONO and ARIS TJAHJOLEKSONO.
Heterotrimeric G proteins composed of α, , and subunits, are bound to the inside surface of the cell membrane. Gα proteins will be activated by extracellular stimuli (biotic or abiotic stress). It is presumably Gα proteins playing an important role in aluminum (Al)-stress tolerance. Soybean cv. Slamet was considered as a tolerant plant to Al and has a single copy of Gα gene. Gamma irradiation to soybean cv. Slamet seeds caused a random chromosomal mutation of second generation plants (designated as M2). The Gα mutant candidates were selected based on stomatal closure in the daylight and dwarf. Gα mutant candidates M4, number 338 and 103 have severe root cell tissues damage compared to non mutant ones. Plant number 338/3S1.174/12/14 (M4) and 103/3S3.103/8/5/1/4 (M5) have a Gα gene mutation at the U3-L4 sequences (exon 5 – exon 11). Whereas, plant number 344/3S1.196/14/1/8 (M4) is assumed as Gα gene mutant. All of those plants were replanted until sixth generation. The candidates of Gα mutant selected have more than fifty percent of closed stomata and dwarf. Four number of M5 plants and three number of M6 plants were selected for Gα mutant identification based on mRNA analysis.
IDENTIFIKASI SECARA MOLEKULAR KANDIDAT MUTAN
Gα
DARI
KEDELAI KULTIVAR SLAMET BERDASARKAN mRNA
HAYATUL FAJRI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul : Identifikasi Secara Molekular Kandidat Mutan
Gα
dari Kedelai Kultivar
Slamet Berdasarkan mRNA
Nama : Hayatul Fajri
NIM : G34062152
Disetujui,
Diketahui,
Ketua Departemen Biologi
Dr. Ir. Ence Darmo Jaya Supena, M.Si
NIP. 19641002 198903 1 002
Tanggal Lulus :
Pembimbing I
Dr. Ir. Utut Widyastuti, M.Si
NIP. 19640517 198903 2 001
Pembimbing II
KATA PENGANTAR
Puji serta syukur penulis ucapkan kepada Allah SWT, atas Rahmat dan HidayahNya penulis dapat menyelesaikan laporan penelitian yang berjudul Identifikasi Secara Molekular Kandidat Mutan Gα dari Kedelai Kultivar Slamet Berdasarkan mRNA.
Penulis menyampaikan ucapan terima kasih kepada Dr. Ir. Utut Widyastuti, M.Si dan Dr. Ir. Aris Tjahjoleksono, DEA sebagai pembimbing atas bimbingan, waktu, nasehat dan sarana yang telah diberikan. Terima kasih kepada Hibah Bersaing Dikti atas nama Dr. Utut Widyastuti yang telah memberikan dana penelitian. Terima kasih kepada kepala Pusat Pengembangan Sumber Daya Hayati dan Bioteknologi (PPSHB) beserta seluruh staf Labolatorium Biotechnological Research Indonesia Netherland (BIORIN) atas sarana, prasarana dan bantuan selama penulis melakukan penelitian. Terima kasih kepada Dr. Ir. Tatik Chikmawati, M.Si atas saran yang diberikan sehingga tulisan ini menjadi lebih baik. Terima kasih kepada Bapak Abdul Mulya, Mbak Pepi Elvavina dan Bapak Adi atas bantuan, nasehat dan kerjasamanya.
Penulis juga menyampaikan ungkapan terima kasih kepada Ibu penulis yang selalu mencurahkan doa, kasih sayang, perhatian dan harapan sehingga penulis mampu menyelesaikan tulisan ini. Terima kasih juga penulis sampaikan kepada kakak: Del Yuza, Ardiansyah, Yulfi Zawarnis dan Hidayatunnismah serta adik Rahma atas dukungan dan perhatiannya. Terima kasih untuk teman-teman Biologi 43 dan Biorin Crews: Bapak Yustinus Ulung, Bapak Muzuni, Ibu Saleha Hanum, Ibu Ratna Yuniati, Ibu Sri Listiyowati, Mbak Ika Atifah Zahro, Mbak Ulfah Mushofa, Mbak Anita Theresia, Mbak Novi, Mbak Nurul, Mbak Medikca, Kak Luria, Kak Muchdar Davis, Indah, Laila, Lita dan Iin atas kerjasama, bantuan, saran, dan kebersamaannya.
Karya ilmiah ini penulis persembahkan untuk Almarhum Ayah tercinta.
Bogor, Juli 2011
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 10 Januari 1988 di Bukittingi. Penulis merupakan anak kelima dari Zuwirda dan Almarhum Yusran. Penulis lulus dari SMA Negeri 6 Kota Bogor pada tahun 2006. Pada tahun yang sama penulis diterima sebagai mahasiswa IPB melalui jalur Ujian Seleksi Masuk IPB (USMI) dan tahun berikutnya diterima di Mayor Biologi.
DAFTAR ISI
Halaman
DAFTAR GAMBAR... viii
DAFTAR TABEL... viii
DAFTAR LAMPIRAN... viii
PENDAHULUAN... 1
Latar belakang penelitian... 1
Tujuan penelitian... 1
BAHAN DAN METODE... 1
Waktu dan tempat... 1
Bahan dan alat... 1
Metode penelitian... 2
Penanaman biji dan seleksi kandidat mutan gen Gα... 2
Isolasi RNA Total... 2
Sintesis cDNA... 2
Identifikasi kandidat mutan dengan PCR... 2
HASIL... 3
PEMBAHASAN... 4
SIMPULAN... 5
SARAN... 5
DAFTAR PUSTAKA... 5
DAFTAR GAMBAR
Halaman
1 Panjang basa nitrogen antar primer cDNA heterotrimerik Gα... 4 2 Ekspresi gen Gα dan aktin generasi M5 pada kandidat mutan Slm1-5, Slm2-5, Slm3-5,
dan Slm4-5... 4
3 Ekspresi gen Gα dan aktin generasi M6 pada kandidat mutan Slm1-6... 4 4 Ekspresi gen Gα dan aktin generasi M6 pada kandidat mutan Slm3-6 dan Slm2-6... 4
DAFTAR TABEL
1 Sekuen basa nitrogen pada primer yang digunakan... 3
2 Keadaan stomata menutup dan tinggi tanaman... 3
DAFTAR LAMPIRAN
1 Hasil seleksi kandidat mutan berdasarkan persentase stomata menutup >50% dan tanaman pendek... 8
2 Komposisi buffer MOPS dan gel agarosa 1% (b/v)... 9
PENDAHULUAN
Latar belakang penelitian
Protein G berdasarkan komposisi dan fungsinya diklasifikasikan menjadi protein G kecil dan protein heterotrimerik G (Ma 1994). Protein heterotrimerik G terdiri atas tiga subunit yaitu α, dan (Gα, G dan G ) (Ma 1994) dan dapat diaktifasi dengan adanya sinyal ekstraseluler (Albert et al. 2002). Protein heterotrimerik G adalah mediator yang mengirimkan sinyal ekstraseluler melalui molekul reseptor ke molekul efektor (Fujisawa et al. 2001). Sinyal ekstraseluler menyebabkan protein heterotrimerik G aktif dan akan mengubah Guanin Diphospat (GDP) yang berlekatan dengan Gα menjadi Guanin Triphospat (GTP). Terikatnya GTP pada Gα menyebabkan Gα berpisah dari dimer G dan kembali membentuk trimer saat inaktif (Perfush-Barbeoch et al. 2004).
Kedelai kultivar Slamet merupakan tanaman toleran aluminium (Al) (Jusuf & Suharsono 2006). Salah satu sinyal ekstraseluler yang dapat mempengaruhi regulasi seluler pada tanaman adalah cekaman Al. Kedelai kultivar Slamet memiliki satu kopi gen Gα (Suharsono & Suharsono 2004). Mastoparan berperan sebagai aktivator protein Gα dan dapat meningkatkan sistem pertahanan tanaman kedelai kultivar Lumut yang peka terhadap cekaman Al (Jusuf & Suharsono 2006), melalui pertambahan panjang akar, penurunan kandungan Al dan kalosa, dan peningkatan integritas membran (Srimulyati 2007).
Hartini (2008) telah melakukan induksi dengan menggunakan irradiasi sinar gamma dosis 0.1 dan 0.3 kGy pada kedelai kultivar Slamet dan Lumut untuk mendapatkan mutan
Gα. Irradiasi tersebut menghasilkan sepuluh kandidat mutan Gα generasi ke-2 (M2) dengan ciri kondisi stomata menutup. Kandidat mutan dari kultivar lumut dihasilkan dari dosis irradiasi 0.3kGy, sedangkan kandidat mutan dari kultivar Slamet dihasilkan dari dosis irradiasi 0.1 dan 0.3 kGy. Dari sepuluh tanaman tersebut, baru satu kandidat mutan yang telah dianalisis secara molekular dan menunjukkan adanya mutasi gen Gα.
Selanjutnya kesepuluh kandidat mutan generasi M2 tersebut ditanam masing-masing sebanyak 10-15 tanaman dan dilakukan seleksi berdasarkan kondisi stomata yang menutup (Perfush-Barbeoch et al. 2004) dan kerdil (Fujisawa et al. 1999; Ma 2001) pada generasi M3. Biji dari kandidat mutan
generasi M3 selanjutnya ditanam untuk mendapatkan tanaman generasi M4. Kandidat mutan M4 untuk setiap nomor dipilih berdasarkan kondisi stomata yang menutup >50% dan pendek. Limbong (2010) telah berhasil menseleksi tiga generasi (M2 sampai M4) dan mendapatkan 26 tanaman yang diduga merupakan kandidat mutan Gα dengan ciri stomata menutup >50%. Identifikasi mutan berdasarkan keberadaan gen Gα telah dilakukan berdasarkan DNA genom dari tanaman kandidat mutan.
Dua tanaman M4 yaitu nomor 338/3S1.174/12/1/4 dan 103/3S3.103/8/5/1/4 merupakan mutan Gα pada ruas U3-L4. Tanaman nomor 344/3S1.196/14/1/8 diduga mengalami mutan gen Gα. Identifikasi mutan
Gα berdasarkan DNA genom tersebut menghasilkan fragmen (bukan gen utuh) gen
Gα. Oleh karena itu, perlu dilakukan identifikasi Gα berdasarkan mRNA sehingga dapat diketahui adanya mutasi pada ruas utuh (full lenght) gen Gα dari ketiga tanaman kandidat mutan tersebut.
Tujuan penelitian
Tujuan penelitian ini adalah untuk mengidentifikasi kandidat mutan protein heterotrimerik Gα dari kedelai toleran Al kultivar Slamet berdasarkan hasil analisis mRNA.
BAHAN DAN METODE
Waktu dan tempat
Penelitian dilaksanakan mulai bulan Februari 2010 sampai dengan bulan Mei 2011 di Laboratorium Biotechnological Research Indonesia-Netherland (BIORIN), Pusat Penelitian Sumber Daya Hayati dan Bioteknologi (PPSHB), Institut Pertanian Bogor.
Bahan dan alat
Bahan tanaman yang digunakan dalam penelitian ini antara lain yaitu biji kedelai kultivar Slamet yang merupakan kandidat mutan heterotrimerik Gα (Limbong 2010). Biji tersebut adalah kandidat mutan generasi ke-5 (M5) dari tanaman generasi ke-4 (M4)
nomor 338/3S1.174/12/1/4,
344/3S1.196/14/1/8, dan generasi ke-6 (M6) dari tanaman M5 nomor 103/3S3.103/8/5/1/4, serta kedelai kultivar Slamet non mutan.
mikro, sentrifus Jouan BRi, vacuum dryer, perangkat elektroforesis, UV transluminator, spektrofotometer, dan perangkat PCR (Polymerase Chain Reaction).
Metode penelitian
Penanaman biji dan seleksi kandidat mutan gen Gα
Biji kedelai ditanam pada lahan seluas 4 x 7 meter, dengan media tanam tanah yang dicampur kompos. Masing-masing nomor ditanam pada sepuluh lubang (satu biji/ lubang) dengan jarak tanam 20 x 40 cm. Pemupukan dilakukan dua minggu setelah tanam dengan dosis Urea/TSP/KCL: 50/100/50 kg/ha. Hama diatasi dengan Acodan dosis 0.5ml/l. Penyemprotan Acodan dilakukan tiga kali selama masa tanam, yaitu 1 kali perbulan. Setelah tanaman memiliki cukup daun (± 3 minggu setelah tanam), dilakukan pengamatan kondisi stomata pada saat pukul 12.00 dengan metode nail polish swath. Selain itu, juga diamati suhu dan kelembaban pada saat pengamatan stomata (Lampiran 1). Pengamatan tinggi tanaman dilakukan pada saat muncul bunga. Tanaman yang tingginya lebih rendah dari pada Slamet non mutan dan jumlah stomata menutup >50% dipilih untuk diisolasi RNAnya.
Isolasi RNA total
Isolasi RNA menggunakan Kit Trizol Invitrogen. Langkah-langkah isolasi yaitu: 500-1000 mg daun segar digerus menggunakan nitrogen cair dan dicampur dengan 800µl Kit Trizol invitrogen. Campuran diinkubasi pada suhu ruang selama 5 menit dan ditambah dengan 200µl kloroform. Campuran dikocok selama 30 detik dan disentrifus dengan kecepatan 9000rpm, selama 15 menit pada suhu 6ºC. Supernatan diambil dan ditambah 500µl isopropanol dan diinkubasi kembali selama 10 menit pada suhu ruang, kemudian disentrifus dengan kecepatan 9000rpm selama 10 menit pada suhu 6ºC. Pelet diambil dan dicuci menggunakan alkohol-DEPC 70% dengan sentrifugasi 5700 rpm selama 5 menit pada suhu 6ºC. Pelet dikeringkan menggunakan vacuum dryer dan dilarutkan dalam 30 µl ddH2O-DEPC 1%, kemudian RNA
dikuantifikasi menggunakan spektrofotometer pada panjang gelombang 260 nm dan 280 nm. Keutuhan RNA di cek dengan elektroforesis agarosa 1% (b/v) menggunakan buffer MOPS 1x (Lampiran 2). Langkah-langkah elektro-foresis yaitu 10µg RNA dicampur dengan 12 µl premix (formamid, formaldehid, MOPS 20x dan ddH2O-DEPC 1%) dan dipanaskan
pada suhu 65ºC selama 10 menit, kemudian didinginkan di dalam es selama 5 menit. RNA dicampur dengan loading dye, dimasukan ke dalam gel dan dimigrasikan menggunakan buffer MOPS selama 30 menit dengan tegangan 100 volt. Selanjutnya gel direndam dalam larutan ethidium bromida 0.5 µg/ml selama 10 menit, dibilas dengan akuades, kemudian pita RNA dilihat menggunakan UV transluminator.
Sintesis cDNA
Sebelum sintesis cDNA, dilakukan pemurnian RNA dari DNA yaitu dengan mencampur 5µg RNA dengan 1.1 µl buffer DNase dan 1 unit enzim DNase. Campuran di inkubasi pada suhu 25ºC selama 8 menit. Selanjutnya dicampur dengan 1µl EDTA 25mM dan diinkubasi pada suhu 65ºC selama 10 menit. RNA murni didinginkan dalam es selama 5 menit. Sintesis cDNA dilakukan dengan metode Suharsono et al. (2002). Lima µl RNA murni dicampur dengan 4µl 5x buffer reaksi, 2 µl primer oligo dT, 1.6 µl dNTP 2.5 mM, 1 unit enzim reverse transcriptase III (Invitrogen), 1 µl dTT (0.1 M) dan ddH2
0-DEPC 1% steril sampai volume akhir 20 µl. Reaksi transkripsi balik (RT) dilakukan pada suhu 30ºC selama 10 menit, 42ºC selama 50 menit, 95ºC selama 5 menit sebanyak satu siklus. Untuk mengetahui keberhasilan sintesis cDNA dilakukan amplifikasi cDNA melalui PCR menggunakan primer aktin yang didesain dari kedelai (Accession V00450) dengan primer forward tepat dikodon awal dari ekson 1 (5’ATGGCAGATGCCGAGGγ’) dan primer reverse tepat pada daerah ekson 2 (5’CAGTTGTGCGACCACTTGCAγ’). PCR dilakukan dengan mencampur 1 µl hasil RT dengan 1 µl buffer taq 10x, 1 µl dNTP 2.5 mM, 10 pmol primer forward, 10 pmol primer reverse, 0.4 µl DMSO, 1 unit enzim taq polymerase dan ddH20 sampai volume akhir 10 µl. Kondisi PCR untuk aktin adalah: denaturasi pra-PCR 95ºC selama 5 menit, denaturasi 94ºC selama 30 detik, penempelan primer 54ºC selama 1 menit, pemanjangan DNA 72ºC selama 1 menit, siklus diulangi sebanyak 35 kali, pemanjangan pasca PCR 72ºC selama 5 menit dan proses pendinginan dilakukan pada suhu 15ºC selama 15 menit.
Identifikasi kandidat mutan dengan PCR
Nama primer Asal Sekuen basa nitrogen(5' - '3) Posisi nukleotida
L27418-SOYGALP1 (L1)
cDNA Gen Gα Kedelai
(Accession: L27418) GCTTCACACTTCACACTTAACACT (F)
111 nukleotida sebelum ATG
L27418-SOYGALP3 (L3)
cDNA Gen Gα Kedelai
(Accession: L27418) GCGCAGAATGACTTTGATTCTT (F)
309 nukleotida sesudah ATG
L27418-SOYGALP4 (L4)
cDNA Gen Gα Kedelai
(Accession: L27418) CTTAGGACCCACTCAAAAACGTTCC (R)
358 nukleotida sebelum TGA
L27418-SOYGALP5 (L5)
cDNA Gen Gα Kedelai
(Accession: L27418) GTTTACCCGCGTCTTACCAA (F)
361 nukleotida sesudah ATG
RGA-U3
DNA Genom Gen Gα Padi (Accession:
L35844)
CTGGCTTTGATGAGGCAGAAC (F) 775 nukleotida sesudah ATG
L35844) (Limbong 2010). Kombinasi primer tersebut yaitu L1-L2, L3-L2, L3-L4, U3-L4, L1-L4 dan L5-L4 (Tabel 1). Komposisi PCR sama dengan komposisi PCR aktin. Kondisi PCR juga sama dengan PCR aktin, tetapi dengan suhu penempelan primer 52ºC. Identifikasi mutan dilakukan dengan membandingkan pita gen pada gel agarosa 2% (b/v).
HASIL
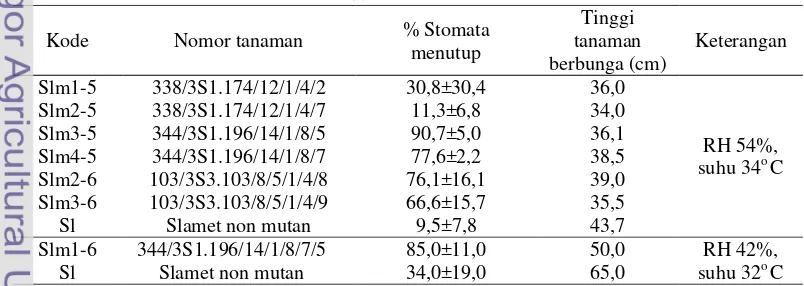
Dua puluh delapan nomor tanaman kandidat mutan M5 dan M6 berhasil ditanam, diamati keadaan stomata dan diukur tingginya (Lampiran 1). Enam tanaman diseleksi berdasarkan keadaan stomata menutup >50% dan lebih pendek dari pada Slamet non mutan (Tabel 2).
RNA total dari tanaman yang terseleksi berhasil diisolasi dan integritasnya dianalisis dengan elektroforesis. RNA total digunakan sebagai cetakan untuk mensintesis cDNA melalui RT-PCR. Identifikasi mutan Gα pada kandidat mutan menggunakan cDNA sebagai cetakan dan lima kombinasi primer yaitu L1-L2, L3-L1-L2, L3-L4, U3-L4, dan L5-L4 (Gambar 1).
Keberhasilan sintesis cDNA dapat diketahui dengan mengamplifikasi gen aktin melalui PCR. Hasil PCR aktin yaitu pita DNA berukuran 450pb sesuai dengan ukuran ekson 1 dan ekson 2 dari gen aktin (Gambar 2, 3 dan 4). Hasil tersebut juga menunjukkan bahwa cDNA yang diperoleh murni tidak terkontaminasi DNA genom.
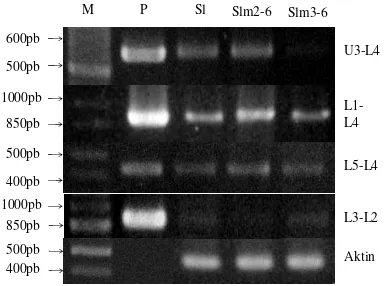
Kandidat mutan M5 dengan kode Slm1-5, Slm2-5, Slm3-5, dan Slm4-5 telah diidentifikasi secara molekular. Kandidat mutan Slm4-5 mengalami mutasi pada ruas L5-L4 berupa penurunan ekspresi gen, namun pada tiga nomor yang lain tidak mengalami mutasi gen Gα (Gambar 2). Kandidat mutan Slm4-5 ditanam kembali sehingga didapatkan kandidat mutan generasi ke-6 yaitu Slm1-6. Kandidat mutan Slm1-6 mengalami mutasi gen Gα pada ruas L1 sampai L3 (Gambar 3). Tanaman Slm1-6 memiliki stomata menutup >50% (Tabel 2) dan lebih pendek dari pada Slamet non mutan (Lampiran 3). Dua kandidat mutan lainnya mengalami mutasi gen Gα dengan penurunan ekspresi gen pada ruas U3-L4 yaitu tanaman Slm3-6 dan pada ruas L3-L2 yaitu Slm2-6. Hal tersebut dapat dilihat dari ketebalan pita dua kandidat tersebut yang lebih tipis dari tanaman non mutan (Gambar 4).
Kode Nomor tanaman % Stomata
menutup
Tinggi tanaman berbunga (cm)
Keterangan
Slm1-5 338/3S1.174/12/1/4/2 30,8±30,4 36,0
RH 54%, suhu 34o C
Slm2-5 338/3S1.174/12/1/4/7 11,3±6,8 34,0
Slm3-5 344/3S1.196/14/1/8/5 90,7±5,0 36,1
Slm4-5 344/3S1.196/14/1/8/7 77,6±2,2 38,5
Slm2-6 103/3S3.103/8/5/1/4/8 76,1±16,1 39,0 Slm3-6 103/3S3.103/8/5/1/4/9 66,6±15,7 35,5
Sl Slamet non mutan 9,5±7,8 43,7
Slm1-6 344/3S1.196/14/1/8/7/5 85,0±11,0 50,0 RH 42%,
suhu 32o C
[image:11.595.105.513.97.259.2]Sl Slamet non mutan 34,0±19,0 65,0
Tabel 1 Sekuen basa nitrogen pada primer yang digunakan
[image:11.595.109.511.611.754.2]Gambar 1 Panjang basa nitrogen antar primer cDNA heterotrimerik Gα
PEMBAHASAN
RNA dari tanaman kandidat mutan yang terseleksi berdasarkan keadaan stomata menutup dan lebih pendek dari pada Slamet non mutan telah berhasil diisolasi. RNA dapat dibagi menjadi dua kelompok yaitu yang berhubungan dengan ekspresi gen (mRNA, tRNA dan rRNA) dan yang tidak berhubungan dengan proses tersebut (RNA
primer). Dari ketiga jenis RNA yang berperan pada ekspresi gen, rRNA memiliki jumlah yang terbesar (83%) di dalam sel (Jusuf 2001). Oleh karena itu, rRNA digunakan sebagai indikator keberhasilan isolasi RNA total. PCR gen aktin dengan cDNA sebagai cetakan menghasilkan pita sebesar 450pb (Gambar 2, 3 dan 4). Kontaminasi cDNA oleh DNA genom ditandai dengan adanya pita yang ukurannya lebih besar (550pb). cDNA
P Sl Slm1-5 Slm2-5 Slm3-5 Slm4-5
L1-L4 L3-L4 L5-L4 Aktin 1000pb 850pb 700pb 600pb 500pb 400pb 500pb 400pb M 1650pb 1000pb L1-L2
P Sl Slm1-6
L3-L4 U3-L4 L1-L4 Aktin M 500pb 600pb 850pb 650pb 1000pb 850pb 500pb 400pb
Gambar 3 Ekspresi gen Gα dan aktin generasi M6 pada kandidat mutan Slm1-6. Marker (M), kontrol positif plasmid SL16 (P), dan kontrol non mutan kedelai kultivar Slamet (Sl).
Gambar 2 Ekspresi gen Gα dan aktin generasi M5 pada kandidat mutan Slm1-5, Slm2-5, Slm3-5, dan Slm4-5. Marker (M), kontrol positif plasmid SL16 (P), dan kontrol non mutan kedelai kultivar Slamet (Sl).
M P Sl Slm2-6 Slm3-6
U3-L4 L1-L4 L5-L4 L3-L2 Aktin 400pb 1000pb 850pb 500pb 850pb 1000pb 500pb 600pb 400pb 500pb
[image:12.595.109.510.83.448.2] [image:12.595.317.510.421.564.2]murni digunakan untuk mengidentifikasi keberadaan gen Gα dengan menggunakan PCR. PCR gen Gα dilakukan dengan beberapa kombinasi primer yang menghasilkan ukuran pita DNA yang berbeda (Gambar 1).
Keracunan Al pada tanaman kandidat mutan nomor 338/3S1.174/12/1 menyebabkan kerusakan jaringan akar sampai ke kortek dan pertumbuhan ujung akar menjadi bengkok (Giok 2010). Mutasi gen Gα dapat menyebabkan perubahan fenotipe tanaman mutan. Kandidat mutan memiliki persentase stomata menutup >50% dan lebih pendek dari pada Slamet non mutan. Penutupan dan pembukaan stomata merupakan akibat dari aktifitas hormon tumbuh seperti asam absisat (ABA) yang menyebabkan stomata menutup pada tanaman (Mishra et al. 2006). Empat kandidat mutan gen Gα generasi M5 yang berhasil diseleksi yaitu Slm1-5, Slm2-5, Slm3-5, dan Slm4-5 (Tabel 2). Tanaman Slm4-5 mengalami mutasi gen Gα pada ruas L5-L4 berupa penurunan ekspresi gen, sedangkan tiga nomor lainnya diduga mengalami mutasi gen Gα (Gambar 2). Hasil identifikasi menggunakan lima kombinasi primer pada dua kandidat mutan, juga menunjukkan adanya penurunan ekspresi gen
Gα pada dua kandidat mutan yaitu Slm3-6 pada ruas U3-L4 dan Slm2-6 pada ruas L3-L2 (Gambar 4). Penurunan ekspresi gen dapat dilihat dari ketebalan pita gen yang lebih tipis dari pita kultivar Slamet non mutan. Tingkat ekspresi gen ditentukan oleh promotor. Mutasi pada promotor dapat menyebabkan penurunan ekspresi gen. Berdasarkan hasil identifikasi Limbong (2010) kandidat mutan nomor 103/3S3.103/8/5/1/4 mengalami mutasi gen
Gα pada ruas U3 dan L4. Hal ini dapat terjadi karena kemungkinan adanya perubahan beberapa basa nitrogen pada situs penempelan primer U3. Mutasi terjadi pada kandidat mutan Slm1-6 yaitu pada ruas L1 sampai L3 (Gambar 4). Tanaman kandidat mutan Slm1-6 memiliki fenotipe lebih pendek dari pada Slamet non mutan dan stomata menutup >50%. Mutasi pada nomor ini diketahui setelah dilakukan pengecekan dengan tiga kombinasi primer yaitu L3-L4, L1-L4 dan U3-L4. Mutasi yang terjadi pada nomor ini adalah hilangnya sebagian atau seluruh sekuen L1 dan L3, sehingga primer L1 dan U3 yang berada di ruas L1 sampai L3 tidak dapat menempel, sedangkan primer yang berada di ruas setelah U3 dapat menempel, sehingga ruas L3-L4 dapat teramplifikasi. Hasil penelitian Limbong (2010) menunjukkan bahwa kandidat mutan M4 dengan nomor
344/3S1.196/14/1/8 tidak mengalami mutasi gen Gα. Hal tersebut menunjukkan bahwa kandidat nomor 344/3S1.196/14/1/8 (M4) memiliki alel gen Gα yang heterozigot terhadap mutan Gα.
SIMPULAN
Tiga nomor tanaman kandidat mutan generasi M6 yang diidentifikasi berdasarkan mRNA, mengalami mutasi gen Gα, yaitu nomor 103/3S3.103/8/5/1/4/9 pada ruas U3-L4, nomor 103/3S3.103/8/5/1/4/8 pada ruas L3-L2 dan nomor 344/3S1. 196/14/1/8/7/5 pada ruas L1 sampai L3.
SARAN
Untuk mendapatkan kandidat mutan perlu dilakukan seleksi dari seluruh biji yang berhasil dipanen. Hal ini dilakukan agar didapatkan kandidat mutan yang memiliki tingkat segregasi lebih kecil. Selain itu, perlu dilakukan sekuensing untuk membuktikan mutasi yang terjadi pada sekuen gen Gα.
DAFTAR PUSTAKA
Alberts et al. 2002. Molecular biology of the cell. New York: Garlan science.
Fujisawa et al. 1999. Supression of heterotrimeric G protein causes abnormal morphology including dwarfism in rice. Proc Natl Acad Sci 96:7575-7580
Fujisawa, Kato, Iwasaki . 2001. Structure and function of heterotrimeric G proteins in plants. Plant Cell Physiol 42(8):789-794 Giok G. 2010. Studi anatomi dan histokimia
pada akar kedelai (Glycine max (L) Merr) kandidat mutan Gα terhadap cekaman aluminium [Skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor
Hartini S. 2008. Induksi mutasi dengan irradiasi sinar gamma pada kedelai (Glycine max (L) Merrill) kultivar Slamet dan Lumut [Tesis]. Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor.
Jusuf M. 2001. Genetika I: Struktur dan Ekspresi Gen. Jakarta: Sagung Seto. Jusuf M, Suharsono. 2006. Perbaikan Genetik
Limbong LM. 2010. Identifikasi molekular kandidat mutan protein heterotimerik G subunit α (Gα) dari kedelai (Glycine max (L) Merrill) toleran aluminium kultivar Slamet [Skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Ma H. 1994. GTP binding proteins in plants: new members of an old family. Plant Mol Biol 26:1611-1636.
Ma H. 2001. Plant G proteins: the different faces of GPA1. Curr Biol 11:R869-R871.
Mishra G, Zhang W, Deng F, Zhao J, Wang Z. 2006. A bifurcating pathway direct abscisic acid effects on stomatal closure and opening in Arabidopsis. Science 312: 264-266.
Perfush-Barbeoch L, Jones AM, Assman SM. 2004. Plant heterotrimeric G protein function: Insight from Arabidopsis and rice mutants. Plant Biol 7:719-731. Srimulyati T. 2007. Keterlibatan protein
heterotrimerik Gα terhadap cekaman aluminium pada kedelai (Glycine max (L.) Merril) melalui studi histokimia [Skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Suharsono U, Fujisawa Y, Kawasaki T, Iwasaki Y, Satoh H, Shimamoto K. 2002. The heterotrimeric G protein subunit acts upstream of the small GTPase Rac in disease resistance of rice. Proc Natl Acad Sci USA 99: 13307-13312.
Lampiran 1 Hasil seleksi tanaman kandidat mutan berdasarkan persentase stomata menutup >50% dan tanaman pendek
*suhu : 34oc RH : 54% Nomor tanaman Stomata
menutup (%)
Tinggi tanaman (cm)
Umur berbunga
(HST)
Jumlah biji
338/3S1.174/12/1/4/1 4.3±4.6 38.5 37.0 0
338/3S1.174/12/1/4/2 30.8±30.4 36.0 37.0 0
338/3S1.174/12/1/4/3 10.3±5.9 46.0 35.0 0
338/3S1.174/12/1/4/4 13.4±8.00 48.3 35.0 0
338/3S1.174/12/1/4/5 21.6±5.7 40.0 37.0 0
338/3S1.174/12/1/4/6 57.4±11.7 41.0 37.0 0
338/3S1.174/12/1/4/7 11.3±6.8 34.0 37.0 0
338/3S1.174/12/1/4/8 0.0±0.0 48.6 35.0 0
344/3S1.196/14/1/8/2 72.3±26.6 29.0 40.0 133
344/3S1.196/14/1/8/3 100.0±0.0 40.6 40.0 53
344/3S1.196/14/1/8/4 91.4±3.4 41.0 40.0 166
344/3S1.196/14/1/8/5 90.7±5.0 36.1 40.0 100
344/3S1.196/14/1/8/6 85.0±3.5 35.6 40.0 120
344/3S1.196/14/1/8/7 77.6±2.2 38.5 40.0 169
344/3S1.196/14/1/8/8 88.3±1.6 41.1 40.0 128
103/3S3.103/8/5/1/4/1 19.6±11.4 31.0 40.0 0
103/3S3.103/8/5/1/4/2 27.4±11.5 30.0 41.0 33
103/3S3.103/8/5/1/4/4 95.7±4.0 34.0 40.0 115
103/3S3.103/8/5/1/4/6 70.4±26.8 37.5 40.0 36
103/3S3.103/8/5/1/4/7 54.1±14.4 38.0 40.0 70
103/3S3.103/8/5/1/4/8 76.1±16.1 39.0 43.0 12
103/3S3.103/8/5/1/4/9 66.6±15.7 35.5 41.0 68
103/3S3.103/8/5/1/4/10 70.2±26.0 31.0 40.0 21
103/3S3.103/8/5/1/4/11 93.6±5.8 33.0 41.0 79
103/3S3.103/8/5/1/4/12 87.9±11.1 34.5 43.0 0
Lampiran 2 Komposisi buffer MOPS 20x dan gel agarosa 1%
A. Buffer MOPS 20x
Morpholino propano sulfonic acid (MOPS) : 42 gram
Natrium asetat pekat : 4.1 gram
EDTA (Tritriplex) : 3.7 gram
Aquades : 1 liter
B. Komposisi gel agarosa 1% (b/v)
Agarosa : 0. 25 gr
20x MOPS : 1.25 ml
Lampiran 3 Tanaman kandidat mutan Slm1-6 (kanan) dan Slamet non mutan (kiri)