PENGEMBANGAN ALTERNATIF TEKNOLOGI
BIOPROSES PEMBUATAN BIOETANOL DARI UBI
KAYU MENGGUNAKAN
Trichoderma viride,
Aspergillus niger
DAN
Saccharomyces cerevisiae
I WAYAN ARNATA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER
INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul :
PENGEMBANGAN ALTERNATIF TEKNOLOGI BIOPROSES
PEMBUATAN BIOETANOL DARI UBI KAYU MENGGUNAKAN
Trichoderma viride, Aspergillus niger DAN Saccharomyces cerevisiae adalah benar merupakan karya sendiri dibawah arahan Komisi Pembimbing dan belum pernah dipublikasikan. Semua sumber data dan informasi telah dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Bogor, September 2009
ABSTRACT
I WAYAN ARNATA. Development of Alternative Bioprocess Technology to Bioethanol Production from Cassava by Trichoderma viride, Aspergillus niger
and Saccharomyces cerevisiae. Supervised by Dwi Setyaningsih and Nur Richana.
Cassava (Manihot utilisima) is one of viable feedstock for bioethanol production that contains starch and fiber. Both components can be hydrolyzed through the acid and enzymatic processes. T. viride is capable of producing cellulase useful for cellulose hydrolysis, while A. niger is able to produce amyloglucosidase for starch hydrolysis. Glucose produced in this step is used by
S. cerevisiaeas fermentation substrate to produce ethanol.
This study aimed at increasing the yield of ethanol concentration from cassava by making bioprocess alternative through mixed cultureT. viride,A. niger
andS. cerevisiaeat both acid and enzyme hydrolisate.
The experiments were performed in three stages. The first stage was the cultivation ofT. virideandA. nigerto determine the stationary phase of maximum spore yield. The second stage was the determination of the produce time needed that gave maximum enzyme activity. Enzyme assay was conducted by measuring the cellulase (CMC-ase, FP-ase) and amyloglucosidase (AMG) activities. The third stage was the hydrolysis and fermentation processes with six alternatives of production process and one control. The first treatment (P1) was using acid hydrolisate and the addition of coculture gradually. The second treatment (P2) was using acid hydrolisate and the addition of mixed culture simultaneously. The third treatment (P3) was using enzyme hydrolisate with SFS coculture addition. The forth treatment (P4) was using enzyme hydrolisate with the addition of crude cellulase enzyme in the saccharification process. The fifth treatment (P5) was using enzyme hydrolisate and the addition of mixed culture gradually. The sixth treatment (P6) was using enzyme hydrolisate with the addition of commercial cellulase enzyme in the saccharification process. The control process (P7) was using enzyme hydrolisate and the addition of monoculture S. cerevisiae. Observations during the fermentation process included changes in the total sugar concentration (Dubois et al., 1956), pH, concentration of the crude fibre residue (AOAC, 1984) and the ethanol concentration (Gas Chromatography).
From the experiments, the stationary phase of maximum spore yield occurred after 7 days of cultivation of T. virideandA. nigerwith maximum spore number of 1.51 x 10 9 and 1.26 x 10 9, respectively. Maximum cellulose and AMG activities obtained after 7 days of fermentation with the CMC-ase activities of 5.05 ± 0.42 U/ml, FP-ase of 4.77 ± 0.72 U/ml and AMG of 62.77 ± 4.49 U/ml.
Mixed culture T. viride, A. niger and S. cerevisiae in the fermentation process of acid hydrolisate either done gradually or with simultaneous saccarification and fermentation was not able to increase ethanol concentration when compared to control. However, ethanol concentration increased significantly with AMG addition of crude and commercial cellulase in the saccharification process. The ethanol concentration increased from 5.36 ± 0.63 % (b/v) in the control to 9.29 ± 1.76 % (b/v) (adding crude cellulase) and 8.92 ± 0.73 % (b/v) (commercial cellulase) or an increase of 73.45 % and 64.42 %, respectively, compared to that of the monoculture. The experiments resulted that the highest ethanol concentration is produced from the fourth treatment (P4) that is 9.29 ± 1.76 % (b/v) with yield of 34.77 % (v/b).
RINGKASAN
I WAYAN ARNATA. Pengembangan Alternatif Teknologi Bioproses Pembuatan Bioetanol Dari Ubi Kayu Menggunakan Trichoderma viride, Aspergillus niger
dan Saccharomyces cerevisiae. Dibimbing oleh Dwi Setyaningsih dan Nur Richana.
Ubi kayu (Manihot utilisima) merupakan salah satu bahan baku pembuatan bioetanol yang mengandung fraksi pati dan serat. Kedua fraksi ini dapat dihidrolisis secara asam maupun enzim. Jenis kapang T. viride mampu menghasilkan selulase yang berguna untuk menghidrolisis serat (selulosa) dan A. niger mampu menghasilkan amiloglukosidase untuk menghidrolisis pati. Hasil hidrolisis berupa glukosa dapat dipergunakan oleh S. cerevisiae sebagai substrat fermentasi untuk menghasilkan etanol.
Penelitian ini bertujuan mendapatkan alternatif teknologi bioproses pembuatan bioetanol terbaik dari ubi kayu menggunakan kultur campuran T. viride, A. nigerdanS. cerevisiaebaik pada hidrolisat asam maupun enzim.
Penelitian dilakukan dalam tiga tahap. Tahap pertama dilakukan kultivasi pertumbuhan T. viride dan A. niger untuk menentukan fase stasioner yang menghasilkan jumlah spora maksimal. Tahap kedua dilakukan untuk menentukan lama proses produksi yang menghasilkan aktivitas enzim maksimal. Pengujian enzim dilakukan dengan mengukur aktivitas selulase (CMC-ase, FP-ase) dan amiloglukosidase (AMG). Tahap ketiga dilakukan proses hidrolisis dan fermentasi dengan enam alternatif proses produksi dan satu proses kontrol. Pengamatan selama proses fermentasi meliputi perubahan konsentrasi total gula (Dubois et al. 1956), pH, konsentrasi serat kasar sisa (AOAC,1984) dan konsentrasi etanol.
Dari hasil penelitian, fase stasioner pertumbuhan spora T. viride dan A. niger diperoleh setelah kultivasi selama 7 hari dengan jumlah spora maksimum masing-masing 1,51 x 10 9/ml dan 1,26 x 10 9/ml. Aktivitas selulase dan AMG maksimum diperoleh setelah fermentasi selama 7 hari dengan aktivitas CMC-ase 5,05 ± 0,42 U/ml, FP ase 4,77 ± 0,72 U/ml dan AMG 62,77 ± 4,49 U/ml.
Penggunaan kultur campuran T. viride, A. niger dan S. cerevisiae pada proses proses fermentasi substrat hidrolisat enzim secara SFS selama 4 hari dapat meningkatkan konsentrasi etanol etanol dari 5,36 ± 0,63 % (b/v) menjadi 7,41 ± 1,79 % (b/v) atau meningkat 38,29 % dibandingkan dengan menggunakan kultur tunggal S. cerevisiae, sedangkan penggunaan kultur campuran yang ditambahkan secara bertahap pada proses fermentasi hanya mampu meningkatkan konsentrasi etanol dari 5,36 ± 0,63 % (b/v) menjadi 6,38 ± 0,83 % (b/v) atau meningkat 19,06 % terhadap penggunaan kultur tunggalS. cerevisiae.
© Hak Cipta milik IPB, tahun 2009
Hak Cipta dilindungi Undang-Undang
1.
Dilarang mengutip sebagian atau seluruhnya karya tulis ini
tanpa mencantumkan atau menyebutkan sumber.
a. Pengutipan
hanya
untuk
kepentingan
pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan,
penulisan kritik atau tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2.
Dilarang mengumumkan dan memperbanyak sebagian atau
PENGEMBANGAN ALTERNATIF TEKNOLOGI BIOPROSES
PEMBUATAN BIOETANOL DARI UBI KAYU
MENGGUNAKAN
Trichoderma viride, Aspergillus niger
DAN
Saccharomyces cerevisiae
I WAYAN ARNATA
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Teknologi Industri Pertanian
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
LEMBAR PENGESAHAN
Judul Penelitian : Pengembangan Alternatif Teknologi Bioproses Pembuatan Bioetanol Dari Ubi Kayu Menggunakan
Trichoderma viride, Aspergillus niger dan
Saccharomyces cerevisiae
Nama Mahasiswa : I Wayan Arnata
N RP : F351070111
Disetujui
Komisi Pembimbing
Dr. Ir. Dwi Setyaningsih, M.Si. Dr. Ir. Nur Richana, M.Si.
Ketua Anggota
Diketahui
Ketua Program Studi a.n. Dekan Sekolah Pascasarjana Teknologi Industri Pertanian Sekretaris Program Magister
Prof. Dr. Ir. Irawadi Jamaran Dr. Ir. Naresworo Nugroho, M.Si.
PRAKATA
Puji syukur penulis pajatkan kehadirat Tuhan Yang Maha Esa, karena atas berkat rahmat-nya penulis dapat menyelesaikan tugas akhir penelititan yang berjudul : “Pengembangan Alternatif Teknologi Bioproses Pembuatan Bioetanol Dari Ubi Kayu MenggunakanTrichoderma viride, Aspergillus niger
danSaccharomyces cerevisiae”tepat pada waktunya.
Pada kesempatan ini penulis mengucapkan terima kasih yang sebesar-besarnya kepada Dr. Ir. Dwi Setyaningsih, M.Si. selaku Ketua Komisi Pembimbing dan Dr. Ir. Nur Richana, M.Si. selaku Anggota Komisi Pembimbing atas bimbingan dan arahannya.
Ayahanda I Nyoman Kader, Ibunda Ni Made Kedri, Istri Nelly Verawati, SP dan Kedua putriku Wayan Arlyana Anantha dan Kadek Arlyatha Anantha tercinta. Keluarga Bapak Petrus Sarijono dan Ibu Lucia Siti Rahayu di Caruban Madiun. Teman-teman di Perhimpunan Mahasiswa Pasca Sarjana Bali “Puhnawacana” DR. drh. Nyoman Suarsana, drh. I Gst Ngurah Sudisma, MSi, drh Made Rai Yasa, MP, Ni Luh Yulianti, STp. M.Si. Diah STp. M.Si, , drh.Yudi A. Ibu Suci, SPt dan Gayeng, ST. Teman-teman dan rekan kerja di FTP Udayana terima kasih atas motivasinya. Seluruh staf pengajar Teknologi Industri Pertanian dan Staf Laboran, staf administrasi di Lingkungan Fateta IPB dan Teman-teman Teknologi Industri Pertanian ’07 dan tidak lupa juga kepada Yuyun, Jihan, Asep, Lily, Mbak Fitri dan Yusup terima kasih atas bantuannya.
Penulis menyadari bahwa karya tulis ilmiah ini masih jauh dari sempurna, oleh karena itu saran dan kritik yang bersifat membangun sangat diharapkan agar dapat memberikan informasi dalam pengembangan karya tulis ini lebih lanjut.
RIWAYAT HIDUP
Penulis dilahirkan di Desa Ungasan, Kuta Selatan Badung Bali pada tanggal 20 Juni 1978 sebagai anak tunggal dari I Nyoman Kader dan Ni Made Kedri. Penulis memasuki jenjang Sekolah Dasar tahun 1984 dan lulus tahun 1990 di SDN 4 Jimbaran, melanjutkan ke jenjang SMP tahun 1990 dan lulus tahun 1993 di SMPN 2 Kuta, jenjang SMA tahun 1993 sampai 1996 di SMAN 5 Denpasar. Penulis melanjutkan kuliah strata 1 tahun 1996 di Program Studi Teknologi Pertanian Universitas Udayana sampai tahun 2001. Dari tahun 2001 sampai 2005 penulis sempat bekerja sebagai pegawai swasta sebelum menjadi staf dosen di Fakultas Teknologi Pertanian Universitas Udayana. Pada tahun 2007 melalui dana beasiswa pendidikan BPPS penulis melanjutkan kuliah strata 2 di Sekolah Pascasarjana, Teknologi Industri Pertanian IPB.
Bogor, September 2009
DAFTAR ISI
Halaman
DAFTAR TABEL... xiii
DAFTAR GAMBAR ... xiv
DAFTAR LAMPIRAN... xv
I. PENDAHULUAN 1.1 Latar belakang ... 1
1.2 Rumusan masalah ... 3
1.3 Tujuan... 3
1.4 Hipotesis ... 3
1.5 Ruang lingkup ... 4
1.6 Kerangka pemikiran ... 5
II. TINJAUAN PUSTAKA 2.1 Ubi Kayu ... 6
2.2 Polisakarida Dalam Ubi Kayu... 8
2.3 Bioetanol ... 9
2.4 Hidrolisis Asam... 13
2.5 Hidrolisis Enzim ... 15
2.6 Sakarifikasi Fermentasi Simultan (SFS) ... 18
III. METODOLOGI PENELITIAN 3.1 Waktu dan Tempat Penelitian ... 20
3.2 Bahan dan Alat ... 20
3.2.1 Bahan... 20
3.2.2 Alat... 20
3.3 Tahapan Penelitian ... 20
3.3.1 Karakteristik ubi kayu ... 21
3.3.3 Persiapan kulturA. Niger... 22
3.3.4 Persiapan kulturS. Cerevisiae... 22
3.3.5 Produksi enzim glukoamilaseA. Niger... 22
3.3.6 Produksi enzim selulaseT. Viride... 22
3.3.7 Pembuatan etanol ... 23
3.3 Teknik Analisis Data... 24
3.4.1 Produksi enzim selulase ... 24
3.4.2 Produksi enzim amiloglukosidase... 25
3.4.3 Proses hidrolisis ... 26
3.4.2 Proses fermentasi ... 26
IV HASIL DAN PEMBAHASAN 4.1 Komposisi kimia ubi kayu ... 28
4.2 Kultivai dan Produksi Enzim ... 30
4.2.1 Kultivasi Trichoderma viride... 31
4.2.2 Produksi enzim selulase ... 32
4.2.3 KultivasiAspergillus niger dan produksi amiloglukosidase... 33
4.3 Proses hidrolisis dan karakteristik hidrolisat ... 35
4.4 Proses fermentasi ... 39
V KESIMPULAN DAN SARAN 5.1 Kesimpulan ... 55
5.2 Saran... 55
DAFTAR TABEL
Halaman
1. Perkembangan produksi ubi kayu Indonesia... 7
2. Sifat fisiko kimia ubi kayu dan tepung ubi kayu ... 8
3. Perbandingan keuntungan dan kelemahan antara concentrated-acid hydrolisisdengandilute-acid hydrolisis... 14
4. Kondisi hidrolisis dan fermentasi tahapan proses ... 24
5. Hasil analisa komposisi kimia ubi kayu... 28
DAFTAR GAMBAR
Halaman
1. Kerangka pemikiran penelitian ... 5
2. Struktur selulosa... 9
3. Mekanisme proses glikolisis ... 19
4. Tahapan penelitian ... 21
5. Inokulum T. viride dan A. niger ... 30
6. Kurva pertumbuhanT. viride,aktivitas CMC-ase dan FP-ase... 31
7. Kurva pertumbuhanA. niger dan aktivitas glukoamilase ... 34
8. Pengaruh hidrolisis terhadap total gula dan gula pereduksi... 37
9. Perubahan pH dan total gula selama fermentasi P1 ... 41
10. Perubahan pH dan total gula selama fermentasi P2 ... 41
11. Perubahan pH dan total gula selama fermentasi P3 ... 42
12. Perubahan pH dan total gula selama fermentasi P4 ... 43
13. Perubahan pH dan total gula selama fermentasi P5 ... 44
14. Perubahan pH dan total gula selama fermentasi P6 ... 45
15. Perubahan pH dan total gula selama fermentasi P7 ... 46
16. Pengaruh jenis perlakuan fermentasi terhadap konsentrasi etanol, rendemen dan efisiensi substrat ... 48
DAFTAR LAMPIRAN
Halaman
1. Prosedur Analisa ... 63
2. Alternatif tahapan proses pembuatan bioetanol ... 70
3. Kurva standar DNS dan total gula ... 75
4. Analisis deskriptif pertumbuhan jumlah spora T. viride dan A. niger... 76
5. Analisa aktivitas enzim selulase dan glukoamilase ... 77
6. Analisis keragaman hasil hidrolisis tepung ubi kayu... 81
7. Perubahan total gula selama fermentasi... 83
8. Perubahan pH selama fermentasi ... 86
9. Hasil produksi etanol ... 89
10. Hasil analisis keragaman terhadap produksi etanol ... 91
I. PENDAHULUAN
1.1 Latar Belakang
Bioetanol merupakan hasil proses fermentasi glukosa dari bahan yang mengandung komponen pati atau selulosa. Kedua komponen ini merupakan homopolimer dari glukosa. Bietanol dapat dipergunakan sebagai salah satu energi alternatif pensubsitusi bensin yang ramah lingkungan jika dibandingkan dengan bahan bakar fosil. Berdasarkan data Departemen Energi dan Sumberdaya Mineral (2007), proyeksi konsumsi etanol untuk mensubsitusi 5 % premium (E5) di Indonesia dari tahun 2007 – 2010 ditargetkan sekitar 5 % dan tahun 2011 - 2015 ditargetkan sekitar 10 % atau sekitar 2,78 juta kL dari total konsumsi.
Berbagai jenis sumber bahan baku bioetanol terdapat di Indonesia, seperti ubi kayu, sagu, ubi jalar dan tetes tebu. Ubi kayu sebagai bahan baku bioetanol mempunyai kelebihan yaitu dapat tumbuh pada lahan yang kurang subur, mempunyai daya tahan tinggi terhadap penyakit dan dapat diatur masa panennya. Perkembangan produksi ubi kayu di Indonesia mengalami peningkatan sekitar 23 % (16 ton menjadi 20 ton) dari tahun 2000 - 2008 (Deptan 2008). Ubi kayu mempunyai kadar karbohidrat sekitar 32 – 35 % yang sebagian besar adalah pati yaitu sekitar 83,8%. Komponen-komponen lain ubi kayu terdiri dari air, protein, lemak, serat dan abu. Serat pada ubi kayu tersusun dari selulosa, hemiselulosa, lignin dan protein. Konversi bahan baku pati ubi kayu menjadi bioetanol menghasilkan rendemen sekitar 16, 67 %, ini berarti setiap pengolahan 1 ton ubi kayu akan menghasilkan 166,7 liter bioetanol (Nurdyastuti 2005). Rendemen yang dihasilkan pada proses pembuatan bioetanol dari ubi kayu sangat tergantung pada kemampuan proses hidrolisis komponen-komponen ubi kayu terutama pati menjadi glukosa, selanjutnya tinggi rendahnya kandungan glukosa hasil hidrolisis akan mempengaruhi proses fermentasi dalam pembentukan etanol.
Hidrolisis pati dan serat dapat dilakukan secara kimiawi atau enzimatik. Hidrolisis kimiawi biasanya menggunakan asam seperti H2SO4 dan HCl. Dalam menghasilkan glukosa, hidrolisis dengan asam akan memotong ikatan pada pati dan serat secara acak. Hidrolisis enzimatik dilakukan dengan menggunakan enzimα-amilase dan amiloglukosidase. Enzim α-amilase dapat menghidrolisis molekul pati baik pada amilosa maupun pada amilopektinnya. Hasil akhir hidrolisis amilosa adalah glukosa dan maltosa, sedangkan hasil akhir hidrolisis amilopektin menghasilkan campuran limit dekstrin, maltosa, isomaltosa dan disertai sedikit glukosa. Enzim amiloglukosidase mampu menghidrolisis pati menjadi molekul-molekul glukosa. Enzim glukoamilase dapat diperoleh dari strainAspergillus nigeratauRhizopus(Tjokroadikoesoemo 1986).
Selulosa mempunyai potensi sebagai bahan baku bioetanol karena komponen selulosa tersusun dari homopolimer D-glukosa. Enzim selulase merupakan kompleks enzim yang dapat mengkatalisis penguraian selulosa menjadi glukosa. Beberapa spesies seperti Trichoderma viride dan Fusarium oxysporum dilaporkan mempunyai kemampuan untuk menghasilkan enzim selulase. Hidrolisis komponen selulosa dan hemiselulosa dalam ubi kayu dapat dilakukan dengan hidrolisis asam maupun enzim. Hidrolisis selulosa akan menghasilkan D-glukosa, sedangkan hemiselulosa akan menghasilkan D-xilosa dan monosakarida lainnya.
Proses pengolahan ubi kayu menjadi bioetanol secara umum dimulai dari proses pencucian, pemarutan, hidrolisis atau dilikuifikasi dengan menggunakan enzimα-milase, kemudian sakarifikasi dengan enzim amiloglukosidase dan fermentasi menggunakanS. cerevisiae. Penggunaan enzim α-amilase dan amiloglukosidase pada proses produksi hanya mampu menghidrolisis fraksi pati menjadi glukosa dengan memutus ikatanα-1,4 dan α-1,6 glikosidik pada fraksi amilosa dan amilopektin, sedangkan fraksi serat atau selulosa yang mempunyai ikatanβ-1,4 glikosidik tidak mampu dihidrolisis oleh enzim-enzim amilolitik (Thomas dan William 1997).
sehingga dapat meningkatkan konsentrasi glukosa. Dengan meningkatnya konsentrasi glukosa diharapkan dapat meningkatkan produksi etanol.
1.2 Rumusan Masalah
Pembuatan bioetanol dari ubi kayu masih dapat ditingkatkan konsentrasi atau rendemen etanolnya dengan aplikasi enzim atau kultur campuran. Adanya kendala harga enzim atau kultur mikroba yang relatif mahal atau sukar didapatkan, maka dalam penelitian ini akan dicoba dengan aplikasi hidrolisis menggunakan asam. Berdasarkan latar belakang di atas maka rumusan permasalahan dalam penelitian ini adalah mengembangkan alternatif teknologi bioproses pembuatan bioetanol dari ubi kayu dengan menggunakan kultur campuran T. viride, A. niger dan S. cerevisiaebaik pada substrat hidrolisat asam maupun enzim.
1.3 Tujuan
Tujuan penelitian ini adalah mendapatkan alternatif teknologi bioproses pembuatan bioetanol terbaik dari ubi kayu menggunakan filtrat enzim selulase kasar dan komersial serta kultur campuran T. viride, A. niger dan S. cerevisiae baik pada substrat hidrolisat asam maupun enzim.
1.4 Hipotesis
T. viride merupakan jenis kapang yang mampu menghasilkan selulase yang berguna untuk menghidrolisis selulosa menjadi glukosa, sedangkan A. niger mampu menghasilkan amiloglukosidase yang berfungsi untuk menghidrolisis fraksi pati menjadi glukosa.
1.5 Ruang lingkup
Untuk mendapatkan hasil yang jelas dan terarah, maka ruang lingkup penelitian ini adalah sebagai berikut:
1. Melakukan kultivasi pertumbuhanT. viride dan A. nigeruntuk menghasilkan spora maksimal.
2. Produksi enzim kasar selulase dengan penginduksi onggok oleh T. viride dan produksi amiloglukosidase dengan penginduksi pati ubi kayu olehA. niger.
3. Proses hidrolisis dilakukan secara kimiawi menggunakan H2SO4 tanpa pemisahan fraksi serat dengan pati dan dibandingkan dengan hasil hidrolisis yang dilakukan secara enzimatis.
4. Proses sakarifikasi dan fermentasi dilakukan dengan sistem batch secara simultan dan bertahap menggunakan kultur campuran T. viride, A. niger dan S. cerevisiae
5
1.6 Kerangka pemikiran
II. TINJAUAN PUSTAKA
2.1 Ubi Kayu
Ubi kayu (Manihot utilissima pohl) berasal dari Benua Amerika dan Bangsa Portugis membawanya ke Afrika dan digunakan sebagai bahan makanan. Ubi kayu saat ini penyebarannya hampir keseluruh dunia dan berkembang di negara-negara yang terkenal wilayah pertaniannya. Ubi kayu ditanam secara komersial di wilayah Indonesia sekitar tahun 1810, setelah sebelumnya diperkenalkan orang Portugis pada abad ke-16 ke Nusantara dari Brasil. Klasifikasi tanaman ubi kayu adalah :
Kingdom : Plantae atau tumbuh-tumbuhan Divisi : Spermatophyta atau tumbuhan berbiji Sub Divisi : Angiospermae atau berbiji tertutup Kelas : Dicotyledoneae atau biji berkeping dua Ordo : Euphorbiales
Famili : Euphorbiaceae
Genus : Manihot
Spesies :Manihot utilissimaPohl ;Manihot esculenta Crantz sin.
Ubi kayu termasuk tumbuhan berbatang pohon lunak atau mudah patah. Ubi kayu berbatang bulat dan bergerigi yang terjadi dari bekas pangkal tangkai daun, bagian tengahnya bergabus dan termasuk tumbuhan yang tinggi. Ubi kayu bisa mencapai ketinggian 1-4 meter. Ubi kayu mempunyai panjang fisik rata-rata bergaris tengah 2-3 cm dan panjang 50-80 cm, tergantung dari jenis ubi kayu yang ditanam. Daging umbinya berwarna putih atau kekuning-kuningan. Ubi kayu biasanya diperdagangkan dalam bentuk masih berkulit. Umbinya mempunyai kulit yang terdiri dari 2 lapis yaitu kulit luar dan kulit dalam. Daging umbi berwarna putih atau kuning. Di bagian tengah daging umbi terdapat suatu jaringan yang tersusun dari serat. Antara kulit dalam dan daging umbi terdapat lapisan kambium.
muda dari jenis yang beracun berguna untuk berbagai macam sayur. Daun yang kering untuk makanan ternak. Batangnya dapat digunakan untuk kayu bakar dan kadang-kadang untuk pagar hidup. Salah satu varietas tanaman ini mempunyai daun yang indah warnanya yang dimanfaatkan sebagai tanaman hias (Syarief 1988).
Ubi kayu mengandung racun yang disebut asam sianida (HCN). Berdasarkan kandungan asam sianidanya, ubi kayu dapat digolongkan menjadi empat yaitu (a) golongan tidak beracun, mengandung HCN 50 mg per kg umbi segar yang telah diparut, (b) beracun sedikit mengandung HCN antara 50 dan 80 mg per kg, (c) beracun, mengandung HCN antara 80 dan 100 mg per kg dan (d) sangat beracun, mengandung HCN lebih besar dari 100 mg per kg. Ubi kayu yang tidak beracun dikenal sebagai ubi kayu manis sedangkan ubi kayu yang beracun disebut ubi kayu pahit.
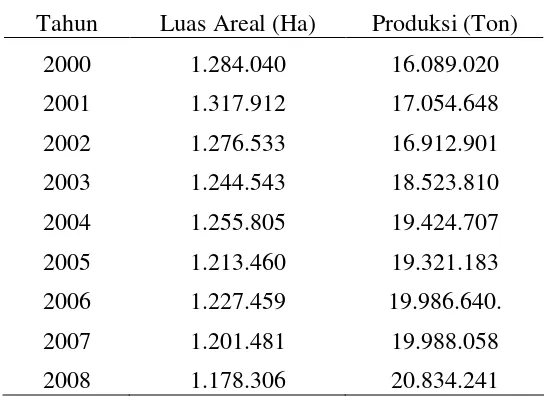
Ubi kayu memiliki kelebihan sebagai bahan baku bioetanol yaitu dapat tumbuh di tanah yang kurang subur, memiliki daya tahan yang tinggi terhadap penyakit, dan dapat diatur waktu panennya. Potensi pengembangan produksi ubi kayu di Indonesia disajikan pada Tabel 1.
Tabel 1. Perkembangan produksi ubi kayu Indonesia Tahun Luas Areal (Ha) Produksi (Ton)
2000 1.284.040 16.089.020
2001 1.317.912 17.054.648
2002 1.276.533 16.912.901
2003 1.244.543 18.523.810
2004 1.255.805 19.424.707
2005 1.213.460 19.321.183
2006 1.227.459 19.986.640.
2007 1.201.481 19.988.058
2008 1.178.306 20.834.241
Sumber : Departemen Pertanian (2008)
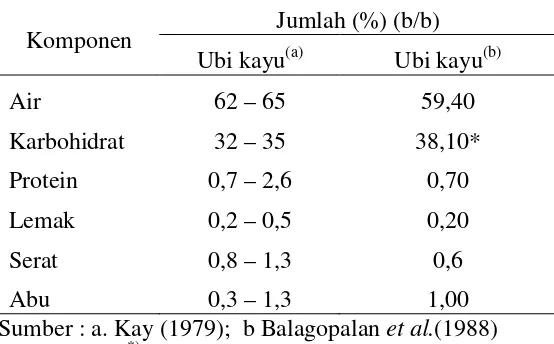
Tabel 2. Sifat fisiko kimia ubi kayu dan tepung ubi kayu Jumlah (%) (b/b)
Komponen
Ubi kayu(a) Ubi kayu(b)
Air 62 – 65 59,40
Karbohidrat 32 – 35 38,10*
Protein 0,7 – 2,6 0,70
Lemak 0,2 – 0,5 0,20
Serat 0,8 – 1,3 0,6
Abu 0,3 – 1,3 1,00
Sumber : a. Kay (1979); b Balagopalanet al.(1988) Keterangan :*)Dihitung berdasarkanby difference
2.2 Polisakarida Dalam Ubi Kayu
Polisakarida yang menyusun ubi kayu terdiri dari pati, selulosa dan hemiselulosa. Pati pada tumbuhan dipergunakan sebagai cadangan makanan yang dapat diuraikan menjadi glukosa dan dikonversikan menjadi energi. Pada saat yang tepat, tubuh tanaman akan mensintesa α-amilase, β-amilase dan R-enzim yang secara bersama-sama dipergunakan untuk memutuskan ikatan-ikatan rantai pati menjadi molekul-molekul glukosa bebas (Tjokroadikoesoemo 1986).
Pati merupakan homopolimer glukosa dengan ikatanα-glikosidik. Pati terdiri dari dua fraksi yang dapat dipisahkan dengan air panas yaitu fraksi amilosa dan amilopektin. Fraksi amilosa sifatnya larut dalam air panas dan fraksi amilopektin bersifat tidak larut. Amilosa mempunyai struktur lurus dengan ikatan α-(1,4)-D-glukosa, sedang amilopektin mempunyai cabang dengan ikatan α-(1,6)-D-glukosa sebanyak 4 – 5 % dari berat total (Winarno 1992). Hidrolisis amilosa menghasilkan maltosa, glukosa dan oligosakarida lainnya. Pada amilopektin sebagian dari molekul-molekul glukosa di dalam rantai percabangannya saling berikatan melalui gugus α-1,6. Ikatan α-1,6 sangat sukar diputuskan, apalagi jika dihidrolisis menggunakan katalisator asam.
menyebabkan selulosa bersifat kristalin, tidak mudah larut dan tidak mudah didegradasi secara kimia maupun mekanis. Rumus bangun selulosa disajikan pada Gambar 2.
Gambar 2 Struktur selulosa
Selulosa bersama-sama dengan hemiselulosa, pektin dan protein berfungsi untuk membentuk struktur jaringan dinding sel tanaman (Holtzapple 1993). Selulosa dapat berasosiasi dengan lignin sehingga sering disebut lignoselulosa. Selulosa, hemiselulosa dan lignin merupakan komponen utama penyusun tanaman yang dihasilkan melalui proses fotosintesis. Komponen-komponen ini dapat diuraikan menjadi komponen-komponen yang lebih sederhana oleh aktifitas mikroorganisme dan dipergunakan sebagai sumber energi (Enari 1983).
2.3 Bioetanol
Bioetanol merupakan etanol atau kependekan dari etil alkohol (C2H5OH) atau sering juga disebut dengan grain alcohol. Etanol berbentuk cairan tidak berwarna dan mempunyai bau khas. Berat jenis pada suhu 15oC sebesar 0,7937 dan titik didihnya 78,32 oC pada tekanan 76 mmHg. Sifat lainnya adalah larut dalam air dan eter dan mempunyai panas pembakaran 328 Kkal.
Bioetanol dapat dipergunakan sebagai bahan bakar alternatif memiliki beberapa keunggulan yaitu mampu menurunkan emisi CO2 hingga 18 %, bioetanol merupakan bahan bakar yang tidak beracun dan cukup ramah lingkungan serta dihasilkan melalui proses yang cukup sederhana yaitu melalui proses fermentasi menggunakan mikrobia tertentu. Bioetanol sebagai bahan bakar memiliki nilai oktan lebih tinggi dari bensin sehingga dapat menggantikan fungsi aditif seperti metil tertiary butyl ether (MTBE) yang menghasilkan timbal (Pb) pada saat pembakaran. Di Indonesia, minyak bioethanol sangat potensial untuk diolah dan dikembangkan karena bahan bakunya merupakan jenis tanaman yang banyak tumbuh di negara ini dan sangat dikenal masyarakat. Tumbuhan yang potensial untuk menghasilkan bioetanol adalah tanaman yang memiliki kadar karbohidrat tinggi atau selulosa, seperti: tebu, nira, sorgum, ubi kayu, garut, ubi jalar, sagu, jagung, jerami, bonggol jagung, dan kayu.
Tahap inti proses pembuatan bioetanol adalah fermentasi gula baik yang berupa glukosa, fruktosa maupun sukrosa oleh yeast atau ragi terutamaS. cerevisiae dan bakteri
Z. mobilis. Pada proses ini gula dikonversi menjadi etanol dan gas karbon dioksida. Secara umum proses pembuatan bioetanol meliputi tiga tahapan, yaitu persiapan bahan baku, fermentasi dan pemurnian. Pada tahap persiapan, bahan baku berupa padatan terlebih dahulu harus dikonversi menjadi larutan gula sebelum difermentasi menjadi etanol. Untuk bahan-bahan yang sudah berada dalam bentuk larutan seperti molase dapat langsung difermentasi. Proses pengecilan ukuran dengan cara menggiling dapat dilakukan sebelum memasuki tahap pemasakan.
Tahap pemasakan meliputi proses likuifikasi dan sakarifikasi. Pada tahap ini tepung dikonversi menjadi gula melalui proses pemecahan menjadi gula kompleks. Pada tahap likuifikasi dilakukan penambahan air dan enzim alpha amilase. Proses ini dilakukan pada suhu 80-90oC. Berakhirnya proses likuifikasi ditandai dengan parameter cairan seperti sup. Tahap sakarifikasi dilakukan pada suhu 50 – 60oC. Enzim yang ditambahkan pada tahap ini adalah enzim glukoamilase. Pada tahap sakarifikasi akan terjadi pemecahan gula kompleks menjadi gula sederhana.
dihasilkan gas CO2 dengan perbandingan stokiometri yang sama dengan etanol yang dihasilkan yaitu 1 : 1. Setelah melalui proses pemurnian, gas CO2 dapat digunakan sebagai bahan baku gas dalam pembuatan minuman berkarbonat.
Fermentasi adalah suatu proses perubahan kimia pada substrat organik, baik karbohidrat, protein, lemak atau lainnya, melalui kegiatan katalis biokimia yang dikenal sebagai enzim dan dihasilkan oleh jenis mikroba spesifik (Prescott dan Dunn 1981). Secara biokimia fermentasi juga dapat diartikan sebagai pembentukan energi melalui senyawa organik. Secara sederhana proses fermentasi alkohol dari bahan baku yang mengandung gula atau glukosa terlihat pada reaksi berikut:
Glukosa 2C2H5OH + 2CO2+ 2 ATP + 5 Kkal
Dari reaksi diatas, 70% energi bebas yang dihasilkan dibebaskan sebagai panas dan secara teoritis 100% karbohidrat diubah menjadi 51,1% etanol dan 48,9 % menjadi CO2. Fermentasi menurut jenis medianya dapat dibedakan menjadi dua, yaitu fermentasi media padat dan media cair. Fermentasi media padat adalah fermentasi yang subtratnya tidak larut dan tidak mengandung air bebas, tetapi cukup mengandung air untuk keperluan mikroba. Fermentasi media cair adalah proses fermentasi yang subtratnya larut atau tersuspensi dalam media cair. Fermentasi media padat umumnya berlangsung pada media dengan kadar air berkisar antara 60-80 %.
Dalam proses fermentasi, glukosa dapat diubah secara anaerobik menjadi alkohol oleh bermacam-macam mikroorganisme. Khamir sering digunakan dalam proses fermentasi etanol, seperti Saccharomyces cerevisiae, S. uvarum, Schizosaccharomyces sp
dan Kluyveromyces sp. Secara umum khamir dapat tumbuh dan memproduksi etanol secara efisien pada pH 3,5-6,0 dan suhu 28-35oC. Laju awal produksi etanol dengan menggunakan khamir akan meningkat pada suhu yang lebih tinggi, namun produktifitas keseluruhan menurun karena adanya pengaruh peningkatan etanol yang dihasilkan. (Ratledge 1991). Khamir yang sering dipergunakan dalam proses fermentasi etanol adalah Saccharomyces cereviseae. Khamir ini bersifat fakultatif anaerobik, tumbuh baik pada suhu 30oC dan pH 4,0 – 4,5 (Oura 1983).
Produksi etanol dari substrat gula oleh khamir Saccharomyces cereviseae
menjadi biomassa, etanol dan CO2. Terdapat dua parameter yang mengendalikan pertumbuhan dan methabolisme khamir dalam keadaan anaeorobik, yaitu konsentrasi gula dan etanol. Secara kinetik glukosa berperan ganda, pada konsentrasi rendah (kurang dari 1 g/l) merupakan substrat pembatas, sedangkan pada konsentrasi tinggi (lebih dari 300 g/l) akan menjadi penghambat (Mangunwidjaja 1994). Pada permulaan proses fermentasi, khamir memerlukan oksigen untuk pertumbuhannya. Setelah terjadi akumulsi CO2 dan reaksi berubah menjadi anaerob, alkohol yang terbentuk akan menghalangi proses fermentasi lebih lanjut setelah konsentrasi alkohol mencapai 13-15 persen volume dan biasanya maksimum 13 persen volume (Prescott dan Dunn 1981). Selama proses fermentasi juga menimbulkan panas, bila tidak dilakukan pendinginan, maka suhu akan terus meningkat sehingga proses fermentasi terhambat (Oura 1983).
Faktor lingkungan seperti suhu, pH, kebutuhan nutrient dan kofaktor perlu diperhatikan dalam kehidupan khamir. Sejumlah kecil oksigen harus disediakan pada proses fermentasi oleh khamir karena oksigen merupakan komponen yang diperlukan dalam biosintesis beberapa asam lemak tidak jenuh. Untuk kebutuhan oksigen dalam proses fermentasi, biasanya diberikan tekanan oksigen 0,05 – 0,10 mm Hg. Jika tekanan oksigen yang diberikan lebih besar dari nilai tersebut, maka konversi akan cenderung kearah pertumbuhan sel. Kebutuhan relatif nutrien sebanding dengan komponen utama sel khamir, yaitu mencakup karbon, oksigen, nitrogen dan hidrogen. Pada jumlah lebih rendah, fosfor, sulfur, potasium dan magnesium juga harus tersedia untuk sintesis komponen-komponen mineral. Beberapa mineral seperti Mn, Co, Cu dan Zn serta faktor pertumbuhan organik seperti asam amino, asam nukleat dan vitamin diperlukan dalam jumlah besar untuk pertumbuhan khamir.
2.4 Hidrolisis Asam
hydrolisis) (Taherzadeh dan Keikhosro 2007). Pemilihan antara kedua metode kimiawi ini didasarkan pada pertimbangan laju hidrolisis, tingkat degradasi, produk dan biaya total produksi. Perbandingan keuntungan dan kelemahan antara concentrated-acid hydrolisisdengandilute-acid hydrolisisdisajikan pada Tabel 3 berikut ini :
Tabel 3 Perbandingan keuntungan dan kelemahan antara concentrated-acid hydrolisis
dengandilute-acid hydrolisis
Metode hidrolisis Keuntungan Kelemahan
Hidrolisis pada suhu rendah dengan konsentrasi asam tinggi
–Dioperasikan pada suhu
rendah
–Rendemen gula tinggi
– Konsentrasi asam
tinggi
–Korosi peralatan –Energi tinggi untuk
pengambilan asam Hidrolisis pada suhu tinggi
dengan konsentrasi asam rendah
–Konsentrasi asam rendah –Waktu tinggal singkat
–Suhu operasi tinggi –Yield gula rendah – Korosi peralatan
Sumber: Taherzadeh dan Keikhosro (2007).
Hidrolisis asam dengan konsentrasi rendah (dilute-acid) dilakukan dalam dua tahap yaitu: pertama, tahap yang melibatkan asam encer untuk menghidrolisis gula dari golongan pentosa umumnya yang terdapat fraksi hemiselulosa. Tahapan ini biasanya menggunakan 1% H2SO4 pada suhu 80-120oC selama 30-240 menit. Tahap kedua menggunakan asam dengan konsentrasi yang lebih tinggi untuk menghidrolisis gula yang berasal dari golongan heksosa seperti selulosa menjadi glukosa, biasanya dilakukan dengan konsentrasi asam 5-20 % H2SO4 dengan suhu mendekati 180 oC. Dengan menggunakan hidolisis bertahap ini, maka kondisi optimum untuk memaksimalkan hasil glukosa dan miminimumkan hasil samping yang tidak diinginkan (Purwadi 2006). Proses pemisahan antara fraksi gula dengan fraksi asam dapat dilakukan dengan proses pertukaran ion dan asam dapat dikonsentrasikan kembali dengan proses evaporasi (Demirbas 2007).
akan menghasilkan glukosa, manosa, xilosa atau campuran senyawa-senyawa fenolik. Selama proses hidrolisis asam gula pentosa akan menghasilkan furfural dan gula heksosa menghasilkan 5-hidroksimetilfurfural (HMF) (Lopez et al.(2004).
Hidrolisis asam dengan konsentrasi rendah dapat dipergunakan sebagai langkah perlakuan awal (pretreatment) untuk proses hidrolisis secara enzimatik. Perlakuan awal hidrolisis enzimatik pada limbah lignoselulosik menggunakan H2SO40,1-1 % pada suhu 140-190oC akan dapat melemahkan ikatan-ikatan selulosa. Pretreatment dapat dilakukan selama 5 menit pada suhu 180 oC atau 30-90 menit pada suhu 120 oC (Taherzadeh dan Karimi 2007)
2.5 Hidrolisis Enzim
Enzim adalah satu atau beberapa gugus polipeptida atau protein yang berfungsi sebagai katalis dalam suatu reaksi kimia. Enzim bekerja dengan cara menempel pada permukaan molekul zat-zat yang bereaksi sehingga dapat mempercepat proses reaksi. Percepatan terjadi karena enzim menurunkan energi pengaktifan yang dengan sendirinya akan mempermudah terjadinya reaksi. Enzim bekerja secara khas, yang artinya setiap jenis enzim hanya dapat bekerja pada satu macam senyawa atau reaksi kimia. Hal ini disebabkan perbedaan struktur kimia tiap enzim yang bersifat tetap. Sebagai contoh, enzimα-amilase hanya dapat digunakan pada proses perombakan pati menjadi glukosa.
Hidrolisis pati dapat menggunakan enzim α-amilase dan glukoamilase. Enzim α -amilase merupakan endo-enzim yang dapat memecah ikatanα-1,4 glikosidik secara acak dibagian dalam molekul baik pada amilosa maupun pada amilopektinnya. Hasil akhir hidrolisis amilosa adalah glukosa dan maltosa dengan perbandingan 13 % dan 17 %, sedangkan hasil akhir hidrolisis amilopektin menghasilkan campuran limit dekstrin bercabang dan tidak bercabang yang terdiri dari hepta, heksa, penta, tetra dan trisakarida juga maltosa dan isomaltosa disertai sedikit glukosa.
memutuskan rantai pati menjadi molekul-molekul glukosa pada bagian non pereduksi, baik pada ikatanα-1,4 danα-1,6 glikosidik (Tjokroadikoesoemo 1986).
Selulosa dapat dikonversi menjadi produk-produk bernilai ekonomi yang lebih tinggi seperti etanol, glukosa dan pakan ternak dengan jalan menghidrolisis selulosa dengan bantuan selulase sebagai biokatalisator atau dengan hidrolisis asam atau basa. Selulase adalah enzim yang dapat mengkatalis terjadinya reaksi hidrolisis selulosa menjadi glukosa. Keuntungan hidrolisisi ensim dibandingkan dengan hidrolisis asam adalah kondisi reaksi ringan dan tidak terjadi reaksi samping yang berarti.
Enzim selulase dapat diproduksi oleh mikroorganisme, seperti T.viride atau T. reesei. Mikroorganisme selulolitik mampu menghasilkan selulase kompleks, yaitu suatu campuran beberapa jenis selulase yang berbeda. Selulase kompleks mampu menghidrolisis kristal selulosa menjadi gula-gula terlarut secara efisien. Beberapa spesies bakteri yang dapat memproduksi enzim selulase dan hemiselulase adalah
Clostridium, Cellumonas, Thermomonospora, Bacillus, Bacteriodes, Ruminococcus,
Erwinia, Acetovibrio, Microbispora dan Streptomyces, dan jamur seperti Tricoderma, Penicillium, Fusarium, Phanerochaete, Humicola dan Schizophillum spp. Walaupun enzim selulase dapat diproduksi oleh berbagai macam mikroorganisme, enzim selulase dari T. reesei atau T viride telah banyak dipelajari dan mempunyai karakteristik yang paling baik.
Enzim selulase kompleks terdiri dari tiga enzim utama yaitu endoglukanase, eksoglukanase dan selobiase. Endoglukanase menghidrolisis ikatan 1,4 β-glikosidik secara acak pada daerah amorf selulosa menghasilkan glukosa, selobiosa dan selodekstrin. Eksoglukanase menghidrolisis selodekstrin dengan memutus unit selobiosa dari ujung rantai polimer. Selobiase menghidrolisis selobiosa dan selo-oligosakarida menjadi glukosa ( Wuet al.2000; Jeewon 1997).
Faktor-faktor yang berpengaruh pada proses hidrolisis enzim diantaranya yaitu kualitas dan konsentrasi substrat, metode perlakuan awal yang diaplikasikan, aktivitas enzim selulase dan kondisi proses hidrolisis seperti suhu dan pH. Suhu dan pH optimum merupakan fungsi dari bahan, sumber enzim dan waktu hidrolisis. Suhu dan pH optimum pada enzim selulase umumnya pada 40 – 50 oC dan pH 4 – 5, sehingga waktu yang digunakan tergantung pada kondisi tersebut.
Hidrolisis enzimatik tongkol jagung yang diberi perlakuan awal H2SO4 1% pada suhu 180 oC kemudian dihidrolisis dengan enzim selulase kasar T. viride dan enzim glukoamilase kasar A.niger yang dilakukan pada suhu 50 oC, pH 4,8 selama 60 jam menghasilkan 45,7 g/L etanol melalui sistem produksi fed batch(Chenet al.2007)
Salah satu faktor utama yang berpengaruh terhadap hasil yang diperoleh dan kecepatan hidrolisis enzimatis adalah substrat. Konsentrasi substrat yang tinggi dapat menyebabkan penghambat yang memperlambat proses hidrolisis. Terjadinya penghambat oleh substrat tergantung pada perbandingan antara banyaknya enzim terhadap banyaknya substrat. Masalah pengadukan dan perpindahan panas juga akan timbul pada substrat yang berkonsentrasi tinggi. Banyaknya enzim yang ditambahkan pada substrat sangat berpengaruh terhadap kecepatan proses hidrolisis. Semakin banyak enzim yang ditambahkan akan semakin cepat proses hidrolisis yang terjadi dan hasil yang diperoleh juga semakin banyak, tetapi semakin tinggi biaya yang harus dikeluarkan. Banyaknya enzim yang ditambahkan pada substrat biasanya 5 – 35 FPU/gram substrat. Pengurangan biaya untuk penyediaan enzim pada proses hidrolisis enzim dapat dilakukan dengan daur ulang enzim selulase. Bercampurnya enzim dalam hidrolisat dan terbentuknya sisa proses yang berupa padatan (kemungkinan lignin) mempersulit proses pemisahan enzim. Alternatif yang dapat dilakukan untuk mengatasi hal tersebut adalah dengan aplikasi imobilisasi enzim selama proses.
2.6Trichoderma viride
yang dapat menghidrolisis selulosa kristalin dan selulosa non kristalin. Pada enzim selulase dapat terjadi sinergisme antara eksoselulase dengan endoselulase, melainkan juga antar eksoselulase.
Pertumbuhan T. viride optimal pada pH sekitar 4,0, sedangkan untuk produksi enzim selulase mendekati ph 3,0. Selama produksi enzim, pH harus dipertahankan dalam kisaran 3,0 – 4,0 karena inaktivasi enzim akan terjadi di bawah pH 2,0. Suhu optimum pertumbuhan sekitar 32 – 35 oC dan untuk produksi enzim sekitar 25 – 28 oC. Karakteristik dari enzim selulase T. viride adalah memiliki pH optimum 4,0 dan akan tetap stabil pada pH 3 – 7. suhu optimum adalah 50 oC dan aktivitasnya akan menurun jika suhunya lebih dari 50oC.
T. viride selain mampu memproduksi enzim selulase, juga dapat menghasilkan enzim endo-1,4-β-xilanase yang dapat mendegradasi xilan. Berat molekul xilanase yang dihasilkan dari T. viride adalah sebesar 22.000 dalton ( Ujiie et al. 1991; Tholudur 1999). Palmvist et al. (1997) dan Larsson et al. (1999), melaporkan Trichoderma
mampu secara simultan melakukan proses detoksifikasi dan produksi enzim secara simultan pada hidrolisat asam yang mengandung senyawa-senyawa inhibitor seperti furfural dan HMF. Kapang ini juga mampu memetabolisme gula dari golongan pentosa maupun heksosa dan tidak terlalu sensitif terhadap material-material lignoselulosik.
2.7 Aspergillus niger
Aspergillus niger termasuk genus Aspergillus, famili Eurotiaceae dan ordo Eurotiales. Kapang ini mempunyai miselium bercabang dan berseptat. Kapang umumnya bersifat aerob dan tumbuh baik pada kisaran suhu 25 – 30 oC, namun genus Aspergillus
dapat tumbuh pada kisaran suhu 35 – 37oC. Kapang ini dapat tumbuh dengan baik pada suhu 30 oC dengan pH optimum 7,0 atau agak asam dan besifat tidak tahan panas. A. niger dalam media pertumbuhan dapat langsung mengkonsumsi molekul-molekul sederhana seperti gula dan komponen lain yang larut disekitar hifa, namun untuk molekul-molekul yang lebih kompleks seperti selulosa, pati dan protein harus dipecah terlebih dahulu sebelum masuk kedalam sel.
enzim akan menurun pada suhu lebih dari 30 oC karena energi respirasi lebih banyak dipergunakan untuk pembentukan spora dari pada untuk membentuk miselium.
A. niger dikenal sebagai kapang penghasil asam sitrat, anilin, pektinase, selulase,
β-1,4-glikan hidrolase, protease, α-amilase, glukoamilase, maltase, β-galaktosidase, α -glukosidase, β-glukosidase, asam glukonat, glukosa oksidase, asam oksalat, fosfodiestrase, ribonuklease, pupulan 4- glukanohidrolase, β-xilosidase, xilanase dan lipase. Glukoamilase dari A. niger menunjukkan bobot molekul berkisar 54-112 k D dan pH optimum berkisar antara 4,0-5,0. Temperatur optimum aktivasi berkisar antara 40 – 65oC ( Selvakumaret al.1996).
2.8 Saccharomyces cerevisiae
Saccharomyces cerevisiae termasuk ke dalam kelas Ascomycetes yang dicirikan dengan pembentukan askus yang merupakan tempat pembentukan askospora. S. serevisiae memperbanyak diri secara aseksual yaitu dengan bertunas (Pelezar dan Chan 1986). Dinding selS. cerevisiae terdiri dari komponen-komonen glukan, manan, protein, kitin dan lemak (Waluyo 2004).
Saccharomyces cerevisiae sering digunakan dalam fermentasi etanol karena sangat tahan dan toleran terhadap kadar etanol yang tinggi (12-18% v/v), tahan pada kadar gula yang cukup tinggi dan tetap aktif melakukan fermentasi pada suhu 4-32oC.S. cerevisiaemempunyai aktivitas optimum pada suhu 30 – 35oC dan tidak aktif pada suhu lebih dari 40 oC. S. cerevisiae dapat memfermentasi glukosa, sukrosa, galaktosa serta rafinosa (Kunkee dan Mardon 1970). Biakan S. cerevisiae mempunyai kecepatan fermentasi optimum pada pH 4,48 (Harrison dan Graham 1970)
Rendemen alkohol dari heksosa dalam fermentasi menggunakan khamir dari genus Saccharomyces dapat mencapai 90 % (Boyles 1984). Proses fermentasi oleh
Glukosa
Glukosa-6-P
Fruktosa-6-P
Fruktosa-1,6-di-P
Gliseraldehida-3-P Dihidroksiaseton fosfat
Gliseraldehida-3-P
1,3-di fosfogliserat
3-fosfogliserat
2-fosfogliserat
Fosfoenolpiruvat
Piruvat
Glukosa
Glukosa-6-P
Fruktosa-6-P
Fruktosa-1,6-di-P
Gliseraldehida-3-P Dihidroksiaseton fosfat
Gliseraldehida-3-P
1,3-di fosfogliserat
3-fosfogliserat
2-fosfogliserat
Fosfoenolpiruvat
Piruvat
Glukosa
Glukosa-6-P
Fruktosa-6-P
Fruktosa-1,6-di-P
Gliseraldehida-3-P Dihidroksiaseton fosfat
Gliseraldehida-3-P
1,3-di fosfogliserat
3-fosfogliserat
2-fosfogliserat
Fosfoenolpiruvat
[image:36.612.164.482.78.418.2]Piruvat
Gambar 3 Mekanisme proses glikolisis
III. METODOLOGI PENELITIAN
3.1 Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan dari bulan Maret sampai Juli 2009, bertempat di Laboratorium Mikrobiologi PAU IPB dan Laboratorium Bioindustri Teknologi Industri Pertanian IPB. Beberapa laboratorium penunjang antara lain Laboratorium Instrumentasi Teknologi Industri Pertanian IPB dan Laboratorium Dasar dan Ilmu Terapan Teknologi Industri Pertanian IPB.
3.2 Bahan dan Alat 3.2.1 Bahan
Bahan baku yang digunakan dalam penelitian ini adalah ubi kayu varietas Darul Hidayah yang diperoleh dari Sukabumi. Mikroorganisme yang digunakan untuk fermentasi adalahSaccharomyces cerevisiae,Trichoderma viridedan Aspergillus niger
yang diperoleh dari Laboratorium Mikrobiologi PAU IPB. Bahan untuk proses likuifikasi, sakarifikasi dan fermentasi meliputi: PDA, NPK, ZA , H2SO4, CMC, AMG,
α-amilase, selulase komersial. Bahan kimia untuk keperluan analisa meliputi: HCl 1 N, larutan schroll H2SO425%, Na2S2O30,1 N, fenol 5%, H2SO4.
3.2.2 Alat
Alat-alat yang digunakan dalam penelitian adalah oven, mixer, timbangan analitik, incubator, otoklaf, termometer, erlenmeyer, pH meter, gelas ukur, alat distilasi, kertas saring, mikropipet, spektrofotometer dan GC (Gas Chromatography)
3.3 Tahapan Penelitian
3.3.1 Karakteristik ubi kayu
Karakterisasi ubi kayu dilakukan dengan tujuan untuk mengetahui karakteristik bahan baku yang meliputi analisis kadar air, lemak, protein, pati, abu, selulosa, hemiselulosa dan serat kasar. Prosedur analisis dapat dilihat pada Lampiran 1.
3.3.2 Persiapan kulturT. viride.
[image:38.595.85.509.358.695.2]Kultur T. viride sebelum dipergunakan disegarkan supaya dapat memproduksi selulase dengan optimal. T. viride digoreskan dalam media PDA (Potato Dextrose Agar) yang dibuat miring pada tabung reaksi dan dibiarkan tumbuh dalam inkubator pada suhu 28oC. Pertumbuhan maksimal T. viride ditentukan dengan membuat kurva pertumbuhan jumlah spora yang diamati setiap hari selama 7 hari. Jumlah spora maksimal yang diperoleh selanjutnya dipergunakan untuk memproduksi enzim selulase.
3.3.3 Persiapan kulturA. niger
Isolat A. niger diperbanyak dan diremajakan dengan menginokulasikannya ke dalam media padat aagr miring PDA, kemudian diinkubasi pada suhu kamar 25 – 28oC selama 5 hari. Media padat dibuat dengan cara diambil 3,9 gram media PDA dilarutkan dalam 100 ml air kemudian disterilisasi pada suhu 121oC selama 15 menit.
3.3.4 Persiapan kulturS. cerevisiae
Isolat yeast Saccharomyces cerevisiae diremajakan pada media PDA dan diinkubasi selama 2 hari. Setelah itu isolat ditumbuhkan lagi pada 50 ml media YMGP yang terdiri dari ekstrak yeast 5 g/l, malt 5 g/l, glukosa 10 g/l dan pepton 5 g/l di dalam erlenmeyer 200 ml. Inkubasi dilakukan pada shaker berkecepatan 125 rpm dengan suhu 30°C selama 24 jam.
3.3.5 Produksi enzim glukoamilaseA. niger
Isolat A. niger yang telah diremajakan disuspensikan dengan larutan buffer fosfat 10 ml untuk setiap tabung agar miring. Larutan spora yang diperoleh dimasukkan ke dalam media fermentasi yang telah disterilkan dengan konsentrasi 10 %. Media fermentasi cair yang dipergunakan terdiri dari substrat tapioka dan larutan mineral. Produksi dilakukan dalam erlenmeyer 250 ml dengan volume kerja 100 ml. Media produksi kemudian ditutup dengan kapas dan disterilisasi dalam otoklaf suhu 121 oC selama 15 menit. Larutan mineral terdiri dari 1 % tapioka, 0,14 % (NH4)2SO4, 0,01 % MgSO4. 7H2O, 0,60 % KH2PO4, 0,20 % K2HPO4, 0,50 ekstrak yeast dan 0,20 pepton. Inkubasi dilakukan pada inkubator goyang dengan kecepatan 100 rpm selama 7 hari pada huhu 30 oC. Pada masa akhir inkubasi dilakukan pengukuran aktivitas enzim glukoamilase (Gomeset al.2005).
3.3.6 Produksi enzim selulaseT. viride
ZnSO4.H2O, 1,25 gr Co(NO3)2. 6H2O. Untuk produksi selulase ditambahkan 5 -10 gr onggok dan 0,5-1,0 gr polipepton untuk 1 liter media.
Proses produksi dilakukan pada suhu 30 oC dan pH 4,0 (Jenie, 1990). Kemampuan untuk menghasilkan enzim selulase diamati dengan mengukur antivitas FP-ase dan CMC-Ase setiap 24 jam selama 7 hari. Kurva aktivitas enzim dibuat dan dipergunakan untuk melihat waktu tercapainya aktivitas maksimum T. viride untuk menghasilkan selulase.
3.3.7 Pembuatan etanol
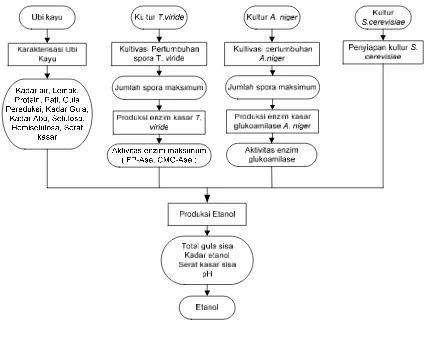
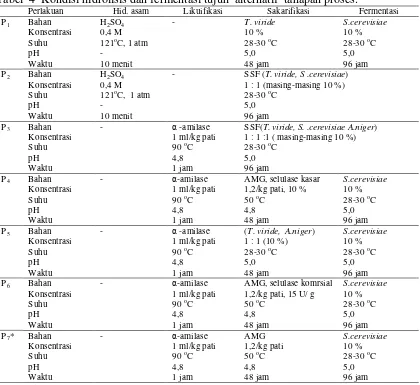
Tahapan pembuatan etanol dalam penelitian dilakukan melalui sistem batch
dengan tujuh perlakuan proses fermentasi (Lampiran 2). Proses hidrolisis dilakukan secara asam dan enzimatik. Penggunaan kultur campuran T. viride, A. niger dan S. cerevisiaedilakukan dengan dua cara yaitu secara SFS dan secara bertahap. Kultur yang ditambahkan sebanyak 10 % dari total volume substrat. Kondisi hidrolisis dan fermentasi tujuh alternatif tahapan proses disajikan pada Tabel 4.
Masing-masing alternatif proses diulang tiga kali dan diamati secara periodik selama 4 hari terhadap perubahan total gula dan pH selama fermentasi. Pada akhir fermentasi dianalisa kadar etanol dan sisa serat kasar. Dari data yang diperoleh dihitung efisiensi fermentasi, rendemen dan efisiensi penggunaan substrat. Pemilihan proses produksi terbaik didasarkan pada produksi etanol tertinggi. Efisiensi fermentasi merupakan persentase perbandingan antara konsentrasi etanol yang diperoleh secara aktual dengan konsentrasi etanol secara teoritis berdasarkan jumlah gula yang dikonsumsi. Rendemen merupakan persentase perbandingan volume etanol yang diperoleh dengan bobot tepung yang dipergunakan.
Rendemen (% v/b) = x100%
kayu ubi ng Bobot tepu aktual secara diperoleh yang etanol Volume
Efisiensi substrat (ds/s) = x100% S S -S o o
Efisiensi fermentasi = x100%
Tabel 4 Kondisi hidrolisis dan fermentasi tujuh alternatif tahapan proses.
Perlakuan Hid. asam Likuifikasi Sakarifikasi Fermentasi P1 Bahan
Konsentrasi Suhu pH Waktu
H2SO4
0,4 M 121oC, 1 atm
-10 menit
- T. viride
10 % 28-30oC 5,0 48 jam
S.cerevisiae
10 % 28-30oC 5,0 96 jam P2 Bahan
Konsentrasi Suhu pH Waktu
H2SO4
0,4 M 121oC, 1 atm
-10 menit
- SSF (T. viride, S .cerevisiae) 1 : 1 (masing-masing 10 %) 28-30oC
5,0 96 jam P3 Bahan
Konsentrasi Suhu pH Waktu
- α-amilase 1 ml/kg pati 90oC 4,8 1 jam
SSF(T. viride, S. .cerevisiae A.niger)
1 : 1 :1 ( masing-masing 10 %) 28-30oC
5,0 96 jam P4 Bahan
Konsentrasi Suhu pH Waktu
- α-amilase 1 ml/kg pati 90oC 4,8 1 jam
AMG, selulase kasar 1,2/kg pati, 10 % 50oC
4,8 48 jam
S.cerevisiae
10 % 28-30oC 5,0 96 jam P5 Bahan
Konsentrasi Suhu pH Waktu
- α-amilase 1 ml/kg pati 90oC 4,8 1 jam
(T. viride, A.niger)
1 : 1 (10 %) 28-30oC 5,0 48 jam
S.cerevisiae
10 % 28-30oC 5,0 96 jam P6 Bahan
Konsentrasi Suhu pH Waktu
- α-amilase 1 ml/kg pati 90oC
4,8 1 jam
AMG, selulase komrsial 1,2/kg pati, 15 U/ g 50oC
4,8 48 jam
S.cerevisiae
10 % 28-30oC
5,0 96 jam P7* Bahan
Konsentrasi Suhu pH Waktu
- α-amilase 1 ml/kg pati 90oC 4,8 1 jam
AMG 1,2/kg pati 50oC 4,8 48 jam
S.cerevisiae
10 % 28-30oC 5,0 96 jam * Kontrol
3.4 Teknik Analisis Data 3.4.1 Produksi enzim selulase
Persamaan model rancangannya sebagai berikut:
ij
Yij i
dimana :
Yij : Parameter pengamatan pada perlakuan lama fermentasi ke-i dan ulangan ke-j
μ : Nilai rata-rata umum
βi : Pengaruh perlakuan lama fermentasi ke-i
εij : Galat percobaan akibat perlakuan lama fermentasi ke-i dan ulangan ke-j i : Taraf lama fermentasi (1, 2,...,7 hari)
j : Ulangan (1, 2, 3)
3.4.2 Produksi enzim amiloglukosidase
Untuk mengetahui pengaruh lama fermentasi terhadap besarnya aktivitas AMG, maka dilakukan Uji F dengan Rancangan Acak Lengkap (RAL) faktor tunggal. Faktornya adalah lama fermentasi dengan tujuh taraf perlakuan yaitu 1 hari, 2 hari, 3 hari, 4 hari, 5 hari, 6 hari dan 7 hari. Masing-masing perlakuan diulang 3 kali. Apabila perlakuan lama fermentasi berpengaruh nyata, maka dilanjutkan dengan Uji Duncan. Parameter yang diamati adalah aktivitas AMG. Persamaan model rancangannya sebagai berikut:
ij
Yij i
dimana :
Yij : Parameter pengamatan pada perlakuan lama fermentasi ke-i dan ulangan ke-j
μ : Nilai rata-rata umum
βi : Pengaruh perlakuan lama fermentasi ke-i
εij : Galat percobaan akibat perlakuan lama fermentasi ke-i dan ulangan ke-j i : Taraf lama fermentasi (1, 2,...,7 hari)
3.4.3 Proses hidrolisis
Untuk mengetahui pengaruh jenis proses hidrolisis terhadap besarnya konsentrasi total gula dan gula pereduksi, maka dilakukan Uji F dengan Rancangan Acak Lengkap (RAL) faktor tunggal. Faktornya adalah jenis proses hidrolisis dengan empat taraf perlakuan yaitu :
1. Hidrolisis menggunakan asam (H2SO40,4 M)
2. Hidrolisis menggunakan enzimα-amilase dan amiloglukosidase
3. Hidrolisis menggunakan enzimα-amilase, AMG komersial dan selulase kasar 4. Hidrolisis menggunakan enzimα-amilase, AMG komersial dan selulase komersial.
Masing-masing perlakuan diulang 3 kali. Apabila perlakuan lama fermentasi berpengaruh nyata, maka dilanjutkan dengan Uji Duncan. Parameter yang diamati adalah konsentrasi total gula dan gula pereduksi. Persamaan model rancangannya sebagai berikut:
ij
Yij i
dimana :
Yij : Parameter pengamatan pada perlakuan jenis proses hidrolisis ke-i dan ulangan ke-j
μ : Nilai rata-rata umum
βi : Pengaruh perlakuan jenis proses hidrolisis ke-i
εij : Galat percobaan akibat perlakuan lama fermentasi ke-i dan ulangan ke-j i : Taraf jenis proses hidrolisis (1, 2, 3,4)
j : Ulangan (1, 2, 3)
3.4.4 Proses fermentasi
Persamaan model rancangannya sebagai berikut:
ij
Yij i
dimana :
Yij : Parameter pengamatan pada perlakuan alternatif proses fermentasi ke-i dan ulangan ke-j
μ : Nilai rata-rata umum
βi : Pengaruh perlakuan alternatif proses fermentasi ke-i
εij : Galat percobaan akibat perlakuan alternatif proses fermentasi ke-i dan ulangan ke-j
IV. HASIL DAN PEMBAHASAN
4.1 Komposisi Kimia Ubi Kayu
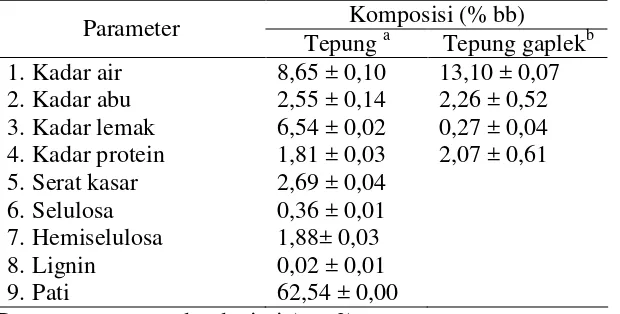
[image:45.595.87.397.323.480.2]Ubi kayu yang dipergunakan dalam penelitian ini adalah ubi kayu varietas Darul Hidayah yang diperoleh dari Daerah Sukabumi, Jawa Barat. Ubi kayu sebelum dipergunakan sebagai substrat fermentasi terlebih dahulu dijadikan tepung dengan ukuran 40 mesh kemudian dianalisis komposisi kimianya, yang meliputi kadar air, abu, lemak, protein dan karbohidrat. Hasil analisis komposisi kimia tepung ubi kayu disajikan pada Tabel 5.
Tabel 5 Hasil analisis komposisi tepung kimia ubi kayu. Komposisi (% bb) Parameter
Tepunga Tepung gaplekb 1. Kadar air
2. Kadar abu 3. Kadar lemak 4. Kadar protein 5. Serat kasar 6. Selulosa 7. Hemiselulosa 8. Lignin 9. Pati
8,65 ± 0,10 2,55 ± 0,14 6,54 ± 0,02 1,81 ± 0,03 2,69 ± 0,04 0,36 ± 0,01 1,88± 0,03 0,02 ± 0,01 62,54 ± 0,00
13,10 ± 0,07 2,26 ± 0,52 0,27 ± 0,04 2,07 ± 0,61
Data : rerata ± standar deviasi (n = 2) a
Analisis proksimat (2009),bRichana et al.(1990).
Kandungan air dalam bahan berpengaruh terhadap kesegaran dan daya simpan. Kandungan air ini dipengaruhi oleh varietas, umur tanam, unsur hara tanah dan iklim. Kandungan air dalam ubi kayu segar sekitar 60,30 % (Padonou et al. 2005). Berdasarkan hasil pengukuran kadar air bahan baku tepung ubi kayu yang dipergunakan mengandung sekitar 8,65 ± 0,10 % (bb), sedangkan menurut Richanaet al. (2004) yaitu sekitar 13,10 ± 0,07 %.
Kadar lemak dan protein pada bahan masing-masing sebesar 6,54 ± 0,02 % (bb) dan 1,81 ± 0,03 % (bb). Kadar lemak pada bahan yang dipergunakan konsentrasinya lebih tinggi dibandingkan dengan kadar lemak yang dilaporkan oleh Richana et al.
(2004) yaitu sekitar 0,27 ± 0,04 %, sedangkan menurut Padonou et al. (2005), kadar lemak tepung ubi kayu sekitar 0,56 % (bb). Kandungan lemak dan protein dalam bahan berpengaruh terhadap karakteristik gelatinisasi dan kekentalan bahan pada saat diolah. Lemak pada bahan yang mengandung pati dapat mengganggu proses gelatinisasi karena dapat membentuk kompleks dengan amilosa sehingga menghambat keluarnya amilosa dari granula pati, sedangkan protein dapat menyebabkan kekentalan pati menurun (Mohamed 2003).
Karbohidrat terdiri dari fraksi pati dan serat kasar. Kedua fraksi ini merupakan bagian penting yang akan dipergunakan sebagai substrat fermentasi. Fraksi serat kasar terutama terdiri dari selulosa, hemiselulosa dan lignin. Pati dan selulosa merupakan homopolimer glukosa yang jika dihidrolisis akan menghasilkan glukosa, sedangkan hemiselulosa akan menghasilkan campuran gula yang terdiri dari glukosa, xilosa, galaktosa, arabinopiranosa, arabinofuranosa dan manosa. Glukosa, manosa dan galaktosa merupakan gula dari golongan heksosa, sedangkan xylosa dan arabinosa merupakan gula dari golongan pentosa (Demirbas 2005; Irawadi 1990). Dari hasil analisis komposisi kimia menunjukkan bahwa ubi kayu yang dipergunakan sebagai bahan dalam penelitian ini mengandung karbohidrat 80,45 ± 0,23 % (bb) dengan kandungan pati 62,35 ± 0,00 % dan serat kasar 2,69 ± 0,04 % (bb). Fraksi serat kasar mengandung selulosa 0,36 ± 0,01 % (bb), hemiselulosa 1,88 ± 0,03 % (bb) dan lignin 0,02 ± 0,01 % (bb). Menurut Richanaet al. (2004), kadar karbohidrat dalam tepung ubi kayu adalah 82,30 ± 0,31 %. Perbedaan dalam komposisi karbohidrat ubi kayu dapat disebabkan oleh adanya perbedaan varietas, umur panen dan musim panen. Wargiono
et al.(2006) menyatakan bahwa hasil ubi segar dan pati ubi kayu dengan umur panen 8 bulan dapat mencapai 16,19 ton/ha dengan hasil pati 2,31 ton/ha, sedangkan pada umur panen 18 bulan dapat mencapai 45,25 ton/ha dengan hasil pati 9,19 ton/ha.
dihidrolisis disebut bagian kristalin. Selulosa mempunyai sifat mengembang (sweeling) jika direaksikan dengan hidroksi logam alkali, garam-garam dalam larutan basa kuat dan senyawa amina. Dari sekian senyawa itu, NaOH paling lazim dipergunakan untuk mengembangkan ikatan selulosa (Irawadi 1990)
Kandungan karbohidrat yang cukup tinggi menunjukkan ubi kayu mempunyai potensi sebagai sumber glukosa dalam substrat fermentasi. Kadar lignin dalam ubi kayu ini relatif kecil. Kandungan serat kasar dapat menurunkan efisiensi hidrolisis sehingga meningkatkan dosis enzim yang diperlukan, sedangkan kandungan lignin yang tinggi dapat menghambat proses hidrolisis karena membentuk kompleks dengan selulosa dan hemiselulosa.
4.2 Kultivasi dan Produksi Enzim
[image:47.595.109.487.433.643.2]Kurva pertumbuhan sangat penting dipergunakan untuk menggambarkan karakteristik pertumbuhan mikroba, sehingga memudahkan proses kultivasi dalam suatu media. Pertumbuhan mikroba dipengaruhi oleh beberapa faktor seperti suhu, pH, aktivitas air, adanya oksigen dan ketersediaan nutrisi makanan.
Gambar 5 InokulumT. viridedanA. niger pada media PDA
Hari ke-2 Hari ke-4 Hari ke-7
4.2.1 Kultivasi Trichoderma viride
Kultivasi pertumbuhan dilakukan untuk menentukan waktu yang diperlukan oleh
T. viride untuk menghasilkan spora maksimum yang selanjutnya akan dipergunakan sebagai kultur dalam media produksi selulase. Kultivasi kultur dilakukan dengan menghitung jumlah spora yang terbentuk setiap hari selama 7 hari.
Pada awal inokulasi terdapat rata-rata jumlah spora 7,08 x 107/ml. Pada akhir hari ke-1 jumlah spora mengalami penurunan yang cukup signifikan karena spora diduga telah mengalami germinasi membentuk miselium berwarna putih (Gambar 5). Pada hari berikutnya mulai terbentuk spora berwarna putih dengan jumlah rata-rata 1,02 x 108/ml. T. viride mengalami fase untuk menghasilkan spora dengan cepat mulai dari hari ke-1 hingga hari ke-3.
[image:48.595.129.471.469.652.2]Setelah hari ke-3, T. viride mulai menunjukkan fase pembentukan spora relatif lambat. Pada fase ini miselium dan spora yang dihasilkan masih berwarna putih kehijauan. Perubahan warna miselium dan spora dari putih menjadi hijau mulai terbentuk setelah kultivasi selama 6 hari. Jumlah spora maksimum dengan rata-rata spora 1,58 x 109/ml terjadi pada hari ke-6. Kurva pertumbuhanT. viridedisajikan pada Gambar 6. 0 1 2 3 4 5 6
0 1 2 3 4 5 6 7
Lama fermentasi (hari)
A k ti v it a s C M C -a s e F P -a s e (U /m l) 0,E+00 4,E+08 8,E+08 1,E+09 2,E+09 2,E+09 J u m la h s p o ra (p e r m l)
CMC-ase (U/ml) PF-ase (U/ml) Jml spora
4.2.2 Produksi enzim selulase
Pada penelitian ini produksi enzim selulase menggunakan Media Andreoti yang dimodifikasi (Jenie 1990). Modifikasi dilakukan dengan mengganti selulosa murni sebagai bahan penginduksi selulase menggunakan onggok ubi kayu. Onggok dapat dipergunakan sebagai bahan penginduksi selulase karena mengandung serat kasar terutama selulosa yang dapat dipergunakan sebagai sumber karbon untuk pertumbuhan mikroba. Selain itu, selulosa juga merupakan senyawa penginduksi sintesis enzim selulase. Kandungan serat kasar onggok rata-rata 6,58 ± 0,08 % (b/b). Konsentrasi onggok yang dipergunakan dalam penelitian ini adalah 10 g/L. Menurut Richanaet al.
(2004), kadar serat kasar onggok adalah 9,7 % yang terdiri dari lignin 1,3 %, xilan 5,8 % dan selulosa 2,61 %.
Penentuan pengaruh lama fermentasi yang menghasilkan aktivitas paling tinggi dilakukan dengan mengukur besarnya aktivitas enzim kasar setiap hari selama 1 minggu. Pengukuran antivitas enzim ini meliputi CMC-ase dan FP-ase. Dari hasil analisis keragaman diperoleh bahwa lama fermentasi memberi pengaruh nyata terhadap besarnya aktivitas CMC-ase dan FP-ase (p-value<0,05) (Lampiran 5)
Pada Gambar 5 menunjukkan bahwa pada awal fermentasi terjadi penurunan aktivitas CMC-ase sampai hari ke-4. Setelah itu aktivitas enzim CMC-ase cenderung mengalami peningkatan dan aktivitas maksimal sebesar 5,05 ± 0,42 U/ml diperoleh setelah fermentasi selama 7 hari atau 1 minggu, namun berdasarkan uji lanjut Duncan, aktivitas CMC-ase ini tidak berbeda nyata pada fermentasi selama 6 hari.
Hal serupa juga ditunjukkan pada besarnya aktivitas FP-ase, dimana pada awal fermentasi juga terjadi penurunan aktivitas sampai hari ke-3 dan pada hari berikutnya mengalami peningkatan. Aktivitas maksimal sebesar 4,77 ± 0,72 U/ml juga diperoleh setelah fermentasi selama 7 hari, namun berdasarkan uji lanjut Duncan, aktivitas FP-ase ini tidak berbeda nyata pada fermentasi selama 6 hari. Jenie (1990) memproduksi selulase kasar dari T. reesei selama 9 hari menggunakan substrat onggok diperoleh besarnya aktivitas CMC-ase 12,7 U/ml dan FP-ase sebesar 0,88 U/ml.
sel. Suhartono (1992) menjelaskan bahwa sintesis enzim ekstraseluler dalam jumlah terbesar secara normal terjadi pada saat sebelum sporulasi, yaitu pada akhir fase eksponensial dan awal fase stasioner. Keadaan tersebut diduga karena pada masa transisi fase eksponensial diikuti dengan penurunan jumlah sumber karbon dalam medium, sehingga sintesis enzim selulase mulai meningkat.
Terjadinya peningkatan aktivitas enzim pada proses fermentasi diduga disebabkan oleh adanya perubahan pH dari pH awal 4,0 menjadi 3,28 pada hari ke-7 Enari (1983) menyebutkan bahwa pH optimal untuk pertumbuhan Trichodermasekitar 4,0, sedangkan untuk produksi selulase mendekati 3,0. Selama produksi enzim, pH harus dipertahankan dalam kisaran 3,0-4,0 karena inaktivasi enzim akan terjadi jika pH berada dibawah 2,0. Penurunan pH yang terjadi pada produksi selulase berhubungan langsung dengan adanya konsumsi karbohidrat yang terdapat pada onggok. Pola perubahan pH selama fermentasi untuk produksi enzim selulase disajikan pada Lampiran 5.
4.2.3 KultivasiAspergillus nigerdan produksi amiloglukosidase
Kultivasi kultur A. niger juga dilakukan selama 7 hari dengan melakukan perhitungan jumlah spora yang terbentuk setiap hari. Pada awal inokulasi terdapat rata-rata jumlah spora 3,47 x 106/ml. Pada akhir hari ke-1 spora mengalami penurunan karena diduga spora mengalami germinasi membentuk miselium berwarna hitam (Gambar 5). Pada hari berikutnya mulai terbentuk spora dengan jumlah rata-rata 3,45 x 108/ml. Mikroba menghasilkan spora dengan cepat mulai hari ke-1 hingga hari ke-4 dengan spora berwarna hitam. Setelah hari ke-4 laju pembentukan spora relatif lambat dan jumlah spora maksimum mulai terjadi pada hari ke-6 dengan jumlah rata-rata spora 1,33 x 109/ml. Kurva pertumbuhanA. nigerdisajikan pada Gambar 7.
Duncan, aktivitas ini berbeda nyata dengan semua perlakuan lama fermentasi (Lampiran 5). 0 20 40 60 80
0 1 2 3 4 5 6 7
Lama fermentasi (Hari)
[image:51.595.143.459.148.326.2]A k ti v it a s A M G (U /m l) -1,00E+08 4,00E+08 9,00E+08 1,40E+09 1,90E+09 J m l s p o ra (p e r m l) AMG spora
Gambar 7 Kurva pertumbuhanA. nigerdan aktivitas glukoamilase
Dari Gambar 7 terlihat bahwa aktivitas enzim glukoamilase pada fermentasi hari ke-1 sampai hari ke-3 mengalami penurunan dari 32,41 U/ml -26,47 U/ml, hal ini dapat disebabkan enzim dalam masa penyesuaian, sedangkan pada hari ke-4 sampai hari ke-7 aktivitas enzim semakin meningkat. Ini menunjukkan semakin lama difermentasi aktivitas glukoamilase semakin besar dalam menghidrolisis pati menjadi monomer glukosa.
Perbedaan aktivitas enzim dengan variasi waktu fermentasi dapat disebabkan meningkatnya aktivitas glukoamilase I pada kompleks glukoamilase A. niger pada kondisi waktu fermentasi yang lebih lama. Hal ini dapat pula disebabkan oleh adanya perubahan pH selama fermentasi (Kombong 2004)
Enzim glukoamilase I merupakan komponen kompleks glukoamilase yang aktif menghidrolisis ikatanα-1,6 glikosidik pada rantai cabang pati dan glukoamilase II aktif menghidrolisis ikatanα-1,4 glikosidik pada rantai lurus pati menjadi monomer glukosa.
segera setelah semua karbohidrat dikonsumsi. Perubahan pH selama fermentasi produksi enzim glukoamilase dapat dilihat pada Lampiran 5.
4. 3 Proses Hidrolisis dan Karakteristik Hidrolisat
Proses pembuatan hidrolisat dilakukan dengan menghidrolisis fraksi pati dan serat yang terkandung pada bahan. Hidrolisis dilakukan dengan tujuan untuk menyediakan glukosa yang akan dipergunakan sebagai substrat S. cerevisiae dalam proses fermentasi. Pada dasarnya prinsip hidrolisis adalah memutuskan rantai polimer bahan menjadi unit-unit monomer yang lebih sederhana. Pemutusan rantai polimer tersebut dapat dilakukan dengan berbagai metode, misalnya secara enzimatis, kimiawi ataupun kombinasi keduanya. Hidrolisis secara enzimatis memiliki perbedaan mendasar dibandingkan hidrolisis secara kimiawi dan fisik dalam hal spesifitas pemutusan rantai polimer bahan. Hidrolisis secara kimiawi dan fisik akan memutus rantai polimer secara acak, sedangkan hidrolisis enzimatis akan memutus rantai polimer secara spesifik pada percabangan tertentu.
Dalam penelitian ini, pembuatan hidrolisat dilakukan dengan 2 cara yaitu hidrolisis menggunakan asam dan enzim. Asam yang dipergunakan adalah H2SO40,4 M dengan waktu hidrolisis 10 menit pada suhu 121oC, tekanan 1 atm. Penggunaan H2SO4 0,4 M diharapkan hanya menghidrolisis fraksi pati tanpa menghidrolisis fraksi seratnya.
Hidrolisis enzimatik mempunyai beberapa keuntungan, yaitu menghasilkan produk yang lebih spesifik sesuai dengan yang diinginkan, kondisi proses dapat dikontrol dan lebih sedikit menghasilkan produk samping. Enzim yang dipergunakan meliputiα-amilase, amiloglukosidase komersial (AMG), selulase komersial dan selulase kasar dari T. viride. Enzim α-amilase dan AMG dipergunakan untuk menghidrolisis fraksi pati menjadi glukosa, sedangkan selulase komersial dan selulase kasar dipergunakan untuk menghidrolisis fraksi serat atau selulosa menjadi glukosa atau gula-gula sederhana lainnya yang merupakan monosakarida.
90 oC selama 1 jam. Dosis enzim yang dipergunakan dalam penelitian ini adalah 1 ml/kg pati (Budiyanto et al. 2005 ). Likuifikasi merupakan proses pencairan pati yang telah mengalami gelatinisasi. Gelatinisasi dapat dilakukan dengan melakukan pemanasan pati di dalam air sehingga granula pati mulai mengembang sehingga kekentalannya meningkat (Thomas dan Atwell 1997). Adanya proses gelatinisasi menyebabkan ikatan-ikatan antar molekul pati lebih lemah sehingga kerja enzim dapat lebih mudah.
[image:53.595.87.509.467.567.2]Tahap sakarifikasi dilakukan pada suhu 50 oC selama 48 jam. Enzim yang ditambahkan pada tahap ini adalah AMG yang berfungsi untuk memutuskan rantai pati menjadi molekul-molekul glukosa pada bagian non pereduksi, baik pada ikatan α-1,4 maupun α-1,6 glikosida dan menghasilkan unit-unit glukosa. Dosis AMG yang dipergunakan sebesar 1,2 ml/kg pati (Budiyanto et al. 2005 ). Beberapa perlakuan hidrolisis pada tahap sakarifikasi ditambahkan enzim selulase baik yang komersial maupun filtrat enzim selulase kasar. Enzim selulase yang ditambahkan pada tahap sakarifikasi ini diharapkan dapat menghidrolisis fraksi serat terutama selulosa yang mempunyai ikatan β-1,4 glikosida untuk menghasilkan glukosa. Karakteristik hasil hidrolisis asam dan enzim disajikan pada Tabel 6.
Tabel 6 Karakteristik hasil hidrolisis asam dan enzim
Hidrolisis Total gula
(% b/v)
Gula reduksi (% b/v)
DE (%) Asam (H2SO40,4 M)
α-amilase, AMG
α-amilase, AMG, selulase kasar
α-amilase, AMG, selulase komersial
38,93 ± 8,09 34,93 ± 10,28 35,59 ± 11,32 36,62 ± 22,23
22,04 ± 4,31 19,50 ± 3,65 21,11 ± 1,94 26,43 ± 2,60
56,63 55,82 59,32 72,17 AMG : Amiloglukosidase komersial, Data : Rerata ± standar deviasi (n = 3)
Gambar



Dokumen terkait
H al ini scjalan
Pada tahap sebelumnya telah dilakukan fermentasi dari dekstrin dan sirup glukosa pati sagu untuk mendapatkan jenis substrat dan konsentrasi substrat yang optimum untuk
Hasil penelitian etanol dari proses fermentasi dilanjutkan destilasi dengan menggunakan substrat limbah kulit ubi kayu dan limbah kulit nanas dengan menggunakan
Peningkatan yang terjadi pada kadar protein limbah padat bioetanol singkong hasil fermentasi ini relatif kecil karena tidak dilakukan pengayaan dengan sumber nutrien lain
Hasil menunjukkan total gula terlarut dan gula reduksi pada sampel SHF1 lebih tinggi dibandingkan dengan sampel yang menggunakan SHF2.. niger mampu
Penelitian ini bertujuan untuk mengetahui pengaruh volume enzim selulase terhadap konsentrasi gula reduksi yang dihasilkan melalui proses hidrolisis serbuk kayu
Hal ini menunjukkan bahwa pada perlakuan ini, substrat yang ditambahkan hanya sedikit yang digunakan untuk pembentukan sel, namun dengan biomassa yang sedikit tersebut mampu
Perlakuan P1 dan P2 merupakan proses fermentasi dengan menggunakan kultur tunggal, dan jika dibandingkan dengan proses fermentasi menggunakan kultur campuran, terlihat bahwa
