KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA PADA
AREAL TANAMAN KARET (STUDI KASUS DI PTPN III
KEBUN BATANG TORU KABUPATEN
TAPANULI SELATAN)
TESIS
Oleh
RIZKY AMELIA DONA SIREGAR
127030010/BIO
PROGRAM PASCASARJANA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS SUMATERA UTARA
MEDAN
KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA PADA
AREAL TANAMAN KARET (STUDI KASUS DI PTPN III
KEBUN BATANG TORU KABUPATEN
TAPANULI SELATAN)
TESIS
Diajukan sebagai salah satu syarat untuk memperoleh
gelar Magister Sains dalam Program Studi Biologi pada Program Pascasarjana Fakultas MIPA Universitas Sumatera Utara
Oleh
RIZKY AMELIA DONA SIREGAR
127030010/BIO
PROGRAM PASCASARJANA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS SUMATERA UTARA
MEDAN
PERNYATAAN ORISINALITAS
KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA PADA AREAL TANAMAN KARET
(STUDI KASUS DI PTPN III KEBUN BATANG TORU KABUPATEN TAPANULI SELATAN)
TESIS
Dengan ini saya nyatakan bahwa saya mengakui semua karya tesis ini adalah hasil kerja saya sendiri kecuali kutipan dan ringkasan yang tiap satunya telah dijelaskan sumbernya dengan benar.
Medan, 15 Juli 2014
PERNYATAAN PERSETUJUAN PUBLIKASI KARYA ILMIAH UNTUK KEPENTINGAN AKADEMIS
Sebagai sivitas akademis Universitas Sumatera Utara, saya yang bertanda tangan di bawah ini:
Nama : Rizky Amelia Dona Siregar
NIM : 127030010
Program Studi : Magister Biologi Jenis Karya Ilmiah : Tesis
Demi pengembangan ilmu pengetahuan, menyetujui untuk memberikan kepada Universitas Sumatera Utara Hak Bebas Royalti Non-Eksklusif (Non-Exclusive Royalty Free Right) atas Tesis saya yang berjudul:
Keanekaragaman Fungi Mikoriza Arbuskula Pada Areal Tanaman Karet (Studi Kasus Di PTPN III Kebun Batang Toru Kabupaten Tapanuli Selatan)
Beserta perangkat yang ada (jika diperlukan). Dengan Hak Bebas Royalti Non-Eksklusif ini, Universitas Sumatera Utara berhak menyimpan, mengalih media, memformat, mengelola dalam bentuk data-base, merawat dan mempublikasikan Tesis saya tanpa meminta izin dari saya selama tetap mencantumkan nama saya sebagai penulis dan sebagai pemegang dan atau sebagai pemilik hak cipta.
Demikian pernyataan ini dibuat dengan sebenarnya.
Medan, 15 Juli 2014
Telah diuji pada Tanggal 15 Juli 2014
PANITIA PENGUJI TESIS
KETUA : Dr. Delvian, S.P., M.P
ANGGOTA : 1. Prof. Dr. Dwi Suryanto, M.Sc
2. Prof. Dr. Erman Munir, M.Sc
ABSTRACT
RIZKY AMELIA DONA SIREGAR. Vesicular Arbuscular Mycorrhizal Diversity of Rubber Plantation Area (Case Study in PTPN III Batang Toru Estate, South Tapanuli). Supervised by DELVIAN and DWI SURYANTO.
This study aims to determine the density of spores, colonization percentage, and types of vesicular arbuscular mycorrhizal in rubber tree in PTPN III Batang Toru Estate with different soil fertility conditions. Soil and root samples were taken from rhizosphere of rubber each 3 plots in afdeling 1, 2, and 4. Isolation, determine the percentage of colonization
Key words: Vesicular arbuscular mycorrhizal, diversity, rubber (Hevea brasiliensis), spore, colonization
ABSTRAK
RIZKY AMELIA DONA SIREGAR. Keanekaragaman Fungi Mikoriza Arbuskula pada Areal Tanaman Karet (Studi Kasus di PTPN III Kebun Batang Toru Kabupaten Tapanuli Selatan). Dibimbing oleh DELVIAN dan DWI SURYANTO.
Penelitian ini bertujuan untuk mengetahui kepadatan spora, persentase kolonisasi, dan tipe fungi mikoriza arbuskula (FMA) pada areal tanaman karet di PTPN III Kebun Batang Toru dengan kondisi kesuburan tanah yang berbeda. Sampel tanah dan akar diambil dari rizosfer tanaman karet masing-masing 3 plot pada afdeling 1, 2, dan 4. Variabel pengamatan yang dilakukan adalah isolasi, penghitungan kolonisasi, dan identifikasi spora FMA. Hasil penelitian menunjukkan bahwa kriteria kepadatan spora dan persentase kolonisasi FMA masing-masing tergolong tinggi dan sedang. Nilai tertinggi dari jumlah spora dan persentase kolonisasi terdapat pada afdeling 1, yaitu 249 dan 319 spora/50 g tanah (lapangan dan
trapping) untuk jumlah spora dan 42,5% untuk persentase kolonisasi. Tipe FMA yang diperoleh adalah 44 tipe Glomus dan 3 tipe Acaulospora.
Kata kunci: Fungi mikoriza arbuskula, keanekaragaman, karet (Hevea brasiliensis), spora, kolonisasi
KATA PENGANTAR
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas rahmat dan karunia-Nya sehingga penulisan tesis ini dapat diselesaikan. Dengan selesainya penulisan tesis ini, penulis mengucapkan terima kasih kepada:
1. Bapak Dr. Delvian, S.P., M.P. dan Bapak Prof. Dr. Dwi Suryanto, M.Sc. sebagai dosen pembimbing atas waktu dan perhatiannya untuk memberikan arahan dan bimbingan dalam penulisan tesis ini.
2. Bapak Prof. Dr. Erman Munir, M.Sc. dan Ibu Dr. It Jamilah, M.Sc. sebagai dosen penguji yang telah banyak memberikan kritik dan saran demi kesempurnaan tesis ini.
3. Pimpinan serta staf pegawai di Departemen Biologi FMIPA USU dan Laboratorium Biologi Tanah Fakultas Pertanian USU.
4. Manajer dan staf pegawai PTPN III Kebun Batang Toru Kabupaten Tapanuli Selatan.
5. Ayahanda dan Ibunda serta seluruh keluarga yang telah memberikan doa dan dukungan dalam penulisan tesis ini.
6. Seluruh rekan dan sahabat yang namanya tidak bisa disebutkan satu persatu. Penulis menyadari bahwa dalam penyusunan tesis ini masih terdapat berbagai kekurangan. Oleh karena itu, penulis mengharapkan kritik dan saran yang bersifat membangun agar tesis ini menjadi lebih baik. Semoga tesis ini dapat menambah wawasan atau pengetahuan bagi para pembaca. Akhir kata, penulis mengucapkan terima kasih.
RIWAYAT HIDUP
DATA PRIBADI
Nama : Rizky Amelia Dona Siregar
Tempat dan Tanggal Lahir : Padangsidimpuan, 22 April 1989
Alamat Rumah : Aspol Sitataring No. 15B Padangsidimpuan
Telpon/Hp : 081396979013
e-mail : rizkyameliadonasiregar@yahoo.co.id
DATA PENDIDIKAN
SD : Negeri 26 Padangsidimpuan Tamat : 2001 SMP : Negeri 1 Padangsidimpuan Tamat : 2004 SMA : Negeri 1 Padangsidimpuan Tamat : 2007
DAFTAR ISI
Halaman
Abstract i
Abstrak ii
Kata Pengantar iii
Riwayat Hidup iv
Daftar Isi v
Daftar Tabel vii
Daftar Gambar viii
Daftar Lampiran ix
BAB I PENDAHULUAN 1
2.1. Tanaman Karet (Hevea brasiliensis) 3
2.2. Fungi Mikoriza Arbuskula (FMA) 3
2.2.1. Anatomi dan Morfologi FMA 5
2.2.2. Simbiosis FMA pada Akar 5
2.2.3. Faktor yang Mempengaruhi Perkembangan FMA 6
2.2.4. Hasil Penelitian Keanekaragaman FMA 8
BAB III METODE PENELITIAN 10
3.1. Waktu dan Tempat 10
3.2. Bahan dan Alat 10
3.3. Pengambilan Sampel Tanah dan Akar 10
3.4. Pengamatan Sampel Tanah dan Akar 11
3.4.1. Ekstraksi Spora dan Identifikasi FMA 11
3.4.2. Kolonisasi FMA pada Akar Tanaman 12
3.5. Pemerangkapan (Trapping) 13
BAB IV HASIL DAN PEMBAHASAN 14
4.1. Sifat Kimia Tanah 14
4.2. Kepadatan Spora FMA 15
4.3. Persentase Kolonisasi FMA 18
BAB V KESIMPULAN DAN SARAN 30
5.1. Kesimpulan 30
5.2. Saran 30
DAFTAR PUSTAKA 31
DAFTAR TABEL
Halaman
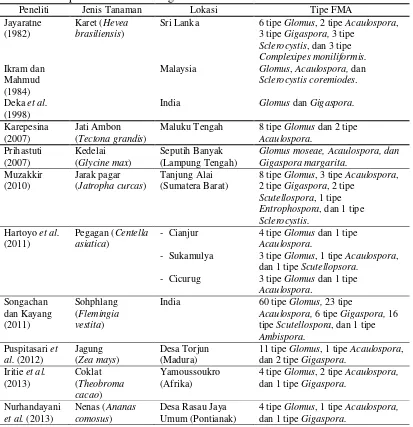
Tabel 1. Hasil penelitian keanekaragaman FMA 9
Tabel 2. Hasil analisis tanah 14
Tabel 3. Tipe dan karakteristik spora FMA lapangan dan trapping 21
DAFTAR GAMBAR
Halaman
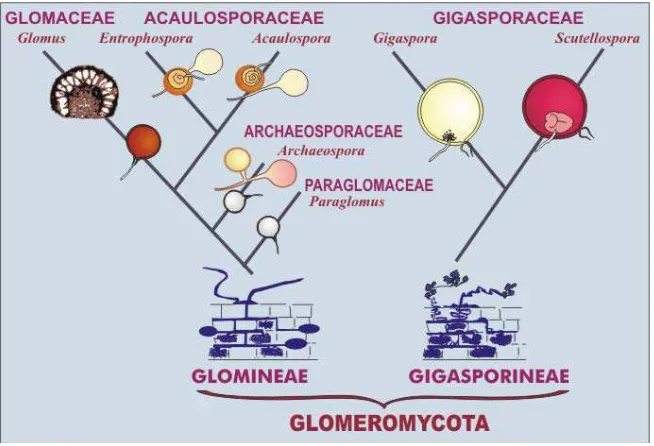
Gambar 1. Klasifikasi FMA 4
Gambar 2. Ilustrasi petak contoh pengambilan sampel
tanah dan akar 11
Gambar 3. Kepadatan spora FMA dari lapangan dan trapping 15 Gambar 4. Kolonisasi akar oleh FMA yang ditandai dengan adanya (a) hifa
dan (b) vesikula 18
Gambar 5. Persentase kolonisasi FMA 18
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Hasil analisis tanah 36
ABSTRACT
RIZKY AMELIA DONA SIREGAR. Vesicular Arbuscular Mycorrhizal Diversity of Rubber Plantation Area (Case Study in PTPN III Batang Toru Estate, South Tapanuli). Supervised by DELVIAN and DWI SURYANTO.
This study aims to determine the density of spores, colonization percentage, and types of vesicular arbuscular mycorrhizal in rubber tree in PTPN III Batang Toru Estate with different soil fertility conditions. Soil and root samples were taken from rhizosphere of rubber each 3 plots in afdeling 1, 2, and 4. Isolation, determine the percentage of colonization
Key words: Vesicular arbuscular mycorrhizal, diversity, rubber (Hevea brasiliensis), spore, colonization
ABSTRAK
RIZKY AMELIA DONA SIREGAR. Keanekaragaman Fungi Mikoriza Arbuskula pada Areal Tanaman Karet (Studi Kasus di PTPN III Kebun Batang Toru Kabupaten Tapanuli Selatan). Dibimbing oleh DELVIAN dan DWI SURYANTO.
Penelitian ini bertujuan untuk mengetahui kepadatan spora, persentase kolonisasi, dan tipe fungi mikoriza arbuskula (FMA) pada areal tanaman karet di PTPN III Kebun Batang Toru dengan kondisi kesuburan tanah yang berbeda. Sampel tanah dan akar diambil dari rizosfer tanaman karet masing-masing 3 plot pada afdeling 1, 2, dan 4. Variabel pengamatan yang dilakukan adalah isolasi, penghitungan kolonisasi, dan identifikasi spora FMA. Hasil penelitian menunjukkan bahwa kriteria kepadatan spora dan persentase kolonisasi FMA masing-masing tergolong tinggi dan sedang. Nilai tertinggi dari jumlah spora dan persentase kolonisasi terdapat pada afdeling 1, yaitu 249 dan 319 spora/50 g tanah (lapangan dan
trapping) untuk jumlah spora dan 42,5% untuk persentase kolonisasi. Tipe FMA yang diperoleh adalah 44 tipe Glomus dan 3 tipe Acaulospora.
Kata kunci: Fungi mikoriza arbuskula, keanekaragaman, karet (Hevea brasiliensis), spora, kolonisasi
BAB I PENDAHULUAN
1.1. Latar Belakang
Fungi mikoriza arbuskula (FMA) merupakan mikroorganisme tanah yang terdapat di rizosfer dan membentuk simbiosis mutualisme dengan tanaman. Fungi mikoriza arbuskula akan memperoleh unsur karbon dari tanaman sedangkan tanaman akan memperoleh unsur hara terutama fosfor (P) dari FMA (Smith dan Read, 2008). Dengan demikian, kemampuan FMA untuk menyediakan unsur hara bagi tanaman dapat meningkatkan pertumbuhan tanaman.
Fungi mikoriza arbuskula telah diketahui berperan dalam merangsang pertumbuhan tanaman khususnya pada tanah yang kurang subur. Peningkatan pertumbuhan tanaman oleh FMA terutama disebabkan oleh penyerapan fosfor. Selain itu, FMA juga dapat meningkatkan penyerapan air, melindungi tanaman inang dari patogen tanah, dan meningkatkan stabilitas tanah (Smith dan Read, 2008).
Fungi mikoriza arbuskula memiliki sifat kosmopolit (tersebar di seluruh dunia) dan dapat menginfeksi hampir semua jenis tanaman. Penyebaran FMA berlangsung secara aktif pada miselium di dalam tanah (Suhardi, 1988). Banyaknya jenis tanaman yang dapat diinfeksi FMA menunjukkan bahwa FMA tidak memiliki spesifikasi tanaman inang. Oleh karena itu, FMA banyak mendapat perhatian dan sering diteliti terutama untuk mempelajari potensinya.
Pemanfaatan FMA pada tanaman karet akan lebih efektif jika diperoleh isolat FMA yang potensial dan spesifik. Hal pertama yang harus diketahui untuk mempelajari potensi suatu organisme adalah keberadaan dan keberagaman dari organisme tersebut. Demikian juga halnya dengan studi keanekaragaman FMA di areal tanaman karet. Dengan adanya data tentang keanekaragaman FMA di areal tanaman karet dapat dilakukan seleksi untuk memperoleh isolat FMA yang potensial dan spesifik pada tanaman karet.
1.2. Perumusan Masalah
Keanekaragaman dan penyebaran FMA sangat bervariasi karena kondisi lingkungannya yang juga berbeda. Pada areal tanaman karet, informasi tentang keanekaragaman FMA sejauh ini masih terbatas. Mansur et al. (2002) menyatakan bahwa hampir 70% kegiatan penelitian FMA diarahkan pada manfaatnya dalam pertumbuhan tanaman dan kurang dari 15% yang mempelajari keanekaragaman FMA pada suatu ekosistem. Eksplorasi tipe-tipe FMA di areal tanaman karet merupakan langkah awal yang penting untuk dapat mengidentifikasi tipe-tipe FMA dominan dan spesifik yang ada. Kegiatan ini sangat penting untuk mendapatkan informasi keanekaragaman tipe-tipe FMA sehingga diperoleh isolat FMA yang potensial. Berdasarkan pernyataan di atas, maka perlu dilakukan penelitian mengenai keanekaragaman FMA pada areal tanaman karet di PTPN III Kebun Batang Toru.
1.3. Tujuan Penelitian
Tujuan penelitian ini adalah untuk mengetahui kepadatan spora, persentase kolonisasi, dan tipe spora FMA pada areal tanaman karet di PTPN III Kebun Batang Toru dengan kondisi kesuburan tanah yang berbeda.
1.4. Manfaat Penelitian
BAB II
TINJAUAN PUSTAKA
2.1. Tanaman Karet (Hevea brasiliensis)
Tanaman karet (Hevea brasiliensis) tergolong jenis tanaman tahunan yang
berasal dari fam
perkebunan yang mampu menciptakan lingkungan sehat karena dapat berfungsi sebagai sumber oksigen serta memiliki nilai ekonomi yang tinggi karena merupakan penghasil lateks maupun kayu.
Tanaman karet tumbuh baik pada daerah tropis, zona 150 LU dan 150 LS, suhu 25-300 C, ketinggian 0-400 m di atas permukaan laut, curah hujan minimal 1500 mm/tahun, dan penyinaran matahari 5-7 jam. Perakaran tanaman karet tersusun atas akar tunggang, akar lateral, dan akar baru. Perkembangan akar dipengaruhi oleh energi yang tersedia dalam jaringan tanaman dan keadaan tanah di lingkungan akar tanaman (kesuburan tanah). Tanah yang cocok untuk tanaman karet adalah aerase dan drainase baik, remah, porus, dapat menahan air, tekstur terdiri atas 35% liat dan 30% pasir, tidak bergambut, kandungan unsur hara (N,P, dan K) cukup, dan pH 4,5-6,5 (Verheye, 2010). Kondisi tanah seperti ini dapat meningkatkan produksi tanaman karet. Oleh karena itu, faktor kesuburan tanah sangat penting untuk pertumbuhan dan produktivitas tanaman.
2.2. Fungi Mikoriza Arbuskula (FMA)
Mikoriza terdiri dari dua kata yang berasal dari bahasa Yunani, yaitu myces
Mikoriza diklasifikasikan menjadi dua subdivisi besar, yaitu ektomikoriza dan endomikoriza. Fungi mikoriza arbuskula (FMA) digolongkan dalam kelompok endomikoriza dengan ciri-ciri: 1) akar yang kena infeksi tidak membesar, 2) lapisan hifa pada permukaan akar tipis, 3) hifa masuk ke dalam individu sel jaringan korteks, dan 4) mempunyai struktur vesikula serta arbuskula. Fungi ini bersifat obligat dan telah dilaporkan dapat bersimbiosis dengan hampir 90% jenis tanaman (Smith dan Read, 2008).
Fungi mikoriza arbuskula termasuk dalam filum Glomeromycota, kelas
Zygomycetes, dan ordo Glomales. Ordo ini terdiri atas 2 sub-ordo, yaitu
Gigasporineae dan Glomineae (INVAM, 2013). Klasifikasi FMA secara lengkap ditunjukkan pada Gambar 1.
2.2.1. Anatomi dan Morfologi FMA
Anatomi FMA dibentuk oleh beberapa struktur sehingga dapat bertahan, tumbuh, dan berkembangbiak pada akar tanaman inang. Struktur tersebut adalah hifa, arbuskula (struktur hifa bercabang-cabang), vesikula (struktur lonjong atau bulat yang mengandung cairan lemak), sel auksilari (hifa pelengkap), dan spora. Spora memiliki klamidospora yang akan terbentuk jika FMA terpisah dengan tanaman inangnya (INVAM, 2013).
Fungi mikoriza arbuskula dapat diidentifikasi secara morfologis dengan melakukan observasi terlebih dahulu terhadap FMA tunggal yang diisolasi dari sampel tanah. Meskipun FMA pada tingkat spesies tidak dapat menggunakan kriteria morfologis karena memiliki morfologi yang hampir sama, namun beberapa spesies memiliki perbedaan dari morfologi vesikel, diameter hifa, dan pola pertumbuhan akar (Abbott, 1982). Dengan demikian, pendekatan morfologis tetap bisa dilakukan dengan tujuan menilai keberhasilan inokulasi di tanah.
2.2.2. Simbiosis FMA pada Akar
Simbiosis FMA berawal dari pergerakan hifa ekstraradikal yang berasal dari perkecambahan spora dalam tanah atau akar terkolonisasi. Hal ini terjadi karena tanaman mengeksudasikan senyawa flavonoid. Selanjutnya, hifa ekstraradikal menyentuh permukaan akar, membentuk appresoria, dan menembus dinding sel akar untuk membentuk hifa intraradikal. Hifa ini tumbuh menjalar di antara sel atau menembus sel epidermis dan akhirnya mengkolonisasi ruang intra dan interseluler korteks akar. Setelah itu, hifa intraradikal berdiferensiasi membentuk arbuskula, vesikula, sel auksilari, dan spora intraradikal.
untuk akar tanaman. Selain itu, hifa ekstraradikal juga mampu menembus pori mikro untuk mendapatkan air yang tidak dapat dijangkau oleh akar karena garis tengahnya yang jauh lebih kecil dibandingkan dengan garis tengah akar. Penyerapan unsur P dan air oleh FMA dipengaruhi oleh jenis FMA, tanaman, dan lingkungan. Hal ini mengindikasikan bahwa kesesuaian fungsional antara FMA dan tanaman tidak selalu berkaitan dengan kolonisasinya (Smith dan Read, 2008).
2.2.3. Faktor yang Mempengaruhi Perkembangan FMA
Faktor-faktor yang mempengaruhi perkembangan FMA adalah sebagai berikut:
a. Suhu
Suhu yang relatif tinggi akan meningkatkan aktivitas FMA sehingga berpengaruh terhadap pertumbuhan tanaman inang. Persentase kolonisasi meningkat pada suhu 300 C, tetapi beberapa simbiosis FMA dengan tanaman berkembang secara normal pada suhu 350 C atau lebih (Smith dan Read, 1997). Aktivitas FMA hanya menurun pada suhu di atas 400
b. Kadar air tanah
C (Mosse, 1981).
Keberadaan FMA dapat menguntungkan tanaman yang tumbuh di daerah kering. Hal ini disebabkan: 1) FMA dapat menurunkan gerakan air sehingga transfer air ke akar meningkat, 2) FMA meningkatkan kadar P tanaman sehingga daya tahan terhadap kekeringan juga meningkat, dan 3) adanya hifa eksternal FMA yang dapat menyerap air dari areal yang lebih jauh (Rothwell, 1984). Penelitian Daniels dan Trappe (1980) menunjukkan bahwa perkecambahan maksimum Glomus epigaeus
c. Derajat keasaman (pH) tanah
Fungi mikoriza arbuskula pada umumnya lebih tahan terhadap perubahan pH tanah. Meskipun demikian, daya adaptasi masing-masing spesies FMA terhadap pH tanah berbeda-beda. Hal ini disebabkan pH tanah mempengaruhi perkecambahan, perkembangan, dan peran FMA terhadap pertumbuhan tanaman. Menurut Daniels dan Trappe (1980), perkecambahan maksimum Glomus epigaeus terjadi pada pH 6-8. Sementara itu, pada spesies yang berbeda Glomus fasciculatus dapat berkembang dengan baik pada tanah asam (Mosse, 1981).
d. Bahan organik
Jumlah maksimum spora ditemukan pada tanah-tanah yang mengandung bahan organik 1-2% dan kandungan spora sangat rendah pada tanah-tanah berbahan organik kurang dari 0,5%. Residu akar mempengaruhi ekologi FMA karena serasah akar yang terkolonisasi FMA merupakan sarana penting untuk mempertahankan generasi FMA dari satu tanaman ke tanaman berikutnya. Serasah akar tersebut mengandung hifa, vesikel, dan spora yang dapat mengkolonisasi FMA (Whiffen, 2007).
e. Cahaya
Cahaya berpengaruh terhadap proses fotosintesis pada tanaman. Fotosintesis yang rendah menyebabkan berkurangnya jumlah karbohidrat di dalam akar sehingga mengurangi persentase kolonisasi FMA. Sebaliknya, kolonisasi meningkat pada intensitas cahaya yang lebih tinggi (Gianinazzi-Pearson dan Gianinazzi, 1983).
f. Ketersediaan hara
dapat meningkatkan derajat infeksi FMA, memperbaiki kesuburan tanah, dan meningkatkan hasil tanaman (Smith dan Read, 1997).
g. Logam berat dan unsur lain
Beberapa spesies FMA diketahui mampu beradaptasi dengan tanah yang tercemar seng (Zn). Infeksi FMA lebih tinggi pada tanah yang mengalami kekahatan Mn daripada yang tidak kahat Mn (Mosse, 1981). Beberapa penelitian lain juga diketahui bahwa FMA tertentu toleran terhadap kandungan Al dan Na yang tinggi (Janoukova et al., 2006).
h. Fungisida
Fungisida merupakan racun kimia yang digunakan untuk membunuh jamur penyebab penyakit pada tanaman. Namun, penggunaan fungisida juga berdampak buruk terhadap FMA. Sukarno et al. (1993) melaporkan bahwa fungisida Benlate dan Ridomil dapat mengurangi jumlah hifa antarsel dan arbuskula. Schreiner dan Bethlenfalvay (1996) menambahkan bahwa aplikasi fungisida seperti Benomyl, PCNB, dan Captan menurunkan persentase kolonisasi akar oleh FMA bila dibandingkan dengan tanpa fungisida.
2.2.4. Hasil Penelitian Keanekaragaman FMA
Tabel 1. Hasil penelitian keanekaragaman FMA
Peneliti Jenis Tanaman Lokasi Tipe FMA Jayaratne
Sclerocystis, dan 3 tipe
Complexipes moniliformis.
Glomus, Acaulospora, dan
Sclerocystis coremiodes.
Glomus dan Gigaspora.
Karepesina
Glomus moseae, Acaulospora, dan Gigaspora margarita.
Scutellospora, 1 tipe
Entrophospora, dan 1 tipe
Sclerocystis. dan 1 tipe Scutellopsora.
3 tipe Glomus dan 1 tipe
Acaulospora, 6 tipe Gigaspora, 16 tipe Scutellospora, dan 1 tipe
Ambispora. dan 2 tipe Gigaspora.
Iritie et al.
dan 1 tipe Gigaspora.
Nurhandayani
BAB III
METODELOGI PENELITIAN
3.1. Waktu dan Tempat
Pengambilan sampel tanah dan akar tanaman karet dilakukan di Perkebunan Karet PTPN III Kebun Batang Toru pada bulan September 2013. Ekstraksi spora, identifikasi, dan penghitungan kolonisasi FMA dilakukan di Laboratorium Biologi Tanah Universitas Sumatera Utara pada bulan September-Desember 2013.
3.2. Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah sampel tanah dan akar tanaman karet; larutan glukosa 60%; larutan Melzer’s; polyvinyl alcohol lactoglycerol
(PVLG); KOH 2,5%; HCL 2%; trypan blue 0,05%; chlorox 5,25%; hyponex merah (25-5-20); dan benih Zea mays. Alat yang digunakan adalah saringan bertingkat dengan ukuran 250, 125, dan 53 µm serta pinset spora.
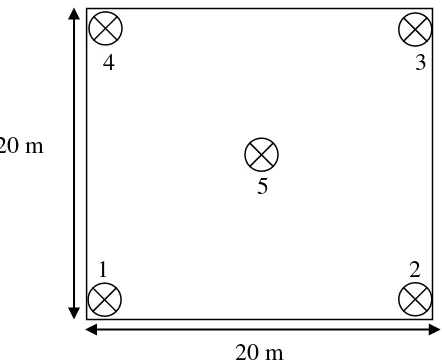
3.3. Pengambilan Sampel Tanah dan Akar
4 3
20 m
5
1 2
20 m
Gambar 2. Ilustrasi petak contoh pengambilan sampel tanah dan akar
3.4. Pengamatan Sampel Tanah dan Akar
Dalam penelitian ini dilakukan identifikasi tipe FMA dan persentase kolonisasi FMA.
3.4.1. Ekstraksi Spora dan Identifikasi FMA
Ekstraksi spora FMA dilakukan untuk memisahkan spora FMA dari sampel tanah sehingga dapat dilakukan identifikasi FMA guna mengetahui jumlah dan tipe spora FMA yang terdapat pada setiap plot. Teknik yang digunakan dalam ekstraksi spora FMA adalah teknik tuang saring yang dilanjutkan dengan teknik sentrifugasi (Brundrett et al., 1996).
yang tertinggal pada saringan paling bawah dipindahkan ke dalam tabung sentrifus. Kemudian hasil saringan tadi ditambahkan dengan larutan glukosa 60% sebanyak 3 mL. Tabung sentrifus ditutup rapat dan disentrifugasi dengan kecepatan 2500 rpm selama 3 menit. Cairan supernatan yang diperoleh dituang ke dalam saringan 53 µm, dicuci dengan air mengalir, dipindahkan ke cawan petri, dan diperiksa di bawah mikroskop untuk penghitungan kepadatan spora serta pembuatan preparat guna identifikasi spora FMA yang ada.
Pembuatan preparat spora menggunakan larutan Melzer’s dan PVLG yang diletakkan secara terpisah pada satu object glass. Spora-spora FMA yang diperoleh dari ekstraksi dihitung jumlahnya, lalu diletakkan dalam larutan Melzer’s dan PVLG. Selanjutnya spora-spora tersebut ditutup dengan cover glass dan diamati dengan mikroskop. Perubahan warna spora dalam larutan Melzer’s adalah salah satu indikator untuk menetukan tipe spora yang ada.
3.4.2. Kolonisasi FMA pada Akar Tanaman
Kolonisasi FMA pada akar tanaman karet ditandai dengan adanya hifa, arbuskula, atau vesikula ketika diamati dengan mikroskop. Tahap ini dilakukan melalui teknik pewarnaan akar (root staining). Langkah pertama adalah memilih sampel akar yang halus dan dicuci dengan air mengalir. Sampel akar yang telah bersih kemudian direndam dalam larutan KOH 2,5% selama 7 hari untuk mengeluarkan seluruh isi sitoplasma dari sel akar. Selanjutnya sampel akar dicuci kembali dengan air mengalir dan direndam dalam larutan HCL 2% selama 2 malam. Setelah itu, sampel akar langsung direndam dalam larutan trypan blue 0,05% selama 24 jam untuk mempermudah pengamatan (Kormanik dan McGraw, 1982).
(+), sedangkan yang tidak menunjukkan kolonisasi diberi tanda negatif (-). Persentase kolonisasi akar oleh FMA dihitung dengan rumus berikut:
% kolonisasi akar = ∑ bidangpandangbertanda (+)
∑ bidangpandangkeseluruhan × 100%
3.5. Pemerangkapan (Trapping)
Tujuan pemerangkapan (trapping) adalah mendapatkan keanekaragaman spora FMA. Langkah pertama adalah mempersiapkan benih jagung (Zea mays) sebagai tanaman inang dan pasir sebagai media tanamnya. Benih jagung terlebih dahulu direndam dalam larutan chlorox 5,25% selama 5 menit, lalu diganti dengan air selama 5 menit. Benih-benih jagung kemudian disemaikan dalam pasir yang telah disterilisasi hingga muncul dua helai daun dan dilanjutkan dengan trapping.
BAB IV
HASIL DAN PEMBAHASAN
4.1. Sifat Kimia Tanah
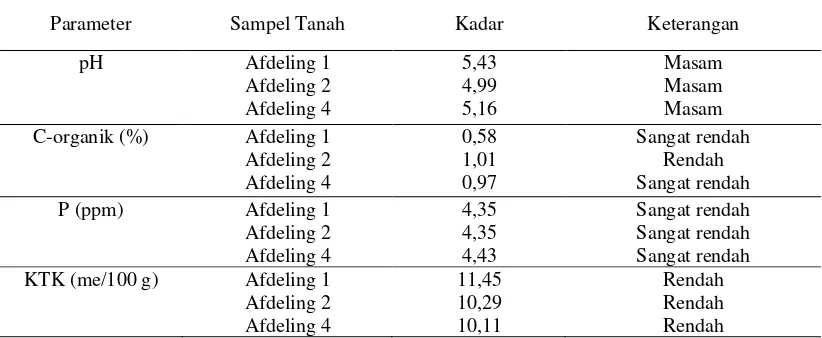
Sifat kimia tanah yang dijadikan sebagai sampel isolasi spora FMA dapat diketahui dengan menganalisis beberapa parameternya, yaitu pH, C-organik, fosfor (P), dan kapasitas tukar kation (KTK). Hasil analisis tanah ditunjukkan pada Tabel 2. Tabel 2. Hasil analisis tanah
Parameter Sampel Tanah Kadar Keterangan
pH Afdeling 1
Berdasarkan hasil analisis tanah, tingkat kesuburan tanah pada afdeling 1, 2, dan 4 tergolong rendah (kurang subur). Kadar pH tanah yang diperoleh menunjukkan bahwa kondisi tanah dalam penelitian ini tergolong masam. Keberadaan FMA pada kondisi tanah seperti ini menunjukkan bahwa FMA mampu beradaptasi dengan kadar pH yang masam. Kemasaman tanah dapat dipengaruhi oleh bahan organik tanah termasuk C-organik. Menurut Utami dan Handayani (2003), kadar C-organik yang rendah menunjukkan kecenderungan pH lebih rendah.
juga rendah. Kapasitas tukar kation merupakan salah satu indikator kesuburan tanah yang dipengaruhi oleh kadar C-organik dalam tanah. Kadar KTK yang rendah menunjukkan rendahnya tingkat kesuburan tanah. Dengan demikian, sifat kimia tanah yang sama di setiap afdeling menunjukkan bahwa kondisi tanahnya juga sama. Kondisi inilah yang dapat mempengaruhi adanya beberapa hasil penelitian yang sama di setiap afdeling.
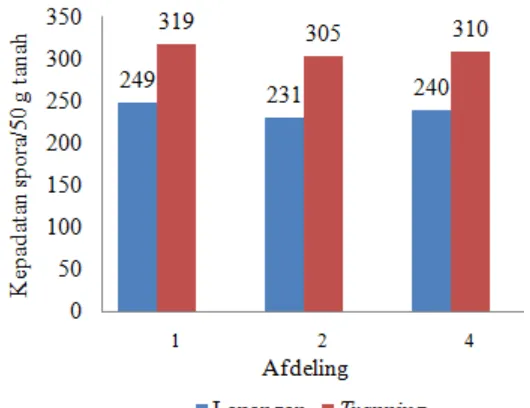
4.2. Kepadatan Spora FMA
Kepadatan spora FMA yang diperoleh di setiap afdeling dari hasil lapangan dan trapping ditunjukkan pada Gambar 3. Jumlah spora tertinggi diperoleh pada afdeling 1, yaitu 249 dan 319 spora/50 g tanah masing-masing dari hasil lapangan dan
trapping. Secara keseluruhan, jumlah spora hasil trapping lebih tinggi dibandingkan lapangan.
Kepadatan spora yang diperoleh pada setiap afdeling dari hasil lapangan dan
trapping masing-masing tinggi dan sangat tinggi (Smith dan Read, 1997). Tingkat kepadatan spora yang sama di setiap afdeling baik lapangan maupun trapping
disebabkan oleh adanya sifat kimia tanah yang sama sehingga menciptakan kondisi lingkungan yang sama di setiap afdeling. Penelitian lain yang juga memperoleh kepadatan spora yang tinggi pada areal tanaman karet, yaitu Jayaratne dan Waidyanatha dalam Akond et al. (2008) dan Souza et al. (2010) masing-masing memperoleh 200-2000 dan 222 spora/50 g tanah. Hal ini menunjukkan bahwa pada lokasi yang berbeda mungkin terdapat sifat kimia tanah yang sama sehingga tingkat kepadatan spora yang diperoleh juga sama. Dengan demikian, sifat kimia tanah yang sama pada afdeling 1, 2, dan 4 merupakan faktor yang menyebabkan tingkat kepadatan spora yang diperoleh pada masing-masing afdeling juga sama.
Jumlah spora hasil trapping di setiap afdeling menunjukkan nilai yang tidak begitu berbeda. Hal ini disebabkan oleh kondisi lingkungan yang sama baik lokasi maupun perlakuan. Jika dibandingkan, jumlah spora hasil trapping lebih tinggi dibandingkan lapangan. Hal ini disebabkan adanya perlakuan stressing (cekaman air) dengan tidak melakukan penyiraman. Perlakuan ini mengakibatkan kondisi perakaran menjadi kering sehingga memacu FMA untuk membentuk spora dalam jumlah yang banyak. Pacioni (1986) menyatakan bahwa pembentukan spora FMA dirangsang oleh kondisi cekaman air. Brundrett (2006) menambahkan bahwa dalam kondisi yang tidak menguntungkan keberadaan mikoriza dapat diamati, yaitu dalam bentuk spora. Hal ini berarti spora banyak diperoleh pada kondisi lingkungan yang tidak optimal seperti perlakuan stressing. Oleh karena itu, jumlah spora hasil trapping lebih tinggi dibandingkan lapangan. Hasil serupa juga diperoleh Iritie et al. (2013), yaitu jumlah spora dari hasil trapping mencapai 4.078 spora/50 g tanah yang sebelumnya hanya berjumlah 324 spora/50 g tanah dari hasil lapangan. Penelitian lainnya juga menunjukkan bahwa jumlah spora hasil trapping lebih tinggi dibandingkan lapangan (Karepesina, 2007 dan Hartoyo et al., 2011).
4.3. Persentase Kolonisasi FMA
Kolonisasi akar tanaman karet oleh FMA dalam penelitian ini ditandai dengan adanya hifa dan atau vesikula (Gambar 4). Persentase kolonisasi yang tertinggi diperoleh pada afdeling 1, yaitu 42,5% (Gambar 5).
(a) (b)
Hifa Vesikula Gambar 4. Kolonisasi akar oleh FMA yang ditandai dengan adanya (a) hifa dan (b) vesikula
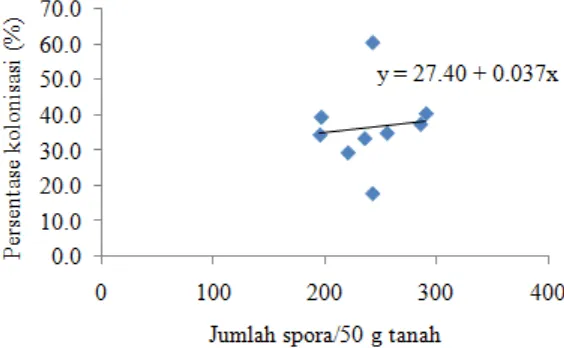
Persentase kolonisasi yang diperoleh pada afdeling 1, 2, dan 4 tergolong sama, yaitu sedang. Menurut Setiadi et al. (1992), persentase kolonisasi tergolong sedang jika berada di antara 26-50%. Ikram dan Mahmud (1984) menyatakan bahwa persentase kolonisasi FMA di areal tanaman karet tidak lebih dari 50%. Persentase kolonisasi yang diperoleh dalam penelitian ini tampaknya sejalan atau berkorelasi positif dengan kepadatan spora (Gambar 6).
Gambar 6. Korelasi persentase kolonisasi dengan kepadatan spora FMA (×) Jumlah spora dan (y) persentase kolonisasi
Berdasarkan gambar di atas tampak bahwa persentase kolonisasi FMA tergantung pada jumlah spora. Persentase kolonisasi akan meningkat 0,037% setiap penambahan 1 spora. Dengan demikian, peningkatan persentase kolonisasi FMA dipengaruhi oleh peningkatan jumlah spora. Hasil serupa yang menunjukkan bahwa persentase kolonisasi sejalan dengan kepadatan spora juga diperoleh dari penelitian lain (Delvian, 2010; Songachan dan Kayang, 2011; Khakpour dan Khara, 2012; Aytok et al., 2013).
penyerapan P dalam bentuk tidak tersedia. Songachan dan Kayang (2011) menunjukkan hasil yang sama dengan penelitian ini, yaitu persentase kolonisasi 30,11%; 43,77% dan 35,21%; 29,51% (sedang) masing-masing diperoleh pada kadar P 0,15% dan 0,47% (sangat rendah).
Selain P, eksudat akar juga mempengaruhi kolonisasi FMA secara tidak langsung. Bonfante dan Genre (2010) menyatakan bahwa eksudat yang dilepaskan oleh akar tanaman dapat menginduksi percabangan hifa sebagai faktor utama untuk meningkatkan kemungkinan kolonisasi. Faktor lain yang juga mempengaruhi kolonisasi adalah curah hujan. Pada saat pengambilan sampel, curah hujan bulanan 269 mm yang berarti tergolong sedang karena berada di antara 101-300 mm (BMKG, 2013). Hal ini menunjukkan adanya serapan air ke dalam tanah yang cukup untuk perkecambahan spora sehingga meningkatkan produksi hifa yang akan mengkolonisasi akar.
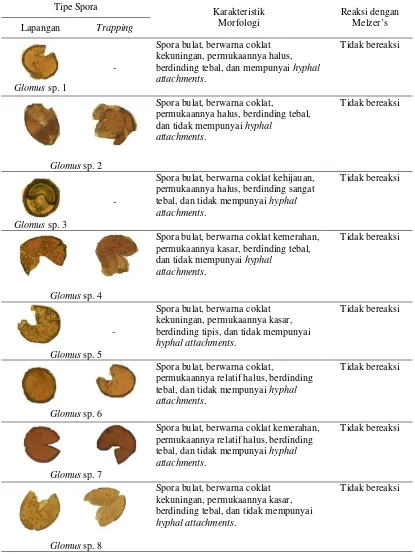
4.4. Tipe dan Karakteristik Spora FMA
Tabel 3. Tipe dan karakteristik spora FMA dari lapangan dan trapping Tipe Spora Karakteristik
Morfologi
Spora bulat, berwarna coklat kekuningan, permukaannya halus, berdinding tebal, dan mempunyai hyphal attachments.
Tidak bereaksi
Glomus sp. 2
Spora bulat, berwarna coklat,
permukaannya halus, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
Glomus sp. 3
-
Spora bulat, berwarna coklat kehijauan, permukaannya halus, berdinding sangat tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
Glomus sp. 4
Spora bulat, berwarna coklat kemerahan, permukaannya kasar, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 5
Spora bulat, berwarna coklat kekuningan, permukaannya kasar, berdinding tipis, dan tidak mempunyai
hyphal attachments.
Tidak bereaksi
Glomus sp. 6
Spora bulat, berwarna coklat,
permukaannya relatif halus, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
Glomus sp. 7
Spora bulat, berwarna coklat kemerahan, permukaannya relatif halus, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
Glomus sp. 8
Spora bulat, berwarna coklat kekuningan, permukaannya kasar, berdinding tebal, dan tidak mempunyai
hyphal attachments.
Lanjutan
Glomus sp. 9
Spora bulat, berwarna coklat
kekuningan, permukaannya relatif halus, berdinding tipis, dan tidak mempunyai
hyphal attachments.
Tidak bereaksi
Glomus sp. 10
Spora bulat, berwarna coklat
kekuningan, permukaannya relatif halus, berdinding tebal, dan tidak mempunyai
hyphal attachments.
Tidak bereaksi
Glomus sp. 11
Spora bulat, berwarna coklat kekuningan, permukaannya halus, berdinding sangat tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
Glomus sp. 12
-
Spora bulat, berwarna coklat,
permukaannya relatif halus, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
Glomus sp. 13
-
Spora bulat, berwarna coklat kemerahan, permukaannya relatif kasar, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
Glomus sp. 14
Spora bulat, berwarna coklat
kekuningan, permukaannya relatif halus, berdinding tipis, dan tidak mempunyai
hyphal attachments.
Tidak bereaksi
Glomus sp. 15
-
Spora bulat lonjong, berwarna coklat kekuningan, permukaannya halus, berdinding tebal, dan tidak mempunyai
hyphal attachments.
Tidak bereaksi
Glomus sp. 16
-
Spora bulat, berwarna coklat kehijauan, permukaannya sangat halus, berdinding tebal, dan tidak mempunyai hyphal attachments.
Lanjutan
Glomus sp. 17
-
Spora bulat, berwarna coklat kehijauan, permukaannya relatif halus, berdinding sangat tebal, dan mempunyai hyphal attachments.
Tidak bereaksi
Glomus sp. 18
-
Spora bulat, berwarna coklat,
permukaannya halus, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 19
Spora bulat, berwarna coklat kehitaman, permukaannya halus, berdinding tebal, dantidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 20
Spora bulat, berwarna coklat kehijauan, permukaannya kasar, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 21
Spora bulat, berwarna coklat,
permukaannya halus, berdinding tipis, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 22
Spora bulat, berwarna coklat kemerahan, permukaannya relatif halus, berdinding tipis, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Glomus sp. 23
Spora bulat, berwarna coklat kehitaman, permukaannya relatif halus, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Glomus sp. 24
Spora bulat, berwarna coklat,
permukaannya halus, berdinding tebal, tidak mempunyai hyphal attachments, dan mempunyai tonjolan.
Lanjutan
-
Glomus sp. 25
Spora bulat lonjong, berwarna coklat, permukaannya halus, berdinding tipis, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 26
Spora bulat lonjong, berwarna coklat, permukaannya halus, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 27
Spora bulat, berwarna coklat kemerahan, permukaannya relatif halus, berdinding tipis, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Glomus sp. 28
Spora bulat, berwarna coklat
kekuningan, permukaannya relatif halus, berdinding tipis, dan tidak mempunyai
hyphal attachments.
Tidak bereaksi
-
Glomus sp. 29
Spora bulat, berwarna coklat kekuningan, permukaannya halus, berdinding sangat tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Glomus sp. 30
Spora bulat, berwarna coklat kemerahan, permukaannya relatif halus, berdinding sangat tebal, dan tidak mempunyai
hyphal attachments.
Tidak bereaksi
-
Glomus sp. 31
Spora bulat, berwarna coklat kehijauan, permukaannya halus, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 32
Spora bulat, berwarna coklat kehijauan, permukaannya relatif halus, berdinding tebal, dan mempunyai hyphal
attachments.
Lanjutan
-
Glomus sp. 33
Spora bulat, berwarna coklat kehitaman, permukaannya halus, berdinding tipis, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 34
Spora bulat, berwarna coklat,
permukaannya halus, berdinding tipis, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 35
Spora bulat, berwarna coklat pekat, permukaannya relatif kasar, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Glomus sp. 36
Spora bulat, berwarna coklat kekuningan, permukaannya kasar, berdinding sangat tebal, dan mempunyai
hyphal attachments.
Tidak bereaksi
-
Glomus sp. 37
Spora bulat, berwarna coklat kehitaman, permukaannya kasar, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 38
Spora bulat, berwarna coklat kekuningan, permukaannya kasar, berdinding tebal (mengkilat), dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Glomus sp. 39
Spora bulat, berwarna coklat kehijauan, permukaannya relatif halus, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Glomus sp. 40
Spora bulat, berwarna orange,
permukaannya kasar, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Lanjutan
-
Glomus sp. 41
Spora bulat, berwarna coklat,
permukaannya relatif kasar, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Glomus sp. 42
Spora bulat, berwarna coklat susu, permukaannya halus, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 43
Spora bulat, berwarna coklat kehijauan, permukaannya halus, berdinding tebal, dan tidak mempunyai hyphal
attachments.
Tidak bereaksi
-
Glomus sp. 44
Spora bulat, berwarna coklat kehijauan, permukaannya relatif kasar, berdinding tebal, dan tidak mempunyai hyphal attachments.
Tidak bereaksi
-
Acaulospora sp. 1
Spora bulat, berwarna coklat, permukaannya relatif halus, dan berdinding tebal (warna kontras).
Bereaksi
-
Acaulospora sp.2
Spora bulat, berwarna kuning, dan berdinding tebal. Permukaannya relatif kasar dan membentuk ornamen seperti kulit jeruk
Bereaksi
-
Acaulospora sp. 3
Spora bulat, berwarna coklat kemerahan, permukaannya relatif halus, dan
berdinding tebal (warna kontras).
Tipe spora yang diperoleh dari hasil trapping lebih beragam dibandingkan lapangan. Hal ini diduga adanya pengaruh faktor jumlah spora yang sebelumnya memang lebih sedikit ditemukan dari lapangan dibandingkan trapping. Jumlah spora yang lebih banyak pada saat isolasi akan memungkinkan kita untuk mendapatkan tipe maupun genus baru pada saat identifikasi. Hasil penelitian ini menunjukkan bahwa banyak tipe baru dari Glomus yang muncul setelah dilakukan trapping, bahkan ada satu genus baru yang ditemukan, yaitu Acaulospora.
Secara keseluruhan, jumlah tipe spora FMA yang dihasilkan dalam penelitian ini adalah 47 tipe spora (44 tipe Glomus dan 3 tipe Acaulospora). Banyaknya tipe
Glomus yang diperoleh menunjukkan bahwa Glomus mempunyai tingkat penyebaran yang lebih tinggi dibandingkan Acaulospora (Tabel 4). Hasil serupa yang menunjukkan bahwa tipe Glomus lebih sering ditemukan daripada tipe lainnya pada areal tanaman karet juga dilaporkan oleh Jayaratne (1982) dan Souza et al. (2010) yang masing-masing memperoleh 6 tipe Glomus, 2 tipe Acaulospora, 3 tipe
Gigaspora, 3tipe Sclerocystis, 3 Complexipes moniliformis dan 4 tipe Glomus, 1 tipe
Acaulospora.Tipe Glomus tampaknya sering ditemukan di areal tanaman karet. Deka
et al. (1998) memperoleh tipe Glomus dan Gigaspora. Ikram dan Mahmud (1984) dalam Deka et al. (1998) memperoleh Glomus, Acaulospora, dan Sclerocystis. Selain itu, keanekaragaman tipe spora yang didominasi oleh Glomus juga diperoleh dari penelitian lain (Karepesina, 2007; Muzakkir, 2010; Hartoyo et al., 2011; Songachan dan Kayang, 2011; Puspitasari et al., 2012; Iritie et al., 2013; dan Nurhandayani et al., 2013). Dengan demikian, dapat disimpulkan bahwa tingginya tingkat penyebaran
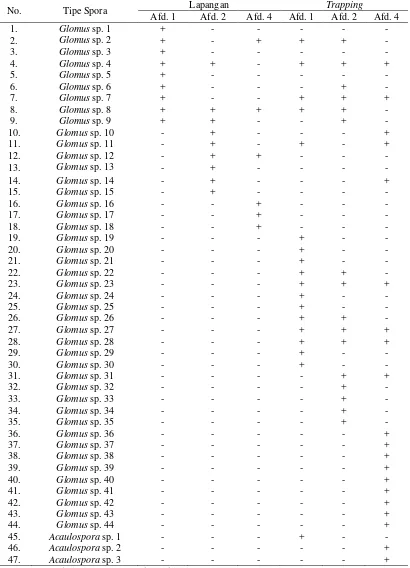
Tabel 4. Kehadiran tipe spora FMA di lapangan dan trapping
No. Tipe Spora Lapangan Trapping
Selain faktor adaptasi, waktu sporulasi (pembentukan spora) juga mempengaruhi adanya perbedaan jumlah tipe spora dari tiap genus. Hal ini dapat dikaitkan dengan waktu pengambilan sampel yang hanya satu kali sehingga tipe-tipe spora dari genus yang berbeda belum tentu terwakili secara keseluruhan. Corryanti (2011) menyatakan bahwa sebaran tipe FMA yang ditemukan terkait pada periode masa dengan kuantitas minimum atau maksimum spora yang dihasilkan. Oleh karena itu, keanekaragaman tipe spora dari genus yang berbeda akan lebih tinggi jika dilakukan lebih dari satu kali pengamatan. Hal ini telah dilaporkan oleh Delvian (2003) yang menunjukkan adanya variasi tipe spora pada suatu petak ukur selama lima kali pengamatan dengan interval waktu 3 bulan.
BAB V
KESIMPULAN DAN SARAN
5.1. Kesimpulan
Dari hasil penelitian ini dapat disimpulkan bahwa:
1. Kepadatan spora FMA di setiap afdeling (lapangan dan trapping) tergolong tinggi. Jumlah spora tertinggi diperoleh pada afdeling 1, yaitu 249 dan 319 spora/50 g tanah masing-masing dari hasil lapangan dan trapping.
2. Persentase kolonisasi FMA di setiap afdeling tergolong sedang dengan persentase tertinggi 42,5% pada afdeling 1.
3. Tipe spora FMA yang diperoleh adalah 44 tipe Glomus dan 3 tipe Acaulospora.
5.2. Saran
Saran yang dapat diberikan penulis, yaitu:
1. Mikoriza dipengaruhi oleh musim. Oleh karena itu, penelitian dengan menggunakan interval waktu selama pengambilan sampel perlu dilakukan untuk memperoleh data keanekaragaman FMA yang lebih valid.
DAFTAR PUSTAKA
Abbot, L.K. 1982. Comparative Anatomy of Vesicular Arbuscular Mycorrhizas Formadon Subterranean Clover. Aust. J. Bot. 30: 485-499.
Abbot, L.K. dan A.D. Robson. 1996. Working with Mycorrhizas in Forestry and Agriculture. ACIAR Monograph.
Akond, M.A., S. Mubassara, M.M. Rahman, S. Alam, dan Z.U.M. Khan. 2008. Status of Vesicular-Arbuscular (VA) Mycorrhizae in Vegetable Crop Plants of Bangladesh. World Journal of Agricultural Sciences 4 (6): 704-708.
Aytok, O., K.T. Yilmaz, I. Ortas, dan H. Cakan. 2013. Changes in Mycorrhizal Spore and Root Colonization of Coastal Dune Vegetation of The Seyhan Delta in The Postcultivation Phase. Turkish Journal of Agriculture and Forestry 37: 52-61. BMKG. 2013. Distribusi Curah Hujan September 2013. Diakses melalui
Bonfante, P. dan A. Genre. 2010. Mechanisms Underlying Beneficial Plant–Fungus Interactions in Mycorrhizal Symbiosis. Nature Communications 1 (48): 1-11. Brundrett, M., N. Bougher, B. Dell, T. Grave, dan N. Malajezuk. 1996. Workingwith
Mycorrizha in Forestry and Agriculture. Australia Centre for Internasional Agricultural Researche (ACIAR). Canberra.
Brundrett, M. 2006. Mycorrhizae-Mutualistic Plant-Fungus Symbioses. (35 pictures). Diakses melal Corryanti. 2011. Jamur Mikoriza Arbuskula Pada Lahan Tanaman Jati
Bertumpangsari Tebu. Jurnal Agrotropika 16 (1): 1-8.
Daniels, B.A. dan J.M. Trappe. 1980. Factors Affecting Spore Germination of The Vesicular-Arbuscular Mycorrhizal Fungus, Glomus epigaeus. Mycologia 72: 457-471.
Delvian. 2003. Keanekaragaman dan Potensial Pemanfaatan Fungi Mikoriza Arbuskula (FMA) di Hutan Pantai. Disertasi. Program Pascasarjana Institut Pertanian Bogor, Bogor.
Delvian. 2010. Keberadaan Cendawan Mikoriza Arbuskula di Hutan Pantai Berdasarkan Gradien Salinitas. Jurnal Ilmu Dasar 11 (2): 133-142.
Ervayenri, Y. Setiadi, N. Sukarno, dan C. Kusmana. 1997. Arbuscular Mycorrhizal Fungi (AMF) Diversity in Peat Soil Influenced by Land Vegetation Types. Proceedings of International Confrence on Mycorrhizas in Sustainable Tropical Agriculture and Forest Ecosystems: 85-90.
Gianinazzi-Pearson, V. dan S. Gianinazzi. 1983. The Physiology of Vesicular-Arbuscular Mycorrhizal Roots. Plant and Soil 71: 197–209.
Giovanetti. M. dan B. Mosse. 1980. An Evaluation of Technique for Meaning Vesikular Mycorrhiza Infection in Roots. New Phytologiest 84: 489-500.
Hartoyo, B., M. Ghulamahdi, L.K. Darusman, S.A. Aziz, dan I. Mansur. Keanekaragaman Fungi Mikoriza Arbuslula pada Rizosfer Tanaman Pegagan. Jurnal Littri 17 (1): 32-40.
Hardjowigeno, S. 1995. Ilmu Tanah. Mediyatama Sarana Perkasa. Jakarta.
Ikram, A. dan A.W. Mahmud. 1984. Endomycorrhizal Fungi in Soils Under Rubber.
Journal of The Rubber Research Institute of Malaysia 32: 198-206.
INVAM. 2013. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. Diakses melalui
April 2013.
Iritie, Z.B., T.B. Tra, Z.G. Noel, K.Z.C. Ghislaine, F.K. Romain, dan A. Zeze. 2012. Arbuscular Mycorrizal Fungi Associated with Theobroma cacao L. in The Region of Yamoussoukro (Cote d’Ivoire). African Journal of Agricultural Research 7 (6): 993-1001.
Janoukova, M., D. Pavlikova, dan M. Vosatka 2006. Potential Contribution of Arbuscular Mychorriza to Cadmium Immobilitation in Soil. Chemosphere 07. 007.
Karepesina, S. 2007. Keanekaragaman Fungi Mikoriza Arbuskula dari Bawah Tegakan Jati Ambon (Tectona grandis Linn. f.) dan Potensi Pemanfaatannya.
Tesis. Program Pascasarjana IPB, Bogor.
Khakpour, O. dan J. Khara. 2012. Spore Density and Root Colonization by Arbuscular Mycorrhizal Fungi in Some Species in The Northwest of Iran.
International Research Journal of Applied and Basic Sciences 3 (5): 977-982. Kormanik, P.P. dan A.C. McGraw. 1982. Quantification of VA Mycrorrhizae in Plant
Root. Di dalam: C. Schenck (Ed.) Methods and Principles of Mycorrhizae Research. The American Phytop. Soc. 46: 37-45.
Mansur, I., Y. Setiadi, dan R. Primaturi. 2002. Status of Research on Mycorrhizas Arbuscula Associated with Tropical Tree Species. Paper Presented at The Fourth International Wood Science Symposium (4th IWSS) LIPI-JSPS Core University Program in The Field of Wood Science. 2-3 September 2002. Research Centre for Physic Indonesian Institute of Science, Serpong, Tangerang, Indonesia.
Mosse, B. 1981. Vesicular Arbuscular Mycorrhiza Research for Tropical Agriculture. Research Buletin 194. College of Agriculture and Human. Resources Honolulu. University of Hawai.
Muzakkir. 2010. Keragaman dan Potensi Pemanfaatan FMA Indigenus Bersama Pupuk Hijau Terhadap Tanaman Jarak Pagar (Jatropha curcas L.) di Lahan Kritis. Disertasi. Program Pascasarjana Universitas Andalas, Padang.
Nurhandayani, R., R. Linda, dan S. Khotimah. 2013. Inventarisasi Jamur Mikoriza Vesikular Arbuskular dari Rhizosfer Tanah Gambut Tanaman Nanas [Ananas comosus (L.) Merr]. Protobiont 2 (3): 146 -151.
Nursanti, R.P. Tamin, dan Hamzah. 2012. Identifikasi Fungi Mikoriza Arbuskular (FMA) di Hutan Lindung Mangrove Pangkal Babu Kabupaten Tanjung Jabung Barat Jambi. Jurnal Penelitian Universitas Jambi Seri Sains 14 (2): 29-34. Pacioni, G. 1986. Sporulation of the VAM Fungi Stimulated by Water Stress in
Natural Conditions. Di dalam: Gianinazzi-Pearson, V. dan S. Gianinazzi (Eds.). Physiological and Genetical Aspect of Mycorrhizae. Proceeding of the 1st Europens Symposium on Mycorrhizae: 713-716.
Puspitasari, D., K.I. Purwani., dan A. Muhibuddin. Eksplorasi Vesicular Arbuscular Mycorrhiza (VAM) Indigenous pada Lahan Jagung di Desa Torjun, Sampang Madura. Jurnal Sains dan Seni ITS 1: 19-22.
Rothwell, F.M. 1984. Agregation of Surface Mine Soil by Interaction Between VAM Fungi and Lignin Degradation Product of Lespedeza. Plant Soil 80: 99-104. Schreiner, R.P. dan G.J. Bethlenfalvay. 1996. Mycorrhizae, Biocides, and Biocontrol.
4. Response of a Mixed Culture of Arbuscular Mycorrhizal Fungi and Host Plant to Three Fungicides. Biol. Fertil. Soils 23: 189-195.
Setiadi, Y., Mansur, dan Achmad. 1992. Mikrobiologi Tanah Hutan. Pusat Antar Universitas Bioteknologi Tanaman Pangan. Institut Pertanian Bogor, Bogor. Sghir, F., M. Chliyeh, W. Kachkouch, M. Khouader, A.O. Touhami, R. Benkirane,
dan A. Douira. 2013. Mycorrhizal Status of Olea europaea spp. oleaster in Morocco. Journal of Applied Biosciences 61: 4478–4489.
Simanungkalit, R.D.M. 1997. Effectiveness of 10 Species of Arbuscular Mycorrhizal (AM) Fungi Isolated from West Java and Lampung on Maize and Soybean. Di dalam: Jenie, U.A. et al., editor. Challenges of Biotechnology in The 21th Century. Proceedings of The Indonesian Biotechnology Conference Vol II: 17-19 Juni 17-1997. Jakarta: The Indonesian Biotechnology Consortium.
Smith, S.E., dan D.J. Read. 1997. Mycorrhizal Symbiosis (Second Edition). Academic
Press. Harcourt Brace & Company Publisher. London.
Smith, S.E. dan D.J. Read. 2008. Mycorrhizal Symbiosis (Third Edition). Academic Press. Great Britain.
Songachan, L.S. dan H. Kayang. 2011. Diversity and Species Composition of Arbuscular Mycorrhizal Fungi in Flemingia vestita Under Shifting and Continuous Cropping System. NeBIO 2 (4): 1-8.
Souza, G.I.A., A.L. Caproni, J.R.D.O. Granha, E.L. Souchie, dan R.L.L. Berbara. 2010. Arbuscular Mycorrhizal Fungi in Agricultural and Forest Systems. Gl. Sci. Technol. 3 (2): 1-9.
Sukarno, N., S.E. Smith, dan E.S. Scott. 1993. The Effect of Fungicides on Vesicular-Arbuscular Mycorrhizal Symbiosis. I. The Effects on Vesicular- Vesicular-Arbuscular Mycorrhizal Fungi and Plant Growth. New Phytol. 25: 139–147.
Utami, S.N. dan S. Handayani. 2003. Sifat Kimia Entisol Pada Sistem Pertanian Organik. Ilmu Pertanian 10 (2): 63-69.
Verheye, W. 2010. Growth and Production of Rubber. In: Verheye, W. (ed.), Land Use, Land Cover and Soil Sciences. Encyclopedia of Life Support Systems (EOLSS), UNESCO-EOLSS Publishers, Oxford, UK. Diakses melalui
Whiffen, L. 2007. Arbuscular Mycorrhizal Fungi and Carbon Sequestration in Soil. A Research Thesis Submitted to Fulfil The Requirements for The Degree of Doctor of Philosophy. School of Biological Sciences, The University of Sydney.
Lampiran 2. Kriteria Penilaian Sifat Kimia Tanah
Sifat Tanah Sangat
Rendah Rendah Sedang Tinggi
Sangat Tinggi C-organik
(%) < 1,00 1,00-2,00 2,01-3,00 3,01-5,00 > 5,00 P-avl Bray-II
(ppm) < 8,00 8,0-15 16-25 26-35 >35
KTK
(me/100 g) < 5 10-16 17-24 25-40 > 40
Sangat
Masam Masam
Agak
Masam Netral
Agak
Alkalis Alkalis pH
(H2
< 4,5
O) 4,5-5,5 5,6-6,5 6,6-7,5 7,6-8,5 >8,5