KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA
DI HUTAN PANTAI SONANG, TAPANULI TENGAH
T E S I S
Oleh
AJA NAZLI YUSNI
077030003/BIO
PROGRAM MAGISTER BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS SUMATERA UTARA
KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA
DI HUTAN PANTAI SONANG, TAPANULI TENGAH
T E S I S
Diajukan untuk melengkapi tugas dan memenuhi syarat mencapai gelar Magister Sains dalam Program Studi Biologi Pada Pada
Program Pascasarjana Universitas Sumatera Utara
Oleh
AJA NAZLI YUSNI
077030003/BIO
PROGRAM MAGISTER BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS SUMATERA UTARA
LEMBAR PERSETUJUAN
Judul Penelitian : KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA DI
HUTAN PANTAI SONANG, TAPANULI TENGAH
Nama Mahasiswa : Aja Nazli Yusni
NIM : 077030003
Program Studi : Biologi
Konsentrasi : Mikrobiologi
Menyetujui
Komisi Pembimbing
Dr. Delvian, SP, MP Ketua
Prof. Dr. Dwi Suryanto, MSc Anggota
Ketua Program Studi Biologi Dekan FMIPA
Prof. Dr. Dwi Suryanto, M.Sc. Prof. Dr. Eddy Marlianto, M.Sc.
Telah diuji
Pada tanggal : 8 Januari 2010
Panitia Penguji Tesis :
Ketua : Dr. Delvian, SP. MP
Anggota : 1. Prof. Dr. Dwi Surianto, M. Sc.
PERNYATAAN
Dengan ini saya menyatakan bahwa tesis saya yang berjudul Keanekaragaman Fungi Mikoriza Arbuskula Di Hutan Pantai Sonang, Tapanuli Tengah adalah benar merupakan hasil karya sendiri dan belum pernah dipublikasikan. Sumber data dan informasi yang digunakan telah dinyatakan dengan jelas dan dapat diperiksa kebenarannya.
Medan, 8 Januari 2010
ABSTRAK
Penelitian ini bertujuan untuk mengetahui keberadaan dan keanekaragaman Fungi Mikoriza Arbuskula (FMA) di hutan Pantai Sonang, Tapanuli Tengah dan hubungannya dengan tingkat salinitas tanah. Sampel tanah diambil dari setiap petak ukur. Untuk isolasi spora digunakan 50 gram tanah sampel dari setiap petak ukur. Isolasi spora FMA menggunakan metode tuang saring untuk penghitungan spora, identifikasi spora dan kolonisasi akar diteliti dengan menggunakan metode pewarnaan. Hasil penelitian menunjukkan bahwa salinitas berpengaruh negatif terhadap keberadaan dan keanekaragaman FMA serta persentase kolonisasi akar. Penurunan tingkat salinitas tanah diikuti dengan peningkatan kepadatan dan keanekaragaman spora serta persentase
kolonisasi akar. Hasil trapping spora dengan tanaman inang Pueraria javanica
menghasilkan jumlah spora yang lebih banyak dibandingkan dengan spora yang diisolasi dari lapangan. Pada penelitian ini didapat 28 jenis spora FMA, 26 jenis merupakan genus
Glomus dan 2 jenis lainnya merupakan genus Acaulospora. Kolonisasi ditemukan pada semua akar tanaman sampel. Persentase kolonisasi secara umum adalah rendah sampai dengan sedang
ABSTRACT
The objective of this study was to find out existence and diversity of arbuscular mycorrhizal fungi (AMF) as well as its relationship with soil salinity level in Sonang coastal forest, Tapanuli Tengah. Soil sample was taken from each plot. For spores isolation, fifty gram of soil sample was taken from each plot. The AMF spores were isolated using filtering-pouring method to calculate the number of spores, identification of spores and colonization of roots were observed using staining method. The result showed that salinity affected negatively on both the existence and diversity of AMF and root colonization. The decrease of soil salinity level was followed by the increasing in density and diversity of spores as well as the percentage of root colonization. The result
of trapping of spores with Pueraria javanica plant produced more number of spores than
the number of spores isolated from the field. There were 28 types of AMF spores found
in this study, 26 types of those were Glomus and two other types were Acaulospora.
Colonization was found in all roots of sample plant. The percentages of colonization were generally classified as low to medium level.
KATA PENGANTAR
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala rahmat dan
karunia-Nya sehingga tesis yang berjudul Keanekaragaman Fungi Mikoriza
Arbuskula Di Hutan Pantai Sonang, Tapanuli Tengah dapat terselesaikan. Tesis ini
dibuat sebagai salah satu syarat untuk menyelesaikan studi pada Program Studi Magister
Biologi Sekolah Pascasarjana Universitas Sumatera Utara Medan.
Selama pelaksanaan penelitian ini penulis telah banyak mendapat bantuan dan
bimbingan dari berbagai pihak. Untuk itu pada kesempatan ini penulis ingin
menyampaikan rasa terima kasih kepada:
1. Gubernur Sumatera Utara dan Ketua BAPPEDA yang telah memberikan Beasiswa S2
kepada penulis di sekolah Pascasarjana Universitas Sumatera Utara.
2. Rektor Universitas Sumatera Utara, Prof.Dr. H. Chairuddin P. Lubis DTM & H atas
kesempatan dan fasilitas yang diberikan kepada kami untuk mengikuti dan
menyelesaikan pendidikan program Magister.
3. Direktur Program Pascasarjana Universitas Sumatera Utara yang dijabat oleh Prof.
Dr. Ir. T. Chairun Nisa B, M.Sc atas kesempatan yang diberikan kepada kami
sehingga kami dapat menjadi mahasiswa Program Magister pada Program
Pascasarjana Universitas Sumatera Utara.
4. Dekan Fakultas Matematika Dan Ilmu Pengetahuan Alam Universitas Sumatera Utara
yang dijabat oleh Prof. Dr. Eddy Marlianto, M.Sc. yang telah memberikan
kesempatan kepada kami sehingga kami dapat menjadi mahasiswa Program Megister
Dalam Program Studi Biologi di Fakultas Matematika Dan Ilmu Pengetahuan Alam
5. Ketua Program Studi Magister Biologi Prof. Dr. Dwi Suryanto, M.Sc yang telah
memberikan bantuan, bimbingan dan arahan kepada kami.
6. Dr. Delvian, SP, MP selaku ketua komisi pembimbing dan Prof. Dr.Dwi Suryanto,
M.Sc selaku pembimbing II, yang dengan penuh perhatian telah memberikan
dorongan, bimbingan, arahan serta bantuan kepada penulis.
7. Prof. Dr. Erman Munir, M.Sc. sebagai penguji I serta Dr. Deni Elfiati, SP. MP
sebagai penguji II yang telah banyak memberikan arahan dan masukan dalam
penyempurnaan penyusunan tesis ini.
8. Drs. Sufrizal Tanjung, M.Si selaku kepala sekolah SMA Negeri 6 Medan yang telah
memberikan kesempatan dan dukungan kepada penulis
9. Suamiku Drs. Ikhwan Faizan Nasution, M.AP dan anak-anakku tersayang yang telah
memberikan motivasi, doa dan mendampingi dengan sabar selama pendidikan demi
keberhasilan studi ini.
Penulis menyadari bahwa tesis ini masih jauh dari sempurna. Oleh karena itu
dengan segala kerendahan hati penulis menerima kritik dan saran yang membangun dari
semua pihak demi kesempurnaan tesis ini. Penulis berharap semoga hasil penelitian ini
dapat bermanfaat bagi pihak yang memerlukannya dan bagi kita semua. Amin.
Medan, Januari 2010
RIWAYAT HIDUP
Penulis dilahirkan di Medan pada tanggal 5 September 1970, sebagai anak kedua
dari lima bersaudara, dari pasangan alm. H. Dt. Azham Yusni dan Hj. T. Zairul Laily.
Tahun 1983 penulis menyelesaikan pendidikan sekolah dasar di SD Pertiwi Medan,
selanjutnya pada tahun 1986 menyelesaikan pendidikan sekolah menengah pertama di
SMP Negeri 9 Medan. Tahun 1989 penulis menyelesaikan pendidikan sekolah menengah
atas di SMA Negeri 3 Medan. Pendidikan sarjana ditempuh di IKIP Negeri Medan,
Fakultas Pendidikan Matematika dan Ilmu Pengetahuan Alam, Jurusan Biologi dan lulus
pada tahun 1994. Kemudian penulis mendapat kesempatan melanjutkan pendidikan ke
Program Magister (S2) Biologi di Sekolah Pascasarjana Universitas Sumatera Medan,
mulai tahun 2007 melalui beasiswa pemerintah Provinsi Sumatera Utara.
Tahun 1992-1995 penulis bertugas sebagai guru honor bidang studi Biologi di SMP
swasta Perguruan Husni Thamrin Medan dan SMA swasta Brigjen Katamso Medan. Pada
tahun 1994 penulis diangkat sebagai guru PNS yang ditugaskan pada SMA Negeri 3
Tebingtinggi hingga tahun 1996. Sejak bulan Juli 1996 sampai dengan sekarang, penulis
dipindahtugaskan dari SMA Negeri 3 Tebingtingi ke SMA Negeri 6 Medan sebagai guru
DAFTAR ISI
Halaman
ABSTRACT ... iii
ABSTRAK ... iv
KATA PENGANTAR ... v
RIWAYAT HIDUP ... vii
DAFTAR ISI ... viii
DAFTAR TABEL ... x
DAFTAR GAMBAR ... xi
DAFTAR LAMPIRAN ... xii
I. PENDAHULUAN ... 1
1.1 Latar Belakang ... 1
1.2 Permasalahan ... 2
1.3 Tujuan Penelitian ... 3
1.4 Manfaat Penelitian ... 3
II. TINJAUAN PUSTAKA ... 4
2.1 Klasifikasi Fungi Mikoriza Arbuskula ... 4
2.2 Struktur Fungi Mikoriza Arbuskula ... 4
2.3 Manfaat Fungi Mikoriza Arbuskula bagi Tanaman Inang ... 6
2.4 Faktor-faktor yang Mempengaruhi Pertumbuhan dan Penyebaran FMA ... 9
2.5 Hutan Pantai ... 14
2.6 Keberadaan dan Keanekaragaman FMA di Daerah Bersalinitas Tinggi ... 15
III. BAHAN DAN METODE ... 17
3.1 Tempat dan Waktu ... 17
3.2 Bahan dan Alat... 17
3.3 Pelaksanaan Penelitian ... 18
IV. HASIL DAN PEMBAHASAN ... 24
4.1 Sifat Kimia Tanah ... 24
4.2 Kepadatan Spora ... 27
4.3 Persentase Kolonisasi ... 44
V. KESIMPULAN DAN SARAN ... 51
5.1 Kesimpulan ... 51
5.2 Saran ... 51
DAFTAR PUSTAKA ... 52
DAFTAR TABEL
Tabel Judul Halaman
1 2 3 4 5 6 7 8
Hasil Analisis Beberapa Sifat Kimia Sampel Tanah...
Keanekaragaman spora FMA yang Ditemukan dari Lapangan Berdasarkan Tingkat Salinitas Tanah...
Keanekaragaman Spora FMA Hasil Trapping Berdasarkan
Tingkat Salinitas Tanah ...
Keanekaragaman karakteristik spora FMA yang ditemukan
dilapangan...
Keanekaragaman karakteristik spora FMA hasil trapping ...
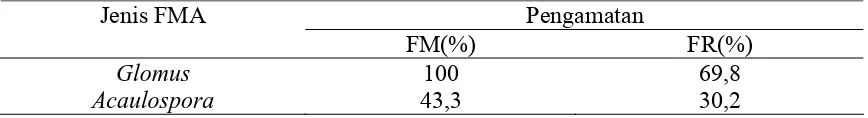
Nilai Frekuensi Mutlak (FM) dan Frekuensi Relatif (FR)
Kehadiran Suatu Genus FMA di Lapangan ...
Nilai Frekuensi Mutlak (FM) dan Frekuensi Relatif (FR)
Kehadiran Suatu Genus FMA pada Pengamatan Hasil Trapping....
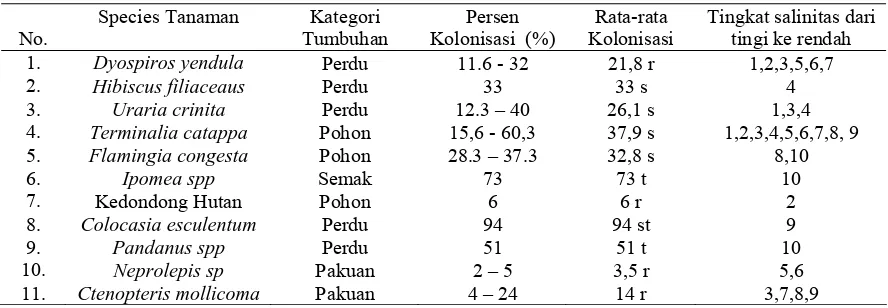
Keberadaan FMA pada Vegetasi Hutan Pantai Sonang ...
DAFTAR GAMBAR
Gambar Judul Halaman
1
2
3
4
5
6
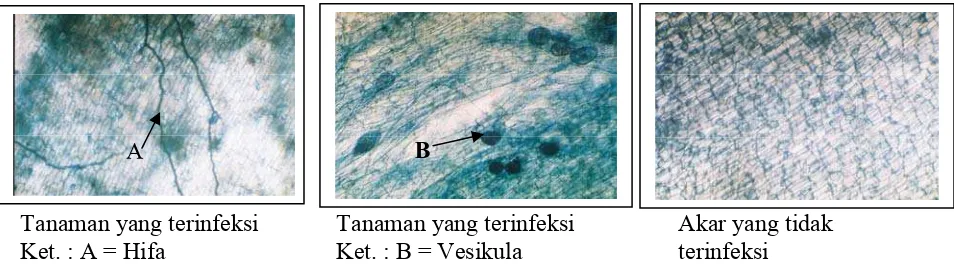
Penampang Longitudinal Akar yang Terinfeksi FMA ...
Jumlah Spora di Lapangan Berdasarkan Tingkat Salinitas
tanah ….……….……….
Jumlah Spora Hasil Trapping Berdasarkan Tingkat Salinitas
Tanah ... …
Infeksi FMA pada Akar Tanaman yang Diamati ...
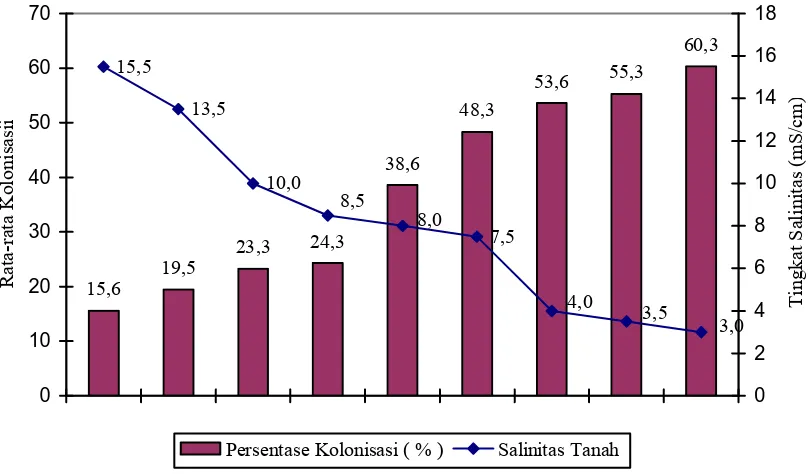
Persentase Kolonisasi FMA Berdasarkan Tingkat Salinitas Tanah ...
Persentase Kolonisasi pada Berbagai Tingkat Salinitas Tanah
pada Satu Individu Tanaman, yaitu tanaman Terminalia catappa..
5
28
32
44
45
DAFTAR LAMPIRAN
Lampiran Judul Halaman
1 2 3 4 5 6 7 8 9 10 11 12
Teknik Pengambilan Sampel Tanah dan Akar ...
Tahapan Ekstraksi untuk Penghitungan Jumlah dan Identifikasi Spora Fungi Mikoriza ...
Pengamatan Kolonisasi Fungi Mikoriza pada Akar Tanaman Sampel ...
Teknis Pembuatan Kultur Trapping Fungi Mikoriza ...
Dokumentasi Penelitian Tanaman Pueraria Javanica dalam
Trapping ...
Kriteria Penggolongan Kadar salinitas dalam Tanah menurut Afandie dan Nasih, 2002 ...
Kriteria Penggolongan pH tanah menurut Sarwono
Hardjowigeno, 1987 ...
Kriteri Penggolongan C Organik menurut Sarwono
Hardjowigeno, 1987 ...
Kriteria Penggolongan N total menurut Sarwono Hardjowigeno, 1987 ...
Kriteria Penggolongan P tersedia menurut Sarwono
Hardjowigeno, 1987 ...
Kriteria Persentase Kolonisasi Akar menurut Setiadi et al.
(1992) ...
Data Hasil Analisis Tanah ...
ABSTRAK
Penelitian ini bertujuan untuk mengetahui keberadaan dan keanekaragaman Fungi Mikoriza Arbuskula (FMA) di hutan Pantai Sonang, Tapanuli Tengah dan hubungannya dengan tingkat salinitas tanah. Sampel tanah diambil dari setiap petak ukur. Untuk isolasi spora digunakan 50 gram tanah sampel dari setiap petak ukur. Isolasi spora FMA menggunakan metode tuang saring untuk penghitungan spora, identifikasi spora dan kolonisasi akar diteliti dengan menggunakan metode pewarnaan. Hasil penelitian menunjukkan bahwa salinitas berpengaruh negatif terhadap keberadaan dan keanekaragaman FMA serta persentase kolonisasi akar. Penurunan tingkat salinitas tanah diikuti dengan peningkatan kepadatan dan keanekaragaman spora serta persentase
kolonisasi akar. Hasil trapping spora dengan tanaman inang Pueraria javanica
menghasilkan jumlah spora yang lebih banyak dibandingkan dengan spora yang diisolasi dari lapangan. Pada penelitian ini didapat 28 jenis spora FMA, 26 jenis merupakan genus
Glomus dan 2 jenis lainnya merupakan genus Acaulospora. Kolonisasi ditemukan pada semua akar tanaman sampel. Persentase kolonisasi secara umum adalah rendah sampai dengan sedang
ABSTRACT
The objective of this study was to find out existence and diversity of arbuscular mycorrhizal fungi (AMF) as well as its relationship with soil salinity level in Sonang coastal forest, Tapanuli Tengah. Soil sample was taken from each plot. For spores isolation, fifty gram of soil sample was taken from each plot. The AMF spores were isolated using filtering-pouring method to calculate the number of spores, identification of spores and colonization of roots were observed using staining method. The result showed that salinity affected negatively on both the existence and diversity of AMF and root colonization. The decrease of soil salinity level was followed by the increasing in density and diversity of spores as well as the percentage of root colonization. The result
of trapping of spores with Pueraria javanica plant produced more number of spores than
the number of spores isolated from the field. There were 28 types of AMF spores found
in this study, 26 types of those were Glomus and two other types were Acaulospora.
Colonization was found in all roots of sample plant. The percentages of colonization were generally classified as low to medium level.
BAB I
PENDAHULUAN
1.1. Latar Belakang
Mikoriza merupakan bentuk hubungan simbiosis mutualisme antara fungi dengan
perakaran tingkat tinggi. Hubungan simbiosis antara inang dengan fungi meliputi
penyediaan fotosintat (karbohidrat) oleh tanaman inang. Sebaliknya, Fungi Mikoriza
Arbuskula (FMA) aktif membantu tanaman dalam penyerapan hara, meningkatkan
ketahanan tanaman terhadap kekeringan dan serangan patogen akar, bahkan dapat
meningkatkan pertumbuhan pada daerah-daerah bekas pertambangan, meningkatkan
produksi hormon dan zat pengatur tumbuh, memperbaiki struktur tanah,
mempertahankan keanekaragaman tumbuhan (Musnamar, 2003; Setiadi, 1999;
Munyanziza et al., 1997; Killham, 1994; Wani et al., 1991; Sieverding, 1991; Imas et
al.,1989).
Keberadaan FMA di alam bersifat kosmopolitan, artinya fungi mikoriza dipastikan
ada dalam kondisi tanah apapun, termasuk tanah di hutan pantai yang berpasir. Mikoriza
ini memiliki kemampuan bersimbiosis dengan hampir 90% jenis tanaman. Fungi
Mikoriza Arbuskula pada umumnya dapat ditemukan pada spesies tanaman tingkat tinggi
yang tumbuh pada berbagai tipe habitat dan iklim. Adapun penyebarannya bervariasi
menurut iklim, lingkungan dan tipe penggunaan lahan (Setiadi, 2001).
Simbiosis FMA telah lama diyakini sangat penting untuk pertumbuhan dan
stabilitas tanaman di daerah pantai (Siguenza et al., 1996) yang umumnya dicirikan
dengan ketersediaan fosfor yang rendah (Nicholson, 1960 dalam Siguenza et al., 1996).
dalam, rambut akarnya sedikit sehingga kurang mampu dalam penyerapan zat hara.
Kondisi lahan yang berpasir, tanah yang bersalinitas tinggi, temperatur permukaan yang
tinggi dan hembusan angin yang kencang yang berakibat evapotranspirasi sangat tinggi
juga sangat mempengaruhi. Daerah yang kondisi seperti ini peranan fungi mikoriza
sangat diperlukan (Siradz et al., 2007).
Belakangan ini FMA mendapat perhatian yang lebih dari para ahli lingkungan dan
pertanian karena merupakan salah satu alternatif teknologi di masa datang yang tidak saja
efektif tetapi juga ramah lingkungan. Keadaan ini telah mendorong sejumlah survei untuk
mempelajari keanekaragaman serta penyebaran FMA di suatu daerah tertentu atau
ekosistem alami (Delvian, 2003; Ruiz–Lozano dan Azcon, 2000; Ragupathy dan
Mahadevan, 1991; Kim dan Weber, 1985).
Penelitian yang ada tentang keberadaan dan keanekaragaman FMA pada ekosistem
alami, khususnya pada tanah salin di Indonesia masih sangat kurang. Padahal keberadaan
dan keanekaragaman FMA sangat penting artinya dalam menanggulangi masalah
salinitas. Hal inilah yang menjadi latar belakang dilakukannya penelitian ini. Dalam
penelitian ini diharapkan dapat diidentifikasi jenis-jenis FMA yang terdapat pada tanah
salin, khususnya di hutan pantai Sonang, Tapanuli Tengah.
1.2 Permasalahan
Diyakini bahwa FMA mempunyai kemampuan dalam menanggulangi masalah
salinitas pada tumbuhan didaerah pantai. Akan tetapi sampai saat ini belum tersedia isolat
yang potensial untuk tanah salin. Bahkan informasi tentang keberadaan dan
keanekaragaman FMA pada ekosistem alami khususnya tanah salin masih sangat kurang,
daerah pantai. Berdasarkan permasalahan tersebut peneliti menganggap perlu dilakukan
penelitian mengenai keanekaragaman fungi mikoriza pada tanah salin dihutan pantai.
1.3. Tujuan Penelitian
Tujuan penelitian ini adalah untuk mengetahui keberadaan dan keanekaragaman
FMA di Hutan Pantai Sonang, Tapanuli Tengah berdasarkan tingkat salinitas tanah.
1.4. Manfaat Penelitian
Memberi informasi mengenai keanekaragaman FMA di Hutan Pantai Sonang,
BAB II
TINJAUAN PUSTAKA
2.1 Klasifikasi Fungi Mikoriza Arbuskula
Fungi mikoriza arbuskula adalah salah satu tipe fungi mikoriza dan termasuk ke
dalam golongan endomikoriza. Fungi mikoriza arbuskula termasuk ke dalam kelas
Zygomycetes, dengan ordo Glomeromycota yang mempunyai 2 sub-ordo, yaitu
Gigasporineae dan Glomineae. Gigasporineae dengan famili Gigasporaceae mempunyai
2 genus, yaitu Gigaspora dan Scutellospora. Glomineae mempunyai 4 famili, yaitu
famili Glomaceae dengan genus Glomus dan Sclerocystis, famili Acaulosporaceae
dengan genus Acaulospora dan Entrophospora, Paraglomaceae dengan genus
Paraglomus, dan Archaeosporaceae dengan genus Archaeospora (INVAM, 2009)
2.2 Struktur Fungi Mikoriza Arbuskula
Karakteristik FMA menurut Scannerini dan Bonfante-Fosolo (1983) adalah sebagai
berikut: sistem perakaran tanaman yang terinfeksi FMA tidak membesar, fungi
membentuk struktur lapisan hifa tipis dan tidak merata pada permukaan akar, hifa masuk
ke dalam individu sel jaringan korteks, dan pada umumnya ditemukan struktur
percabangan hifa yang disebut arbuskula (arbuscules) dan struktur khusus berbentuk
oval yang disebut vesikula (vesicles). Anatomi sederhana dari FMA dapat dilihat pada
Gambar 1. Penampang Longitudinal Akar yang Terinfeksi FMA
(Sumber: Brundrett et al., 1994)
Arbuskula merupakan suatu struktur mirip haustorium pada jamur pathogen yang
dibentuk oleh hifa tipis interselular 2-3 hari setelah infeksi dan mempunyai banyak
percabangan halus. Arbuskula berfungsi sebagai tempat pertukaran metabolit antara fungi
dan tanaman inang. Adanya arbuskula sangat penting untuk mengidentifikasi bahwa telah
terjadi infeksi pada akar tanaman (Scannerini dan Bonfante-Fosolo, 1983; Bonfante
Fosolo, 1984). Selanjutnya dikatakan bahwa seluruh endofit dan yang termasuk genus
Gigaspora, Scutellospora, Glomus, Sclerocystis, dan Acaulospora mampu membentuk
Selain arbuskula, FMA juga memiliki vesikula yang terbentuk melalui
penggelembungan hifa internal, kebanyakan berbentuk bulat telur dan berisi banyak
lipida sehingga dapat berfungsi sebagai cadangan makanan (Suhardi, 1989). Pendapat
lain tentang fungsi dari vesikula ini, yaitu sebagai organ istirahat, karena jumlahnya
meningkat pada saat tanaman tua atau saat tanaman akan mati. Vesikula ditemukan baik
di dalam maupun di luar lapisan kortek parenkhim dan tidak semua FMA membentuk
vesikula dalam akar inangnya, seperti Gigaspora dan Scutellospora vesikulanya
ekstra-radikal dan tidak teratur (Bonfante-Fosolo, 1984; Abbott dan Robson, 1982)
2.3 Manfaat Fungi Mikoriza Arbuskula Bagi Tanaman Inang
Adanya simbiosis mutualistik antara FMA dengan perakaran tanaman dapat
membantu pertumbuhan tanaman menjadi lebih baik, terutama pada tanah-tanah marjinal.
Hal ini disebabkan FMA efektif dalam meningkatkan penyerapan unsur hara makro dan
mikro ( Killham, 1994; Abbott dan Robson, 1984 ), meningkatkan daya tahan tanaman
terhadap serangan patogen (Wani et al., 1991), meningkatkan ketahanan terhadap
kekeringan dan dapat membantu pertumbuhan tanaman pada daerah yang tercemar logam
berat (Munyanziza et al., 1997), meningkatkan produksi hormon dan zat pengatur
tumbuh (Imas et al., 1989), memperbaiki struktur tanah (Wright dan Uphadhyaya, 1998;
Sieverding, 1991) dan mempertahankan keanekaragaman tumbuhan (Setiadi, 1999).
2.3.1. Meningkatkan penyerapan unsur hara makro dan mikro
Jaringan hifa eksternal dari mikoriza akan memperluas bidang serapan air dan hara.
Disamping itu ukuran hifa yang lebih halus dari bulu-bulu akar memungkinkan hifa bisa
kondisi kadar air tanah yang sangat rendah (Killham, 1994). Serapan air yang lebih besar
oleh tanaman bermikoriza,juga membawa unsur hara yang mudah larut dan terbawa oleh
aliran masa seperti N, K dan S, sehingga serapan unsur tersebut juga makin meningkat.
Disamping serapan hara melalui aliran masa, serapan P yang tinggi juga disebabkan
karena hifa. Fungi juga mengeluarkan enzim fosfatase yang mampu melepaskan P dari
ikatan-ikatan spesifik, sehingga tersedia bagi tanaman. Menurut Abbott dan Robson
(1984), akar yang bermikoriza dapat meningkatkan kapasitas pengambilan hara karena
waktu hidup akar yang dikolonisasi diperpanjang dan derajad percabangan serta diameter
akar diperbesar, sehingga luas permukaan absorbsi akar diperluas.
2.3.2 Meningkatkan daya tahan tanaman terhadap serangan patogen
Mikoriza dapat meningkatkan pertumbuhan tanaman melalui perlindungan
tanaman dari patogen akar dan unsur toksik (Wani et al., 1991). Mikoriza menggunakan
hampir semua kelebihan karbohidrat dan eksudat lainnya, sehingga tercipta lingkungan
yang tidak cocok untuk patogen. Fungi mikoriza dapat mengeluarkan antibiotik yang
dapat mematikan patogen. Akar tanaman yang sudah diinfeksi fungi mikoriza, tidak
dapat diinfeksi oleh fungi patogen yang menunjukkan adanya kompetisi.
2.3.3. Meningkatkan ketahanan terhadap kekeringan
Tanaman yang bermikoriza lebih tahan kekeringan dari pada yang tidak
bermikoriza dan akan cepat kembali pulih setelah periode kekeringan berakhir. Hal ini
dimungkinkan karena hifa FMA masih mampu menyerap air pada pori-pori tanah pada
saat akar tanaman sudah tidak mampu. Selain itu penyerapan hifa di dalam tanah sangat
2.3.4. Membantu pertumbuhan tanaman pada daerah yang tercemar logam berat
Mikoriza juga dapat melindungi tanaman dari ekses unsur tertentu yang bersifat
racun seperti logam berat (Munyanziza et al., 1997). Mekanisme perlindungan terhadap
logam berat dan unsur beracun yang diberikan mikoriza dapat melalui efek filtrasi,
menonaktifkan secara kimiawi atau penimbunan unsur tersebut dalam hifa fungi.
2.3.5. Meningkatkan produksi hormon dan zat pengatur tumbuh
Menurut Imas et al. (1989), FMA dapat meningkatkan produksi hormon
pertumbuhan seperti auksin, sitokinin, dan gibberelin dan juga zat pengatur tumbuh
seperti vitamin bagi tanaman inangnya. Auksin berfungsi memperlambat proses penuaan
akar sehingga fungsi akar sebagai penyerap unsur hara dan air akan bertahan lebih lama.
2.3.6. Memperbaiki struktur tanah
Fungi mikoriza arbuskula memiliki peranan yang penting dalam perbaikan
struktur tanah yaitu dengan cara mengikat partikel-partikel tanah melalui perkembangan
miselliumnya yang intensif (Sieverding, 1991). Wright dan Uphadhyaya (1998)
mengatakan bahwa FMA menghasilkan senyawa glycoprotein glomalin yang sangat
berkorelasi dengan peningkatan kemantapan agregat. Konsentrasi glomalin lebih tinggi
ditemukan pada tanah-tanah yang tidak diolah dibandingkan dengan yang diolah.
Glomalin dihasilkan dari sekresi hifa eksternal bersama enzim-enzim dan senyawa
polisakarida lainnya. Pengolahan tanah menyebabkan rusaknya jaringan hifa sehingga
2.3.7. Mempertahankan keanekaragaman tumbuhan
Fungi Mikoriza Arbuskula memiliki kemampuan berasosiasi dengan hampir 90%
jenis tanaman, maka FMA berperan dalam mempertahankan stabilitas keanekaragaman
tumbuhan dengan cara transfer nutrisi dari satu akar tumbuhan ke akar tumbuhan lainnya
yang berdekatan melalui struktur yang disebut Bridge Hypae (Setiadi, 1999).
2.4. Faktor-Faktor Yang Mempengaruhi Pertumbuhan Dan Penyebaran FMA
Faktor-faktor yang mempengaruhi perkembangan dan penyebaran FMA adalah
sebagai berikut :
2.4.1. Faktor musim
Di beberapa habitat ditemukan bahwa pembentukan spora FMA bersifat musiman
(Moreira et al., 2007; Delvian, 2006 a; Abbott dan Gazey, 1994). Umumnya studi tentang
variasi musiman dalam populasi FMA didasarkan pada jumlah spora yang diisolasi
(Abbott dan Gazey, 1994). Apakah puncak produksi spora terjadi pada musim
semi-musim panas atau semi-musim dingin-semi-musim gugur tampaknya berhubungan dengan iklim dan
tanaman tetapi penurunan jumlah spora yang nyata terjadi selama musim dingin.
Selanjutnya inokulum yang mampu bertahan selama musim dingin akan segera
mengkolonisasi akar tanaman pada musim semi. Jumlah spora meningkat selama musim
pertumbuhan dan kemudian menurun dengan berlalunya musim semi.
Pada kondisi basah atau banyak hujan umumnya persentase kolonisasi meningkat
dan pembentukan spora baru berkurang. Hal ini disebabkan karena kelembaban tanah yng
tinggi pada kondisi basah akan merangsang perkecambahan spora dan terbentuknya
pembentukan spora baru akan meningkat dan persentase kolonisasi akan menurun.
Kondisi kering akan merangsang pembentukan spora yang banyak sebagai respon alami
dari FMA serta upaya untuk mempertahankan keberadaannya di alam (Delvian b, 2006).
Kolonisasi akar ditemukan lebih efektif pada musim penghujan pada bulan oktober
sampai maret (Moreira et al., 2007).
2.4.2. Faktor lingkungan
Mikoriza sangat dipengaruhi oleh faktor lingkungan seperti: suhu, kadar air tanah,
pH tanah, bahan organik, cahaya dan ketersediaan hara, logam berat dan unsur lain, serta
fungisida .
A. Suhu
Suhu berpengaruh terhadap tahapan infeksi yakni pada perkembangan spora di
tanah, penetrasi hifa ke dalam sel akar dan perkembangan hifa di dalam korteks akar.
Kisaran suhu yang sesuai untuk perkembangan FMA adalah 30°C namun untuk
kolonisasi yang terbaik adalah 28° – 34°C , sedang perkembangan bagi vesikula pada
suhu 35°C ( Schenk dan Schroder, 1974 ).
B. Kadar air tanah
Status air tanah dapat berpengaruh baik langsung atau tidak langsung terhadap
FMA. Penjenuhan air tanah yang lama berpotensi mengurangi pertumbuhan dan infeksi
jamur mikoriza karena kondisi anaerob (Safir dan Duniway, 1982). Percobaan–percobaan
telah dilakukan pada tanah-tanah dengan berbeda-beda kadar airnya. Daniels dan Trappe
(1980) menggunakan Glomus epigaeum dikecambahkan pada lempung berdebu (silt
loam) pada berbagai kandungan air. G. epigaeum ternyata berkecambah paling baik pada
Terdapat juga fakta bahwa potensial air yang rendah dapat juga menurunkan infeksi
FMA secara drastis (Safir dan Duniway, 1982). Perkecambahan Gigaspora sp, akan
menurun pada kondisi potensial air yang rendah (-31 bar) ini juga mempengaruhi
pertumbuhan hifanya. Pada -10 bar perkecambahan sangat drastis menurun (Islami dan
Wani, 1995).
Mikoriza berkembang baik bila tidak ada hambatan aerase. Oleh karena itu
mikoriza dapat berkembang lebih baik pada tanah berpasir dibandingkan tanah berliat
atau gambut (Islami dan Wani 1995).
C. pH tanah
Fungi pada umumnya lebih tahan terhadap perubahan pH tanah. Meskipun
demikian daya adaptasi masing-masing species FMA terhadap pH tanah berbeda-beda,
tergantung pada adaptasi FMA terhadap lingkungan. pH dapat berpengaruh langsung
terhadap aktivitas enzim yang berperan dalam perkecambahan, perkembangan dan peran
mikoriza terhadap pertumbuhan tanaman (Maas dan Nieman, 1978). Glomus fasciculatus
berkembang biak pada pH masam. Demikian pula peran G. fasciculatus di dalam
meningkatkan pertumbuhan tanaman pada tanah masam menurun akibat pengapuran.
Pada pH 5,1 dan 5,9 G. fasciculatus menampakkan pertumbuhan yang terbesar. G.
fasciculatus memperlihatkan pengaruh yang lebih besar terhadap pertumbuhan tanaman
justru kalau pH 5,1. G. mosseae biasanya pada tanah alkali dapat berkecambah dengan
baik yaitu pada pH 6-9. Sedangkan spora dari Gigaspora corolloidea dan G. heterogama
dari jenis yang lebih asam dapat berkecambah dengan baik pada pH 4-6, Glomus
Perubahan pH tanah melalui pengapuran biasanya berdampak merugikan bagi
perkembangan FMA asli yang hidup pada tanah tersebut sehingga pembentukan mikoriza
menurun. Untuk itu tindakan pengapuran harus dibarengi tindakan inokulasi dengan
FMA yang cocok agar pembentukan mikoriza terjamin.
D. Bahan organik
Bahan organik merupakan salah satu komponen tanah yang penting disamping air
dan udara. Bahan organik dapat mengurangi kemampuan berkecambah dari spora baik
pada misellium agar maupun media tanah tidak steril. Ketersediaan nitrogen dan fosfat
yang rendah akan mendorong pertumbuhan mikoriza. Akan tetapi kandungan bahan
organik yang terlalu rendah atau tinggi menghambat pertumbuhan mikoriza.
Perkecambahan spora tidak hanya bergantung pada species dari FMA tetapi juga
kandungan bahan organik di dalam tanah (Islami dan Wani, 1995).
E. Cahaya dan ketersediaan hara
Peningkatan intensitas sinar dan panjang hari meningkatkan kolonisasi akar dan
produksi spora. Penyinaran dengan periode 12 jam atau lebih mungkin lebih penting
daripada intensitas sinar yang besar dengan periode penyinaran yang pandek di dalam
meningkatkan kolonisasi akar, tetapi dengan panjang hari penyinaran yang sesuai dengan
peningkatan intensitas sinar dapat meningkatkan kolonisasi (Moreira et al., 2007).
Adanya naungan yang berlebihan terutama untuk tanaman yang senang cahaya akan
mengurangi infeksi akar dan produksi spora, selain itu respon tanaman terhadap FMA
akan berkurang. Hal ini disebabkan adanya hambatan pertumbuhan dan perkembangan
internal hifa dalam akar yang berakibat pada terbatasnya perkembangan eksternal hifa
F. Logam berat dan unsur lain
Pada percobaan dengan menggunakan tiga jenis tanah dari wilayah iklim sedang
didapatkan bahwa pengaruh menguntungkan karena adanya FMA menurun dengan
naiknya kandungan Al dalam tanah. Aluminium diketahui terhambat munculnya jika
kedalam larutan tanah ditambahkan kalsium (Ca). Jumlah Ca di dalam larutan tanah
ternyata mempengaruhi perkembangan FMA. Tanaman yang ditumbuhkan pada tanah
yang memiliki derajat infeksi FMA yang rendah. Hal ini mungkin karena peran Ca 2+
dalam memelihara integritas membran sel.
Beberapa species FMA diketahui mampu beradaptasi dengan tanah yang tercemar
seng (Zn), tetapi sebagian besar species FMA peka terhadap kandungan Zn yang tinggi.
Pada beberapa penelitian lain diketahui pula bahwa strain-strain species FMA tertentu
toleran terhadap kandungan Mn, Al, dan Na yang tinggi (Janouskuva et al., 2006).
G. Fungisida
Fungisida merupakan racun kimia yang dibuat untuk membunuh fungi penyebab
penyakit pada tanaman, akan tetapi selain membunuh fungi penyebab penyakit, fungisida
juga dapat membunuh mikoriza, dimana pemakaian fungisida ini menurunkan
pertumbuhan dan kolonisasi serta kemampuan mikoriza dalam menyerap P.
2.4.3 Tipe penggunaan lahan
Hutan alami yang terdiri dari banyak species tanaman dan umur yang tidak seragam
sangat mendukung perkembangan mikoriza. Konservasi hutan untuk lahan pertanian akan
mengurangi keragaman jenis dan jumlah propagul fungi, karena perubahan spesies
multi spesies berubah menjadi hutan monokultur dengan umur seragam sangat
berpengaruh terhadap jumlah dan keragaman mikoriza ( Setiadi, 2001 ).
Praktek pertanian seperti pengolahan tanah, sistem penanaman, ameliorasi dengan
bahan organik, pemupukan dan penggunaan pestisida sangat berpengaruh terhadap
keberadaan mikoriza (Zarate dan Cruz, 1995). Pengolahan tanah yang intensif akan
merusak jaringan hifa eksternal fungi mikoriza. Penelitian McGonigle dan Miller (1993),
menunjukkan bahwa pengolahan tanah minimum akan meningkatkan populasi mikoriza
dibandingkan pengolahan tanah konvensional. Dalam budidaya tradisional, pengolahan
tanah berulang-ulang dan panen menyebabkan erosi hara dan bahan organik dari lahan
tersebut dan ini berpengaruh terhadap populasi mikoriza. Dalam pertanian modern yang
menggunakan pupuk dan pestisida berlebihan (Rao, 1994) serta terjadinya kompaksi
tanah oleh alsintan (Mc Gonigle dan Miller, 1993) berpengaruh negatif terhadap
pembentukan mikoriza.
2.5. Hutan Pantai
Tipe ekosistem hutan pantai terdapat didaerah-daerah kering tepi pantai dengan
kondisi tanah berpasir atau berbatu dan terletak di atas garis pasang tertinggi. Di daerah
seperti itu pada umumnya jarang tergenang oleh air laut, namun sering terjadi atau
terkena angin kencang dengan hembusan garam (Arief, 1994).
Spesies-spesies pohon yang pada umumnya terdapat dalam ekosistem hutan pantai
antara lain Barringtonia speciosa, Terminalia catappa, Callophyllu inophyllum, Hibiscus
tiliaceus, Thespenia populnea, Casuarina equisetifolia, dan Pisonia grandis (Direktorat
kadang-kadang terdapat juga species pohon Hernandia peltata, Manilkara kauki, dan
Sterculia foetida (Arief, 1994).
2.6. Keberadaan dan keanekaragaman FMA di daerah bersalinitas tinggi
Hubungan antara salinitas tanah dan keberadaan FMA telah diteliti oleh beberapa
peneliti (Delvian, 2003; Kim dan Weber, 1985; Ragupathy dan Mahadevan, 1991).
Delvian (2003) melakukan survey keberadaan FMA pada daerah pantai di Hutan Cagar
Alam Leuweng Sancang Kabupaten Garut, Jawa Barat. Kepadatan spora meningkat
sejalan dengan menurunnya salinitas tanah, dimana kepadatan spora terendah terdapat
pada tanah dengan tingkat salinitas 10,2 mmho/cm dan kepadatan spora tertinggi terdapat
pada tanah dengan tingkat salinitas 6,1 mmho/cm. Persentasi kolonisasi juga meningkat
dengan menurunnya tingkat salinitas tanah, dimana persentasi kolonisasi terendah
terdapat pada tingkat salinitas 10,2 mmho/cm dan tingkat kolonisasi tertinggi terdapat
pada tingkat salinitas 5,9 mmho/cm. Selain mempengaruhi kepadatan spora dan
kolonisasi, tingkat salinitas tanah juga mempengaruhi jumlah tipe spora FMA, dimana
penurunan tingkat salinitas tanah menghasilkan peningkatan jumlah tipe spora FMA.
Penghitungan frekuensi mutlak dan frekuensi relatif kehadiran setiap spora FMA didapat
spora tipe Glomus mempunyai frekuensi mutlak dan frekuensi relative kehadiran tertinggi
(100% dan 37,98%) yang diikuti oleh Acaulospora (76% dan 28,82%), Sclerocystis (54%
dan 20,38%) dan Gigaspora (34% dan 12,81%).
Kim dan Weber (1985) melakukan survey keberadaan FMA pada halofit sepanjang
transect Danau Playa, Utah dan menyimpulkan bahwa kepadatan spora FMA
berhubungan erat dengan salinitas tanah dimana kepadatan spora FMA akan menurun
Ragupathy dan Mahadevan (1991) mempelajari pengaruh gradien salinitas
terhadap distribusi CMA di Kodikkarai Reserve Forest, Tamil Nadu, India menyatakan
bahwa salinitas akan menekan pembentukan spora FMA dan kolonisasi akar tanaman.
Penurunan tingkat salinitas tanah dari 7,0 mmho/cm menjadi 2,0 mmho/cm menghasilkan
peningkatan kepadatan spora antara 51-1.052 spora per 100 gram tanah dan persentase
kolonisasi sebesar 5-85%.
Beberapa peneliti melaporkan hasil identifikasi jenis FMA yang ditemukan di
lokasi yang mereka survey. Menurut Delvian (2003), jenis FMA yang kebanyakan
terdapat di tanah bergaram adalah Glomus spp. Menurut Ruiz-Lozano dan Azcon (1996),
dikemukakan bahwa FMA seperti Glomus spp mampu hidup dan berkembang di bawah
kondisi salinitas yang tinggi dan menunjukkan pengaruh yang nyata terhadap penurunan
kehilangan hasil karena salinitas. Namun menurut Al-Kariki (2000), mekanisme
perlindungannya belum diketahui dengan pasti, tetapi diduga disebabkan karena
meningkatnya serapan hara immobil seperti P, Zn, dan Cu. Delvian (2003), menyatakan
bahwa vegetasi Hutan Pantai Cagar Alam Leuweung Sancang Kabupaten Garut, Jawa
Barat mempunyai asosiasi yang tinggi dengan FMA, yaitu sekitar 95% dari tanaman yang
diobservasi mampu membentuk asosiasi mikoriza dengan persentase kolonisasi yang
BAB III
BAHAN DAN METODE
3.1. Tempat dan Waktu
Pengambilan sampel tanah dan anakan (akar) tanaman dilakukan di kawasan hutan
Pantai Sonang Kabupaten Tapanuli Tengah Sumatera Utara pada bulan April 2009.
Ekstraksi spora, identifikasi dan penghitungan kolonisasi FMA pada akar tanaman
sampel dilakukan di Laboratorium Biologi Tanah Fakultas Pertanian, Universitas
Sumatera Utara.
3.2. Bahan dan Alat
Untuk ekstraksi dan identifikasi spora FMA dibutuhkan bahan berupa tanah dan
akar tanaman dari hutan pantai, larutan glukosa 60%, larutan Melzer’s sebagai bahan
pewarna spora, larutan PVLG sebagai bahan pengawet spora. Sedangkan untuk
pewarnaan akar dibutuhkan bahan-bahan kimia antara lain; KOH 10%, HCL 2%, larutan
pewarna (Staining: lacto glycerol, asam laktat dan trypan blue), dan aquades.
Alat yang digunakan untuk pengambilan tanah dan akar tanaman sampel adalah tali
plastik, cangkul, kantung plastik, spidol, kertas label, gunting serta tabung film.
Sedangkan peralatan untuk pengamatan di laboratorium adalah saringan 710 µm, 425
µm, 125 µm, 45 µm, tabung sentrifuse, cawan petri, pinset spora, mikroskop binokuler,
3.3. Pelaksanaan Penelitian
Sebelum pengambilan sampel pertama sekali yang perlu dilakukan adalah
penetapan areal pengambilan sampel, kemudian dilanjutkan dengan pembuatan jalur.
Jalur dibuat sepanjang 200 m dengan lebar 5 m dari garis pantai menuju ke daratan. Jalur
dibagi dalam 10 petak dengan ukuran panjang setiap petak 20 m dan lebar 5 m. Jumlah
jalur yang dibuat sebanyak 3 jalur dengan jarak antar jalur sekitar 200 m. Teknik
pengambilan sampel tanah dan akar disajikan pada Lampiran 1.
Teknik pengambilan sampel tanah dan akar ini mengacu pada metode Kim dan
Weber (1985) dan Ragupathy dan Mahadevan (1991), yaitu metoda jalur atas dasar
gradien salinitas. Pada masing-masing petak dalam jalur diambil sampel tanah sebanyak
600–700 g dari zona rizosfir, yaitu pada kedalaman 0–20 cm. Selain itu juga diambil 3
jenis anakan yang dominan pada setiap petak ukur untuk mempelajari kolonisasi FMA
pada setiap petak ukur. Dari sampel tanah yang diambil juga dilakukan analisa kimia
untuk mengetahui tingkat salinitas tanah dan beberapa sifat kimia contoh tanah,
diantaranya N, P, pH dan C organik.
3.3.1. Ekstraksi dan Identifikasi Fungi Mikoriza Arbuskula
Ekstraksi spora FMA dilakukan untuk memisahkan spora FMA dari sampel tanah
sehingga dapat dilakukan identifikasi FMA guna mengetahui jumlah dan genus spora
FMA yang terdapat pada setiap petak sampel. Teknik yang digunakan dalam
mengekstraksi spora FMA adalah teknik tuang–saring dari Pacioni (1992) dan akan
dilanjutkan dengan teknik sentrifugasi dari Brundrett et al. (1996). Prosedur kerja teknik
tuang–saring adalah sebagai berikut: pertama mencampurkan tanah sampel sebanyak 50 g
disaring dalam satu set saringan dengan ukuran 710 µm, 425 µm, 125 µm dan 45 µm
secara berurutan dari atas ke bawah. Dari saringan bagian atas disemprot dengan air kran
untuk memudahkan bahan saringan lolos. Kemudian saringan paling atas dilepas dan
saringan kedua kembali disemprot dengan air kran. Setelah saringan kedua dilepas
sejumlah tanah sisa yang tertinggal pada saringan terbawah dipindahkan ke dalam tabung
sentrifuse.
Hasil saringan dalam tabung sentrifuse ditambahkan dengan glukosa 60% yang
diletakkan pada bagian bawah dari larutan tanah dengan menggunakan pipet. Tabung
sentrifuse ditutup rapat dan disentrifuse dengan kecepatan 2500 rpm selama 3 menit.
Selanjutnya cairan pada tabung sentrifuse dituang ke dalam saringan 45 µm dan
diusahakan supernatannya tidak ikut, lalu dicuci dengan air mengalir (air kran) untuk
menghilangkan glukosa. Endapan yang tersisa dalam saringan di atas dituangkan ke
dalam cawan petri dan kemudian diamati di bawah mikroskop binokuler untuk
penghitungan kepadatan spora dan pembuatan preparat guna identifikasi spora FMA yang
ada.
Bahan pewarna Melzer’s dan pengawet PVLG yang digunakan sebagai pembuat
preparat spora diletakkan secara terpisah pada satu kaca preparat. Spora-spora FMA yang
diperoleh dari ekstraksi setelah dihitung jumlah diletakkan dalam larutan Melzer’s dan
PVLG dan jenis spora FMA yang ada di kedua larutan ini sama. Selanjutnya spora-spora
tersebut dipecahkan secara hati-hati dengan cara menekan kaca penutup preparat
menggunakan ujung lidi. Perubahan warna spora dalam larutan Melzer’s adalah salah
satu indikator untuk menentukan tipe spora yang ada. Secara skematis alur kerja dalam
3.3.2. Kolonisasi FMA pada Akar Tanaman Sampel
Kolonisasi FMA pada akar ditandai dengan terdapatnya hifa, arbuskula dan
vesikula atau salah satu dari ketiganya pada akar sampel. Setiap bidang pandang
mikroskop yang menunjukkan tanda kolonisasi diberi simbol (+) dan yang tidak terdapat
tanda kolonisasi diberi simbol (-). Pengamatan kolonisasi FMA pada akar tanaman
sampel dilakukan melalui teknik pewarnaan akar (staining). Metoda yang digunakan
untuk pembersihan dan pewarnaan akar sampel adalah metoda dari Kormanik dan
McGraw (1982). Langkah pertama adalah memilih akar-akar halus dengan diameter
kurang dari 1 mm (Rajapakse dan Miller Jr., 1992) segar dan dicuci dengan air mengalir
hingga bersih.
Sampel akar yang telah dibersihkan, kemudian digunting dan dimasukkan kedalam
tabung film, lalu diberi larutan KOH 10% (sampai akar terendam) dan dibiarkan selama
lebih kurang 24 jam sehingga akar berwarna putih atau pucat. Tujuannya adalah untuk
mengeluarkan semua isi sitoplasma dari sel akar sehingga akan memudahkan pengamatan
struktur infeksi FMA. Larutan KOH kemudian dibuang dan akar contoh dicuci pada air
mengalir selama 5-10 menit. Selanjutnya akar sampel direndam dalam larutan HCl 2%
dan diinapkan selama satu malam berfungsi untuk mengasamkan akar, sehingga waktu
pewarnaan nantinya pengikatan terhadap warna lebih kuat. Larutan HCl 2% kemudian
dibuang dengan mengalirkannya secara perlahan-lahan. Selanjutnya akar sampel
direndam dalam larutan Trypan blue 0,05% selama 24 jam. Kemudian larutan Trypan
blue dibuang dan diganti dengan larutan lacto glycerol untuk proses destaining
Penghitungan persentase kolonisasi akar menggunakan metoda panjang akar
terkolonisasi (Giovannetti dan Mosse, 1980). Secara acak diambil potong-potongan akar
yang telah diwarnai dengan panjang ± 1 cm sebanyak 10 potongan akar dan disusun pada
kaca preparat, untuk setiap tanaman sampel dibuat dua preparat akar. Potongan-potongan
akar pada kaca preparat diamati untuk setiap bidang pandang. Secara skematis alur kerja
pengamatan kolonisasi fungi mikoriza pada akar tanaman sampel dapat dilihat pada
lampiran 3. Derajat/persentase kolonisasi akar dihitung dengan menggunakan rumus:
% kolonisasi akar =
n keseluruha pandang bidang da ber pandang bidang _ _ ) _( tan _ _ x 100%3.3.3. Pemerangkapan (Trapping)
Pemerangkapan ini dilakukan karena pada saat pengambilan sampel langsung
dilapangan biasanya mengandung banyak mikroorganisme yang tidak diinginkan dan
mungkin pada saat itu mikoriza belum sporulasi, sehingga dengan dilakukannya
pemerangkapan diharapkan nantinya penelitian ini memperoleh keanekaragaman spora
mikoriza yang baik dan dalam jumlah yang cukup. Teknik trapping yang digunakan
dalam penelitian ini mengikuti metoda Brundrett et al, (1994) dengan menggunakan pot
kultur terbuka.
1. Persiapan media tumbuh
Pasir dicuci sampai bersih guna menghilangkan kotoran yang ada. Pasir yang tidak
bersih berdampak negatif terhadap perkembangan FMA. Setelah itu pasir direndam
dalam larutan NaCl selama 24 jam.
2. Persiapan tanaman inang
dalam larutan chlorox 5% selama 5-10 menit sebagai upaya sterilisasi permukaan.
Kemudian direndam dalam air hangat selama ± 24 jam untuk memecahkan dormansi
yang mungkin terjadi. Selanjutnya benih-benih tersebut disemaikan dalam tempat
persemaian selama ± 10 hari atau telah muncul dua helai daun. Setelah itu dapat langsung
dilakukan penanaman.
3. Pembuatan kultur
Media tanam yang digunakan berupa campuran sampel tanah sebanyak ± 50 g dan
pasir sebanyak ±150 g. Cara pembuatan kultur sebagai media tanam adalah pot diisi
dengan pasir sampai setengah volume pot, kemudian dimasukkan sampel tanah dan
terakhir ditutup dengan pasir sehingga media tersusun atas pasir-sampel tanah-pasir. Dari
setiap sampel tanah dibuat 3 pot kultur sehingga terdapat 90 pot kultur.
Kegiatan berikutnya meliputi penyiraman, pemberian hara, dan pengendalian hama
secara manual. Dalam pemberian hara digunakan larutan Hyponex merah (25-5-20)
dengan konsentrasi 1 g/l. Pemberian larutan ini dilakukan setiap minggu sebanyak 20 ml
tiap pot kultur. Pemeliharaan tanaman dilakukan selama 2 bulan, selanjutnya tanaman
dibiarkan sampai mati. Pemanenan dilakukan untuk mengamati spora-spora baru.
Variabel-variabel yang diamati adalah perubahan tingkat salinitas tanah selama
pemeliharaan. Teknis pembuatan kultur trapping fungi mikoriza dapat dilihat pada
Lampiran 4
3.3.4. Pengamatan
Hasil yang dianalisa dengan metoda deskriptif kuantitatif, yaitu dengan menyajikan
tabel-tabel hasil identifikasi genus-genus FMA serta nama tanaman yang menjadi
diperoleh dihitung frekuensi mutlak (FM), frekuensi relatif (FR). Secara rinci rumus
tersebut adalah sebagai berikut:
1. Frekuensi Mutlak (FM - %)
Σ Petak ditemukan genus A
Frekuensi Mutlak (FM) = x 100%
Σ Petak keseluruhan
2. Frekuensi Relatif (FR – 100%)
Frekuensi genus A
BAB IV
HASIL DAN PEMBAHASAN
4.1. Sifat Kimia Tanah
Hasil analisis beberapa sifat kimia sampel tanah menunjukkan bahwa secara umum
semua tingkatan mempunyai variasi yang sangat kecil dalam hal sifat kimia kecuali
tingkat salinitas tanah. Hasil analisis beberapa sifat kimia sampel tanah disajikan pada
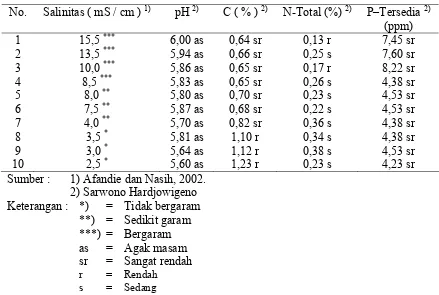
[image:41.612.83.522.302.597.2]Tabel 1.
Tabel 1. Hasil Analisis Beberapa Sifat Kimia Sampel Tanah
No. Salinitas ( mS / cm ) 1) pH 2) C ( % ) 2) N-Total (%) 2) P–Tersedia 2)
(ppm)
1 15,5 *** 6,00 as 0,64 sr 0,13 r 7,45 sr
2 13,5 *** 5,94 as 0,66 sr 0,25 s 7,60 sr
3 10,0 *** 5,86 as 0,65 sr 0,17 r 8,22 sr
4 8,5 *** 5,83 as 0,65 sr 0,26 s 4,38 sr
5 8,0 ** 5,80 as 0,70 sr 0,23 s 4,53 sr
6 7,5 ** 5,87 as 0,68 sr 0,22 s 4,53 sr
7 4,0 ** 5,70 as 0,82 sr 0,36 s 4,38 sr
8 3,5 * 5,81 as 1,10 r 0,34 s 4,38 sr
9 3,0 * 5,64 as 1,12 r 0,38 s 4,53 sr
10 2,5 * 5,60 as 1,23 r 0,23 s 4,23 sr
Sumber : 1) Afandie dan Nasih, 2002.
2) Sarwono Hardjowigeno Keterangan : *) = Tidak bergaram **) = Sedikit garam ***) = Bergaram
as = Agak masam
sr = Sangat rendah
r = Rendah
s = Sedang
Dari data yang diperoleh tampak bahwa tanah Hutan Pantai Sonang Tapanuli
Tengah dari nomor urut 1 sampai 7 merupakan tanah bergaram dengan nilai salinitas
berkisar antara 15,5–4,0, sedangkan tanah dari nomor urut 8 sampai 10 termasuk tanah
salinitasnya > 4 mS / cm dan dikatakan bebas garam jika tanah berada pada salinitas < 4
mS / cm.
Nilai salinitas suatu lokasi ditentukan oleh konsentrasi dari NaCl, CaCl2, KCl,
NaNO3, Ca(NO3)2, Na2SO4, Ca(SO4), K2SO4. Garam-garam ini dapat berasal dari
batuan induk, air irigasi atau air laut. Untuk daerah pantai sumber utama salinitas tanah
adalah air laut, dimana NaCl adalah penyusun utamanya (Chapman, 1975; Afandie dan
Nasih, 2002). Kandungan Na dan Cl dalam air laut menurut Carter (1975) masing-masing
adalah 30.61% dan 55,04%.
Hasil pengukuran pH tanah menunjukkan bahwa pH memiliki kisaran antara
5,60-6,00. Menurut Hardjowigeno (1987) tanah dengan kisaran 5,60–6,00 tergolong agak
masam. Dalam penelitian ini sulit diketahui pengaruh pH tanah terhadap keberadaan dan
keanekaragaman FMA karena tingkat keasaman pH tanah dari nomor urut 1 sampai
dengan 10 adalah sama, yaitu agak masam.
Kondisi pH mempengaruhi syarat tumbuh tanaman, sehingga mempengaruhi
pertumbuhan dan perkembangan FMA. Hal ini disebabkan karena FMA merupakan
simbion obligat sehingga semua faktor yang mempengaruhi tanaman inang juga
mempengaruhi FMA sebagai simbionnya. Artinya kondisi yang cukup bagi pertumbuhan
dan perkembangan inang memberikan pertumbuhan dan perkembangan terbaik bagi
FMA. Dalam penelitian ini belum dapat diketahui secara pasti apakah pH mempengaruhi
keberadaan dan keanekaragaman FMA, mengingat pH masih dalam kelompok yang sama
yaitu kelompok asam sedang. Barbara dan Daniels (1984), menyebutkan bahwa masih
disebabkan pH optimum untuk keberadaan FMA berbeda-beda untuk masing-masing
spesies FMA dan untuk lingkungan yang berbeda-beda pula.
Hasil analisis terhadap C organik pada sampel tanah menunjukkan bahwa tanah
pada daerah Pantai Sonang termasuk tanah kurang subur. Hal ini dapat dilihat dari nilai C
organik yang terdapat pada sampel tanah dengan nilai antara 0,64–1,23%. Menurut
Hardjowigeno (1987), tanah yang memiliki persen C-organik antara < 1,00 termasuk ke
dalam harkat sangat rendah dan C-organik 1,00 – 2,00 termasuk ke dalam harkat rendah.
Berdasarkan hasil analisis C organik menunjukkan bahwa peningkatan C organik
dari nomor urut 1 sampai 7 tergolong ke dalam harkat sangat rendah dan nomor urut 8
sampai dengan 10 tergolong ke dalam harkat rendah. Menurut Sutanto (2005) kandungan
bahan organik tanah biasanya diukur berdasarkan kandungan C organik. Kandungan
bahan organik ini dipengaruhi oleh akumulasi bahan asli dan dekomposisi yang sangat
tergantung pada kondisi lingkungan (vegetasi, iklim, batuan, praktek pertanian).
Meningkatnya C organik ini kemungkinan disebabkan oleh jumlah dan volume tanaman
yang digugurkan oleh tanaman, dimana sampai batas tertentu semakin banyak sisa
tanaman yang digugurkan, makin subur tanahnya. Dengan kata lain semakin rendah
tingkat salinitas, maka keadaan tanah semakin baik dalam mendukung pertumbuhan
tanaman.
Hasil analisis terhadap N pada sampel tanah Pantai Sonang berkisar antara
0,13-0,38 %. Menurut Hardjowigeno (1987) tergolong ke dalam tingkatan rendah sampai
sedang. Islami dan Wani (1995), menyatakan bahwa kondisi tanah yang dapat
mempengaruhi perkembangan mikoriza salah satunya adalah ketersediaan hara tanaman.
sampai 10 tergolong ke dalam tingkat sedang, sehingga sulit menentukan apakah N
mempengaruhi keberadaan dan keanekaragaman serta daya infeksi akar.
Hasil analisis terhadap P tersedia pada sampel tanah Pantai Sonang berkisar
4.23-8.22 ppm yang tergolong sangat rendah. Menurut Hardjowigeno (1987) bahwa P tersedia
< 10 ppm termasuk ke dalam golongan sangat rendah. Dari analisis ini sulit diketahui
apakah P tersedia mempengaruhi keberadaan dan keanekaragaman FMA serta daya
infeksi akar, mengingat dalam penelitian ini unsur P tersedia dalam semua tingkat nomor
urut masih dalam satu harkat, yaitu sangat rendah. Namun menurut Lynch (1983); Islami
dan Utomo (1995) bahwa infeksi akar berkurang ketika ketersediaan P meningkat di
tanah.
Husin et al. (2000) menyatakan bahwa kesuburan tanah (unsur N dan P tersedia),
kadar air, drainase tanah dan pH tanah berpengaruh terhadap perkembangan FMA. FMA
dapat berkembang dengan baik pada tanah yang mempunyai kandungan P lebih rendah
dan aerasi yang lebih baik. Pernyataan ini dibuktikan oleh Habte and Soedarjo (1996)
dalam penelitiannya yang menginokulasi FMA jenis Glomus aggregatum ke tanaman
Acacia mangium. Hasil penelitiannya diperoleh bahwa pada konsentrasi P tersedia yang
lebih rendah (0,002 me/100 g) dengan kisaran pH 4,3 – 6, inokulasi FMA memiliki
pengaruh yang lebih besar terhadap persen kolonisasi dan status P dalam jaringaan
tanaman, bila dibandingkan dengan inokulasi FMA pada kondisi pH yang sama namun
konsentrasi P yang lebih tinggi (0,008 me/100 g).
4.2. Kepadatan Spora
Hasil penghitungan kepadatan spora di lapangan (jumlah spora per 50 gram tanah)
bahwa kepadatan spora di lapangan meningkat sejalan dengan menurunnya kadar
salinitas tanah. Kepadatan spora terendah di lapangan ditemukan pada tingkat salinitas
15,5 mS/cm (10 spora) dan tertinggi pada tingkat salinitas 2,5 mS/cm (129 spora).
10 17 23 30 36 60 78 84 97 129 15.5 13.5 10.0 8.5 8.0 7.5 4.0 3.5 3.0 2.5 0 20 40 60 80 100 120 140 K epa da ta n S p ora ( pe
r 50 g t
an ah ) 0 2 4 6 8 10 12 14 16 18 T in g k at S al in it as ( m S /c m )
Kepadatan Spora (per 50 g tanah) Tingkat Salinitas
Gambar 2 : Jumlah Spora di Lapangan Berdasarkan Tingkat Salinitas Tanah
Dari data di atas dapat dikatakan bahwa tingginya salinitas tanah ini berpengaruh
negatif terhadap kepadatan spora FMA. Terjadinya penurunan salinitas tanah
menghasilkan peningkatan kepadatan spora FMA sepanjang jalur pengamatan. Hasil ini
sejalan dengan penelitian yang dilakukan oleh Ragupathy dan Mahadevan (1991) yang
mempelajari pengaruh gradien salinitas terhadap distribusi FMA di Kodikkarai Reserve
Forest, Tamil Nadu, India dan secara tegas menyatakan bahwa salinitas menekan
pembentukan spora FMA. Pada hasil penelitiannya ini diperoleh data bahwa penurunan
tingkat salinitas tanah dari 7,0 mmho/cm menjadi 2,0 mmho/cm menghasilkan
peningkatan kepadatan spora antara 51–1.052 spora per 100 gram tanah. Hasil penelitian
keanekaragaman cendawan mikoriza arbuskula di Hutan Pantai Cagar Alam Leuweung
Sancang, Pameungpeuk Kabupaten Garut. Hasil penelitiannya menyebutkan bahwa
kepadatan spora meningkat sejalan dengan menurunnya salinitas tanah. Secara umum
Brundrett et al. (1996) menyimpulkan bahwa salinitas merupakan salah satu faktor tanah
yang menyebabkan kurangnya spora FMA di tanah disamping faktor pH tanah,
kekeringan, pencucian, atau iklim yang ekstrim, dan kehilangan lapisan tanah bagian
atas, atau kurangnya tanaman inang.
Peningkatan jumlah spora yang selalu diikuti dengan turunnya tingkat salinitas
tanah ini disebabkan oleh faktor lingkungan yaitu tingkat salinitas tanah dan tanaman
inang (Johnson-Green et al., 1995; Siguenza et al., 1996). Dimana salinitas tanah dapat
mempengaruhi setiap tahapan perkembangan FMA, mulai dari perkecambahan spora
(Hirrel, 1981) sampai tahapan pembentukan spora baru (Pacioni, 1986). Penundaan atau
penghambatan fase perkecambahan spora akibat tingginya konsentrasi garam terlarut
dalam larutan tanah menunda atau menghambat pertumbuhan hifa sehingga pada
akhirnya menunda kolonisasi akar tanah dan pembentukan simbiosis (Juniper dan Abbott,
1993). Penundaan itu akhirnya akan mempengaruhi proses pembentukan spora dan
jumlah spora yang dihasilkan.
Kondisi salinitas tanah juga sangat mempengaruhi tingkat ketersediaan air bagi
pertumbuhan dan perkembangan tanaman yang pada akhirnya juga mempengaruhi
perkembangan FMA yang terdapat pada perakaran tanaman sebagai simbionnya.
Terdapat beberapa penelitian tentang pengaruh ketersediaan air terhadap sporulasi FMA,
antara lain Sieverding dan Toro (1988) yang melaporkan bahwa, sporulasi sejumlah FMA
pertumbuhan tanaman Cassava yang diinokulasi dengan 7 jenis FMA yang berbeda.
Terdapat 2 perlakuan air: yaitu basah, dimana tanah dipertahankan pada kapasitas
lapangan ( kadar air 33%), dan kering, dimana pot diairi sampai kapasitas lapangan, lalu
kadar airnya menurun sampai 15% dan kemudian diairi kembali sampai kapasitas lapang.
Jumlah total spora yang terbentuk dari semua jenis FMA yang digunakan secara
signifikan akan menurun pada kondisi kering, kecuali pada Scutellospora heterogama
yang relatif meningkat dengan perlakuan pengeringan. Hal ini terjadi karena perlakuan
kering akan menurunkan produksi bahan kering tanaman, maka produksi spora FMA juga
akan menurun.
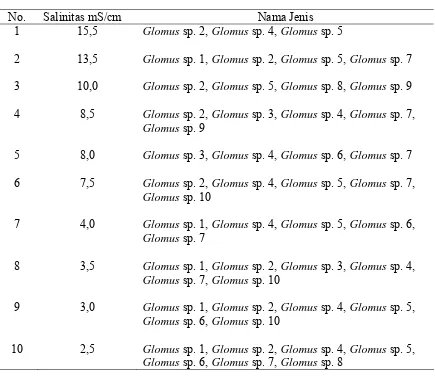
Selain mempengaruhi kepadatan spora, tingkat salinitas tanah juga mempengaruhi
keanekaragaman spora FMA yang ditemukan (Tabel 2). Hasil identifikasi di lapangan
menunjukkan bahwa secara umum penurunan tingkat salinitas tanah menghasilkan
peningkatan keanekaragaman spora FMA, dimana jenis Glomus sp. 2 dan Glomus sp. 4
merupakan jenis spora yang mempunyai daerah penyebaran pada tingkat salinitas yang
lebih luas. Berdasarkan pengamatan spora di lapangan dijumpai 10 jenis spora FMA,
yang mana kesemuanya adalah Glomus sp. Jenis-jenis lainnya seperti Acaulospora sp.,
Gigaspora sp., Scutellospora sp dan Enthrospora sp tidak dijumpai pada pengamatan
Tabel 2. Keanekaragaman spora FMA yang Ditemukan dari Lapangan Berdasarkan Tingkat Salinitas Tanah.
No. Salinitas mS/cm Nama Jenis
1 2 3 4 5 6 7 8 9 10 15,5 13,5 10,0 8,5 8,0 7,5 4,0 3,5 3,0 2,5
Glomus sp. 2, Glomus sp. 4, Glomus sp. 5
Glomus sp. 1, Glomus sp. 2, Glomus sp. 5, Glomus sp. 7
Glomus sp. 2, Glomus sp. 5, Glomus sp. 8, Glomus sp. 9
Glomus sp. 2, Glomus sp. 3, Glomus sp. 4, Glomus sp. 7,
Glomus sp. 9
Glomus sp. 3, Glomus sp. 4, Glomus sp. 6, Glomus sp. 7
Glomus sp. 2, Glomus sp. 4, Glomus sp. 5, Glomus sp. 7,
Glomus sp. 10
Glomus sp. 1, Glomus sp. 4, Glomus sp. 5, Glomus sp. 6,
Glomus sp. 7
Glomus sp. 1, Glomus sp. 2, Glomus sp. 3, Glomus sp. 4,
Glomus sp. 7, Glomus sp. 10
Glomus sp. 1, Glomus sp. 2, Glomus sp. 4, Glomus sp. 5,
Glomus sp. 6, Glomus sp. 10
Glomus sp. 1, Glomus sp. 2, Glomus sp. 4, Glomus sp. 5,
Glomus sp. 6, Glomus sp. 7, Glomus sp. 8
Penurunan jumlah jenis spora FMA pada tingkat salinitas tanah yang tinggi, diduga
berhubungan dengan toleransi setiap jenis FMA terhadap tingkat salinitas tanah (Flower
et al., 1997). Toleransi FMA terhadap salinitas tanah ditentukan oleh kemampuan setiap
jenis untuk melakukan osmoregulasi atau penyesuaian osmotik agar potensial osmotik
dalam sel FMA lebih rendah dari pada larutan tanah.
Hasil pengamatan dari kultur pemerangkapan (trapping) menunjukkan bahwa
kepadatan dan keanekaragaman spora meningkat dibandingkan spora di lapangan,
mempunyai daerah penyebaran pada tingkat salinitas yang lebih luas. Hasil penghitungan
kepadatan spora dan keanekaragaman spora hasil trapping (jumlah dan keanekaragaman
spora per 50 gram tanah) berdasarkan tingkat salinitas tanah dapat dilihat pada Gambar 3
dan Tabel 3.
15 23 37 54 74 106 125 131 145 177 15,5 13,5 10,0 8,5 8,0 7,5 4,0 3,5 3,0 2,5 0 20 40 60 80 100 120 140 160 180 200 K ep ada ta n S por a ( p er 50 g ta na h ) 0 2 4 6 8 10 12 14 16 18 T ingk at S al ini ta s ( m S /c m )
Kepadatan Spora (per 50 g tanah) Tingkat Salinitas
Gambar 3 : Jumlah Spora Hasil Trapping Berdasarkan Tingkat Salinitas Tanah
Dari Gambar 3 di atas tampak bahwa kepadatan spora hasil trapping meningkat
sejalan dengan menurunnya kadar salinitas tanah. Hasil trapping menunjukkan bahwa
jumlah kepadatan spora terendah hasil trapping ditemukan pada tingkat salinitas 15,5
mS/cm (15 spora / 50 g tanah) dan tertinggi pada tingkat salinitas 2,5 mS/cm (177 spora /
Tabel 3. Keanekaragaman spora FMA Hasil Trapping Berdasarkan Tingkat Salinitas tanah
No. Salinitas Nama Jenis
1 2 3 4 5 6 7 8 9 10 15,5 13,5 10,0 8,5 8,0 7,5 4,0 3,5 3,0 2,5
Glomus sp. 2, Glomus sp. 4, Glomus sp. 5, Glomus sp. 17, Glomus sp. 26
Glomus sp. 1, Glomus sp. 2, Glomus sp. 5, Glomus sp. 7, Glomus sp. 26, Acaulospora sp. 2
Glomus sp. 2, Glomus sp. 5, Glomus sp. 8, Glomus sp. 9, Glomus sp. 21, Glomus sp. 22
Glomus sp. 2, Glomus sp. 3, Glomus sp. 4, Glomus sp. 7, Glomus sp. 9, Glomus sp. 24
Glomus sp. 3, Glomus sp. 4, Glomus sp. 6, Glomus sp. 7, Glomus sp. 18, Glomus sp. 25
Glomus sp. 2, Glomus sp. 4, Glomus sp. 5, Glomus sp. 7, Glomus sp. 10, Glomus sp. 18, Glomus sp. 24
Glomus sp. 1, Glomus sp. 4, Glomus sp. 5, Glomus sp. 6, Glomus sp. 7, Glomus sp. 13, Glomus sp. 18, Glomus
sp. 19, Glomus sp. 20, Acaulospora sp. 1
Glomus sp. 1, Glomus sp. 2, Glomus sp. 3, Glomus sp. 4, Glomus sp. 7, Glomus sp. 10, Glomus sp. 14, Glomus
sp. 15, Glomus sp. 16, Glomus sp. 18, Glomus sp. 21,
Acaulospora sp. 1
Glomus sp. 1, Glomus sp. 2, Glomus sp. 4, Glomus sp. 5, Glomus sp. 6, Glomus sp. 10, Glomus sp. 11, Glomus
sp. 12, Glomus sp. 17, Glomus sp. 18, Glomus sp. 26,
Acaulospora sp. 1
Glomus sp. 1, Glomus sp. 2, Glomus sp. 4, Glomus sp. 5, Glomus sp. 6, Glomus sp. 7, Glomus sp. 8, Glomus
sp. 18, Glomus sp. 20, Glomus sp. 23, Glomus sp. 24,
Meningkatnya jumlah dan keanekaragaman spora hasil trapping diduga karena
FMA dari lapangan banyak yang belum bersporulasi sehingga dengan dilakukannya
trapping, diharapkan FMA yang ada akan bersporulasi sehingga akan terbentuk
keanekaragaman dan jumlah FMA yang lebih banyak serta data yang diperoleh juga lebih
akurat. Menurut Delvian (2006 b) pada kondisi basah atau banyak hujan umumnya
persentase kolonisasi meningkat dan pembentukan spora baru berkurang. Hal ini
disebabkan karena kelembaban tanah yang tinggi pada kondisi basah akan merangsang
keberadaan dan keanekaragaman FMA serta terbentuknya kolonisasi dengan tanaman
inang. Sebaliknya pada kondisi kering atau sedikit hujan pembentukan spora baru akan
meningkat dan persentase kolonisasi akan menurun. Kondisi kering akan merangsang
pembentukan spora yang banyak sebagai respon alami dari FMA serta upaya untuk
mempertahankan keberadaannya di alam.
Pada identifikasi hasil trapping dijumpai 28 jenis spora FMA. Dua puluh enam jenis
spora diantaranya merupakan genus Glomus sp dan 2 jenis spora merupakan genus
Acaulospora sp. Jenis-jenis mikoriza lainnya seperti Gigaspora sp., Scutellospora sp dan
Enthrospora sp tidak dijumpai pada pengamatan tipe spora hasil trapping. Identifikasi ini
dilakukan berdasarkan perbedaan ciri, karakteristik morfologi (bentuk ketebalan dinding
sel, ada tidaknya sublending hifa, kehalusan permukaan) dan reaksi spora terhadap
pewarna melzers.
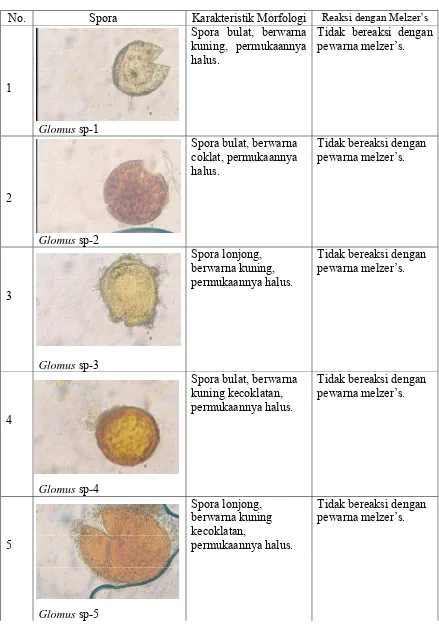
Keanekaragaman karakteristik spora FMA yang ditemukan di lapangan ditampilkan
pada Tabel 4 dan keanekaragaman karakteristik spora FMA yang ditemukan hasil
Tabel 4. Keanekaragaman dan karakteristik spora FMA yang ditemukan di lapangan
No. Spora Karakteristik Morfologi Reaksi dengan Melzer’s
1
Glomus sp-1
Spora bulat, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
2
Glomus sp-2
Spora bulat, berwarna coklat, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
3
Glomus sp-3
Spora lonjong, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
4
Glomus sp-4
Spora bulat, berwarna kuning kecoklatan, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
5
Spora lonjong, berwarna kuning kecoklatan,
permukaannya halus.
No. Spora Karakteristik Morfologi Reaksi dengan Melzer’s
6
Glomus sp-6
Spora bulat, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
7
Glomus sp-7
Spora bulat, berwarna kuning, permukaannya agak kasar.
Tidak bereaksi dengan pewarna melzer’s.
8
Glomus sp-8
Spora bulat, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
9
Glomus sp-9
Spora bulat, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
10
Glomus sp-10
Spora bulat, berwarna kuning, permukaannya halus.
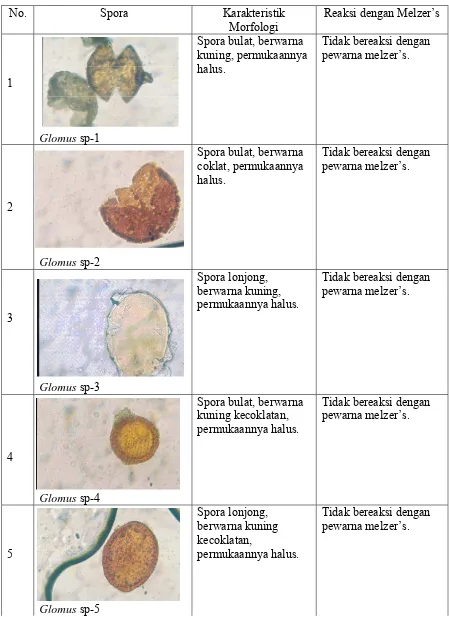
Tabel 5. Keanekaragaman dan karakteristik spora FMA hasil trapping
No. Spora Karakteristik
Morfologi
Reaksi dengan Melzer’s
1
Glomus sp-1
Spora bulat, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
2
Glomus sp-2
Spora bulat, berwarna coklat, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
3
Glomus sp-3
Spora lonjong, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
4
Glomus sp-4
Spora bulat, berwarna kuning kecoklatan, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
5
Spora lonjong, berwarna kuning kecoklatan,
permukaannya halus.
No. Spora Karakteristik Morfologi
Reaksi dengan Melzer’s
6
Glomus sp-6
Spora bulat, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
7
Glomus sp-7
Spora bulat, berwarna kuning, permukaannya agak kasar.
Tidak bereaksi dengan pewarna melzer’s.
8
Glomus sp-8
Spora bulat, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
9
Glomus sp-9
Spora bulat, berwarna kuning, permukaannya halus.
Tidak bereaksi dengan pewarna melzer’s.
10
Glomus sp-10
Spora bulat, berwarna kuning, permukaannya halus. <