KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA
(FMA) DI BAWAH TEGAKAN TANAMAN JABON
(Anthocephalus cadamba Miq.) DI PEMALANG JAWA
TENGAH
TRIA AMELIA
DEPARTEMEN SILVIKULTUR FAKULTAS KEHUTANAN INSTITUT PERTANIAN BOGOR
BOGOR 2013
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul: Keanekaragaman Fungi Mikoriza Arbuskula (FMA) di bawah Tegakan Tanaman Jabon (Anthocephalus cadamba Miq.) di Pemalang Jawa Tengah adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Oktober 2013
Tria Amelia E44090023
ABSTRAK
TRIA AMELIA. Keanekaragaman Fungi Mikoriza Arbuskula (FMA) di Bawah Tegakan Tanaman Jabon (Anthocephalus cadamba Miq.) di Pemalang Jawa Tengah. Dibimbing oleh SRI WILARSO BUDI R.
Mikoriza merupakan suatu bentuk simbiosis mutualisme antara fungi dan akar tanaman. FMA memiliki berbagai fungsi, salah satunya adalah peningkatan penyerapan hara terutama P. Sampel akar dan tanah yang digunakan dalam penelitian ini diambil dari bawah tegakan tanaman jabon Pemalang, di daerah pantai dan pegunungan. Pengamatan kolonisasi FMA pada akar tanaman dilakukan melalui teknik pewarnaan akar, sedangkan isolasi spora dilakukan dengan menggunakan metode tuang basah. Spora diidentifikasi dengan melihat bentuk morfologi dan reaksi spora setelah ditetesi larutan Melzer. Hasil pengamatan menunjukkan bahwa rata-rata persentase infeksi akar terbesar terdapat pada lokasi PML 2.2 sebesar 73.33% sedangkan rata-rata persentase infeksi akar terendah terdapat pada lokasi PML 2.5 yaitu 46.89%. Dalam penelitian ini ditemukan 3 genus spora yaitu Glomus, Acaulospora dan Scutellospora. Jenis Glomus merupakan yang paling dominan dan terdapat pada setiap lokasi. Kepadatan populasi spora tertinggi yaitu di lokasi PNK 1. Pertumbuhan tanaman jabon di daerah pantai lebih baik dibandingkan dengan daerah pegunungan.
Kata kunci: fungi mikoriza arbukula, kolonisasi, keanekaragaman, kepadatan spora
ABSTRACT
TRIA AMELIA. Diversity of Arbuscular Mycorrhizal Fungi (AMF) under a stand of Jabon Plants (Anthocephalus cadamba Miq.) in Pemalang, Central Java. Supervised by SRI WILARSO BUDI R.
Mycorrhiza is a form of symbiosis mutualism between fungi and the roots of plants. Arbuscular mycorrhizal fungi (AMF) has some functions, one of them is increasing nutrients absorption, especially P. The sample of root and soil used in this research taken from a stand of jabon plants in Pemalang, at the coastal and mountainous area. Observation colonization on plant roots carried out through staining roots and isolation of spores from soil samples is done by using the wet sieving and decanting method. Spores were identified by looking at the spores reaction after etched with a Melzer solution. The result of the research show that the biggest average of root colonization, is found at PML 2.2 is 73.33%. Where as the lowest average of root colonization is found at PML 2.5, is 46.89. In this research is found 3 type of Genus that is Glomus, Acaulospora and Scutellospora. Glomus is the most dominant and are found in all location. The highest density of spores population is at PNK 1. The growth of plants in a coastal area better than a mountainous region.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
pada
Departemen Silvikultur
KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA
(FMA) DI BAWAH TEGAKAN TANAMAN JABON
(Anthocephalus cadamba Miq.) DI PEMALANG JAWA
TENGAH
TRIA AMELIA
DEPARTEMEN SILVIKULTUR FAKULTAS KEHUTANAN INSTITUT PERTANIAN BOGOR
BOGOR 2013
Judul Skripsi : Keanekaragaman Fungi Mikoriza Arbuskula (FMA) di bawah Tegakan Tanaman Jabon (Anthocephalus cadamba Miq.) di Pemalang Jawa Tengah
Nama : Tria Amelia NIM : E44090023
Disetujui oleh
Dr Ir Sri Wilarso Budi R, MS Dosen Pembimbing
Diketahui oleh
Prof Dr Ir Nurheni Wijayanto, MS Ketua Departemen Silvikultur
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala nikmat dan karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan November 2012 ini ialah mengenai fungi mikoriza arbuskula, dengan judul Keanekaragaman Fungi Mikoriza Arbuskula (FMA) di bawah Tegakan Tanaman Jabon (Anthocephalus cadamba Miq.) di Pemalang Jawa Tengah.
Terima kasih penulis ucapkan kepada Bapak Dr Ir Sri Wilarso Budi R, MS. selaku pembimbing yang telah banyak memberi saran. Di samping itu, ungkapan terima kasih juga disampaikan kepada ayah, ibu, serta seluruh keluarga, atas segala doa dan kasih sayangnya, kepada sahabat dan teman yang selalu memberi dukungan dan semangat sehingga karya ilmiah ini berhasil diselesaikan.
Semoga karya ilmiah ini bermanfaat.
Bogor, Oktober 2013 Tria Amelia
DAFTAR ISI
DAFTAR TABEL iv DAFTAR GAMBAR v PENDAHULUAN 1 Latar Belakang 1 Tujuan Penelitian 2 Manfaat Penelitian 2 METODE 2Waktu dan Tempat 2
Bahan Penelitian 2
Alat Penelitian 2
Prosedur Penelitian 3
Analisis Data 4
HASIL DAN PEMBAHASAN 5
Sifat Fisik dan Kimia Tanah di Bawah Tegakan Jabon Pemalang 5
Persentase Infeksi Akar 6
Kepadatan Spora 9
Keanekaragaman Spora 10
Kelimpahan Spora 12
Frekuensi Spora 13
Rata-rata Diameter dan Tinggi Tanaman Jabon di Pantai dan Pegunungan 14
SIMPULAN DAN SARAN 15
DAFTAR PUSTAKA 15
DAFTAR TABEL
1 Karakteristik sifat fisik dan kimia tanah 5
2 Persentase infeksi akar pada jabon 7
3 Hasil analisis regresi antara sifat kimia tanah dengan kepadatan spora 9 4 Keragaman spora di bawah tegakan jabon Pemalang 11
DAFTAR GAMBAR
1 Penampang akar jabon yang terinfeksi 6
2 Rata-rata persentasi infeksi FMA pada akar jabon untuk setiap lokasi 7
3 Kepadatan spora tiap lokasi 10
4 Glomus sp. 11
5 Acaulospora sp. 12
6 Scutellosporasp. 12
7 Kelimpahan spora FMA di bawah tegakan jabon Pemalang 13 8 Frekuensi sebaran genus spora FMA di bawah tegakan jabon Pemalang 14 9 Rata-rata tinggi tanaman jabon di pantai dan pegunungan 14 10 Rata-rata diameter tanaman jabon di pantai dan pegunungan 15
DAFTAR LAMPIRAN
1 Kriteria presentase kolonisasi akar 18
1
PENDAHULUAN
Latar Belakang
Mikrobiologi di dalam tanah merupakan suatu hal yang perlu dipelajari karena dalam substansi penyusunnya ada yang bersifat patogen dan ada pula yang bersifat komersial. Mikrobiologi terdiri dari berbagai macam jenis, di antaranya adalah bakteri, aktinomisetes, fungi, alga, dan protozoa. Salah satu di antaranya sebagian besar mempengaruhi interaksi biologi di alam dan biasanya terdapat di bawah tegakan pohon-pohon hutan. Asosiasi yang terdapat pada struktur tanaman tingkat tinggi dikenal dengan istilah mikoriza atau disebut juga dengan istilah fungi akar. Mikoriza dapat ditemukan langsung di bawah akar pohon tingkat tinggi, contohnya jabon. Jabon (Anthocephalus cadamba) merupakan salah satu jenis tanaman yang dapat berasosiasi dengan fungi mikoriza arbuskula.
Mikoriza merupakan suatu bentuk simbiosis mutualistik antara fungi dan akar tanaman. Hampir pada semua jenis tanaman terdapat bentuk simbiosis ini. Umumnya mikoriza dibedakan dalam tiga kelompok, yaitu: endomikoriza, ektomikoriza dan ektendomikoriza. Peranan penting FMA dalam pertumbuhan tanaman adalah kemampuannya untuk menyerap unsur hara baik makro maupun mikro. Selain itu akar yang mempunyai mikoriza dapat menyerap unsur hara dalam bentuk terikat dan yang tidak tersedia bagi tanaman. Hifa eksternal pada mikoriza dapat menyerap unsur fosfat dari dalam tanah dan segera diubah menjadi senyawa polifosfat.
Menurut Santoso et al. (2007) mikoriza sangat penting dalam kehutanan karena dapat meningkatkan kegiatan rehabilitasi hutan dan lahan terdegradasi. Akar yang diinfeksi oleh mikoriza ini lebih sering berasimilasi dengan fosfat dan pohon dapat tumbuh baik tanpa terjadi defisiensi fosfor. Selain itu, tanaman kehutanan yang bermikoriza mempunyai beberapa keunggulan jika dibandingkan dengan tanaman yang tidak bermikoriza, di antaranya yaitu tahan terhadap serangan patogen akar, tahan terhadap kekeringan, pemanenan unsur hara optimal dan pertumbuhan anakan akan jauh lebih cepat (Fakuara 1988).
Penggalian potensi keanekaragaman mikoriza dalam suatu tipe ekosistem sangat diperlukan karena peranan mikoriza dalam kestabilan ekosistem tersebut. Berdasarkan hasil review Wang and Qiu (2006) ditemukan sekitar 2.469 dari 3.617 jenis tanaman yang berasosiasi dengan FMA, 55 jenis di antaranya adalah jenis yang termasuk dalam famili Rubiaceae. Tanaman jabon yang termasuk dalam famili Rubiaceae tidak termasuk dalam 55 jenis yang sudah diketahui asosiasi dan respon pertumbuhannya terhadap inokulasi FMA. Hal ini menunjukan bahwa informasi penelitian internasional maupun nasional tentang asosiasi simbiotik FMA dengan jenis tersebut masih kurang, khususnya penelitian tentang keanekaragaman fungi mikoriza di bawah tegakan jabon masih sangat sedikit. Oleh karena itu dilakukan penelitian mengenai keanekaragaman fungi mikoriza arbuskula di bawah tegakan jabon guna untuk pengelolaan dan pemanfaatan yang lebih baik ke depannya.
2
Tujuan Penelitian
Penelitian ini bertujuan untuk menganalisis keberadaan dan keanekaragaman fungi mikoriza arbuskula di bawah tegakan jabon di Pemalang Jawa Tengah yang berpotensi bagi pertumbuhan semai jabon.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat dijadikan sebagai sumber informasi mengenai keberadaan dan keanekaragaman jenis fungi mikoriza arbuskula di bawah tegakan jabon di Pemalang Jawa Tengah yang dapat digunakan untuk pengelolaan dan pemanfaatan selanjutnya.
METODE
Waktu dan Tempat
Penelitian ini berlangsung selama 6 bulan yaitu dari bulan November 2012 sampai dengan Februari 2013 dan dilanjutkan bulan Mei 2013 sampai dengan Juni 2013. Analisis akar dilaksanakan di Laboratorium Silvikultur, Departemen Silvikultur, Fakultas Kehutanan Institut Pertanian Bogor. Pengamatan spora dilaksanakan di Laboratorium Bioteknologi Hutan PPSHB IPB dan Laboratorium Mikrobiologi Tanah Pusat Penelitian dan Pengembangan Sumber Daya Hutan Kementrian Kehutanan RI di Bogor serta pengambilan sample tanah dan akar dilakukan di Pemalang Jawa Tengah.
Bahan
Bahan yang digunakan dalam penelitian ini adalah contoh tanah dan akar tanaman dari bawah tegakan jabon Pemalang Jawa Tengah, larutan glukosa 60%. KOH 2.5%, HCL 0.1N, larutan trypan blue 0.02%, gliserin 50%, H2O2, larutan
Melzer dan aquades.
Alat
Alat-alat yang digunakan adalah mikroskop stereo, cawan petri, kantong plastik, spido,. kertas label, saringan spora (saringan bertingkat tiga yaitu 250 μm, 126 μm dan 63 μm), sentrifus, pinset spora, mikroskop binokuler, kaca preparat, cover glass, tabung film, tabung gelas, kuteks, pipet, timbangan, gunting akar.
3
Prosedur Penelitian Pengambilan Sample Tanah
Contoh tanah yang diambil yaitu dari bawah tegakan tanaman jabon yang tersebar di beberapa lokasi dengan ketinggian dan umur yang berbeda. Lokasi pertama yaitu di daerah pantai dengan umur tanaman 2 tahun, 2.2 tahun dan 2.5 tahun. Kemudian lokasi kedua yaitu daerah pegunungan dengan umur tanaman 2 tahun dan 1 tahun. Tanah diambil dari bawah tanaman jabon secara komposit dari empat sisi yang berbeda. Setiap lokasi terdiri atas 3 plot. Dari masing-masing plot diwakili oleh 3 tanaman jabon yang dipilih secara acak. Selain contoh tanah juga dilakukan pengambilan contoh akar tanaman.
Analisis Infeksi akar
Pengamatan kolonisasi FMA pada akar tanaman dilakukan melalui teknik pewarnaan akar (staining). Pewarnaan akar dilakukan dengan metode Brundrett et al. (1996). Tahapan pewarnaan tersebut ialah sebagai berikut: akar serabut yang berasal dari lapangan dipisahkan dari tanah kemudian dimasukkan ke dalam botol film. Setelah itu akar direndam dengan alkohol 70% kemudian dibilas dengan air sampai bersih. Lalu akar direndam dengan KOH 2.5% dan dimasukkan ke dalam oven dengan suhu ±90 ⁰C selama 30-40 menit. Perendaman dengan KOH dilakukan untuk mengeluarkan semua isi sitoplasma dari sel akar sehingga akan memudahkan pengamatan struktur infeksi FMA. Setelah selesai dipanaskan, akar dikeluarkan dan dicuci dengan air. Jika akar belum bening, maka dapat ditambahkan H2O2 kemudian dikocok sampai akar berwarna bening, tetapi jika
akar berwarna bening maka dapat langsung direndam dengan HCl. Akar direndam dengan HCl 0.1N selama ±10 menit. Setelah itu HCl diganti dengan trypan blue kemudian akar dimasukkan kembali ke dalam oven selama 30-40 menit dengan suhu ±90 ⁰C. Setelah itu trypan blue dibuang, kemudian akar dicuci dan akar direndam dengan larutan gliserin 50% untuk proses pengurangan warna. Akar siap diamati. Untuk pengamatan akar, dilakukan dengan memotong akar yang telah diwarnai sepanjang 1 cm, kemudian akar ditata di atas preparat dan ditutup dengan cover glass. Jumlah akar tiap preparat sebanyak 15 potong. Infeksi akar dapat dilihat melalui adanya vesikula, arbuskula, hifa maupun spora yang menginfeksi akar.
Isolasi Spora
Isolasi spora FMA dilakukan dengan menggunakan metoda tuang basah menurut Gerdermann dan Nickolson (1963), dilanjutkan dengan metode sentrifugasi menurut Brundrett et al. (1996). Untuk isolasi spora tanah yang digunakan masing-masing sebanyak 10 gram. Prosedur kerjanya adalah sebagai berikut: tanah yang sudah ditimbang sebanyak 10 gram dimasukkan dalam gelas kemudian ditambahkan ± 2/3 air. Larutan tersebut diaduk hingga rata sampai tanah hancur dan didiamkan ±1 menit. Tanah yang sudah diaduk dituangkan di atas saringan spora tiga tingkat dengan diameter lubang 250 μm, 125 μm, dan 63 μm (ketika proses penyaringan berlangsung air kran harus terus mengalir). Penyaringan dilakukan sebanyak 3–5 kali ulangan dengan prosedur yang sama. Setelah penyaringan dilakukan. tanah yang menempel pada saringan yang berukuran 125 μm dan 63 μm dikumpulkan di satu ujung (saringan dimiringkan).
4
Tanah-tanah tersebut dituangkan ke dalam tabung sentrifuge. Kemudian ditambahkan larutan glukosa 60% sampai tabung terisi 2/3 nya. Kocok terlebih dahulu tabung sentrifus sebelum dimasukkan ke dalam mesin sentrifus. Sentrifugasi dilakukan selama 1 menit dengan kecepatan 2500 rpm. Setelah itu larutan tersebut dituangkan pada saringan yang berukuran 63 μm. Larutan dicuci dengan air mengalir yang berfungsi untuk menghilangkan larutan gula yang masih tertinggal. Spora yang menempel pada saringan dipindahkan ke dalam cawan petri untuk diidentifikasi. Spora siap diamati dibawah mikroskop untuk dihitung jumlah sporanya.
Identifikasi Spora FMA
Spora yang didapat dari contoh tanah yang diambil diidentifikasi dengan melihat bentuk morfologi berdasarkan INVAM (2013) dan reaksi spora setelah ditetesi larutan Melzer. Identifikasi spora berupa penentuan genus spora. Spora diletakkan di preparat, lalu ditetesi dengan larutan Melzer. Spora dipecahkan untuk melihat reaksi antara lipid yang ada di dalam spora dengan larutan Melzer. Pengamatan dilakukan dengan menggunakan rumus sebagai berikut:
Kepadatan spora = Jumlah spora 10 gram tanah
Kekayaan spora = Jumlah genus pada 10 gram tanah Kelimpahan Relatif = Jumlah genus x 100%
total spora
Frekuensi = Jumlah sampel ditemukan spora x 100% total sample
Infeksi Mikoriza
Perhitungan persentase infeksi akar dilakukan dengan meletakan 15 potongan akar dengan panjang masing-masing 1 cm pada kaca objek dan ditutup dengan cover glass, kemudian diamati di bawah mikroskop. Perhitungan infeksi akar dapat menggunakan rumus (Giovannety dan Mosse 1980) :
∑ ∑
Analisis Data
Analisis data dilakukan secara deskriptif dan pengujian beda nilai tengah untuk ragam populasi tidak diketahui pada taraf kepercayaan 95% dengan keputusan uji apabila thitung jatuh pada wilayah kritik maka H0 ditolak (terima H1).
Pengolahan data analisis regresi sifat kimia tanah dilakukan dengan menggunakan program komputer minitab16.
5
HASIL DAN PEMBAHASAN
Sifat Fisik dan Kimia Tanah di Bawah Tegakan Jabon
Keberadaan mikoriza dalam tanah dapat mempengaruhi sifat fisik dan kimia tanah. Data sifat fisik dan kimia tanah di bawah tegakan tanaman jabon Pemalang Jawa Tengah dapat dilihat pada Tabel 1.
Tabel 1 Karakteristik sifat fisik dan kimia tanah
Lokasi pH N-total (%) C (%) C/N P-tersedia (ppm) Tekstur H2O KCl PNK 1 5.7 5.1 0.43 4.27 10 4 lempung berpasir PNK 2 5.6 5.1 0.41 4.19 10 5 lempung berpasir PML 2 7.6 6.8 0.12 1.08 9 4.4 lempung berliat PML 2.2 7.6 6.9 0.09 0.93 10 3.5 lempung berdebu PML 2.5 7.5 6.7 0.07 0.68 10 6.3 lempung
PNK 1 = pegunungan umur 1 tahun; PNK 2 = pegunungan umur 2 tahun; PML 2 = pantai umur 2 tahun; PML 2.2 = pantai umur 2.2 tahun; PML 2.5 = pantai umur 2.5 tahun.
Kelima asal tanah ini berasal dari 2 tempat, yaitu daerah pegunungan (PNK 1 dan PNK 2) dan daerah pantai (PML 2, PML 2.2, dan PML 2.5). Berdasarkan Tabel 1 tanah yang berasal dari pegunungan yaitu PNK 1 dan PNK 2 memiliki tekstur yang sama yaitu lempung berpasir, sedangkan untuk daerah pantai memiliki tekstur yang berbeda-beda yaitu lembung berliat, lempung berdebu dan lempung. Berdasarkan data analisis kimia tanah diketahui bahwa kisaran pH H2O
tanah yang terdapat di daerah pegunungan tergolong agak masam sedangkan untuk di daerah pantai kisaran pH H2O tanah tergolong netral sampai agak alkalis.
pH KCl yang terdapat pada lokasi berkisar antara netral sampai agak alkalis. Nisbah C/N mempengaruhi ketersediaan unsur hara di dalam tanah sehingga menjadi parameter untuk menentukan kesuburan tanah. Tanah pada lokasi penelitian ini tergolong tidak subur berdasarkan kriteria dari analisis sifat kimia tanah yang sudah dilakukan karena nisbah C/N tergolong rendah.
Berdasarkan data analisis kimia tanah di atas juga dapat diketahui bahwa ketersediaan unsur P di dalam tanah tergolong sangat rendah. Rendahnya kandungan unsur P dalam tanah menyebabkan tumbuhan mampu bersimbiosis dengan FMA. Selain itu dapat dilihat bahwa ketersediaan unsur N di daerah pegunungan tergolong sedang yaitu 0.43 dan 0.41, sedangkan untuk daerah pantai tergolong rendah yaitu 0.12 sampai sangat rendah yaitu 0.09 dan 0.07. Unsur N sangat berperan penting dalam pertumbuhan tanaman. Apabila tingkat nitrogen dalam suatu tanah tinggi, maka akan berpengaruh negatif terhadap pembentukan dan perangsangan pertumbuhan mikoriza di dalam tanah tersebut. Kriteria penilaian sifat-sifat kimia tanah tersebut dapat dilihat pada lampiran 2 menurut Hardjowigeno (1995).
6
Persentase Infeksi pada Akar
Menurut Brundrett et al. (1996) mikoriza adalah suatu bentuk hubungan simbiosis mutualistik antara fungi dengan akar tanaman. Kedua simbion sama-sama mendapat keuntungan. Dalam hal ini, fungi mikoriza dapat membantu tumbuhan dalam penyerapan air dan hara mineral dari dalam tanah, sedangkan fungi mendapatkan bahan-bahan organik dari tumbuhan yang bersangkutan. Akar tanaman akan mengeluarkan cairan karbohidrat yang akan dimanfaatkan oleh fungi pembentuk mikoriza untuk sumber energi. Hifa fungi bertindak sebagai akar-akar rambut dan berfungsi menyerap air serta hara dalam tanah, kemudian memberikannya kepada tanaman (Fakuara 1988). Akar yang terinfeksi ditunjukkan denga adanya struktur yang dibentuk oleh FMA yaitu hifa, vesikula dan arbuskula. Untuk lebih jelasnya dapat dilihat pada Gambar 1.
Gambar 1 Penampang akar jabon yang terinfeksi. arbuskula (a) vesikula (b) hifa (c)
Berdasarkan hasil pengamatan, pada akar tanaman sampel anakan jabon ditemukan adanya asosiasi antara akar dengan fungi mikoriza yang membentuk kolonisasi. Penginfeksian mikoriza ini ditandai dengan adanya hifa yang menembus sel epidermis melalui permukaan akar atau rambut-rambut akar. sehingga kelihatan jelas bagian yang terinfeksi yaitu bagian hifa, vesikula dan arbuskula. Arbuskula terbentuk secara intraseluler dan merupakan tempat terjadinya pertukaran hara antara inang dan fungi. Vesikel kebanyakan berbentuk bulat dan mengandung lipida, biasanya terbentuk di ujung hifa dan diperkirakan sebagai organ penyimpanan sementara (Dommergues dalam Sastrahidayat 2011). Vesikula juga berfungsi sebagai propagul (organ reproduktif). Ciri utama arbuskula mikoriza adalah terdapatnya arbuskula di dalam korteks akar. Awalnya fungi tumbuh di antara sel-sel korteks, kemudian menembus dinding sel inang dan berkembang di dalam sel (Brundrett et al. 1996). Bagi fungi mikoriza, hifa berfungsi mendukung fungsi reproduksi serta untuk transportasi karbon serta hara lainnya ke dalam spora, selain fungsinya untuk menyerap unsur hara dari dalam tanah. Pada kondisi lapangan keaktifan maksimal simbiosis antara mikoriza dengan pohon tidak dapat diketahui. Sistem perakaran pada tanaman tingkat tinggi berasosiasi tidak hanya dengan mengubah lingkungan organik dan anorganik tetapi juga dengan sistem metabolik dari mikroorganisme (Yusniar 2011). Arbuskula mikoriza merupakan salah satu tipe mikoriza paling umum yang membentuk struktur fungi yang berupa vesikel dan arbuskula di dalam jaringan korteks akar (Power dan Bagyaraj dalam Sastrahidayat 2011). Perkembangan infeksi dari mikoriza biasanya dimulai dengan apresorium oleh hifa eksternal pada
7
permukaan akar yang terinfeksi dalam tanah. Struktur dari akar yang terinfeksi pada dasarnya tidak menunjukkan tanda terinfeksi seperti pada ektomikoriza. Akar tanaman menciptakan lingkungan yang cocok bagi kehidupan mikroba dalam tanah. Lingkungan dan faktor biotik diketahui memiliki pengaruh terhadap pembentukan FMA dan proses infeksi terhadap inang. Hasil pengamatan infeksi FMA pada akar tanaman jabon yang berasal dari Pemalang Jawa Tengah diperoleh jumlah presentasi infeksi akar yang berbeda tiap lokasi seperti terlihat pada Gambar 2 dan Tabel 2.
Gambar 2 Rata-rata persentasi infeksi FMA pada akar jabon untuk setiap lokasi (PNK 1 = pegunungan umur 1 tahun; PNK 2 = pegunungan umur 2 tahun; PML 2 = pantai umur 2 tahun; PML 2.2 = pantai umur 2.2 tahun; PML 2.5 = pantai umur 2.5 tahun)
Tabel 2 Persentase infeksi akar pada jabon setiap lokasi
Lokasi Rata-rata Kriteria persentase infeksi*
PNK 1 57.78 Tinggi
PNK 2 60.37 Tinggi
PML 2 65.00 Tinggi
PML 2.2 73.33 Tinggi
PML 2.5 48.89 Sedang
*Sumber: Setiadi et al. (1992).
Berdasarkan Tabel 2 dapat diketahui persentase rata-rata kolonisasi akar terbesar terdapat pada lokasi PML 2.2 yaitu sebesar 73.33% dan persentase rata-rata terkecil terdapat pada lokasi PML 2.5 yaitu 48.89%. Kriteria persentase infeksi dikelompokan dalam empat kelas (Lampiran 1) berdasarkan Setiadi (1992). Peranan utama dari mikoriza adalah serapan unsur hara dan translokasi, khususnya fosfor. Manfaat dari asosiasi antara tanaman inang dengan mikoriza antara lain yaitu meningkatkan penyerapan unsur hara, lebih tahan terhadap kekeringan, tahan terhadap serangan patogen akar dan memperbaiki struktur tanah.
Berdasarkan hasil analisis kimia tanah, lokasi PML 2.2 memiliki kandungan fosfor terkecil dan lokasi PML 2.5 memiliki kandungan fosfor terbesar.
57.78 60.37 65.00 73.33 46.89 0 10 20 30 40 50 60 70 80 PNK 1 PNK 2 PML 2 PML 2.2 PML 2.5 R ata -r ata k o lo n is asi (%) Lokasi
8
Berdasarkan data analisis kimia tanah kandungan fosfor di PNK 1 lebih tinggi dibandingkan dengan PNK 2 dan nilai rata-rata infeksi akarnya PNK 1 lebih kecil yaitu 57.78% dibandingakan dengan PNK 2 yaitu 60.37%. Kandungan fosfor di lokasi daerah pantai secara berturut-turut dari yang terkecil yaitu PML 2.2, PML 2, PML 2.5, sedangkan untuk nilai rata-rata infeksi akarnya berturut-turut adalah 73.33%, 57.78% dan 48.89%. Hal ini sesuai dengan pernyataan yang dikemukakan oleh Harley dan Smith (1983) bahwa fungi mikoriza dapat berkembang dengan baik pada tanah yang mempunyai fosfor lebih rendah, sebaliknya kandungan fosfor dalam tanah tinggi, maka fungi mikoriza yang terdapat di sekitar rizosfer sedikit. Tanaman-tanaman yang tipe perakarannya kasar dan rambut akarnya kurang ternyata lebih sering terinfeksi mikoriza dan pertumbuhannya lebih tergantung kepada adanya mikoriza tersebut (Paul dan Clark 1980 dalam Syamsiyah 2008).
Adanya fungi mikoriza sangat penting bagi ketersediaan unsur hara seperti P, Mg, K, Fe dan Mn untuk pertumbuhan tanaman. Hal ini terjadi melalui pembentukan hifa pada permukaan akar yang berfungsi sebagai perpanjangan akar terutama di daerah yang kondisinya miskin unsur hara, pH rendah dan kurang air. Manfaat fungi mikoriza ini secara nyata terlihat jika kondisi tanahnya miskin hara atau kondisi kering, sedangkan pada kondisi tanah yang subur peran fungi ini tidak begitu nyata (Lakitan 2000).
Fungi mikoriza arbuskula dapat ditemukan hampir pada semua jenis tanah dan dapat memperbaiki pertumbuhan tanaman pada tanah-tanah yang tidak subur atau kekurangan unsur hara. Fungi mikoriza biasanya tersebar dengan berbagai cara. Mikoriza secara fisik terbentuk karena fungi pembentuk mikoriza mengeluarkan spora, kemudian spora tersebut berkecambah menjadi hifa dan masuk ke dalam akar tanaman sehingga terbentuklah mikoriza. Penyebaran aktif miselia melalui tanah. Setelah infeksi di akar, hifa berkembang di daerah perakaran pada tanah dan terbentuk struktur fungi, di antaranya yaitu miselium eksternal akar yang merupakan organ yang sangat penting dalam penyerapan unsur hara dan mentransfer ke tanaman. Keanekaragaman spesies mikoriza di ekosistem alami di daerah tropika sangat tinggi. Hutan alami yang terdiri dari banyak spesies tanaman dan umur yang tidak seragam sangat mendukung perkembangan mikoriza. Populasi mikoriza umumnya lebih tinggi pada tanah yang diolah dibandingkan dengan tanah perawan (hutan). Pada tanah lapisan atas, jumlah fungi mikoriza tinggi dan berkurang dengan semakin dalam dan tidak ditemukan pada kedalaman yang tidak dapat dijangkau akar. Sebaran spesies fungi mikoriza juga bervariasi menurut iklim, lingkungan edafik dan penggunaan lahan (Baon dalam Sastrahidayat 2011).
Rata-rata persentase infeksi akar di daerah pegunungan lebih kecil dibandingkan dengan daerah pantai. Fungi mikoriza bersifat aerobik atau membutuhkan oksigen yang cukup. Kekurangan okisgen menghambat perkembangan baik tumbuhan maupun simbiosis mikorizanya. Pada tegakan di daerah pegunungan, intensitas cahaya matahari yang sampai ke lantai hutan rendah, hal ini dipengaruhi oleh adanya kerapatan tajuk. Kondisi seperti ini menyebabkan kelembabannya cukup tinggi dan pasokan oksigen di dalam tanah rendah, sehingga menyebabkan infeksi mikorizanya rendah. Hasil analisis regresi antara sifat kimia tanah dengan nilai rata-rata infeksi akar dapat dilihat pada Tabel 3.
9
Tabel 3 Hasil analisis regresi antara sifat kimia tanah dengan nilai rata-rata infeksi akar di daerah pantai
Sifat Kimia
Tanah Persamaan
N-total Nilai infeksi akar = 32.6 + 312 N (%). = 33.7%. Pr>F = 0.606
C-organik Nilai infeksi akar = 15.5 – 51.4 C (%). = 59.1 %. Pr>F= 0,441
P-tersedia Nilai infeksi akar = 107 – 9.46 P (ppm). = 100.0%. Pr>F = 0.004
N=nitrogen; C=carbon; P=fosfor; = koefisien determinasi; Pr>F=peluang.
Berdasarkan hasil regresi pada Tabel 3 dapat dilihat nilai koefisien determinasi (R2) N-total terhadap nilai infeksi akar sebesar 33.7%, artinya N-total memiliki tingkat hubungan yang rendah terhadap infeksi akar (Sugiyono 2011). Berdasarkan uji ANOVA diperoleh nilai Pr>F sebesar 0.606, artinya N-total tidak berpengaruh terhadap infeksi akar di daerah pantai. Nilai koefisien determinasi (R2) C-organik terhadap nilai infeksi akar sebesar 59.1%, artinya C-organik memiliki tingkat hubungan yang sedang terhadap infeksi akar. Berdasarkan uji ANOVA diperoleh nilai Pr>F sebesar 0.441, artinya C-organik tidak berpengaruh terhadap infeksi akar di daerah pantai. Nilai koefisien determinasi (R2) fosfor terhadap nilai infeksi akar sebesar 100%, artinya fosfor memiliki tingkat hubungan yang sangat kuat terhadap infeksi akar. Berdasarkan uji ANOVA diperoleh nilai Pr>F sebesar 0.004, artinya P-tersedia berpengaruh terhadap infeksi akar di daerah pantai. Pada daerah pegunungan, data tidak dapat diolah menggunakan analisis regresi menggunakan program, hal ini disebabkan data tidak mencukupi. Namun secara deskriptif dapat dilihat bahwa kandungan P berpengaruh pada persentase infeksi akar. Semakin rendah kandungan P dalam tanah, maka akar yang terinfeksi semakin tinggi.
Setelah dilakukan pengujian hipotesis beda nilai tengah untuk nilai infeksi akar di daerah pantai dan pegunungan pada tingkat nya 5%, didapatkan hasil keputusan uji bahwa thitung jatuh di luar wilayah kritik maka H0 diterima. Dengan
demikian dapat disimpulkan bahwa pada tingkat nyata 5%, berdasarkan contoh yang ada, pernyataan bahwa nilai tengah infeksi akar daerah pantai sama dengan nilai tengah infeksi akar daerah pegunungan dapat diterima. Hal ini diduga karena selain kondisi fisik dan kimia tanah, kondisi lingkungan yang mempengaruhi daerah pantai dan daerah pegunungan berbeda.
Kepadatan Spora
Kepadatan spora merupakan jumlah spora yang dijumpai pada saat pengamatan. Hasil pengamatan menunjukan bahwa rata-rata kepadatan spora per 10 gram tanah adalah 34.13/10 gram tanah. Jumlah spora yang didapat diperoleh dari contoh tanah yang diambil dari lapangan sebanyak 10 gram tiap lokasinya dengan menggunakan metode tuang basah. Pada umumnya pada waktu spora membentuk miselium di sekeliling akar, yang menghambat perkembangan miselium bagian luar atau pertumbuhan akar adalah miskinnya kandungan unsur hara. Faktor kesuburan tanah sangat mempengaruhi jumlah spora yang berkembang. Jumlah spora FMA yang dijumpai pada masing-masing lokasi dapat dilihat pada Gambar 3.
10
Gambar 3 Kepadatan spora berdasarkan ketinggian dan umur tanaman (PNK 1 = pegunungan umur 1 tahun; PNK 2 = pegunungan umur 2 tahun; PML 2 = pantai umur 2 tahun; PML 2.2 = pantai umur 2.2 tahun; PML 2.5 = pantai umur 2.5 tahun)
Dari data di atas dapat diketahui bahwa kepadatan spora per 10 gram tanah berbeda-beda tiap lokasi, hal ini sesuai dengan Rainiyati (2007) yaitu perbedaan kepadatan spora kemungkinan dipengaruhi oleh perbedaan lingkungan (jenis tanah, hara tanaman, ketinggian tempat, cahaya) dan musim pada saat pengambilan sample tanah. Gambar 3 menujukan jumlah rata-rata spora tertinggi ditemukan di daerah pegunungan yaitu pada lokasi PNK 2 (45 spora) dan jumlah rata-rata spora terendah yaitu pada lokasi PNK 2.2 dan PML 2 (26 spora).
Setelah dilakukan pengujian hipotesis beda nilai tengah untuk nilai kepadatan spora di daerah pantai dan pegunungan pada tingkat nyata 5%, didapatkan hasil keputusan uji bahwa thitung jatuh di luar wilayah kritik maka H0
diterima. Dengan demikian dapat disimpulkan bahwa pada tingkat nyata 5%, berdasarkan contoh yang ada, pernyataan bahwa nilai tengah kepadatan spora di daerah pantai sama dengan nilai kepadatan spora di daerah pegunungan dapat diterima. Seperti halnya pada infeksi akar, hal tersebut diakibatkan kondisi fisik, kimia tanah dan lingkungan yang berbeda serta genus spora itu sendiri. Berdasarkan data diatas dapat dilihat bahwa rata-rata jumlah spora di pegunungan lebih besar dibandingkan dengan daerah pantai. Kepadatan spora tinggi pada saat infeksi akar kecil. Artinya dapat disimpulkan bahwa dari hasil penelitian ini, kepadatan spora tidak berhubungan erat dengan infeksi akar. Hal ini sesuai dengan pernyataan Tuheteru (2003) bahwa antara infeksi akar dan jumlah spora yang dihasilkan tidak memiliki korelasi yang erat, sehingga spora yang banyak belum tentu persentase infeksi akar akan tinggi pula.
Keanekaragaman Spora
Keanekaragaman spora yang ditemukan pada setiap lokasi pengamatan berbeda-beda. Dalam penelitian ini identifikasi FMA hanya dibatasi hingga tingkat genus. Genus terbanyak yang ditemukan yaitu genus Glomus. Beberapa genus yang ditemukan di lokasi penelitian dapat dilihat pada Tabel 4.
27 45 26 26 44 0 10 20 30 40 50 PNK 1 PNK 2 PML 2 PML 2.2 PML 2.5 Ju m lah s p o ra p er 1 0 g ram Lokasi
11
Tabel 4 Keanekaragaman spora di bawah tegakan jabon Pemalang
Sample Tanah Nilai Keanekaragaman FMA
PNK 1 2 (Glomus, Scutellospora)
PNK 2 3 (Glomus, Scutellospora, Acaulospora) PML 2 3 (Glomus, Scutellospora, Acaulospora) PML 2.2 2 (Glomus, Acaulospora)
PML 2.5 2 (Glomus, Scutellospora)
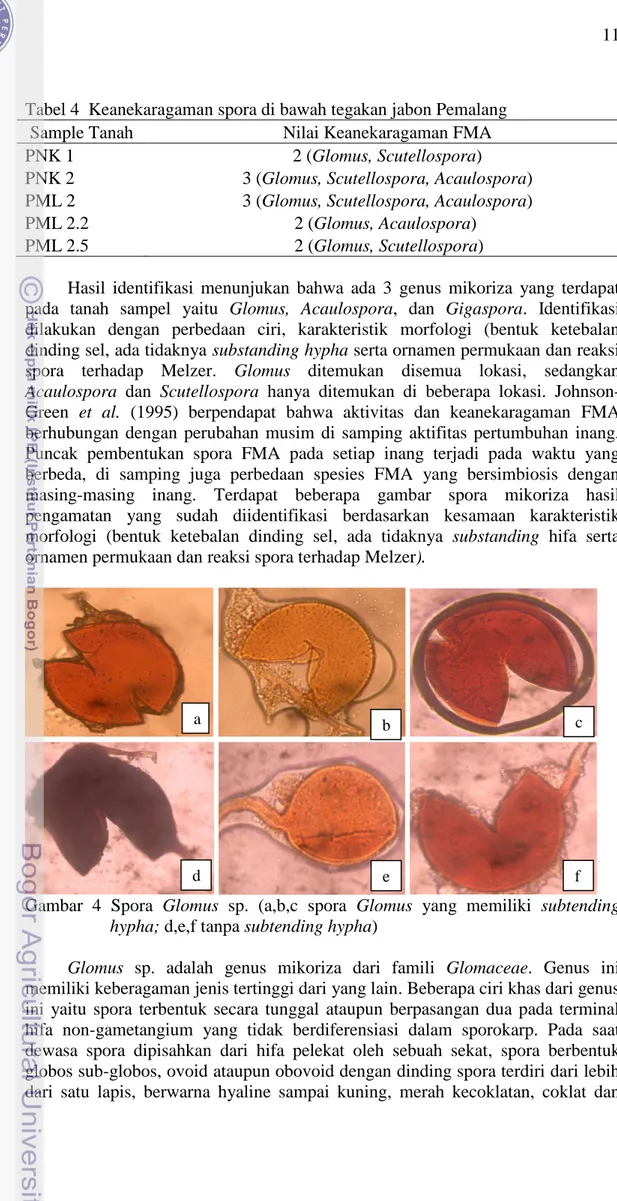
Hasil identifikasi menunjukan bahwa ada 3 genus mikoriza yang terdapat pada tanah sampel yaitu Glomus, Acaulospora, dan Gigaspora. Identifikasi dilakukan dengan perbedaan ciri, karakteristik morfologi (bentuk ketebalan dinding sel, ada tidaknya substanding hypha serta ornamen permukaan dan reaksi spora terhadap Melzer. Glomus ditemukan disemua lokasi, sedangkan Acaulospora dan Scutellospora hanya ditemukan di beberapa lokasi. Johnson-Green et al. (1995) berpendapat bahwa aktivitas dan keanekaragaman FMA berhubungan dengan perubahan musim di samping aktifitas pertumbuhan inang. Puncak pembentukan spora FMA pada setiap inang terjadi pada waktu yang berbeda, di samping juga perbedaan spesies FMA yang bersimbiosis dengan masing-masing inang. Terdapat beberapa gambar spora mikoriza hasil pengamatan yang sudah diidentifikasi berdasarkan kesamaan karakteristik morfologi (bentuk ketebalan dinding sel, ada tidaknya substanding hifa serta ornamen permukaan dan reaksi spora terhadap Melzer).
Gambar 4 Spora Glomus sp. (a,b,c spora Glomus yang memiliki subtending hypha; d,e,f tanpa subtending hypha)
Glomus sp. adalah genus mikoriza dari famili Glomaceae. Genus ini memiliki keberagaman jenis tertinggi dari yang lain. Beberapa ciri khas dari genus ini yaitu spora terbentuk secara tunggal ataupun berpasangan dua pada terminal hifa non-gametangium yang tidak berdiferensiasi dalam sporokarp. Pada saat dewasa spora dipisahkan dari hifa pelekat oleh sebuah sekat, spora berbentuk globos sub-globos, ovoid ataupun obovoid dengan dinding spora terdiri dari lebih dari satu lapis, berwarna hyaline sampai kuning, merah kecoklatan, coklat dan
a b c
12
hitam. Berukuran antara 20–400 μm (INVAM 2010). Selain itu Glomus sp. tidak bereaksi dengan larutan Melzer. Glomus berkembang dengan baik pada pH 5.5 sampai 6.5 (Sastrahidayat 2011).
Gambar 5 Spora Acaulospora sp. (a,b Acaulospora sp. dengan saccule; c Acaulospora sp. dengan cicatrix)
Acaulospora sp. adalah genus mikoriza yang termasuk dalam famili Acaulosporaceae. Genus ini memiliki beberapa ciri khas antara lain yaitu memiliki 2–3 dinding spora, spora terbentuk di sisi samping leher sporiferous saccule, berbentuk globos hingga elips, berwarna hyaline, kuning, ataupun merah kekuningan,berukuran antara 100–400 μm (INVAM 2013). Selain itu ciri khas Acaulospora sp. adalah dinding spora bagian luar tidak bereaksi dengan larutan Melzer sedangkan bagian dalam bereaksi dengan larutan Melzer. Menurut Moreira et al. (2007) pada ekosistem hutan asli Acaulospora mempunyai keanekaragaman jenis yang paling tinggi. Acaulouspora berkembang dengan baik pada pH 5.0 (Sastrahidayat 2011).
Gambar 6 Spora Scutellospora sp. memiliki germination shield
Scutellospora sp. adalah genus mikoriza dari famili Gigasporaceae. Proses perkembangan Scutellospora sama dengan Gigaspora. Untuk membedakan dengan genus Gigaspora, pada Scutellospora terdapat lapisan kecambah. Bila berkecambah hifa ke luar dari lapisan kecambah tadi. Spora bereaksi dengan larutan Melzer secara menyeluruh. Warna sporanya merah coklat ketika bereaksi dengan larutan Melzer. Ukuran sporanya rata-rata 165 μm (INVAM 2013).
Kelimpahan Spora
Kelimpahan relatif ialah jumlah genus yang terdapat pada lokasi pengamatan dibagi dengan total spora yang ada pada lokasi pengamatan dan
13
dikali 100%. Besarnya kelimpahan genus spora FMA di bawah tegakan tanaman jabon Pemalang Jawa Tengah dapat dilihat pada Gambar 7.
Gambar 7 Kelimpahan spora FMA di bawah tegakan jabon Pemalang (PNK 1 = pegunungan umur 1 tahun; PNK 2 = pegunungan umur 2 tahun; PML 2 = pantai umur 2 tahun; PML 2.2 = pantai umur 2.2 tahun; PML 2.5 = pantai umur 2.5 tahun)
Glomus memiliki nilai kelimpahan relatif lebih tinggi dibandingkan dengan Scutellospora dan Acaulospora di setiap lokasi. Genus Glomus memiliki kelimpahan paling tinggi. Dominannya Glomus ini menunjukan bahwa Glomus bersifat adaptif dan menunjukan toleransi yang tinggi pada tanah di bawah tegakan tanaman Jabon karena kemampuan Glomus untuk dapat tumbuh pada kisaran lingkungan yang lebih luas. Glomus mempunyai jenis yang paling banyak dan mempunyai kemampuan adaptasi pertumbuhan terhadap lingkungan yang lebih besar dibandingkan dengan genus lainnya. Tumbuh baiknya Glomus ini dapat disebabkan karena Glomus dapat tumbuh dengan baik di daerah tersebut serta penyebaran genus yang sangat luas dan memiliki kesesuaian yang optimal terhadap faktor lingkungan yang ada.
Frekuensi Spora
Fungi mikoriza arbuskula dapat ditemukan hampir pada semua jenis tanah dan dapat memperbaiki pertumbuhan tanaman pada tanah-tanah yang tidak subur atau kekurangan unsur hara. Fungi mikoriza biasanya tersebar dengan berbagai cara. Frekuensi spora berkaitan dengan penyebaran spora pada lokasi tempat pengambilan sampel tanah. Jumlah spora juga sangat dipengaruhi oleh suhu dan kelembaban. Semakin tinggi suatu daerah, maka suhunya akan semakin rendah. Suhu yang rendah dapat berpengaruh negatif terhadap tanaman, di antaranya yaitu dapat menurunkan laju fotosintesis dan dapat mengurangi serapan unsur hara. Kelembaban yang tinggi menandakan kandungan air yang tinggi dan suhu yang rendah pada suatu lahan hutan. akibatnya unsur hara akan semakin rendah akibat proses pencucian pada lahan tersebut. Frekuensi spora di bawah tegakan jabon Pemalang Jawa Tengah disajikan pada Gambar 8.
92.77 81.62 90.59 94.67 95.56 0.00 7.35 4.71 5.33 0.00 7.23 11.03 4.71 0.00 4.44 0 25 50 75 100 PNK 1 PNK 2 PML 2 PML 2.2 PML 2.2 Kelim p ah an r elatif ( %) Lokasi Glomus Acaulospora Scutellospora
14
Gambar 8 Frekuensi sebaran genus spora FMA dibawah tegakan Jabon Pemalang (PNK 1 = pegunungan umur 1 tahun; PNK 2 = pegunungan umur 2 tahun; PML 2 = pantai umur 2 tahun; PML 2.2 = pantai umur 2.2 tahun; PML 2.5 = pantai umur 2.5 tahun)
Gambar 8 menunjukkan bahwa genus Glomus memiliki frekuensi paling besar pada setiap lokasi, hingga frekuensinya mencapai 85.71%. Hal tersebut menjelaskan bahwa genus tersebut terdapat dan menyebar di setiap lokasi pengamatan. Glomus merupakan genus yang mempunyai jenis paling banyak dan juga kemampuannya yang dapat tumbuh pada kisaran yang luas. Berdasarkan Gambar 8 genus Acaulospora memiliki nilai frekuensi spora yang terendah.
Rata-rata Diameter dan Tinggi Tanaman Jabon
Dari hasil rekapitulasi data tinggi dan diameter tanaman di setiap lokasi, diketahui rata-rata tinggi dan diameter tanaman di daerah pantai dan daerah pegunungan. Hasilnya dapat dilihat pada Gambar 9 dan Gambar 10.
Gambar 9 Rata-rata tinggi tanaman jabon di pantai dan pegunungan (PNK = lokasi pegunungan; PML = lokasi pantai)
85.71 72.00 80.95 85.71 85.71 0.00 8.00 4.76 0.00 0.00 14.29 20.00 14.29 14.29 14.29 0 25 50 75 100 PNK 1 PNK 2 PML 2 PML 2.2 PML 2.5 Fre k u en si ( %) Lokasi Glomus Acaulospora Scutellospora 3.44 11.43 0 2 4 6 8 10 12 14 PNK PML R ata -r ata tin g g i p o h o n ( m ) Lokasi
15
Gambar 10 Rata-rata diameter tanaman jabon di pantai dan pegunungan (PNK = lokasi pegunungan; PML = lokasi pantai)
Gambar 9 dan Gambar 10 menunjukkan bahwa rata-rata diameter dan tinggi tanaman jabon di daerah pegunungan lebih kecil dibandingkan dengan rata-rata diameter dan tinggi tanaman di daerah pantai. Salah satu faktor yang menyebabkan perbedaan ketinggian tersebut adalah suhu di pegunungan yang lebih rendah dan terbentuknya awan atau kabut yang kerap menyelimuti atap tajuk. Pengaruh ketinggian tempat terhadap pertumbuhan pohon bersifat tidak langsung (Soedomo 1984). Artinya perbedaan ketinggian tempat akan mempengaruhi keadaan lingkungan tumbuh pohon, terutama suhu, kelembaban, O2 di udara dan keadaan tanah. Keadaan lingkungan ini akhirnya mempengaruhi
pertumbuhan pohon. Kekurangan okisgen menghambat perkembangan baik tumbuhan maupun simbiosis mikorizanya. Pada tegakan di daerah pegunungan, intensitas cahaya matahari yang sampai ke lantai hutan rendah. Kondisi seperti ini menyebabkan kelembabannya cukup tinggi dan suplai oksigen di dalam tanah rendah, sehingga menyebabkan pertumbuhan pohon tidak optimal.
Dalam penelitian Pendry dan Proctor (1996) dalam Kusmana et al. (2008) di Bukit Belalong (913 mdpl) yang merupakan sebuah gunung kecil di Brunei menemukan bahwa struktur tegakan hutan tropis dataran rendah dengan ukuran pohon yang besar ditemukan sampai pada ketinggian 750 m dpl. dan di atas ketinggian tersebut terjadi perubahan dengan ditemukan ukuran pohon yang semakin mengecil. Mereka menyatakan bahwa perubahan struktur hutan ini disebabkan oleh perubahan ketinggian tempat yang pada gilirannya menyebabkan menurunnya suhu udara.
4.35 14.21 0 2 4 6 8 10 12 14 16 PNK PML R ata -r ata d iam eter p o h o n ( cm ) Lokasi
16
SIMPULAN DAN SARAN
Simpulan
1. Keanekaragaman fungi mikoriza arbuskula (FMA) di bawah tegakan tanaman jabon Pemalang Jawa Tengah diantaranya terdapat 3 genus yaitu Glomus, Acaulospora dan Scutellospora. Lokasi di daerah pegunungan memiliki kepadatan spora yang lebih besar dibandingkan dengan lokasi di daerah pantai.
2. Genus Glomus memiliki nilai kelimpahan relatif dan frekuensi yang tertinggi dibandingkan dengan genus lainnya. Persentase rata-rata kolonisasi akar di daerah pantai lebih besar yaitu 59% dan termasuk dalam kriteria infeksi tinggi, sedangkan rata-rata infeksi di daerah pegunungan yaitu 62% dan termasuk kriteria sedang tinggi.
Saran
Perlu dilakukan penelitian lanjutan mengenai potensi mikoriza terhadap pertumbuhan tanaman Jabon untuk setiap genus.
DAFTAR PUSTAKA
Adawiyah. 2009. Status keanekaragaman fungi mikoriza arbuskula berdasarkan tinggi gradien salinitas di hutan pantai pulau Pandang Kabupaten Batu Bara Sumatera Utara [tesis]. Medan (ID): Universitas Sumatera Utara.
Brundrett M, Boucher N, Dell NB, Gove T, Malajczuk N. 1996. Working with Mycorrhizas in Forestry and Agriculture. Canberra (AU): Australian Centre for International Agriculture Research.
Fakuara Y. 1988. Mikoriza, Teori dan Kegunaannya dalam Praktik. Bogor (ID): Fakultas Kehutanan IPB.
Gerdemann JW, Nicolson TH. 1963. Spores of mycorrhizal endogene spesies extracted from soil by wet selving and decanting. Trans Brit Mycol Soc. 46:235-244.
Giovannetti M, Mosse B. 1980. An evaluation of techniques for measuring vesicular-arbuscular mycorrhizal infection in roots. New Phytol. 84:489-500. Johnson-Green PC, Kenkel NC, Booth T. 1995. The distribution and phenology of
arbuscular mycorrhizae along an inland salinity gradient. Can.J.Bot. 73:1318-1327.
Hardjowigeno S. 1995. Ilmu Tanah. Jakarta (ID): Akademika Pr.
Harley JL, Smith SE. 1983. Mycorrhizal Symbiosis. New York (US): Academic Pr.
[INVAM] International culture collection of (vesicular) arbuscular mycorrhizal fungi. 2013. Reference cultures of species (vesicular) arbuscular mycorrhizal
17
fungi [Internet].[diunduh 2013 Jun 20].Tersedia pada: http://invam.caf.wvu.edu /Myco-info/Taxonomy/clasification.htm.
Kusmana C, Wiharto M, Prasetyo LB, Partomihardjo T. 2008. Distribusi kelas diameter pohon pada berbagai tipe vegetasi di Gunung Salak, Bogor, Jawa Barat. Jurnal Ilmu Pertanian Indonesia 13(2):95-102.
Lakitan B. 2000. Dasar-dasar Fisiologi Tumbuhan. Jakarta (ID): PT Raja Grafindo Persada.
Moreira, Dilmar, Tsai SM. 2007. Biodiversity dan distribution of arbuscular mycorrhizal fungi in Araucaria angustifolia forest. Journal Agriculture 64:393-399.
Rainiyati. 2007. Status dan keanekaragaman fungi mikoriza arbuskula (FMA) pisang raja nangka dan potensi pemanfaatannya untuk peningkatan produksi pisang asal kultur jaringan di Kabupaten Merangin, Jambi [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Santoso E, Turjaman M, Irianto RSB. 2007. Aplikasi mikoriza untuk meningkatkan kegiatan rehabilitasi hutan dan lahan terdegradasi. Di dalam: Siran AS, Bismark M, Samsoedin I, Suhaendi H, Pratiwi, Haryono, Mardiah, editor. Konservasi dan Rehabilitasi Sumberdaya Hutan; 2006 Sep 20; Padang, Indonesia. Bogor (ID): Pusat Penelitian dan Pengembangan Kehutanan. hlm 71-80.
Sastrahidayat IR. 2011. Rekayasa Pupuk Hayati Mikoriza dalam Meningkatkan Produksi Pertanian. Malang (ID): UB Pr.
Setiadi Y. 1992. Mikoriza dan Pertumbuhan Tanaman. Departemen Pendidikan dan Kebudayaan Direktorat Jendral Pendidikan Tinggi Pusat Antar Universitas IPB. Bogor (ID): Pusat Antar Universitas Institut Pertanian Bogor.
Soedomo S. 1984. Studi hubungan sifat-sifat tanah dan fisiografi dengan peninggi Pinus mercusii Jungh et de Vriese [tesis]. Bogor (ID): Institut Pertanian Bogor.
Sugiyono. 2011. Statistika untuk Penelitian. Bandung (ID): Penerbit Alfabeta.
Syamsiyah S. 2008. Respon tanaman padi gogo (Oryza sativa L.) terhadap stres air dan inokulasi mikoriza [skripsi]. Bogor (ID): Institut Pertanian Bogor Tuheteru FD. 2003. Aplikasi asam humat tergadap sporulasi FMA dari bawah
tegakan alami sengon [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Wang B, Qiu YL. 2006. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 16:299-363.
Warisno, Dahana K. 2011. Peluang Investasi Jabon Tanaman Masa Depan. Jakarta (ID): PT Gramedia Pustaka Utama.
Yusniar M. 2011. Keanekaragaman fungi mikoriza arbuskula di bawah tegakan sengon (Paraserianthes falcataria) [skripsi]. Sumatra (ID): Universitas Sumatra Utara.
18
Lampiran 1 Kriteria persentase kolonisasi akar *
No. Persentase kolonisasi akar (%) Keterangan
1 0 – 25 Rendah
2 26 – 50 Sedang
3 51 – 71 Tinggi
4 76 – 100 Sangat Tinggi
*Sumber: Setiadi et al. (1992).
Lampiran 2 Kriteria penilaian sifat-sifat kimia tanah* Sifat tanah Satuan Sangat
rendah Rendah Sedang Tinggi
Sangat Tinggi C (Carbon) % <1,00 1,00-2,00 2,01-3,00 3,01-5,00 >5,00 N (Nitrogen) % <0,10 0,10-0,20 0,21-0,50 0,51-0,75 >0,75 C/N --- <5 5-10 11-15 16-25 >25 P2O5 Total % <0,03 0,03-0,06 0,06-0,079 0,08-0,10 >0,10 P2O5 Eks HCL % <0,021 0,021-0,039 0,040-0,060 0,061-0,100 >0,100 P-avl Bray II ppm <8,0 8,0-1,5 16-25 26-35 >35 P- avl Troug ppm <20 20-39 40-60 61-80 >80 P- avl Olsen ppm <10 10-25 26-45 46-60 >60 K2O eks-HCL % <0,03 0,03-0,06 0,07-0,11 0,12-0,20 >0,20 MgO- eks HCL % <0,05 0,05-0,09 0,10-0,20 0,21-0,30 >0,30 MnO- eks HCL % <0,05 0,05-0,09 0,10-0,20 0,21-0,30 >0,30 K- tukar me/100 <0,10 0,10-0,20 0,30-0,50 0,60-0,100 >1,00 Na- tukar me/100 <0,10 0,10-0,30 0,40-0,70
0,80-0,100 >1,00 Ca- tukar me/100 <2,0 2,0-5,0 6,0-10,0 11,0-20,0 >20 Mg- tukar me/100 <0,40 0,40-1,00 1,10-2,00 2,10-8,00 >8,00 KTK (CEC) me/100 <5 5-16 17-14 25-80 >40 KB(BS) % <20 20-35 36-50 51-70 >70 Kej .Al % <10 10-20 21-30 31-60 >60 EC (Nedeco) mmhos/ cm --- --- 2,5 2,6-10 >10
19 Lanjutan Lampiran 2 pH Sangat masam Masam Agak masam Netral Agak alkalis Alkalis H2O <4,5 4,5-5,5 6,5-6,5 6,6-7,5 7,6-8,5 >8,5 KCl <2,5 2,5-4,0 ---- 4,1-6,0 6,1-6,5 >6,5 *Sumber: Hardjowigeno (1995).
20
RIWAYAT HIDUP
Penulis dilahirkan di Tasikmalaya pada tanggal 19 Juli1990 dari ayah Uyu Suprayadi dan ibu Munawaroh. Penulis adalah putri pertama dari dua bersaudara. Tahun 2009 penulis lulus dari SMA Negeri 1 Tasikmalaya dan pada tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk IPB dan diterima di Departemen Silvikultur, Fakultas Kehutanan.
Selama menjadi mahasiswa, penulis aktif di beberapa organisasi, yaitu Lingkungan Seni Gentra Kaheman, Badan Eksekutif Mahasiswa Fakultas Kehutanan (BEM-E) Divisi Kemahasiswaan pada tahun 2010/2011, Himpunan Profesi Tree Grower Community pada tahun 2010/2012. Selain itu, penulis juga aktif di berbagai kepanitiaan yang diselenggarakan oleh beberapa organisasi tingkat perguruan tinggi maupun nasional. Bulan Februari sampai dengan April, penulis mengikuti Praktek Kerja Profesi di KPH Tasikmalaya, Perum Perhutani Unit III Jawa Barat dan Banten dengan Judul Teknik dan Managemen Penanaman Jati Plus Perhutani (JPP) di KPH Tasikmalaya Perum Perhutani Unit III Jawa Barat Dan Banten. Penulis juga pernah mengikuti program kreativitas mahasiswa (PKM).
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan, penulis melakukan penelitian yang berjudul Keanekaragaman Fungi Mikoriza Arbuskula (FMA) di Bawah Tegakan Tanaman Jabon (Anthocephalus cadamba Miq.) di Pemalang Jawa Tengah dibawah bimbingan Dr Ir Sri Wilarso Budi R, MS.