STATUS DAN KEANEKARAGAMAN FUNGI MIKORIZA
ARBUSKULA BERDASARKAN GRADIEN SALINITAS
DI HUTAN PANTAI PULAU PANDANG,
BATU BARA, SUMATERA UTARA
T E S I S
Oleh :
ADAWIYAH
077030001/BIO
SEKOLAH PASCASARJANA
UNIVERSITAS SUMATERA UTARA
STATUS DAN KEANEKARAGAMAN FUNGI MIKORIZA
ARBUSKULA BERDASARKAN GRADIEN SALINITAS
DI HUTAN PANTAI PULAU PANDANG,
BATU BARA, SUMATERA UTARA
T E S I S
Diajukan Sebagai Salah Satu Syarat Untuk Memperoleh
Gelar Magister Sains dalam Program Studi Biologi pada Sekolah
Pascasarjana Universitas Sumatera Utara
Oleh
ADAWIYAH
077030001/BIO
SEKOLAH PASCASARJANA
UNIVERSITAS SUMATERA UTARA
Judul Tesis : STATUS DAN KEANEKARAGAMAN FUNGI MIKORIZA ARBUSKULA BERDASARKAN
GRADIEN SALINITAS DI HUTAN PANTAI PULAU PANDANG, BATU BARA, SUMATERA UTARA
Nama Mahasiswa : ADAWIYAH Nomor Pokok : 077030001 Program Studi : Biologi
Menyetujui Komisi Pembimbing
(Dr. Delvian, SP. MP) (Prof. Dr. Dwi Suryanto, M. Sc) Ketua Anggota
Ketua Program Studi Direktur
Prof. Dr. Dwi Suryanto, M. Sc Prof. Dr. Ir. T. Chairun Nisa B, M. Sc
Telah diuji pada
Tanggal: 08 September 2009
PANITIA PENGUJI TESIS
Ketua : Dr. Delvian, SP. MP
Anggota : 1. Prof. Dr. Dwi Suryanto, M.Sc
2. Dr. Ir. Edy Batara Mulya Siregar, MS
ABSTRAK
Penelitian ini bertujuan untuk mengetahui keanekaragaman dan jenis fungi mikoriza di hutan pantai berdasarkan gradien salinitas. Contoh tanah sebanyak 50 g diambil dari Pantai Pulau Pandang Kabupaten Batu Bara dari setiap petak ukur 20 x 5 m. Contoh tanah disaring dengan satu set saringan bertingkat dan spora yang didapat diidentifikasi. Hasil pengamatan menunjukkan bahwa di lokasi penelitian terdapat 246 spora yang termasuk kedalam 3 genus mikoriza yaitu Glomus, Acaulospora dan Gigaspora. Jenis Glomus merupakan yang paling dominan dan terdapat pada setiap petak ukur. Kepadatan populasi spora makin meningkat sejalan dengan meningkatnya gradien salinitas. Hasil trapping dengan tanaman inang Pueraria javanica menunjukkan jumlah spora yang tinggi dibandingkan dengan spora yang diisolasi dari lapangan. Semua akar contoh tanaman terinfeksi fungi mikoriza. Fungi mikoriza mampu membentuk asosiasi dengan akar tanaman hutan pantai dengan persentase kolonisasi yang beragam, paling rendah dalam akar tanaman Ipomoea pescaprae (13%) dan yang paling tinggi (83,9%) dalam akar tanaman Terminalia catappa.
ABSTRACT
The objective of research was to know the mycorrhiza diversity in coastal forest based on salinity gradient. A 50 g of soil sample has been taken from the Pulau Pandang Beach, District of Batu Bara, from each plot of 20 x 5 m size. The soil sample was filtered through a set of gradient filter, and the spora found was then identified. The result of observation indicated that in location of research, there were 246 spores categorized into three genus of mycorrhizae; Glomus, Acaulospora and Gigaspora. Glomus was the most dominant type and found in each plot. The density of spora population increased with increased salinity gradient. The trapping result with host of Pueraria javanica indicated a high number of spora in comparison to the spora isolated from the site. All roots of plant sample have been infected by mycorrhiza fungi. The mycorrhizae fungi could form the association with roots of coastal forest plants of various colonization percentage, the lowest percentage was in plant roots of Ipomoea pescaprae (13%) and the highest percentage (83,9%) was in plant roots of Terminalia catappa.
DAFTAR TABEL
Nomor Judul Halaman
1
Klasifikasi fungi mikoriza arbuskula ………...
Tingkat salinitas tanah pada lokasi penelitian ………
Hasil analisis sifat kimia contoh tanah dan kriterianya menurut
Pusat penelitian tanah (1983) dan BPP dalam Harjowigeno…..
Jumlah tipe spora berdasarkan tingkat salinitas tanah ..………
Jumlah spora dari lapangan ………...
Jumlah spora hasil pemerangkapan (trapping) ……….
Nilai frekuensi mutlak (FM-%) dan frekuensi relatif (FR-%) kehadiran suatu jenis fungi mikoriza pada setiap petak ukur di
lapangan
...
Nilai frekuensi mutlak (FM-%) dan frekuensi relatife (FR-%)
Kehadiran suatu jenis fungi mikoriza pada hasil trapping ...
Karakteristik spora yang ditemukan dari lapangan …………..
Karakteristik spora yang ditemukan dari trapping …………...
DAFTAR GAMBAR
Nomor Judul Halaman
1
2
3
4
5
6
Penampang longitudinal akar yang terinfeksi fungi mikoriza (Brundrett et al., 1994)
Filogeni perkembangan dan taksonomi ordo
Glomeromycota (INVAM, 2009) ………...
Kepadatan spora dari lapangan pada berbagai tingkat salinitas tanah ………
Kepadatan spora hasil trapping pada berbagai tingkat salinitas tanah ………
A. Penampang akar Buchanania arborescen, v (vesikula) B. Penampang akar Terminalia catappa, h (hifa) ……….
Hubungan persentase kolonisasi mikoriza dengan tingkat salinitas tanah ………...
6
7
25
27
36
DAFTAR LAMPIRAN
Skematis ekstraksi dan identifikasi spora fungi mikoriza..
Skematis kolonisasi fungi mikoriza pada akar tanaman sampel ………...
Skematis teknis pembuatan kultur trapping fungi
mikoriza ………...
Dokumentasi penelitian
Gambar 1. Tanaman Pueraria javanica dalam trapping fungi mikoriza ………...
Gambar 2. Lokasi pengambilan sampel (tepi pantai Pulau Pandang) ...
Gambar 3. Lokasi pengambilan sampel (dalam hutan pantai Pulau Pandang) ...
Gambar 4. Peta lokasi penelitian pantai Pulau Pandang ...
KATA PENGANTAR
Bismillahirrahmannirrahim
Puji dan syukur kehadirat Allah SWT yang telah melimpahkan rahmat
dan karunia-Nya sehingga tesis ini yang berjudul “Status dan keanekaragaman
fungi mikoriza arbuskula berdasarkan gradien salinitas di hutan pantai Pulau
Pandang, Batu
Bara, Sumatera Utara” dapat diselesaikan tepat pada waktunya. Tesis ini dibuat
sebagai salah satu syarat untuk menyelesaikan studi pada Program Studi Magister
Biologi Sekolah Pascasarjana Universitas Sumatera Utara Medan.
Selama pelaksanaan penelitian ini penulis telah banyak mendapat
bantuan dan bimbingan dari berbagai pihak. Untuk itu pada kesempatan ini
penulis ingin menyampaikan rasa terima kasih kepada :
1. Dr. Delvian, SP. MP selaku ketua komisi pembimbing dan Prof. Dr. Dwi
Suryanto
selaku pembimbing II, sekaligus sebagai ketua Departemen Biologi, atas
segala
bantuan, bimbingan dan arahan yang telah diberikan kepada penulis.
2. Dr. Ir. Edy Batara Mulya Siregar, MS sebagai penguji I serta Dr. Deni
Elfiati,
SP. MP sebagai penguji II yang telah banyak memberikan arahan dan
masukan
3. Pemerintah Provinsi Sumatera Utara melalui BAPPEDA yang telah
memberikan
kesempatan dan bantuan finansial kepada penulis untuk mengikuti perkuliahan
Sekolah Pascasarjana ini.
4. Drs. Sofyan Alwi, M.Hum selaku kepala sekolah SMA Harapan 1 Medan
dan Ibnu Rusdi S.Pd. M.si yang telah memberikan dukungan dan motivasi bagi
penulis.
5. Orang tuaku Hasan Mansyur dan Mahinun juga saudara kandungku serta
seluruh
keluarga yang telah memberikan motivasi dan doa untuk menyelesaikan tesis
ini.
6. Suamiku tercinta Ir. Ms. Birgantara dan anak-anakku tersayang yang
memberikan
motivasi, doa dan mendampingi dengan sabar selama pendidikan
demi
keberhasilan studi ini.
Penulis menyadari bahwa tesis ini masih jauh dari sempurna. Oleh karena
itu dengan segala kerendahan hati menerima kritik dan saran yang membangun
dari semua pihak. Penulis berharap semoga hasil penelitian ini dapat bermanfaat
bagi pihak yang memerlukannya. Akhir kata semoga bermanfaat bagi kita semua.
Amin.
RIWAYAT HIDUP
Penulis dilahirkan di Tg. Tiram pada tanggal 15 Januari 1969,
sebagai
anak keempat dari tiga belas bersaudara, dari pasangan Hasan Mansyur dan
Mahinun.
Tahun 1983 penulis menyelesaikan pendidikan Sekolah Dasar di SD
Negeri No. 5 Tg. Tiram tahun 1983, selanjutnya pada tahun 1986 menyelesaikan
pendidikan Sekolah Menengah Pertama di SMP Negeri No. 1 Tg. Tiram. Tahun
1989 penulis menyelesaikan pendidikan Sekolah Menengah Atas di SMA swasta
UISU Medan.
Pendidikan sarjana ditempuh di Jurusan Biologi, Fakultas Keguruan Ilmu
Pendidikan UISU Medan, lulus pada tahun 1994. Kemudian penulis mendapatkan
kesempatan untuk melanjutkan pendidikan ke Program Magister (S-2) Biologi
di Sekolah Pascasarjana Universitas Sumatera Utara Medan, mulai tahun 2007
melalui Beasiswa Pemerintahan Propinsi Sumatera Utara.
Tahun 1992-1997 penulis bertugas sebagai guru bidang studi Biologi di
SMA Taman Siswa Medan, pada tahun 1994-1997 bertugas sebagai guru bidang
studi Biologi di SMA UISU Medan. Tahun 1997 penulis mengajar di SMA
ABSTRAK
Penelitian ini bertujuan untuk mengetahui keanekaragaman dan jenis fungi mikoriza di hutan pantai berdasarkan gradien salinitas. Contoh tanah sebanyak 50 g diambil dari Pantai Pulau Pandang Kabupaten Batu Bara dari setiap petak ukur 20 x 5 m. Contoh tanah disaring dengan satu set saringan bertingkat dan spora yang didapat diidentifikasi. Hasil pengamatan menunjukkan bahwa di lokasi penelitian terdapat 246 spora yang termasuk kedalam 3 genus mikoriza yaitu Glomus, Acaulospora dan Gigaspora. Jenis Glomus merupakan yang paling dominan dan terdapat pada setiap petak ukur. Kepadatan populasi spora makin meningkat sejalan dengan meningkatnya gradien salinitas. Hasil trapping dengan tanaman inang Pueraria javanica menunjukkan jumlah spora yang tinggi dibandingkan dengan spora yang diisolasi dari lapangan. Semua akar contoh tanaman terinfeksi fungi mikoriza. Fungi mikoriza mampu membentuk asosiasi dengan akar tanaman hutan pantai dengan persentase kolonisasi yang beragam, paling rendah dalam akar tanaman Ipomoea pescaprae (13%) dan yang paling tinggi (83,9%) dalam akar tanaman Terminalia catappa.
ABSTRACT
The objective of research was to know the mycorrhiza diversity in coastal forest based on salinity gradient. A 50 g of soil sample has been taken from the Pulau Pandang Beach, District of Batu Bara, from each plot of 20 x 5 m size. The soil sample was filtered through a set of gradient filter, and the spora found was then identified. The result of observation indicated that in location of research, there were 246 spores categorized into three genus of mycorrhizae; Glomus, Acaulospora and Gigaspora. Glomus was the most dominant type and found in each plot. The density of spora population increased with increased salinity gradient. The trapping result with host of Pueraria javanica indicated a high number of spora in comparison to the spora isolated from the site. All roots of plant sample have been infected by mycorrhiza fungi. The mycorrhizae fungi could form the association with roots of coastal forest plants of various colonization percentage, the lowest percentage was in plant roots of Ipomoea pescaprae (13%) and the highest percentage (83,9%) was in plant roots of Terminalia catappa.
BAB I
PENDAHULUAN
1.1 Latar Belakang
Mikoriza merupakan hubungan simbiosis mutualisme antara fungi dengan
perakaran tanaman tingkat tinggi. Kehadiran fungi mikoriza arbuskula (FMA)
penting bagi ketahanan suatu ekosistem, stabilitas tanaman dan pemeliharaan serta
keragaman tumbuhan dan meningkatkan produktivitas tanaman (Moriera et al.,
2007). Selain itu mikoriza membantu kerja perakaran tanaman, mikoriza juga
mampu meningkatkan toleransi tanaman terhadap keadaan lingkungan yang tidak
menguntungkan seperti kekeringan dan salinitas (Brundrett et al., 1996; Delvian,
2003). Fungi mikoriza arbuskula merupakan salah satu jenis fungi tanah yang
memiliki tingkat penyebaran tinggi, karena kemampuannya bersimbiosis dengan
hampir 90% jenis tanaman. Fungi mikoriza pada umumnya dapat ditemukan pada
spesies tanaman tingkat tinggi yang tumbuh pada berbagai tipe habitat dan iklim.
Adapun penyebarannya bervariasi menurut iklim, lingkungan dan tipe
penggunaan lahan (Setiadi, 2001).
Keberadaan fungi mikoriza di alam bersifat kosmopolitan, artinya fungi
mikoriza hampir pasti ada dalam kondisi tanah apapun, seperti di hutan pantai
yang berpasir fungi mikoriza masih dapat tumbuh. Tanah hutan pantai memiliki
faktor pembatas yang berpengaruh terhadap keberadaan fungi mikoriza antara lain
kondisi tanah yang memiliki kadar salinitas yang tinggi (Siradz et al., 2007).
Pada umumnya tanaman asli hutan pantai produktifitasnya sangat rendah.
sedikit sehingga kurang mampu dalam penyerapan zat hara. Kondisi lahan yang
berpasir, temperatur permukaan yang tinggi dan hembusan angin yang kencang
yang berakibat evapotranspirasi sangat tinggi juga sangat mempengaruhi. Daerah
yang kondisi seperti ini peranan fungi mikoriza sangat diperlukan (Siradz et al.,
2007).
Peranan fungi mikoriza pada lahan pasir pantai yaitu membantu dalam
pembentukan agregat tanah. Hifa eksternal mikoriza dapat mengikat butiran pasir
sehingga terbentuk agregat. Agregasi meningkat dengan meningkatnya
perkembangan fungi mikoriza arbuskula, terutama terdapat pada daerah yang
berdekatan dengan zona akar tanaman pionir (Siradz et al., 2007). Peranan
mikoriza pada tanah salin antara lain membantu pertumbuhan tanaman dalam hal
memperbaiki nutrisi tanaman dengan meningkatkan serapan hara terutama fosfor,
sebagai pelindung hayati dan membantu meningkatkan resistensi tanaman
terhadap kekeringan (Brundrett et al., 1996).
Meskipun telah diketahui bahwa peranan fungi mikoriza dalam
pertumbuhan tanaman pada kondisi salin sangat penting, namun jenis-jenis fungi
mikoriza asal tanah salin belum dipelajari secara lengkap. Hal ini ditunjukkan
oleh kebanyakan penelitian pemanfaatan fungi mikoriza pada kondisi salin
menggunakan fungi mikoriza yang berasal dari tanah tidak salin (Hirrel dan
Gerdemann, 1980; Delvian, 2003). Pemanfaatan fungi mikoriza pada tanah salin
merupakan alternatif lain dalam menanggulangi masalah rendahnya produktivitas
tanaman pada tanah salin, disamping itu penggunaan fungi mikoriza ini tidak
diperoleh dan keterbatasan informasi tentang fungi mikoriza pada kondisi salin
maka perlu adanya upaya untuk mengetahui keberadaan dan keanekaragaman
fungi mikoriza arbuskula berdasarkan gradien salinitas.
1.2 Permasalahan
Status fungi mikoriza pada ekosistem hutan pantai kurang mendapatkan
perhatian, padahal hasil penelitian menunjukkan fungi mikoriza dapat membantu
pertumbuhan tanaman pada daerah pantai berpasir yang kondisi tanahnya salin
(Al- Karaki, 2000; Ruiz-Lozano dn Azcon, 2000; Delvian 2003), Tanah yang
bersalinitas tinggi merupakan faktor pembatas budidaya pertanian pada tanah
salin karena cekaman salinitas dapat menghambat pertumbuhan hampir semua
jenis tanaman.
Pemanfaatan mikoriza pada tanah salin sangat penting untuk meningkatkan
pertumbuhan tanaman pada ekosistem pantai, akan tetapi sampai saat ini belum
ada isolat yang berasal dari tanah salin, maka perlu dilakukan upaya dan langkah
awal mempelajari keanekaragaman mikoriza.
Informasi tentang status dan keanekaragaman fungi mikoriza pada hutan
pantai sangat diperlukan sebagai bahan untuk menentukan langkah pengelolaan
dan pemanfaatannya. Berdasarkan permasalahan tersebut perlu dilakukan
penelitian mengenai keanekaragaman fungi mikoriza pada tanah salin di hutan
pantai.
Penelitian ini bertujuan untuk mengetahui status dan keanekaragaman
fungi mikoriza di hutan pantai Pulau Pandang dan hubungannya dengan tingkat
salinitas tanah.
1.4 Manfaat Penelitian
Hasil penelitian ini diharapkan dapat dijadikan sebagai sumber
informasi
mengenai status dan keanekaragaman jenis fungi mikoriza di hutan pantai, yang
berguna untuk pengelolaan dan pemanfaatan selanjutnya.
BAB II
TINJAUAN PUSTAKA
2.1 Fungi Mikoriza Arbuskula
Fungi mikoriza arbuskula merupakan suatu bentuk asosiasi antara jamur
dengan akar tumbuhan tingkat tinggi, yang mencerminkan adanya interaksi
fungsional yang saling menguntungkan antara suatu tumbuhan dengan satu atau
lebih galur mikobion dalam ruang dan waktu. Fungi mikoriza termasuk golongan
endomikoriza. Tipe fungi ini dicirikan oleh hifa yang intraseluler yaitu hifa yang
menembus ke dalam korteks dari satu sel kesel yang lain (Manan, 1993). Diantara
sel-sel terdapat hifa yang membelit atau struktur hifa yang bercabang-cabang
yang
disebut arbuskula. Pembengkakan yang terbentuk pada hifa yang berbentuk oval
disebut vesikula. Arbuskula merupakan tempat pertukaran metabolit antara jamur
dan tanaman. Adanya arbuskula sangat penting untuk mengidentifikasi bahwa
telah terjadi infeksi pada akar tanaman (Scannerini dan Bonfante-Fosolo, 1983
dalam Delvian, 2003), sedangkan vesikula merupakan organ penyimpan makanan
dan berfungsi sebagai propagul (organ reproduktif). Selanjutnya dikatakan bahwa
seluruh endofit dan yang termasuk genus Gigaspora, Scutellospora, Glomus,
Sclerocystis dan Acaulospora mampu membentuk arbuskula. Anatomi sederhana
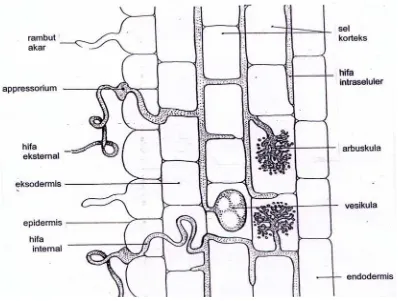
dapat dilihat pada Gambar 1.
Gambar 1. Penampang longitudinal akar yang terinfeksi fungi mikoriza (Brundrett
et al., 1994)
Vesikula menurut Abbott dan Robson (1982), berbentuk globosa dan
berasal dari menggelembungnya hifa internal dari fungi mikoriza. Vesikula
ditemukan baik di dalam maupun di luar lapisan kortek parenkim. Tidak semua
fungi mikoriza membentuk vesikula dalam akar inangnya, seperti Gigaspora dan
Scutellospora. Banyak pendapat tentang fungsi dari vesikula ini, yaitu sebagai
organ reproduksi atau organ yang berfungsi sebagai tempat penyimpanan
makanan yang kemudian diangkut ke dalam sel (Delvian, 2003).
menembus dinding sel inang dan berkembang di dalam sel (Brundrett et al.,1996).
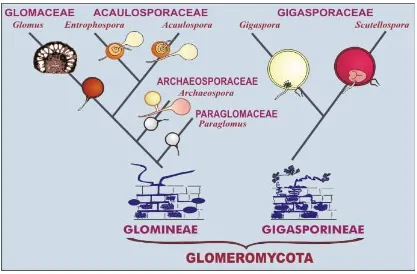
Perkembangan dan taksonomi mikoriza dapat dilihat pada Gambar 2, sedangkan
klasifikasi fungi mikoriza menurut INVAM (2009) dapat dilihat pada Tabel 1.
Gambar 2. Filogeni perkembangan dan taksonomi ordo Glomeromycota (INVAM,
2009)
Tabel 1. Klasifikasi fungi mikoriza arbuskula
Ordo Sub Ordo Famili Genus
2.2 Peranan Fungi Mikoriza Arbuskula
Adanya fungi mikoriza sangat penting bagi ketersediaan unsur hara seperti
P, Mg, K, Fe dan Mn untuk pertumbuhan tanaman. Hal ini terjadi melalui
pembentukan hifa pada permukaan akar yang berfungsi sebagai perpanjangan akar
terutama di daerah yang kondisinya miskin unsur hara, pH rendah dan kurang air.
Akar tanaman bermikoriza ternyata meningkatkan penyerapan seng dan sulfur
dari dalam tanah lebih cepat daripada tanaman yang tidak bermikoriza (Abbot dan
Robson 1984). Manfaat fungi mikoriza ini secara nyata terlihat jika kondisi
tanahnya miskin hara atau kondisi kering, sedangkan pada kondisi tanah yang
subur peran fungi ini tidak begitu nyata (Setiadi, 2001; Lakitan, 2000).
Menurut Siradz et al., (2007), hampir semua tanaman asli lahan pantai
terinfeksi oleh fungi mikoriza. Hubungan antara jumlah spora dengan
pertumbuhan tanaman menunjukkan hubungan positif dalam hal menyerap unsur
hara. Hubungan yang positif tersebut cukup memberikan indikasi yang jelas
tentang peluang penggunaan fungi mikoriza untuk meningkatkan pertumbuhan
tanaman, membantu memperbaiki dan meningkatkan sifat-sifat struktur agregat
tanah.
Menurut Marx (1982), akar tanaman yang terbungkus oleh fungi mikoriza
menyebabkan akar tersebut terhindar dari serangan penyakit dan hama. Infeksi
patogen terhambat, disamping itu fungi mikoriza menggunakan semua kelebihan
dan eksudat akar lainnya, sehingga tercipta lingkungan yang tidak cocok bagi
2.3 Distribusi dan Ekologi Fungi Mikoriza Arbuskula
Fungi mikoriza biasanya tersebar dengan berbagai cara. Penyebaran aktif
miselia melalui tanah, setelah infeksi di akar hifa berkembang di daerah perakaran
pada tanah dan terbentuk struktur fungi, diantaranya miselium eksternal akar
merupakan organ yang sangat penting dalam menyerap unsur hara dan
mentransferkan ke tanaman, sedangkan penyebaran pasif dapat dilakukan oleh
beberapa hewan dan juga angin (Setiadi, 2001). Penyebaran fungi mikoriza
melalui inokulasi agak berkurang pada tanah yang sudah bermikoriza, tetapi
meningkat pada tanah yang tidak bermikoriza.
Perbedaan lokasi dan rizosfer menyebabkan perbedaan keanekaragaman
spesies dan populasi fungi mikoriza, misalnya yang didominasi oleh fraksi
lempung berdebu merupakan tanah yang baik bagi perkembangan Glomus (Baon,
1998), begitu juga dengan tanah mangrove yang bercirikan tanah berlumpur dan
cenderung liat hanya Glomus sp. yang dapat hidup, sedangkan tanah yang berpasir
genus Acaulospora dan Gigaspora ditemukan dalam jumlah yang tinggi. Sebaran
kedua genus tersebut ternyata berkebalikan apabila ditinjau posisinya dari garis
pantai. Kepadatan populasi Acaulospora meningkat sejalan dengan jarak dari
garis pantai, artinya makin jauh dari garis pantai populasi Acaulospora makin
tinggi. Kecenderungan sebaliknya diperlihatkan oleh Gigaspora yang makin jauh
dari garis pantai populasinya semakin menurun (Siradz et al., 2007).
Menurut Moreira (2007), pada ekosistem hutan asli Acaulospora
Glomus macrocarpum yang menunjukkan jumlah spora yang paling banyak,
sedangkan daerah yang dihutankan kembali jenis yang paling banyak adalah
Glomus macrocarpum dan Archeospora gerdemanni. Jenis-jenis ini
menyesuaikan diri pada lingkungan dan menunjukkan toleransi yang tinggi dan
adaptasi yang berbeda.
2.4 Faktor Yang Mempengaruhi Keberadaan FMA
Keberadaan spora FMA dipengaruhi oleh beberapa faktor lingkungan
seperti :
1. Cahaya
Adanya naungan yang berlebihan terutama untuk tanaman yang senang
cahaya dapat mengurangi infeksi akar dan produksi spora, selain itu respon
tanaman terhadap fungi mikoriza akan berkurang. Hal ini disebabkan adanya
hambatan pertumbuhan dan perkembangan internal hifa dalam akar yang
berakibat terbatasnya perkembangan eksternal hifa pada rizosfer (Setiadi, 2001).
2. Suhu
Suhu berpengaruh terhadap infeksi yakni pada perkembangan spora,
penetrasi hifa pada sel akar dan perkembangan pada korteks akar, selain itu suhu
juga berpengaruh pada ketahanan dan simbiosis. Semakin tinggi suhu semakin
besar terbentuknya kolonisasi dan meningkatnya produksi spora. Schenk dan
yakni pada suhu 30oC tetapi untuk koloni miselia terbaik berada pada suhu 28–
34oC, sedangkan perkembangan bagi vesikula pada suhu 35oC.
3. Kandungan air tanah
Kandungan air tanah dapat berpengaruh baik secara langsung atau tidak
langsung terhadap infeksi dan pertumbuhan fungi mikoriza. Pengaruh secara
langsung tanaman bermikoriza dapat memperbaiki dan meningkatkan kapasitas
serapan air. Sedangkan pengaruh tidak langsung karena adanya miselia eksternal
menyebabkan fungi mikoriza efektif dalam mengagregasi butir-butir tanah,
kemampuan tanah menyerap air meningkat. Penjenuhan air tanah yang lama
berpotensi mengurangi pertumbuhan dan infeksi fungi mikoriza karena kondisi
yang anaerob. Daniels dan Trappe (1980) menggunakan Glomus epigaeum
dikecambahkan pada lempung berdebu pada berbagai kandungan air. Glomus
epigaeum ternyata berkecambah paling baik pada kandungan air di antara
kapasitas lapang dan kandungan air jenuh.
4. pH Tanah
Fungi mikoriza pada umumnya lebih tahan terhadap perubahan pH tanah.
Meskipun demikian adaptasi masing-masing spesies fungi mikoriza terhadap pH
tanah berbeda-beda, karena pH tanah mempengaruhi perkecambahan,
perkembangan dan peran mikoriza terhadap pertumbuhan tanaman (Maas dan
Nieman, 1978).
pH optimum untuk perkembangan fungi mikoriza berbeda-beda
berpengaruh langsung terhadap aktivitas enzim yang berperan dalam
perkecambahan spora fungi mikoriza. Misalnya Glomus mosseae biasanya pada
tanah alkali dapat berkecambah dengan baik pada air atau pada soil extract agar
pada pH 6-9. Spora Gigaspora coralloidea
dan Gigaspora heterogama dari jenis yang lebih tahan asam dapat berkecambah
dengan baik pada pH 4-6. Glomus epigaeum perkecambahannya lebih baik pada
pH 6-8.
5. Bahan organik
Bahan organik merupakan salah satu komponen dalam tanah yang penting
disamping air dan udara. Jumlah spora FMA berhubungan erat dengan kandungan
bahan organik dalam tanah. Jumlah maksimum spora ditemukan pada tanah-tanah
yang mengandung bahan organik 1-2% sedangkan pada tanah-tanah berbahan
organik kurang dari 0,5% kandungan spora sangat rendah (Pujianto, 2001).
6. Logam berat dan unsur lain
Adanya logam berat dalam larutan tanah dapat mempengaruhi
perkembangan mikoriza. Beberapa spesies mikoriza arbuskular diketahui mampu
beradaptasi dengan tanah yang tercemar seng (Zn), tetapi sebagian besar spesies
mikoriza peka terhadap kandungan Zn yang tinggi. Pada beberapa penelitian lain
diketahui pula strain-strain fungi mikoriza tertentu toleran terhadap kandungan
2.5 Fungi Mikoriza Arbuskula dalam Tanah Salin
Salah satu faktor lingkungan yang mempengaruhi pertumbuhan
tanaman
adalah salinitas tanah. Tanah bersalinitas tinggi biasanya banyak ditemukan di
daerah mangrove dan hutan pantai. Pengaruh salinitas paling umum adalah
terhambatnya pertumbuhan tanaman. Peningkatan konsentrasi garam dalam tanah
menyebabkan terjadinya perubahan morfologi dan fisiologi tanaman dengan
metabolisme yang abnormal akibat kandungan garam di jaringan tanaman, selain
itu terjadi penurunan potensial osmotik tanah sehingga menyulitkan penyerapan
air dan hara bagi tanaman, merusak kloroplas dan mengganggu proses fotosintesis
yang akhirnya menekan pertumbuhan dan produksi tanaman (Khattak et al.,
1991).
Kadar garam yang tinggi dalam larutan tanah di daerah perakaran tanaman
menyebabkan tekanan osmotik yang tinggi dan berkurangnya ketersediaan unsur
kalium bagi tanaman (Bernstein, 1981 dalam Delvian, 2003). Untuk mengetahui
bagaimana pengaruh salinitas terhadap pembentukan fungi mikoriza perlu
diketahui bagaimana pengaruh salinitas terhadap pertumbuhan tanaman inang.
Beberapa studi menyimpulkan bahwa pembentukan fungi mikoriza menurun
dengan bertambahnya salinitas tanah. Peningkatan level salinitas tanah
menyebabkan terjadinya penurunan pertumbuhan tajuk sehingga mengakibatkan
penurunan area fotosintesis pada tanaman (Hirrel dan Gerderman, 1980 dalam
Menurut Ruiz-Lozano dan Azcoon, (2000), dikemukakan bahwa fungi
mikoriza seperti Glomus sp mampu hidup dan berkembang pada kondisi salinitas
yang tinggi. Beberapa penelitian menunjukkan bahwa fungi mikoriza dapat
meningkatkan pertumbuhan tanaman yang ditanam pada habitat salin. Tanaman
bawang merah yang diinokulasikan dengan fungi mikoriza dari spesies Glomus
ternyata memiliki berat bulbus dan bobot kering bawang serta total serapan hara
yang lebih tinggi dibandingkan dengan tanaman yang tidak diinokulasikan baik
pada tingkat salinitas rendah (-0,06 Mpa), sedang (-0,20 Mpa) dan tinggi (-0,4
Mpa). Namun demikian infeksi fungi mikoriza cenderung menurun secara linier
BAB III
BAHAN DAN METODA
3.1 Tempat dan Waktu
Pengambilan contoh tanah dan akar tanaman dilakukan di kawasan hutan
pantai Pulau Pandang Kabupaten Batu Bara pada bulan Maret 2009. Ekstraksi dan
identifikasi spora serta penghitungan kolonisasi fungi mikoriza pada akar tanaman
contoh dilakukan di Laboratorium Biologi Tanah dan Laboratorium Bioteknologi
Kehutanan Fakultas Pertanian Universitas Sumatera Utara.
3.2 Bahan dan Alat
Dalam penelitian ini digunakan contoh tanah dan akar tanaman dari
hutan pantai. Untuk ekstraksi dan identifikasi spora fungi mikoriza digunakan
bahan berupa larutan glukosa 60%, larutan Melzers sebagai bahan pewarna spora
dan larutan polyvinyl lacto glycerol (PVLG) sebagai bahan pengawet spora.
Sedangkan untuk pewarnaan akar digunakan bahan-bahan kimia antara lain, yaitu
KOH 10%, HCl 2%, larutan pewarna (Staining: gliserol, asam laktat dan trypan
blue), dan aquades.
Alat-alat yang digunakan untuk pengambilan contoh tanah dan akar
tanaman adalah kompas, tali plastik, cangkul, kantong plastik dan spidol serta
kertas label, sedangkan peralatan untuk pengamatan di laboratorium adalah
pinset spora, mikroskop binokuler, mikroskop cahaya, kaca preparat, dan kaca
penutup.
3.3 Pengambilan Contoh Tanah dan Akar
Pengambilan contoh tanah dan akar tanaman menggunakan metoda jalur
atas dasar gradien salinitas. Jalur dibuat sepanjang 120 m dengan lebar 5 m dari
garis pantai menuju ke daratan. Jalur dibagi dalam 6 petak dengan ukuran panjang
setiap petak 20 m dan lebar 5 m. Jumlah jalur yang dibuat sebanyak 3 jalur
dengan jarak antar jalur sekitar 200 m. Pada masing-masing petak dalam jalur
diambil contoh tanah sebanyak 600-700 g dari zona rizosfir, yaitu pada kedalaman
0-20 cm. Selain itu juga diambil 3 jenis anakan yang dominan pada setiap petak
ukur untuk mempelajari kolonisasi fungi mikoriza pada setiap petak ukur. Dari
contoh tanah yang diambil juga dilakukan analisis tingkat salinitas tanah dengan
metoda daya hantar listrik.
3.4 Ekstraksi dan Identifikasi Spora Fungi Mikoriza Arbuskula
Ekstraksi spora fungi mikoriza dilakukan untuk memisahkan spora dari
sampel tanah dan mengidentifikasinya yaitu dengan teknik tuang saring dari
Pacioni (1992) dan dilanjutkan dengan sentrifuse (Brundrett et al., 1996).
Prosedur kerja secara lengkap adalah sebagai berikut: tanah sebanyak 50 g
dicampur dengan 200-300 ml liter air dan diaduk sampai butiran tanah hancur.
Campuran tanah dan air tersebut disaring dalam 1 set saringan dengan ukuran 425
µm, 212 µm, 106 µm dan 53 µm secara berurutan dari atas ke bawah. Dari
lolos. Partikel yang tertahan pada saringan terbawah dipindahkan ke dalam tabung
sentrifuse lalu tambahkan larutan glukosa 60% yang diletakkan pada bagian
bawah dari larutan tanah dengan menggunakan pipet. Tabung sentrifuse ditutup
rapat dan disentrifugasi dengan kecepatan 2500 rpm selama 3 menit, kemudian
larutan supernatan dituang ke dalam saringan 53 µm dan dicuci dengan air untuk
menghilangkan glukosa yang tersisa dalam saringan lalu dituangkan ke dalam
cawan petri dan kemudian diamati bawah mikroskop. Selanjutnya spora yang
diperoleh dihitung jumlahnya, kemudian diletakkan dalam larutan pewarnaan
Melzers dan pengawetan polyvinil lacto glyserol yang terpisah pada satu kaca
preparat. Spora tersebut dipecahkan secara hati-hati dengan menekan kaca
penutup dengan menggunakan ujung lidi, adanya perubahan warna spora adalah
salah satu indikator untuk menentukan jenis spora.
3.5 Kolonisasi FMA pada Akar Tanaman Sampel
Akar halus segar dengan diameter 0,5 mm dicuci dengan air mengalir
sampai
bersih, lalu akar sampel direndam dalam larutan KOH 10% selama 24 jam.
Kemudian larutan KOH dibuang dan akar dicuci dengan air lalu direndam
dengan larutan HCl 2% selama 24 jam (Kormanik dan McGraw, 1982).
Selanjutnya akar sampel direndam dalam larutan trypan blue dan digantikan
dengan larutan lacto glycerol untuk proses destaining (pengurangan warna).
Penghitungan persentase kolonisasi akar menggunakan metode panjang akar
kaca preparat (Giovanneti dan Mosse, 1980). Secara skematis alur kerja kolonisasi
fungi mikoriza pada akar tanaman disajikan pada lampiran 2.
Persentase kolonisasi akar dihitung dengan menggunakan rumus.
%
3.6 Pemerangkapan (Trapping culture)
Teknik pemerangkapan digunakan mengikuti metode Brundreet et al.,
(1994).
Setiap contoh tanah dibuat 3 pot kultur sehingga terdapat 90 pot kultur. Media
tanam pot kultur berupa campuran contoh tanah 50 g dan pasir sebanyak 150 g,
selanjutnya benih Purieria javanica ditaruh dalam lubang tanam yang sudah diisi
dengan pasir-tanah dan ditutup lagi dengan pasir (lampiran 3). Tanaman diberi
larutan NaCl dengan konsentrasi yang sesuai dengan data di lapangan dan
frekuensi pemberian 1 x 2 minggu. Perlakuan ini bertujuan untuk mengurangi
kemungkinan terjadinya perubahan salinitas tanah dibandingkan dengan
ekosistem aslinya. Setiap 2 hari sekali disiram dan diberi hara Hyponex merah
dengan konsentrasi 1 g. l-1 setiap minggu. Pemeliharaan tanaman dilakukan
selama 8 minggu, selanjutnya dibiarkan tanaman sampai mati, setelah itu
3.7 Pengamatan
Hasil pengamatan yang diperoleh dilakukan secara deskriptif dan
menyajikan
tabel-tabel hasil identifikasi genus-genus fungi mikoriza serta nama tanaman
yang menjadi inangnya. Parameter pengukuran adalah sebagai berikut:
1). Tingkat salinitas tanah dan pH
Untuk mengetahui tingkat salinitas tanah maka dilakukan pengukuran
dengan menggunakan metode daya hantar listrik.
2). Jumlah spora dan tipe spora
Untuk menghitung kepadatan spora dan tipe spora maka dilakukan dengan
pengamatan preparat sesuai dengan yang sudah dijelaskan pada poin 3.4. Selain
itu data yang diperoleh dihitung frekuensi mutlak (FM), frekuensi relatif (FR),
secara rinci rumus tersebut adalah sebagai berikut:
1. Frekuensi Mutlak (FM - %)
2. Frekuensi Relatif (FR – 100%)
3). Persentase kolonisasi akar
Penghitungan kolonisasi akar menggunakan metode panjang akar seperti
BAB IV
HASIL DAN PEMBAHASAN
4.1 Salinitas tanah dan sifat kimia tanah
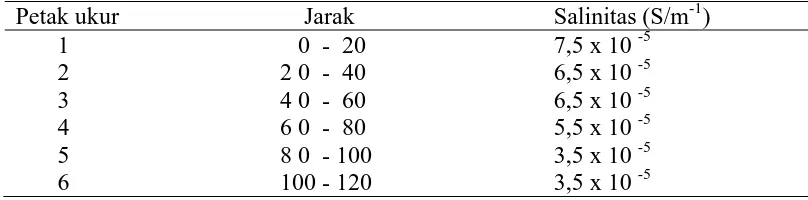
Data yang diperoleh menunjukkan bahwa terjadi penurunan tingkat salinitas
tanah seiring dengan semakin jauhnya letak petak pengambilan contoh tanah dari
garis pantai. Tingkat salinitas tertinggi terdapat pada petak ukur 1 (0-20 m) dari
garis pantai dan terendah pada petak ukur 6 (100-120 m) dari garis pantai. Hasil
pengukuran salinitas tanah disajikan pada Tabel 2.
Tabel 2. Tingkat salinitas tanah pada lokasi penelitian
Petak ukur Jarak Salinitas (S/m-1)
Tanah hutan pantai Pulau Pandang mempunyai tingkat salinitas yang
berkisar antara 3,5-7,5 x 10-5 S/cm. Menurut Chapman (1975) nilai salinitas suatu
lokasi ditentukan oleh konsentrasi NaCl, NaCO3 atau garam-garam Mg. Untuk
daerah pantai sumber utama salinitas tanah adalah air laut, NaCl adalah penyusun
utamanya. Kandungan Na dan Cl dalam air laut menurut Carter (1975)
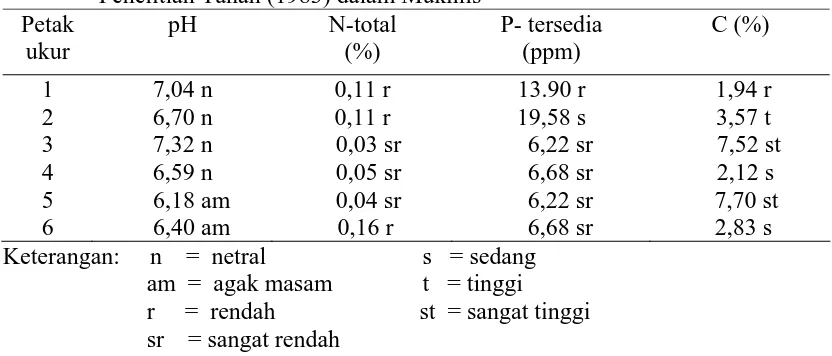
masing-masing adalah 30,61% dan 55,04%. Adanya perbedaan antara pH, N, P tersedia
dan C organik dari
data yang diperoleh menunjukkan variasi dalam hal sifat kimia dari tiap petak
tanah. Hasil analisis dari contoh tanah diperlihatkan pada Tabel 3.
Hasil analisis kimia tanah menunjukkan bahwa variasi sifat-sifat kimianya
cukup berarti. Nilai pH tanah tampaknya tidak berhubungan dengan tingkat
salinitas tanah karena adanya peningkatan salinitas tanah tidak diikuti oleh
peningkatan pH. Konsentrasi P tersedia dalam tanah erat hubungannya dengan
sifat kimia tanah lainnya, khususnya pH yaitu makin tinggi pH tanah maka
ketersediaan P di dalam tanah makin besar (Winarso dan Setiawati, 2003).
Kandungan fosfor agak tinggi pada petak ukur 1 dan 2 dibandingkan dengan petak
ukur yang lain. Pada petak ini ditemui jumlah spora sedikit. Apabila kadar fosfor
dalam kandungan tanah tinggi, biasanya fungi mikoriza terdapat sedikit di sekitar
rizosfer, dan sebaliknya fungi mikoriza dapat berkembang dengan baik pada tanah
yang mempunyai fosfor lebih rendah dan aerasi tanah yang lebih baik (Husin et
Fungi mikoriza membantu penyerapan air dan hara terutama fosfor. Tingkat
kolonisasi diatur oleh fosfor dan nitrogen. Tanah yang kurang subur, kolonisasi
akan maksimal terutama bila pada tanah tersebut tersedia fosfor. Pada tanah yang
unsur fosfor sedikit maka mikoriza akan bekerja maksimal. Kolonisasi mikoriza
menurun seiring dengan meningkatnya kesuburan tanah (Delvian, 2003).
Miselium mikoriza dapat menyerap hara terutama fosfor melalui enzim
Pospatase. Adanya enzim ini ion-ion fosfor yang terikat kuat pada mineral tanah
seperti aluminium dapat diuraikan sehingga fosfor lebih tersedia di tanah dan
dapat diserap oleh tanaman (Daniels, 1984). Unsur hara yang lain juga
mempengaruhi pertumbuhan mikoriza. Tingkat nitrogen tanah yang tinggi
berpengaruh negatif terhadap pembentukan dan perangsangan pertumbuhan fungi
mikoriza, pengaruh nitrogen terhadap fungi mikoriza juga dipengaruh kuat oleh
ketersediaan fosfor rendah atau tinggi didalam tanah. Pemupukan nitrogen dapat
mengurangi infeksi mikoriza, namun pada tingkat fosfor yang sedang didalam
tanah penambahan nitrogen meningkatkan infeksi fungi mikoriza (Safir dan
Duniway 1982).
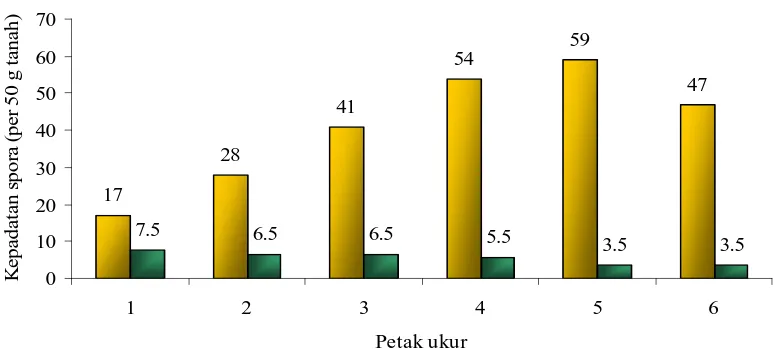
4.2 Kepadatan Spora
Hasil pengamatan menunjukkan kepadatan spora meningkat sejalan dengan
menurunnya salinitas tanah meskipun pada petak terakhir terjadi penurunan
jumlah spora yang diperoleh. Jumlah spora terendah ditemukan pada petak 1 (17
spora). Data pengamatan spora (jumlah spora per 50 gram tanah) dan
hubungannya dengan tingkat salinitas dapat dilihat pada Tabel 4.
Tabel 4. Jumlah tipe spora berdasarkan tingkat salinitas tanah
Petak ukur Salinitas (S.m-1) Jumlah spora
Dari data yang diperoleh ternyata pada petak 5 jumlah spora lebih tinggi
daripada
petak 6. Adanya perbedaan ini diduga pada petak ukur 6 spora belum
banyak
bersporulasi dan ada kecenderungan dipengaruhi oleh musim (curah hujan).
Keanekaragaman dan kepadatan spora fungi mikoriza selalu berubah dengan
perubahan waktu pengamatan, jenis inang dan tingkat salinitas. Hal ini
menunjukkan bahwa keanekaragaman fungi mikoriza dipengaruhi oleh faktor
perubahan musim seperti curah hujan dan tanaman inang (Siguenza et al., 1996).
Hasil pengamatan isolasi spora di lapangan berdasarkan gradien salinitas
ditemukan 3 jenis spora, pada petak ukur terdepan dari garis pantai dijumpai jenis
dan jumlah spora yang sedikit. Hal ini diduga dipengaruhi oleh tingginya tingkat
salinitas, artinya tingginya salinitas tanah berpengaruh negatif terhadap kepadatan
populasi spora. Menurut Junifer dan Abbot (1993) salinitas tanah mempengaruhi
Hasil identifikasi menunjukkan ada 3 genus mikoriza yang berkembang
pada lahan pasir pantai Pulau Pandang yaitu Glomus, Acaulospora, dan
Gigaspora
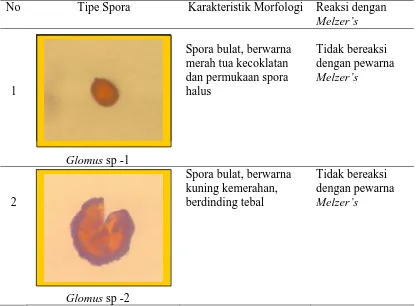
(Tabel 5). Identifikasi dilakukan berdasarkan perbedaan ciri, karakteristik
morfologi (bentuk ketebalan dinding sel, ada tidaknya sublending hifa, kehalusan
permukaan dan reaksi spora terhadap Melzers.
Tabel 5. Jumlah spora dari lapangan
Petak ukur Glomus Acaulospora Gigaspora
1
Sebaran anggota genus Glomus merata pada setiap petak ukur. Kepadatan
populasi spora makin meningkat sejalan dengan berkurangnya tingkat salinitas
tanah. Genus Glomus memiliki kepadatan tinggi. Hal ini menunjukkan bahwa
Glomus bersifat adaptif dan menunjukkan toleransi yang tinggi pada ekosistem
pantai yang berpasir. Studi keanekaragaman fungi mikoriza pada tanah salin telah
banyak dilakukan oleh peneliti, juga menunjukkan bahwa Glomus adalah jenis
dengan jumlah spora yang dominan (Siradz, 2007; Delvian, 2003; Koske dan
Tews, 1987). Menurut Moreira (2007) jenis Glomus menunjukkan toleransi yang
tinggi pada semua habitat di alam, karena jenis Glomus ditemui dalam jumlah
yang besar pada beberapa ekosistem. Nilai kepadatan spora yang diisolasi dari
pada Gambar 3, dari gambar tersebut tampak bahwa kepadatan spora meningkat
sejalan dengan menurunnya salinitas tanah.
17
Kepadatan spora (per 50 g tanah) Salinitas tanah (S.m-1)
Gambar 3. Kepadatan spora dari lapangan dan hubungannya dengan salinitas
tanah.
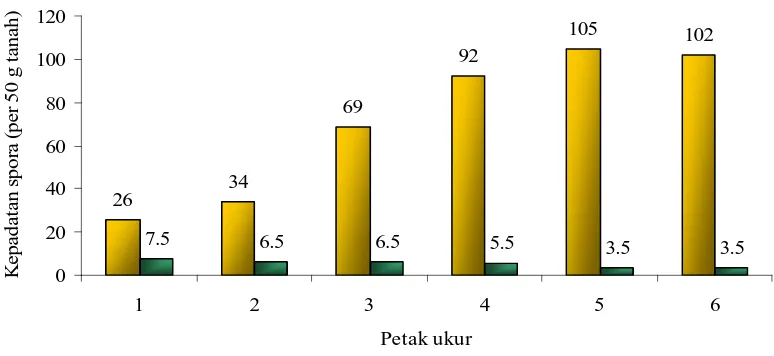
Hasil pengamatan dari pemerangkapan (trapping) menunjukkan bahwa
kepadatan populasi spora sangat meningkat dibandingkan dengan populasi spora
di lapangan, disebabkan karena fungi mikoriza yang diisolasi di lapangan telah
berasosiasi dengan Pueraria javanica sebagai tanaman inangnya. Hal ini
menunjukkan bahwa fungi mikoriza yang pada saat diisolasi di lapangan diduga
belum bersporulasi sehingga dengan dilakukan pemerangkapan memberikan
kesempatan propagul fungi mikoriza yang masih dorman untuk tumbuh dan
berkembang membentuk spora, sehingga keanekaragaman dan jumlah fungi
mikoriza dijumpai lebih banyak dan mendapatkan data yang akurat. Hasilnya
pada Tabel 6.
Tabel 6. Jumlah spora hasil pemerangkapan (trapping)
Petak ukur Glomus Acaulospora Gigaspora
1 26 - - 2 32 1 1 3 64 3 2 4 87 3 2 5 99 4 2 6 95 4 3
Data hasil pemerangkapan (trapping) juga menunjukkan bahwa jenis spora
didominasi oleh Glomus. Hal ini menunjukkan bahwa jenis Glomus adalah jenis
yang dominan pada Hutan Pantai Pulau Pandang. Tingkat salinitas tanah juga
mempengaruhi jumlah jenis spora fungi mikoriza yang ditemukan (Gambar 4).
Berdasarkan gambar 4 dapat dilihat bahwa jumlah spora hasil trapping sangat
meningkat dibandingkan dengan jumlah spora hasil isolasi dari lapangan (Gambar
3). Kepadatan populasi spora juga makin meningkat sejalan dengan menurunnya
salinitas tanah dan meningkatnya jarak dari garis pantai kearah pedalaman.
Kim dan Weber (1985) menyimpulkan bahwa kepadatan spora fungi
mikoriza berhubungan erat dengan salinitas tanah, kepadatan spora fungi mikoriza
akan menurun sejalan dengan peningkatan salinitas tanah. Hal ini karena fungi
mikoriza berhubungan erat dengan tanaman inang. Fungi mikoriza dalam
simbiosisnya sangat tergantung pada nutrisi dari karbohidrat hasil fotosintesis
tanaman inang, Pengaruh salinitas terhadap fotosintesis menyebabkan terjadinya
perubahan konsentrasi osmotik dari cairan daun, potensial air dan pembukaan
dan perkembangan fungi mikoriza yang terdapat pada perakaran tanaman.
(Thomson et al., 1990).
26
Kepadatan spora (per 50 g tanah) Salinitas tanah (S.m-1)
Gambar 4. Kepadatan spora hasil trapping dan hubungannya dengan salinitas
Hasil penghitungan Frekuensi mutlak (FM) dan Frekuensi relatif (FR) jenis
spora fungi mikoriza di lapangan dan trapping menunjukkan peran yang hampir
sama, seperti yang tampak pada Tabel 7 dan 8. Dari data ini dapat dilihat bahwa
spora fungi mikoriza jenis Glomus mempunyai FM dan FR tertinggi, begitu juga
dengan hasil data yang diperoleh dari hasil trapping. Tingginya jumlah Glomus
yang ditemukan diduga karena jenis Glomus lebih banyak daripada jenis spora
lainnya. Dari 172 jenis fungi mikoriza yang sudah diidentifikasi diketahui Glomus
adalah jenis yang paling dominan sehingga berpengaruh terhadap sebarannya di
FM FR FM FR FM FR FM FR FM FR FM FR
berapa tipe spora fungi mikoriza dan deskripsinya dalam contoh tanah dari
isolasi di lapangan dapat dilihat pada Tabel 9.
Tabel 9. Karakteristik spora yang ditemukan di lapangan
7
11
Acaulospora sp - 3
Acaulospora sp -3
dan berlapis-lapis bagian dalam spora berwarna lebih gelap
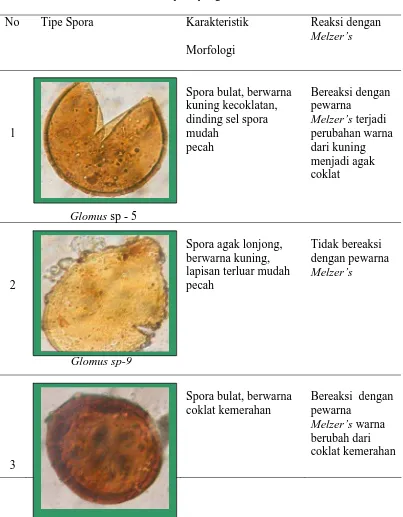
Beberapa tipe spora fungi mikoriza dan deskripsinya dalam contoh tanah dari
hasil trapping dapat dilihat pada Tabel 10.
Tabel 10. Karakteristik spora yang ditemukan dari trapping
No Tipe Spora Karakteristik
Acaulospora sp -4
16
Glomus sp -19
Spora bulat, berwarna coklat kemerahan, permukaan spora mudah pecah
Bereaksi dengan pewarna
4.3 Persentase kolonisasi akar
Hasil pengamatan akar tanaman hutan pantai Pulau Pandang ditemukan
adanya asosiasi akar dengan fungi mikoriza membentuk kolonisasi. Penginfeksian
mikoriza dapat ditandai dengan adanya hifa menembus sel epidermis melalui
permukaan akar atau rambut-rambut akar, sehingga kelihatan jelas hifa dan
vesikula (Gambar 5). Pada pengamatan ini tidak dijumpai arbuskula.
v
v A
A
v
B
h
B
h
Gambar 5. A. Penampang akar Buchanania arborescen, v (vesikula) B. Penampang
akar Terminalia catappa, h (hifa)
Persentase kolonisasi akar yang terinfeksi mikoriza beragam, persentase
paling rendah dalam akar tanaman Ipomoea pescaprae (13%) dan yang
(83,9%) dalam akar tanaman Terminalia catappa. Kolonisasi fungi mikoriza pada
akar tanaman paling tinggi terdapat pada tingkat salinitas yang rendah. Spesies
yang sama pada tingkat salinitas yang berbeda mempunyai persentase kolonisasi
yang berbeda pula tergantung pada tingkat salinitasnya, seperti disajikan pada
Tabel 11.
Hasil pengamatan menunjukkan bahwa persentase kolonisasi pada akar
tanaman inang bervariasi. Kolonisasi meningkat dengan semakin jauhnya letak
tanaman dari garis pantai walaupun ada kecendrungan beberapa tanaman terjadi
fluktuasi, seperti ditampilkan pada Gambar 6. Hasil tersebut menunjukkan bahwa
tanaman yang berada pada petak ukur terdepan mempunyai persentase kolonisasi
yang rendah dan makin meningkat dengan semakin jauhnya letak tanaman dari
garis pantai, walaupun ada beberapa petak ukur yang jauh dari garis pantai
memiliki derajat infeksi yang sedikit.
14.3
Rata-rata kolonisas i (%) Salinitas tanah (S.m -1)
Gambar 6. Hubungan persentase kolonisasi mikoriza dengan tingkat salinitas
tanah
No Spesies
Persentase kolonisasi fungi mikoriza bervariasi dan berfluktuasi pada setiap
tanaman dalam pengambilan contoh tanah. Penurunan persentase kolonisasi fungi
mikoriza pada perakaran tanaman dengan adanya peningkatan salinitas tanah
diduga disebabkan oleh perubahan fisiologi tanaman yang akan mempengaruhi
simbionnya secara langsung maupun tidak langsung.
Variasi kolonisasi fungi mikoriza dipengaruhi oleh tingkat salinitas tanah.
Salinitas berpengaruh negatif terhadap perkecambahan spora dan perkembangan
hifa (Junifer dan Abbot, 2003). Lain halnya dengan pendapat Rozema et al.
(1986) salinitas yang meningkat tidak terlalu mempengaruhi tingkat infeksi
mikoriza tetapi di bawah salinitas yang rendah (150 mM NaCl) yang dikombinasi
dengan kondisi-kondisi penggenangan air persentase infeksi mikoriza mengalami
penurunan. Hal ini diduga disebabkan karena keadaan penggenangan air yang
bersifat anaerobik.
Tingkat kolonisasi fungi mikoriza tergantung pada pertumbuhan tanaman
dan produksi nutrisi karbohidrat dalam tanaman inang, adanya faktor yang
mempengaruhi produksi karbohidrat dan translokasinya ke akar bisa
mempengaruhi jumlah kolonisasi mikoriza (Thomson et al.,1990). Menurut Abbot
et al. (1992) setiap jenis fungi mikoriza mempunyai pola kolonisasi yang berbeda
Kolonisasi fungi mikoriza pada beberapa tanaman yang tahan pada garam telah
banyak dilaporkan di lapangan (Hirrel dan Gerdemann, 1980). Menurut Setiadi
(2001), kriteria persentase kolonisasi akar dari 51-75 termasuk tinggi sedangkan
persentase kolonisasi mikoriza pada akar tanaman di hutan pantai berkisar
BAB V
KESIMPULAN DAN SARAN
5.1 Kesimpulan
1. Status dan keanekaragaman fungi mikoriza di hutan pantai Pulau
Pandang dipengaruhi oleh salinitas tanah
2. Penurunan tingkat salinitas tanah diikuti oleh peningkatan kepadatan spora
dan persentase kolonisasi fungi mikoriza pada akar tanaman.
3. Jenis-jenis fungi mikoriza yang terdapat di pantai Pulau Pandang adalah
Glomus, Acaulospora dan Gigaspora. Spora mikoriza yang dominan
ditemui adalah jenis Glomus.
4. Persentase kolonisasi mikoriza pada akar tanaman di hutan pantai
berkisar antara rendah sampai tinggi
5. Hasil trapping menunjukkan bahwa kepadatan spora lebih tinggi daripada
spora yang diisolasi dari lapangan.
5.2 Saran
1. Dalam studi keanekaragaman fungi mikoriza perlu diadakan trapping,
karena pada saat eksplorasi fungi mikoriza di lapangan mungkin
banyak mikoriza yang belum bersporulasi sehingga dengan adanya
trapping akan diperoleh keanekaragaman fungi mikoriza lebih banyak.
2. Hasil penelitian ini hanya mendapatkan data keanekaragaman mikoriza
belum mengenai potensi mikoriza, jadi perlu dilanjutkan studi potensi
DAFTAR PUSTAKA
Abbott LK dan Robson AD. 1982. The role of VA mycorrhizae fungi agriculture and the selection of fungi for inoculation. Aust. J. Agric. Res. 33 : 389
1984. The effect of mycorrhizae on plant growth. Hlm: 113-130. Dalam: Powell CL dan Bagyaraj DJ. (Eds). Vesicular- Arbuscular mycorrhiza. CRC Press. Inc. Boca Raton. Florida.
Abbot LK, Robson AD, Jasper DA, dan Gazey C. 1992. What is the role of VA mycorrhyzal hypae in soil?. Hlm: 37-41. Dalam: Read D J, D H Lewis, A H Fitter, dan I J Alexander (Eds). Mycorrhizas in ecosystem. C.A.B. International.
Al-karaki GN. 2000 Growth, water use efficiency, and mineral acquisition by tomato cultivars grown under salt stress. J. Plant. 23 : 1-8.
Baon JB 1998. Peranan mikoriza VA pada kopi dan kakao. Makalah disampaikan dalam workshop aplikasi fungi mikoriza arbuskula pada tanaman pertanian, perkebunan dan kehutanan. Bogor.
Bernstein L 1981. Effects of salinity dan sodicity on plant growth. Annu. Rev. Phytopathol. 13 : 295-312.
Brundrett MC, Bougher N, Dells B, Grove T, dan Malajozuk N. 1996. Working with mycorrhizas in forestry and agriculture. ACIAR. Canberra. 374 hlm.
Brundrett MC, Melville L dan Peterson L. 1994. Practical methods in mycorrhyza research. Mycology publication. Ontario, Canada. 161 hlm.
Carter DL 1975. Problems of salinity in agriculture. Dalam: A. Poljakoff-Mayber dan J. Gale (Eds) Plants in saline environments. Springer – Verlag - Berlin, Heildelberg. New York. Hlm: 25-38.
Champman VJ 1975. The salinity problem in general, its importance and distribution with special reference to natural halophytes. Dalam: A. Poljakoff-Mayber dan J. Gale (Eds). Plants in saline environments. Springer-verlag – Berlin, Heildelberg. New York. Hlm: 7-24.
Daniels BAH dan Trappe JM 1980 Factors affecting spora germination of the VAM fungus, Glomus epigaeus. Mycology. 72 : 457- 463.
Delvian. 2003. Keanekaragaman dan potensi pemanfaatan cendawan mikoriza arbuskula (CMA) di Hutan Pantai [Disertasi]. Program Pasca Sarjana, Institut Pertanian Bogor. Bogor.
Gale J, Kohl HC dan Hagan RM. 1967. Changes in water balance and photosynthesis on onion plants under saline conditions. Physiologia. 20: 408-420.
Giovannetti M dan Mosse B. 1980. An evaluation of technigue for measuring vesicular-arbuscular mycorrhizal infection in roots. New Phytol 84 : 489-500.
Gusmeizal. 1997. Pengujian toleransi bibit beberapa klon karet dengan dan tanpa inokulasi CMVA terhadap tingkat salinitas tanah. Program Pasca Sarjana Universitas Sumatera Utara. Medan.
Hirrel MC dan Gerdermann JW. 1980. Improved growth of onion and bell pepper in saline saoils by soils by two vesicular-arbuscular mycorrhyzal fungi. Soil Sci. Soc. Am. J. 44 : 654-655.
Husin EF, S Syafei M. Kasim dan R Hartawan. 2000. Respon pertumbuhan bibit Acasia mangium di persemaian terhadap mikoriza dan rhizobium. Prosiding pemanfaatan fungi mikoriza sebagai agen bioteknologi ramah lingkungan dalam meningkatkan produktivitas lahan di bidang kehutanan, perkebunan dan pertanian di era millennium baru. 21-23 April 2000. Bogor.
INVAM. 2009. International culture collection of (vesicular) arbuscular mycorrhizal Fungi. http ://invam. caf. wvu. Edu/Myco - info/Taxonomy/classification.htm. [15.04.2009].
Janouskova M; Pavlikova D; Vosatka M. 2006. Potensial contribution of arbuscular mycorrhiza to cadmium immobili sation in soil. Chemosphere 65 (11): 1959 - 1965.
Junifer S dan Abbott LK. 1993. Vesicular-arbuscular mycorrhyzas and soil salinity Mycorrhyza. 4 : 45-57.
Khattak MS, Marziah M, dan Syed MA. 1991. Effect of increasing levels of salinity on selected enzyme activities in rice cell suspension culture. Trans Malaysia Soc. Malaysia.
Kormanik PP dan McGraw AC. 1982. Quantification of VA mycorrhiza in plant root. Dalam N.C. Schenk (Ed). Methods and principles of mycorrhiza research. The American Phytop. Soc. 46 : 37-45
Koske RE dan Tews LL. 1987. Vesicular-arbuscular mycorrhizal fungi of wisconsin sandy soil. Mycologia. 73 : 289-300
Lakitan B. 2000. Dasar-dasar fisiologi tumbuhan. PT. Raja Grafindo Persada. Jakarta.
Maas EV dan RH Nieman. 1978. Physiology of plant tolerance to salinity. Dalam GA Jung (Ed). Crop tolerance to suboptimal land conditions. ASA Spec. Pub. Hlm: 277-299.
Manan S. 1993. Pengaruh mikoriza pada pertumbuhan semai Pinus merkusi di persemaian. Kuliah silvikultur umum. Fakultas Kehutanan IPB. Bogor. Hlm 247-261.
Marx DH. 1982. Mycorrhiza in interaction with other microorganism. In Method dan Principles of mycorrhizal research. The Am. Phyt. Soc Minessota.
Moreira, Dilmar dan Tsai SM. 2007. Biodiversity dan distribution of arbuscular mycorrhizal fungi in Araucaria angustifolia forest. Journal agriculture vol. 64 : 393-399.
Mukhlis. 2007. Analisis tanah tanaman Universitas Sumatera Utara. Press. Medan.
Pacioni G. 1992. Wet sieving and decanting techniques for the extraction of spores of VA mycorrhyzal fungi. Dalam : Norris JR, DJ Read and AK Varma (Eds). Methods in Microbiology. Vol. 24. Academic Press Inc. San Diego Hlm: 317-322.
Poljakoff-Mayber A dan Gale J. 1975. Morphological dan anatomical changes in plant as a response to salinity stress. Dalam : Poljakoff - Mayber A
dan Gale Gale J (Eds). Plants in saline environments. Springer-Verlag Berlin Heidelberg New York. Hlm: 97-117.
Rozema J, W. ARP, Van Esbroek M. 1986. Vesicular arbuscular mycorrhiza in salt marsh plants in response to soil salinity and flooding and the significance to the water relations. Hlm: 657-660.
Safir GR dan JM Duniway. 1982. Evolution of pland Response to colonization by vesicular arbuscular mycorrhizae fungi in NC Schenks (ed). Methoda and Principled of mycorrhizae research.The American Phytopathology Society. St. Paul.
Scannerini S dan Bonfante-Fosolo P. 1983. Comparative ultrastructural analysis of mycorrhyzal associations. Can. J. Bot. 61: 917-922
Schenck NC dan Schroder VN 1974. Temperature response of endogone micorrhiza on soybean roots. Mycologia. 66 : 71.
Setiadi Y. 2001. Peranan mikoriza arbuskula dalam reboisasi lahan kritis di Indonesia. makalah seminar penggunaan CMA dalam sistem pertanian organik dan rehabilitas lahan. Bandung. 21-23 April 2001.
Siguenza C, Espejel l dan Allen EB. 1996. Seasonality of mycorrhizae in coastal sand dunes of Baja California. Mycorrhiza. 6 : 151-157
Siradz SA dan S Kabirun. 2007. Pengembangan lahan marginal pesisir pantai dengan bioteknologi masukan rendah. Jurnal Ilmu Tanah dan Lingkungan. Jurusan Tanah, Fakultas Pertanian UGM. 7 : 83-92.
Smith SE dan D, Read DJ. 1997. Mycorrhizal symbiosis. Second edition. Academic Press. Harcourt Brace dan Company Publisher. London. Hlm: 32-79
Tan KH. 1991. Principles of soil chemistry. Marcel Dekker. Madison Vanue New York Inc.
Thomson BD, Robson AD dan Abbot LK 1990. Mycorrhizas formed by Gigaspora calospora and Glomus fasciculatum on subterranean clover in relation to soluble carbohydrate concentration in root. New Phytol. 114 : 217-225.
Lampiran 1. Skematis ekstraksi dan identifikasi spora fungi mikoriza
Ekstraksi mikoriza
PVLG
PVLG + Melzer
Identifikasi mikoriza
Lampiran 2. Skematis kolonisasi fungi mikoriza pada akar tanaman sampel
Pengamatan kolonisasi mikoriza
Akar di rendam dalam KOH 10%
larutan Trypan blue KOH 10%
Mikroskop Akar Sampel
Rendam dalam HCL 2%
larutan Glycerol
Gambar 1. Tanaman Pueraria javanica dalam trapping fungi mikoriza
Gambar 2. Lokasi pengambilan sampel (tepi pantai Pulau Pandang)
Lokasi penelitian
Gambar 4. Peta lokasi penelitian pantai Pulau Pandang
Lampiran 5. Kriteria persentase kolonisasi akar menurut Setiadi et al. (1992)
No Persentase kolonisasi (%) keterangan
1 0 – 25 rendah