PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis “Keefektifan Kutukebul dalam Menularkan Virus Penyebab Penyakit Kuning pada Tanaman Tomat” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan mau pun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Februari 2010
ABSTRACT
EVA DWI FITRIASARI. Effectiveness of Whiteflies in Transmitting Viruses causing Yellowing Disease in Tomato Plants. Under direction of GEDE SUASTIKA, ALI NURMANSYAH, and DEWI SARTIAMI.
Yellowing disease outbreaks had been occurring in tomato crops in highland of West Java since 2007. The outbreaks were associated with high populations of whiteflies. Symptoms consist mainly of interveinal yellowing that developed initially on lower leaves and then progress to the upper part of the plant. Affected plants are less vigorous and yield less due to reduced fruit growth and delayed ripening. Identifications using Reverse transcription-polymerase chain reaction (RT-PCR) revealed that the yellowing diseases on tomato in West Jawa were associated with infection of two different viruses that were tomato chlorosis virus (ToCV) and tomato infectious chlorosis virus (TICV), both of which belong to the members of genus Crinivirus, family Closteroviridae. Whiteflies collected from tomato fields in Bogor and Cianjur were identified, based on morphologies of their puparium, as Bemisia tabaci and Trialeurodes vaporariorum. T. vaporariorum was found efficiently transmitted both ToCV and TICV, but B. tabaci transmitted ToCV only.
RINGKASAN
EVA DWI FITRIASARI. Keefektifan Kutukebul dalam Menularkan Virus Penyebab Penyakit Kuning pada Tanaman Tomat. Dibimbing oleh GEDE SUASTIKA, ALI NURMANSYAH, dan DEWI SARTIAMI.
Penyakit kuning pada tanaman tomat merupakan salah satu penyakit yang di kategorikan sebagai “new emerging disease” akibat pengaruh pemanasan global. Pada tahun-tahun belakangan ini, penyakit kuning mulai banyak dilaporkan terjadi di berbagai negara penghasil tomat dunia. Gejala penyakit kuning terlihat menguning pada jaringan di antara tulang daun terutama dar i daun-daun tua, mirip dengan gejala yang disebabkan oleh kekurangan unsur hara. Telah dilaporkan bahwa dua spesies virus yang berbeda yaitu ToCV dan TICV dapat terlibat dalam menginduksi penyakit kuning ini. Gejala penyakit yang diinduksi oleh ToCV dilaporkan tidak dapat dibedakan dengan gejala yang diinduksi oleh infeksi TICV. Selain itu, gejala yang muncul juga akan sama bila kedua virus ini bersama-sama menginfeksi tanaman tomat. Menurut beberapa peneliti di belahan lain dunia, terdapat beberapa spesies kutukebul yang mengkolonisasi tanaman tomat. Spesies kutukebul yang ditemukan tersebut adalah T. vaporariorum, T. abutilonea, dan B. tabaci. Data detail mengenai kutukebul pada tanaman tomat dan asosiasinya dengan Crinivirus penyebab penyakit kuning belum tersedia di Indonesia. Oleh karena itu, penelitian ini bertujuan untuk mengidentifikasi virus penyebab penyakit kuning di Indonesia, mengidentifikasi ulang kutukebul yang mengkolonisasi tanaman tomat di lapangan, dan menetapkan keefektifan spesies kutukebul dalam menularkan virus penyebab penyakit kuning.
memanjang, lingulanya memanjang berbentuk lidah, submarginnya tidak mempunyai barisan papila, serta vasiform orifice berbentuk segitiga dan memanjang. Puparium T. vaporariorum berbentuk ovoid, lebih tebal dibandingkan dengan B. tabaci. T. vaporariorum mempunyai barisan papila pada submarginnya, lingulanya membulat, dan tidak mempunyai rambut dorsal.
Pada penularan ini digunakan jumlah kutukebul yang berbeda yaitu 1, 3, 5, 7 dan 10 untuk setiap tanaman tomat yang diinokulasi. Dari hasil uji penularan menjelaskan bahwa B. tabaci hanya dapat menularkan ToCV, sedangkan T. vaporariorum dapat menularkan ToCV maupun TICV. Keefektifan penularan ditentukan oleh jumlah kutukebul walaupun dengan tingkat persentase tanaman terinfeksi yang beragam. Semakin sedikit jumlah kutukebul per tanaman, semakin rendah jumlah tanaman terinfeksi. Semakin banyak jumlah kutukebul yang digunakan maka semakin tinggi tingkat penularan yang terjadi. TICV dengan 10 ekor T. vaporariorum penularan dapat dicapai hingga 100%, namun untuk ToCV perlu lebih dari 10 ekor untuk mencapai 100% penularan. ToCV efektif ditularkan baik oleh B. tabaci maupun T. vaporariorum, sedangkan TICV efektif ditularkan hanya oleh T. vaporariorum tetapi tidak dapat ditularkan oleh B. tabaci. Menurut beberapa penelitian di negara lain bahwa kespesifikan virus dan vektornya sangat ditentukan oleh reseptor yang ada pada stilet serangga dengan CP dari virus bersangkutan. Selubung protein minor pada ToCV, yang membentuk bagian ekor/ujung virion memiliki peranan dalam penularan dengan kutukebul.
Hak Cipta milik IPB, tahun 2010
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
KEEFEKTIFAN KUTUKEBUL DALAM MENULARKAN
VIRUS PENYEBAB PENYAKIT KUNING
PADA TANAMAN TOMAT
EVA DWI FITRIASARI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Keefektifan Kutukebul dalam Menularkan Virus Penyebab Penyakit Kuning pada Tanaman Tomat
Nama : Eva Dwi Fitriasari
NIM : A352070061
Disetujui Komisi Pembimbing
Dr. Ir. Gede Suastika, M.Sc. Ketua
Dr. Ir. Ali Nurmansyah, M.Si. Anggota
Dra. Dewi Sartiami, M.Si. Anggota
Diketahui
Ketua Program Studi Mayor Fitopatologi
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc.
Dekan Sekolah Pascasarjana
Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
PRAKATA
Segala puji bagi Allah SWT semesta alam, atas ramat dan karunia-Nya lah penulis dapat menyelesaikan penelitian dan penulisan tesis dengan judul Keefektifan Kutukebul dalam Menularkan Virus Penyebab Penyakit Kuning pada Tanaman Tomat. Sebagian dari penelitian ini dibiayai oleh Hibah Kompetitif Penelitian Sesuai Prioritas Nasional [Ditjen Dikti Depdiknas] No.343/SP2H/PP/DP2M/VI/2009 dan dari kerjasama dengan Utsunomiya University, Japan melalui Dr. Ir. Gede Suastika, M.Sc.
Penulis ucapkan terima kasih dengan tulus kepada Bapak Dr. Ir. Gede Suastika, M.Sc. (Ketua Komisi Pembimbing), Bapak Dr. Ir. Ali Nurmansyah, M.Si., dan Ibu Dra. Dewi Sartiami, M.Si. (Anggota Komisi Pembimbing) yang telah memberi saran, petunjuk, koreksi, dan bimbingan kepada penulis dalam menyelesaikan penelitian dan penulisan tesis ini. Nasehat dan arahan dari para pembimbing tidak akan pernah dilupakan, serta semangat dan kerja keras pembimbing akan selalu saya contoh. Terima kasih kepada Bapak Dr. Ir. Kikin Hamzah Mutaqin, M.Si. selaku Penguji Luar Komisi yang telah memberi masukan dan saran atas penulisan tesis ini.
Penulis juga ucapkan terima kasih kepada teman-teman yang telah membantu penelitian ini dalam pengambilan sampel, pengerjaan laboratorium: Donnarina Simanjuntak, Ibu Rika Meliansyah, Alis Mukhlis. Kepada teknisi dan laboran Lab. Virologi: Mba Tuti Legiastuti dalam membantu mengerjakan deteksi virus, Pak Mpud dalam mambantu menyediakan peralatan penelitian, informasi, masukan dan juga Pak Edi; laboran Lab. Biosistematika: Ibu Aisyah yang bersedia membantu dalam proses identifikasi kutukebul; teknisi lapangan di Darmaga Pak Sodik yang membantu menyediakan peralatan penelitian; di Cianjur Pak Ateng yang telah membantu proses pengerjaan penelitian di lapangan.
Terima kasih atas dukungan dan semangatnya dari teman-teman di lab: Ka Elsa, Mba Lia, Mba Pipiet, Pak Sayuthi, Faishol (Mahasiswa S1 angkatan ‘43), Mba Cici, Mba Devi, Bu Ifa, Bu Rita, dan Pak Irwan. Tak lupa penulis ucapkan terima kasih sebesar-besarnya dan kepada merekalah penelitian ini kupersembahkan: orangtuaku Ir. Darwanto dan Dewi Tri Puji Astuti; kakakku Elly; dan adik-adikku Intan, Herlina, Indah, dan Sita yang selalu memberi dukungan, motivasi, dan doanya kepada penulis. Keluarga merupakan segalanya yang terbaik buat hidup saya yang selalu memberi dukungan dan dorongan semangat sehingga penyelesaian studi ini berjalan sesuai harapan. Nasehat dan semangat dari orangtua akan selalu diingat agar ke depannya penulis bisa menjadi orang yang lebih mandiri dan dewasa.
Akhirnya penulis berharap semoga karya ilmiah ini bermanfaat bagi pembaca dan perkembangan ilmu pengetahuan.
RIWAYAT HIDUP
Penulis dilahirkan di Karawang pada tanggal 20 Juni 1985 dari ayah Ir. Darwanto dan Ibu Dewi Tri Puji Astuti. Penulis merupakan putri kedua dari enam bersaudara.
DAFTAR ISI
Hubungan Virus dengan Serangga Vektornya ... 11
BAHAN DAN METODE ... 13
Tempat dan Waktu ... 13
Survei Penyakit Kuning pada Pertanaman Tomat ... 13
Identifikasi Virus yang Berasosiasi dengan Penyakit Kuning ... 13
Ekstraksi RNA ... 13
Reaksi RT ... 14
PCR ... 14
Identifikasi Kutukebul yang Mengkolonisasi Pertanaman Tomat ... 15
Penularan Virus Penyebab Penyakit Kuning dengan Kutukebul ... 15
Inokulum Virus ... 15
Kutukebul ... 16
Penularan Virus oleh Kutukebul ... 16
HASIL DAN PEMBAHASAN ... 17
Survei Penyakit Kuning pada Tanaman Tomat di Jawa Barat ... 17
Virus- Virus yang Berasosiasi dengan Gejala Kuning pada Tanaman Tomat ... 19
Kutukebul yang Mengkolonisasi Pertanaman Tomat di Jawa Barat . 20 Penularan Virus Penyebab Penyakit Kuning dengan Kutukebul ... 23
KESIMPULAN DAN SARAN ... 29
Kesimpulan ... 29
Saran ... 29
DAFTAR TABEL
Halaman
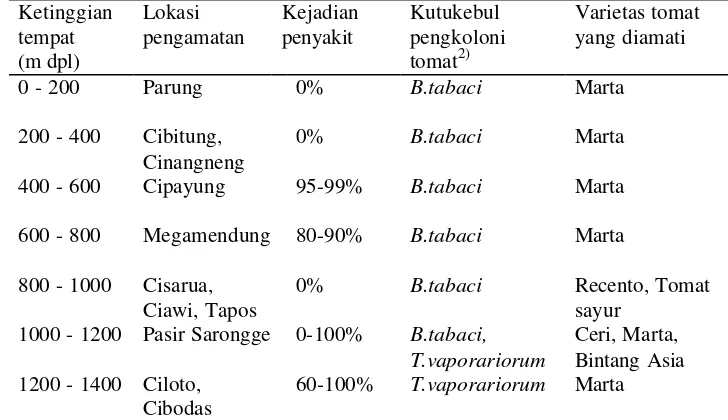
1 Kejadian penyakit kuning pada tanaman tomat menurut ketinggian
tempat di daerah Bogor dan Cianjur ... 18 2 Penularan ToCV dan TICV melalui T. vaporariorum dan B. tabaci dari tanaman sumber virus terinfeksi ganda ... 23 3 Keefektifan B. tabaci menularkan ToCV pada tomat dari tanaman sumber virus terinfeksi tunggal ... 25 4 Keefektifan T. vaporariorum menularkan ToCV dan TICV pada tomat
DAFTAR GAMBAR
Halaman
1 Gejala kuning pada tanaman tomat mulai terjadi dari daun-daun bawah kemudian berkembang ke arah pucuk (atas). Klorosis terutama terjadi
pada jaringan di antara tulang daun (interveinal chlorosis, bawah) ... 19 2 Hasil amplifikasi DNA genom virus menggunakan pasangan primer
spesifik ToCV (lajur 1 dan 2) dan spesifik TICV (lajur 3 dan 4) terhadap sampel daun yang diambil dari tanaman tomat bergejala kuning dari Cipanas (lajur 1), Pacet (lajur 2 dan 3), dan Cikajang (lajur 4). Lajur M adalah 1 kb DNA leader (Qiagen, Germany) ... 20 3 Imago B. tabaci (kiri) dan T. vaporariorum (kanan) ... 21 4 Preparat puparium T. vaporariorum (kiri) dan B. tabaci (kanan): (a) seta kauda, (b) vasiform orifice, (c) rambut dorsal, (d) pinggiran trakea, (e) basal tungkai tengah dan belakang, (f) ruas abdomen VII, (g) submargin, (h) lingula ... 22 5 Gejala kuning pada tanaman tomat yang telah diinokulasi dengan ToCV (kiri) dan TICV (kanan) ... 24 6 Hasil amplifikasi DNA TICV dan ToCV dengan metode RT-PCR
menggunakan sepasang primer TICV dan ToCV. M (marker 1 kb); lajur 1 (negatif TICV), lajur 2 - 9 (positif TICV); lajur 14, 15, 17
PENDAHULUAN
Latar Belakang
Penyakit kuning pada tanaman tomat merupakan salah satu penyakit yang di kategorikan sebagai “new emerging disease” akibat pengaruh pemanasan global (Dovas et al. 2002; Segev et al. 2004). Pada tahun-tahun belakangan ini, penyakit kuning mulai banyak dilaporkan terjadi di berbagai negara penghasil tomat dunia (Louro et al. 2000; Dalmon et al. 2008). Gejala penyakit kuning terlihat menguning pada jaringan di antara tulang daun terutama dari daun-daun tua, mirip dengan gejala yang disebabkan oleh kekurangan unsur hara (Navas-Castillo et al. 2000; Accotto et al. 2001). Pada kunjungan di beberapa lokasi di sentra produksi tomat dataran tinggi di Bogor, Cianjur, dan Garut menemukan gejala penyakit kuning dengan ciri-ciri yang sama dengan yang telah dilaporkan di negara lain (Tsai et al. 2004; Dalmon et al. 2005). Serangan penyakit ini sudah dirasakan pengaruhnya oleh petani setempat mulai tahun 2007 (Hasil wawancara dengan petani di lokasi pengamatan). Pengaruh yang sangat dirasakan petani adalah mengecilnya ukuran buah tomat dan proses pemasakan buah yang tidak normal sehingga tidak bernilai ekonomi. Pengaruh yang sama juga dilaporkan oleh Wisler et al. (1998).
Penyakit kuning dilaporkan diinduksi oleh dua spesies virus yang berbeda yaitu tomato infectious chlorosis virus (TICV) dan tomato chlorosis virus (ToCV) (Dalmon et al. 2005). Gejala penyakit yang diinduksi oleh ToCV dilaporkan tidak dapat dibedakan dengan gejala yang diinduksi oleh infeksi TICV (Dovas et al. 2002). Selain itu, gejala yang muncul juga akan sama bila kedua virus ini bersama-sama menginfeksi tanaman tomat (Wintermantel et al. 2008).
masing-masing berukuran 7.8 dan 8.2 kb. Menurut Martelli et al. (2000), RNA 1 mengkode dua jenis protein yang terlibat dalam replikasi virus; sedangkan RNA 2 mengandung beberapa gen yaitu untuk sebuah protein kecil yang hidrofobik (small hydrophobic protein), sebuah protein berukuran sekitar 60 kDa, dan dua jenis protein mantel yaitu coat protein (CP) dan coat proteinminor (CPm). ToCV dan TICV merupakan anggota genus Crinivirus yang dapat ditularkan oleh kutukebul (whitefly) dan terpisah dari kelompok lain dari anggota famili Closteroviridae yang mempunyai serangga vektor kutudaun (aphids).
Virus-virus pada genus Crinivirus telah diketahui tidak dapat ditularkan melalui cairan perasan tanaman sakit ataupun melalui benih (Martelli et al. 2002). Oleh sebab itu kutukebul menjadi agens utama penyebar penyakit kuning di lapangan. Hal ini bersesuaian dengan penemuan Navas-Castillo et al. (2000) yang menyatakan bahwa tingkat kejadian penyakit kuning di lapangan berkorelasi positif dengan tingkat populasi kutukebul.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengidentifikasi virus penyebab penyakit kuning di Indonesia, mengidentifikasi ulang kutukebul yang mengkolonisasi tanaman tomat di lapangan, dan menganalisis keefektifan spesies kutukebul dalam menularkan virus penyebab penyakit kuning.
Manfaat Penelitian
TINJAUAN PUSTAKA
Tomato infectious chlorosis virus (TICV)
TICV pertama kali ditemukan di lahan tomat California tahun 1993 (Duffus et al. 1994) dan setelah itu ditemukan pula di beberapa lahan tomat di Italia (Vaira et al. 2000; Parrella & Scassillo 2006), Spanyol (Font et al. 2002), Yunani (Dovas et al. 2002), dan Perancis (Dalmon et al. 2005). Di Asia, TICV telah terdeteksi pada tanaman tomat di Indonesia dan Jepang (Verhoeven et al. 2003; Hartono et al. 2003). Virus ini diketahui sebagai virus yang ditransmisikan oleh kutukebul pada tanaman tomat yang terinfeksi (Klaassen et al. 1995). Virus ini ditransmisikan khusus oleh kutukebul T. vaporariorum dan memiliki periode persisten selama 4 hari (Duffus et al. 1994; Wisler et al. 1996). Penyebaran virus ini sangat bergantung pada bantuan kutukebul T. vaporariorum untuk menularkan virus pada jaringan floem.
TICV berbentuk panjang lentur, partikel berfilamen dengan ukuran sekitar 650 nm. Closterovirus ini menginduksi penyakit kuning pada bagian jaringan floem (Duffus et al. 1996). TICV merupakan kelompok genom bipartit RNA untai tunggal (ssRNA), dengan panjang genom RNA 1 dan RNA 2 berturut-turut yaitu 7.8 dan 7.4 kb (Liu et al. 2000).
Penyakit kuning pada tanaman tomat telah menyebar di sentra pertanaman tomat di Magelang, Jawa Tengah. Penyakit dengan gejala yang sama juga telah dilaporkan di Purwakarta, Jawa Barat. Hasil pengamatan lapangan di Magelang mencatat rata-rata intensitas penyakit kuning-ungu antara 30% hingga 80%
Gejala penyakit kuning pada tanaman tomat meliputi menguningnya daun, nekrosis, daun menggulung ke bawah, daun mengering dan rapuh diikuti dengan kehilangan hasil yang parah. Kehilangan hasil terjadi karena area fotosintesis pada daun yang berkurang. Gejala kuning tampak pada daun bagian bawah, pada daun bagian tengah, dan bahkan pada daun bagian atas (Anfoka & Abhary 2007).
Gejala penyakit kuning yang disebabkan TICV tidak mudah dikenali karena virus-virus tersebut berada pada daun tua dan berkelompok dengan gejala yang disebabkan oleh kekurangan nutrisi (Dovas et al. 2002). Gejala virus ini secara alami terbatas pada jaringan floem, dan dengan konsentrasi virus rendah menyebabkan diagnosis sulit dilihat pada tanaman inang yang terinfeksi. Walaupun tidak tampak jelas gejala pada bunga dan buah tomat, tetapi produksi buah berkurang dengan mengecilnya ukuran buah dan menurunnya jumlah buah akibat berkurangnya daerah fotosintesis (Li et al. 1998).
Tomato chlorosis virus (ToCV)
ToCVpertama kali tersebar di negara bagian Florida sejak 1989 dan diduga virus ini sudah tersebar di Indonesia saat ini. Virus ini dengan cepat menyebar ke seluruh dunia dan sampai saat ini keberadaannya telah dilaporkan di banyak negara seperti di Taiwan (Tsai et al. 2004), Spanyol (Navas-Castillo et al. 2000; Lozano et al. 2006), Yunani (Kataya et al. 2008), dan Perancis (Masse et al. 2008). Virus ini ditransmisikan oleh kutukebul T. vaporariorum,B. tabaci biotipe A dan B, serta T. abutilonea dengan periode retensi 1-2 hari. ToCV dit ularkan oleh ketiga spesies serangga vektor di atas dengan sangat efisien secara semi-persisten, sehingga kejadian penyakit kuning pada tanaman tomat sangat dipengaruhi oleh kepadatan populasi dan aktivitas serangga ini (Jacquemond et al. 2008). Efisiensi penularan bervariasi oleh vektor B. tabaci biotipe B (juga dikenal sebagai B. argentifolii Bellows & Perring) dan T. abutilonea yang lebih efisien menularkan ToCV daripada B. tabaci biotipe A atau T. vaporariorum (Wisler & Duffus 2001).
yang terlibat dalam replikasi virus; sedangkan RNA 2 mengandung beberapa gen yaitu sebuah protein kecil yang hidrofobik, sebuah protein berukuran sekitar 60 kDa, dan dua jenis protein mantel yaitu CP dan CPm. Selubung protein minor (CPm) pada ToCV, yang membentuk bagian ekor/ujung virion memiliki peranan dalam penularan dengan kutukebul. CPm dari ToCV memiliki kespesifikan dengan reseptor T. vaporariorum dan B. tabaci. Menurut Wintermantel (2006), kespesifikan virus dan vektornya sangat ditentukan oleh reseptor yang ada pada stilet serangga dengan CP dari virus bersangkutan.
Infeksi ToCV pada tanaman tomat menyebabkan klorosis yang pada awalnya terjadi pada daun-daun bagian bawah kemudian berkembang ke bagian pucuk, pada lamina daun di antara tulang daun mengalami klorosis (interveinal yellowing). Setelah munculnya vektor kutukebul, gejala ini berkembang dalam beberapa minggu. Daun menjadi tebal dan keriting, dan mudah rapuh jika dipatahkan. Virus ini dapat menyebar dengan cepat ke pertanaman di sekitar sumber virus sesuai dengan aktivitas kutukebul sebagai vektornya sehingga kejadian penyakit dalam satu kebun petani sering ditemukan mencapai lebih dari 90% (Navas-Castillo et al. 2000). Kehilangan hasil akibat infeksi virus ini di lapangan menyebabkan ukuran buah mengecil, jumlah buah berkurang saat panen, dan menurunnya umur tanaman (Wintermantel 2004).
Infeksi Ganda (TICV dan ToCV)
Persebaran TICV dan ToCV bergantung pada kutukebul. Kedua virus tersebut ditularkan kutukebul terpisah dari kelompok lain dari anggota famili Closteroviridae yang mempunyai serangga vektor kutudaun. Adanya infeksi ganda yang ditularkan oleh T. vaporariorum dan B. tabaci menandakan bahwa infeksi oleh satu virus tidak mencegah infeksi oleh virus yang kedua. Epidemik Crinivirus tidak hanya dipengaruhi oleh perpindahan serangga vektor tetapi juga oleh faktor persaingan pada tanaman inang. Kemungkinan bahwa persaingan masing-masing virus yang menyebabkan kemunculan penyakit kuning dipengaruhi oleh jenis/spesies tanaman inang dan umur tanaman inang (Dalmon et al. 2008).
termasuk gulma Picris achioides L., tembakau Nicotiana glauca G., Cynara cardunculus L. dan beberapa tanaman hias yaitu tanaman cina aster dan petunia. Gejala penyakit yang disebabkan oleh TICV lebih parah pada tanaman indikator Nicotiana branthamina dan N. clevelandi berupa bercak klorosis pada tulang daun dibandingkan dengan yang diinduksi oleh ToCV. Gejala penyakit kuning yang disebabkan ToCV pada tanaman inang yaitu klorosis pada interval tulang daun tetapi tidak nekrosis, sedangkan TICV terlihat klorosis dan nekrosis (Duffus et al. 1994). Menurut Wisler et al. (1998) gejala penyakit kuning yang terinfeksi kedua virus tersebut terlihat daun menguning pada interval tulang daun, bercak nekrosis, daun menggulung ke bawah kemudian berkembang ke bagian atas. Penelitian tentang kisaran inang pada tanaman selada mengindikasikan bahwa TICV menginfeksi selada sedangkan ToCV tidak mampu menginfeksi tanaman selada (Parrella 2007). Tanaman kentang di rumah kaca rentan terhadap infeksi TICV dan ToCV yang mirip dengan gejala yang diinduksi oleh potato leafroll luteovirus (Famili Lutoeviridae, Genus Luteovirus) walaupun lahan tanaman kentang yang terinfeksi oleh kedua virus tersebut belum dilaporkan (Wisler et al. 1998).
Kutukebul
Kutukebul merupakan kelompok serangga yang berukuran kecil berwarna putih dan bertubuh lunak. Kutukebul termasuk ke dalam Famili Aleyrodidae, superfamili Aleyrodoidea, subordo Sternorrhyncha, ordo Hemiptera (Carver et al. 1991). Famili Aleyrodidae memiliki dua sub famili, yaitu Alerodicinae dan Aleyrodinae (Martin 1987). Saat ini jumlah spesies serangga famili Aleyrodidae yang pernah dideskripsikan sebanyak 1200 spesies.
telah menempel pada daun-daun itu dapat dijadikan media hidup cendawan lain yaitu Capnodium sp. atau dikenal sebagai embun jelaga (Hoddle 2004).
Siklus hidup kutukebul terdiri dari empat fase perkembangan yaitu telur, nimfa, pupa, dan imago. Kutukebul bereproduksi secara seksual atau partenogenesis. Serangga betina yang sudah dibuahi oleh serangga jantan meletakkan telurnya di permukaan daun. Ketika telur menetas, nimfa instar pertama kutukebul akan bergerak untuk mencari tempat menghisap makanan yang sesuai dan menetap disana. Pada stadia nimfa akhir, kutukebul menghentikan aktivitas makannya dan membentuk semacam kulit pupa sebagai tempat perlindungan proses menuju imago. Stadia ini disebut puparium, setelah melewati fase pupa kutukebul menjadi imago (Kalshoven 1981).
Beberapa tahun belakangan, kutukebul telah menjadi masalah utama bagi para petani di seluruh dunia. Walaupun kutukebul dianggap sebagai grup serangga tropis, spesies berbahaya ini banyak sekali ditemukan di seluruh belahan lain dunia, terutama di daerah beriklim subtropis. Sepanjang abad ke-20, B. tabaci dan T. vaporariorum telah menjadi vektor patogen-patogen tertentu baik di rumah kaca maupun di lahan terbuka di wilayah beriklim hangat (Martin et al. 2000). Stadia nimfa dan imago kutukebul merupakan stadia yang menyebabkan kerusakan tanaman (Morales 2001).
Sebagai vektor, kutukebul dilaporkan dapat menularkan sekitar tujuh kelompok virus yaitu closterovirus, geminivirus, carlavirus, potyvirus, nepovirus, luteovirus, dan virus DNA yang berbentuk batang (Markham et al. 1994). Di antara kelompok virus tersebut yang paling banyak ditularkan adalah closterovirus (Famili Closteroviridae, Genus Crinivirus) dan geminivirus (Famili Geminiviridae, Genus Begomovirus) (Muniyappa & Reddy 1983, Wisler et al. 1998).
Trialeurodes vaporariorum. T. vaporariorum umumnya dikenal sebagai
lilin hampir sejajar dengan permukaan daun. Imago ini sayapnya seperti tenda mengikuti seluruh tubuhnya, sayap menutupi tubuh dan sayap itu berbentuk segitiga (Smith 2009).
Selama instar empat atau instar akhir (pupa) susunan mata dan jaringan tubuh menjadi terlihat, larva menebal dan muncul dari permukaan daun. Instar akhir ini sering disebut pupa karena imago kutukebul muncul dari fase ini. Lama stadium pupa berkisar antara 3-7 hari. Ciri khas pupa T. vaporariorum terutama pada bagian-bagian seperti: lingula, vasiform orifice, submargin papila. T. vaporariorum mempunyai barisan papila pada submarginnya, basal tungkai tengah dan belakang mempunyai seta yang kecil dan halus, dan lingulanya membulat (Martin 1987).
Imago T. vaporariorum umumnya menetap pada daun-daun muda yang dekat pada titik tumbuh tanaman, dan meletakkan telur di tempat tersebut. Imago bertahan hidup pada suhu 22 sampai 25 oC, sedangkan pada suhu di atas 30 oC imago tidak mampu berkembang dan suhu di atas 35 oC imago akan mati (Smith 2009). Xie et al. (2006) menjelaskan bahwa T. vaporariorum memiliki kemampuan beradaptasi pada suhu dingin pada semua fase perkembangan dibandingkan dengan B. tabaci.
Semua stadia hidupnya selain telur dan pupa menyebabkan kerusakan tanaman dengan memakan langsung, memasukkan stiletnya ke dalam tulang daun dan mengekstrak makanan dari sap floem (Wintermantel 2004). Cara makan nimfa yaitu dengan menghisap sap dari tanaman, protein ekstrak, nutrisi dan mengeluarkan gula, dimana keberadaan sap tersebut disebut dengan embun madu. Sebagai produk makanan, embun madu yang dikeluarkan dapat menjadi sumber utama kerusakan. Pada populasi yang tinggi, sejumlah embun madu yang diproduksi menyebabkan tanaman atau buah-buahan menjadi lengket, dan pertumbuhan embun jelaga menyebabkan terbatasnya sistem fotosintesis menjadi terhalang. Karakteristik yang lebih berpotensi berbahaya yaitu kemampuan imago menularkan beberapa virus tanaman. T. vaporariorum mampu menularkan beberapa penyakit virus tanaman (Cardona 2002).
Bemisia tabaci. B. tabaci dikenal dengan nama umum kutukebul kapas,
kutukebul ini yaitu: tubuh imago berwarna kuning, sayap tertutup oleh tepung berwarna putih, panjang tubuh 1-1,5 mm. Serangga ini memiliki siklus hidup sekitar 2 sampai 3 minggu (Kalshoven 1981).
Nimfa dan pupa berwarna keputih-putihan dan bentuknya bervariasi tergantung pada substratnya, memiliki panjang 0,7 mm (Kalshoven 1981). Nimfa instar pertama tidak mempunyai peran yang penting dalam penyebaran virus tanaman (Costa 1969). Pada pupa terdapat dua bintik merah yang merupakan bakal mata pada fase imago. Kedua bintik merah itu terlihat melalui integumennya yang transparan (Hill 1987). Bentuk pupa bulat memanjang, bagian toraks agak melebar, dan cembung. Ruas-ruas abdomen tampak jelas. Pinggir kantung pupa tidak rata dan pada bagian dorsal terdapat tujuh pasang duri dan satu pasang pada ujung anal. Vasiform orifice berbentuk segitiga dan memanjang, operculum menutupi hampir seluruh bagian dari vasiform orifice (Martin 1987).
Lama hidup imago betina sekitar 6 hari (Kalshoven 1981), tetapi juga bisa sampai 60 hari, sedangkan lama hidup imago jantan umumnya lebih pendek, yaitu antara 9-17 hari. Sayapnya terdiri dari dua pasang dan transparan seperti tenda dengan posisi saat istirahat terlihat menyempit ke depan (CABI 1999).
B. tabaci ini beradaptasi pada suhu hangat yakni berkisar 14 sampai 35 oC, dengan suhu optimum sekitar 25 sampai 30 oC. Pada tanaman tomat, imago mampu hidup selama 10-15 hari pada suhu 28 sampai 30 oC, sedangkan imago betina mampu memproduksi sekitar 195 telur pada suhu 25 oC (Smith 2009). Hal ini menunjukkan bahwa aktivitas terbesar B. tabaci menyerang tanaman terjadi pada musim panas dan untuk daerah tropis populasi serangga ini banyak dijumpai pada musim kemarau.
Hubungan Virus dengan Serangga Vektornya
Sebagian besar virus tanaman ditularkan oleh serangga vektor dan tergantung pada perilaku dan kapasitas penyebaran vektor tersebut untuk menyebarkan virus dari tanaman ke tanaman. Serangga vektor virus tanaman memiliki beberapa ordo (Hemiptera, Coleoptera, Thysanoptera, Orthoptera, Dermaptera, Lepidoptera, Diptera), akan tetapi Hemiptera merupakan kelompok vektor yang paling penting pada virus tanaman. Ordo-ordo tersebut dibagi ke dalam tiga subordo: Heteroptera (kepik), Auchenorrhyncha (tonggeret dan wereng) dan Sternorrhyncha (kutudaun, kutukebul, kutuputih, kutu loncat). Sebagian besar anggota subordo Auchenorrhyncha dan Sternorrhyncha termasuk vektor virus tanaman (Carver et al. 1991; Borror et al. 1996). Dari 1200 spesies kutukebul yang telah diteliti, hanya empat spesies (B. tabaci, T. vaporariorum, T. abutilonea, dan T. ricini Misra) ditemukan menularkan virus tanaman (Jones 2003).
Interaksi antara protein selubung virus dengan kutukebul terjadi saat penempelan partikel virus dengan reseptor sehingga virus dapat tertular. Virus akan berada dalam tubuh serangga vektor saat diakuisisi. Virus menuju sel epitel saluran pencernaan dan berasosiasi dengan kelenjar saliva serangga. Virus bersirkulasi dalam tubuh serangga sampai akhirnya virus mencapai ke stilet dan masuk ke dalam tanaman sehat saat vektor makan cairan floem. Virus tersebut memerlukan waktu akuisisi dan inokulasi satu jam hingga satu hari dan periode laten satu hari hingga beberapa minggu dalam tubuh serangga (Gray & Banerjee 1999).
BAHAN DAN METODE
Tempat dan Waktu
Kegiatan survei dilakukan di sentra produksi tomat di daerah Bogor, Cianjur, dan Garut; identifikasi kutukebul dilakukan di Laboratorium Biosistematika Serangga; identifikasi virus dilakukan di Laboratorium Virologi Tumbuhan; dan penetapan hubungan virus-kutukebul dilakukan di Rumah Kaca, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian dilakukan dari Agustus 2008 sampai November 2009.
Survei Penyakit Kuning pada Pertanaman Tomat
Kegiatan survei dilakukan di daerah Bogor dan Cianjur yang mempunyai ketinggian tempat dari beberapa meter di atas permukaan laut (m dpl) sampai lebih dari 1300 m dpl. Pada setiap rentang ketinggian tempat diamati empat sampai enam kebun tomat petani. Kemudian setiap kebun diamati gejala penyakit kuning dan kejadian penyakit dihitung dengan membagi jumlah tanaman contoh yang memperlihatkan gejala dengan jumlah tanaman yang diamati. Selanjutnya diamati kutukebul yang mengkolonisasi pertanaman tomat setempat.
Identifikasi Virus yang Berasosiasi dengan Penyakit Kuning
Untuk mengetahui virus yang berasosiasi dengan penyakit kuning pada tanaman tomat di daerah Jawa Barat, maka dilakukan deteksi virus melalui RT-PCR.
Ekstraksi RNA. RNA total diekstraksi dari jaringan daun tanaman tomat
ml baru. Kemudian ditambahkan 0,5 vol etanol 96% ( ± 225µl) dicampur dengan menaik turunkan pipet. Dimasukkan sampel (± 650 µl) termasuk endapan yang terbentuk ke dalam Rneasy mini column pink, ditempatkan pada tabung koleksi 2 ml. Tutup dengan baik, lalu disentrifuse 10000 rpm 15 detik. Cairan dibuang pada tabung koleksi. Ditambahkan 700µl bufer RW1 ke dalam Rneasy column. Disentrifuse 10000 rpm 15 detik untuk mencuci column. Rneasy column dipindahkan ke tabung koleksi 2 ml baru, lalu dipipet 500 µl bufer RPE ke dalam Rneasy column. Disentrifuse 10000 rpm 15 detik. Cairan dibuang pada tabung koleksi. Gunakan kembali tabung koleksi, lalu ditambahkan 500 µl bufer RPE. Sentrifuse 10000 rpm 2 menit. Untuk meyakinkan bahwa column telah kering, coloumn dipindahkan pada tabung koleksi baru, disentrifuse 10000 rpm 1 menit. 40 µl Rnase free water ditambahkan ke dalam Rneasy column, didiamkan 10 menit. disentrifuse 10000 rpm 1 menit. Siapan RNA total digunakan sebagai template dalam reaksi RT-PCR.
Reaksi RT. Reaksi RT dilakukan dengan volume total 25 µl yang
mengandung 3 µl RNA total, 0,75 pmol random primer, 500 mM dNTP, 5 mM MgCl2, 4 µl bufer RT (250 mM Tris- HCl, pH 8,3; 375 mM KCl; 15 mM MgCl2; 50 mM DTT), 20 unit RNasin ribonuclease inhibitor (Promega, Madison, WI), dan 65 unit MMLV RT (Promega, Madison, WI). Reaksi RT dilakukan pada suhu 42 oC selama 60 menit dan dilanjutkan inaktivasi pada 95 oC selama 5 menit.
ditambahkan 25 ml buffer TAE 0,5x (Tris Asetat EDTA). Kemudian dipanaskan dalam microwave sampai larutan berwarna bening. Larutan agar didinginkan hingga suhu 60 oC selama 10 menit lalu ditambahkan 1,25 µl etidium bromida; kemudian diaduk dan dituang dalam cetakan gel agarose. Gel didiamkan selama 30 menit sampai gel mengeras. Gel ditempatkan di dalam bak elektroforesis posisi sumur pada kutub negatif. Sampel cDNA 10 µ l dicampur dengan 2 µ l loading dye, marker 1 kb sebanyak 5 µ l dicampur dengan 1 µ l laoding dye. Sampel dan marker tersebut dimasukkan ke dalam sumur gel elektroforesis lalu dimigrasikan pada tegangan 50 volt selama 45 menit. Visualisasi DNA hasil PCR dilakukan dengan transluminator UV kemudian dipotret dengan kamera digital.
Identifikasi Kutukebul yang Mengkolonisasi Pertanaman Tomat
Identifikasi spesies kutukebul dilakukan dengan cara membuat preparat kantung puparium dengan metode tanpa pemanasan berdasarkan metode Martin (1999). Puparium kutukebul didehidrasi dengan merendam dalam alkohol 95% selama 10 menit dan kemudian dalam asam asetat glasial selama 10 menit. Setelah dicuci dengan akuades, kantung puparium direndam dalam carbol xylene selama satu menit dan kembali dicuci dengan akuades dan siap diwarnai. Pewarnaan dilakukan dengan perendaman dalam campuran asam asetat glasial dan asam fuksin (1:1) selama satu malam. Untuk mengurangi kepekatan pewarnaan, kantung puparium dimasukkan berturut-turut ke dalam alkohol 80% dan 100% masing-masing selama 10 menit. Selanjutnya, preparat diletakkan di cawan sirakus dan direndam selama 10 menit dalam minyak cengkeh. Kantung puparium dipindahkan dan diletakkan di atas kaca objek. Selanjutnya, balsam kanada diteteskan sebanyak satu tetes dan ditutup dengan kaca penutup. Identifikasi spesies kutukebul dilakukan mengikuti Martin (1987).
Penularan Virus Penyebab Penyakit Kuning dengan Kutukebul
Inokulum virus. Virus TICV dan ToCV yang digunakan dalam penelitian
RT-PCR bahwa terinfeksi tunggal oleh ToCV, atau oleh TICV, dan terinfeksi ganda ToCV dan TICV.
Kutukebul. Kutukebul disiapkan dengan mengumpulkan puparium dari
pertanaman tomat di daerah Bogor dan Cianjur. Puparium dikumpulkan secara terpisah dari koloni kutukebul yang berbeda (dilihat dari ukuran dan cara hinggap kutukebul). Puparium dari koloni kutukebul yang berbeda kemudian masing-masing diletakkan pada tanaman tomat yang dipelihara dalam kurungan yang berbeda. Imago yang keluar beberapa hari berikutnya dipelihara dan dibiarkan meletakkan telur pada tanaman tomat yang sama. Tanaman tomat yang telah mengandung sejumlah telur kutukebul dipindahkan ke kurungan baru yang telah berisi tanaman tomat sehat lain. Pemeliharaan dilakukan sampai diperoleh imago baru yang merupakan imago bebas virus dan kemudian digunakan dalam studi penularan virus.
Penularan virus oleh kutukebul. Penularan dilakukan dengan membiarkan
HASIL DAN PEMBAHASAN
Survei Penyakit Kuning pada Tanaman Tomat di Jawa Barat
Survei yang telah dilakukan di daerah Jawa Barat yaitu di Bogor, Cianjur, dan Garut menemukan bahwa penyakit kuning pada tanaman tomat telah banyak terjadi. Gejala penyakit kuning lebih banyak terlihat pada tanaman tomat yang ditanam di daerah dengan ketinggian sedang mulai dari 400 m dpl sampai ketinggian sekitar 1400 m dpl (Tabel 1). Gejala penyakit kuning pada tanaman tomat terjadi mulai dari daun-daun terbawah kemudian secara perlahan berkembang ke arah daun-daun bagian atas dan pada akhirnya seluruh daun memperlihatkan gejala penyakit kuning. Penyakit kuning ditandai dengan perubahan warna menjadi kuning pada lamina di antara tulang-tulang daun (Gambar 1). Kadang-kadang bagian yang klorotik berkembang menjadi nekrotik karena kematian jaringan tanaman. Di samping itu, beberapa daun terutama daun-daun bagian atas maupun pucuk berubah warna menjadi ungu. Tekstur daun-daun juga berubah menjadi kurang elastis, daun menjadi tebal dan keriting, serta rapuh bila diremas. Gejala ini berkembang dalam beberapa minggu setelah munculnya vektor kutukebul. Walaupun tidak tampak jelas gejala pada bunga dan buah tomat, tetapi produksi buah berkurang dengan mengecilnya ukuran buah dan menurunnya jumlah buah akibat berkurangnya daerah fotosintesis. Gejala semacam ini sama dengan yang sudah dideskripsikan oleh beberapa peneliti terdahulu (Accotto et al. 2001).
Kutukebul merupakan agens utama penyebar penyakit kuning di lapangan. Hal ini terlihat dari hasil pengamatan di lapangan menemukan dua spesies kutukebul yaitu B. tabaci dan T. vaporariorum pada tanaman tomat yang terserang penyakit kuning. Hal ini bersesuaian dengan penemuan Navas-Castillo et al. (2000) yang menyatakan bahwa tingkat kejadian penyakit kuning di lapangan berkorelasi positif dengan tingkat populasi kutukebul. Oleh karena itu, penyakit kuning ini diketahui tidak dapat ditularkan melalui cairan perasan tanaman sakit ataupun melalui benih (Martelli et al. 2002).
Kutukebul pengkoloni tanaman tomat dengan ketinggian dimulai dari 0 m dpl sampai 1000 m dpl didominasi oleh B. tabaci, ketinggian 1000 sampai 1200 m dpl oleh kedua kutukebul tersebut yaitu B. tabaci dan T. vaporariorum, sedangkan ketinggian diatas 1200 m dpl hanya didominasi oleh T. vaporariorum. Xie et al. (2006) menjelaskan bahwa T. vaporariorum memiliki kemampuan beradaptasi pada suhu dingin pada semua fase perkembangan dibandingkan dengan B. tabaci sehingga hal tersebut diduga mempengaruhi pola distribusi kedua spesies tersebut. Tabel 1 Kejadian penyakit kuning pada tanaman tomat menurut ketinggian tempat
di daerah Bogor dan Cianjur1) Ketinggian
0% B.tabaci Recento, Tomat
sayur
1) Pengamatan dari 4 sampai 6 kebun petani tomat pada setiap kreteria ketinggian tempat; 2)
Gambar 1 Gejala kuning pada tanaman tomat mulai terjadi dari daun-daun bawah kemudian berkembang ke arah pucuk (atas). Klorosis terutama terjadi pada jaringan di antara tulang daun (interveinal chlorosis, bawah)
Virus-Virus yang Berasosiasi dengan Gejala Kuning pada Tanaman Tomat
Pada saat ini, deteksi virus pada tanaman lebih banyak dilakukan melalui uji serologi karena lebih murah dan juga akurat. Namun demikian, untuk virus-virus yang belum tersedia antiserumnya termasuk antiserum ToCV dan TICV, maka deteksi dilakukan melalui pendekatan molekuler terutama RT-PCR. Deteksi dengan RT-PCR perlu didesain sepasang primer yang spesifik untuk virus target. Berdasarkan analisa sekuen ToCV dan TICV yang diunduh dari GenBank, maka
telah didesain sepasang primer ToCV- CP-F (5’
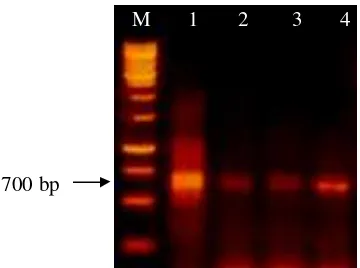
Gambar 2 Hasil amplifikasi DNA genom virus menggunakan pasangan primer spesifik ToCV (lajur 1 dan 2) dan spesifik TICV (lajur 3 dan 4) terhadap sampel daun yang diambil dari tanaman tomat bergejala kuning dari Cipanas (lajur 1), Pacet (lajur 2 dan 3), dan Cikajang (lajur 4). Lajur M adalah 1 kb DNA leader (Qiagen, Germany)
Seperti yang ditampilkan pada Gambar 2, RT-PCR yang dilakukan terhadap jaringan daun yang diambil dari tanaman tomat bergejala kuning di lapangan berhasil dilakukan dengan primer spesifik ToCV maupun TICV dan terbentuk pita yang jelas dengan ukuran sekitar 700 bp. Hasil deteksi ini mengindikasikan dengan jelas bahwa penyakit kuning yang terjadi pada tanaman tomat di daerah Jawa Barat berasosiasi dengan infeksi ToCV dan/atau TICV. Hasil penelitian ini juga mengkonfirmasi bahwa kedua virus ini sudah ada di daerah Jawa Barat. Hartono & Wijonarko (2007) juga pernah melaporkan bahwa TICV sudah ditemukan di daerah Yogyakarta.
Kutukebul yang Mengkolonisasi Pertanaman Tomat di Jawa Barat
Survei yang telah dilakukan pada pertanaman tomat di daerah Jawa Barat ditemukan dua jenis koloni kutukebul dari imago yang tampak berbeda dari ukuran maupun cara hinggapnya. Satu kelompok kutukebul mempunyai imago yang berukuran relatif lebih kecil (sekitar 1 mm) dan hinggap dengan sayap ke dalam sedangkan kelompok lainnya berukuran sekitar 1,5 mm dan hinggap dengan sayap agak ke luar (Gambar 3). Identifikasi lebih lanjut berdasarkan morfologi pada masing-masing pupariumnya menggunakan kunci identifikasi (Martin 1987) menemukan bahwa spesies kutukebul kelompok pertama adalah B. tabaci, sedangkan kelompok kedua adalah T. vaporariorum.
M 1 2 3 4
Imago B. tabaci berwarna kuning dengan disertai sayap yang ditutupi oleh sekresi berupa tepung berwarna putih, dengan panjang tubuh 1-1,5 mm. Sayapnya terdiri dari dua pasang dan transparan seperti tenda dengan posisi saat istirahat terlihat menyempit ke depan.
Imago T. vaporariorum secara umum menetap pada daun-daun muda dekat pada titik tumbuh tanaman, dan meletakkan telur di tempat tersebut. Imago kutukebul panjangnya 1-2 mm dengan warna tubuh kekuningan dan memiliki empat sayap diselimuti lilin hampir sejajar dengan permukaan daun. Imago dewasa ini sayapnya seperti tenda mengikuti seluruh tubuhnya, menyembunyikan tubuhnya dan memperlihatkan sayap yang berbentuk segitiga.
Gambar 3 Imago B. tabaci (kiri) dan T. vaporariorum (kanan)
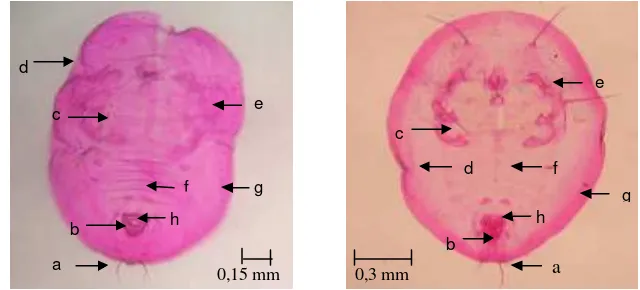
Puparium T. vaporariorum berbentuk ovoid, lebih tebal dibandingkan dengan B. tabaci.Ciri khas T. vaporariorum terutama pada bagian-bagian seperti: lingula, vasiform orifice, submargin papila. T. vaporariorum mempunyai barisan papila pada submarginnya, basal tungkai tengah dan belakang mempunyai seta yang kecil dan halus, lingulanya membulat, tidak mempunyai rambut dorsal, serta pinggiran trakeanya tidak seperti sisir (Gambar 4). Permukaan dorsal tanpa pola duri yang kokoh. Kantung pupa pucat, kadang-kadang dengan tanda kehitaman. Hasil penelitian ini memastikan bahwa kutukebul yang mengkoloni tanaman tomat di Jawa Barat adalah B. tabaci dan T. vaporariorum. Hasil yang sama didapatkan oleh Rahayuwati (2009) yang telah mengidentifikasi kutukebul pada beberapa tanaman tomat di daerah lain di Indonesia.
Gambar 4 Preparat puparium T. vaporariorum (kiri) dan B. tabaci (kanan): (a) seta kauda, (b) vasiform orifice, (c) rambut dorsal, (d) pinggiran trakea, (e) basal tungkai tengah dan belakang, (f) ruas abdomen VII, (g) submargin, (h) lingula
Hasil identifikasi kantung pupa tersebut kemudian dilanjutkan dengan pengumpulan dan perbanyakan serangga vektor dari pupa T. vaporariorum dan B. tabaci yang diambil di bawah permukaan daun tomat. Pupa diletakkan selama 2-3 hari pada kurungan, imago segera muncul, dan imago tersebut yang digunakan sebagai serangga vektor TICV dan/atau ToCV. Serangga vektor yang digunakan dalam penularan virus ini yaitu imago yang telah bebas virus karena virus tidak ditularkan pada generasi berikutnya.
Penularan Virus Penyebab Penyakit Kuning dengan Kutukebul
Hasil penelitian ini telah memastikan bahwa ToCV dan TICV berasosiasi dengan kejadian penyakit kuning pada tanaman tomat. Demikian juga B. tabaci dan T. vaporariorum merupakan dua spesies kutukebul yang ditemukan mengkoloni tanaman tomat di lapangan. Penelitian selanjutnya dilakukan untuk mengetahui asosiasi antara virus dan kutukebul. Pada pengujian pertama, setiap spesies kutukebul diuji kemampuannya untuk menularkan virus dari tanaman tomat terinfeksi ganda, sumber ToCV dan TICV ke bibit tomat sehat.
Hasil penelitian seperti yang ditampilkan pada Tabel 2 memperlihatkan bahwa bila B. tabaci digunakan sebagai serangga penular maka hanya ToCV yang terdeteksi pada tanaman tomat uji. Akan tetapi, bila T. vaporariorum digunakan dalam penularan maka terdeteksi ToCV atau TICV pada tanaman tomat uji baik secara terpisah maupun secara bersama-sama. Dari hasil penularan ini, dapat dis impulkan bahwa B. tabaci hanya dapat menularkan ToCV, sedangkan T. vaporariorum dapat menularkan ToCV maupun TICV. Hasil yang sama juga telah didapatkan oleh Wisler & Duffus (2001) yang menyatakan bahwa ToCV dapat ditularkan oleh B. tabaci biotipe A, biotipe B (juga dikenal sebagai B. argentifolii), T. abutilonea dan T. vaporariorum, sedangkan TICV tidak dapat ditularkan oleh B. tabaci.
Tabel 2 Penularan ToCV dan TICV melalui T. vaporariorum dan B. tabaci dari tanaman sumber virus terinfeksi ganda1)
Spesies
Tanaman sumber virus adalah tanaman tomat yang terinfeksi oleh ToCV dan TICV dan telah diverifikasi melalui RT-PCR; 2) Jumlah imago kutukebul yang digunakan adalah 10 ekor per tanaman uji; 3) Infeksi virus diverifikasi melalui RT-PCR dua minggu setelah penularan dan tanaman yang terinfeksi menunjukkan gejala kuning (Gambar 5)
dengan virus yang ditularkan oleh T. vaporariorum dan B. tabaci. Selubung protein minor pada ToCV, yang membentuk bagian ekor/ujung virion memiliki peranan dalam penularan dengan kutukebul (Wintermantel et al. 2005). Terdapat interaksi antara protein selubung virus dengan kutukebul saat terjadi penempelan partikel virus dengan reseptor sehingga virus dpat tertular. Virus persisten, saat diakuisisi akan berada dalam tubuh serangga vektor. Virus berada pada sel epitel saluran pencernaan dan berasosiasi dengan kelenjar saliva serangga. Virus bersirkulasi dalam tubuh serangga sampai akhirnya virus sampai ke stilet dan masuk ke dalam tanaman sehat saat vektor makan cairan floem. Virus tersebut memerlukan waktu akuisisi dan inokulasi satu jam hingga satu hari dan periode laten satu hari hingga beberapa minggu (Gray & Banerjee 1999).
Gejala TICV dan/atau ToCV pada tanaman tomat yaitu daun menguning hanya terbatas pada tulang daun dan tulang daun tampak terlihat berwarna hijau (Gambar 5). Gejala penyakit yang diinduksi oleh ToCV dilaporkan tidak dapat dibedakan dengan gejala yang diinduksi oleh infeksi TICV (Dovas et al. 2002). Selain itu, gejala yang muncul juga akan sama bila kedua virus ini bersama-sama menginfeksi tanaman tomat (Wintermantel et al. 2008).
Gambar 5 Gejala kuning pada tanaman tomat yang telah diinokulasi dengan ToCV (kiri) dan TICV (kanan)
produksi buah berkurang dengan mengecilnya ukuran buah dan menurunnya jumlah buah.
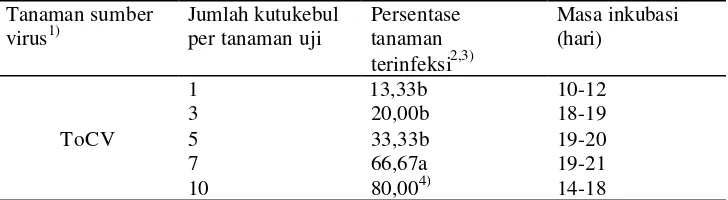
Pengujian selanjutnya dilakukan untuk mengetahui keefektifan B. tabaci dalam menularkan ToCV dan keefektifan T. vaporariorum dalam menularkan ToCV dan TICV. Pada penularan ini digunakan jumlah kutukebul yang berbeda yaitu 1, 3, 5, dan 7 untuk setiap tanaman tomat yang diinokulasi. Tabel 3 memperlihatkan hasil penularan ToCV oleh B. tabaci. Persentase tanaman terinfeksi pada uji penularan ToCV melalui kutukebul B. tabaci dengan jumlah 1, 3, dan 5 ekor kutukebul per tanaman menunjukkan hasil yang tidak berbeda nyata pada taraf nyata 5% (tanaman yang terinfeksi berturut-turut 13,33%; 20,00%; 33,33%). Akan tetapi persentase tanaman terinfeksi pada perlakuan 1-5 kutukebul per tanaman ini berbeda nyata dengan persentase tanaman terinfeksi pada perlakuan 7 kutukebul per tanaman dengan persentase tanaman terinfeksi semakin meningkat menjadi 66,67%.
Tabel 3 Keefektifan B. tabaci menularkan ToCV pada tomat dari tanaman sumber virus terinfeksi tunggal
Infeksi virus diverifikasi melalui RT-PCR (Gambar 6); 2) Rata-rata dari 3 ulangan;
3)Angka yang diikuti huruf yang berbeda pada kolom yang sama berbeda nyata (uji selang
ganda Duncan a = 0,05); 4) Perlakuan tidak diuji dengan Duncan (jumlah tanaman berbeda dengan perlakuan jumlah kutukebul 1,3,5, dan 7)
ini menjelaskan adanya kespesifikan vektor untuk dua jenis Crinivirus tersebut (Wisler et al. 1998).
Hasil di atas dengan jelas memperlihatkan bahwa semakin banyak jumlah B. tabaci yang digunakan maka persentase penularan ToCV semakin tinggi. Namun dengan jumlah B. tabaci 7 ekor per tanaman uji, tingkat penularan belum mencapai 100% dan bahkan bila menggunakan 10 ekor serangga per tanaman uji pun tingkat penularannya hanya mencapai 80%. Untuk mencapai tingkat penularan 100% tampaknya perlu menambah jumlah serangga lebih dari 10 ekor per tanaman uji.
ToCV memiliki kekhususan antara virus tanaman yang ditularkan oleh 4 spesies kutukebul, dari 2 genus (Dalmon et al. 2008). Hal ini erat kaitannya dengan kemampuan serangga vektor memperoleh virus, konsentrasi virus dalam inangnya, dan kemampuan virus untuk melewati dinding usus tengah serta bertahan di dalam hemolimfa serangga (Swenson 1967).
Tabel 4 Keefektifan T. vaporariorum menularkan ToCV dan TICV pada tomat dari tanaman sumber virus terinfeksi tunggal
Tanaman sumber
Infeksi virus diverifikasi melalui RT-PCR (Gambar 6); 2) Rata-rata dari 3 ulangan;
3)
Angka yang diikuti huruf yang berbeda pada kolom yang sama berbeda nyata (uji selang ganda Duncan a = 0,05); 4) Perlakuan tidak diuji dengan Duncan (jumlah tanaman berbeda dengan perlakuan jumlah kutukebul 1,3,5, dan 7)
kejadian penyakit oleh 7 kutukebul per tanaman (60,00%), sedangkan persentase tanaman terinfeksi pada perlakuan 3 dan 5 kutukebul per tanaman (26,67%; 40,00%) tidak berbeda nyata dengan perlakuan 1 dan 7 kutukebul per tanaman.
Pada pengujian penularan ToCV melalui kutukebul T. vaporariorum menunjukkan bahwa kejadian penyakit oleh 1 kutukebul per tanaman (6,67%) lebih rendah dibandingkan pada perlakuan 3 dan 5 kutukebul per tanaman dengan kejadian penyakit berturut-turut 26,67% dan 33,33%. Hal ini juga ditunjukkan pada perlakuan 7 kutukebul per tanaman dengan kejadian penyakit 60% menunjukkan hasil berbeda nyata dengan perlakuan 3 dan 5 kutukebul per tanaman.
Keefektifan penularan ditentukan oleh jumlah kutukebul walaupun dengan tingkat persentase tanaman terinfeksi yang beragam. Semakin sedikit jumlah kutukebul per tanaman, semakin rendah jumlah tanaman terinfeksi. Hasil penelitian Duffus et al. (1996) juga mendapatkan bahwa untuk satu ekor kutukebul per tanaman uji, rata-rata jumlah tanaman terinfeksi hanya 8%. Semakin banyak jumlah T. vaporariorum yang digunakan maka semakin tinggi juga tingkat penularan yang terjadi. Untuk TICV dengan 10 ekor T. vaporariorum penularan dapat dicapai hingga 100%, namun untuk ToCV perlu lebih dari 10 ekor untuk mencapai 100% penularan. Hasil penelitian Dalmon et al. (2008) menunjukkan bahwa perlakuan 10-60 kutukebul per tanaman mampu menularkan ToCV dengan jumlah tanaman terinfeksi 100%.
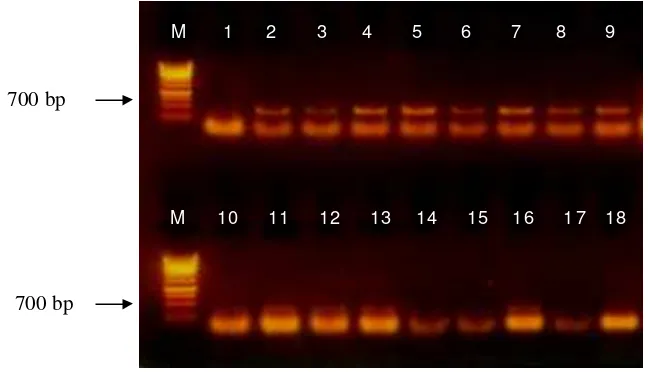
Gambar 6 Hasil amplifikasi DNA TICV dan ToCV dengan metode RT-PCR menggunakan sepasang primer TICV dan ToCV. M (marker 1 kb); lajur 1 (negatif TICV), lajur 2 - 9 (positif TICV); lajur 14, 15, 17 (negatif ToCV), lajur 10 -13, 16, 18 (positif ToCV)
M 1 2 3 4 5 6 7 8 9
M 10 11 12 13 14 15 16 1 7 18
700 bp
KESIMPULAN DAN SARAN
Kesimpulan
Dari hasil penelitian ini dapat disimpulkan bahwa: (1) Penyakit kuning pada tanaman tomat sudah banyak terjadi di daerah sentra produksi tomat terutama di dataran tinggi Jawa Barat; (2) Dua spesies ToCV dan TICV dari genus Crinivirus, ditemukan berasosiasi dengan penyakit kuning pada tanaman tomat; dan (3) ToCV efektif ditularkan baik oleh B. tabaci maupun T. vaporariorum, sedangkan TICV efektif ditularkan hanya oleh T. vaporariorum tetapi tidak dapat ditularkan oleh B. tabaci.
Saran
DAFTAR PUSTAKA
Accotto GP, Vaira AM, Vecchiati M, Finetti Sialer MM, Gallitelli D and Davino M. 2001. First report of tomato chlorosis virus in Italy. Plant Disease 85: 1208.
Anfoka GH, Abhary MK. 2007. Occurrence of Tomato infectious chlorosis virus
(TICV) in Jordan. Bulletin OEPP/EPPO 37: 186–190.
Aidawati N, Hidayat SH, Suseno R, Sosromarsono S. 2002. Transmission of an Indonesian Isolate of Tobacco leaf curl virus (Geminivirus) by Bemisia tabaci Genn. (Hemiptera: Aleyrodidae). Plant Pathol. J. 18(5) : 231-236. Borror DJ, Triplehorn, Johnson. 1996. Pengenalan Pelajaran Serangga. Edisi ke-6.
Partosoedjono S, penerjemah. Yogyakarta: Gadjah Mada University Press. Terjemahan dari: An Introduction to the Study of Insects.
Botha J, Hardie D, Power G. 2000. Spiraling whitefly Aleurodicus disperses. Exotic Threat to Western Australia. Fact Sheet no. 18/2000.
[CABI] Centre of Agriculture and Biological Institute. 1999. Crop Protection Compendium. London: CABI.
Cardona C. 2002. Whiteflies and whitefly-borne Viruses in the Tropic. Di dalam Whiteflies as pests of Annual Crops in the Tropical Highlands of Latin America. hlm 269-273.
Carver M, Gross GF, Woodward TE. 1991. Hemiptera (bugs, leafhoppers, cicadas, aphids, scale insects, etc). Di dalam The Insect of Australia. Edisi ke-2. hlm: 429-509.
Costa AS. 1969. Whiteflies as Virus Vectors. Dalam K Maramorosch (ed.). viruses, Vectors, and Vegetation. John Willey and Sons (Interscience), New York: 659 p.
Dalmon A, Bouyer S, Cailly M et al., 2005. First report of tomato chlorosis virus and tomato infectious chlorosis virus in France. Plant Disease 89, 1243. Dalmon A, Fabre F, Guilbaud L, Lecoq H, Jackuemond M. 2008. Comparative
whitefly transmission of tomato chlorosis virus and tomato infectious chlorosis virus from single or mixed infections. Plant Pathology.
Dovas, C. I., Katis, N. I., and Avgelis, A. D. 2002. Multiplex detection of criniviruses associated with epidemics of a yellowing disease of tomato in Greece. Plant Dis. 86:1345-1349.
Duffus JE, Liu HY, Coben S. 1994. Partial characterization of a new closterovirus, the causal agent of cucurbit yellow stunting disorder. Page 49 in Sweetpotato Whitefly: 1994 Supplement to the Five- Year Plan US Dep. Agric. Res. Serv. Publ.112.
Fereres A, Moreno A. 2009. Behavioural aspects influencing plant virus transmission by homopteran insects. Virus Research 141: 158–168.
Font I, Marti´nez-Culebras P, Jorda MC, Louro D, Vaira AM, Accotto GP. 2002. First report of Tomato infectious chlorosis virus in Spain. Plant Dis 86:696. Forbes AR. 1969. The stylets of the green peach aphid, Myzus persicae
(Homoptera: Aphididae). Can. Entomol. 101, 31–41.
Gray SM, Banerjee N. 1999. Mechanism of arthropod transmission of plant and animal viruses. Microbiol Mol Biol Rev 3: 128-148.
Hartono S, Natsuaki T, Sayama H, Atarashi H, Okuda S. 2003. Yellowing disease of tomatoes caused by Tomato infectious chlorosis virus newly recognized in Japan. J Gen Plant Pathol 69:61–64.
Hartono S, Wijonarko A. 2007. Karakterisasi Biologi Molekuler Tomato Infectious Chlorosis Virus Penyebab Penyakit Kuning pada Tanaman Tomat di Indonesia. Jurnal Akta Agrosia Edisi Khusus 2: 139 – 146.
Hill D. 1987. Agriculture Insect Pest of the Tropics and Their Control. Cambridge University Press.
Hoddle MS. 2004. The Biology and Management of Silverleaf Whitefly. Bemisia argentifolii Bellows and Perring (Homoptera: Aleyrodidae) on Greenhouse Grown Ornamentals. http://www.biocontrol.ucr.edu/bemisia.html#biology. [19 Januari 2009].
Isaacs R, Willis MA, Byrne DN. 1999. Modulation of whitefly take-off and flight orientation by wind speed and visual cues. Physiol. Entomol. 24, 311–318. Jacquemond M, Verdin E, Dalmon A, Guilbaud L, Gognalons P. 2008.
Serological and molecular detection of tomato chlorosis virus and tomato infectious chlorosis virus in tomato. Plant Pathology.: 1365-3059.
Jones DR, 2003. Plant viruses transmitted by whiteflies. European Journal of Plant Pathology 109: 195–219.
Kalshoven LGE. 1981. The Pests of Crop in Indonesia. PA van der Laan, penerjemah. Jakarta: Ichtiar Baru-van Hoouve.
Kataya ARA, Stavridou E, Farhan K. Livieratos IC. 2008. Nucleotide sequence analysis and detection of a Greek isolate of tomato chlorosis virus. Plant Pathology 57: 819–824.
Kennedy JS, Day MF, Eastop VF. 1962. A Conspectus of Aphids as Vectors of Plant Viruses. Commonwealth Institute of Entomology, London.
Kessing JLM, Mau RFL. 2009. Trialeurodes vaporariorum. http// www. Extento.hawaii.edu/dbase/crop/type/t.vap.htm. [Januari 2009].
Li RH, Wisler GC, Liu HY, and Duffus JE. 1998. Comparison of diagnostic techniques for detecting tomato infectious chlorosis virus. Plant Dis. 82:84-88.
Liu H- Y, Wisler GC, Duffus JE. 2000. Particle lengths of whitefly-transmitted criniviruses. Plant Disease 84: 803-805.
Louro, D., Accotto, G. P., and Vaira, A. M. 2000. Occurrence and diagnosis of tomato chlorosis virus in Portugal. Eur. J. Plant Pathol. 106:589-592. Lozano G, Moriones E, Navas-Castillo J. 2006. Complete nucleotide sequence of
the RNA2 of the crinivirus tomato chlorosis virus. Archives of Virology 151: 581-587.
Lozano G, E. Moriones, J. Navas-Castillo. 2007. Complete sequence of the RNA1 of a European isolate of tomato chlorosis virus. Archives of Virology 152: 839–841.
Masse D, Lefeuvre P, Delatte H, Karime ALA, Hostachy B, Reynaud B, Lett JM. 2008. Tomato chlorosis virus: first report in Mayotte Island. Plant Pathology 57: 388.
Markham PG, Bedford ID, Liu S, Pinner MS. 1994. The transmission of geminiviruses by Bemisia tabaci. Pesticide Science 42: 123-128.
Martelli GP, Agranovsky AA, Bar-Joseph M, Boscia D, Candresse T, Coutts RHA, Dolija VV, Duffus JE, Falk BW, Gonsalves D, Jelkmann W, Karasev AV, Minafra A, Murant A, Namba S, Niblett CL, Vetten HJ and Yoshikawa N. 2000. In: van Regenmortel MHV, Fauquet CM, Bishop DHL, Carstens EB, Estes MK, Lemon SM, Maniloff J, Mayo MA, McGeoch DJ, Pringle CR and Wickner RB (eds) Virus Taxonomy, Seventh Report of the International Committee on Taxonomy of Viruses (pp 943–952) Academic Press, San Diego.
Martelli GP, Agranovsky AA, Bar-Joseph M. 2002. The family Closteroviridae revised. Archives of Virology 147: 2039-2044.
Martin JH. 1987. An Identificaton guide to common whitefly pest spesies of the world (Hompotera: Aleyrodidae). Trop Pest Manag 33(4): 298 – 322. Martin JH. 1999. The whitefly fauna of Australia (Sternorrhyncha: Aleyrodidae)
a taxonomic account anf identification guide. CSIRO Entomologycal Technical Paper 38. 197 hlm.
Martin JH, Mifsud D, Rapisarda C. 2000. The whiteflies (Hemiptera: Aleyrodidae) of Europe and Mediterranean basin. Buletin of Entomological Research 90: 407-448.
Morales FJ. 2001. Conventional breeding for resistance to Bemisia tabaci -transmitted geminiviruses. Crop Prot 20: 825-843.
Muniyappa V, Reddy DVR. 1983. Transmission of Cowpea mild mottle virus by Bemisia tabaci in non persistent manner. Plant Dis 67: 391 – 393.
Navas-Castillo, J., Camero, R., Bueno, M., and Moriones, E. 2000. Severe yellowing outbreaks in tomato in Spain associated with infections of Tomato chlorosis virus. Plant Dis. 84:835-837.
Parrella G. 2007. Interveinal Yellowing Caused by tomato infectious chlorosis virus in Lettuce and Escarole in Southern Italy. J. Phytopathology 156: 190– 192.
Parrella G, Scassillo L. 2006. Nuove segnalazioni di Crinivirus su pomodoro in coltivazioni protette della Campania e della Calabria. Inf Fitopat 6:33–34. Rahayuwati S. 2009. Variasi morfologi puparium dan DNA penyandi gen
mitokondria sitokrom oksidase I Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). [tesis]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor. Segev, L., Wintermantel, W. M., Polston, J. E., and Lapidot, M. 2004. First report
of tomato chlorosis virus in Israel. Plant Dis. 88:1160.
Smith PE. 2009. Whitefly: identification and biology in New Zealand greenhouse tomato crops. Factsheet 1. Horticulture New Zeland. Fresh tomato product group.
Swenson KG. 1967. Plant virus transmission by insect. Di dalam: Maramorosch K, Koprowski H, Editor. Methods in Virology. Academic Press. Hlm 267-307.
Sylvester ES. 1956. Beet yellows virus transmission by the Green peach aphid. J. Econ. Entomol. 49, 789–800.
Tsai WS, Shih SL, Green SK, Hanson P. 2004. First report of the occurrence of tomato chlorosis virus and tomato infectious chlorosis virus in Taiwan. Plant Disease 88, 311.
Verhoeven JTJ, Willemen TM, Roenhorst JW, van der Vlugt RAA. 2003. First report of tomato infectious chlorosis virus in tomato in Indonesia. Plant Dis 87:872.
Vaira AM, Accotto GP, Vecchiati M, Bracaloni M. 2002. tomato infectious chlorosis virus causes leaf yellowing and reddening of tomato in Italy. Phytoparasitica 30:290–294.
Watson MA, Robers FM. 1939. A comparative study of the transmission of Hyoscyamus virus 3, potato virus Y and cucumber virus 1 by the vectors Myzuspersicae (Sulz), M. circumflexus (Buckton), and Macrosiphum gei (K). Proc. R. Soc. Lond. B 127, 543–577.
Wintermantel WM, Cortez AA, Anchieta AG, Gulati-Sakhuja A, Hladky LL. 2008. Co- infection by two Crinivirus alters accumulation of each virus in a host-specific manner and influences efficiency of virus transmission. Phytopathology doi:10.1094/PHYTO-98-12-1340.
Wintermantel WM, Wisler GC, 2006. Vector specificity, host range, and genetic diversity of tomato chlorosis virus. Plant Disease 90, 814–9.
Wintermantel WM, Wisler GC, Anchieta AG, Liu H-Y, Karasev AV, Tzanetakis IE. 2005. The complete nucleotide sequence and genome organization of tomato chlorosis virus. Archives of Virology 150: 2287-2298.
Wintermantel WM. 2004. Emergence of greenhouse whitefly (Trialeurodes vaporariorum) transmitted criniviruses as threats to vegetable and fruit production in North America. APSnet feature (www.apsnet.org/online/feature/whitefly/).
Wisler GC, Liu HY, Klaassen VA, Duffus JE, Falk BW. 1996. tomato infectious chlorosis virus has a bipartite genome and induces phloem limited inclusions characteristic of the closteroviruses. Phytopathology 86:622–626. Wisler, GC., Duffus, JE., Liu, HY. and Li, RH. 1998. Ecology and epidemiology
of whitefly-transmitted closteroviruses. Plant Disease 82: 270-280.
Wisler GC, Li RH, , Lowry DS, Duffus JE. 1998. tomato clorosis virus: a new whitefly-transmitted, phloem-limited, bipartite closterovirus of tomato. Phytopatology 88: 402-409.
Wisler GC, Duffus JE. 2001. Transmission properties of whitefly-borne criniviruses and their impact on virus epidemiology. In: Harris KF, Smith OP, Duffus JE (eds), Virusi nsect–plant interactions. Academic Press, San Diego, pp 293–308.