A mathematical model predicting the frequency of aberrant
rearrangements in the T-cell receptor gene
Edward G. Brooks *
Child Health Research Center,Department of Pediatrics,Uni6ersity of Texas Medical Branch,301Uni6ersity Boule6ard,Gal6eston, TX77555–0366,USA
Received 8 March 1999; received in revised form 18 May 1999; accepted 22 June 1999
Abstract
The T-cell receptor (TCR) genetic loci undergo an orderly process of recombination in ontogeny in order to generate a diverse array of antigen receptors. Normally occurring, out-of-frame and incomplete rearrangements produce non-productive TCR transcripts. Abnormalities in the rearrangement process occur at very low frequencies but may predominate in inborn errors of recombination. Detecting these abnormalities in surviving pools of lymphocytes is difficult and typically focuses on identification of abnormally rearranged alleles or on detecting abnormalities in recombinase proteins. Thus, there currently exists no rapid screening method to identify aberrant V(D)J recombination. To address this issue, a mathematical model was developed to predict the error rate from the measured proportions of different non-productive TCR alleles. Since the proportions of different non-productive rearrangements vary in a characteristic fashion in response to abnormalities in the recombination process, the mathematical model presented here provides a tool to indirectly assess the error rate of TCR recombination. The model was applied to a group of patients with Omenn’s syndrome, most of whom had an unknown primary defect. The results indicate that these patients had a \90% rate of aberrant TCR recombination. © 1999 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:T-cell receptor; Recombination; Recombinase; Activating gene
www.elsevier.com/locate/biosystems
1. Introduction
The goal of the current study was to develop a mathematical model that could predict the fre-quency of aberrant recombinations that occur in T-cell receptor (TCR) rearrangements during
thymic ontogeny. The model is based on the frequency of non-productive TCR alleles that oc-cur under normal and abnormal conditions. It is presumed that abnormal recombination events oc-cur at low frequencies in normal individuals. However, in genetic errors of the recombination machinery, such as those identified within the murine SCID defect, the level of recombinatorial errors increases to the point where few, if any, T or B cells bearing productive rearrangements
sur-* Corresponding author. Tel.: +1-409-7723129; fax: + 1-409-7721761.
E-mail address:[email protected] (E.G. Brooks)
vive to populate the thymus, bone marrow and peripheral lymphoid organs. In some conditions, putative defects in TCR and immunoglobulin re-arrangement mechanisms may be suspected but unproven. The model described here identifies patterns of non-productive rearrangements within surviving T cells, that predict the degree to which recombination errors occurred during thymic on-togeny without the necessity of a priori identifica-tion of the specific abnormalities in the process. Currently, investigations of these defects rely on methods such as sequencing of abnormal products of rearrangement or using artificial recombination cassettes transfected into cell lines bearing poten-tial defects in recombination machinery (Ichihara et al., 1988; Schwarz et al., 1991; Hsieh et al., 1993; Taccioli et al., 1993; Abe et al., 1994). In the first case, it is difficult to detect recombination abnormalities, since many result in DNA double strand breaks, and target the cell for death, thus removing that abnormality from the surviving T-cell population (Cantoni et al., 1996). The sec-ond method is technically difficult and is usually applied to cases where obvious defects are sus-pected (Nicolas et al., 1998). The mathematical relationships developed here, when applied to sur-viving T-cell populations, should reflect patterns of expression of non-productive TCR alleles that would indicate normal or defective recombination mechanisms. Rapid methods are available to quickly identify the proportions of non-produc-tive alleles and thus provide a convenient tool for the detection of novel errors. One such method, the inverse PCR, was applied to a population of patients with the Omenn’s syndrome, the results of which are presented here (Brooks et al., 1999).
2. Results
The TCR genetic loci undergo somatic recom-bination during ontogeny to generate a diverse array of receptors (Chien et al., 1984). Each of these genetic loci, TCRA, TCRB, TCRG, and TCRD are made up of gene clusters of variable (V), diversity (D), joining (J), and constant (C) region gene segments. Within the human TCR loci, there are 60 TCRB and TCRA variable
segments, 2 TCRB D segments, and 13 TCRB J segments, and 60 TCRA J gene segments with which to recombine (Clark et al., 1984; Yoshikai et al., 1984, 1985, 1986; Hayday et al., 1985; Toyonaga et al., 1985; Concannon et al., 1986; Tillinghast et al., 1986; Lai et al., 1988) TCR proteins are encoded by in-frame VDJC tran-scripts (for b and d chains) or a VJC transcript (foraandgchains). To produce these transcripts, the VDJ or VJ segments are produced via recom-bination. Initially, D to J rearrangement brings a D gene in proximity to a J segment, and subse-quently a V region gene segment is recombined to the DJ segment, giving a final VDJ rearrangement at the TCRB and TCRD loci. During recombina-tion, the V to D, and D to J joints recombine imperfectly, allowing both exonucleolytic loss of nucleotides and insertion of nucleotides within V(D)J joints by terminal deoxynucleotidyl trans-ferase (TdT) (Bogue et al., 1992; Gilfillan et al., 1993). As a result of the random nature of both the loss and addition of nucleotides within re-combining joints, there is only a 1/3 chance of recombination occurring in the proper reading frame (in-frame rearrangements).
rearrange-ment of it’s first allele with two possible out-comes: (1) an in-frame and productive VDJ rear-rangement (symbolized by (I)); or (2) an out-of-frame and non-productive VDJ rearrange-ment (symbolized by (c)). There is a probability of 1/3 for an in-frame (I) and 2/3 for an out-of-frame (c) rearrangement. However, errors in recombination may create abnormal recombina-tion events such as recombinarecombina-tion of V or D segments into intronic sequences, translocations, or unrepaired double strand breaks. These erro-neously recombined alleles will be designated as ‘abnormal’ (symbolized by (X)). The frequency of recombination events yielding abnormally recom-bined alleles, (X), will be represented by the sym-bol,A.
If abnormal recombination events occur at a given frequency, A, then the three possible allelic configurations and their frequencies after the first allele rearranges may be represented as follows:
X/G
c/G I/G
Allelic configuration
A
Frequency(f) 1/3(1−A) 2/3(1−A)
In the above scenario, the frequencies of the two normal rearrangements (I) and (c) are reduced by the occurrence of abnormal rearrangements,A, and thus their new frequencies become the product of (1−A) and 1/3 or 2/3, respectively.
As a result of the process of allelic exclusion at the TCRB locus, if the first allele rearranges pro-ductively, rearrangement of the second allele is inhibited. This process normally inhibits the V to D rearrangement yielding a truncated (T) DJ rearrangement. Rarely, D to J rearrangements may not occur, leaving the TCRB locus in an unrearranged state, but for the purposes of these calculations, this possibility will be considered negligible, and all allelically inhibited rearrange-ments at the TCRB locus will be assumed to result in truncated DJ rearrangements (T). Thus, all cells bearing productive rearrangements (I/G) on the first allele create a truncated (T) rearrange-ments on the second allele giving rise to an I/T configuration. Abnormal recombinations rarely occur in the ‘normal’ setting, so for the purposes of this exercise, that baseline frequency will be
considered negligible, and thus, in the baseline state, whereA=0, the frequency of I/T would be 1/3(1−A)=1/3.
If the first allele rearranges non-productively (c or X), then the second allele undergoes full rearrangement to yield three possibilities each,
c/I, c/c, or c/X, or X/I, X/c, or X/X. The frequencies of the various allelic combinations become the products of the individual allelic fre-quencies (Fig. 1). In the baseline state, A=0, there are no alleles bearing abnormal rearrange-ments (X), and the probability of an in-frame productive rearrangement on either allele (I/T+ c/I) is: 1/3(1−A)+2/3(1−A)1/3(1−A)=0.56, and the probability of an out-of-frame non-pro-ductive rearrangement on both alleles (c/c) is: (2/3(1−A))2=0.44. These baseline values agree
with previously published mathematical models and experimental results (Malissen et al., 1992). In Fig. 2, The frequency of these various
combina-Fig. 1. Allelic combinations and frequency formlas of TCRB gene rearrangements. The frequency of each rearrangement is shown below each combination. G, germline; I, full length in-frame VDJ rearrangement; T, truncated DJ rearrangement;
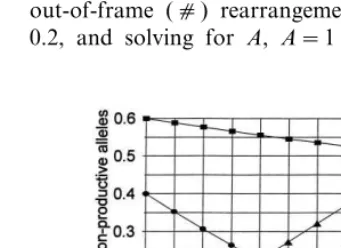
Fig. 2. Frequency of TCRB gene allelic combinations. As the frequency (A) of abnormal rearrangements (X) increases, the frequencies of all other possible combinations decrease pro-portionately. The frequencies of the possible allelic combina-tions for the first and second allelic rearrangements are shown. T, truncated DJ rearrangement; c, full length VDJ out-of-frame rearrangement; X, abnormal rearrangement.
value is approached, the frequency of surviving out-of-frame rearrangements (fsc) approaches
zero and those of both truncated (fsT) and
abnor-mal (fsX) rearrangements approach 0.5.
If all abnormal rearrangements produce double stranded breaks, such as those that occur in the murine SCID defect, then those T cells bearing abnormal rearrangements would be targeted for cell death mechanisms (Obe et al., 1992; Cantoni et al., 1996). In that case, only those T cells bearing a productive rearrangement (I) paired with a truncated (T) or out-of-frame (c) rear-rangement would survive. As the frequency of the abnormal rearrangements approaches 100% the predominant non-productive rearrangements would be truncated (T) (Fig. 4). To express this as a ratio, fsS=(fsT/fsc)=[(2/3)(1−A)(1/3)(1− A)]/[(1/3)(1−A)], which can be simplified to (2/
3)(1−A). Thus, the ratio of c/T can be expressed as a linear relationship (Fig. 5). These ratios can be used to determine the theoretical value of Ain an experimental setting where Ais unknown. For instance, if all TCRB non-produc-tive rearrangements identified within a given ex-perimental sample yield 100 truncated (T) and 20 out-of-frame (c) rearrangements, then c/T=
0.2, and solving for A, A=1−3/2(c/T)=0.7. tions (ordinate) are plotted as a function of the
frequency of abnormal recombination events (A) on the abscissa. Therefore, as the frequency of abnormal rearrangements (A) increases, the rela-tive frequencies of other rearrangements decrease. Combinations of alleles bearing both abnormal (X) and normal (I or c) rearrangements initially increase as a result of increasing A until A\0.5
(Fig. 2). The theoretical maximum for the allelic combinations of c/X+X/c is: 2/3(1−A)A+
A(2/3(1−A)=0.33. Likewise, the theoretical maximum of the allelic combination, X/I is:A(1/
3(1−A))=0.0833.
Since, at least one productive TCR rearrange-ment is a requirerearrange-ment for T-cell survival, the sum (S) of the surviving frequencies (fs) of allelic
combinations I/T, c/I, and X/I would be equiva-lent to the sum of the frequencies of non-produc-tive rearrangements in surviving T cells or:
fsS=(f(I/T)+f(c/I)+f(X/I))=fsT+fsc+ fsX=1. Those frequencies are shown in Fig. 3.
What is apparent, is that the frequency of out-of-frame rearrangements decreases at a faster rate than the truncated rearrangements. In Fig. 3, the values of the various allelic combinations that would occur at 100% abnormal rearrangements (A) cannot practically be attained, since that value would yield no productive rearrangements and, thus, no surviving T cells. However, as that
Fig. 4. Frequency of TCRB gene rearrangements in surviving T cells. In this figure, it is assumed that those T cells bearing abnormal rearrangements (X), such as the double strand breaks seen in the murine SCID mutation, would be targeted for cell death and, thus, would not appear in the surviving T-cell populations. T, truncated DJ rearrangement; c, full length VDJ out-of-frame rearrangement.
Table 1
Analysis of Omenn’s syndrome patients
Case TCRb(c/T) TCRa(c/T)
0/2 6/4
1
2 1/7 3/2
3 0/1 1/3
6/0
4 0/5
6/2 0/0
5
6 0/2 0/2
3/3
7 0/2
25/16=1.6 1/19=0.05
Total
control to ensure that the technique used in the study, the inverse PCR, did not skew the results towards increasing numbers of truncated (T) rearrangements.
3. Discussion
The mathematical model presented here was designed to predict the frequency of allelic combi-nations of TCRB non-productive gene rearrange-ments. The predictions are based upon assumptions of the orderly process by which suc-cessive rearrangements occur within a given T cell, principles of allelic exclusion, and the surviv-ability of T cells bearing abnormal rearrange-ments. The initial assumptions are that there is a 1/3 chance of rearranging in the correct reading frame and that subsequent TCRB rearrangements are inhibited by the presence of a productive TCR. These assumptions have been previously tested and proven (Malissen et al., 1992; Ferradini et al., 1993). It will be important to establish the validity of the model in an experimental situation where the frequency (A) of abnormal rearrange-ments (X) can be varied in a reliable fashion.
The model could be useful in several ways, such as the existence of cases of murine SCID with ‘leaky defects’. In murine SCID, there is a defect in the recombination machinery, as a result of a mutation in the DNA dependent protein kinase catalytic subunit, that creates a block in the rear-rangement (Schuler et al., 1986; Chang et al., 1993; Miller et al., 1995; Araki et al., 1997). However, some mouse strains with SCID have This can interpreted to suggest that 70% of all
rearrangement events were abnormal.
We recently employed this model in analyzing the TCRs from a group of patients with the Omenn’s syndrome (Table 1) (Brooks et al., 1999). In that study, the c/T ratio for TCRB was 0.05, which when applied to the model would indicate an overall error frequency (A)=92.5%. The c/T ratio for TCRA in that study was 1.6. Because of a lack of strict allelic exclusion the TCRA c/T ratio is greater than TCRB. The TCRA data thus provides a convenient internal
small numbers of T and B cells with productive antigen receptors. The ‘leakiness’ of the defect may imply that either the defect is not complete, that there may be a compensatory mechanisms unre-lated to the specific defect, or that there are genetic revertants within the T-cell population (Bosma et al., 1988; Kotloff et al., 1993). Application of the model to the surviving T cells in these animals may help to explain their origin.
The model was applied to a population of patients with a form of SCID known as Omenn’s syndrome (Brooks et al., 1999). The model pre-dicted that a \90% error rate existed during the
ontogeny of the TCR’s. Many patients with the Omenn’s syndrome have been shown to have defects in the recombinase complex (partial RAG defects) (Villa et al., 1998). A high error rate of TCR recombination would explain the small num-bers of surviving oligoclonal T cells in these patients and is consistent with a postcleavage role for RAG proteins in V(D)J recombination (Ramsden et al., 1997; Besmer et al., 1998; Shockett and Schatz, 1999).
Alternatively, if the initiation of recombination is faulty, as may be the case in certain RAG defects, then it is possible that the explanation for the increased proportion of truncated rearrangements in circulating T cells, may be as a result of a lack of initiation and completion of the rearrangement at the second TCR locus. If that were the case, one might expect a similar effect at the TCR a locus, but there appear to be abundant numbers of TCR
a out-of-frame rearrangements. In contrast, there does appear to be an increased number of unrear-ranged TCRbgenes in some patients. This could suggest a problem with initiation. We are aware of the status of the RAG genes in only two patients, patient number 4 (normal) and patient number 1 (DiGeorge Syndrome, 22q11 deletion) (Brooks et al., 1999). Thus, a correlation of RAG function with the proportion of non-productive alleles in surviving T cells in these patients cannot be con-cluded at this time, but is the subject of an ongoing investigation.
The utility of the model in situations of overt recombination defects and severe immune defi-ciency is obvious. However, application of the model may prove quite useful in situations in which
the recombinatorial error rate is increased, but not to the point of overt immune deficiency. The T- and B-cell repertoires are normally so large as to preclude the ability to detect a reduction in the repertoire as a marker of partial recombination defects. In these situations, the ratios of out-of-frame to truncated rearrangements, as predicted in the model would be a sensitive measure of partial abnormalities.
Acknowledgements
Sergei Atamas, Judah Rosenblatt, Randall M. Goldblum, and Nancy Van Houten for critical review of the manuscript. E. Brooks was supported by a National Institutes of Health Clinical Investi-gator Award, K08 AI01064.
References
Abe, T., Tsuge, I., Kamachi, Y., Torii, S., Utsumi, K., Akahori, Y., Ichihara, Y., Kurosawa, Y., Matsuoka, H., 1994. Evi-dence for defects in V(D)J rearrangements in patients with severe combined immunodeficiency. J. Immunol. 152, 5504 – 5513.
Araki, R., Fujimori, A., Hamatani, K., Mita, K., Saito, T., Mori, M., Fukumura, R, Morimyo, M., Muto, M., Itoh, M., Tatsumi, K., Abe, M., 1997. Nonsense mutation at Tyr-4046 in the DNA-dependent protein kinase catalytic subunit of severe combined immune deficiency mice. Proc. Natl. Acad. Sci. USA 94, 2438 – 2443.
Besmer E., Mansilla-Soto J., Cassard S., Sawchuk D.J., Brown G., Sadofsky M., Lewis S.M., Nussenzweig M.C., Cortes P., 1998. Hairpin coding end opening is mediated by RAG1 and RAG2 proteins. Molecular Cell 2, 817 – 828.
Bogue, M., Gilfillan, S., Benoist, C., Mathis, D., 1992. Regula-tion of N-region diversity in antigen receptors through thymocyte differentiation and thymus ontogeny. Proc. Natl. Acad. Sci. USA 89, 11011 – 11015.
Bosma G.C., Fried M., Custer R.P., Carroll A., Gibson D.M., Bosma M.J., 1988. Evidence of functional lymphocytes in some (Leaky)scidmice. J. Exp. Med, 167, 1016 – 1033. Brooks, E.G., Filipovich, A.H., Padgett, J.W., Mamlock, R.,
Goldblum, R.M., 1999. T-cell receptor analysis in Omenn’s syndrome: Evidence for defects in gene rearrangement and assembly. Blood 93, 242 – 250.
Chang, C., Biedermann, K.A., Mezzina, M., Brown, J.M., 1993. Characterization of the DNA double strand break repair defect inscidmice. Cancer Res. 53, 1244 – 1248. Chien, Y., Gascoigne, N.R.J., Kavaler, J., Lee, N.E., Davis,
M.M., 1984. Somatic recombination in a murine T-cell receptor gene. Nature 309, 322 – 326.
Clark, S.P., Yoshikai, Y., Taylor, S., Siu, G., Hood, L., Mak, T.W., 1984. Identification of a diversity segment of human T-cell receptor b-chain, and comparison with the analogous marine element. Nature 311, 387 – 389. Concannon, P., Pickering, L.A., Kung, P., Hood, L., 1986.
Diversity and structure of human T-cell receptorb-chain variable region genes. Proc. Natl. Acad. Sci. USA 83, 6598 – 6602.
Ferradini, L., Roman Roman, S., Azogui, O., Genevee, C., Viel, S., Hercend, T., Triebel, F., 1993. The use of an-chored polymerase chain reaction for the study of large numbers of human T-cell receptor transcripts. Mol. Im-munol. 30, 1143 – 1150.
George, J.F. Jr., Schroeder, H.W. Jr., 1992. Developmental regulation of D beta reading frame and junctional diversity in T-cell receptor-beta transcripts from human thymus. J. Immunol. 148, 1230 – 1239.
Gilfillan, S., Dierich, A., Lemeur, M., Benoist, C., Mathis, D., 1993. Mice lacking TdT: mature animals with an immature lymphocyte repertoire. Science 261, 1175 – 1178.
Hayday, A.C., Diamond, D.J., Tanigama, G., Heilig, J.S., Folsom, V., Saito, H., Tonegawa, S., 1985. Unusual organ-isation and diversity of T-cell receptora-chain genes. Na-ture 316, 828 – 832.
Hsieh, C.L., Arlett, C.F., Lieber, M.R., 1993. V(D)J recombi-nation in ataxia telangiectasia, Bloom’s syndrome, and a DNA ligase I-associated immunodeficiency disorder. J. Biol. Chem. 268, 20105 – 20109.
Ichihara, Y., Matsuoka, H., Tsuge, I., Okada, J., Torii, S., Yasui, H., Kurosawa, Y., 1988. Abnormalities in DNA rearrangements of immunoglobulin gene loci in precursor B cells derived from x-linked agammaglobulinemia patient and a severe combined immunodeficiency patient. Im-munogenetics 27, 330 – 337.
Kotloff, D.B., Bosma, M.J., Ruetsch, N.R., 1993. V(D)J re-combination in peritoneal B cells of leaky scid mice. J. Exp. Med. 178, 1981 – 1994.
Lai, E., Concannon, P., Hood, L., 1988. Conserved organiza-tion of the human and murine T-cell receptor b-gene families. Nature 331, 543 – 546.
Malissen, M., Trucy, J., Jouvin Marche, E., Cazenave, P.A., Scollay, R., Malissen, B., 1992. Regulation of TCR alpha and beta gene allelic exclusion during T- cell development. Immunol. Today 13, 315 – 322.
Miller, R.D., Hogg, J., Ozaki, J.H., Gell, D., Jackson, S.P., Riblet, R., 1995. Gene for the catalytic subunit of mouse DNA-dependent protein kinase maps to the scid locus. Proc. Natl. Acad. Sci. USA 92, 10792 – 10795.
Nicolas, N., Moshous, D., Cavazzana-Calvo, M., Papado-poulo, D., de Chasseval, R., Le Deist, F., Fischer, A., De Villartay, J.P., 1998. A human severe combined
im-munodeficiency (SCID) condition with increased sensitivity to ionizing radiations and impaired V(D)J rearrangements defines a new DNA recombination/repair deficiency. J. Exp. Med. 188, 627 – 634.
Obe, G., Johannes, C., Schulte-Frohlinde, D., 1992. DNA double-strand breaks induced by sparsely ionizing radia-tion and endonucleases as critical lesions for cell death, chromosomal aberrations, mutations and oncogenic trans-formation, [Review] [127 refs]. Mutagenesis 7, 3 – 12. Ramsden, D.A., Paull, T.T., Gellert, M., 1997. Cell-free V(D)J
recombination. Nature 388, 488 – 491.
Schuler, W., Weiler, I.J., Schuler, A., Phillips, R.A., Rosen-berg, N., Mak, T.W., Kearney, J.F., Perry, R.P., Bosma, M.J., 1986. Rearrangement of antigen receptor genes is defective in mice with severe combined immune deficiency. Cell 46, 963 – 972.
Schwarz, K., Hansen Hagge, T.E., Knobloch, C., Friedrich, W., Kleihauer, E., Bartram, C.R., 1991. Severe combined immunodeficiency (SCID) in man: B cell-negative (B-) SCID patients exhibit an irregular recombination pattern at the JH locus. J. Exp. Med. 174, 1039 – 1048.
Shockett, P.E., Schatz, D.G., 1999. DNA hairpin opening mediated by RAG1 and RAG2 proteins. Mol. Cell. Biol. 9, 4159 – 4166.
Taccioli, G.E., Rathbun, G., Oltz, E., Stamato, T., Jeggo, P.A., Alt, F.W., 1993. Impairment of V(D)J recombination in double-strand break repair mutants. Science 260, 207 – 210.
Thompson, S.D., Manzo, A.R., Pelkonen, J., Larche, M., Hurwitz, J.L., 1991. Developmental T-cell receptor gene rearrangements: relatedness of the alpha/beta and gamma/
delta T-cell precursor. Eur. J. Immunol. 21, 1939 – 1950. Tillinghast, J.P., Behlke, M.A., Loh, D.Y., 1986. Structure
and diversity of the human T-cell receptor b-chain variable region genes. Science 233, 879 – 883.
Toyonaga, B., Yoshikai, Y., Vadasz, V., Chin, B., Mak, T.W., 1985. Organization and sequences of the diversity, joining, and constant region genes of the human T-cell receptorb
chain. Proc. Natl. Acad. Sci. USA 82, 8624 – 8628. Villa, A., Santagata, S., Bozzi, F., Giliani, S., Frattini, A.,
Imberti, L., Gatta, L., Ochs, H., Schwartz, K., No-tarangelo, L., Vezzoni, P., Spanopoulou, E., 1998. Partial V(D)J recombination activity leads to Omenn Syndrome. Cell 93, 885 – 896.
Yoshikai, Y., Anatoniou, D., Clark, S.P., Yanagi, Y., Sang-ster, R., Van den Elsen, P., Terhorst, C., Mak, T.W., 1984. Sequence and expression of transcripts of the human T-cell receptorb-chain genes. Nature 312, 521 – 524.
Yoshikai, Y., Clark, S.P., Taylor, S., Sohn, U., Wilson, B.I., Minden, M.D., Mak, T.W., 1985. Organization and se-quences of the variable, joining and constant region genes of the human T-cell receptor a-chain. Nature 316, 837 – 840.
Yoshikai, Y., Kimura, N., Toyonaga, B., Mac, T.W., 1986. Sequences and repertoire of human T-cell receptora-chain variable region genes in mature T lymphocytes. J. Exp. Med. 164, 90 – 103.