4 II TINJAUAN PUSTAKA
2.1. Xilan
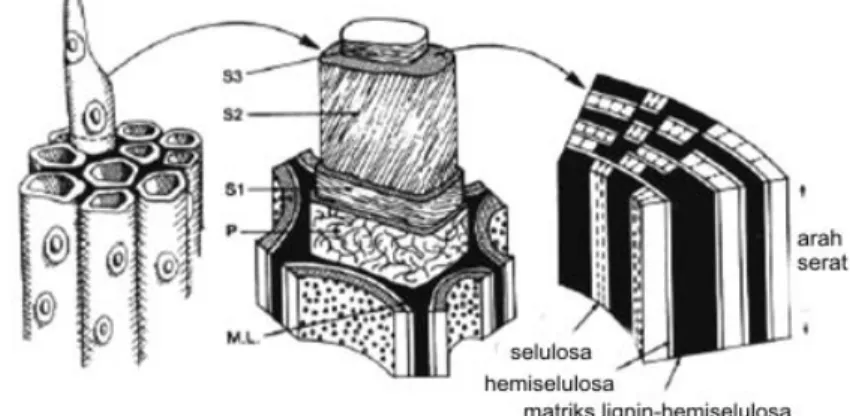
Lignoselulosa merupakan komponen utama tanaman yang menggambarkan jumlah sumber bahan organik yang dapat diperbaharui. Lignoselulosa terdiri atas selulosa, hemiselulosa, lignin dan beberapa bahan ekstraktif lain. Semua komponen lignoselulosa terdapat pada dinding sel tanaman. Xilan merupakan bagian hemiselulosa, yang merupakan komponen penyusun dinding sel tanaman kedua terbesar setelah selulosa. Susunan dinding sel tanaman terdiri atas lamela tengah (M), dinding primer (P) serta dinding sekunder (S) yang terbentuk selama pertumbuhan dan pendewasaan sel yang terdiri atas lamela transisi (S1), dinding sekunder utama (S2) dan dinding sekunder bagian dalam (S3) (Gambar 1).
Gambar 1. Konfigurasi dinding sel tanaman (Perez et al. 2002).
Hemiselulosa relatif lebih mudah dihidrolisis dengan asam menjadi monomer yang mengandung glukosa, manosa, galaktosa, xilosa dan arabinosa. Hemiselulosa mengikat lembaran serat selulosa membentuk mikrofibril yang meningkatkan stabilitas dinding sel. Hemiselulosa juga berikatan silang dengan lignin membentuk jaringan kompleks dan memberikan struktur yang kuat.
Xilan adalah polisakarida non pati yang menghasilkan monomer gula sederhana berupa xilooligosakarida, xilobiosa, dan xilosa selama proses hidrolisis oleh enzim xilanase. Xilan ditemukan dalam fraksi hemiselulosa pada
5 tumbuhan bersama dengan arabinan, galaktan, manan dan asam uronat (Leeson dan Summers 2001). Xilan terikat pada selulosa, pektin, lignin dan polisakarida lainnya dalam angiosperma untuk membentuk dinding sel tanaman.
Xilan dengan rantai utama homopolimer unit β-D-xilopiranosil yang terikat melalui ikatan (1→4)-β- glikosidik merupakan heteropolimer yang dihubungkan dengan rantai samping dari gula yang lain, umumnya rantai tunggal dari (4-O-metil)-α-D-asam glukuronat (pada dikotil dan gimnosperma) atau pada satu atau lebih α-L-arabinofuranosil (pada rumput) (Singleton dan Sainsbury 2001). Rantai xilan bercabang dan strukturnya tidak terbentuk kristal sehingga lebih mudah dimasuki pelarut dibandingkan dengan selulosa (Richana 2002).
Xilan terdapat hampir pada semua tanaman, kebanyakan dijumpai pada tanaman tahunan dan limbah-limbah pertanian seperti tongkol jagung, bagas tebu, jerami padi, dedak gandum, dan biji kapas (Subramaniyan dan Prema 2002). Xilan sebagai komponen utama penyusun polisakarida hemiselulosa pada tanaman kandungannya mencapai sekitar 30-35% berat kering totalnya. Xilan kayu keras dari golongan Angiospermae kandungannya lebih tinggi daripada kayu lunak dari golongan Gymnospermae dengan jumlah secara berturut-turut sekitar 15-30% dan 7-12% dari berat kering total (Beg et al. 2001).
2.2. Enzim Xilanase
Xilanase (1,4-ß-D-xylan xylanohidrolase, EC 3.2.1.8) merupakan hemiselulase utama yang menghidrolisis ikatan β-1,4 rantai xilan. Enzim Xilanase merupakan enzim ekstraseluler yang dapat menghidrolisis secara acak ikatan glikosidik internal (ß1,4) rantai utama xilan (hemiselulosa) menjadi gula penyusunnya xilosa dan xilooligosakarida (Eriksson et al. 1990). Xilanase memotong ikatan xilosidik internal pada kerangka xilan dan β-xilosidase melepaskan residu xilosil melalui pemotongan bagian ujung dari xilooligosakarida (Saha 2003).
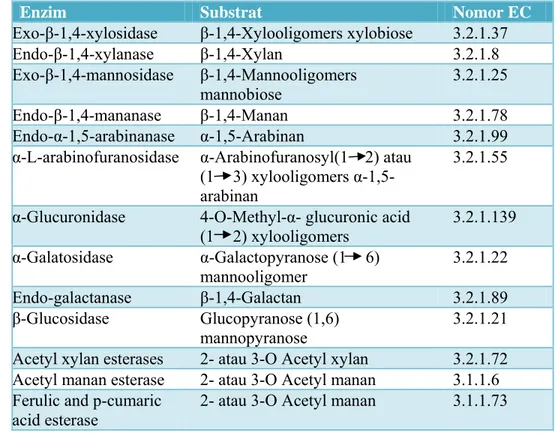
Hidrolisis hemiselulosa juga membutuhkan enzim pelengkap yang bekerja secara sinergis dalam menguraikan xilan dan manan (Tabel 1). Xilanase dapat diklasifikasikan berdasarkan substrat yang dihidrolisis, yaitu β-xilosidase,
6 eksoxilanase, dan endoxilanase. Enzim β-xilosidase mampu menghidrolisis xilooligosakarida rantai pendek menjadi xilosa. Eksoxilanase memutus rantai polimer xilosa (xilan) pada ujung pereduksi, sehingga menghasilkan xilosa sebagai produk utama dan sejumlah oligosakarida rantai pendek. Endoxilanase mampu memutus ikatan β 1-4 pada bagian dalam rantai xilan secara teratur. Xilanase pada umumnya merupakan protein kecil dengan berat molekul 15 kDa – 30 kDa, aktif pada suhu 55oC dengan pH 9 (Yang et al.1988; Yu et al. 1991). Tabel 1. Hemiselulase dan jenis substrat yang dihidrolisis (Howard et al. 2003)
Pada suhu 60oC dan pH netral, xilanase yang dihasilkan oleh Streptomyces thermoviolaceus OPC-520 bersifat lebih stabil (Tsujibo et al. 1992). Xilanase dari bakteri Streptomyces.sp (strain Lb 24D) mempunyai aktifitas tinggi pada kisaran pH 5-8, dengan pH optimal 6,5 (Rawashdeh et al. 2005).
Penambahan xilanase pada pakan dasar gandum dapat menurunkan viskositas digesta dan meningkatkan pertambahan bobot badan ayam broiler pada usia 6 minggu hingga 14,72% dan 2,60% (Chiang et al. 2005). Xilanase dapat menurunkan viskositas digesta dengan cara menghidrolisis arabinoxilan
Enzim Substrat Nomor EC
Exo-β-1,4-xylosidase β-1,4-Xylooligomers xylobiose 3.2.1.37
Endo-β-1,4-xylanase β-1,4-Xylan 3.2.1.8 Exo-β-1,4-mannosidase β-1,4-Mannooligomers mannobiose 3.2.1.25 Endo-β-1,4-mananase β-1,4-Manan 3.2.1.78 Endo-α-1,5-arabinanase α-1,5-Arabinan 3.2.1.99 α-L-arabinofuranosidase α-Arabinofuranosyl(1 2) atau
(1 3) xylooligomers α-1,5-arabinan
3.2.1.55
α-Glucuronidase 4-O-Methyl-α- glucuronic acid (1 2) xylooligomers 3.2.1.139 α-Galatosidase α-Galactopyranose (1 6) mannooligomer 3.2.1.22 Endo-galactanase β-1,4-Galactan 3.2.1.89 β-Glucosidase Glucopyranose (1,6) mannopyranose 3.2.1.21
Acetyl xylan esterases 2- atau 3-O Acetyl xylan 3.2.1.72 Acetyl manan esterase 2- atau 3-O Acetyl manan 3.1.1.6 Ferulic and p-cumaric
7 menjadi arabinosa dan xilosa, sehingga mudah dimanfaatkan oleh unggas (Choct 1997). Perlakuan panas selama proses pelleting pada industri pakan mempunyai beberapa keuntungan yaitu, dapat melarutkan sebagian serat larut dalam air seperti glukan, arabinoxilan dan pektin (Thomas et al. 1998) dan melarutkan serat kasar sebesar 8,7% (Vranjes dan Wenk 1995), sehingga dapat meningkatkan kecernaan bahan pakan. Penambahan enzim pada pakan mensyaratkan karakter enzim yang stabil selama proses pelleting. Penggunaan enzim xilanase termostabil diharapkan dapat memenuhi tuntutan tersebut.
2.3. Mekanisme Pemecahan Xilan
Aktifitas xilanase menyebabkan proses hidrolisis xilan. Beberapa model telah diusulkan untuk menjelaskan mekanisme pemecahan xilan oleh xilanase. Secara umum proses hidrolisis merupakan hasil dari retensi atau inversi pusat anomerik monomer gula pereduksi dari rantai karbohidrat. Hal ini akan melibatkan satu atau dua kondisi transisi kimiawi. Transfer Glikosil pada umumnya adalah hasil dari substitusi nukleofil pada karbon jenuh dari pusat anomer dan digantikan baik oleh retensi maupun inversi dari konfigurasi anomerik. Sebagian besar enzim penghidrolisis polisakarida, seperti selulase dan xilanase dikenal sebagai enzim yang menghidrolisis substratnya dengan cara retensi atom C1 dari konfigurasi anomeriknya. Hal ini melibatkan mekanisme pemindahan ganda untuk retensi anomerik produk (Kulkarni et al. 1999).
Mekanisme pemindahan ganda (double displacement) melibatkan beberapa hal sebagai berikut: (1). Katalis asam yang memprotonasi substrat; (2) gugus karboksil dari enzim terbentuk (3) ikatan kovalen glikosil enzim antara (intermediate) dengan karboksilat tersebut dimana konfigurasi anomer dari gula berseberangan dengan substrat; (4) ikatan kovalen intermediate ini dicapai dari kedua sisi melalui kondisi transisi yang melibatkan ion-ion oksokarbonium ; (5) berbagai variasi interaksi non kovalen lebih banyak menyebabkan laju pendegradasian (Subramaniyan dan Prema 2002).
Berdasarkan studi kristalografi xilopentaose binding xilanase A dari Pseudomonas fluorescense, Leggio et al. (2000) menyatakan mekanisme enzim yang mengkombinasikan konsep klasik seperti diatas dan fakta yang diturunkan
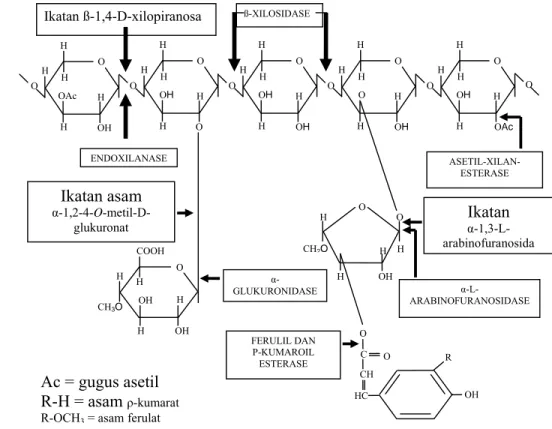
8 dari studi mereka. Berdasarkan kajian tersebut (1) xilan dikenali dan diikat oleh xilanase sebagai tangan kiri heliks tiga lipatan, (2) residu xilosil pada subsite-1 didistorsikan dan ditarik menuju residu katalitik dan ikatan glikosidik diregangkan dan putus membentuk intermediate kovalen enzim-substrat, (3) bentukan intermediate ini diserang oleh molekul air teraktivasi, mengikuti mekanisme klasik retensi glikosil hidrolase dan produk dilepaskan. Struktur xilan dan mekanisme pemecahannya oleh enzim disajikan pada Gambar 2.
Gambar 2. Struktur xilan dengan residu pengganti dan letak ikatan yang dipecah oleh enzim xilanolitik (Beg et al. 2001).
Beberapa laporan mengenai pola hidrolitik xilanase dari Bacillus spp. sebagian besar menghasilkan xilobiosa, xilotriosa, dan xilotetraosa, dan pembentukan xilosa berlangsung hanya pada proses perpanjangan inkubasi. Xilanase A dan B dari Trichoderma reesei dan xilanase C dan D dari Trichoderma harzianum pada kombinasi yang berbeda, menunjukkan interaksi sinergis pada substrat xilan yang berbeda. Kombinasi beberapa xilanase lebih
ENDOXILANASE O H H H H OH OH COOH CH3O Ikatan asam α-1,2-4-O-metil-D-glukuronat Ikatan α-1,3-L-arabinofuranosida α-L-ARABINOFURANOSIDASE ASETIL-XILAN-ESTERASE α-GLUKURONIDASE H O H H H O OH CH2O O C O CH HC OH R FERULIL DAN Ρ-KUMAROIL ESTERASE
Ikatan ß-1,4-D-xilopiranosa ß-XILOSIDASE
Ac = gugus asetil R-H = asam ρ-kumarat R-OCH3 = asam ferulat
H O H O H H H OAc OH H O H H H H OH O O H O H H H H OH OH O H O H H H H O OH O H O H H H H OH OAc O O
e d d x r x d N x d d 2 d b B k m m efektif darip dan Maring dari Bacillu xilobiosa, xi residu xilo xilotetraosa, disukai untu Namun dem secara cepa xilosa, xilob diacu dalam dapat dilihat 2.4. Dedak Deda dari hasil ik bahan kerin BETN (karb kaya akan f mengandung menyebabka pada xilanase er 1999 dia s circulans ilotriosa dan opiranosida , diluar subs uk membuat mikian xilana at menjadi biosa dan x m Subramani t pada Gamb Gambar 3. S ( Gandum (P ak gandum m kutan pengol ng 86,66%, p bohidrat) 60 fosfor tetapi g vitamin A an dedak ga e tunggal unt acu dalam S WL-12 men n xilotetraos untuk m strat lain ya t jenuh semu ase I dari ba xilotetraosa ilotriosa seb iyan dan Pr bar 3. Struktur tiga (http://grayla Pollard) merupakan s ahan gandum protein kasa 0,32% dan a i rendah kad dan D tetap andum tidak tuk menghid Subramaniya nghidrolisis sa. Enzim i membentuk ang dicoba, ua situs pen akteri yang s dan perpa bagai produk rema 2002) a dimensi xil ab.jhu.edu/~ salah satu ba m. Kandung ar 13,91%, s abu 3,14% ( dar Ca-nya, i kaya akan k dapat dik drolisis holo an dan Prem xilan menja ni membutu komplek merupakan ngikatan (bin sama mampu anjangan ink k akhirnya ). Struktur t lanase tampa ~mdaily/capr ahan pakan t gan nutrisi d serat kasar 1 (Pantaya 20 yaitu P=1,2 vitamin B. konsumsi se selulosa pinu ma 2002). X adi produk u uhkan minim produktif, substrat yan nding site) e u mendegrad kubasi men (Esteban et tiga dimensi ak atas ri/gallery/T1 ternak yang dedak gandu 16,49%, lem 003). Dedak 29%;Ca=0,1 Tingginya s cara berlebi 9 us (Wong ilanase II utamanya mal empat sehingga ng paling enzim ini. dasi xilan nghasilkan al. 1982 i xilanase 7.png). diperoleh um adalah mak 2,8%, k gandum 3% tidak erat kasar ihan oleh
10 ayam broiler, karena akan menganggu efektifitas digesta dan absorbsi yang akhirnya dapat mengganggu penyerapan energi dan protein ransum.
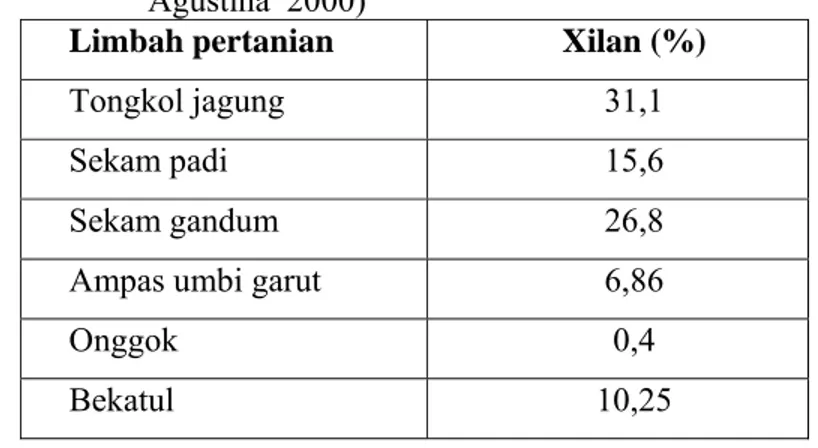
Serat kasar merupakan komponen penyusun dinding sel tanaman. Polisakarida non pati merupakan komponen serat kasar yang terdiri atas selulosa, polimer non selulosa dan pektin polisakarida. Polimer non selulosa terdiri atas arabinoxylan, β-glukan, manan, galaktan dan xiloglukan (Choct 1997). Komponen - komponen tersebut banyak terdapat dalam bahan pakan seperti barley, oat, gandum, rye dan triticale yang telah dibuktikan dapat menghambat kecernaan pati, nutrisi lain dan meningkatkan viskositas digesta (Campbell dan Bedford 1992). Leeson dan Summers (2001) menyatakan batas penggunaan pollard bagi ayam muda petelur maksimal 8%, ayam breeding petelur maksimal 15%, breeding ayam pedaging 10%, dan ayam pedaging usia 4-8 minggu batas penggunaan pollard 20%. Kadar xilan dari beberapa sumber limbah agroindustri dapat dilihat pada Tabel 2.
Tabel 2. Kadar xilan berbagai limbah industri pertanian (Parajo 2004; Agustina 2000)
Limbah pertanian Xilan (%)
Tongkol jagung 31,1
Sekam padi 15,6
Sekam gandum 26,8
Ampas umbi garut 6,86
Onggok 0,4
Bekatul 10,25
2.5. Pemurnian Protein
Tahap awal proses pemurnian enzim/protein adalah ekstraksi enzim dari organisme sumbernya. Secara umum metode ekstraksi yang dipilih bukan hanya bergantung pada tipe sel dan lokasi enzim didalam sel atau dikeluarkan dari sel, namun juga pada karakteristik enzim yang akan diekstrak. Pada tahap ini pemecahan sel biasanya dilakukan secara mekanik seperti penggerusan, disrupsi sel menggunakan manik-manik kaca atau sonikasi (Palmer 1981). Tahap
11 berikutnya adalah proses klarifikasi dan pengendapan protein enzim yang diinginkan dari larutan. Klarifikasi dilakukan untuk memisahkan enzim dari partikel sel debris dan partikel substrat. Pengendapan protein enzim bertujuan untuk meningkatkan konsentrasi enzim, mengurangi volume larutan dan memisahkan enzim target dari protein pengotor yang lain.
Prosedur pemurnian enzim lanjutan adalah menggunakan teknik kromatografi kolom. Prinsip kromatografi kolom yaitu pengaliran suatu cairan melalui kolom yang mengandung bahan pengisi dan substanta yang ingin dipisahkan menjadi beberapa komponen dengan adanya perbedaan terhadap daya ikat bahan pengisi. Berdasarkan daya kerja partikel pengisi terhadap substanta yang ingin dipisahkan, kromatografi dapat digolongkan ke dalam jenis absorbsi, pertukaran ion, filtrasi gel dan interaksi biokimia (Suhartono 1989).
2.5.1. Kromatografi Filtrasi Gel.
Kromatografi filtrasi gel merupakan metode pemisahan dalam pemurnian protein, peptida dan oligonukleotida berdasarkan pada ukuran partikel. Molekul-molekul tersebut masuk melalui pori-pori gel matrik dan terdifusi dalam matrik berdasarkan besar kecilnya ukuran molekul. Molekul dengan ukuran kecil terdifusi lebih jauh dalam pori-pori gel oleh karenanya terelusi lebih lambat. Molekul yang berukuran besar tidak masuk ke dalam pori-pori permukaan gel sehingga terelusi lebih cepat. Teknik kromatografi filtrasi gel ini dapat digunakan untuk penentuan bobot molekul, pemisahan komponen-komponen dalam suatu campuran, atau untuk menghilangkan komponen garam dari suatu larutan (desalting).
Beberapa jenis matrik yang digunakan dalam filtrasi gel adalah dekstran, akrilamid, agarosa dan polistiren. Sephadex (gel dekstran) bersifat tahan terhadap garam atau basa pada konsentrasi tinggi, tetapi akan rusak oleh asam kuat ( dibawah pH 2) dan oksidator kuat. Contoh jenis Sephadex adalah Sephadex G-25, Sephadex G-50, Sepadex G-75 dan Sephadex G-100. Huruf G menunjukkan bahwa Sephadex tersebut dikembangkan dengan air sedangkan nomor dibelakangnya menunjukkan pengembangan tersebut, misalnya 25 kali, 50 kali , 75 kali dan seterusnya (Suhartono 1989).
12 2.5.2. Kromatografi Pertukaran Ion.
Kromatografi pertukaran ion adalah metode pemisahan senyawa-senyawa pada matriks yang tidak larut dan mengandung ion-ion labil yang mampu saling bertukar posisi dengan ion-ion medium sekitarnya. Bahan penukar ion dapat bersifat anion atau kation bergantung sifat afinitasnya terhadap ion positif atau negatif (Nur et al. 1989).
Teknik kromatografi ini memerlukan fase diam yang biasanya merupakan polimer terhidratasi yang bersifat tidak larut seperti selulosa, dextran dan agarosa. Gugus ion diimobilisasikan didalam matriks. Beberapa gugus penukar anion adalah aminoetil (AE-), kuaternari aminoetil (QAE-) dan dietilaminoetil (DEAE-), sedangkan gugus penukar kation antara lain sulfopropil (SP-), metil sulfonat dan karboksimetil (CM-) (Coligan et al. 2003).
Dasar dari kromatografi pertukaran ion adalah ion bermuatan dapat bebas dipertukarkan dengan ion yang memiliki tipe muatan sama. Protein yang memiliki gugus bermuatan negatif dapat dipertukarkan dengan ion klorida. Mula-mula gugus fungsional matriks yang bermuatan negatif mengikat ion dari bufer (misalnya Na+), pada saat sampel dimasukkan ke dalam kolom, maka
protein yang bermuatan positif akan menggantikan ion Na+ sedangkan protein yang bermuatan negatif atau netral tidak terikat. Protein yang tidak terikat akan dibilas dengan menggunakan bufer. Protein yang terikat pada matriks kolom akan dilepaskan dengan cara membilas kolom menggunakan bufer yang mengandung NaCl atau KCl. Pembilasan dilakukan dengan meningkatkan konsentrasi NaCl atau KCl secara bertahap sehingga protein yang memiliki ikatan lemah dengan matriks akan lepas terlebih dulu dan diikuti oleh protein yang memiliki ikatan yang lebih kuat.
2.5.3. Kromatografi Afinitas.
Kromatografi afinitas merupakan suatu metode pemisahan protein/enzim berdasarkan interaksi yang spesifik antara pasangan senyawa yang dimurnikan seperti makromolekul enzim dengan substrat, kofaktor, allosterik, efektor atau inhibitor.
13 Prinsip kromatografi afinitas adalah adanya suatu ligan yang terikat secara kovalen pada matriks tidak larut dalam air yang akan mengikat salah satu atau beberapa campuran yang memiliki afinitas spesifik terhadap ligan tersebut. Komponen yang tidak memiliki afinitas akan melaju terus dan molekul-molekul yang terikat dapat dilepaskan dari ligan cara elusi menggunakan larutan bufer. Kromatografi interaksi hidrofobik dalam pemurnian enzim menggunakan prinsip perbedaan hidrofobisitas (polaritas) antara protein target dan matriks dalam kolom. Matriks yang bersifat non-polar (hidrofobik) akan memisahkan protein enzim yang bersifat hidrofobik dari protein atau molekul lain yang bersifat hidrofilik (polar) (Suhartono 1989). Pemurnian enzim dengan menggunakan beberepa teknik kromatografi kolom seperti yang diuraikan diatas, disajikan pada Gambar 4.
Gambar 4. Pemurnian enzim dengan berbagai teknik kromatografi kolom. (http://tainano.com/Molecular2520Biology2520Glossary.files/imag e047.gif).
14 Teknik yang sering digunakan untuk memurnikan xilanase dari mikroba adalah kromatografi kolom. Berbagai teknik pemurnian ini akan memberikan hasil dan tingkat kemurnian yang bervariasi. Beberapa peneliti telah melakukan pemurnian enzim xilanase dengan berbagai teknik pemurnian seperti yang tercantum dalam Tabel 3.
Tabel 3. Teknik pemurnian xilanase dari beberapa jenis mikroba.
Isolat Xilanase Teknik pemurnian Referensi
Melanocarpus abomyces endo-1,4-ß xilanase Kromatografi interaksi hidrofobik (HIC) Gupta et al. 2002 Staphylococcus
sp SG-13 endo-1,4-ß-xilanase Kromatografi pertukaran anion, filtrasi gel
Gupta et al. 2000
Bacillus sp
endo-1,4-ß-xilanase Kromatografi pertukaran anion, filtrasi gel
Sapre et al. 2005
Streptomyces lividans
Asetil xilan
esterase Kromatografi penukar kation Dupont et al. 1996 Aeromonas caviae W-61 endo-1,4-ß-xilanase Kromatografi pertukaran ion, filtrasi gel Dung et al. 1993
Humicola grisea endo-1,4-ß-xilanase Kromatografi pertukaran ion, interaksi hidrofobik Lucena-Neto dan Ferreira-Filho 2004 Trichoderma reesei Α
glucuronidase Kromatografi pertukaran ion, HIC dan filtrasi gel
Lappalainen et al. 2000
Streptomyces galbus NR
endo-1,4-ß-xilanase amonium sulfat Nagieb 2004 Kansoh dan Thermomyces
lanuginosus
endo-1,4-ß-xilanase
Kromatografi filtrasi gel dan pertukaran ion
Cesar dan Mrsa 1996
2.6. Kinetika Pertumbuhan Mikroba
Persamaan kinetika yang digunakan berdasarkan persamaan Luedecking-Piret (1959), dimana studi kinetika berdasarkan persamaan tersebut telah digunakan oleh Weiss dan Ollis (1980) untuk produksi kultivasi gum xanthan dan oleh Multhuvelayudham dan Viruthagiri (2007) untuk memproduksi selulase. Persamaan yang dihasilkan adalah laju penggunaan substrat (dS/dt),
15 laju pembentukan produk (dP/dt) dan laju pembentukan massa (dX/dt) sebagai berikut:
Laju Pembentukan Biomassa (Persamaan logistik).
Persamaan yang digunakan merupakan bentuk penurunan dari bentuk logistik sebagai berikut:
dx/dt = f(x)
dx/dt = µ X (1 – X/X max)...(1)
dengan µ = laju pertumbuhan spesifik dan X maks = konsentrasi biomassa maksimum. Bila persamaan (1) diintegralkan menggunakan Xo= X (t=0) akan menghasilkan kurva sigmoid X(t), baik fase eksponential maupun fase stationer, dalam hubungan sebagai berikut :
X(t) = Xo[eµt/1-(Xo/Xmaks) (1-eµt)]...(2) Laju Pembentukan Produk : Persamaan Luedecking-Piret
Pembentukan produk mengikuti persamaan:
dP/dt = nX + m (dx/dt)...(3)
dengan m adalah tetapan pembentukan produk yang berasosiasi dengan pertumbuhan atau pembentukan produk, n adalah tetapan pembentukan produk yang tidak berasosiasi dengan pertumbuhan atau pembentukan produk pada fase stasioner, P adalah konsentrasi produk (g/l) dan Po adalah produk pada saat t=0. Pada model ini n dievaluasi dari data fase stationer (dx/dt=0), m mengikuti dP/dt sepanjang fase eksponensial awal. Integrasi persamaan (3) menggunakan persamaan (2) untuk X(t), memberikan persamaan dengan dua kondisi awal (Xo,Po), kondisi akhir (Xmaks) dan tiga parameter (µ, n dan m):
P(t) = Po + mXo (eµt/1-[(Xo/Xmaks) (1-eµt)]) + n (Xmaks/µ) ln (1-
[Xo/(Xmaks(1-eµt))])...(4)
n = (dP/dt)stat ...(5) Xmaks
Laju Penggunaan Substrat : Modifikasi Luedeking-Piret
Laju penggunaan substrat merupakan fungsi dari pertumbuhan biomassa, pembentukan produk dan pemeliharaan sel. Bentuk persamaan kinetikanya diasumsikan sebagai bentuk linier dari kombinasi ketiga parameter tersebut (Weiss dan Ollis 1980).
16 dS/dt = -(1/Yx)(dX/dt) – (1/Yp)(dP/dt) – KeX
pertumbuhan produk dan pemeliharaan sel : dS/dt = -α dX/dt – βX dimana α = 1/Yx/s + m/Yp/s dan ß = n/Yp/s + Ke
dengan memasukkan nilai diatas pada persamaan kurva pertumbuhan dan pemeliharaan, maka diperoleh persamaan
dS/dt = -(1/Yx/s + m/Yp/s ) dX/dt – (n/Yp/s + Ke )X ...(6)
dengan demikian penggunaan substrat juga akan mengikuti persamaan Luedecking-Piret dengan asumsi tetap, seperti persamaan pembentukan produk (persamaan 3). Substitusi persamaan (2) pada persamaan (6) dan mengintegralkannya, akan menghasilkan :
So-S(t) = ((/Yx/s + m/Yp/s))(X-Xo) + (n/Yp/s + Ke ) Xmaks/µ (ln (1-(Xo/Xmaks)
(1-eµt)))... (7) atau
So-S(t) = α (X-Xo) + ( ß Xmaks/µ) ln (1-(Xo/Xmaks)(1-eµt)) ... (8) Dimana So adalah konsentrasi substrat awal (g/l) dan S(t) adalah konsentrasi substrat sisa pada saat t jam. Sedangkan α adalah tetapan penggunaan substrat yang berasosiasi dengan pertumbuhan biomassa dan β adalah tetapan penggunaan substrat yang tidak berasosiasi dengan pertumbuhan, tetapi berasosiasi dengan pembentukan produk dan pemeliharaan sel. Nilai ß diperoleh dari persamaan
ß = dS/dt stationer ... (9) Xmaks
2.7. Staphylococcus aureus
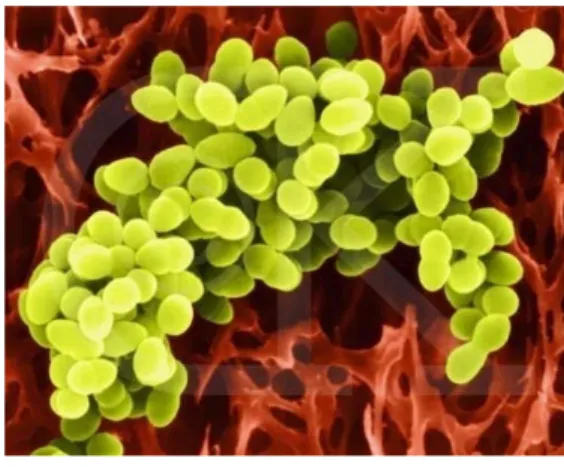
Staphylococcus aureus dapat diklasifikasikan dalam domain Bacteria, filum Firmicutes, kelas Bacilli, ordo Bacillales dan famili Staphylococcaceae. S.aureus adalah bakteri yang bersifat anaerobik fakultatif, Gram-positif, koagulase positif, katalase positif dan menghasilkan asam laktat. Sel staphylococci ini berbentuk bulat berdiameter 1µm, berwarna kuning terang dan bergerombol seperti anggur (Gambar 5). Pada tahun 1884, Rosenbach menjelaskan bahwa terdapat dua jenis koloni berwarna pada staphylococci yaitu Staphylococcus aureus (kuning) dan Staphylococcus albus (putih). S. albus ini
17 sekarang bernama S. epidermidis. Terdapat lebih dari 20 spesies Staphylococcus yang digambarkan dalam Bergey's Manual (2001), namun hanya S. aureus dan S.epidermidis yang banyak berinteraksi dengan manusia. S.aureus dapat memproduksi enzim katalase dan dapat mengubah H2O2 menjadi air dan
oksigen. Uji katalase inilah yang membedakan staphylococci dari enterococci dan streptococci (http://www.textbookofbacteriology.net/staph.html).
S. aureus pertama kali ditemukan oleh ahli bedah Sir Alexander Ogston di Aberdeen, Skotlandia pada tahun 1880 dari abses luka operasi. Bakteri ini dikenal secara umum sebagai bakteri penyebab infeksi pada luka di kulit, mulai dari infeksi ringan maupun penyebab penyakit menahun seperti meningitis, pneumonia, endocarditis dan Toxic Shock Syndrome (TSS). Pada ternak S.aureus dikenal sebagai penyebab mastitis (peradangan kelenjar susu/ambing).
Gambar 5. Koloni Staphylococcus.aureus
(http://202.114.65.51/fzjx/wsw/newindex/tuku/MYPER/zxj/z xjimage).
Berbagai komponen S.aureus yang berperan dalam mekanisme infeksi adalah : (1) Polisakarida dan protein yang merupakan substansi penting di dalam dinding sel, seperti protein adhesion hemaglutinin dan glikoprotein fibronectin. Protein ini berperan dalam proses kolonisasi bakteri pada jaringan inang; (2) Invasin yang berperan dalam penyebaran bakteri di dalam jaringan, misalnya leukocidin, kinase, hyaluronidase; (3) Kapsul yang dapat menghambat fagositosis oleh leukosit polimorfonuklear: (4) Protein A, koagulase dan koagulase yang terikat pada sel (clumping factor) untuk menghindarkan diri dari
18 respon imun inang. Protein A adalah protein permukaan yang umum terdapat pada S.aureus dan memiliki kemampuan untuk berikatan dengan bagian Fc immunoglobulin G (IgG) berbagai species mamalia dan juga pada beberapa species protein A ini dapat berikatan dengan bagian Fc IgA dan IgM (Kusunoki et al. 1992). Studi peran protein A terhadap tingkat virulensi S.aureus menunjukkan bahwa bakteri yang tidak memiliki protein A menunjukkan pengurangan virulensi terhadap infeksi subkutan dan peritoneal pada mencit, tapi tidak pada mastitis (Foster 1992, Arbuthnott et al. 1983); (5) Substansi biokimia, seperti : karotenoid dan katalase, dapat membuat bakteri bertahan hidup dalam fagosit; (6) Toksin yang dapat merusak membran sel dan jaringan inang (Todar 1998).
S.aureus juga memiliki beberapa faktor virulen lain seperti beberapa jenis enzim yang dihasilkan yaitu (1) Koagulase, yang mengkonversi fibrinogen menjadi fibrin dan mencegah aktifitas fagositosis, karena sel granulosit sulit menembus gumpalan fibrin; (2) Hyaluronidase, adalah eksoenzim yang ditemukan lebih dari 90% pada S.aureus dan mampu menghidrolisis asam hyaluronat yang merupakan substansi jaringan ikat. Hal ini akan menyebabkan infeksi meluas; (3) Lipase, enzim ini bersifat lipolitik dan tidak memerlukan substrat yang spesifik. Lipase aktif pada berbagai macam substrat termasuk plasma lemak dan minyak yang terakumulasi di permukaan tubuh. Materi ini digunakan untuk mempertahankan diri dan aktifitas paling besar terdapat di kelenjar sebasea; (4) Nuklease, enzim nuclease yang diproduksi S.aureus bersifat kuat, tahan pendidihan dan hanya aktif dengan kehadiran ion kalsium. Nuklease adalah fosfodiesterase bersifat endo dan eksonukleolitik yang memotong DNA atau RNA menjadi 3’fosfomononukleotida (Joklik et al.1980 diacu dalam Abrar 2001).