KEANEKARAGAMAN SPESIES RAYAP DAN BAKTERI
SIMBIONNYA: STUDI KASUS DI KAMPUS
IPB DRAMAGA, BOGOR
NADZIRUM MUBIN
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Keanekaragaman Spesies Rayap dan Bakteri Simbionnya: Studi Kasus di Kampus IPB Dramaga, Bogor adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2013
Nadzirum Mubin NIM A34090014
ABSTRAK
NADZIRUM MUBIN. Keanekaragaman Spesies Rayap dan Bakteri Simbionnya: Studi Kampus di Kawasan IPB Dramaga, Bogor. Dibimbing oleh IDHAM SAKTI HARAHAP dan GIYANTO.
Sebagai negara tropis, Indonesia kaya dengan keanekaragaman hayati. Keanekaragaman spesies rayap cukup tinggi di negara ini, mencakup 200 jenis spesies yang sudah teridentifikasi dibandingkan dengan 2000 spesies rayap yang ada di seluruh dunia. Rayap bersimbiosis dengan mikroorganisme pencerna selulosa, sehingga hubungan antara keanekaragaman mikrob dengan keanekaragaman spesies rayap sangat berkorelasi. Tujuan dari penelitian ini adalah menginventarisasi keanekaragaman spesies rayap dan bakteri simbionnya di Kampus IPB Dramaga. Metode yang digunakan adalah pembuatan stasiun pengamatan di tanah dan transek dengan ukuran 10 m x 10 m yang berlokasi di Kampus IPB Dramaga untuk mengoleksi rayap contoh. Mikrob simbion diekstraksi dari saluran pencernaan belakang (proktodeum) masing-masing spesies rayap dengan teknik pengenceran berseri dan pewarnaan pada media biakan. Bakteri hasil isolasi dikarakterisasi berdasarkan Gram dan morfologinya. Hasil yang diperoleh menunjukkan bahwa ditemukan 4 spesies rayap dari famili Termitidae (Macrotermes gilvus, Capritermes mohri, Odontotermes javanicus, dan Microtermes insperatus) dan 2 spesies dari famili Rhinotermitidae (Coptotermes curvignathus dan Schedorhinotermes javanicus). Bakteri simbion dari hasil ekstraksi saluran pencernaan terdiri dari 34 isolat dari famili Termitidae dan 9 isolat dari famili Rhinotermitidae berdasarkan karakter Gram dan morfologi.
ABSTRACT
NADZIRUM MUBIN. Species Diversity of Termite and Bacterial Symbiont: Cases Study in Campus of IPB Dramaga, Bogor. Supervised by IDHAM SAKTI HARAHAP and GIYANTO.
As tropical country, Indonesia is very rich with biodiversity. Termite species diversity is quite high in this country, i.e around 200 spesies compared to 2000 spesies for the whole world. Since termites must have a symbioses with microbes to digest cellulose, so spesies diversity of those microbes must be correlated with termites spesies diversity. The objective of this research was to inventory spesies diversity of termites and their bacterial symbiont in Campus IPB Dramaga. Termites were collected through observation station installed in the ground and transect 10 m x 10 m which were set in Campus IPB Dramaga area. Symbiont microbes were extracted from proctodeum of each termites spesies with serial dilution technique and staining on culture media. Bacteria kolonied and characterized by Gram and morphology. The results showed that 4 species of termites belong to family Termitidae (Macrotermes gilvus, Capritermes mohri, Odontotermes javanicus, and Microtermes insperatus) and 2 species belong to family Rhinotermitidae (Coptotermes curvignathus and Schedorhinotermes javanicus) were founds. Symbiont bacteria extracted from proctodeum of those termites consist of 34 isolates from Termitidae and 9 isolates from Rhinotermitidae based on Gram and morphological characteristic.
© Hak Cipta milik IPB, tahun 2013 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis dalam bentuk apa pun tanpa izin IPB.
KEANEKARAGAMAN SPESIES RAYAP DAN BAKTERI
SIMBIONNYA: STUDI KASUS DI KAMPUS
IPB DRAMAGA, BOGOR
NADZIRUM MUBIN
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Program Studi Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul skripsi: Keanekaragaman Spesies Rayap dan Bakteri Simbionnya: Studi Kasus di Kampus IPB Dramaga, Bogor
Nama : Nadzirum Mubin
NIM : A34090014
Disetujui oleh
Dr Ir Idham Sakti Harahap, MSi Dr Ir Giyanto, MSi
Pembimbing I Pembimbing II
Diketahui oleh
iiDr Ir Abdjad Asih Nawangsih, MSiii Ketua Departemen Proteksi Tanaman
PRAKATA
Puji dan syukur kehadirat Allah SWT yang telah memberikan karunia dan hidayah-Nya sehingga penulis dapat menyelesaikan skripsi yang berjudul “Keanekaragaman Spesies Rayap dan Bakteri Simbionnya: Studi Kasus di Kampus IPB Dramaga, Bogor”. Skripsi ini dibuat untuk memenuhi tugas akhir sebagai syarat untuk memenuhi gelar Sarjana Pertanian di Institut Pertanian Bogor, Fakultas Pertanian, Departemen Proteksi Tanaman.
Tugas akhir ini dapat terselesaikan dengan baik karena bantuan berbagai pihak. Untuk itu dengan penuh hormat, cinta, dan kasih sayang penulis mengucapkan terima kasih kepada Ayahanda H. Ahmad Romlan dan Ibunda Hj. Nau’ul Masdariyah, serta keluarga besar penulis yang telah mendoakan dan memberikan dukungan yang luar biasa kepada penulis. Bapak Idham Sakti Harahap dan Bapak Giyanto selaku dosen pembimbing skripsi yang telah banyak memberikan masukan, motivasi, dan bimbingan. Bapak Swastiko Priyambodo selaku dosen pembimbing akademik yang banyak memberikan motivasi dan bimbingan; teman-teman penulis kosan “PODJOK” yaitu Nicko Surya, Tri Setyawan, Aris Pracoyo, Tody, Samsi, dan Zendy; dan teman-teman angkatan 46 Proteksi Tanaman yang selalu membantu dan memberikan semangat sehingga dimudahkan dalam penyelesaian tugas akhir ini.
Semoga kebaikan dan perhatian yang telah diberikan memperoleh balasan yang lebih baik dari Allah SWT. Penulis juga menyadari bahwa skripsi ini masih jauh dari kesempurnaan, namun penulis berharap karya ini dapat memberikan manfaat bagi yang memerlukan, khususnya di bidang ilmu proteksi tanaman.
Bogor, Juni 2013
DAFTAR ISI
Halaman PENDAHULUAN 1 Latar Belakang 1 Tujuan 4 Manfaat 4BAHAN DAN METODE 5
Tempat dan Waktu 5
Bahan dan Alat 5
Metode Penelitian 5
Eksplorasi dan Identifikasi Rayap 5
Pengambilan Sampel Rayap 5
Pemeliharaan Koleksi Rayap 6
Identifikasi Rayap 6
Pengamatan Tipe Habitat 7
Isolasi dan Karakterisasi Bakteri Simbion dari Saluran Pencernaan
Rayap 8
Persiapan Media 8
Isolasi Bakteri Simbion 8
Karakterisasi Bakteri Simbion 9
HASIL DAN PEMBAHASAN 10
Keanekaragaman Spesies Rayap di Lapangan 10
Hubungan Spesies Rayap yang Ditemukan di Berbagai Tipe Habitat 12
Hasil Pengamatan Tanah 12
Hasil Pengamatan Vegetasi Penutup Tanah 13
Hasil Pengamatan Persentase Naungan 15
Isolasi dan Karakterisasi Bakteri Simbion dari Saluran Pencernaan Rayap 18 Isolasi Bakteri Simbion dari Saluran Pencernaan Rayap 18 Karakterisasi Bakteri Simbion dari Pencernaan Rayap 18
KESIMPULAN DAN SARAN 23
Kesimpulan 23
Saran 23
DAFTAR PUSTAKA 25
viii
DAFTAR TABEL
No Halaman
1 Lokasi pengumpanan dan transek di Kampus IPB Dramaga 6
2 Jenis dan jumlah rayap yang ditemukan dari lapangan 11 3 Hubungan spesies rayap yang ditemukan dengan karakteristik habitatnya 12 4 Tipe tanah dan pH pada lokasi pengambilan sampel rayap 13
5 Jenis-jenis vegetasi penutup tanah di lapangan 14
6 Kawasan dengan berbagai naungan 18
7 Hasil isolasi dan karakterisasi bakteri simbion dari saluran pencernaan
x
DAFTAR GAMBAR
No Halaman
1 Peletakkan kayu umpan. (a) skema pengumpanan (kayu, karton, dan
para-lon; kayu tonggak) (b) Pengumpanan di lapang 7
2 Skema transek 7
3 Persentase naungan tajuk (a) rendah 0-25%, (b) sedang 26-50%, (c) tinggi
51-75%, dan (d) sangat tinggi 76-100% 8
4 Rayap yang ditemukan dari famili Termitidae (a) M. gilvus (mayor), (b) M. gilvus (minor), (c) M. insperatus, (d) C. mohri, dan (e) O. javanicus
(perbesaran 100x) 10
5 Rayap yang ditemukan dari family Rhinotermitidae (a) S. javanicus (mayor), (b) S. javanicus (minor), dan (c) C.curvignathus (perbesaran
100x) 10
6 Jenis tanah (a) regosol (hitam) (b) latosol (coklat) 13
7 Gulma yang ditemukan di lapangan (a) A. compressus, (b) C. aridus 15 8 Hubungan persentase naungan dengan jenis rayap yang ditemukan 17 9 Hasil isolasi bakteri simbion dari protodeum (a) koloni sebelum
dimurnikan (b) koloni setelah dimurnikan 18
10 Morfologi koloni bakteri yang berbentuk bulat 19
11 Elevasi bakteri hasil ekstraksi proktodeum (a) bentuk cembung pada koloni bakteri, (b) skema cembung, (c) bentuk kawah pada koloni
bakteri, dan (d) skema kawah 20
12 Visualisasi bakteri hasil ekstraksi proktodeum (a) putih susu, (b) putih,
xii
DAFTAR LAMPIRAN
No Halaman
1 Data pengamatan dari lapangan 31
2 Jumlah rayap yang ditemukan pada persentase naungan yang berbeda 33 3 Jenis-jenis rayap dari lapangan dan deskripsinya berdasarkan kunci
PENDAHULUAN
Latar Belakang
Indonesia merupakan negara tropis yang kaya akan keanekaragaman tumbuhan yang sangat mendukung tempat hidup rayap. Sekitar 80% daratan Indonesia merupakan habitat yang baik untuk perkembangan rayap (Nandika et al. 2003). Rayap termasuk ke dalam ordo Blattodea famili Termitidae (Inward et al. 2007) yang terdiri atas 2000 spesies yang tersebar di dunia dan baru sekitar 200 spesies teridentifikasi di Indonesia sampai tahun 1970 (Tarumingkeng 1971).
Keanekaragaman spesies rayap di Pulau Jawa belum terinventarisasi seluruhnya. Pada tahun 1990-an dilakukan inventarisasi oleh beberapa peneliti yaitu Nandika et al. (1997) meneliti jenis rayap dan penyebarannya di wilayah DKI Jakarta dan Bandung. Penelitian tersebut menemukan 9 spesies rayap yaitu Microtermes insperatus, M. incertoides, Macrotermes gilvus, Odontotermes
javanicus, O. malaccensis, Schedorhinotermes javanicus, Coptotermes
curvignathus, C. haviliandi, C. kalshoveni, C. heimi, dan C. travians. Pada tahun 2003, Primanda et al. meneliti keanekaragaman spesies rayap di Kampus Universitas Indonesia, Depok dan menemukan 6 spesies yaitu M. insperatus, M. gilvus, O. javanicus, O. grandiceps, S. javanicus, dan C. curvignathus dengan persebaran yang acak.
Pertanaman karet, kelapa sawit, kakao, dan sengon di Kampus IPB Dramaga sangat berpotensi menjadi habitat rayap. Rayap menyukai semua jenis kayu karena kayu tersusun oleh selulosa yang merupakan makanan utama rayap. Herlinda et al. (2010) telah meneliti tentang serangan rayap C. curvignathus pada pertanaman karet di Sumatera Selatan dan menemukan bahwa rayap jenis ini sangat berpotensi merusak pohon-pohon karet yang sedang berproduksi sehingga dapat menurunkan produktivitas dan juga dapat menyebabkan kematian pada pohon yang diserang. Hasil penelitian pada perkebunan kelapa sawit di Sarawak, Malaysia menemukan 13 spesies rayap yaitu C. curvignathus, C. sepangensis, C. gestroi, Parrhinotermes aequalis, S. brevialatus, S. javanicus, S. sarawakensis, Pericapritermes dolchocepalus, P. latigathus, Nasutitermes haviliandi, N. langinasoides, N. matangensiformis, dan Havilanditermes atripennis (Bong et al. 2012). Rayap juga menyerang fase pembibitan kakao, sehingga sangat berisiko jika terserang pada fase tersebut (Du et al. 2009). Keng (2006) telah meneliti tentang rayap di perkebunan kakao yang dibandingkan dengan hutan primer di Taman Bukit Tawau, Sabah. Berdasarkan hasil penelitian tersebut diperoleh data bahwa rayap juga menyerang perkebunan kakao meskipun tingkat keragamannya lebih rendah jika dibandingkan dengan hutan primer. Kelimpahan rayap lebih banyak pada pertanaman kakao sehingga serangannya menjadi berat. Hal tersebut disebabkan oleh sumber makanannya yang melimpah.
Menurut Tarumingkeng (1971) identifikasi rayap lebih mudah dimulai dari tingkat famili hingga pada tingkat spesies. Rayap di kawasan tropis seperti Indonesia sudah banyak diteliti dan terdapat beberapa famili yaitu Kalotermitdae, Rhinotermitidae, dan Termitidae. Famili Kalotermitidae merupakan kelompok rayap yang menyerang dan bersarang pada pohon yang masih hidup atau bersarang pada kayu kering yang tidak berhubungan dengan tanah. Kelompok rayap jenis tersebut biasa disebut dengan rayap kayu kering (Shelton et al. 2000).
2
Spesies rayap kayu kering yang biasa menyerang di permukiman salah satunya adalah Cryptotermes cynocephalus. Spesies rayap ini akan menyerang permukiman yang bangunannya terbuat dari kayu. Famili kedua adalah Rhinotermitidae. Ciri khas dari famili ini adalah adanya keping sklerit pada toraks yang berbebntuk datar. Famili ini sering bersarang di dalam kayu atau bahan lain yang mengandung selulosa yang terdapat pada permukaan tanah. Jenis rayap dari famili ini yang sering menyerang permukiman dan perkebunan adalah dari genus Schedorhinotermes dan Coptotermes. Genus Coptotermes mempunyai ciri khas yaitu pada saat koloninya merasa terganggu maka prajuritnya akan mengeluarkan cairan seperti susu yang fungsinya untuk melumpuhkan musuh. Cairan ini keluar dari fontanel yang ada di depan kepala (Bong et al. 2012). Famili ketiga adalah Termitidae, merupakan kelompok rayap dengan jumlah spesies yang paling banyak. Ciri dari famili Termitidae adalah adanya keping sklerit pada pronotum berbentuk seperti pelana; pusat sarangnya berada di dalam tanah dan membuat galeri cendawan berbentuk bunga karang; dan biasanya membuat gundukan-gundukan tanah. Contoh genus yang dikenal banyak menyerang perkebunan adalah Macrotermes, Odontotermes, dan Microtermes.
Rayap dan semut merupakan kelompok serangga sosial. Keduanya mempunyai karakterisitik yang hampir sama. Masyarakat sering menyebutkan bahwa rayap adalah semut putih atau white ant, meskipun antara rayap dan semut tidak mempunyai hubungan kekerabatan. Berdasarkan taksonominya, rayap merupakan ordo dari Blattodea (Inward et al. 2007) sedangkan semut merupakan kelompok dari ordo Hymenoptera. Selain itu dari sisi morfologi semut mempunyai penggentingan antara torak dan abdomen yang merupakan ciri khas dari kelompok tabuhan (Hymenoptera: Apocrita). Penggentingan itu disebut petiol sedangkan pada rayap tidak ada penggentingan bahkan sering tidak terlihat jelas batas antara torak dan abdomen. Umumnya rayap mencari makan pada wilayah yang tertutup (kriptik) dicirikan dengan tabung kembara yang berasal dari tanah membentuk saluran-saluran yang menghubungkan antara tabung kembara satu dengan yang lainnya, sedangkan pada semut cenderung mencari makan pada wilayah yang terbuka. Makanan rayap berupa lignoselulosa yang terdiri atas selulosa, lignin, dan hemiselulosa. Selulosa tersusun atas polimer glukosa yang kaya akan serat sedangkan semut memakan bahan organik yang mengandung gula-gulaan yang tersusun atas polisakararida.
Serangga sosial mempunyai perbedaan tugas dalam suatu koloninya atau biasa disebut dengan kasta. Rayap mempunyai sistem pembagian kasta, yaitu kasta reproduktif, prajurit, dan pekerja. Kasta reproduktif bertugas untuk kawin dan bertelur yang terdiri atas raja dan ratu sebagai reproduktif primer. Kasta reproduktif terbagi menjadi dua yaitu primer dan neoten (sekunder). Kasta reproduktif primer berasal dari rayap bersayap (alate) atau laron, sedangkan reproduksi neoten muncul ketika ratu atau raja mati atau hilang dalam koloni sehingga kasta reproduksi neoten ini akan muncul (Nandika et al. 2003). Kasta prajurit mempunyai ciri khas tidak kawin, mata tereduksi, dan tugasnya hanya mempertahankan atau melindungi koloni jika terjadi serangan dari musuh. Kasta ini dicirikan dengan berkembangnya mandibel yang maju ke depan yang biasa digunakan untuk menyerang musuh yang menyerang koloninya. Kelompok rayap Nasutitermitinae merupakan kelompok rayap yang mempunyai keunikan dibandingkan dengan kelompok rayap yang lain. Kasta prajuritnya mempunyai
3 mandibel yang tidak berkembang dengan baik, tetapi fontanel (dahi) dari kepala yang lebih berkembang. Fungsi fontanel ini hampir sama sebagai pertahanan, yaitu dapat mengeluarkan senyawa kimia yang dapat melumpuhkan musuh.
Kasta pekerja merupakan kasta yang terbanyak pada suatu koloni yaitu sekitar 80%. Kasta pekerja tidak mempunyai mata dan mandul. Tugasnya hanya mencari dan memberikan makan ke ratu dan prajurit. Kasta pekerja mampu mencerna kayu yang mengandung banyak selulosa. Bantuan mikrob di dalam tubuh rayap memudahkan dalam proses pencernaan. Mikrob yang membantu dalam proses pencernaan mengeluarkan enzim selulolitik untuk memudahkan dalam mendegradasi selulosa dari kayu.
Menurut Klepzig et al. (2009) tipe-tipe hubungan antara serangga dan mikroorganisme simbionnya sangat luas. Hubungan ini mempunyai variasi yang banyak seperti mutualisme, komensalisme, atau parasitisme. Rayap merupakan salah satu contoh yang mempunyai simbiosis mutualisme antara rayap dengan organsime simbionnya. Rayap mempunyai hubungan yang spesifik terhadap mikroorganisme simbionnya seperti bakteri, cendawan, dan protozoa. Hubungan ini mempunyai manfaat baik secara langsung maupun tidak langsung. Hubungan simbiosis pada rayap ini dapat bersifat membantu dalam pencernaan makanan, penyerapan nutrisi, maupun dalam melindungi dari musuh alaminya. Menurut Sapp (1994) simbiosis merupakan kehidupan bersama pada organisme yang berbeda.
Rayap terdiri dari kumpulan spesies yang beragam, secara garis besar dibagi menjadi dua yaitu rayap tingkat rendah dan rayap tingkat tinggi. Rayap tingkat rendah bersimbiosis dengan sebagian besar populasi prokariot dan protista (eukariot bersel tunggal). Rayap tingkat tinggi hanya terdiri dari famili Termitidae akan tetapi jenisnya lebih dari tiga perempat dari semua jenis spesies yang ada dan bersimbiosis dengan sebagian besar kelompok bakteri. Asosiasi dari protista selulolitik pada pencernaan rayap tingkat rendah diketahui sebagai contoh dari simbiosis mutualisme. Protista menghasilkan asetat dari partikel selulosa atau endositosis kayu, hasil asetat tersebut diserap oleh rayap sebagai energi dan sumber karbon (Upadhyaya et al. 2012).
Beberapa spesies rayap yang bersimbiosis dengan bakteri misalnya pada rayap tingkat tinggi yaitu spesies Nasutitermes lujae (Famili Termitidae). Spesies rayap ini ditemukan bakteri anaerobik dengan karakteristiknya antara lain bakteri termasuk kelompok gram positif, bergerak dengan flagel, dan berbentuk oval. Permukaan koloni bakteri tumbuh dengan optimal pada suhu 37 0C dan pH 7.5 merupakan karakteristik dari bakteri Clostridium termitidis yang ditemukan pada N. lujae (Hethener et al. 1992). Spesies tingkat rendah seperti Coptotermes formosanus (Famili Rhinotermoitidae), ditemukan lima jenis tipe morfologi (tiga jenis koloni bakteri asam laktat dan yang lainnya dari genus Dysgonomonas) (Husseneder et al. 2009). Penelitian Adams et al. (2005) pada C. formosanus ditemukan bakteri Serratia marcescens, Enterobacter aerogens, E. cloacae, dan Citrobacter farmeri. Menurut Upadhyaya et al. (2012) beberapa strain yang diisolasi dari beberapa rayap tingkat rendah pada tiga media yang berbeda diperoleh bakteri Citrobacter, Enterobacter, dan Cellulomonas.4Hal ini menunjukkan bahwa terdapat kelompok bakteri yang berperan dalam proktodeum pada rayap tingkat rendah. Cho et al. (2010) meneliti proktodeum pada rayap
4
Reticulitermes speratus (Rhinotermitidae) dan menemukan bakteri yang toleran terhadap oksigen yaitu Dyella sp., Chryseobacterium sp., dan Bacillus sp..
Rayap tingkat tinggi dan tingkat rendah mempunyai hubungan yang hampir sama yaitu bersimbiosis dengan bakteri. Menurut Nandika et al. (2003) kontribusi bakteri dibedakan atas dasar tingkatan rayap. Pada rayap tingkat tinggi dan tingkat rendah berhubungan terhadap perilakunya dalam mencari makan maupun perilaku lainnya. Penelitian sebelumnya masih menguraikan simbion bakteri yang sebagian besar bersimbiosis pada rayap tingkat tinggi dan protozoa pada rayap tingkat rendah. Penelitian ini bertujuan untuk dapat mengetahui hubungan antara bakteri pada rayap tingkat tinggi (Termitidae) dan pada rayap tingkat rendah (Rhinotermitidae dan Kalotermitidae).
Tujuan
Penelitian ini bertujuan mengeksplorasi dan menginventarisasi keanekaragaman spesies rayap dan bakteri simbionnya di kampus IPB Dramaga, Bogor.
Manfaat
Penelitian ini diharapkan dapat digunakan sebagai sumber informasi tentang keanekaragaman spesies rayap dan bakteri simbionnya yang ada kampus IPB Dramaga, Bogor.
5
BAHAN DAN METODE
Tempat dan Waktu
Eksplorasi rayap dilakukan di kampus IPB Dramaga Bogor dengan membuat stasiun pengamatan dan transek. Selanjutnya proses identifikasi rayap dilakukan di Laboratorium Biosistematika Serangga. Isolasi dan karakterisasi bakteri dari hasil ekstraksi saluran pencernaan rayap dilakukan di Laboratorium Bakteriologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian ini dilaksanakan pada bulan September 2012 sampai Maret 2013.
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini yaitu kayu pinus sebagai umpan utama, kertas karton sebagai umpan sekunder, pipa paralon, rayap (kasta prajurit dan pekerja), alkohol 70%, media NA (Nutrient Agar) sebagai media tumbuh bakteri.
Alat-alat yang digunakan dalam penelitian ini adalah kuas, wadah plastik, mikroskop stereo, GPSmap 60CSx, mikroskop cahaya, pinset, mikropistil, mikropipet, laminar flow, vortex, shaker, autoklaf, tabung reaksi, dan tabung eppendorf.
Metode Penelitian
Eksplorasi dan Identifikasi Rayap Pengambilan Sampel Rayap
Pembuatan stasiun pengamatan. Stasiun pengamatan atau lokasi tempat
pengamatan rayap dibuat untuk memudahkan dalam pengamatan dan pengambilan rayap jika dibutuhkan pada saat ekstraksi bakteri. Pembuatan stasiun rayap dilakukan dengan menentukan lokasi terlebih dahulu (Tabel 1) kemudian disiapkan kayu sebagai umpan untuk rayap. Kayu yang digunakan yaitu kayu pinus karena lebih disukai oleh rayap (Mahmudi 2008). Kayu-kayu disiapkan dan dibentuk menjadi berbentuk ajir atau tonggak kayu berukuran 2 x 2 x 40 cm yang ditancapkan pada lokasi yang sudah dijadikan target lokasi stasiun pengamatan. Kemudian kayu dengan ukuran 2 x 2 x 10 cm digunakan untuk umpan yang ada di dalam paralon. Disiapkan juga kertas karton sebagai umpan sekunder agar kayu tidak habis termakan. Lokasi yang dijadikan target pemasangan stasiun pengamatan digali dan dipasangi paralon yang berisi kayu dan karton. Pengumpanan ditunggu selama dua minggu sebelum dilakukan tahap berikutnya (isolasi bakteri sombion).
Transek rayap. Transek dilakukan di lokasi-lokasi yang sama dengan
lokasi pengumpanan rayap (stasiun pengumpanan) (Tabel 1). Transek dibuat dengan luasan transek 100 m2 atau 10 x 10 m (Gambar 2). Pada luasan 100 m2
tersebut dilakuan pencarian rayap secara manual yaitu dengan mengambil seluruh jenis rayap yang ditemukan kemudian disimpan untuk koleksi.
6
Pemeliharaan Koleksi Rayap
Rayap yang sudah diperoleh dari pengumpanan dan transek, kemudian dimasukkan ke dalam botol dan diberi label pembeda yaitu tanggal dan lokasi pengambilan. Hal tersebut dimaksudkan untuk dapat memudahkan perlakuan lanjutan. Kasta pekerja disimpan dalam keadaan hidup atau segar agar mikrob simbionnya tetap hidup. Pada kasta pekerja ini akan dilakukan proses lanjutan yaitu ekstraksi saluran pencernaan untuk mengisolasi bakteri simbionnya.
Kasta prajurit yang diperoleh dari lapangan langsung dimasukkan ke dalam botol kaca berisi alkohol 70%. Menurut Sceffrahn (1994) biasanya hanya bagian sayap dan mandibel rayap pada saat spesimen kering yang mempunyai kondisi yang tidak berubah. Kasta prajurit ini yang akan digunakan dalam identifikasi rayap.
Tabel 1 Lokasi pengumpanan dan transek di Kampus IPB Dramaga
Lokasi Jumlah
umpan
Jumlah transek
Koordinat
Taman Proteksi Tanaman 2 2 S: 06
033.561’
E: 106043.766’
Arboretrum ARL 3 3 S: 06
033.654’
E: 106043.726’
Kebun karet Rektorat 10 5 S: 06
033.640’
E: 106043.476’
Kebun sengon Rektorat 4 4 S: 06
033.601’
E: 106043.431’ Halaman kandang FAPET
(Fakultas Peternakan) 3 3 S: 06033.328’ E: 106043.057’ Halaman Belakang FKH 3 3 S: 06 033.483’ E: 106043.183’ Perumahan dosen 3 3 S: 06 033.229’ E: 106043.428’
Kebun kakao Cikabayan 7 3 S: 06
033.140’
E: 106043.060’
Kebun kelapa sawit Cikabayan 9 5 S: 06
033.119’
E: 106043.997’
Kebun Masjid Al Hurriyah 3 3 S: 06
033.343’
E: 106043.496’
Kebun karet FPIK 3 5 S: 06
033.443’ E: 106043.579’ Agrimart 1 1 S: 06 033.566’ E: 106043.877’ Identifikasi Rayap
Identifikasi dilakukan pada kasta prajurit dan kasta reproduktif (laron). Kasta prajurit mempunyai karakteristik mandibel yang berbeda dengan kasta
7 prajurit dari spesies yang lainnya begitu juga dengan kasta reproduksi (laron). Laron juga mempunyai penanda khusus pada venasi sayapnya.
Menurut Kadarsah (2005) langkah awal yang dilakukan adalah identifikasi secara umum untuk mengetahui jenis-jenis rayap yaitu berdasarkan (1) ukuran tubuh rayap kasta prajurit, pekerja, dan reproduktif (laron) dan (2) bentuk mandibel pada kasta prajurit. Kemudian rayap hasil koleksi dari lapang diidentifikasi dari tingkat famili sampai tingkat spesies. Identifikasi dilakukan menggunakan kunci identifikasi Tarumingkeng (1971), kunci identifikasi Ahmad (1959), dan Tho (1992).
Gambar 1 Peletakan kayu umpan (a) skema pengumpanan (kayu, karton, dan paralon; kayu tonggak) (b) pengumpanan di lapang
Gambar 2 Skema transek
Pengamatan Tipe Habitat
Parameter tipe habitat yang diamati yaitu tipe tanah, tipe vegetasi penutup tanah, dan persentase naungan. Tipe tanah diamati dengan cara mengukur tingkat keasaman dan jenis tanahnya. Tipe vegetasi penutup tanah diamati dominansi dari vegetasi penutup tanah di lokasi pengamatan rayap. Sedangkan tingkat persentase naungan diamati dengan cara mengamati tajuk pohon yang menutupi lokasi pengamatan rayap dengan memberikan persen penutupan tajuk dengan persentase
b a
8
rendah (0-25%), sedang (26-50%), tinggi (51-75%), dan sangat tinggi (76-100%) (Gambar 3).
Gambar 3 Persentase penutupan tajuk (a) rendah 0-25%, (b) sedang 26-50%, (c) tinggi 51-75%, dan (d) sangat tinggi 76-100%
Isolasi dan Karakterisasi Bakteri Simbion dari Saluran Pencernaan Rayap Persiapan Media
Pembuatan media NA. NA (Nutrient Agar) merupakan media khusus
untuk pertumbuhan bakteri, namun tidak spesifik pada bakteri tertentu. Bahan-bahan yang digunakan untuk pembuatan media NA yaitu agar 50 g, peptone 3 g, beef extract 5 g, dan air akuades 1 L. Kemudian dimasukkan ke dalam erlenmenyer, ditutup menggunakan alumunium foil. Setelah itu media NA disterilisasi menggunakan autoklaf dengan suhu 120 0C pada tekanan 1 atm
selama 20 menit.
Isolasi Bakteri Simbion dari Saluran Pencernaan Rayap
Isolasi bakteri simbion pada saluran pencernaan dilakukan pada rayap pekerja. Rayap pekerja diambil dari lapangan sebanyak 3 ekor. Rayap pekerja yang sudah diambil dari lapangan dicuci dengan air steril. Setelah itu rayap disterilisasi permukaan tubuhnya dengan alkohol 70% dan diulang sebanyak 3 kali. Proses sterilisasi permukaan tubuh rayap berfungsi untuk mematikan atau meminimalkan mikrob kontaminan yang ada di permukaan tubuhnya. Setelah disterilisasi permukaan, saluran pencernaan bagian belakang (proktodeum) rayap diambil dengan cara dikelupas bagian kutikula abdomennya. Menurut Bignell (2000) di bagian proktodeum terdapat banyak aktivitas dari mikrob (bakteri dan protozoa) untuk mencerna selulosa.
9 Saluran pencernaan belakang yang sudah diperoleh kemudian digerus di dalam tabung eppendorf menggunakan mikropistil dan ditambahkan air steril 1 mL. Kemudian sebanyak 1 mL dari tabung eppendorf diambil dan ditambahkan ke dalam tabung reaksi yang berisi air steril 9 mL. Dilakukan pengenceran berseri yaitu 10-1, 10-2, 10-3, 10-4, dan 10-5. Diambil sebanyak 0.5 mL dari tabung reaksi untuk dilakukan platting ke dalam media NA. Hasil platting diinkubasikan pada suhu 25 0C selama 24 sampai 48 jam sebelum dilanjutkan pada tahap karakterisasi berdasarkan Gram, bentuk, pinggiran, warna, dan elevasi.
Karakterisasi Bakteri Simbion pada Saluran Pencernaan Rayap
Pengamatan jenis bakteri dengan menguji jenis gram. Pengujian jenis Gram merupakan langkah awal untuk memudahkan dapat pengklasifikasian sebelum proses identifikasi lebih lanjut. Pengujian Gram dapat dilakukan dengan dua cara yaitu dengan menggunakan metode pewarnaan Gram dan KOH 3%.
Uji menggunakan Pewarnaan Gram. Pengujian yang pertama
menggunakan metode teknik pewarnaan Gram atau Staining Gram dengan menggunakan pewarna secara bertahap. Teknik ini ditemukan oleh HCJ Gram pada tahun 1884. Teknik ini menggunakan crystal violet sebagai pewarna primer, iodine sebagai pengikat, alkohol 95% sebagai pendegradasi warna, dan safranin sebagai pewarna tandingan.
Sebanyak satu tetes air steril diteteskan di atas kaca preparat, kemudian diambil sebanyak satu inokulum dari biakan bakteri dan diaduk sampai merata dan dikeringanginkan. Setelah kering, kemudian diteteskan crystal violet dan ditunggu 1 menit dan dibilas dengan akuades, kemudian dikeringanginkan kembali. Jika sudah kering diteteskan larutan iodine dan ditunggu selama 2 menit dan dibilas dengan akuades dan dikeringanginkan kembali. Setelah kering, langkah selanjutnya adalah dicuci dengan etanol 95%, dibilas dengan akuades, dan dikeringanginkan kembali. Jika sudah kering diteteskan pewarna tandingan (safranin) dan ditunggu selama 10 detik, dibilas dengan akuades dan dikeringanginkan kembali. Setelah kering, koloni bakteri hasil pewarnaan dapat dilihat di bawah mikroskop.
Uji menggunakan KOH 3%. Metode ini merupakan metode konfirmasi
dari teknik pengujian menggunakan Staining Gram (Power 1995). Pengujian ini dilakukan dengan cara meteteskan larutan KOH 3% di atas kaca preparat, kemudian diambil satu inokulum bakteri murni menggunakan jarum inokulum dan dicampurkan secara merata dengan larutan KOH 3%.
Indikator kelompok bakteri Gram positif pada pengujian dengan KOH 3% yaitu koloni yang dioleskan pada cairan KOH terlihat akan memencar, sedangkan pada Gram negatif koloni bakteri yang diuji akan berlendir.
Karakterisasi bakteris simbion selanjutnya yaitu dari hasil ekstraksi yang telah tumbuh kemudian dilakukan isolasi. Isolasi dilakukan berdasarkan karakter morfologi yang berbeda (bentuk, pinggiran, elevasi, dan warna).
HASIL DAN PEMBAHASAN
Keanekaragaman Spesies Rayap di Lapangan
Berdasarkan hasil idetifikasi rayap yang ditemukan dari lapangan diperoleh 6 spesies rayap dari kawasan permukiman maupun perkebunan yaitu 4 spesies dari famili Termitidae yaitu M. gilvus, M. insperatus, C. mohri, dan O. javanicus; dan 2 spesies dari famili Rhinotermitidae yaitu S. javanicus dan C. curvignathus (Gambar 4 dan 5)
Gambar 4 Rayap yang ditemukan dari famili Termitidae (a) M. gilvus (mayor), (b) M. gilvus (minor), (c) M. insperatus, (d) C. mohri, dan (e) O. javanicus (perbesaran 100 kali)
Gambar 5 Rayap yang ditemukan dari famili Rhinotermitidae (a) S. javanicus (mayor), (b) S. javanicus (minor), dan (c) C.curvignathus
(perbesaran 100 kali)
Spesies rayap dari famili Termitidae lebih mendominasi jika dibandingkan dengan jenis rayap dari famili Rhinotermitidae yaitu 4 spesies berbanding 2 spesies. Rayap yang diperoleh dari lapangan dari famili Termitidae lebih banyak yaitu berjumlah 46 (kumulatif 4 spesies) atau 79.31% sedangkan rayap dari famili Rhinotermitidae memiliki jumlah yang lebih sedikit saat ditemukan di lapang yaitu 12 (kumulatif dari 2 spesies) atau 20.69% (Tabel 2). Famili Termitidae merupakan kelompok rayap tingkat tinggi yang mempunyai jumlah 3/4 dari jumlah seluruh rayap (Nandika et al. 2003). Sedangkan pada rayap tingkat rendah
a b c
d e
11 memiliki famili lebih banyak (Rhinotermitidae, Serritermitidae, Kalotermtidae, Archotermopsidae, Hodotermtidae, Stolotermitidae, Kalotermitidae, dan Mastotermitidae) namun jumlahnya di lapangan lebih sedikit jika dibandingkan dengan kelompok rayap tingkat tinggi.
Tarumingkeng (1971) mendeskripsikan jenis rayap berdasarkan lokasi bersarangnya, famili Termitidae merupakan jenis rayap yang bersarang di dalam tanah terutama berdekatan dengan bahan organik yang mengandung selulosa. Contoh rayap dari famili Termitidae yang umum menyerang bangunan adalah M. gilvus, M. insperatus, dan O. javanicus. Jenis-jenis rayap ini mampu menyerang objek yang berjarak hingga 200 m dari sarangnya. Famili Rhinotermitidae merupakan jenis rayap subteranean yang umumnya hidup di dalam tanah yang banyak mengandung bahan organik yang telah mati atau membusuk, kayu yang telah mati maupun yang masih hidup. Jenis rayap subteranean yang banyak merusak bangunan adalah jenis C. curvignathus dan S. javanicus. Perilaku rayap ini hampir mirip dengan rayap tanah M. gilvus, tetapi berbeda dalam kemampuan untuk bersarang di dalam kayu yang diserangnya.
Tabel 2 Jenis dan jumlah rayap yang ditemukan dari lapangan
Famili Jumlah Termitidae M. gilvus 12 M. insperatus 15 C. mohri 5 O. javanicus 14 Rhinotermtidae S. javanicus 7 C. curvignathus 5 total 58
Jenis rayap yang ditemukan di lapangan merupakan jenis-jenis rayap yang berpotensi merusak bangunan kecuali rayap jenis C. mohri yang merupakan rayap tanah yang potensi merusaknya kecil. Rayap tanah C. mohri merupakan rayap pemakan tanah yang sangat berpengaruh terhadap habitat. Tingkat gangguan dan kekayaan spesies rayap berkorelasi negatif terhadap kelimpahan dan kekayaan spesies rayap. Hasil penelitian Pribadi et al. (2011) tidak menemukan rayap pemakan tanah pada kawasan permukiman di lereng gunung Selamet, Jawa Tengah. Tidak ditemukan rayap pemakan tanah di area tersebut disebabkan oleh tingkat dispersal rayap yang rendah, sehingga kemampuannya untuk mengkolonisasi habitat sekitarnya tidak luas. Rayap pemakan tanah sangat sensitif dengan kawasan yang terganggu. Kondisi habitat yang ideal untuk jenis rayap pemakan tanah adalah hutan tropis yang rapat penutupan tajuknya (Eggleton et al. 2002).
Berbeda dengan spesies C. mohri, rayap spesies M. gilvus mempunyai potensi merusak bangunan yang tinggi. Menurut Tarumingkeng (1971) M. gilvus adalah spesies yang banyak ditemukan di permukiman. M. gilvus mempunyai potensi merusak yang tinggi. Spesies rayap M. gilvus juga banyak ditemukan di
12
permukiman dengan tingkat gangguan yang tinggi dan termasuk ke dalam kawasan yang terbuka.
Hubungan Antara Spesies Rayap yang Ditemukan di Beberapa Tipe Habitat
Setiap spesies rayap mempunyai perilaku yang berbeda-beda. Aktivitas rayap sangat tergantung pada habitat yang ditempatinya karena perilaku dasar rayap dalam mencari makan untuk koloninya. Selain itu, keberadaan rayap juga dipengaruhi oleh beberapa hal yaitu tipe tanah, tipe vegetasi, dan persentase tajuk pohon sebagai naungan.
Tabel 3 Hubungan antara spesies rayap yang ditemukan dengan karakteristik habitatnya
Jenis rayap pH tanah Jenis Tanah Vegetasi penutup Persentase naungan Termitidae M. gilvus 4, 5, 6 1 1, 3, 4, 5 1, 2, 3, 4 M. insperatus 4, 5, 6 1 1, 2, 3, 4, 5, 6 1, 2, 3, 4 O. javanicus 4, 5, 6 1, 2 1, 2, 3, 4, 5, 7 1, 2, 3, 4 C. mohri 4, 5 1 1, 4, 5 2, 3, 4 Rhinotermitidae S. javanicus 5 1 1, 4, 5 1, 2, 3, 4 C. curvignathus 5 1 1, 4, 5 1, 3, 4
Keterangan: Jenis tanah (1: Latosol, 2: Regosol), vegetasi penutup tanah (1: A. compressus, 2: A.
gangetica, 3: S. plicata, 4: C. lappacea, 5: C. aridus, 6: D. adscendens, 7: Pannicum sp.), dan
persentase naungan (1: 0-25%, 2: 25-50%, 3: 51-75%, dan 4: 76-100%)
Hasil Pengamatan Tanah
Kampus IPB Dramaga dengan luas + 270 ha mempunyai jenis tanah latosol dan regosol (Gambar 6). Tanah latosol dicirikan dengan tanah yang banyak mengandung zat besi dan aluminium. Tanah ini berwarna merah hingga kuning, sehingga sering disebut tanah merah. Tumbuhan yang dapat hidup di tanah latosol antara lain seperti padi, palawija, sayuran, buah-buahan, karet, cengkih, cokelat, kopi, dan kelapa sawit. Sedangkan tanah regosol adalah tanah berbutir kasar dan berasal dari material gunung api. Tanah regosol berupa tanah aluvial yang baru diendapkan. Material jenis tanah ini berupa abu vulkan dan pasir vulkan. Tanah regosol sangat cocok ditanami padi, tebu, palawija, tembakau, dan sayuran. Tanah yang dijadikan tempat pengambilan contoh diuji tingkat keasamannya, rata-rata pH tanahnya di bawah 7 (Tabel 4).
Hubungan spesies rayap dengan pengamatan parameter tanah yaitu pada semua jenis rayap ditemukan pada pH 5 dan tipe tanah latosol (Tabel 3). Famili Termitidae sebagian besar ditemukan pada tingkat keasaman tanah 4, 5, dan 6. Hanya pada jenis C. mohri yang tidak ditemukan pada pH tanah 6. Famili Rhinotermitidae hanya ditemukan pada tingkat keasaman 5 dengan jenis tanah latosol. Pada tanah regosol hanya ditemukan jenis rayap O. javanicus sedangkan jenis rayap yang lain tidak ditemukan pada jenis tanah tersebut.
13 Tanah bagi rayap merupakan tempat hidup dan dapat melindungi rayap dari suhu dan kelembaban yang sangat ekstrim (Nandika et al. 2003). Keberadaan dari rayap tersebut dapat membantu meningkatkan kesuburan tanah karena kemampuan dalam mencerna bahan organik menjadi hara yang dibutuhkan oleh makluk hidup lain seperti tumbuhan. Umumnya rayap menyukai tipe tanah yang mengandung liat seperti tanah latosol karena memiliki kandungan bahan organik yang tinggi. Kawasan IPB yang menjadi lokasi pengambilan contoh sebagian besar bertipe tanah latosol dengan tingkat keasaman 4 sampai 6. Hanya dua lokasi yang bertipe tanah regosol yaitu Agrimart dan Masjid Al-Hurriyyah dengan tingkat keasaman mencapai 6 (Tabel 4).
Tabel 4 Lokasi, pH, dan jenis tanah di lapangan
No Lokasi Tanah
pH Jenis tanah
1 Agrimart, Kebun Masjid Al-Hurriyyah 6 Regosol
(hitam) 2 Perumahan Dosen, halaman kandang FAPET,
Halaman belakang FKH
4 Latosol
(coklat merah) 3 Kebun kakao Cikabayan, kebun kelapa sawit
Cikabayan, kebun karet Rektorat, kebun karet FPIK, arboretrum ARL
5 Latosol
(coklat merah) 4 Kebun sengon Rektorat, gladiator, halaman
parkiran FEMA, halaman perpustakaan LSI
6 Latosol
(coklat merah)
Gambar 6 Jenis tanah (a) regosol (hitam) (b) latosol (coklat)
Hasil Pengamatan Vegetasi Penutup Tanah
Vegetasi penutup tanah berpengaruh terhadap tipe tanah sehingga keanekaragaman jenis rayap yang ada juga akan berpengaruh. Semakin banyak vegetasi penutup tanah menjadi indikator bahwa tanah tersebut subur dan kaya akan bahan organik. Rayap tanah dan subterranean lebih menyukai kondisi habitat yang ideal dengan suhu dan kelembaban stabil jika dibandingkan dengan kondisi lahan terbuka dengan iklim mikro ekstrim yang akan mempengaruhi mikrohabitat
14
di sekitarnya. Vegetasi penutup tanah mempengaruhi perilaku rayap. Adanya vegetasi penutup tanah akan membuat iklim mikro yang lebih stabil di atas tanah sehingga kelembaban akan terjaga. Sistem penutupan tanah akan menguntungkan terhadap perilaku rayap, yaitu akan meningkatkan siklus kelembaban dan stabilitas fisik tanah. Vegetasi penutup tanah juga menjadi penghalang (barier) agar tekstur tanah tidak tereduksi menjadi homogen dan mengurangi aliran permukaan tanah akibat hujan (Lamaourex et al. 2012).
Dominansi vegetasi penutup tanah yang diamati banyak ditemukan dari jenis gulma (Tabel 5). Gulma merupakan tanaman yang tumbuh sendiri tanpa bantuan manusia yang secara langsung maupun tidak langsung bersaing untuk mendapatkan unsur hara dengan tanaman budidaya sehingga keberadaannya mengganggu dan merugikan bagi tanaman budidaya. Jenis gulma yang ditemukan yaitu Axonopus compressus, Asystasia gangetica, Setaria plicata, Centhotheca lappacea, Cyslosorus aridus, Digitaria ascendens, dan Panicum sp.
Tabel 5 Jenis-jenis vegetasi penutup tanah di lapangan No Vegetasi
penutup tanah Lokasi pengamatan
1 A. compressusa Kebun Masjid Al-Hurriyyah, kebun kakao
Cikabayan, kebun kelapa sawit Cikabayan, halaman belakang FKH, kebun karet Rektorat, arboretrum ARL, gladiator, halaman parkiran FEMA
2 A. gangetica Agrimart, perumahan dosen, kebun sengon Rektorat
3 S. plicata Kebun kelapa sawit Cikabayan, halaman
perpustakaan LSI
4 C. lappacea Kebun kelapa sawit Cikabayan, kebun karet FPIK,
5 C. aridusa Kebun karet Rektorat, kebun karet FPIK
6 D. adscendensa Kebun sengon Rektorat
7 Pannicum sp. Agrimart 2
Keterangan: apopulasinya mendominasi pada kawasan tersebut
Hubungan antara Jenis rayap yang ditemukan dengan vegetasi penutup tanah, sebagian besar didominasi oleh gulma A. compressus, C. lappacea,dan C. aridus. Sebagian besar jenis rayap ditemukan pada lokasi yang mempunyai dominansi vegetasi penutup tanahnya dari ketiga gulma tersebut. Jenis rayap M. insperatus dan O. javanicus ditemukan pada vegetasi yang hampir sama, bedanya hanya pada satu jenis gulma saja yaitu M. insperatus ditemukan pada gulma D. adscendens sedangkan O. javanicus tidak ditemukan pada gulma tersebut tetapi ditemukan pada dominansi gulma Pannicum sp. (Tabel 4).
Vegetasi penutup tanah yang didominasi oleh gulma tersebut banyak tersebar di seluruh kawasan IPB. Gulma mempunyai kemampuan dispersi yang tinggi, selain itu mempunyai kemampuan mengkolonisasi kawasan yang cukup luas. Hal tersebut membuat gulma mampu tumbuh pada beberapa kawasan. Jenis gulma A. compressus merupakan gulma yang memiliki ciri dengan daya dispersi dan kolonisasi yang tinggi. Gulma ini banyak ditemukan di beberapa kawasan karena kemampuan adaptasinya yang tinggi terhadap cekaman lingkungan. Gulma
15 yang mampu bertahan pada cekaman lingkungan yang tinggi merupakan gulma yang berbahaya untuk tanaman budidaya karena akan sangat merugikan dalam persaingan unsur hara. Jenis gulma C. aridus merupakan gulma jenis paku-pakuan yang hidup di hutan belantara. Kemampuan hidupnya bergantung pada kondisi iklim mikro di sekitarnya. Iklim mikro yang cocok seperti kelembaban tinggi dan suhu rendah akan mendukung kehidupannya. Gulma jenis paku-pakuan ini banyak ditemukan di kawasan perkebunan karet karena pohon karet mempunyai tajuk yang tinggi sehingga akan membuat iklim mikro di bawahnya menjadi stabil. Karakterisitik gulma ini adalah bentuk daun yang lebar, daun berwarna hijau muda sampai tua dan tingginya dari 20-70 cm (Gambar 7). Kemudian jenis gulma D. ascendens merupakan jenis gulma jenis rumput-rumputan yang banyak tumbuh di perkebunan sengon Rektorat. Dominansinya meluas pada kawasan perkebunan tersebut. Gulma lain yang ditemukan antara lain A. gangetica, S. plicata, C. lappacea, dan Pannicum sp.. Gulma-gulma tersebut mempunyai dominansi yang lebih rendah dibandingkan dengan jenis D. adscendens.
Gulma merupakan tanaman yang sering merugikan tanaman budidaya, tetapi di sisi lain gulma yang menjadi tanaman penutup tanah yang mempunyai peranan penting dalam mengurangi aliran permukaan pada saat hujan. Hujan yang turun dan jatuh ke tanah akan menyebabkan erosi tanah, tetapi dengan adanya pengikatan tanah oleh gulma menyebabkan aliran permukaan yang membawa tanah tidak akan terjadi. Selain itu pengikatan tanah oleh gulma akan membuat iklim mikro di dalam tanah menjadi stabil karena tanah tidak langsung tersinari oleh matahari secara langsung. Iklim mikro tanah yang stabil akan meningkatkan mikrofauna, sehingga akan mendukung kehidupan rayap dengan ketersediaan makanan dan kondisi habitat yang stabil.
Gambar 7 Gulma yang ditemukan di lapangan (a) A. compressus , (b) C. aridus
Hasil Pengamatan Persentase Naungan
Rayap tanah banyak ditemukan di lapangan karena kawasan IPB masih terdapat banyak kawasan yang belum terbuka atau terganggu habitatnya. Keberadaan rayap di lapangan sangat berpengaruh terhadap habitat sekitar. Hutan primer, perkebunan, dan permukiman secara berurutan merupakan daerah yang tidak terganggu hingga yang paling terganggu. Tingkat gangguan habitat didasarkan pada intensitas aktivitas manusia terhadap habitat tersebut. Semakin intensif penggunaan lahan oleh manusia maka habitat tersebut akan semakin terganggu. Kawasan dengan celah (gap) penutup tanah, semakin lahan itu terbuka maka lahan tersebut akan semakin terganggu. Jenis rayap yang ditemukan
16
sebagian besar pada lokasi dengan tingkat persentase naungan 51-100% (Tabel 4) atau tingkat naungan yang tinggi (Gambar 8).
Semakin tinggi persentase naungan maka akan semakin banyak jumlah rayap yang ditemukan (Gambar 9). Persentase naungan rendah (0-25%) rata-rata jumlah rayap yang ditemukan sedikit dan didominasi oleh rayap O. javanicus. Rayap jenis O. javanicus ditemukan sebanyak 6 ekor pada kondisi naungan yang rendah. Hal ini dikarenakan rayap dari spesies O. javanicus. mempunyai kemampuan mengkolonisasi habitat yang tinggi pada tingkat gangguan yang tinggi. Berbeda dengan jenis rayap lain yang ditemukan yang relatif sedikit ditemukan pada naungan yang rendah.
Rata-rata semakin tinggi naungan semakin banyak jenis rayap yang ditemukan. Persentase naungan sedang (26-50%) hanya ditemukan 4 jenis rayap dan mempunyai jumlah yang berbeda pada setiap spesies rayap. Persen naungan tinggi (51-75%) dan naungan yang sangat tinggi (76-100%) ditemukan 6 jenis rayap. Pada naungan tinggi rayap yang ditemukan relatif sama yaitu pada kisaran 2, 3, dan 4. Berbeda pada naungan yang sangat tinggi, jenis rayap yang ditemukan ada 6 ekor tetapi jumlah rayap yang ditemukan berbeda-beda. Jenis rayap yang mendominasi pada naungan sangat tinggi yaitu oleh rayap jenis M. gilvus dan M. insperatus.
Persentase naungan sangat berpengaruh terhadap jumlah individu dan jumlah jenis rayap yang ditemukan. Semakin tinggi naungan semakin banyak jenis rayap yang ditemukan, tetapi jumlah individu rayap yang ditemukan relatif stabil pada naungan yang tinggi (51-75%) dibandingkan naungan yang sangat tinggi (76-100%). Hal ini disebabkan oleh kondisi naungan yang mempunyai iklim mikro relatif stabil sehingga lebih disukai oleh rayap. Iklim mikro yang relatif stabil akan meningkatkan mikrofauna tanah yang akan mendukung kehidupan rayap.
Lokasi yang menjadi tempat pengamatan mempunyai persentase naungan yang berbeda-beda. Lokasi dengan naungan 0-25% (naungan rendah) yaitu halaman parkiran FEMA, halaman parkiran FEM, halaman perpustakaan LSI, masjid Al-Hurriyyah, perumahan Dosen, dan halaman kandang FAPET; lokasi dengan persentase naungan 26-50% (naungan sedang) yaitu kebun sengon Rektorat dan Belakang FKH; persentase naungan 51-75% (naungan tinggi) yaitu kebun karet Rektorat dan Arboretrum ARL; sedangkan persentase naungan 76-100% (naungan sangat tinggi) yaitu kebun karet FPIK, kebun kakao Cikabayan, dan kebun sawit Cikabayan. Semakin besar persentase naungan berkorelasi positif dengan jumlah rayap yang ditemukan (Tabel 6). Kawasan dengan gap atau celah yang kecil sangat berpengaruh terhadap ketersediaan mikrohabitat yang sangat mendukung kehidupan rayap (Jones et al. 2003).
Kawasan dengan naungan yang lebih kecil akan menyebabkan sinar matahari akan langsung mengenai tanah. Hal ini akan meningkatkan iklim mikro yang ekstrim dalam tanah, sehingga suhu dan kelembaban tanah akan meningkat. Suhu dan kelembaban tanah yang tidak stabil menyebabkan mikrofauna tanah akan menjauh dan mencari lokasi dengan kondisi suhu dan kelembaban yang lebih stabil. Mikrohabitat yang terganggu akan berpengaruh juga terhadap aktivitas makan rayap. Tanah yang menjadi tempat hidup berbagai mikrofauna menjadi tidak sesuai untuk ditempati lagi. Rayap tanah akan semakin menghindari
17 kawasan yang terbuka dan akan mencari kawasan dengan kondisi habitat yang lebih stabil.
Gambar 8 Hubungan persentase naungan dengan jumlah rayap yang ditemukan
Penurunan keanekaragaman spesies rayap selain disebabkan oleh naungan yang lebih kecil, juga disebabkan oleh adanya sistem budidaya monokultur. Di utara Australia, keanekaragaman jenis semut di savana lebih sedikit jika dibandingkan di daerah hutan hujan, hal ini karena hutan hujan mempunyai komposisi penyusun pepohonan yang lebih banyak jika dibandingkan dengan savana (Andersen et al. 2008). Sistem budidaya monokultur akan menurunkan komunitas rayap di dalamnya karena mikrohabitat pendukung rayap kurang beragam. Semakin kompleks penyusun suatu ekosistem maka akan semakin beragam jenis organisme yang akan ditemui. Pada sistem monokultur, ketersedian sumber makanan melimpah untuk komunitas rayap tetapi mempunyai keragaman jenis makanan yang lebih sedikit dibandingkan dengan di kawasan hutan yang mempunyai beragam jenis pohon.
Kebun karet, kebun sawit, kebun sengon, dan kebun kakao merupakan suatu kawasan dengan sistem budidaya yang monokultur. Pada sistem budidaya monokultur rayap yang ditemukan tidak terlalu beragam jika dibandingkan dengan jenis rayap yang ditemukan di kawasan hutan dengan tingkat gangguan dan komposisi penyusun yang beragam (Jones et al. 2003). Kawasan IPB yang dulunya masih banyak pohon-pohon hutan sekarang lebih banyak permukiman dan perkebunan, sehingga jenis yang monokultur akan berpengaruh terhadap keanekaragaman jenis rayap yang ditemukan.
0 1 2 3 4 5 6 7 0-25 26-50 51-75 76-100 Ju m la h ra y ap (e kor) Persentase naungan (%) Macrotermes gilvus Odontotermes javanicus Microtermes insperatus Capritermes mohri Coptotermes curvignathus Schedorhinotermes javanicus
18
Tabel 6 Kawasan dengan berbagai naungan
Lokasi Jumlah rayap (ekor) Persentase naungan (%)
Halaman parkiran FEMA 2 15 a
Halaman parkiran FEM 1 15 a
Agrimart 1 20 a
Halaman perpustakaan LSI 1 20 a
Masjid Al Huriyyah 1 25 a
Perumahan Dosen 1 15 a
Halaman kandang FAPET 3 10 a
Kebun karet Rektorat 9 70 c
Kebun sengon Rektorat 2 50 b
Gladiator 1 60 c
Kebun kakao Cikabayan 4 85 d
Kebun sawit Cikabayan 6 80 d
Kebun karet FPIK 7 80 d
Belakang FKH 7 45 b
Arboretrum ARL 6 60 c
Keterangan: persentase naungan a = 0-25%, b =26-50%, c = 51-75%, dan d = 76-100%
Isolasi dan Karakterisasi Bakteri Simbion dari Saluran Pencernaan Rayap
Isolasi Bakteri Simbion dari Saluran Pencernaan Rayap
Rayap yang diperoleh dari lapangan kemudian diektraksi untuk mendapatkan bakteri simbionnya. Bakteri simbion diisolasi dari saluran pencernaan rayap bagian belakang (proktodeum) dan diperoleh jenis-jenis koloni yang ditemukan (Gambar 9). Jenis-jenis koloni yang ditemukan kemudian dipisahkan untuk mendapatkan koloni tunggalnya. Hal ini untuk memudahkan dalam karakterisasi.
Gambar 9 Hasil isolasi bakteri simbion dari proktodeum (a) koloni sebelum dimurnikan (b) koloni setelah dimurnikan
Karakterisasi Bakteri Simbion dari Saluran Pencernaan Rayap
Menurut Bignell (2006) semua rayap mempunyai pencernaan belakang (proktodeum) yang berisi sebagian besar oleh mikroorganisme dalam sistem pencernaannya. Bentuk, ukuran, dan perbedaan tingkat pada proktodeum semua
19 bervariasi tergantung taksanya. Proktodeum merupakan bagian dari pencernaan yang paling belakang yang menjadi saluran terakhir. Proktodeum ini menjadi tempat akumulasi semua sari-sari makanan yang diproses dengan bantuan enzim selulase. Enzim selulase membantu menghancurkan selulosa yang dimakan oleh rayap. Enzim ini dikeluarkan oleh mikrob simbion yang ada di dalam proktodeum untuk mendegradasi selulosa dari kayu yang dimakan oleh rayap. Proporsi mikroorganisme di dalam proktodeum lebih besar dibandingkan dinding kutikula atau duri-duri (spines) yang menonjol pada lumen (Bignell 2000).
Pengamatan karakter pertama yaitu melihat hasil pengujian Gram dari pewarnaan Gram dan KOH 3%. Bakteri hasil pewarnaan termasuk dalam kelompok Gram positif ditunjukkan dengan perubahan sel bakteri menjadi ungu dan termasuk Gram negatif dengan perubahan warna sel menjadi merah. Hal ini didasarkan pada tebal atau tipisnya dinding sel penyusun dari bakteri. Dinding sel bakteri Gram positif lebih tebal dibandingkan dengan dinding sel bakteri Gram negatif. Pada pewarnaan crystal violet, pewarna primer tersebut akan masuk ke dalam dinding-dinding sel bakteri Gram positif hingga proses pewarnaan yang terakhir. Berbeda dengan bakteri Gram negatif, dinding selnya lebih tipis sehingga akan tercuci pada tahap degradasi warna oleh etanol 95% dan proses pewarnaan safranin (pewarna tandingan) dinding sel Gram negatif akan terwarnai menjadi merah. Sedangkan pada pengujian menggunakan metode KOH, indikatornya sama yaitu berdasarkan dinding sel. Kelompok bakteri Gram positif mempunyai dinding sel yang tersusun atas lapisan peptidoglikan yang tebal (lebih dari 50%), sedangkan pada dinding sel bakteri Gram negatif hanya berdinding tipis (sepasang membran, lapisan sitoplasma dan lapisan luar). Dinding sel penyusun bakteri Gram negatif yang tipis akan memudahkan dalam pelisisan dinding selnya.
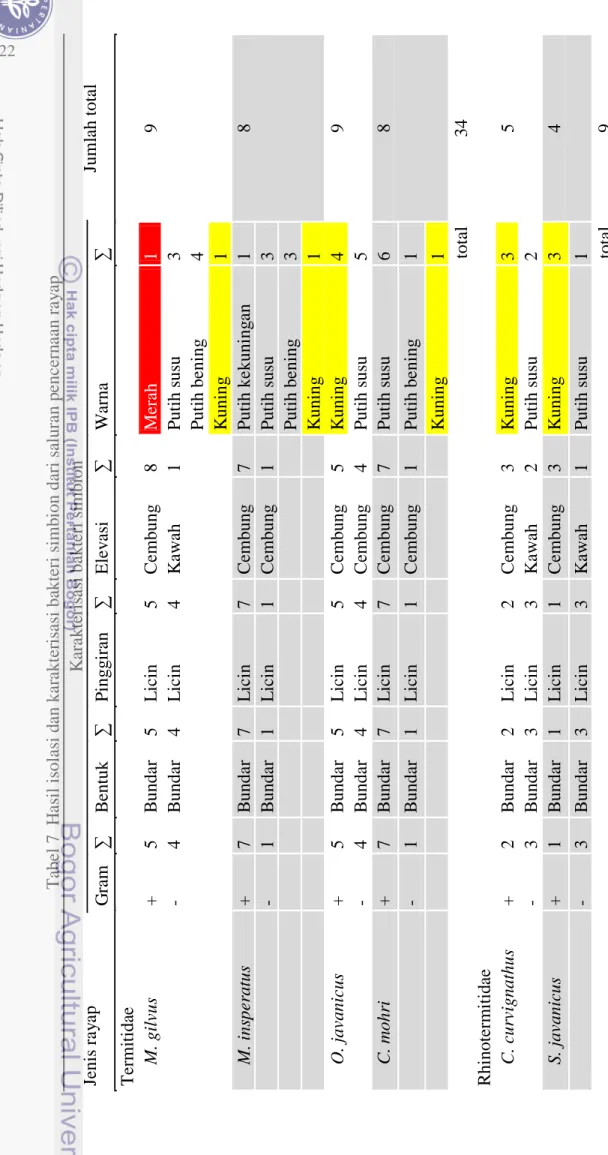
Ditemukan dua jenis Gram pada kedua famili rayap yang diujikan yaitu Gram positif dan negatif. Dari total isolat yang diamati, jenis Gram positif lebih banyak ditemukan pada famili Termitidae, sedangkan famili pada Rhinotermitidae ditemukan lebih banyak jenis bakteri Gram negatif dari total isolat yang diamati. Rata-rata dari koloni semua spesies rayap dari Famili Termitidae, ditemukan jenis bakteri Gram positif yang lebih mendominasi dibandingkan bakteri Gram negatif. Sebaliknya dengan isolat dari spesies rayap dari Famili Rhinotermitidae yang lebih didominasi oleh bakteri Gram negatif dibandingkan dengan bakteri Gram positif (Tabel 7). Meskipun ada perbedaan pada kedua jenis bakteri, tetapi bakteri jenis Gram positif dan negatif mempunyai peranan yang hampir sama yaitu membantu dalam proses pendegradasian selulosa.
Pengujian Gram pada bakteri merupakan langkah awal sebelum proses idenfikasi bakteri selanjutnya. Sebanyak 37 isolat bakteri Gram positif dan 16 isolat Gram negatif dari isolat total kedua famili. Famili Termitidae mempunyai 34 isolat (24 isolat Gram positif dan 10 isolat Gram negatif) sedangkan famili Rhinotermitidae sebanyak 9 isolat (3 isolat Gram positif dan 6 isolat Gram negatif).
Pengamatan selanjutnya yaitu karakter bentuk dan pinggiran pada hasil isolasi bakteri simbion menunjukkan bahwa semua koloni yang ditemukan mempunyai bentuk bulat dengan pinggiran halus (Gambar 10). Proktodeum yang sudah diektraksi dari masing-masing spesies rayap tidak ditemukan bentuk dan pinggiran yang berbeda antara spesies rayap satu dengan yang lainnya. Pengamatan karakter selanjutnya adalah bentuk elevasi dari koloni bakteri
20
simbion. Bentuk elevasi dapat diamati jika dilihat dari permukaan samping (lateral). Hasil isolasi bakteri simbion diperoleh dua jenis elevasi yaitu berbentuk cembung dan kawah (Gambar 11).
Gambar 10 Morfologi koloni bakteri yang berbentuk bulat
Bentuk elevasi cembung umumnya terdapat pada semua koloni dari semua spesies rayap. sedangkan bentuk elevasi kawah hanya ditemukan pada spesies rayap M. gilvus, C. curvignathus, dan S. javanicus. Spesies M. gilvus hanya ditemukan satu koloni yang berbentuk kawah, sedangkan koloni lainnya berbentuk cembung. Spesies C. curvignathus ditemukan dua koloni yang mempunyai elevasi berbentuk kawah, sedangkan tiga koloni lainnya berbentuk cembung. Ditemukan satu koloni berbentuk elevasi kawah pada spesies S. javanicus, sedangkan tiga koloni lainnya berbentuk cembung (Tabel 7).
Gambar 11 Elevasi koloni bakteri hasil ekstraksi proktodeum (a) bentuk cembung pada koloni bakteri, (b) skema cembung, (c) bentuk kawah pada koloni bakteri, dan (d) skema kawah
Elevasi yang ditemukan umumnya berbentuk cembung, sedangkan elevasi berbentuk kawah hanya ditemukan sedikit dari total koloni hasil isolasi bakteri simbion. Famili Termitidae hanya ditemukan elevasi bentuk kawah pada spesies M. gilvus saja, sedangkan spesies yang lain tidak ditemukan. Berbeda halnya dengan famili Rhinotermitidae yang ditemukan pada kedua spesies (C. curvignathus dan S. javanicus). Bentuk elevasi kawah ditemukan pada kedua spesies.
Pengamatan karakter warna pada koloni hasil isolasi bakteri simbion ditemukan empat jenis warna. Visual warna yang terlihat yaitu warna merah, kuning, putih susu, dan putih bening (Gambar 12). Warna putih susu dan putih bening umumnya terdapat pada semua spesies rayap, begitu juga dengan warna kuning.
21
Gambar 12 Visualisasi koloni bakteri hasil ekstraksi proktodeum (a) putih susu, (b) putih, (c) merah, dan (d) kuning
Warna kuning ditemukan pada semua spesies rayap tetapi jumlah koloni yang menampakkan warna kuning berbeda-beda. Spesies M. gilvus, M. insperatus, dan C. mohri hanya ditemukan satu koloni, sedangkan pada O. javanicus ditemukan empat koloni berwarna kuning. Pada spesies C. curvignathus dan S. javanicus ditemukan tiga koloni. Penampakkan warna merah hanya ditemukan pada spesies rayap M. gilvus yaitu sebanyak satu koloni. Spesies yang lain tidak ditemukan penampakkan warna merah pada koloni hasil isolasi bakteri simbion dari proktodeum masing-masing spesies rayap.
T abe l 7 H as il i sol as i d an ka ra kt er is as i ba kt eri s im b ion da ri s al ura n p enc ern aa n ra y ap Je ni s ra y ap K ara kt eri sa si ba kt eri s im b ion Jum la h t ot al G ra m ∑ B ent uk ∑ P ingg ira n ∑ E le v as i ∑ W arna ∑ T erm it ida e M. gi lv us + 5 B unda r 5 L ic in 5 Ce m bung 8 M era h 1 9 - 4 B unda r 4 L ic in 4 K aw ah 1 P ut ih s us u 3 P ut ih be ni ng 4 K uni n g 1 M. i ns pe rat us + 7 B und ar 7 L ic in 7 Ce m bung 7 P ut ih ke kuni ng an 1 8 - 1 B unda r 1 L ic in 1 Ce m bung 1 P ut ih s us u 3 P ut ih be ni ng 3 K uni n g 1 O . j av ani cus + 5 B unda r 5 L ic in 5 Ce m bung 5 K uni n g 4 9 - 4 B unda r 4 L ic in 4 Ce m bung 4 P ut ih s us u 5 C. m ohr i + 7 B unda r 7 L ic in 7 Ce m bung 7 P ut ih s us u 6 8 - 1 B unda r 1 L ic in 1 Ce m bung 1 P ut ih be ni ng 1 K uni n g 1 t ot al 34 Rhi not erm it id ae C. c ur vi gnat hus + 2 B unda r 2 L ic in 2 Ce m bung 3 K uni n g 3 5 - 3 B unda r 3 L ic in 3 K aw ah 2 P ut ih s us u 2 S. j av ani cus + 1 B unda r 1 L ic in 1 Ce m bung 3 K uni n g 3 4 - 3 B unda r 3 L ic in 3 K aw ah 1 P ut ih s us u 1 tot al 9 22
23
KESIMPULAN DAN SARAN
Kesimpulan
Keanekaragaman spesies rayap yang diperoleh dari sampling (stasiun pengamatan dan transek) di kawasan Kampus IPB Dramaga, Bogor diperoleh enam spesies rayap yaitu M. gilvus, O. javanicus, M. insperatus, C. mohri, C. curvignathus, dan S. Javanicus. Pengamatan rayap pada beberapa tipe habitat sangat berpengaruh terhadap persentase naungan dibandingkan habitat tanah dan vegetasi penutup tanah. Semakin tinggi persentase naungan semakin tinggi jumlah rayap yang ditemukan. Sedangkan jenis tanah, pH tanah, jenis vegetasi penutup tanah umumnya menunjukkan jumlah hasil (rayap yang ditemukan) yang sama. Rayap yang diperoleh dari lapangan kemudian diekstraksi bagian saluran pencernaan belakang dan diperoleh keragaman bakteri sebanyak 34 isolat dari famili Termitidae dan 9 isolat dari famili Rhinotermitidae berdasarkan karakter Gram dan morfologinya.
Saran
Perlu adanya pembuatan stasiun pengamatan yang mewakili setiap lokasi di IPB untuk mengetahui sebaran dan daya jelajahnya dan pemanfaatan bakteri simbion untuk pengembangan ilmu pengetahuan yang lebih luas.
25
DAFTAR PUSTAKA
Adams L, Boopathy R. 2005. Koloniion and characterization of enteric bacteria from the proktodeum of Formosan termite. Bioresource Tech. 96:1592– 1598.
Ahmad M. 1959. Key to The Indomalayan Termites. Lahore (PK): University of The Punjabi.
Andersen AN, Ingen LTV, Campos RI. 2008. Contrasting rainforest and savanna ant in monsoonal northern Australia: a rainforest patch in a tropical savanna lanscape [abstract]. Australian Journal of Zoology [internet]. 2013 Mar 19. 55:363-369. Tersedia pada: http://www.mendeley.com/research/contrasting- rainforest-savanna-ant-faunas-monsoonal-northern-australia-rainforest-patch-tropical-sav/. doi: 10.1071/ZO07066.
Bignell DE. 2000. Introduction to symbiosis. Di dalam: Abe T, Bignell DE, Higashi M, editor. Termites: evolution, sociality symbioses. Dordrecht (US): Kluwer Academic Publisher. hlm 189-208.
Bong JCF, King PJH, Ong KH, Mahadi NM. 2012. Termite assemblages in oil palm plantation in Sarawak, Malaysia. J of Entomol. 9:62-72. doi: 10.3923/je.2012.68.78.
Cho MJ, Kim YH, Shin K, Kim YK, Kim YS, Kim TJ. 2010. Symbiotic adaptation of bacteria in the gut of Reticulitermes speratus: Low endo-b-1,4-glucanase activity. Biochem and Biophy Research Com.. 395 (2010):432–435. doi:10.1016/j.bbrc.2010.04.048.
Du LV, Truc NH. 2009. Termite bio-control on cacao seedling: vertiver grass application. Sanrem CRSP. 6:1-6.
Eggleton P, Bignell DE, Hauser S, Dibog L, Norgrove L, Madong B. 2002. Termite diversity across an anthropogenic disturbance gradient in the humid forest zone of West Africa. Agri Ecosys and Envir. 90:189 - 202.
Herlinda S, Septiana R, Irsan C, Adam T, Thalib R. 2010. Populasi dan serangan rayap (Coptotermes curvignathus) pada pertanaman karet di Sumatera Selatan. Prosiding Seminar Nasinoal; 2010 Des 13-14. hlm 528-534.
Hethener P, Brauman A, Garcia J. 1992. Clostridium termitidis sp. Nov., a cellulolytic bacterium from the gut of the wood-feeding termite, Nasutitermes lujae. System Appl Microbiol. 15:52-58.
Husseneder C, Berestecky JM, Grace JK. 2009. Changes in composition of culturable bacteria community in the gut of the formosan subterranean termite depending on rearing conditions of the host. Annals Of The Entomol Soc of America. 102: 498-507.
Inward D, Beccaloni G, Eggleton P. 2007. Death of an order: a comprehensive mplecular phylogenetic study confirms that termites are eusosial cockroaches. Biol Lett. 3:331-335.
Jones DT, Susilo FX, Bignell DE, Hardiwinoto S, Gillison AN, Eggleton P. 2003. Termites assemblage collapse along a land-use intensification gradient in lowland central Sumatra, Indonesia. J of App Eco. 40:380-391.
26
Keng WM.2006. Spesies comparison of termite (Isoptera) in primary forest of Tawau Hill Park, Sabah and adjacent cocoa plantation area.[thesis]. Sabah (MY): University Malaysia Sabah.
Klepzig KD, Adams AS, Handelsman J, Raffa KF. 2009. Symbioses: A Key Driver of Insect Physiological Processes, Ecological Interactions, Evolutionary Diversification, and Impacts on Humans. Envir Entomol. 38:67-77.
Lamaourex S, O’Kane MA. 2012. Effect of termites on soil cover system performance. Mine Clousure. 432:446.
Mahmudi A. 2008. Keragaman lignin terlarut asam (acid soluble lignin) pada empat jenis kayu cepat tumbuh [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Nandika D, Rismayadi Y, Diba F, Harun JP. 2003. Rayap: Biologi dan Pengendaliaannya. Surakarta (ID) : Muhammadiyah University Press. Pribadi T, Raffiudin R, Harahap IS. 2011. Termites community as environmental
bioindicators in highlands: a case study in eastern slopes of Mount Slamet, Central Java. Biodiversitas. 12:235-240.
Sapp J. 1994. Evolution By Association: A History of Symbiosis. Oxford (US): University Press.
Sceffrahn RH, Su NY. 1994. Key To Soldier and Winged Adult Termites (Isoptera) of Florida. Florida Entomol Online. 77: 4.
Shelton TG, Foshee W, Appel AG. 2000. Drywood termite biology, identification, and control. Alabama Cooper Exten Syst. 1-3.
Tarumingkeng RC. 1971. Biologi dan Pengendalian Rayap Perusak Kayu Indonesia. Bogor (ID): LPPK No 138.
Tho YP. 1992. Termites of Peninsular Malaysia. Kirton LG, editor. Kepong, Kuala Lumpur (MY): Malayan Forest Records.
Upadhyaya SK, Manandhar A, Mainali H, Anaya R, Pokhrel, Rijal A, Barun, Pradhan, Koirala B. 2012. Koloniion and Characterization of Cellulolytic Bacteria from Gut of Termite. Rentech Symposium Compendium. 1:14-18.