TINJAUAN PUSTAKA
Morfologi Lambung Domba
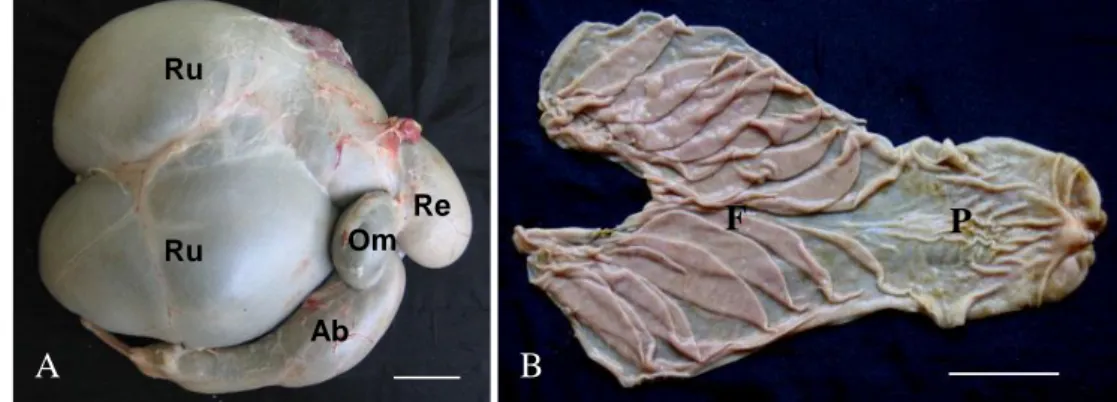
Lambung domba terdiri dari dua bagian utama, yakni lambung depan yang tidak berkelenjar (proventrikulus) dan lambung kelenjar. Lambung depan terdiri dari rumen, retikulum, dan omasum yang pada bagian mukosanya tersusun atas epitel pipih banyak lapis yang melakukan pencernaan secara mekanis, sedangkan lambung kelenjarnya adalah abomasum. Abomasum berada pada sisi kanan dari ruang abdomen. Fungsi abomasum dapat dianalogikan sama dengan fungsi lambung monogastrik pada berbagai spesies (Dyce et al. 1996). Kapasitas lambung domba dewasa berkisar antara 15 sampai dengan 18 liter dengan komposisi kapasitas rumen sekitar 71%, retikulum 8%, omasum 2%, dan abomasum 19% (Getty 1975). Gambaran morfologi abomasum dapat dilihat pada Gambar 1.
Gambar 1 Proporsi ukuran lambung (A) dan mukosa abomasum domba (B) Ru = rumen, Re = retikulum, Om = omasum, Ab = abomasum F = fundus, P = pilorus (Bar = 5 cm) (Sumber: Putra 2009).
Abomasum memiliki tiga daerah kelenjar yaitu: kardia, fundus, dan pilorus. Daerah kelenjar kardia adalah area sempit penghasil mukus pada kranial abomasum. Daerah kelenjar fundus merupakan daerah kelenjar lambung yang paling luas dan terdiri dari sel epitel permukaan yang menghasilkan mukus, sel leher mukus yang memproduksi cairan penetral, parietal cells atau oxyntic cells yang menghasilkan asam klorida (HCl), chief cells yang memproduksi prekursor enzim seperti pepsinogen, dan sel enteroendokrin sebagai penghasil hormon-hormon pencernaan (Dellmann & Eurell 1998) Daerah kelenjar pilorus menghasilkan mukus, untuk menetralisasi keasaman cairan lambung (chyme)
A
B
Ru Ru Re Om AbF
P
yang terletak di kaudal abomasum dan tersambung dengan duodenum pada bagian distal. Pilorus mengatur perpindahan chyme dari lambung ke duodenum dan mencegah aliran balik dari duodenum ke lambung (Banks 1993; Bergman et al. 1996; Colville & Bassert 2002).
Sel-sel pada mukosa abomasum pada ruminansia mensekresikan sedikitnya tiga macam enzim pemecah protein yaitu pepsin, khimosin, dan gastriksin. Enzim-enzim tersebut bersifat asam dan termasuk kedalam golongan endopeptidase yang disekresikan dalam bentuk inaktif, masing-masing pepsinogen, prokhimosin dan progastriksin (Heishi et al. 1995). Enzim akan diubah menjadi bentuk aktif oleh HCl yang diproduksi oleh sel-sel parietal pada bagian fundus menjadi pepsin, khimosin, dan gastriksin (Dellman & Eurell 1998). Pada daerah kelenjar fundus domba umur dewasa muda, sel-sel yang imunoreaktif terhadap pepsinogen terdeteksi dalam jumlah banyak, sedangkan sel-sel yang imunoreaktif terhadap prokhimosin jumlahnya sedang (Fitriyani 2006).
Bahan Bioaktif Rennet
Rennet merupakan bahan bioaktif hasil ekstraksi abomasum ruminansia muda, khususnya sapi yang digunakan sebagai starter dalam proses pembuatan keju karena mengandung enzim khimosin dengan kadar tinggi. Rennet ini digunakan sebagai bahan awal dalam produksi dadih-dadih keju (Cheeseman 1981). Rennet yang berasal dari hewan yang lebih tua memiliki kandungan pepsin yang lebih tinggi dibandingkan dengan rennet hewan muda yang tinggi kandungan khimosin. Tingginya pepsin menyebabkan perbedaan dalam proses koagulasi susu dan juga dalam cita rasa keju yang dihasilkan, seperti tekstur keju yang lebih lembek dan adanya rasa pahit (Kilara dan Iya 1984). Ekstrak rennet dari anak sapi mengandung 88-94% khimosin dan 6-12% pepsin, dan sebaliknya ekstrak yang berasal dari sapi dewasa yaitu mengandung 90-94 % pepsin dan hanya 6-10% khimosin (Broome dan Limsowtin 1998).
Pemanfaatan rennet selain dari sapi muda juga telah dikembangkan, seperti rennet dari kambing muda (Bolen et al. 2003), tanaman, dan mikroba yang direkayasa secara genetik (genetic modified organism = GMO) (Cmegar & Cruegar 1984; Teuber 1993).
Rennet yang diperoleh dari spesies hewan yang berbeda memiliki kemampuan mengkoagulasikan susu yang bervariasi.
Rennet mikroba yang banyak digunakan sebagai pengganti rennet sapi muda berasal dari mikroorganisme seperti kapang dan bakteri. Mikroorganisme yang telah digunakan dalam pembuatan rennet GMO antara lain Bacillus polymyxa, B. subtilis, B. cereus, Endothea parasiticus, Mucor pusillus, dan Mucor miehei. Biaya untuk memproduksi rennet dari mikroba lebih murah dibandingkan dengan pembuatan rennet konvensional yang diekstrak dari lambung ruminansia. Rennet mikroba bersifat seperti enzim tripsin dan pH optimum untuk aktivitasnya berkisar antara 7-8. Akan tetapi, produk rennet ini mempunyai sedikit rasa pahit, sehingga kurang disukai, walaupun keju yang dihasilkan sangat mirip dengan produk-produk keju komersial lainnya (Daulay 1990).
Berbagai tanaman juga diketahui mengandung bahan yang dapat diisolasi untuk menghasilkan senyawa sejenis rennet yang dapat menggumpalkan susu. Penggunaan “rennet” tanaman dalam pembuatan keju, misalnya penggunaan getah pohon ara (Ficus carica), papain dari pohon dan buah pepaya, bromelin dari nanas, dan rezin dari biji jarak. Beberapa ekstrak tanaman tersebut mempunyai aktivitas proteolitik yang terlalu kuat, sehingga menghasilkan cita rasa yang sangat pahit pada keju. Menurut Daulay (1990), ekstrak dari beberapa jenis tanaman akan menghasilkan ekstrak kasar yang membentuk suatu kombinasi koagulasi asam dan enzim sehingga cocok digunakan terutama untuk pembuatan dadih keju.
1. Enzim Khimosin
Khimosin atau rennin adalah suatu enzim proteolitik yang termasuk dalam golongan protease yang bersifat asam, sehingga akan aktif pada pH asam. Khimosin diidentifikasi berdasarkan residu asamnya (aspartat atau glisin) sebagai khimosin A dan B yang masing-masing memiliki pH optimum 4.2 dan 3.7 (Atallah 2007). Khimosin dihasilkan oleh sel utama (chief cell) pada mukosa fundus lambung anak hewan yang masih menyusu (Kumar et al. 2006). Enzim ini mempunyai fungsi spesifik untuk mengkoagulasikan susu sehingga susu tertahan lebih lama di saluran pencernaan dan mengoptimalkan proses pencernaan. Khimosin banyak digunakan dalam industri keju khususnya dalam proses awal untuk menggumpalkan susu (Bowen 1996).
Khimosin dengan kadar tinggi ditemukan pada hewan yang masih mengonsumsi susu induk, Sekresinya semakin menurun sejalan dengan
pertambahan umur hewan. Pada hewan ruminansia yang telah lepas sapih dan mulai mengonsumsi pakan hijauan, produksi khimosin akan digantikan dengan pepsin (Daulay 1990). Pada usia di atas satu minggu produksi khimosin secara gradual akan menurun drastis, namun pada ruminansia produksi khimosin tidak pernah benar-benar terhenti walaupun telah memasuki usia dewasa (Fox 1993).
Enzim ini disekresikan dalam bentuk inaktif proenzim yakni prokhimosin yang akan diaktivasi oleh HCl menjadi khimosin (Dellman dan Eurell 1998). Prokhimosin anak sapi memiliki berat molekul 40.8 kDa dan khimosin 35.6 kDa (Atallah 2007). Menurut Suhartono (1992), aktivasi prokhimosin melibatkan pemotongan ujung terminal-N prokhimosin dan mengakibatkan penurunan berat molekul prokhimosin dari 36 kDa menjadi khimosin dengan berat molekul 31 kDa. Enzim khimosin stabil pada pH 5.3 sampai dengan 6.3.
2. Enzim Pepsin
Pepsin merupakan salah satu dari tiga enzim proteolitik utama di dalam sistem pencernaan hewan dewasa, selain chymotripsin dan tripsin. Enzim pepsin seperti halnya khimosin, dihasilkan oleh chief cell mukosa lambung yang secara alami juga terdapat dalam rennet ruminansia. Enzim pepsin disintesis dalam bentuk prekursor pepsinogen dengan berat molekul 42 kDa. Pepsinogen kemudian dapat diaktivasi melalui dua jalur, pertama melalui aktivasi oleh HCl. Protein yang masuk kedalam lambung akan merangsang pengeluaran hormon gastrin yang selanjutnya merangsang pengeluaran HCl oleh sel parietal mukosa lambung, serta sekresi pepsinogen dari sel utama. Aktivasi kedua melalui proses autokatalisis oleh pepsin itu sendiri. Kedua jalur aktivasi tersebut membentuk pepsin dengan berat molekulnya yang menurun menjadi 35 kDa dan stabil pada pH 5.0-5.5 (Suhartono 1992). Sementara menurut Harrow dan Mazur (1958) titik isolistrik pepsin yaitu 2.85. Berdasarkan Winarno (1983), berat molekul dari pepsin adalah 33 kDa yang mempunyai 321 residu asam amino, sangat stabil pada pH 5.0-5.3, dan sangat aktif pada pH 1-4 dengan keaktifan optimum pada pH 1.8.
Kuantitas pepsin dipengaruhi oleh umur dan jenis pakan hewan. Pedet yang hanya mengonsumsi susu mengandung sekitar 6-12 % pepsin pada
ekstrak abomasumnya, sedangkan pedet yang telah mengonsumsi makanan padat (selain susu) akan mengalami peningkatan kandungan enzim pepsin dan pengurangan kadar khimosin. Hasil koagulasi susu oleh pepsin akan menghasilkan waktu koagulasi yang lama, koagulan yang lunak, kehilangan lemak dalam whey, terbentuk peptida pahit, serta tekstur dan dadih (curd) keju yang lebih lunak (Kilara dan Iya 1984). Komplemen khimosin dengan pepsin merupakan salah satu alternatif dalam industri pengolahan keju. Campuran khimosin dan pepsin dianggap paling optimum dan paling sering dipilih karena sulitnya mendapatkan khimosin murni.
Koagulasi Susu
Susu terdiri atas bahan-bahan yang terdispersi dalam air, terutama kalsium, fosfat, dan protein. Bagian protein susu dibagi menjadi dua fraksi, yaitu kasein dan protein serum (whey protein). Kasein merupakan protein utama dalam susu yang dapat mencapai 80% dari total protein susu. Kasein termasuk ke dalam golongan fosfoprotein dengan berat molekul 20-30 kDa. Kasein akan bergabung dengan ion kalsium (Ca2+) membentuk agregat koloid yang disebut misel (Suhartono 1992). Kasein terdiri atas empat fraksi protein yaitu alpha (α), beta (β), kappa (κ), dan gamma (γ) yang mempunyai sifat khusus yaitu mudah menggumpal oleh adanya pengasaman atau penambahan rennet (Daulay 1990).
Proses koagulasi susu dapat terjadi akibat aktivitas enzim, asam, dan mikroba. Koagulasi susu oleh enzim terutama aktivitas enzim khimosin menjadi prioritas dalam industri keju. Susu digumpalkan oleh khimosin menjadi koagulan (curd) yang lunak dan lembut, serta memisahkan cairannya (whey) dari curd dengan kecepatan yang seragam. Selain khimosin, pepsin juga dapat menggumpalkan susu, tetapi koagulannya lebih sensitif terhadap perubahan pH dan suhu, serta dapat menyebabkan dadih berasa pahit (Daulay 1990). Menurut Suhartono (1992), kekuatan aktivitas koagulasi enzim khimosin lebih tinggi dibandingkan dengan enzim protease lainnya, seperti pepsin dan khimotripsin.
Proses koagulasi dengan reaksi enzimatis terjadi karena penambahan rennet yang bereaksi dengan kappa kasein akan memecah ikatan fenilalanin-metionin menghasilkan para-kasein dan menghancurkan aktivitas penstabilannya terhadap αs-kasein dan β-kasein. Pemecahan ikatan ini akan menyebabkan terpisahnya komponen yang bersifat hidrofilik dari para-kasein dan terbentuknya ikatan dengan ion Ca2+ yang melakukan penggabungan dengan komponen susu
lainnya membentuk curd yang terpisah dari whey (Goenardjoadi 1988, Daulay 1990). Sedangkan proses koagulasi susu nonenzimatis terjadi karena penambahan senyawa asam. Menurut Van Slyke et al. (1949) diacu dalam Widyowatie (1980) dengan bertambahnya kandungan asam pada susu, akan terjadi pembentukan asam laktat dari laktosa karena aktivitas bakteri. Asam laktat akan menyebabkan perubahan sifat fisik dan kimia dari kasein susu. Netralisasi muatan negatif dari kasein oleh ion H+ dari asam laktat akan menyebabkan tercapainya pH isolistrik kasein yang mengakibatkan protein terkoagulasi. Pengumpalan akan sempurna bila semua muatan kasein menjadi netral.
Pembentukan curd setelah penambahan khimosin pada susu terjadi melalui dua tahap. Pada tahap pertama, terjadi peningkatan komponen nitrogen terlarut karena aktivitas khimosin. Komponen nitrogen terlarut ini berasal dari molekul κ-kasein. Pada tahap kedua, terjadi agregasi misel kasein yang telah berubah secara enzimatik, sehingga terbentuk struktur gel (Suhartono 1992). Κappa-kasein kemudian terurai menjadi dua bagian pada ikatan asam amino fenilalanin dan metionin pada nomor 105-106 menjadi makropeptida ρ-κ-kasein dan makropeptida κ-kasein. Apabila sampai 90% κ-kasein telah terhidrolisis, maka terjadi agregat protein yang meningkatkan viskositas susu sampai terbentuk struktur gel. Produk makropeptida yang dihasilkan dari proses hidrolisis bersifat larut air, sedangkan struktur ρ-κ-kasein mengendap. Putusnya ikatan peptida antara fenilalanin dan metionin pada κ-kasein ini mengakibatkan hilangnya kestabilan misel kasein, sehingga fraksi kasein yang lain ikut mengendap (Suhartono 1992).
Pemisahan Protein
Protein maupun peptida memiliki komponen kompleks yang memerlukan proses sekuensi untuk mendapatkan komponen biologisnya (Morris 1976). Hal inilah yang kemudian melatarbelakangi perkembangan proses pemisahan protein. Faktor-faktor yang mempengaruhi pemisahan protein adalah molekul, muatan, serta sifat hidrofobiknya (Harrow & Mazur 1958). Pada awalnya proses pemisahan protein dilakukan dengan metode sederhana memisahkan fase yakni dengan metode filtrasi dan pemisahan fase liquid dengan pengeringan (drying). Pada perkembangannya, metode separasi protein dilakukan dengan cara yang
lebih progresif yakni dengan kristalisasi dan distilasi. Kesulitan dalam mengaplikasikan teknik pemisahan protein dipengaruhi oleh substansi yang ingin diteliti terdiri dari material yang sangat kompleks dan beragam, selain itu juga disebabkan bioavaibilitasnya yang sangat kecil di alam sehingga membutuhkan teknik pemisahan dengan sensitivitas yang tinggi (Morris 1976). Pemurnian protein merupakan proses yang dilakukan untuk mendapatkan isolat homogen dari suatu substansi. Pemurnian dikatakan berhasil apabila tidak dapat lagi membuktikan bahwa suatu material terdiri dari lebih dari satu substansi. Hal ini hanya dapat dilakukan dengan mengaplikasikan berbagai teknik pemisahan protein dan dilanjutkan dengan sistem analisa yang sesuai. Proses analisa tersebut dapat dilakukan secara biologis maupun kimiawi (Morris 1976).
Teknik yang banyak dikembangkan dalam pemisahan protein terbagi menjadi dua metode mayor, yakni chromatographic dan non-chromatografic. Metode non-chromatographic dalam hal ini antara lain elektroforesis, presipitasi, serta filtrasi membran. Berdasarkan Shetty et al. (2006) kromatografi merupakan suatu teknik purifikasi dimana komponen dari sampel dipisahkan berdasarkan kemampuan masing-masing komponen tersebut untuk berinteraksi dengan fase gerak ataupun fase diam yang dilalui sampel. Metode purifikasi chromatographic terdiri atas dua fase, yaitu fase diam (stationary phase) dan fase bergerak (mobile phase). Fase diam dapat berbentuk padat, gel, cair atau campuran padat dan cair, sementara fase gerak dapat berbentuk cair atau gas dan mengalir melewati fase diam. Semua metode chromatographic bekerja dengan dasar keseimbangan yang dicapai antara fase diam dan fase gerak.
Metode non-chromatographic elektroforesis adalah suatu teknik pemisahan senyawa berdasarkan kecepatan migrasi senyawa yang bermuatan dibawah pengaruh medan listrik (Caprette 1996). Dengan teknik elektroforesis molekul-molekul biologis yang memiliki berat molekul tinggi seperti karboidrat, lipid, asam nukleat, dan kompleks lipid-karbohidrat atau kompleks lipid-protein dapat dipisahkan berdasarkan berat molekulnya (Djuwita 2004).
Salah satu teknik pengembangan dari elektroforesis adalah gel elektroforesis yang menggunakan basis selulosa-asetat. Gel elektroforesis dapat diklasifikasikan berdasarkan media gel yang digunakan, yakni starch gel,. polyacrylamide gel, discontinuous gel electrophoresis, sodium dodecyl sulfate-polyaccrylamide electrophoresis, dan agarose gel electrophoresis.
Liquid Isoelectric Focusing Rotofor® (BioRad)
Isoelectric focusing (IEF) merupakan teknik non-chromatografic elektroforesis untuk memisahkan molekul berdasarkan perbedaan muatan listrik dengan prinsip mobilitas elektrolit dalam fungsi pH (Boyer 1986). Protein, termasuk didalamnya enzim membawa muatan listrik, baik positif, negatif, maupun netral. Muatan listrik dari suatu protein dipengaruhi oleh pH lingkungan di sekitar protein tersebut.
Molekul yang akan difraksinansi dipreparasi ke dalam medium dengan pH gradien tertentu yang difasilitasi dengan penggunaan amfolit yang akan memperluas rentang pH dalam medium. Arus listrik dari sumber listrik mengalir dalam medium membentuk ujung-ujung kutub elektroda; anoda yang bermuatan positif di salah satu ujung dan katoda yang bermuatan negatif di ujung lainnya. Molekul yang membawa muatan negatif akan bermigrasi melalui gradien pH menuju elektroda bermuatan positif, sedangkan molekul yang bermuatan positif akan bermigrasi ke arah sebaliknya (Boyer 1986). Hal ini akan terus berlangsung sampai pada titik isolistrik. Titik isolistrik (isoelectric point = pI) yaitu suatu kondisi dimana selisih muatan listrik pada permukaan protein adalah nol. Pada kondisi inilah susu menggumpal dengan baik menjadi dadih karena tidak ada muatan yang tertinggal untuk mempertahankan kasein dalam suspensi.
Gambar 2 Alat Rotofor®, terdiri atas focusing chamber (A) yang terdiri dari elektroda katoda (1), dan elektroda anoda (2); harvesting apparatus (B), yang terdiri dari collection tubes (1), dan vaccum vein (2).
Teknik IEF merupakan langkah awal dalam proses pemurniaan protein. Protein dipisahkan berdasarkan titik isolistrik dan dianalisa kandungan proteinnya dengan SDS-PAGE berdasarkan berat molekulnya. Liquid IEF Rotofor® mengisolasi protein dengan cara meningkatkan gradien pH linear dalam medium
A
B
1
2
1
yang beraliran listrik. Protein yang bersifat asam dengan muatan positif akan bermigrasi menuju wilayah katoda yang bermuatan negatif. Perpindahan tersebut juga diikuti oleh pelepasan ion hidrogen [H+] sampai tercapai selisih muatan nol dan protein akan berhenti bermigrasi. Apabila protein memperoleh muatan secara tidak sengaja akibat penyebaran dalam gradien pH, maka medan beraliran listrik tersebut akan mengembalikan protein tersebut pada rentang pH yang sesuai dengan titik isolistriknya. Protein yang tefraksinasi kemudian dipanen yang selanjutnya mengalami proses pemurnian lanjutan menggunakan gel elektroforesis (Perrit et al. 1992).
1. Preparasi Sampel
Faktor yang mempengaruhi keberhasilan pemisahan protein dalam penyiapan sampel antara lain konsentrasi garam, homogenitas, dan kelarutan. Sampel yang mengandung konsentrasi garam melebihi 10mM harus mengalami proses salting-out (pelepasan garam) sebelum penambahan amfolit ke dalam sampel. Hal tersebut untuk memastikan kisaran pH amfolit dapat diperluas hingga mencapai rentang pada focusing chamber (Gambar 2) dan voltase optimal dapat diaplikasikan. Konsentrasi garam yang tinggi dalam sampel akan bermigrasi menuju kutub-kutub anoda dan katoda yang dapat mengurangi efektivitas pemisahan. Penambahan buffer atau larutan penyangga akan meningkatkan konduktivitas dan menurunkan resolusi sampel. Efek lain dari penambahan buffer akan menekan gradien pH pada wilayah pKa buffer [BioRad].
Sampel yang tidak homogen dan keruh harus dihomogenisasi terlebih dahulu melalui filtrasi maupun sentrifugasi untuk memisahkan debris yang dapat menghalangi membran anion maupun kation. Kelarutan sampel menunjukkan kemampuan presipitasi dalam gel IEF analitik. Peningkatan kelarutan dapat dilakukan dengan penambahan urea 3-5 M yang telah diionisasi sebelumnya [BioRad].
2. Analisa Fraksi
Analisa fraksi dilakukan untuk menentukan kandungan protein yang terdapat dalam fraksi yang telah dipanen. Analisa yang paling umum adalah dengan menggunakan SDS-PAGE atau IEF gel dalam rentang pH 3-10. Metode lain yang dapat diaplikasikan untuk menganalisis fraksi bergantung
pada jenis protein meliputi uji khusus dan pengujian menggunakan metode immunoblotting. Kandungan amfolit dalam sampel dapat mempengaruhi kualitas pengujian. Pemisahan amfolit dari sampel dapat melalui teknik dialisa, presipitasi garam dengan ammonium sulfat, dan teknik kromatografi.
Elektroforesis Gel Poliakrilamid
Elektroforesis digunakan untuk memisahkan campuran asam nukleotida ataupun protein berdasarkan pergerakan molekul-molekul yang bermuatan dibawah pengaruh medan listrik. Molekul-molekul biologis yang memiliki berat molekul tinggi seperti karbohidrat, lipid, asam nukleat, dan kompleks lipid-karbohidrat atau kompleks lipid-protein dapat dipisahkan berdasarkan berat molekulnya. Prinsip perpindahan muatan mengakibatkan molekul akan bermigrasi ke arah katoda atau anoda, bergantung dari muatannya. Banyak molekul biologis seperti asam amino, peptida, protein, nukleotida dan asam nukleat memiliki muatan listrik yang besarnya tergantung pada jenis molekul, pH, dan komponen medium pelarutnya (Djuwita 2004).
Elektroforesis gel melalui agarose atau poliakrilamid merupakan metode yang paling umum dan sering dipergunakan dalam penelitian maupun biologi molekuler terapan. Secara umum, gel agarose lebih ditujukan untuk pemisahan molekul yang berukuran besar, sedangkan gel poliakrilamid untuk molekul yang berukuran lebih pendek (Djuwita 2004). Elektroforesis gel dapat digunakan untuk menentukan atau mendeteksi berbagai hal berikut: berat molekul (BM) suatu bahan (fragmen DNA, RNA atau protein); banyaknya jenis protein dalam suatu sampel misalnya serum albumin; terjadinya pemalsuan bahan atau kerusakan bahan, ada tidaknya suatu infeksi virus atau bibit penyakit lainnya dengan cara mendeteksi antibodi yang terbentuk.
Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) adalah metode yang banyak diaplikasikan untuk analisa campuran protein secara kualitatif, khususnya untuk melihat hasil purifikasi protein dan menentukan estimasi jumlah dan berat molekul protein. SDS poliakrilamid gel elektroforesis terbatas pada berat molekul 10.000 – 200.000 Dalton (Boyer 1986). Menurut Wilson dan Walker (1999), sodium dodecyl sulphate (SDS) (CH3-(CH2)10-CH2OSO3-Na+) merupakan detergen yang tidak bermuatan (anionik). SDS-PAGE dilakukan pada pH sekitar netral. SDS akan membentuk kompleks dengan protein dan kompleks ini bermuatan negatif karena
gugus-gugus anion dari SDS. Kompleks SDS-protein yang lebih besar mempunyai mobilitas yang lebih kecil, sedangkan kompleks yang lebih kecil memiliki mobilitas yang lebih besar.
Berat molekul protein dapat ditentukan dengan kalibrasi menggunakan standar protein yang telah diketahui berat molekulnya (marker) (Nur & Adijuwana 1989; Rybicki & Purves 2000). Penentuan berat molekul suatu fraksi dapat dilakukan dengan cara kualitatif dan kuantitatif. Cara kualitatif dilakukan dengan menggunakan patokan pita standar protein terhadap pita sampel, sedangkan cara kuantitatif dapat dilakukan dengan menghitung mobilitas relatif.