II.
TINJAUAN PUSTAKA
A. KEDELAI
Kedelai merupakan tanaman kacang-kacangan yang termasuk dalam famili Leguminosa, subfamili Papilionidae, genus Glycine dan spesies max, sehingga nama latinnya dikenal sebagai
Glycine max (Koswara 1992). Struktur biji kedelai terdiri atas 3 bagian utama, yaitu kulit biji (hull),
keping biji (kotiledon) dan hipokotil (Wolf 1989). Struktur biji kedelai secara lengkap dapat dilihat pada Gambar 1.
Gambar 1. Struktur biji kedelai (Anonima 2010)
Secara umum kedelai terdiri atas 8% kulit biji, 90% kotiledon dan 2% hipokotil (Handono, 1985). Kulit biji terdapat pada lapisan permukaan luar yang disusun oleh beberapa lapisan sel, sedangkan kotiledon adalah permukaan sel yang ditutupi oleh sel epidermis serta mengandung protein dan minyak pada bagian dalam (Wolf 1989).
Penampakan fisik kedelai memiliki keragaman yang cukup luas. Warna, ukuran, bentuk biji, sifat fisik maupun sifat kimia kacang kedelai sangat bervariasi. Perbedaan ini dipengaruhi oleh faktor varietas dan keadaan lingkungan tanamnya (Smith dan Circle 1977).
Seluruh bagian kedelai termasuk daun, batang dan bijinya dapat dimanfaatkan untuk pangan, obat dan pakan. Bagian keping biji (kotiledon) merupakan bagian yang paling umum untuk diolah menjadi berbagai produk olahan pangan. Hal tersebut disebabkan tingginya kandungan protein dan lemaknya (Wolf dan Cowan 1971)
1. Komposisi Kimia Kedelai
Kedelai mengandung jumlah protein yang bervariasi antara 38% hingga 49% (Saidu 2005) Menurut Liu (1997), protein kedelai mengandung asam amino essensial yang lengkap dengan methionin sebagai asam amino pembatas. Leusin, isoleusin, lisin dan valin merupakan asam amino yang paling tinggi yang terkandung di dalam kedelai. Kadar protein kedelai yang tinggi menjadikan tanaman ini memiliki kualitas yang sama dengan protein hewani.
Biji kedelai terdiri dari 7.3% kulit, 90.3% kotiledon dan 2.4% hipokotil. (Koswara 1992). Komposisi kimia bagian bji kedelai dapat dilihat pada Tabel 1.
4
Tabel 1. Komposisi kimia kedelai dan bagian-bagiannya (% BK)Bagian kedelai Protein (%) Lemak (%) Kadar abu (%) Karbohidrat (%)
Biji utuh 34.9 18.1 4.9 34.8
Kotiledon 42.8 22.8 5.0 29.4
Kulit 8.8 1.0 4.3 85.9
Hipokotil 40.0 11.4 4.4 43.4
Sumber : Kawamura (1977)
Selain mengandung protein, kacang kedelai juga memiliki kandungan lemak yang cukup tinggi. Kacang kedelai mengandung sekitar 18-20% lemak dan 85% dari jumlah tersebut terdiri dari asam lemak tak jenuh. Disamping itu, di dalam lemak kedelai terkandung beberapa fosfolipida yaitu lesitin, sepalin, dan lipositol (Koswara, 1992). Menurut Syarief dan Irawati (1988), lemak kedelai mengandung asam linoleat dan oleat, 10% palmitat, stearat dan palmitat (masing-masing 2%)
Menurut Smith dan Circle (1977), vitamin-vitamin yang terdapat dalam kacang kedelai antara lain vitamin A, vitamin B terutama niasin, riboflavin, dan tiamin, vitamin D, E dan K. Sedangkan mineral yang dikandungnya antara lain Ca, P, Fe, Na, K dan yang terdapat dalam jumlah kecil Mg, Mn, Za, Co, Cu, Se dan F.
Kandungan karbohidrat dalam kedelai sekitar 30% yang terdiri dari 15% karbohidrat tak dapat larut (insoluble carbohydrate) dan 15% karbohidrat yang dapat larut (soluble carbohydrate). Kedelai juga memiliki isoflavone dan zat anti-nutrisi seperti saponin, fosfolipid, protease inhibitor, fitat dan tripsin inhibitor (Saidu 2005). Selain itu kedelai mengandung senyawa penyebab off-flavor, yaitu glukosida, saponin, esterogen dan senyawa penyebab alergi (Koswara 1992).
2. Protein Kedelai
Komponen kimia tertinggi dalam kedelai adalah protein, yaitu antara 38% hingga 49% (Saidu, 2005). Sekitar 90% protein kedelai adalah globulin yang terdapat sebagai protein cadangan, sisanya merupakan enzim-enzim intraseluler (lipoksigenase, urease dan amilase), hemaglutinin, protein inhibitor dan lipoprotein membran (Kinsella 1979).
Globulin merupakan protein terpenting pada kedelai. Protein ini tidak larut dalam air di sekitar titik isoelektriknya tetapi akan segera larut dengan penambahan garam seperti natrium klorida atau kalsium klorida. Globulin larut dalam larutan garam encer pada pH di atas atau di bawah titik isoelektriknya (Pearson 1983). Pada pH sekitar 4.2-4.6, kelarutan protein kedelai mencapai minimum; kisaran pH tersebut dikenal sebagai titik isoelektrik protein kedelai (Berk 1992). Menurut Zayas (1997), kelarutan protein kedelai dalam air meningkat dengan meningkatnya pH dari 6 ke 8 dan suhu dari 10 oC sampai 70 oC. Ketika suhu meningkat, struktur protein terbuka (unfold) menjadi rantai lurus sehingga memungkinkan terjadinya peningkatan interaksi antara protein dan air dan kelarutan protein kedelai pun ikut meningkat.
Menurut Wolf dan Cowan (1975), protein kedelai terdiri dari campuran komponen-komponen yang mempunyai berat molekul 8 sampai 600 kilo Dalton. Melalui ultrasentrifugasi, protein kedelai dapat digolongkan menjadi empat golongan utama, yaitu protein 2S, 7S, 11S dan 15S (Tabel 2.). Protein kedelai juga dapat digolongkan ke dalam 4 fraksi berdasarkan kelarutannya, yaitu albumin (larut dalam air), globulin (larut dalam larutan garam), prolamin (larut dalam alkohol 70%) dan glutelin (larut dalam basa encer) (Belitz dan Grosch 1999).
5
Tabel 2. Komponen fraksi hasil ultrasentrifusa dari ekstrak protein kedelaiFraksi Persentase Komponen BM (kDa)
2S 22 Tripsin inhibitor 8 – 21.5 Sitokrom C 12 7S 37 Hemaglutinin 110 Lipoksigenase 102 β - amilase 61.7 7 S globulin 180 – 210 11S 31 11S globulin 350 15S 11 Polimer 600
Sumber : Wolf dan Cowan (1975)
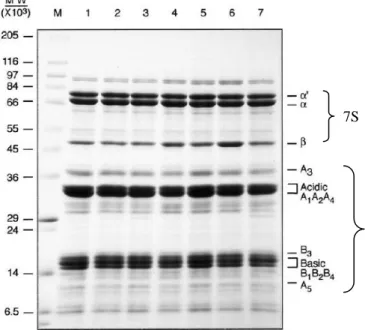
Globulin 7S dan 11S merupakan dua komponen utama protein cadangan biji kedelai. Kedua fraksi ini disebut sebagai protein cadangan karena tidak mempunyai aktivitas biologis kecuali sebagai asam amino cadangan untuk germinasi biji (Murphy 1985). Protein 7S dan 11S merupakan dua protein utama yang menyusun globulin dengan jumlah masing-masing sekitar 37% dan 31% dari total protein kedelai (Wolf dan Cowan 1975). Baik globulin 7S maupun globulin 11S terdiri atas subunit-subunit protein. Glisinin atau protein 11S tersusun atas polipeptida asam dan basa yang saling dihubungkan oleh ikatan disulfida. Sedangkan β-konglisinin atau protein 7S, merupakan protein dengan struktur trimer yang terdiri atas 3 tipe subunit (α‟, α dan β) (Liu et al. 2008). Identifikasi protein 7S dan 11S biasanya menggunakan elektroforesis, yang dapat menampilkan pita protein berdasarkan bobot molekulnya. Hasil publikasi Mujoo et al. (2003) mengenai profil protein tujuh varietas kedelai (SDS-PAGE) dapat dilihat pada Gambar 2.
Gambar 2. Profil protein tujuh varietas kedelai (SDS-PAGE) 1: Vinton-81; 2: S-20F8; 3: HP-204; 4: IA-2034;
5: Steyer; 6: IA-2020; 7: S-2020; M : marker (Mujoo et al. 2003). 7S
6
a. Fraksi 11 S (Glisinin)
Glisinin merupakan protein heksamer (AB)6 dengan berat molekul berkisar 300-380 kDa. Subunit-subunit glisinin terdiri atas polipeptida asam (A) dan polipeptida basa (B) yang dihubungkan oleh ikatan disulfida (Blazek 2008). Polipeptida asam glisinin memiliki berat molekul sekitar 35 kDa, sedangkan polipetida basanya memiliki berat molekul sekitar 20 kDa (Mujoo et al. 2003). Glisinin mengandung paling sedikit 20 ikatan disulfida dan dua grup sulfhidril per mol protein (Draper dan Catsimpoolas 1978 dikutip oleh Nakamura et al. 1984). Ikatan disulfida ini dapat menyebabkan glisinin lebih tahan terhadap denaturasi. Struktur kuarterner glisinin stabil karena adanya interaksi elektrostatik dan hidrofobik serta ikatan disulfida (Badley et al. 1975; Peng et al. 1984). Skematik struktur molekuler glisinin dapat dilihat pada Gambar 3.
Gambar 3. Skematik struktur molekuler glisinin (monomer, trimer, dan heksamer); A: polipeptida asam; B : polipeptida basa; : ikatan disulfida
(Renkema et al. 2001).
b. Fraksi 7 S (β-konglisinin)
Struktur kuarterner globulin 7S tersusun atas oleh 3 subunit yaitu, α‟, α dan β yang dihubungkan melalui interaksi hidrofobik dengan berat molekul sekitar 180 kDa (Kinsella 1985). Subunit α‟ memiliki berat molekul sekitar 72 kDa, sedangkan α dan β memiliki berat molekul masing-masing sekitar 68 dan 52 kDa (Mujoo et al. 2003). Kombinasi subunit-subunit tersebut memberikan berat molekul sekitar 180 kDa tergantung dari subunit-subunit penyusunnya (Blazek 2008). Fraksi 7S merupakan glikoprotein yang tidak mengandung grup sufhidril dan kandungan asam amino sulfurnya sedikit sekali (Nielson 1985 dikutip oleh Yu dan Damodaran 1991). Menurut Lewis dan Chen (1978) β-konglisinin merupakan glikoprotein yang mengandung 3.8-5.4% karbohidrat. Jenis gula yang terdapat dalam protein ini adalah manosa dan glukosamin. Struktur molekuler β-konglisinin dapat dilihat pada Gambar 4.
Gambar 4. Struktur molekuler β-konglisinin (Renkema et al. 2001)
7
B. GELASI Vs KOAGULASI
1. Gelasi Protein
Menurut Liu et al. (2008), protein kedelai memiliki banyak sifat fungsional yang telah dipelajari dengan sangat luas. Sifat fungsional itu diantaranya adalah kemampuan larut, kemudahan terdenaturasi oleh panas, kemampuan membentuk gel, emulsifier, kemampuan membentuk busa, kemampuan mengikat air (water holding capacity(WHC)), pembentuk karakteristik struktur, sifat reologi dan kemampuan membentuk tekstur.
Pada umumnya, gelasi protein merupakan tahapan yang penting dalam menghasilkan produk pangan dengan mutu tekstur yang baik. Karakteristik mutu suatu produk pangan, khususnya sifat tekstur dan juiciness, ditentukan melalui kapasitas gelasi protein. Gel dapat bervariasi dalam hal sifat reologinya yaitu kekerasan, kelengketan, kohesivitas, dan adesivitas. Dalam hal ini, protein sering digunakan untuk menghasilkan sifat tekstur tertentu melalui fenomena gelasi protein.
Sifat gelasi protein berhubungan dengan agregasi protein. Gelasi protein terjadi ketika protein beragregasi membentuk jaringan (Tay et al. 2005). Menurut Schmidt (1981) yang dikutip oleh Zayas (1997), gelasi protein adalah fenomena agregasi protein di mana interaksi polimer-polimer dan polimer-solven setimbang sehingga jaringan atau matriks tersier terbentuk. Agregasi protein sendiri menurut Tay et al. (2005) dapat terjadi melalui proses pemanasan, pengaturan pH atau pengaturan kekuatan ionik dalam larutan protein.
Gel terbentuk ketika protein yang strukturnya terbuka sebagian (unfold) terurai menjadi segmen-segmen polipeptida yang kemudian berinteraksi pada titik tertentu untuk membentuk jaringan ikatan silang tiga dimensi. Protein dengan struktur unfold, dimana struktur sekundernya mengalami perubahan, diperlukan pada proses gelasi protein. Perubahan ini dapat terjadi melalui perlakuan panas, asam, alkali dan urea (Zayas 1997).
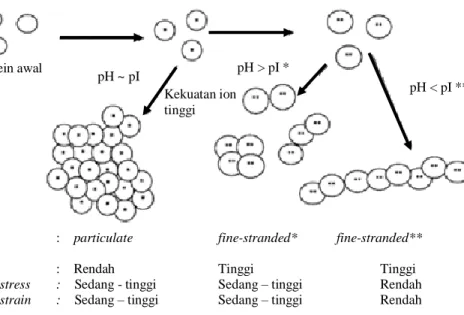
Menurut Zayas (1997), pada proses pembentukan gel, transisi dari bentuk alami menjadi bentuk terdenaturasi merupakan prekursor penting dalam interaksi protein-protein. Jaringan gel baru akan terbentuk setelah sebagian protein mengalami denaturasi. Pembentukan gel protein merupakan hasil dari ikatan hidrogen, interaksi ionik dan hidrofobik, ikatan Van der Waals, dan ikatan kovalen disulfida. Stuktur dan karakteristk fisik gel pada beberapa pH dapat dilihat pada Gambar 5.
Gambar 5. Struktur dan karakteristik fisik gel pada beberapa pH ( Foegeding 2005 ) Protein awal pH < pI ** Kekuatan ion tinggi pH > pI * pH ~ pI
Tipe Gel : particulate fine-stranded* fine-stranded**
WHC : Rendah Tinggi Tinggi
Fracture stress : Sedang - tinggi Sedang – tinggi Rendah
8
Gel yang berbentuk fine-stranded memiliki penampakan gel yang transparan. Gel tipe ini terbentuk saat kondisi pH diatas atau dibawah pI dan saat kekuatan ioniknya rendah. Gel yang terbentuk saat pH diatas maupun dibawah pI memiliki karakteristik fisik yang berbeda. Gel yang terbentuk pada pH yang rendah memiliki karakteristik fisik yang lemah dan rapuh, sedangkan gel yang terbentuk pada pH yang tinggi memiliki karakter fisik yang kuat dan elastis. Tipe gel yang kedua yaitu particulate yang memiliki penampakan buram. Gel berbentuk particulate terbentuk saat minimumnya gaya tolak menolak seperti saat pH mendekati pI atau saat kekuatan ioniknya tinggi. Gel ini memiliki ukuran partikel yang besar dan WHC yang rendah sehingga teksturnya lebih kuat dibandingkan gel fine-stranded yang terbentuk pada pH tinggi,Sifat gelasi protein kedelai sering dihubungkan dengan keberadaan protein 7S dan 11S yang merupakan penyusun utama protein globulin kedelai. Kandungan protein 11S dan rasio 11S/7S dilaporkan memberikan korelasi positif terhadap kekerasan gel dari protein kedelai (Mujoo et al. 2003). Nakamura et al. (1984) yang dikutip oleh Yoshida et al. (1992) melaporkan bahwa kekerasan gel dari globulin 11S berbeda-beda antara varietas yang berbeda pada konsentrasi globulin yang sama. Mereka juga menunjukkan bahwa kekerasan gel meningkat sebanding dengan kandungan dari suatu subunit asam yang berberat molekul tinggi dalam total globulin 11S.
Telah diketahui bahwa fraksi dengan berat molekul yang tinggi dalam suatu polimer akan membentuk gel dengan kekerasan yang lebih tinggi dibandingkan fraksi dengan berat molekul yang lebih rendah (Watase dan Nishinari 1983 yang dikutip oleh Yoshida et al. 1992).
Menurut Corredig (2006), gel yang diperoleh dari isolasi glisinin (11S) memberikan karakter gel yang lebih keras dibandingkan gel yang diperoleh dari β-konglisinin (7S), dan struktur jaringan yang terbentuk memiliki perbedaan antar keduanya, tergantung dari komposisi protein. Blazek (2008) melaporkan bahwa rasio 11S/7S mempengaruhi karakter kekerasan dan elastisitas gel. Glisinin berkontribusi terhadap peningkatan kekerasan dan kekokohan gel, sedangkan β-konglisinin memberikan pengaruh terhadap elastisitas gel yang dihasilkan.
Gel dari globulin 11S yang dibuat dengan penambahan koagulan CaSO4 lebih keras dibandingkan gel dari globulin 7S. Fenomena tersebut berhubungan dengan kandungan ikatan disulfida yang lebih banyak pada globulin 11S. Hal serupa terjadi pada gel yang dibuat dengan penambahan koagulan Glucono δ Lactone (GDL) (Hashizume et al. 1975 yang dikutip oleh Kohyama dan Nishinari 1993).
2. Koagulasi Protein
Koagulasi didefinisikan sebagai interaksi acak molekul-molekul protein yang menyebabkan pembentukan agregat-agregat protein baik bersifat larut ataupun tidak larut (Meng et al. 2002). Koagulasi dapat terjadi melalui penambahan bahan penggumpal protein (koagulan).
Koagulasi susu kedelai merupakan langkah yang paling penting dalam proses pembuatan curd sekaligus menjadi tahapan paling sulit untuk dikendalikan karena merupakan hasil interaksi yang kompleks dari berbagai variabel (Prabhakaran et al. 2006; Blazek 2008). Penggunaan jenis maupun konsentrasi koagulan yang berbeda akan mempengaruhi rendemen, sifat tekstur dan flavor curd yang berbeda pula (Blazek 2008; Mujoo 2003). Menurut Obatolu (2007), proses koagulasi susu kedelai dipengaruhi oleh interaksi kompleks antara jenis kedelai, suhu pemasakan susu kedelai, volume, kandungan padatan, pH, jenis dan jumlah koagulan serta waktu koagulasi.
Menurut Blazek (2008), kurangnya jumlah koagulan yang digunakan untuk koagulasi akan menyebabkan pengendapan protein menjadi tidak sempurna serta menyulitkan proses pemisahan whey dan curd. Jumlah koagulan yang kurang juga akan menghasilkan pembentukan struktur matriks curd yang renggang karena tidak sempurnanya pengendapan, akibatnya curd yang terbentuk terlalu lunak
9
(Obatolu 2007). Sebaliknya, kelebihan jumlah koagulan akan membuat tekstur curd kedelai menjadi keras dan mengurangi palatabilitas.Perbedaan jenis koagulan yang digunakan akan menghasilkan perbedaan kandungan air di dalam curd. Hal ini disebabkan karena pembentukan struktur jaringan gel oleh koagulan dipengaruhi oleh perbedaan kekuatan anion dan kation terhadap kemampuan pengikatan air (WHC) dalam gel protein kedelai. Oleh karena itu, konsentrasi koagulan dan jenis anion ini mempengaruhi kekerasan
curd yang dihasilkan (Prabhakaran 2006).
Rendemen pembentukan curd juga dipengaruhi oleh penggunaan koagulan. Semakin lambat kemampuan koagulan dalam mengkoagulasi susu akan memberikan rendemen curd yang lebih baik karena agregat protein akan memerangkap air lebih banyak di dalam curd. Sebaliknya, koagulan yang mengkoagulasikan protein lebih cepat, kurang memerangkap air sehingga curd yang dihasilkan lebih sedikit (Obatolu 2007). Peningkatan temperatur koagulasi dan kecepatan pengadukan sesaat setelah penambahan koagulan juga akan menurunkan rendemen curd dan mempengaruhi kekerasan curd yang terbentuk (Blazek 2008).
Menurut Shurtleff dan Aoyagi (1984), bahan penggumpal protein kedelai dalam pembuatan tahu dapat digolongkan menjadi beberapa golongan, yaitu : 1) golongan garam klorida atau nigari; 2) golongan garam sulfat; 3) golongan lakton; dan 4) golongan asam. Beberapa contoh koagulan penggumpal protein kedelai dapat dilihat pada Tabel 3.
Tabel 3. Beberapa golongan bahan penggumpal (koagulan) protein kedelai
Golongan Contoh yang umum dipakai
Garam klorida (nigari) MgCl2.6H2O, air laut, CaCl2, CaCl2.2H2O
Garam sulfat CaSO4. 2H2O, MgSO4.7H2O
Lakton C6H10O6 (glukono-δ-lakton)
Asam Asam laktat, asam asetat, sari buah jeruk
Sumber : Shurtleff dan Aoyagi (1984)
Nigari alami diekstrak dari air laut dengan menghilangkan sebagian besar garam (NaCl) dan
air. Koagulan jenis ini mengandung komponen mineral air laut alami terutama magnesium klorida. Penggunaan koagulan jenis nigari membutuhkan waktu pembuatan tahu yang cukup lama karena koagulan jenis ini harus ditambahkan sedikit demi sedikit dan perlahan-lahan, akibatnya dibutuhkan teknik yang baik dalam pembuatan tahu. Selain itu, penggunaan koagulan nigari akan menghasilkan tahu dengan tekstur yang cenderung kurang lembut (Shurtleff dan Aoyagi, 1984).
Garam sulfat merupakan golongan koagulan yang paling banyak digunakan dalam pembuatan
curd protein kedelai (Shurtleff dan Aoyagi, 1979). Koagulan ini akan terdispersi perlahan di dalam
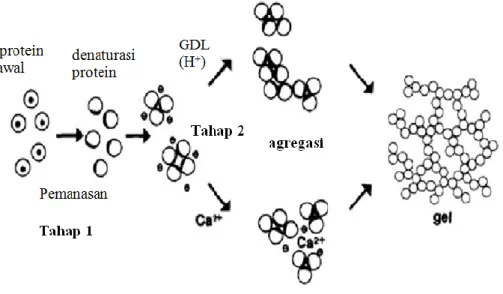
susu kedelai sehingga memberikan waktu koagulasi yang lambat (Shurtleff dan Aoyogi, 1984). Koagulan sulfat mengkoagulasi protein kedelai dengan cara membentuk jembatan antar molekul protein dan meningkatkan ikatan silang polimer sehingga terjadi agregasi protein (Obatolu, 2007). Pemakaian GDL sebagai koagulan akan menurunkan pH susu kedelai dan menyebabkan agregasi dari protein terdenaturasi dengan meningkatkan sifat hidrofobik dan ketidaklarutan (Kohyama dan Nishinari, 1993). Ilustrasi mekanisme gelasi dengan koagulan kalsium sulfat dan GDL dapat dilihat pada Gambar 6
.
Pengendapan menggunakan koagulan asam akan menurunkan pH sistem dan memungkinkan agregasi protein terjadi (Obatolu, 2007). Melalui proses pemanasan susu kedelai, sebagai prasyarat terbentuknya gel, struktur molekul protein kedelai akan terbuka (unfold), akibatnya ikatan hidrogen (-SH), ikatan disulfida (S-S), dan sisi rantai asam amino hidrofobik akan terekspos. Selanjutnya,
10
dengan penambahan koagulan, misalnya koagulan asam, muatan negatif molekul protein akan berkurang akibat protonasi COO- pada residu asam amino. Sebagai akibatnya, molekul-molekul protein akan cenderung saling mendekat karena memiliki muatan yang sama. Keadaan ini membuat ikatan hidrogen (-SH), ikatan disulfida (S-S) serta interaksi hidrofobik terjadi secara intermolekul. Reaksi ini memfasilitasi terjadinya agregasi protein membentuk struktur jaringan tiga dimensi gelcurd (Liu et al 2004).
Gambar 6. Mekanisme gelasi protein dengan koagulan kalsium sulfat dan GDL (Kohyama et al., 1995)
Bahan penggumpal lainnya yang biasa digunakan dalam pembuatan tahu secara tradisional adalah whey tahu. Whey tahu ini merupakan hasil pengepresan yang didiamkan semalam pada suhu kamar. Whey tersebut akan mengalami fermentasi oleh bakteri asam laktat yang dapat menggumpalkan protein kedelai menjadi tahu (Subardjo et al 1987).
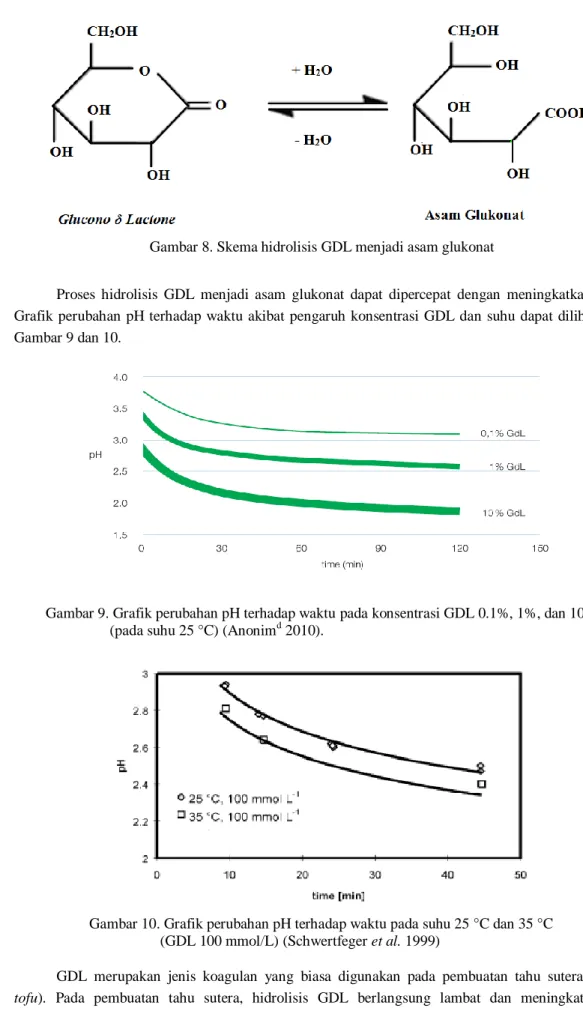
Glucono δ Lactone (GDL) adalah ester siklik netral asam glukonat yang berbentuk serbuk
kristal putih. Asam glukonat diproduksi oleh fermentasi aerobik dari sumber karbohidrat. Setelah fermentasi, asam glukonat dimurnikan dan mengkristal menjadi GDL. Struktur GDL dapat dilihat pada Gambar 7.
Gambar 7. Struktur Glucono δ Lactone (D-Gluconic acid δ lactone)
Saat dilarutkan, GDL dapat larut dengan cepat dan terhidrolisis menjadi asam glukonat. Gugus karbonil pada asam glukonat yang terbentuk cenderung tidak stabil dan membentuk COO- dan H+, adanya H+ ini lah yang menyebabkan penurunan pH lingkungan. Skema hidrolisis GDL menjadi asam glukonat dapat dilihat pada Gambar 8.
α
β
γ
11
Gambar 8. Skema hidrolisis GDL menjadi asam glukonatProses hidrolisis GDL menjadi asam glukonat dapat dipercepat dengan meningkatkan suhu. Grafik perubahan pH terhadap waktu akibat pengaruh konsentrasi GDL dan suhu dapat dilihat pada Gambar 9 dan 10.
Gambar 9. Grafik perubahan pH terhadap waktu pada konsentrasi GDL 0.1%, 1%, dan 10% (pada suhu 25 °C) (Anonimd 2010).
Gambar 10. Grafik perubahan pH terhadap waktu pada suhu 25 °C dan 35 °C (GDL 100 mmol/L) (Schwertfeger et al. 1999)
GDL merupakan jenis koagulan yang biasa digunakan pada pembuatan tahu sutera (silken
12
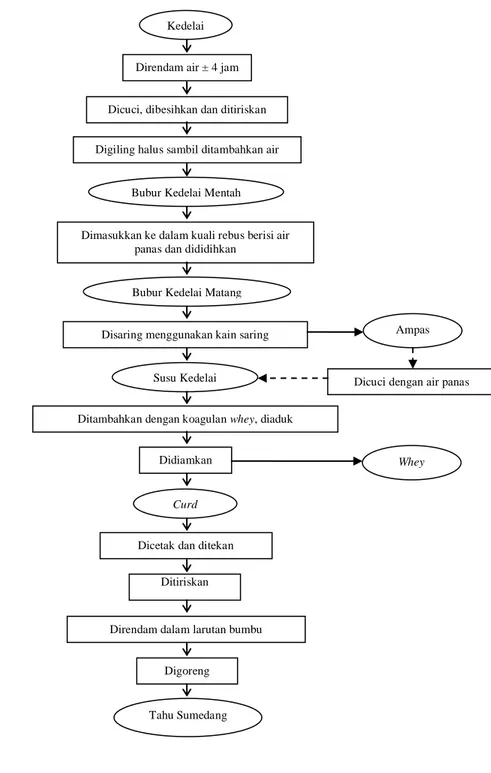
Dicuci, dibesihkan dan ditiriskan Direndam air ± 4 jam
Digiling halus sambil ditambahkan air
Dimasukkan ke dalam kuali rebus berisi air panas dan dididihkan
Disaring menggunakan kain saring
Dicuci dengan air panas
Ditambahkan dengan koagulan whey, diaduk
Didiamkan
Dicetak dan ditekan Ditiriskan
Direndam dalam larutan bumbu
Digoreng
Tahu Sumedang Kedelai
Bubur Kedelai Matang
Susu Kedelai
Ampas
Whey
Curd
Bubur Kedelai Mentah
meningkatnya suhu inkubasi. Walaupun mekanisme koagulasi karena adanya penurunan pH, proses koagulasi yang lambat menyebabkan curd yang dihasilkan memiliki tekstur yang lebih halus dibandingkan curd yang dihasilkan dengan menggunakan koagulan jenis asam.
Proses pembuatan tahu sutera cukup berbeda dengan proses pembuatan tahu press. Perbedaan proses tersebut pula yang menyebabkan perbedaan karakteristik tahu yang dihasilkan. Skema pembuatan tahu press dan tahu sutera dapat dilihat pada Gambar 11 dan 12.
Gambar 11. Skema pembuatan tahu Sumedang (tahu press)di pabrik „Diazara Tresna‟ (Fahmi 2010)
13
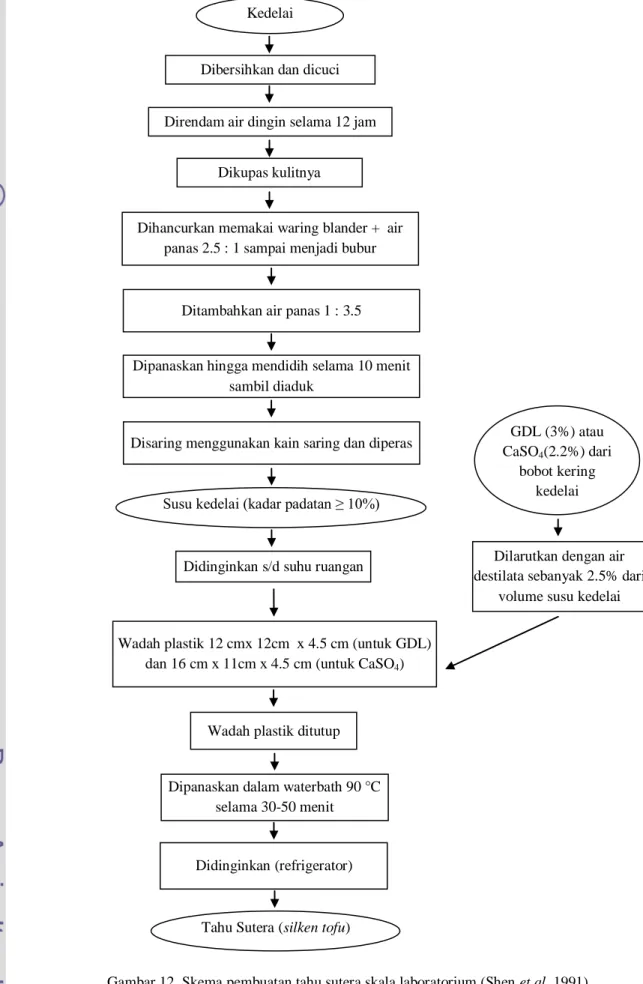
Gambar 12. Skema pembuatan tahu sutera skala laboratorium (Shen et al. 1991)Dilarutkan dengan air destilata sebanyak 2.5% dari
volume susu kedelai Kedelai
Dibersihkan dan dicuci
Direndam air dingin selama 12 jam
Ditambahkan air panas 1 : 3.5
Dipanaskan hingga mendidih selama 10 menit sambil diaduk
Disaring menggunakan kain saring dan diperas
Susu kedelai (kadar padatan ≥ 10%) Dikupas kulitnya
Dihancurkan memakai waring blander + air panas 2.5 : 1 sampai menjadi bubur
Didinginkan s/d suhu ruangan
GDL (3%) atau CaSO4(2.2%) dari
bobot kering kedelai
Wadah plastik 12 cmx 12cm x 4.5 cm (untuk GDL) dan 16 cm x 11cm x 4.5 cm (untuk CaSO4)
Wadah plastik ditutup
Dipanaskan dalam waterbath 90 °C selama 30-50 menit
Didinginkan (refrigerator)
14
3. Curd Kedelai
Curd merupakan hasil penggumpalan protein dalam larutan susu. Gel dari protein kedelai ini,
atau yang dikenal sebagai curd, memiliki kemampuan untuk membentuk matriks yang mampu menahan air, lemak, polisakarida, flavor dan komponen lainnya (Zayas 1997). Secara konvensional
curd yang berasal dari susu kedelai ini dikenal sebagai tahu. Tahu adalah endapan protein yang dibuat
dengan cara mengendapkan susu kedelai dengan koagulan sehingga dihasilkan endapan yang kompak, putih dan bertekstur lembut (Watanabe 1974).
Tahapan pembuatan tahu terdiri atas dua tahap utama, yaitu pembuatan susu kedelai dan tahap koagulasi (penggumpalan) susu kedelai sehingga terbentuk curd yang selanjutnya dipress membentuk tahu (Shurtleff dan Aoyagi 1984). Kedelai yang akan dibuat susu terlebih dahulu direndam dalam air bersih (dengan perbandingan tertentu dengan berat kacang kedelai) dengan tujuan untuk melunakkan struktur seluler kedelai sehingga mempermudah dan mempercepat penggilingan serta menghasilkan ekstrak optimum. Lamanya perendaman perlu diperhatikan, karena perendaman yang terlalu singkat akan membuat biji kedelai sulit pecah ketika penggilingan, sedangkan bila terlalu lama akan terjadi pembentukan busa pada permukaan air rendaman akibat fermentasi kedelai (Subardjo et al. 1987).
Kacang kedelai yang telah direndam kemudian digiling dengan penambahan air sehingga dihasilkan bubur kedelai. Tujuan penggilingan ini adalah untuk memperkecil ukuran-ukuran partikel sehingga meningkatkan efektivitas ekstraksi protein kedelai selama pemasakan (Shurtleff dan Aoyagi 1984).
Bubur kedelai hasil penggilingan harus segera dimasak secepatnya. Penundaan pemasakan sampai 30 menit saja dapat menurunkan rendeman tahu (Subardjo et al. 1987). Menurut Supriatna (2005), untuk menghasilkan sari kedelai yang optimal dari segi kualitas dan kuantitasnya, bubur kedelai terlebih dahulu dimasak sebelum akhirnya disaring. Menurut Liu et al.(2004), pemanasan optimal dalam pembuatan susu kedelai dilakukan selama 3-10 menit setelah mendidih yang tujuannya untuk mengekstrak protein kedelai dan mendenaturasi protein serta memudahkan proses koagulasi. Fungsi lain dari pemanasan dalam pembuatan susu kedelai adalah mengurangi bau langu, menginaktifasi antitripsin, meningkatkan daya cerna dan menambah daya awet produk (Koswara 1992). Selama pemasakan perlu dilakukan pengadukan secara kontinyu agar tidak terjadi kegosongan. Pemasakan yang terlalu lama perlu dihindari karena selain menurunkan nlai gizi dan rasa tahu, tekstur tahu yang diperoleh pun menjadi kurang kompak dan tahu berwarna agak kecokelatan (Subardjo et al. 1987).
Bubur kedelai kemudian disaring, penyaring yang biasa digunakan dipabrik tahu adalah penyaring berbahan kain blacu berwarna putih. Hasil dari penyaringan ini adalah susu kedelai. Proses selanjutnya adalah penggumpalan protein susu kedelai dengan penambahan koagulan. Menurut Shurtleff dan Aoyogi (1986), penambahan bahan penggumpal sebaiknya dilakukan setelah susu kedelai mencapai suhu 70-90 oC, hal ini tergantung dari jenis bahan penggumpal yang digunakan. Gumpalan protein yang terbentuk kemudian dipress dan dicetak. Menurut Shurtleff dan Aoyagi (1984), untuk mendapatkan hasil yang baik pengepressan dilakukan pada tekanan sebesar 0.15-0.21 psi selama 15-20 menit.
Obatolu (2007) melaporkan bahwa perbedaan karakteristik tekstur, khususnya kekerasan, dapat dihubungkan dengan kandungan air di dalam tahu. Tahu dengan kekerasan tinggi memiliki kemampuan menahan air (WHC) yang rendah. Hal ini disebabkan oleh curd yang terbentuk lebih rapat sehingga kemampuannya dalam menahan air berkurang. Sebaliknya tahu yang lunak memiliki matriks yang renggang sehingga air dapat terperangkap dalam jumlah yang lebih banyak. Tahu yang lunak memiliki kandungan air yang tinggi yaitu antara 84 hingga 90%. Tahu dengan kandungan air
15
yang tinggi secara visual akan memberikan penampakan yang lembut sedangkan tahu dengan kandungan air yang rendah cenderung memiliki penampakan yang kasar.C.
TEKSTUR
Tekstur merupakan aspek penting dalam penilaian mutu produk pangan oleh konsumen selain penampakan dan flavor. Menurut Smith (2004), tekstur menjadi faktor kunci penerimaan konsumen atas produk pangan. Bourne (2002), yang dikutip oleh Smith (2004), mendefinisikan sifat tekstur produk pangan sebagai sekelompok karakteristik fisik yang: (1) diperoleh dari elemen struktural produk pangan, (2) dipersepsikan oleh indera peraba, (3) berhubungan dengan deformasi, disintegrasi, dan gaya yang diberikan serta (4) diukur secara obyektif sebagai fungsi dari massa, waktu dan jarak.
Persepsi manusia terhadap tekstur tidak hanya ditentukan ketika produk pangan berada di dalam mulut. Faktor lain seperti penampakan dan pengaruh indera pendengaran juga memberikan persepsi tentang tekstur suatu produk (Kilcast 2004). Persepsi tekstur yang diterima oleh manusia melalui indera peraba dapat diperoleh melalui dua cara, yaitu: somesthesis (secara taktil) yang merupakan respon yang diperoleh manusia melalui sentuhan dari kulit, dan kinesthesis yaitu respon yang diterima melalui aktivitas otot dan tendon. Stimulus sentuhan (somesthesis)dapat dilakukan melalui pengujian produk pangan menggunakan tangan dan jari sedangkan kontak oral (kinesthesis) diperoleh melalui pengujian di dalam mulut akibat aktivitas bibir, lidah, langit-langit mulut dan gigi (Kilcast 1999).
Analisis tekstur produk pangan dapat dilakukan secara organoleptik menggunakan indera manusia ataupun secara instrumen menggunakan alat. Analisis tekstur secara organoleptik memberikan hasil yang subyektif dan beragam, tergantung pada penilaian yang diberikan oleh panelis dalam pengujian. Sebaliknya, analisis secara instrumen akan memberikan hasil yang lebih akurat karena bersifat obyektif (Peleg 1983). Menurut Smewing (1999), analisis tekstur dapat dilakukan menggunakan alat atau instrumen seperti Instron, LFRA Texture Analyser, dan Stable Micro System TA-XT2i Texture Analyser. Umumnya, karakteristik tekstur curd secara obyektif dianalisis menggunakan instrumen texture analyser TA-XT2i dengan metode Texture Profile Analysis (TPA).
Menurut Szczesniac (1987) yang dikutip oleh Faridi dan Faubion (1990), tekstur merupakan atribut sensori yang hanya dipersepsikan, dijelaskan dan diukur dengan indera manusia seperi peraba, penglihatan dan pendengaran. Oleh karena itu, perlu dilakukan analisis sensori terhadap tekstur curd tersebut. Analisis sensori terhadap curd, termasuk dalam penilaian deskriptif yang membutuhkan panelis terlatih dalam bidang mutu tekstur curd. Menurut Kemp et al. (2009), analisis deskriptif hanya membutuhkan 6-18 panelis terlatih dengan kemampuan sensori yang baik dan telah menerima pelatihan. Kem et al. (2009) menyebutkan bahwa tujuan pelatihan panelis tidak hanya meningkatkan kemampuan panelis dalam mendeteksi, membedakan dan mendeskripsikan sampel, melainkan juga meningkatkan kepercayaan diri dan mengurangi ragam antar penelis.
Texture Profile Analysis (TPA) merupakan bentuk penilaian obyektif dari analisis tekstur
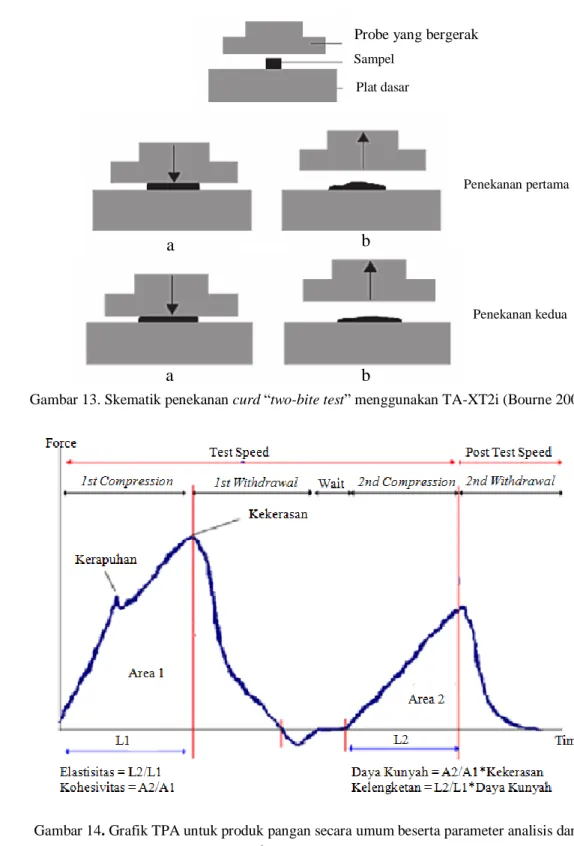
secara sensori. Pada TPA, probe akan melakukan kompresi sebanyak dua kali terhadap sampel. Hal ini dapat dianalogikan sebagai gerakan mulut pada saat mengunyah atau menggigit makanan (Larmond 1976). Oleh karena itu, TPA disebut juga sebagai “two-bite test”. Skematik penekanan curd “two-bite test” menggunakan TA-XT2i dapat dilihat pada gambar 13.
Larmond (1976), menyatakan bahwa analisis menggunakan TPA merupakan analisis yang
multipoint karena hanya dengan sekali analisis akan diperoleh nilai dari beberapa parameter tekstur.
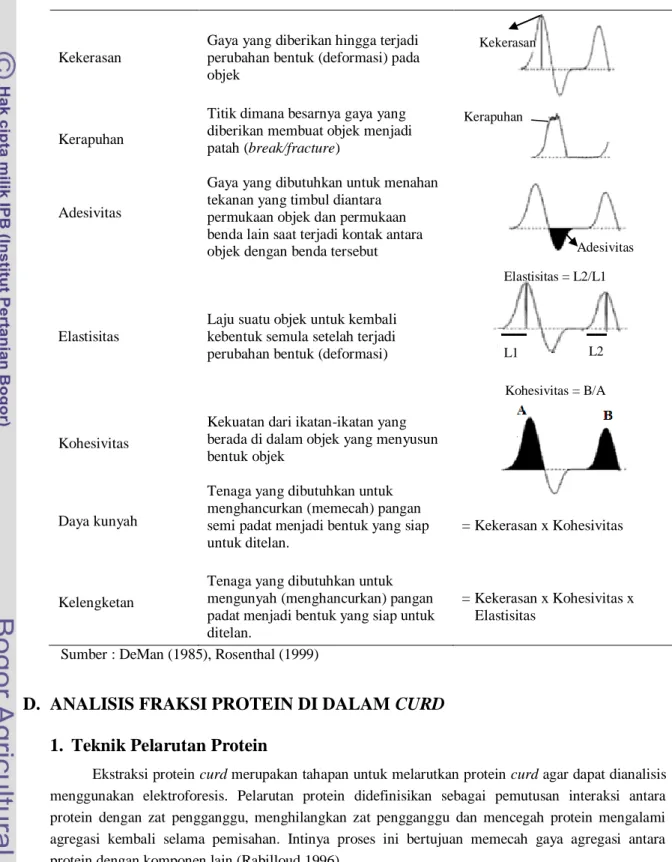
Parameter tekstur yang dapat diukur menggunakan TPA meliputi kekerasan, kerapuhan, elastisitas, kohesivitas, adesivitas, daya kunyah dan kelengketan. Grafik hasil pengukuran tekstur pangan secara umum dengan metode TPA dan perhitungan parameter mekanik dapat dilihat pada Gambar 14.
16
Gambar 13. Skematik penekanan curd “two-bite test” menggunakan TA-XT2i (Bourne 2002)Gambar 14. Grafik TPA untuk produk pangan secara umum beserta parameter analisis dan perhitungannya (Anonimc, 2010)
Szczesniak (1963) yang dikutip Faridi dan Faubion (1990) menyatakan bahwa parameter-parameter tekstur yang digunakan untuk mengklasifikasikan atribut tekstur secara sensori terdiri atas tiga kategori, diantaranya: 1) karakteristik mekanikal, yaitu reaksi bahan pangan terhadap tekanan yang dipersepsikan oleh indra kinestetik, meliputi kekerasan, kohesivitas, viskositas dan kerenyahan; 2) karakteristik geometrikal, yaitu karakteristik yang berhubungan dengan ukuran, bentuk dan orientasi partikel yang dipersepsikan oleh syaraf pengecap dalam mulut atau dengan sentuhan meliputi
a
a
b
b
Penekanan pertama Penekanan kedua Probe yang bergerakSampel Plat dasar
17
gritty, grainy, flaky, stringy, dan smooth; dan 3) karakteristik lain, meliputi atribut mouthfeel yang
berhubungan dengan persepsi terhadap lemak dan air selama pengunyahan dan penelanan. Beberapa karakteristik mekanikal dan definisinya dapat dilihat pada Tabel 4.
Tabel 4. Beberapa karakteristik mekanikal dan definisi dari grafik TPA
Karakteristik Definisi Sensorial Definisi Instrumental
Kekerasan
Gaya yang diberikan hingga terjadi perubahan bentuk (deformasi) pada objek
Kerapuhan
Titik dimana besarnya gaya yang diberikan membuat objek menjadi patah (break/fracture)
Adesivitas
Gaya yang dibutuhkan untuk menahan tekanan yang timbul diantara
permukaan objek dan permukaan benda lain saat terjadi kontak antara objek dengan benda tersebut
Elastisitas
Laju suatu objek untuk kembali kebentuk semula setelah terjadi perubahan bentuk (deformasi)
Kohesivitas
Kekuatan dari ikatan-ikatan yang berada di dalam objek yang menyusun bentuk objek
Daya kunyah
Tenaga yang dibutuhkan untuk menghancurkan (memecah) pangan semi padat menjadi bentuk yang siap untuk ditelan.
= Kekerasan x Kohesivitas
Kelengketan
Tenaga yang dibutuhkan untuk mengunyah (menghancurkan) pangan padat menjadi bentuk yang siap untuk ditelan.
= Kekerasan x Kohesivitas x Elastisitas
Sumber : DeMan (1985), Rosenthal (1999)
D. ANALISIS FRAKSI PROTEIN DI DALAM CURD
1. Teknik Pelarutan Protein
Ekstraksi protein curd merupakan tahapan untuk melarutkan protein curd agar dapat dianalisis menggunakan elektroforesis. Pelarutan protein didefinisikan sebagai pemutusan interaksi antara protein dengan zat pengganggu, menghilangkan zat pengganggu dan mencegah protein mengalami agregasi kembali selama pemisahan. Intinya proses ini bertujuan memecah gaya agregasi antara protein dengan komponen lain (Rabilloud 1996).
Kekerasan L1 Kerapuhan L2 Elastisitas = L2/L1 Kohesivitas = B/A Adesivitas
18
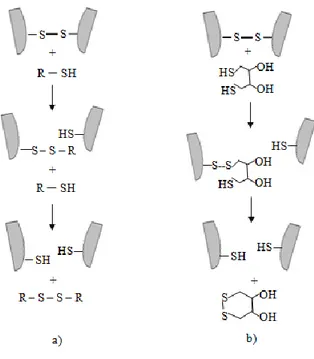
Umumnya rantai polipeptida tidak terikat dalam kompleks biologis oleh ikatan kovalen, kecuali ikatan disulfida, dan ikatan yang dibentuk oleh transglutaminase (TAG). TAG menginisiasi terbentuknya ikatan amida yang secara kimia tak dapat dibedakan dari ikatan peptida. Ikatan ini tidak dapat dipecah selain dengan mendegradasi protein tersebut menjadi bentuk asam aminonya. Sementara itu, ikatan disulfida merupakan ikatan kovalen yang dapat dengan mudah dirusak tanpa harus mendegradasi protein. Sebagai ikatan kovalen, ikatan disulfida dapat diputus secara kimiawi, namun reagen perusak harus dapat masuk dan kontak dengan ikatan disulfida tersebut. Sementara itu, ikatan disulfida terletak di dalam struktur protein sehingga dibutuhkan proses denaturasi untuk membuka sebagian struktur protein (unfold) dan reagen perusak pun dapat menyerang ikatan disulfida. Reagen perusak yang dapat memutus ikatan disulfida, antara lain merkaptoetanol, tiogliserol, sisteamin, ditiotreitol (DTT), atau ditioeritritol (DTE) (Rabilloud 1996).Prinsip dasar memutus ikatan disulfida adalah dengan menambahkan tiol bebas secara berlebih. Alkil tiol seperti 2-mercaptoethanol biasanya digunakan pada konsentrasi yang cukup tinggi (0.2 M) untuk memastikan kecukupan penggantian tiol yang berasal dari protein (Rabilloud 1996). Mekanisme pemutusan ikatan disulfida dapat dilihat pada Gambar 15.
Gambar 15. Mekanisme pemutusan ikatan disulfida dengan tiol bebas, a) reduksi dengan monotiol bebas (2-mercaptoethanol) dan b) reduksi dengan siklik ditiol (DTT) (Rabilloud 1996)
2. Teknik Elektroforesis Dalam Analisis Protein
Elektroforesis didefinisikan sebagai migrasi molekul atau partikel bermuatan di dalam larutan atau medium melalui pengaruh medan listrik (Nielsen 2003). Migrasi partikel bermuatan tersebut dapat terjadi karena perbedaan muatan total, ukuran dan bentuk partikel (Pomeranz dan Meloan 1994). Metode analisis elektroforesis protein merupakan metode analisis yang memisahkan molekul protein berdasarkan berat molekulnya (Bolag dan Edelstein 1991).
Teknik elektroforesis telah banyak digunakan dalam analisis protein untuk menentukan tingkat kemurnian sampel, berat molekul maupun titik isoelektrik (Copeland 1994), untuk menentukan komposisi protein dari suatu produk pangan (Nielsen 2003).
19
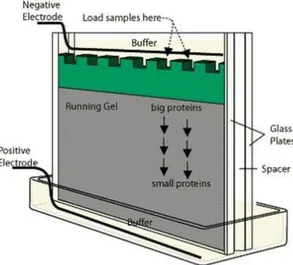
Pemisahan protein berdasarkan muatannya tergantung pada karakter asam dan basa protein. Hal ini ditentukan oleh jumlah dan jenis rantai samping (gugus R) yang dapat terionisasi dalam rantai polipeptida serta pH lingkungan. Pada pH lingkungan yang lebih besar daripada pH isoelektriknya (pI), protein akan memiliki muatan negatif sehingga migrasi protein akan menuju anoda yang bermuatan positif. Sebaliknya, bila pH lingkungan di bawah pI, muatan protein menjadi positif yang membuatnya akan bermigrasi menuju katoda yang bermuatan negatif (Autran 1996). Hal inilah yang menjadi dasar pemisahan protein dengan elektroforesis.Metode elektroforesis protein yang paling umum dan banyak dilakukan adalah SDS-PAGE (Sodium Dodecyl Sulfate-Polyacrylamide Gel Electropho-resis). SDS-PAGE merupakan teknik elektroforesis dalam sistem buffer diskontinyu yang menggunakan dua tipe gel sebagai medianya, yaitu stacking gel dan separating gel. Sistem buffer yang diskontinyu membuat sampel terkonsentrasi dalam stacking gel sehingga menghasilkan resolusi yang lebih baik ketika pemisahan protein terjadi di
separating gel (Garfin 1990). Skematik Elektroforesis SDS-PAGE dapat dilihat pada Gambar 16.
Gambar 16. Skematik electroforesis SDS-PAGE (Anonimb)
Gel poliakrilamid dibentuk dari hasil ko-polimerisasi monomer akrilamid (CH2=CH-CO-NH2) dengan bantuan senyawa yang bertindak sebagai cross-linking agent yaitu N,N‟-metilen-bisakrilamid (CH2=CH-CO-NH-CH2-NH-CO-CH=CH2). Mekanisme polimerisasi akrilamid tersebut dikatalisis oleh TEMED (tetrametiletilendiamin) dan APS (amonium persulfat). TEMED akan menyebabkan pembentukan radikal bebas dari amonium persulfat yang mengakibatkan reaksi pembentukan akrilamid aktif. Akrilamid aktif ini akan bereaksi dengan akrilamid lainnya membentuk rantai polimer yang panjang. Hasil dari polimerisasi ini adalah terbentuknya gel dengan struktur jala dari rantai akrilamid. Ukuran pori dan jala gel tersebut ditentukan oleh jumlah akrilamid yang digunakan per unit volumenya dan derajat ikatan silangnya (Garfin 1990; Autran 1996).
Sodium dodecyl sulfate (SDS) adalah detergen anionik yang paling umum digunakan dalam
elektroforesis. SDS memiliki dua fungsi, yaitu : (1) untuk memisahkan protein-protein yang beragregasi, hidrofobik atau memiliki kelarutan yang rendah, seperti membran protein; dan (2) memisahkan protein berdasarkan bentuk, ukuran dan berat molekulnya. SDS menyelimuti protein dengan muatan negatif serta mengikat protein dengan rasio yang konstan, yaitu 1.4 g SDS per gram polipeptida (Garfin 1990; Autran 1996).
Interaksi SDS dengan protein akan merusak seluruh ikatan non-kovalen protein sehingga struktur protein akan terbuka. Selanjutnya, penggunaan reducing agent seperti 2-merkaptoetanol atau
20
ditiothreitol akan membantu mendenaturasi protein melalui pemutusan ikatan disulfida pada protein sehingga memecahnya menjadi subunit-subunit protein. Akibatnya, mobilitas elektroforetik dari kompleks detergen-polipeptida hanya merupakan fungsi dari berat molekul protein (Garfin 1990).Penggunaan buffer dalam elektroforesis gel dapat digunakan dengan dua sistem, yaitu kontinyu (homogenous) dan diskontinyu (multiphasic) (Copeland 1994). Perbedaan mendasar pada sistem diskontinyu adalah penggunaan dua gel dalam satu slab, yaitu stacking gel dan separating gel. Buffer dan konsentrasi akrilamid yang digunakan pada kedua jenis gel tersebut berbeda (Boyer 1993). Pada
stacking gel digunakan buffer dengan pH 6.8 dan konsentrasi akrilamid yang lebih rendah (ukuran
pori besar) sedangkan pada separating gel digunakan buffer dengan pH 8.8 dan konsentrasi akrilamid yang tinggi (ukuran pori kecil) (Wilson dan Walker 2000). Hal ini akan menghasilkan pemisahan yang baik dengan pita yang tajam karena protein terkonsentrasi pada stacking gel dan mengalami resolusi yang tinggi pada separating gel.
3. Interpretasi Pita Protein
Gel hasil elektroforesis menunjukkan pita-pita protein dengan berat molekul yang berbeda. Protein dengan berat molekul yang lebih besar akan tertahan diatas, sedangkan protein dengan berat molekul yang lebih kecil akan berada dibawah. Penentuan berat molekul pita protein sampel berdasarkan pita protein marker yang digunakan dapat menggunakan persamaan regresi antara mobilitas relatif (Rf) protein marker dengan logaritma dari berat molekul marker yang telah diketahui. Nilai Rf tersebut dirumuskan sebagai :
jarak migrasi protein
Persamaan regresi marker tersebut dapat digunakan untuk menentukan berat molekul pita protein sampel ( dengan y = log BM protein dan x = nilai Rf pita protein). Contoh persamaan regresi marker dapat dilhat pada Gambar 17.
Gambar 17. Contoh persamaan regresi marker
Interpretasi pita protein berdasarkan berat molekul ini umumnya dibandingkan dengan profil protein sejenis yang berasal dari pustaka lain. Profil protein kedelai dengan SDS-PAGE baik total protein maupun hasil pengisolasian protein 11S dan 7S sudah banyak dipublikasikan. Proses isolasi protein 11S dan 7S yang sering digunakan adalah metode Thanh dan Shibasaki (1976). Hasil publikasi Mujoo et al. (2003) mengenai profil protein isolasi 11S dan 7S dapat dilihat pada Gambar 18.
21
Gambar 18. Profil protein 11S dan 7S dari tujuh varietas kedelai dengan SDS-PAGEAngka ganjil = 11S dan angka genap = 7S
Berdasarkan profil protein isolasi 11S dan 7S tersebut dapat digunakan untuk menginterpretasikan pita protein sejenis. Gambar diatas menunjukkan baik protein 11S maupun 7S memiliki beberapa pita protein yang sama dengan intensitas yang berbeda. Sehingga penentuan subunit pada masing-masing fraksi protein berdasarkan pita protein yang dominan (pita yang lebih tebal). Dimana protein 11S memiliki subunit golongan Asam (A1, A2 ,A3, A4, A5) dan Basa (B1,B2,B3,B4) dan protein 7S memiliki subunit α΄, α dan β. Namun berat molekul subunit-subunit pada protein 11S maupun 7S merupakan suatu kisaran, sehingga ada beberapa literatur yang menyatakan berat molekul yang berbeda-beda.