IV. HASIL DAN PEMBAHASAN
4.1. Studi distribusi spasial tumbuhan sagu di P. Seram, Maluku
Hasil analisis tutupan lahan (land cover) menggunakan metode klasifikasi terbimbing (supervised classification) menunjukkan bahwa tumbuhan sagu terdistribusi pada wilayah pesisir di dataran rendah pada tanah-tanah endapan, di tempat-tempat yang berdekatan dengan sungai, lembah-lembah bukit dengan total luas areal mencapai 18.239 ha (Lampiran 1). Apabila luas areal sagu ini dibandingkan dengan luas P. Seram 1,8 juta ha, maka luas tutupan sagu hanya mencapai satu persen. Dalam rangka uji akurasi dilakukan pengumpulan Ground
Control Points (GCP). Tingkat akurasi klasifikasi cukup tinggi mencapai 76 %
(Gambar 9) berdasarkan overall accuracy (Producer’s Accuracy = 73 % dan
User’s Accuracy = 68 %) . Tingkat akurasi ini dipengaruhi oleh awan dan kondisi
topografi sehingga tumbuhan sagu tidak terdeteksi dengan baik pada citra. Selain itu diduga karena terdapat tumbuhan sagu yang tumbuh bercampur dengan vegetasi pohon dan/atau ternaungi sehingga nilai dijitalnya bercampur dengan nilai dijital tumbuhan di atasnya. Hambatan lain yang bisa terjadi adalah pertumbuhan sagu yang tidak teratur dengan luas klaster yang relatif terbatas yakni tidak mencukupi ukuran 3 x 3 piksel. Semua klaster sagu dengan ukuran luas melebihi ukuran tersebut apabila tidak terdapat hambatan lain, misalnya karena adanya awan, maka klaster tersebut tampak pada citra. Resolusi citra landsat-5TM minimal yang dapat terdeteksi pada citra sebanyak sembilan piksel karena dilakukan focal scan dengan ukuran window 3 x 3.
Peta distribusi spasial tumbuhan sagu di P. Seram apabila dikaitkan dengan sifat-sifat lahan, maka dapat dikemukakan bahwa tumbuhan sagu menyukai kondisi lahan dengan ciri-ciri yaitu : 1) lahan datar-curam, 2) dekat pesisir, 3) dekat sungai, 4) pada tanah-tanah aluvial (Entisol dan Inceptisol), dan 5) pada ketinggian tempat antara 0-250 m dpl. Sifat-sifat lahan ini selanjutnya disebut sebagai kondisi habitat tumbuhan sagu yang sesuai, sedangkan kondisi lahan yang tidak termasuk dalam kategori di atas dikategorikan sebagai kondisi habitat yang kurang, bahkan tidak sesuai untuk pertumbuhan dan pengembangan sagu.
4.1.1. Distribusi sagu pada berbagai elevasi
Overlay antara distribusi sagu dan Digital Elevation Model (DEM) ASTER dan observasi lapangan menunjukkan bahwa kebanyakan tumbuhan sagu tumbuh di dataran rendah pada elevasi (ketinggian tempat) ≤ 250 meter dari atas permukaan laut (m dpl) (Tabel 10 dan Gambar 10). Lahan pada ketinggian ini merupakan habitat yang banyak ditemukan tumbuhan sagu, mencapai 99,98 %. Bintoro (2008) mengemukakan bahwa pada tegakan sagu alami di berbagai daerah di Indonesia seperti Papua, Maluku, Sulawesi, Kalimantan, dan Sumatera banyak ditemukan sagu tumbuh pada ketinggian tempat mencapai 300 m dpl. Secara umum tumbuhan sagu dapat tumbuh sampai mencapai ketinggian tempat 1000 m dpl, tetapi pertumbuhan sagu terbaik berada pada ketinggian antara 0 - 400 m dpl. Pada ketinggian yang lebih besar pertumbuhannya terhambat dan memiliki kandungan pati rendah. Pada ketinggian tempat di atas 600 m dpl pertumbuhan sagu memendek, hanya mencapai tinggi sekitar enam meter. Atas dasar ketinggian tempat ini, berdasarkan hasil survey BPPT (1982) dilaporkan bahwa tumbuhan sagu di Maluku pada umumnya ditemukan tumbuh pada ketinggian antara 0-20 m dpl.
Berdasarkan fakta di atas, maka dapat dikemukakan bahwa tumbuhan sagu yang tumbuh dan berkembang di P. Seram kondisi habitat yang sesuai terletak pada ketinggian tempat antara 0-250 m dpl (Lampiran 2). Pada ketinggian tempat yang lebih besar dari 250 m dpl tumbuhan sagu yang tumbuh pada ketinggian tersebut tidak mencapai setengah persen. Hal ini berarti bahwa secara alami tumbuhan sagu tidak dapat beradaptasi dengan baik pada ketinggian yang melebihi 250 m dpl. Kalaupun terdapat sagu, luas klasternya kecil-kecil atau hanya terdiri atas beberapa rumpun. Klaster-klaster seperti dengan menggunakan citra Landsat tidak dapat terdeteksi. Kelas elevasi dan distribusi sagu pada berbagai elevasi di P. Seram disajikan pada Tabel 10.
Tabel 10. Distribusi sagu pada berbagai kelas elevasi di P. Seram, Maluku Ketinggian tempat
(m dpl)
Luas P. Seram
(ha) % Luas sagu (ha) %
0 - 125 617.800,32 34,64 13.385,61 73,39 125 - 250 324.529,29 18,19 4.814,28 26,39 250 - 375 236.379,87 13,25 39,87 0,22 375 - 500 174.499,83 9,78 0,00 0,00 500 - 625 126.367,02 7,08 0,00 0,00 625 - 750 91.214,10 5,11 0,00 0,00 750 - 825 43.930,98 2,46 0,00 0,00 825 - 1000 68.282,82 3,83 0,00 0,00 > 1000 100.680,75 5,64 0,00 0,00 Jumlah 1.783.684,98 100,00 18.239,76 100,00
Pada Tabel di atas tampak bahwa dari sisi ketinggian tempat di atas permukaan laut, lahan yang terletak pada ketinggian antara 0-250 m dpl sebanyak 942.329,61 ha atau setara 52,83 % dari total luas lahan di P. Seram yang mencapai 1,8 juta hektar. Hal ini berarti bahwa secara parsial berdasarkan ketinggian tempat, maka sekitar 50 % lahan sagu di P. Seram dapat dikatakan sesuai sebagai habitat tumbuhan sagu. Pada luas lahan tersebut tumbuh dan berkembang sekitar 18.239 ha tumbuhan sagu. Distribusi sagu di P. Seram pada berbagai ketinggian tempat yaitu : 1) pada ketinggian 0-125 m dpl sebanyak 13,39 ribu ha (73,39 %), 2) 125-250 m dpl 4,81 ribu ha (26,39 %). Apabila luas ini dikaitkan dengan luas lahan yang terdapat pada ketinggian antara 0-250 m dpl, maka dapat dikatakan bahwa hanya sekitar 1,94 % lahan yang ditumbuhi sagu, sisanya berupa jenis vegetasi yang lain. Overlay distribusi sagu pada berbagai ketinggian tempat di P. Seram tersaji pada Gambar 8.
4.1.2. Distribusi sagu pada berbagai kemiringan
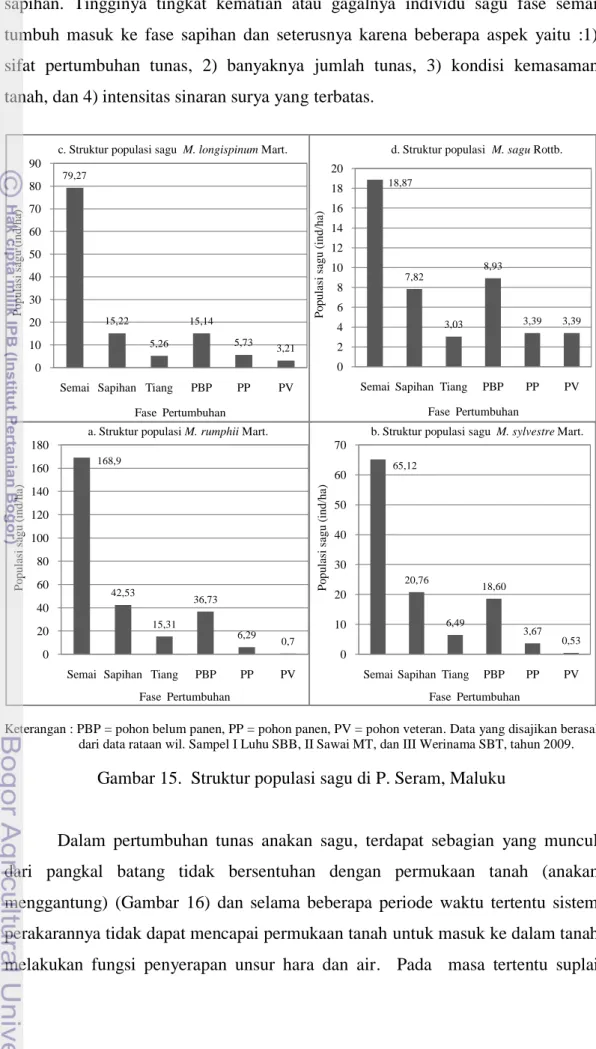
Kondisi topografi lahan di P. Seram pada umumnya berbukit dan berlereng. Berdasarkan sifat lahan ini, tumbuhan sagu banyak ditemukan tumbuh pada kondisi lahan dengan kemiringan rendah atau memiliki slope antara kecil-sedang, berkisar antara 0-40 % (Tabel 11 dan Gambar 11). Kondisi kemiringan lereng ini merupakan sifat lahan yang sesuai sebagai habitat untuk pertumbuhan dan perkembangan sagu (Lampiran 3). Pada kemiringan lereng yang lebih besar dari 40 persen tidak sesuai bagi pertumbuhan sagu. Secara parsial berdasarkan kemiringan lereng, di P. Seram terdapat sekitar 336.922,65 ha setara dengan 18,96
% luas lahan yang sesuai sebagai habitat tumbuhan sagu. Pada luas lahan 336,9 ribu ha ini telah tumbuh sebanyak 18,239 ha sagu. Distribusinya pada berbagai kemiringan lereng yaitu : 1) kemiringan 0-88 % (datar) sebanyak 9,17 ribu ha (50 %), 2) 8-15 % (landai) 2,79 ribu ha (15 %), 3) 15-25 % (agak curam) 2,85 ribu ha (16 %), 25-40 % (curam) 2,68 ribu ha (15 %). Dengan kata lain sebanyak 98 % tumbuhan sagu di P. Seram tumbuh dan berkembang pada kemiringan lereng antara 0-40 %. Sisanya sebesar empat persen tumbuh pada kemiringan >40 %. Tabel 11. Distribusi sagu pada berbagai kelas kemiringan lereng di P. Seram,
Maluku
Kemiringan Lereng Luas P. Seram
(ha) % Luas sagu (ha) % 0 - 8 % (Datar) 336.922,65 18,96 9.169,20 50,00 8 - 15 % (Landai) 270.211,77 15,21 2.787,39 15,00 15 - 25 % (Agak curam) 408.540,78 23,00 2.845,35 16,00 25 - 40 % (Curam) 541.066,77 30,45 2.681 15,00 >40% (Sangat curam) 220.146,39 12,39 756,72 4,00 Jumlah 1.776.888,36 100,00 18.239,66 100,00
4.1.3. Distribusi sagu pada berbagai sistem lahan
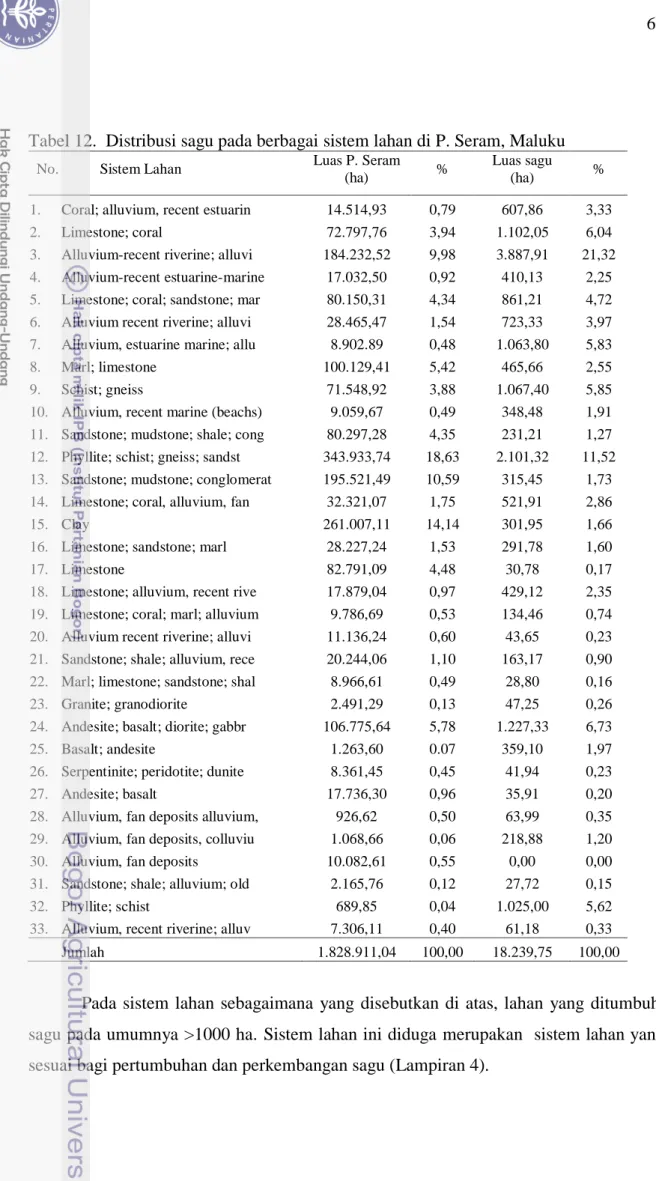
Sistem lahan (land system) berkaitan dengan bahan induk pembentukan tanah. Oleh karena itu sistem lahan menentukan jenis tanah yang terbentuk di suatu tempat. Berdasarkan analisis sistem lahan (land system), tumbuhan sagu di P. Seram kebanyakan terdistribusi pada sistem lahan dari bahan-bahan seperti : 1) Limestone; coral, seluas 607,86 ha (3,33 %), 2) Alluvium-recent riverine; alluvi, 1.102,05 ha (6,04 %), 3) Alluvium, estuarine marine; allu, 3.887,91 ha (21,32 %), 4) Schist; gneiss, 1.067,40 ha (5,85 %), 5) Phyllite; schist; gneiss; sandst, 2.101,32 ha (11,52 %), 6) Andesite; basalt; diorite; gabbr, 1.227,33 ha (6,73 %), dan 7) Phyllite; schist 1.025,00 ha (5,62 %) (Tabel 12 dan Gambar 12). Jumlah luas lahan pada sistem lahan ini mencapai 11.474 ha, setara 62,64 % dari total lahan sagu pulau Seram 1,8 juta ha.
Tabel 12. Distribusi sagu pada berbagai sistem lahan di P. Seram, Maluku No. Sistem Lahan Luas P. Seram
(ha) %
Luas sagu
(ha) %
1. Coral; alluvium, recent estuarin 14.514,93 0,79 607,86 3,33 2. Limestone; coral 72.797,76 3,94 1.102,05 6,04 3. Alluvium-recent riverine; alluvi 184.232,52 9,98 3.887,91 21,32 4. Alluvium-recent estuarine-marine 17.032,50 0,92 410,13 2,25 5. Limestone; coral; sandstone; mar 80.150,31 4,34 861,21 4,72 6. Alluvium recent riverine; alluvi 28.465,47 1,54 723,33 3,97 7. Alluvium, estuarine marine; allu 8.902.89 0,48 1.063,80 5,83 8. Marl; limestone 100.129,41 5,42 465,66 2,55 9. Schist; gneiss 71.548,92 3,88 1.067,40 5,85 10. Alluvium, recent marine (beachs) 9.059,67 0,49 348,48 1,91 11. Sandstone; mudstone; shale; cong 80.297,28 4,35 231,21 1,27 12. Phyllite; schist; gneiss; sandst 343.933,74 18,63 2.101,32 11,52 13. Sandstone; mudstone; conglomerat 195.521,49 10,59 315,45 1,73 14. Limestone; coral, alluvium, fan 32.321,07 1,75 521,91 2,86
15. Clay 261.007,11 14,14 301,95 1,66
16. Limestone; sandstone; marl 28.227,24 1,53 291,78 1,60
17. Limestone 82.791,09 4,48 30,78 0,17
18. Limestone; alluvium, recent rive 17.879,04 0,97 429,12 2,35 19. Limestone; coral; marl; alluvium 9.786,69 0,53 134,46 0,74 20. Alluvium recent riverine; alluvi 11.136,24 0,60 43,65 0,23 21. Sandstone; shale; alluvium, rece 20.244,06 1,10 163,17 0,90 22. Marl; limestone; sandstone; shal 8.966,61 0,49 28,80 0,16 23. Granite; granodiorite 2.491,29 0,13 47,25 0,26 24. Andesite; basalt; diorite; gabbr 106.775,64 5,78 1.227,33 6,73 25. Basalt; andesite 1.263,60 0.07 359,10 1,97 26. Serpentinite; peridotite; dunite 8.361,45 0,45 41,94 0,23 27. Andesite; basalt 17.736,30 0,96 35,91 0,20 28. Alluvium, fan deposits alluvium, 926,62 0,50 63,99 0,35 29. Alluvium, fan deposits, colluviu 1.068,66 0,06 218,88 1,20 30. Alluvium, fan deposits 10.082,61 0,55 0,00 0,00 31. Sandstone; shale; alluvium; old 2.165,76 0,12 27,72 0,15 32. Phyllite; schist 689,85 0,04 1.025,00 5,62 33. Alluvium, recent riverine; alluv 7.306,11 0,40 61,18 0,33
Jumlah 1.828.911,04 100,00 18.239,75 100,00
Pada sistem lahan sebagaimana yang disebutkan di atas, lahan yang ditumbuhi sagu pada umumnya >1000 ha. Sistem lahan ini diduga merupakan sistem lahan yang sesuai bagi pertumbuhan dan perkembangan sagu (Lampiran 4).
4.1.4. Distribusi sagu pada berbagai jenis tanah
Berdasarkan sistem lahan, apabila dikaitkan dengan jenis tanah yang terbentuk di P. Seram, maka tampak bahwa tumbuhan sagu banyak berkembang pada lima jenis tanah yatitu : 1) entisols, 2) alfisols, 3) inceptisols, 4) oxisols, dan 5) ultisols (Tabel 13). Ciri yang dipakai untuk menjelaskan jenis-jenis tanah tersebut adalah berdasarkan empat huruf pada bagian akhir tiap-tiap jenis tanah, sesuai ketentuan penamaan menurut Keys to Soil Taxonomy (Soil Survey Staff USDA 1999). Jenis-jenis tanah ini yang selanjutnya dapat dikategorikan sebagai tanah-tanah yang sesuai bagi pertumbuhan dan perkembangan sagu (Lampiran 5).
Distribusi sagu paling banyak pada jenis tanah entisols dan inceptisols. Jenis tanah ini merupakan tanah yang belum lama terbentuk atau tanah yang baru mulai berkembang. Diduga bahan-bahan penyusunnya berasal dari bahan endapan baru yang terbawa dari bagian ketinggian terbawa oleh aliran permukaan (run off), atau berasal dari endapan banjir apabila terjadi hujan. Menurut Keys to
Soil Taxonomy (Soil Survey Staff USDA 1999) tanah-tanah ini merupakan jenis
tanah baru (recent). Penamaan ini sepadan dengan jenis tanah Aluvial menurut sistem klasifikasi tanah yang dikembangkan oleh Pusat penelitian Tanah Bogor (Hardjowigeno 2003). Jenis tanah Entisols memiliki ciri antara lain selalu jenuh air dan matriksnya tereduksi pada semua horizon di bawah kedalaman 25 cm dari permukaan tanah. Sedangkan jenis tanah Inceptisols memiliki ciri antara lain mulai terbentuk struktur dan di dalam kedalaman 50 cm dari permukaan tanah mineral mengandung cukup besi fero (Fe2+) aktif untuk dapat memberikan reaksi positif terhadap alpha, alpha-dipyridil ketika tanah tidak sedang diirigasi (Soil Survey Staff USDA 1999). Pada Gambar 13 tampak sebaran jenis tanah yang sesuai bagi habitat tumbuhan sagu.
Tabel 13. Distribusi sagu pada berbagai jenis tanah di P. Seram, Maluku
No. Jenis tanah Luas (ha) (%)
1. Troporthens; Tropudalfs; Tropop 607,86 3,33
2. Rendolls; Eutropepts; Tropudalfs 1.665,63 9,13
3. Tropaquepts; Fluvaquents 3.887,91 21,32
4. Hydraquents; Sulfaquuents 410,13 2,25
5. Rendolls; Tropudalfs; Eutropepts 861,21 4,72
… lanjutan
7. Tropaquepts; Eutropepts; Tropud 1.063,80 5,83 8. Eutropepts; Dystropepts; Troport 465,66 2,55
9. Dystropepts; Tropudults 2.377,17 13,03
10. Tropopsamments; Tropaquents 348,48 1,91
11. Tropudults; Dystropept 247,23 1,36
12. Humitropepts; Dystropepts; Tropa 2.101,32 11,52 13. Dystropepts; troporthents; Tropu 315,45 1,73
14. Rendolls; Eutropepts 552,69 3,03
15. Eutropepts; Paleudults; Troudal 301,95 1,66
16. Eutropepts; Tropudults 291,78 1,60
17. Tropofluvents; Tropaquepts 43,65 0,24
18. Tropudults; Tropudalfs; Dystrope 163,17 0,90 19. Eutropepts; Dystropepts; tropudda 28,80 0,16
20. Eutropepts; Dystropepts 359,10 1,97
21. Dystropepts; Eutropepts; Tropudu 41,94 0,23
22. Haplorthox; Acrorthox; Dystrpep 35,91 0,20
23. Dystropepts; Tropudults; Troport 63,99 0,35 24. Dystropepts; Dystrandepts; Tropa 218,88 1,20 25. Dystropepts; Dystrandepts; Tropu 27,72 0,15 26. tropaquents; Tropopluvents; Fluva 1.025,01 5,62
27. Eutropepts; Tropaquepts; Tropfl 9,99 0,05
Jumlah 18239,76 100,00
4.1.4. Distribusi sagu pada berbagai jarak dari sungai
Tumbuhan sagu banyak juga yang tumbuh di bagian pinggir kiri-kanan sungai yang datar atau berupa rawa-rawa yang senantiasa terendam air, baik permanen ataupun temporer (Gambar 14). Hasil observasi lapangan ditemukan bahwa tidak semua kawasan kiri-kanan sungai terdapat tumbuhan sagu. Bagian kiri-kanan sungai yang biasanya dapat ditemukan tumbuhan sagu adalah bagian pinggir sungai yang tanahnya terdapat liat, memiliki tekstur halus, atau mengandung bahan organik yang memadai yang ditunjukkan oleh perubahan warga coklat kehitaman. Pada bagian pinggir sungai yang didominasi partikel pasir, jarang atau bahkan tidak ditemukan sagu. Louhenapessy (1993) mengemukakan bahwa tumbuhan sagu dapat bertumbuh dengan baik pada tanah-tanah berpasir apabila mengandung bahan organik yang cukup.
Hasil overlay distribusi sagu dengan jarak dari pinggir kiri-kanan sungai (buffer sungai) menunjukkan bahwa terdapat sekitar 10.834,44 ha (59,40 %) tumbuhan sagu yang tumbuh pada sisi pinggir kiri-kanan sungai berjarak <300. Hal ini berarti bahwa terdapat sekitar 7.405,36 ha tumbuhan sagu yang tumbuh pada jarak > 300 m (40,60 %) dari sisi kiri-kanan sungai (Tabel 14).
Tabel 14. Distribusi sagu pada berbagai jarak dari sungai di P. Seram, Maluku
No. Jarak dari sungai Luas (ha) (%)
1. 0 - 300 m 10.834,44 59,40
2. > 300 m 7.405,36 40,60
Jumlah 18.239,80 100,00
Pada Tabel di atas tampak bahwa tidak semua areal sagu di P. Seram terdapat di tempat yang berdekatan dengan sungai pada jarak sekitar 300 m. Artinya terdapat sebagian tumbuhan sagu di luar jarak tersebut. Hal ini berarti bahwa buffer sungai untuk keperluan mengetahui distribusi tumbuhan sagu pada sisi kiri-kanan sungai jaraknya perlu diperlebar.
4.2. Studi Autekologi tumbuhan sagu di P. Seram, Maluku
4.2.1. Struktur populasi
Struktur populasi tumbuhan sagu (Metroxylon spp.) dalam kajian ini diuraikan menurut fase pertumbuhan berdasarkan kriteria yang dikembangkan BPPT (1982 dalam Haryanto dan Pangloli 1992) dengan modifikasi. Fase pertumbuhan dimaksud meliputi fase semai (seedling), sapihan (sapling), tiang (pole), dan pohon (trees). Secara teoritis dikenal tiga struktur populasi yang dapat ditemukan dalam pertumbuhan suatu organisme. Pemisahan ini pada umumnya didasarkan pada tingkatan umur organisme, yaitu struktur populasi menurun, stabil, dan muda (Wirakusumah 2003). Struktur populasi menurun dicirikan oleh jumlah populasi yang tinggi pada fase umur post-reproduktif, berkurang pada fase
reproduktif, dan semakin sedikit pada fase pre-reproduktif. Struktur populasi
stabil dicirikan oleh jumlah populasi yang rendah pada fase post-reproduktif,
makin bertambah pada fase reproduktif, dan makin bertambah lagi pada fase
pre-reproduktif. Struktur populasi yang berikut adalah struktur populasi muda, yaitu
struktur populasi yang dicirikan oleh jumlah individu rendah pada fase
post-reproduktif, sedang pada fase produktif, dan sangat banyak pada fase pre-reproduktif.
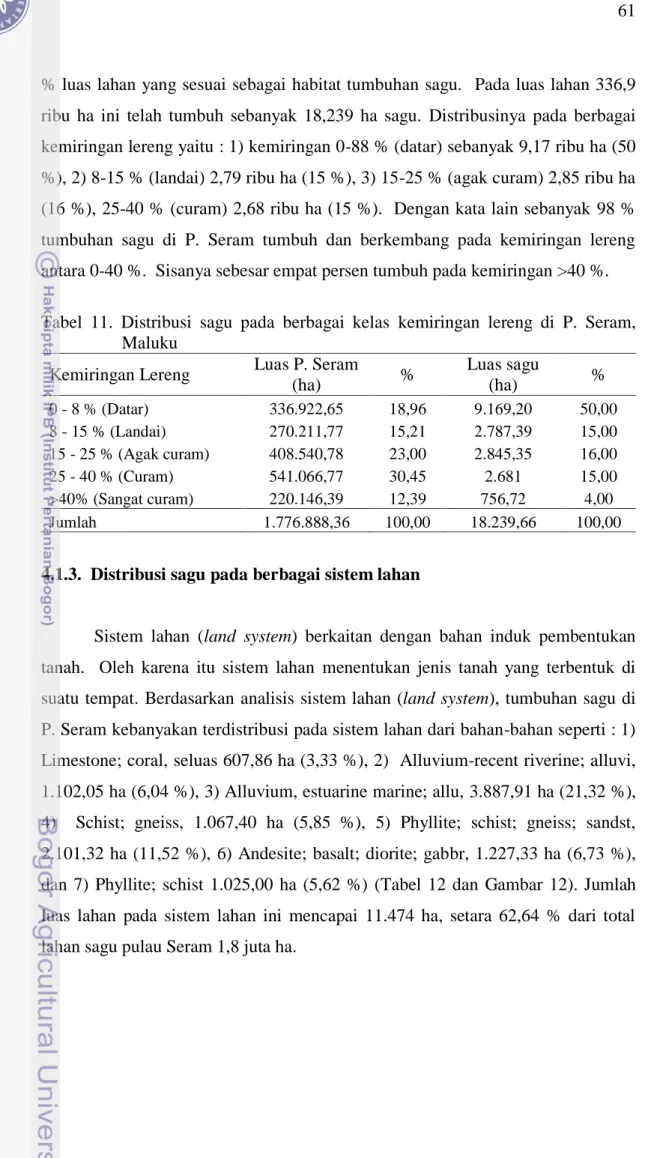
Struktur populasi sagu yang ditemukan di P. Seram Maluku secara umum menyerupai struktur populasi muda yaitu populasi yang memiliki jumlah individu paling banyak pada fase semai, kemudian berkurang secara drastis pada fase sapihan, tiang, dan sedikit meningkat pada fase pohon (Gambar 15). Terjadinya peningkatan populasi pada fase pohon diduga karena tiga alasan yaitu : 1) fase pohon menghambat pertumbuhan sagu pada fase dibawahnya sehingga terjadi gap antara sagu fase tiang dan pohon belum panen. Hambatan ini berlangsung sampai dengan tumbuhan sagu fase pohon tersebut berbunga. Ketika tumbuhan sagu sampai pada fase berbunga, maka ukuran daun mengecil atau tangkai dan anak-anak daun memendek. Pada masa itu tidak lagi terjadi penambahan daun yang kemudian mengarah ke kematian. Hambatan tersebut dapat pula berakhir apabila tumbuhan sagu fase pohon telah dipanen, 2) sejak akhir fase tiang sampai dengan awal fase pohon panen diperlukan periode waktu sekitar 5-7 tahun, dengan asumsi masak panen sagu sekitar 15 tahun, 2) kriteria ukuran tinggi sekitar dua meter bagi pertumbuhan sagu selama fase tiang patut dikoreksi menjadi tiga meter atau memerlukan periode waktu sekitar empat tahun, dan Apabila tiga aspek sebagaimana tersebut di atas dimasukkan dalam pembangunan struktur populasi tumbuhan sagu, kemudian ditarik garis pola pertumbuhan (trend line) maka akan membentuk huruf ‘J’ terbalik. Hasil ini mirip dengan temuan Rostiwati et al. (2008) yang memperoleh trend line yang sama dengan struktur populasi sagu berdasarkan data rumpun sagu alam di Papua dan Maluku.
Dalam struktur populasi sagu di P. Seram secara umum, dimana jumlah individu rata-rata fase semai mencapai 83,04 ind/ha, apabila dibandingkan dengan jumlah individu rata-rata fase sapihan sebanyak 21,59 ind/ha, terlihat bahwa terjadi pengurangan jumlah individu sagu mencapai 76,18 %. Dengan kata lain hanya sebanyak 23,82 % individu semai sagu yang sukses tumbuh mencapai fase
sapihan. Tingginya tingkat kematian atau gagalnya individu sagu fase semai tumbuh masuk ke fase sapihan dan seterusnya karena beberapa aspek yaitu :1) sifat pertumbuhan tunas, 2) banyaknya jumlah tunas, 3) kondisi kemasaman tanah, dan 4) intensitas sinaran surya yang terbatas.
Keterangan : PBP = pohon belum panen, PP = pohon panen, PV = pohon veteran. Data yang disajikan berasal dari data rataan wil. Sampel I Luhu SBB, II Sawai MT, dan III Werinama SBT, tahun 2009.
Gambar 15. Struktur populasi sagu di P. Seram, Maluku
Dalam pertumbuhan tunas anakan sagu, terdapat sebagian yang muncul dari pangkal batang tidak bersentuhan dengan permukaan tanah (anakan menggantung) (Gambar 16) dan selama beberapa periode waktu tertentu sistem perakarannya tidak dapat mencapai permukaan tanah untuk masuk ke dalam tanah melakukan fungsi penyerapan unsur hara dan air. Pada masa tertentu suplai
168,9 42,53 15,31 36,73 6,29 0,7 0 20 40 60 80 100 120 140 160 180
Semai Sapihan Tiang PBP PP PV
P o pul as i sa gu (in d/ ha ) Fase Pertumbuhan a. Struktur populasi M. rumphii Mart.
65,12 20,76 6,49 18,60 3,67 0,53 0 10 20 30 40 50 60 70
Semai Sapihan Tiang PBP PP PV
P o pul as i sa gu (in d/ ha ) Fase Pertumbuhan
b. Struktur populasi sagu M. sylvestre Mart. 79,27 15,22 5,26 15,14 5,73 3,21 0 10 20 30 40 50 60 70 80 90
Semai Sapihan Tiang PBP PP PV
P o pul as i sa gu (in d/ ha ) Fase Pertumbuhan
c. Struktur populasi sagu M. longispinum Mart.
18,87 7,82 3,03 8,93 3,39 3,39 0 2 4 6 8 10 12 14 16 18 20
Semai Sapihan Tiang PBP PP PV
Po p u la si sa g u ( in d /h a ) Fase Pertumbuhan d. Struktur populasi M. sagu Rottb.
makanan dari pohon induk ke tunas anakan tersebut terhenti. Dengan terhentinya suplai makanan ini, maka kebutuhannya tidak mencukupi untuk melangsungkan pertumbuhannya, selanjutnya menyebabkan kematian tunas anakan.
Gambar 16. Anakan sagu M. rumphii Mart. yang tumbuh menggantung Tumbuhan sagu senantiasa menghasilkan jumlah tunas anakan dalam jumlah relatif banyak, sehingga memungkinkan terjadinya persaingan. Persaingan dapat terjadi diantara sesama tunas anakan maupun persaingan dengan individu yang tumbuh lebih awal. Dalam rumpun sagu sangat mungkin terjadi kompetisi atau persaingan diantara sesama individu semai itu sendiri dan fase di atasnya. Persaingan yang dimaksud berkaitan dengan komponen di atas tanah (atmosfer) seperti udara, cahaya, ruang, dan komponen di dalam tanah seperti air, oksigen, dan unsur hara.
Hasil pengukuran intensitas sinaran surya di dekat rumpun sagu menunjukkan bahwa hanya sekitar 12,40 % setara 206,53 lux sinaran surya yang masuk sampai di dekat rumpun sagu. Pada ruang terbuka mencapai 1675,29 lux. Hal ini karena terdapat hambatan masuknya cahaya matahari oleh tajuk rumpun
tumbuhan sagu itu sendiri. Keterbatasan intensitas sinaran ini dapat berdampak terhadap pertumbuhan anakan sagu dan dapat berakhir dengan kematian. Sinaran surya merupakan sumber energi utama bagi kehidupan, dan berperan dalam proses fotosintesis. Apabila intensitas sinaran surya terbatas maka proses fotosintesis tidak dapat berlangsung secara maksimal. Implikasi berikutnya adalah hasil fotosintesis (fotosintat) akan berkurang. Dengan demikian kebutuhan tunas anakan tidak dapat terpenuhi, selanjutnya dapat menyebabkan kematian anakan sagu.
Tumbuhan sagu selain tumbuh pada lahan kering, sebagian besar tumbuh pada kondisi habitat tergenang, baik temporer maupun permanen. Lahan yang tergenang memungkinkan kondisinya menjadi tereduksi. Artinya terjadi kekurangan oksigen di dalam air dan tanah, karena diserap oleh akar dan organisme yang hidup di air maupun dalam tanah melalui proses respirasi. Dalam proses respirasi terjadi pembebasan molekul karbondioksida (CO2). Dengan
pertambahan waktu akan terjadi akumulasi CO2 dalam air dan dalam tanah.
Akumulasi ini memungkinkan terjadi reaksi dengan molekul air (H2O)
menghasilkan senyawa asam karbonat (H2CO3). Senyawa ini memberikan akses
masam ke dalam tanah maupun air, sehingga tingkat kemasaman akan meningkat (pH turun).
Hasil analisis pH (KCl) tanah atau kemasaman potensial dapat mencapai 4,3. Artinya apabila kondisi tanah terendam air dalam periode waktu cukup lama, kemasaman tanah berpotensi turun mencapai angka tersebut. Dalam kaitan dengan kematian tunas anakan sagu, diduga sebagian tunas anakan yang baru muncul tidak dapat bertahan hidup pada kondisi kemasaman yang rendah, dapat pula terjadi melalui pengaruh kemasaman rendah terhadap sistem perakaran tunas anakan yang masih muda. Kemasaman dapat bersifat mengikis sehingga dapat merusak sel-sel akar terutama yang masih muda. Akar tunas anakan yang mengalami kerusakan, berdampak terhadap pertumbuhan secara keseluruhan dan dapat menyebabkan kematian anakan.
Kematian anakan sagu dapat pula disebabkan karena keracunan Fe dan Al. Hasil pengukuran Fe dan Al di dalam tanah mencapai 3,08 % dan 4,99 %, termasuk kategori sangat tinggi menurut kriteria BPT Bogor (2005). Brady (1990)
mengemukakan bahwa konsentrasi unsur mikro seperti Fe yang terlalu tinggi dalam tanah dapat bersifat toxic. Fakta penelitian mengenai kematian semai pada kondisi habitat tergenang atau tereduksi menunjukkan bahwa jumlah populasi semai pada kondisi habitat tergenang temporer air payau dan tergenang permanen hanya mencapai 162 ind/ha dan 264 ind/ha. Jumlah populasi ini hanya separoh dibandingkan dengan populasi semai pada habitat lahan kering dan tergenang temporer air tawar masing-masing sebesar 345,28 ind/ha dan 416,90 ind/ha.
Hasil penelitian jumlah populasi rumpun sagu di P. Seram ditemukan sebanyak 176,55 rumpun/ha (Gambar 17 dan Lampiran 6). Pada setiap rumpun sagu tidak selalu dapat ditemukan rumpun yang memiliki semua fase pertumbuhan berupa pohon, tiang, sapihan, dan semai. Komposisi fase pertumbuhan yang sering dijumpai pada suatu rumpun sagu di lapangan dalam komunitas sagu alami adalah sebagai berikut : semai, sapihan, pohon; semai, pohon; semai, tiang; semai, sapihan; dan/atau hanya semai saja. Pada sejumlah rumpun sagu yang mencapai 176,55 rumpun/ha, ditemukan fase pohon sebanyak 106,06 ind/ha. Jumlah populasi pohon ini termasuk tumbuhan sagu fase masak panen dan yang telah melampaui fase masak panen (pohon veteran). Hal ini berarti bahwa sekitar 60 % rumpun sagu yang memiliki fase pohon. Dengan kata lain bahwa sekitar 40 % rumpun tumbuhan sagu di P. Seram tidak terdapat fase pohon.
Terlihat pada Gambar 17 bahwa dalam struktur populasi tumbuhan sagu, fase tiang memiliki jumlah populasi paling sedikit hanya sekitar 30,3 ind/ha, atau hanya 17,2 % dari jumlah populasi rumpun yang terbentuk. Sedangkan fase sapihan mencapai 49,3 %, dan pada fase semai jumlah populasinya paling tinggi yakni mencapai dua kali lipat dari jumlah rumpun. Hal ini menunjukkan bahwa pada setiap rumpun tumbuhan sagu dapat ditemukan lebih dari satu individu fase semai, bahkan di lapangan untuk jenis tanaman sagu tuni dan sylvestre ditemukan ada yang dapat mencapai lebih dari 10 individu semai (rata-rata 6-8), tetapi pada fase yang lain tidak selalu terdapat pada setiap rumpun.
Keterangan : PBP = pohon belum panen, PP = pohon panen, PV = pohon veteran. Data yang disajikan berasal dari data rataan wil. Sampel I Luhu SBB, II Sawai MT, dan III Werinama SBT, tahun 2009. Gambar 17. Populasi sagu menurut jumlah rumpun dan fase pertumbuhan
4.2.2. Kelimpahan
Kelimpahan sagu (Metroxylon spp.) dalam uraian ini diletakkan pada kelimpahan menurut spesies yang dipahami secara umum. Parameter yang dipergunakan untuk menjelaskan kelimpahan jenis adalah menurut jumlah rumpun dan indeks nilai penting (INP). Dari lima spesies sagu yang ditemukan di P. Seram yaitu M. rumphii Mart., M.sylvestre Mart., M. longispinum Mart., M.
microcanthum Mart. dan M. sagu Rottb. Khusus spesies M. microcanthum Mart.
tidak diuraikan dalam pembahasan dikarenakan jumlah individunya sangat terbatas yakni tidak mencapai satu persen.
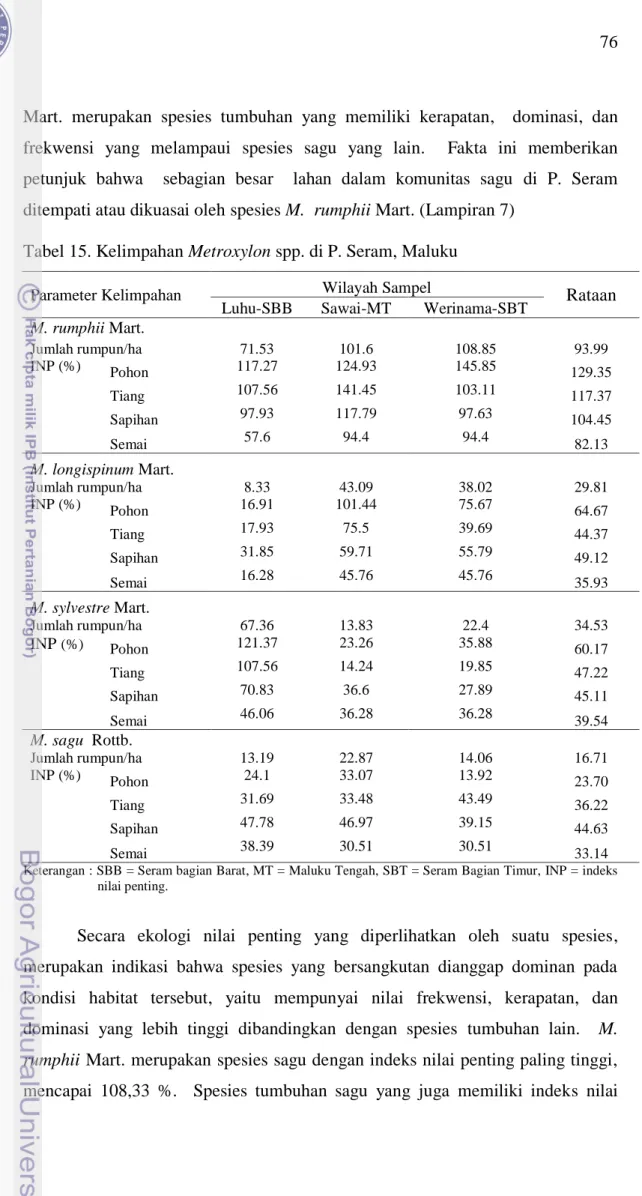
Hasil perhitungan jumlah populasi rumpun sagu dan indeks nilai penting empat spesies sagu menunjukkan bahwa M. rumphii Mart. memiliki jumlah individu paling banyak dan indeks nilai penting paling tinggi diantara semua spesies sagu, bahkan diantara semua spesies tumbuhan dalam komunitas sagu di P. Seram (Tabel 15 dan Gambar 18). Hal ini berarti bahwa spesies M. rumphii
176,6 335,71 87,1 30,27 80,5 19,08 6,48 0 50 100 150 200 250 300 350 400
Rumpun Semai Sapihan Tiang PBP PP PV
P o pul as i sa gu (i n d/h a) Fase Pertumbuhan
Mart. merupakan spesies tumbuhan yang memiliki kerapatan, dominasi, dan frekwensi yang melampaui spesies sagu yang lain. Fakta ini memberikan petunjuk bahwa sebagian besar lahan dalam komunitas sagu di P. Seram ditempati atau dikuasai oleh spesies M. rumphii Mart. (Lampiran 7)
Tabel 15. Kelimpahan Metroxylon spp. di P. Seram, Maluku
Parameter Kelimpahan Wilayah Sampel Rataan
Luhu-SBB Sawai-MT Werinama-SBT
M. rumphii Mart. Jumlah rumpun/ha 71.53 101.6 108.85 93.99 INP (%) Pohon 117.27 124.93 145.85 129.35 Tiang 107.56 141.45 103.11 117.37 Sapihan 97.93 117.79 97.63 104.45 Semai 57.6 94.4 94.4 82.13 M. longispinum Mart. Jumlah rumpun/ha 8.33 43.09 38.02 29.81 INP (%) Pohon 16.91 101.44 75.67 64.67 Tiang 17.93 75.5 39.69 44.37 Sapihan 31.85 59.71 55.79 49.12 Semai 16.28 45.76 45.76 35.93 M. sylvestre Mart. Jumlah rumpun/ha 67.36 13.83 22.4 34.53 INP (%) Pohon 121.37 23.26 35.88 60.17 Tiang 107.56 14.24 19.85 47.22 Sapihan 70.83 36.6 27.89 45.11 Semai 46.06 36.28 36.28 39.54 M. sagu Rottb. Jumlah rumpun/ha 13.19 22.87 14.06 16.71 INP (%) Pohon 24.1 33.07 13.92 23.70 Tiang 31.69 33.48 43.49 36.22 Sapihan 47.78 46.97 39.15 44.63 Semai 38.39 30.51 30.51 33.14
Keterangan : SBB = Seram bagian Barat, MT = Maluku Tengah, SBT = Seram Bagian Timur, INP = indeks nilai penting.
Secara ekologi nilai penting yang diperlihatkan oleh suatu spesies, merupakan indikasi bahwa spesies yang bersangkutan dianggap dominan pada kondisi habitat tersebut, yaitu mempunyai nilai frekwensi, kerapatan, dan dominasi yang lebih tinggi dibandingkan dengan spesies tumbuhan lain. M.
rumphii Mart. merupakan spesies sagu dengan indeks nilai penting paling tinggi,
penting cukup tinggi yaitu M. sylvestre Mart. 48,71 % dan M. longispinum Mart. 51,77 %. Sedangkan dua spesies sagu yang lain indeks nilai pentingnya < 35 %. Dengan demikian dapat dikemukakan bahwa kemampuan menguasai ruang spesies M. rumphii Mart. dua kali lebih besar dibandingkan sagu M. sylvestre Mart. dan M. longispinum Mart. Sedangkan kemampuan menguasai ruang spesies
M. rumphii Mart. apabila dibandingkan dengan M. sagu Mart. tiga kali lebih
tinggi. Secara keseluruhan kemampuan spesies sagu di P. Seram dalam menguasai ruang dalam komunitas sagu alami mencapai 70-90 %, khusus spesies
M. rumphii Mart. kemampuan menempati ruangnya sekitar 43,12 %.
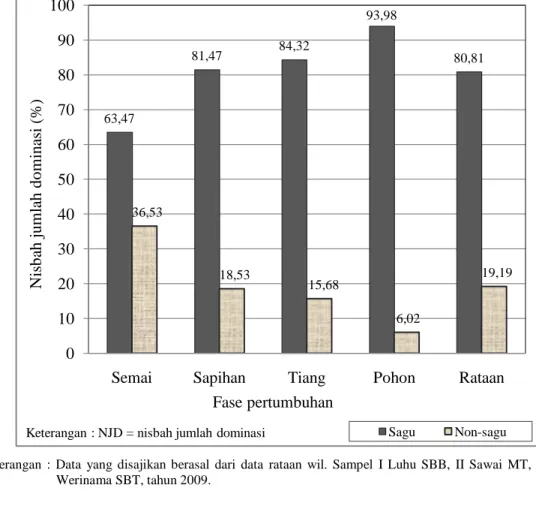
Keterangan : Data yang disajikan berasal dari data rataan wil. Sampel I Luhu SBB, II Sawai MT, dan III Werinama SBT, tahun 2009.
Gambar 18. NJD spesies sagu dan non-sagu di P. Seram, Maluku
Pada Gambar 18 diperlihatkan nisbah jumlah dominasi (NJD = INP/3) spesies sagu secara ekologi merupakan spesies dominan dalam menguasai habitat. Sedangkan spesies tumbuhan lain (bukan sagu) memiliki nilai NJD yang rendah. Fenomena seperti ini merupakan gambaran umum yang dijumpai pada tipe
63,47 81,47 84,32 93,98 80,81 36,53 18,53 15,68 6,02 19,19 0 10 20 30 40 50 60 70 80 90 100
Semai Sapihan Tiang Pohon Rataan
N is b ah j u m la h d o m in as i (% ) Fase pertumbuhan
vegetasi yang mengarah kepada kondisi klimaks dan stabil. Menurut Mueller dan Ellenberg (1974 dalam Setiadi 2005) dikemukakan bahwa komposisi bervegetasi hutan alami yang telah terbentuk dalam jangka panjang akan memperlihatkan fisiognomi, fenologi, dan daya regenerasi yang lambat dan cenderung mantap, sehingga dinamika floristik komunitas hutan tidak terlalu nyata dan menyolok. Dalam konteks ini pergantian generasi atau regenerasi spesies seakan-akan tidak tampak, akibatnya jarang dijumpai spesies tertentu yang kemudian muncul dominan, karena semua spesies telah beradaptasi dalam jangka waktu lama.
4.2.3. Pertumbuhan, perkembangbiakan, dan mekanisme pembentukan rumpun
a. Pertumbuhan
Dalam uraian untuk menjelaskan pertumbuhan sagu, maka siklus pertumbuhan sagu dipisahkan atas beberapa fase pertumbuhan menurut kriteria sebagaimana yang telah dikemukakan sebelumnya, yaitu fase semai (seedling). sapihan (sapling), tiang (pole), dan pohon (trees).
Fase semai diawali dari keluarnya tunas sampai batang utama terbentuk (masa roset). Pada saat muncul tunas anakan, keluar satu tangkai daun, disusul dengan tangkai daun berikutnya. Setelah terbentuk anakan sempurna jumlah tangkai daun berkisar antara 3-4 tangkai dengan panjang sekitar 50-80 cm. Anakan yang terus membesar, diikuti dengan bertambahnya jumlah tangkai daun antara 5-6 tangkai, dengan panjang antara 1-2 m. Menjelang masa roset, jumlah tangkai daun berkisar antara 6-8 tangkai dengan panjang sekitar 3-4 m, dan setelah masa roset jumlah tangkai daun berkisar antara 7-9 tangkai, dengan panjang dapat mencapai 5 m. Anakan sagu yang keluar dari bagian pangkal batang memiliki jumlah yang bervariasi, tergantung jenisnya. Spesies M. rumphii Mart. dan M. sylvestre Mart. biasanya memiliki jumlah anakan yang lebih banyak mencapai 7-8 anakan. Sedangkan spesies sagu yang lain relatif lebih sedikit, jumlah anakan spesies M. longispinum Mart. sekitar 6-7 anakan, M. sagu Rottb. 3-4 anakan, dan yang paling sedikit ditemukan pada spesies M. microcanthum Mart. hanya sekitar 2-3 anakan.
Tunas anakan sagu yang muncul tidak semuanya sukses tumbuh menjadi anakan fase bibit (fase yang lebih besar). Sebagian besar mengalami kematian selama masa pertumbuhan awal. Berdasarkan letak tunas anakan sagu pada bagian pangkal batang, ditemukan dua tipe yaitu : 1) tunas anakan yang keluar dari bagian pangkal batang di bawah permukaan tanah atau sejajar permukaan tanah, dan 2) letak tunas anakan beberapa centimeter dari atas permukaan tanah. Tunas anakan ini biasanya gagal untuk tumbuh dan berkembang ke fase selanjutnya.
Selain dari bagian pangkal batang, tunas anakan sagu muncul pula dari rhyzome yang memanjang. Pada rhyzome ini, tunas anakan keluar dari bagian bawah atau sejajar permukaan tanah dan ada pula yang keluar dari bagian atas, tidak bersinggungan dengan permukaan tanah. Tunas anakan sagu yang disebut terakhir paling rentan terhadap kematian (Gambar 19). Sejak mulai muncul tunas anakan pada pangkal batang, sampai terbentuk batang di permukaan tanah (masa roset) diperlukan waktu sekitar tiga tahun.
Gambar 19. Anakan sagu yang tumbuh pada bagian atas rhyzome
Setelah terbentuk batang pokok di permukaan tanah, pertumbuhan sagu masuk pada fase berikutnya yaitu fase sapihan (sapling). Fase ini ditandai
dengan mulai terbentuk batang sempurna sampai tinggi batang mencapai ukuran sekitar dua meter. Masa pertumbuhan selama fase ini diperlukan waktu sekitar tiga tahun (Sjachrul 1993). Dengan demikian masa pertumbuhan sejak muncul tunas anakan sampai akhir fase ini memerlukan waktu sekitar enam tahun. Pada fase ini jumlah pelepah daun berkisar antara 8-12 tangkai dengan panjang sekitar 5-6 meter. Pada spesies M. rumphii dan M. sylvestre panjang pelepah daun dapat mencapai 7-7,5 meter, dengan jumlah anak daun dapat mencapai 140 helai. Pada fase ini tumbuhan sagu sudah mulai menghasilkan tunas anakan.
Setelah mencapai umur lebih dari enam tahun, tumbuhan sagu masuk ke fase tiang (pole). Fase ini berlangsung sekitar tiga tahun, dan selama masa periode fase ini terjadi pertambahan tinggi batang yang bertambah sekitar tiga meter, sehingga total tinggi batang sampai dengan akhir fase tiang sekitar lima meter (Sjachrul 1993). Sejak mulai muncul tunas anakan sampai berakhir fase ini dibutuhkan waktu sekitar sembilan tahun. Sejak fase ini sering kali terjadi pertambahan diameter batang, sehingga ukuran batang relatif lebih besar daripada ukuran batang sagu di bagian pangkal. Selama fase tiang jumlah pelepah daun berkisar antara 8-12 tangkai dengan jumlah anak daun dapat mencapai 180 helai. Panjang pelepah daun dapat mencapai 7,5–8,0 meter, bahkan pada jenis sagu sylvestre yang pertumbuhannya bagus, panjang pelepah daun dapat mencapai 9-10 meter. Pada akhir fase ini tinggi batang sagu telah mencapai lima meter.
Setelah mencapai umur sekitar sembilan tahun, maka tumbuhan sagu telah masuk ke fase pohon (trees) . Pada fase ini tinggi batang sagu telah mencapai lebih dari lima meter (Sjachrul 1993). Terdapat jenis sagu tertentu seperti spesies rotang dan makanaro sudah dapat dipanen pada tinggi batang mencapai 7-8 meter. Dari fase tiang sampai mencapai ukuran tinggi batang tersebut diperlukan waktu sekitar 3-4 tahun. Dengan demikian tumbuhan sagu secara alami baru dapat dipanen setelah mencapai umur paling kurang 12 tahun . Pada jenis-jenis sagu yang memiliki ukuran tinggi pohon antara 15-20 meter, diperkirakan untuk sampai pada masa panen diperlukan masa pertumbuhan sekitar 12-15 tahun. Jenis-jenis sagu yang dapat mencapai ukuran tinggi tersebut seperti spesies M.
rumphii Mart., M. sylvestre Mart., dan M. sagu Rottb. Selama fase ini jumlah
meter, dan jumlah anak daun mencapai 200 helai. Meskipun demikian, menjelang masa pembungaan ukuran pelepah daun dan anak-anak daun mulai memendek, dan pada masa pembungaan jumlah pelepah daun tidak lagi mengalami penambahan.
b. Perkembangbiakan dan mekanisme pembentukan rumpun
Tumbuhan sagu berkembangbiak dengan menggunakan organ generatif dan vegetatif. Perkembangbiakan dengan organ generatif menggunakan buah atau biji. Di dalam buah terdapat biji yang dapat berkecambah, tumbuh, dan berkembang membentuk individu baru. Agen penyebaran sagu melalui organ buah/biji ini diduga terjadi melalui perantaraan air, karena buah sagu dilengkapi dengan sabut yang dapat menjadikan buah sagu terapung, kemudian terbawa mengikuti arus pergerakan air. Buah sagu yang jatuh di sekitar pohon, bila terjadi hujan dan tinggi muka air meningkat, maka buah sagu akan terapung, kemudian terbawa dari bagian muara sungai atau bagian kaki bukit terangkut mengikuti arus genangan ke bagian pesisir. Apabila aliran genangan masuk sampai ke bagian pesisir pantai, maka buah sagu akan terbawa arus ke berbagai tempat dan memungkinkan terdampar pada sisi pesisir pulau lain. Jika kondisi lingkungan mendukung untuk perkecambahan biji, maka dari biji akan tumbuh anakan dan seterusnya menjadi generasi baru di tempat tersebut.
Agen penyebaran buah sagu yang lain seperti melalui perantaraan hewan, menurut hemat penulis kemungkinan itu sangat kecil dikarenakan tidak terdapat bagian buah atau biji yang disukai sebagai makanan hewan liar. Argumen ini memiliki kelemahan karena penulis tidak memiliki fakta empiris atau sandaran hasil-hasil penelitian yang pernah dilakukan sebelumnya.
Dalam melakukan observasi atau penelitian lapangan di P. Seram selama lebih kurang enam bulan sejak Maret-Agustus 2009, tidak ditemukan fakta adanya biji sagu yang berkecambah di bawah tegakan pohon yang tumbuh dengan kerapatan tinggi. Pada tempat yang lain di sekitar hutan sagu dimana terjadi pembukaan hutan oleh masyarakat petani pengolah sagu, ditemukan biji-biji sagu yang berkecambah dengan baik dalam jumlah terbatas (Gambar 20). Fakta ini menunjukkan bahwa biji sagu akan mengalami hambatan perkecambahan apabila
tidak mendapat penyinaran surya yang cukup atau tidak mendapat penyinaran surya secara langsung. Hal ini berarti bahwa untuk mematahkan dormansi biji sagu diperlukan intensitas sinaran surya yang cukup. Apabila intensitas sinaran surya tidak terpenuhi, maka biji sagu akan tetap mengalami dormansi atau gagal berkecambah.
Gambar 20. Pertumbuhan anakan sagu melalui biji
Kueh (1977 dalam Flach 1983) melaporkan hasil penelitian perkecambahan biji sagu yang berasal dari Serawak, sebagian besar biji tidak dapat berkecambah, terdapat beberapa biji yang berkecambah namun memiliki mutu yang rendah. Disebutkan bahwa kegagalan perkecambahan itu kemungkinan karena biji-biji sagu tersebut tidak mendapatkan cahaya yang cukup atau cadangan makanan telah berkurang karena tersimpan dalam waktu yang cukup lama.
Cara perkembangbiakan sagu yang kedua adalah melalui organ vegetatif berupa anakan, stolon, atau rhyzome yang muncul dari bagian pangkal batang. Anakan sagu biasanya muncul dari pangkal batang di bagian bawah permukaan tanah, sejajar permukaan tanah, atau tidak menyentuh permukaan tanah. Tipe anakan yang disebut terakhir pada umumnya gagal untuk tumbuh menjadi individu baru ke fase pertumbuhan berikutnya. Agen penyebaran sagu dengan
organ ini yang paling banyak terjadi melalui perantaraan manusia terutama dalam praktek budidaya.
Dalam perkembangbiakan secara vegetatif ini, apabila anakan sagu tumbuh berdekatan dengan pohon induk, maka rumpun terbentuk memiliki jumlah individu yang relatif banyak. Di lapangan ditemukan rumpun sagu spesies tertentu seperti M. rumphii Mart. dan M. sylvestre Mart. dapat terdiri dari 15-20 individu. Rumpun sagu dengan jumlah individu sebanyak itu memiliki kemampuan penguasaan ruang dapat mencapai 10 m2. Pertumbuhan rhyzome (percabangan basal) yang memanjang menjauh dari pohon induk, terpisah dari rumpunnya pada jarak antara 1,5 – 2,0 m dapat tumbuh dan berkembang membentuk rumpun sendiri (Gambar 21). Dalam mekansime pembentukan rumpun ini, percabangan basal tumbuh membesar, setelah mencapai fase sapihan mulai diikuti dengan terbentuknya tunas. Tunas-tunas ini selanjutnya tumbuh membesar, secara bersama-sama dengan pohon induk membentuk rumpun baru.
Gambar 21. Percabangan basal yang menjauh dari pohon induk
Percabangan basal akan mengalami pemanjangan, menjauh dari pohon induk atau rumpun apabila terdapat hambatan dalam pertumbuhannya, atau
terdapat kondisi yang tidak memungkinkan untuk segera tumbuh vertikal membentuk individu baru. Hambatan yang dapat menyebabkan percabangan basal memanjang jauh dari pohon induk antara lain adalah adanya tumpukan tangkai daun kering yang jatuh berserakan. Tumpukan ini memunculkan hambatan sinaran surya yang dapat memacu pergerakan tunas ke arah vertikal sebagai bagian dari mekansime fototropisme. Dengan kata lain terjadi hambatan terhadap sinaran surya sehingga pemunculan tunas ke arah vertikal terhambat, sehingga pada periode tertentu tumbuh secara horizontal, setelah cahaya cukup kemudian tumbuh secara vertikal. Dalam perkembangan selanjutnya diikuti dengan pertumbuhan tunas-tunas baru membentuk suatu rumpun lagi.
4.2.4. Sifat morfologi sagu
Sagu (Metroxylon spp.) merupakan jenis tumbuhan palem tropika basah, apabila diamati secara morfologi memiliki bentuk arsitektur pohon model Tomlinson. Model arsitektur ini dicirikan oleh adanya beberapa sumbu yang terbentuk dari percabangan basal, yaitu percabangan yang berkembang dari munculnya mata tunas di bagian pangkal batang, kemudian memanjang atau menjalar ke arah samping menyerupai akar (rhyzome) seterusnya membentuk individu baru (caulomere). Masing-masing kaulomer pada awalnya dihasilkan dari dasar batang yang semula jumlahnya hanya satu, terus bertambah muncul dari permukaan tanah (Halle & Oldeman 1975).
Tumbuhan sagu memiliki sistem perakaran yang serupa dengan perakaran jenis tumbuhan palem pada umumnya yaitu sistem akar serabut. Akar keluar dari hampir seluruh permukaan pangkal batang di bawah permukaan tanah, bahkan sampai beberapa sentimeter di atas permukaan tanah. Akar keluar secara padat menyebar memanjang ke arah horizontal dan vertikal. Perakaran tumbuhan sagu menyebar horizontal sampai radius 3-4 meter dari pangkal batang. Sedangkan jangkauan jelajah ke dalam tanah dapat lebih dari satu meter, namun akumulasi perakaran paling banyak terdapat pada kedalaman tanah sekitar 60 cm. Diameter akar rata-rata sekitar satu centimeter, berwarna coklat muda sampai coklat tua. Sepanjang akar terdapat bagian menyerupai buku, dengan jarak yang bervariasi antara 3-5 cm. Ujung akar pada umumnya berwarna putih kekuningan atau
kemerahan. Hampir pada seluruh permukaan akar tumbuh bulu-bulu akar dengan panjang antara 3-5 cm dan diameter 1,5-2,0 mm.
Batang sagu berbentuk bulat, ukuran diameter bervariasi, tergantung spesies. Spesies M. rumphii Mart. dan M. sylvestre Mart. memiliki ukuran diameter batang berkisar antara 55-75 cm, M. longispinum Mart. dan M.
microcanthum Mart. memiliki diameter sekitar 40-55 cm. Sedangkan spesies M. sagu Rottb. pada umumnya memiliki ukuran diameter sekitar 50-65 cm. Tinggi
batang sagu bebas daun juga bervariasi tergantung spesies. Spesies M. rumphii Mart. dan M. sylvestre Mart. memiliki ukuran tinggi batang antara 15-20 meter,
M. longispinum Mart. sekitar 10-12 meter, M. microcanthum Mart. 7-10 meter,
dan M. sagu Rottb. berkisar antara 12-15 meter. Susunan penampang batang dari bagian luar ke dalam terdiri dari kulit luar yang tipis, kulit keras, serat kasar, dan empulur. Pada sisi bagian luar batang terdapat tanda bekas pelepah daun. Jarak antara bekas pelepah yang satu dengan yang lain secara vertikal berkisar antara 10-15 cm. Warna empulur bervariasi tergantung jenis sagu mulai dari putih sampai kemerahan. Spesies M. rumphii Mart. mempunyai empulur berwarna putih agak kemerahan, M. sylvestre Mart. kemerahan, M. longispinum Mart. merah muda, M. microcanthum Mart. berwarna merah muda, dan M. sagu Rottb. berwarna putih. Menurut Haryanto dan Pangloli (1992) dikemukakan bahwa bahwa berat kulit batang sagu berkisar antara 17-25 % dari berat batang, sisanya berupa berat empulur sekitar 75-83 % dan perbandingan antara berat kulit batang dan empulur selama masa pertumbuhan sagu relatif tetap.
Daun tumbuhan sagu terdiri dari pelepah (tangkai daun), anak daun, tulang daun, dan spesies M. rumphii Mart. terdapat duri (spine) yang menempel pada hampir seluruh bagian pangkal pelepah. Duri-duri tersebut juga terdapat pada bagian belakang tangkai daun. Duri juga terdapat pada sisi pinggir anak daun berupa duri-duri halus. Pangkal pelepah daun sagu menempel mengitari batang. Di sepanjang tangkai daun tumbuh anak daun menyirip berhadapan atau agak berhadapan. Jarak tata letak anak daun pada tangkainya berkisar antara 5-10 cm. Pelapah atau tangkai daun berjumlah antara 12-16 tangkai dan memiliki ukuran panjang sekitar 5-7 m. Pada spesies M. rumphii Mart. dan M. sylvestre Mart. ukuran tangkai daun dapat mencapai delapan meter bahkan lebih. Setiap tangkai
daun dewasa terdapat sekitar 50-80 pasang anak daun. Anak daun memiliki ukuran panjang antara 100-180 cm, dan lebar 10-15 cm. Flach (1983) mengemukakan bahwa tanaman sagu memiliki tangkai mencapai 18 tangkai dengan ukuran antara 5-7 m. Setiap bulan terbentuk satu tangkai daun, dan umur rata-rata tangkai daun diperkirakan sekitar 12-18 bulan. Tangkai daun berwarna hijau muda, kemudian berubah warna menjadi hijau kekuningan dan selanjutnya tangkai dan anak-anak daun menguning, mengering dan gugur. Daun sagu yang masih muda pada umumnya berwarna hijau muda, kadang-kadang berwarna hijau keunguan. Dengan bertambah umur secara berangsur-angsur berubah warna menjadi hijau tua, kuning, dan coklat apabila telah mengering.
Tumbuhan sagu diperkirakan mulai berbunga pada umur sekitar 10-12 tahun, diikuti dengan pembentukan buah. Masa pembungaan diawali dengan munculnya tanda-tanda seperti tangkai daun dan anak-anak daun memendek, ukuran lebar menyempit dan pelepah tangkai daun menunjukkan perubahan warna menjadi hijau kekuningan. Tomlinson (1990 dalam Flach 1997) melakukan deskripsi pembungaan Metroxylon spp. dilaporkan bahwa palem ini melengkapi proses pertumbuhan dengan membentuk pembungaan, merupakan indikasi akan berakhirnya masa pertumbuhan yang diakhiri dengan kematian.
Bunga sagu merupakan bunga majemuk yang keluar dari ujung atau pucuk batang, berwarna merah kecoklatan. Bunga bercabang banyak seperti tanduk rusa yang terdiri dari cabang primer, sekunder, dan tersier. Pada cabang tersier terdapat sepasang bunga jantan dan bunga betina. Diduga penyerbukan tumbuhan sagu berlangsung secara silang (Flach 1977 dalam Haryanto dan Pangloli 1992). Buah sagu berbentuk bulat menyerupai buah salak. Kulit buah berupa sisik yang tersusun secara diagonal. Di dalam buah terdapat biji yang sifatnya fertil. Dikemukakan juga bahwa waktu antara bunga mulai muncul sampai fase pembentukan buah, berlangsung sekitar dua tahun.
4.2.5. Preferensi habitat dan adaptasi tumbuhan sagu
Secara umum tipe habitat sagu dapat dipisahkan menjadi dua kategori yaitu 1) tipe habitat lahan kering dan 2) tipe habitat lahan tergenang, berupa rawa-rawa yang tergenang secara temporer maupun permanen. Tipe habitat kedua atas
dasar karakteristiknya dapat dipisahkan lebih lanjut menjadi beberapa tipe habitat yaitu : 1) tipe habitat tergenang air payau yaitu tipe habitat yang dicirikan oleh adanya pasang-surut sehingga genangannya bersifat temporer, merupakan habitat yang berdekatan atau berbatasan dengan vegetasi nipah (mangrove). Pada umumnya terdapat di bagian belakang nipah, dari bagian pesisir ke arah daratan. Tumbuhan sagu pada tipe habitat ini biasanya mengalami perendaman atau tergenang apabila terjadi air pasang, dan kondisi habitatnya mengering jika terjadi air surut, 2) tipe habitat tergenang sementara oleh air hujan yaitu tipe habitat dimana genangannya sangat ditentukan oleh ada-tidaknya hujan. Apabila terjadi hujan habitat sagu mengalami genangan selama beberapa waktu, pada umumnya sekitar satu sampai dua minggu atau paling lama satu bulan. Apabila tidak terjadi hujan maka kondisi habitatnya mengering, 3) tipe habitat tergenang permanen, yaitu tipe habitat sagu yang mengalami genangan pada periode waktu relatif cukup lama, biasanya lebih dari satu bulan. Air genangan bisa berasal dari air hujan atau air sungai, dan 4) tipe habitat lahan kering, artinya kondisi habitat sagu tidak pernah mengalami genangan air, apakah dari air hujan, sungai atau air laut. Kondisi lahan pada tipe habitat ini pada umumnya kemiringan lahan agak datar, sehingga tidak memungkinkan air sungai, air laut ataupun air hujan yang jatuh tidak menyebabkan genangan tetapi mengalami run off masuk ke sungai atau kolam yang dapat menampung sejumlah air, seringkali masuk ke tipe habitat tergenang tidak permanen air tawar atau ke tipe habitat permanen (Lampiran 8).
Di dalam wilayah P. Seram terdapat lima spesies sagu yaitu : 1) M.
rumphii Mart., 2) M. sylvestre Mart., 3) M. longispinum Mart., 4) M. microcanthum Mart., dan 5) M. sagu Rottb. Hasil penelitian menunjukkan bahwa
tidak semua spesies sagu dapat tumbuh pada setiap tipe habitat. Dari lima spesies tumbuhan sagu yang tumbuh dan berkembang dalam wilayah P. Seram, hanya tiga spesies sagu yang ditemukan tumbuh pada semua tipe habitat yaitu M.
rumphii Mart., M. sylvestre Mart. dan M. longispinum Mart. (Tabel 16). Dua
spesies tumbuhan sagu yang lain yakni M. microcanthum Mart. dan M. sagu Rottb. ditemukan pada tipe habitat terbatas. Spesies M. microcanthum Mart. hanya ditemukan tumbuh pada tipe habitat lahan kering (TTG), sedangkan spesies
air tawar (T2AT) dan tergenang permanen (TPN). Hal ini menunjukkan bahwa tiga spesies tumbuhan sagu yang disebutkan pada bagian awal memiliki kemampuan untuk berinteraksi dengan berbagai tipe habitat, sedangan dua spesies yang lain memiliki kemampuan untuk berinteraksi dengan tipe habitat yang relatif terbatas.
Tabel 16. Populasi rumpun dan fase pertumbuhan sagu pada tipe habitat berbeda di P. Seram, Maluku tahun 2009
No Spesies sagu
Tipe Habitat
Rataan
TTG T2AT T2AP TPN
ind/ha % ind/ha % ind/ha % ind/ha % ind/ha % Populasi rumpun 1. M. rumphii 103,26 58,9 124,33 50,1 62,08 64,2 61,20 37,3 87,72 52.6 2. M. longisp. 28,37 15,8 26,01 10,5 20,00 20,7 36,04 22,0 27,60 17.2 3. M.sylvestre 37,95 22,9 85,10 34,3 14,58 15,1 11,58 7,1 37,30 19.8 4. M. microc. 4,27 2,5 0,00 0,0 0,00 0,0 0,00 0,0 1,07 0.6 5. M. sagu 0,00 0,0 12,50 5,0 0,00 0,0 55,19 33,7 16,92 9.7 Jumlah 173.85 100,0 247,94 100,0 96,67 100,0 164,0 100,0 170,6 100,0 Populasi semai 1. M. rumphii 186,35 54,00 195,56 46,9 159,38 65,6 90,69 34,3 158,0 50.2 2. M. longisp. 53,4 15,5 49,74 11,9 40,00 16,5 77,24 29,2 55,1 18.3 3. M.sylvestre 96,46 27,9 157,71 37,8 43,75 18,0 41,24 15,6 84,79 24.8 4. M. microc. 9,06 2,6 0,00 0,0 0,00 0,0 0,00 0,0 2,27 0.7 5. M. sagu 0,00 0,0 13,89 3,3 0,00 0,0 55,63 21,0 17,38 6.1 Jumlah 345.28 100,0 416,90 100,0 162,08 100,0 264,8 100,0 317,5 100,0 Populasi sapihan 1. M. rumphii 25,97 57,0 25,64 45,1 24,38 73,6 16,77 27,6 23,19 50.8 2. M. longisp. 6,46 14,2 7,68 13,5 7,50 22,6 17,48 28,8 9,78 19.8 3. M.sylvestre 11,01 24,2 18,88 33,2 1,25 3,8 3,30 5,4 8,61 16.7 4. M. microc. 2,12 4,7 0,00 0,0 0,00 0,0 0,00 0,0 0,53 1.2 5. M. sagu 0,00 0,0 4,63 8,2 0,00 0,0 23,16 38,2 6,95 11.6 Jumlah 45.56 100,0 56,83 100,0 33,13 100,0 60,71 100,0 49,06 100,0 Populasi tiang 1. M. rumphii 10,66 58,8 11,04 47,4 2,50 60,0 8,50 33,8 8,17 50.0 2. M. longisp. 1,11 6,1 3,57 15,3 1,67 40,0 8,06 32,1 3,60 23.4 3. M.sylvestre 5,80 32,0 6,39 27,4 0,00 0,0 0,00 0,00 3,05 14.9 4. M. microc. 0,56 3,1 0,00 0,0 0,00 0,0 0,00 0,00 0,14 0.8 5. M. sagu 0,00 0,0 2,31 9,9 0,00 0,0 8,55 34,0 2,72 11.0 Jumlah 18.13 99,9 23,32 99,9 4,17 99,9 25,11 99,9 17,68 99,9 Populasi pohon 1. M. rumphii 57,36 55,7 41,55 43,0 15,00 72,0 24,95 26,6 34,71 49.3 2. M. longisp. 12,95 12,6 14,73 15,3 5,83 28,0 22,84 25,3 14,09 20.3 3. M.sylvestre 29,10 28,2 33,38 34,6 0,00 0,0 4,92 6,9 16,85 17.4 4. M. microc. 3,65 3,5 0,00 0,0 0,00 0,0 0,00 0,0 0,91 0.9 5. M. sagu 0,00 0,0 6,94 7,2 0,00 0,0 29,44 41,3 9,1 12.1 Jumlah 103,06 100,0 96,60 100,0 20,83 100,0 82,14 100,0 75,66 100,0
… lanjutan
Populasi pohon masak panen
1. M. rumphii 8,65 50,4 8,09 32,6 0,83 33,3 7,3 26,2 6,22 35.6 2. M. longisp. 5,42 28,1 6,62 26,7 1,67 66,7 9,73 34,9 5,86 39.1 3. M.sylvestre 3,89 21,5 7,48 30,1 0,00 0,0 0,00 0,0 2,84 12.9 4. M. microc. 0,00 0,0 0,00 0,0 0,00 0,0 0,00 0,0 0,00 0.0 5. M. sagu 0,00 0,0 2,63 10,6 0,00 0,0 10,82 38,9 3,36 12.4 Jumlah 17,95 100,0 24,82 100,0 2,50 100,0 27,85 100,0 18,28 100,0 Populasi pohon veteran
1. M. rumphii 1,63 30,1 0,44 6,9 0,00 0,0 4,44 24,3 1,63 15.3 2. M. longisp. 3,23 59,6 2,83 44,7 0,50 100 8,46 46,2 3,76 62.6 3. M.sylvestre 0,56 10,3 0,44 6,9 0,00 0,0 0,76 4,1 0,44 5.3 4. M. microc. 0,00 0,0 0,00 0,0 0,00 0,0 0,00 0,0 0,00 0.0 5. M. sagu 0,00 0,0 2,63 41,5 0,00 0,0 4,65 25,4 1,82 16.7 Jumlah 5,42 100,0 6,34 100,0 0,50 100,0 18,32 100,0 7,64 100,0 Keterangan : M. longisp = M. longispinum, M. micrc = M. microcanthum, ind = individu, ha = hektar, TTG = lahan
kering, T2AT = tergenang temporer air tawar, T2AP = tergenang temporer air payau, TPN = tergenang permanen. Data yang disajikan berasal dari data rataan wil. Sampel I Luhu SBB, II Sawai MT, dan III Werinama SBT, tahun 2009.
Dalam kaitan dengan habitat tergenang, Levitt (1980) mengemukakan bahwa penggenangan dapat memunculkan tiga macam cekaman secara berurutan yaitu : 1) cekaman tekanan turgor sebagai akibat potensi air meningkat, 2) cekaman defisit oksigen, dan 3) cekaman ionik oleh unsur mangan (Mn2+) dan besi (Fe2+). Jika terjadi penggenangan, mula-mula memunculkan cekaman air yakni peningkatan cekaman turgor, diikuti dengan cekaman sekunder berupa kekurangan oksigen dalam air, implikasi berikut adalah terjadi cekaman ionik. Ketika oksigen di dalam air berkurang, maka potensial oksidasi-reduksi menurun, implikasi selanjutnya adalah terjadi akumulasi Mn2+ dan Fe2+ yang bersifat meracun (toxic). Tumbuh-tumbuhan pada kondisi cekaman karena genangan, maka akan menciptakan resistensi (katahanan) melalui penghindaran (avoidance) dan toleransi (tolerance). Penghindaran terhadap cekaman defisit oksigen terjadi melalui pembesaran ruang antar sel (intercellular space) misalnya dengan meningkatkan volume perakaran yang dapat mencapai 70 %. Sedangkan toleransi terhadap cekaman berlangsung melalui penghindaran terhadap akumulasi senyawa yang bersifat toxic atau toleransi terhadap akumulasi senyawa itu. Skema adaptasi tumbuhan pada kondisi tergenang yang menyebabkan cekaman terhadap defisit oksigen tersaji pada gambar berikut (Gambar 22).
Apabila interaksi tumbuhan sagu dengan tipe habitat ini dijadikan ukuran atau acuan untuk menjelaskan kemampuan adaptasi tumbuhan sagu terhadap tipe
habitatnya, tampak bahwa kemampuan adaptasi diantara spesies sagu dengan tipe habitat relatif berbeda. M. rumphii Mart. secara keseluruhan mempunyai kemampuan adaptasi yang kuat pada semua tipe habitat, yang ditunjukkan melalui jumlah rumpun dan jumlah populasi semua fase pertumbuhan yang lebih tinggi dibandingkan spesies sagu lain.
Gambar 22. Diagram ketahanan tumbuhan terhadap kondisi defisit oksigen (Levitt 1980)
Selain spesies M. rumphii Mart., dua spesies sagu yang juga mempunyai kemampuan adaptasi yang kuat dengan tipe habitatnya adalah spesies M. sylvestre Mart. dan M. longispinum Mart. Spesies M. sylvestre Mart. mempunyai kemampuan adaptasi yang kuat pada tipe habitat lahan kering (TTG) dan tergenang temporer air tawar (T2AT), yang ditunjukkan oleh jumlah populasi rumpun dan fase pertumbuhan yang lebih tinggi dibandingkan dengan spesies M.
longispinum Mart. Sedangkan pada tipe habitat tergenang temporer air payau
(T2AP) dan tergenang permanen (TPN) populasi spesies M. sylvestre Mart. tidak lebih tinggi daripada M. microcanthum Mart. Dengan kata lain pada dua tipe habitat yang disebut terakhir ini, spesies M. longispinum Mart. mempunyai kemampuan adaptasi yang lebih baik dibandingkan dengan spesies M. sylvestre Mart. Spesies M. sagu Rottb. hanya ditemukan tumbuh pada tipe habitat T2AT dan TPN. Hal ini menunjukkan bahwa spesies tumbuhan sagu ini memiliki daya adaptasi yang cukup baik pada kondisi habitat tergenang, tetapi tidak pada air payau karena pada tipe habitat tergenang tidak permanen air payau spesies sagu ini tidak ditemukan. Fakta ini memberikan petunjuk bahwa spesies M.
Ketahanan defisit oksigen
Penghindaran cekaman Toleransi cekaman
Perbanyak akar, menambah ruang antar sel, meningkatkan transport
gas dari tajuk
Toleransi terhadap akumulasi senyawa beracun Penghindaran terhadap akumulasi senyawa beracun
microcanthum Mart. memiliki daya adaptasi yang sangat terbatas atau sempit
terhadap berbagai tipe habitat.
Berdasarkan jumlah populasi tumbuhan sagu, tampak bahwa spesies sagu yang memiliki daya adaptasi yang tinggi sampai sempit terhadap berbagai kondisi habitat secara berurutan sebagai berikut M. rumphii Mart. > M. longispinum Mart. > M. sylvestre Mart. > M. sagu Rottb. > M. microcanthum Mart. Dalam hubungan ini, maka spesies M. rumphii Mart. dapat dikategorikan sebagai spesies sagu yang memiliki tingkat toleransi yang luas/lebar (eury tolerance) terhadap kondisi habitatnya. spesies tumbuhan sagu M. Longispinum Mart., M. Sylvestre Mart., dan M. sagu Rottb. dikategorikan sebagai jenis sagu dengan tingkat toleransi sedang (meso tolerance). Sedangkan spesies M. microcanthum Mart. dikategorikan sebagai spesis sagu yang memiliki tingkat toleransi sempit (steno
tolerance). Indikator untuk menjelaskan lebar atau sempitnya tingkat toleransi
masing-masing jenis ini didasarkan pada ada/atau tidak-adanya suatu spesies pada setiap habitat dan banyak atau sedikitnya jumlah populasi pada masing-masing habitat.
Dalam kaitan itu maka dapat dikemukakan bahwa spesies M. rumphii merupakan spesies tumbuhan sagu yang memiliki kemampuan pertumbuhan dan daya adaptasi yang tinggi terhadap berbagai tipe habitat. Spesies M. sylvestre Mart., M. longispinum Mart., dan M. sagu Rottb. merupakan spesies sagu yang dapat dikategorikan sebagai spesies yang memiliki kemampuan tumbuh tinggi tetapi daya adaptasi yang terbatas. Sedangkan spesies M. microcanthum Mart. merupakan spesies tumbuhan sagu yang memiliki kemampuan pertumbuhan dan daya adaptasi yang lebih terbatas lagi.
Apabila tipe habitat ini diurutkan ke dalam tingkat marjinalisasi habitat, terkait dengan jumlah populasi rumpun masing-masing spesies sagu pada setiap tipe habitat, maka akan didapatkan urutan marjinalisasi habitat sebagai berikut : tergenang temporer air payau (T2AP) > tergenang permanen (TPN) > lahan kering (TTG) > tergenang temporen air tawar (T2AT). Artinya tipe habitat tergenang temporer air payau (T2AP) memiliki tingkat marjinal yang lebih tinggi dibandingkan dengan tingkat marjinal tipe habitat tergenang permanen (TPN), dan tipe habitat ini lebih marjinal dibandingkan dengan tipe habitat lahan kering
(TTG). Kondisi habitat yang sangat rendah tingkat marjinalnya adalah tipe habitat tergenang temporer air tawar (T2AT). Ukuran untuk menjelaskan tinggi atau rendahnya tingkat marjinalisasi ini didasarkan pada banyak atau sedikitnya jumlah populasi rumpun sagu yang tumbuh pada setiap tipe habitat. Pada tipe habitat tergenang temporer air payau, jumlah populasi rumpun sagu hanya mencapai 96,67 ind/ha, tipe habitat tergenang permanen 164,02 ind/ha, tipe habitat lahan kering 173,85 ind/ha, sedangkan tipe habitat tergenang temporer air tawar jumlah populasinya mencapai 247,94 ind/ha. Diagram ringkasan adaptasi sagu pada berbagai tipe habitat disajikan pada Gambar 23.
Keterangan : TTG = lahan kering; T2AT = tergenang temporer air tawar; T2AP = tergenang temporer air payau; TPN = tergenang permanen; secara horizontal tidak terdapat jenis yang sama. Gambar 23. Diagram ringkasan adaptasi spesies sagu pada berbagai tipe habitat
4.2.6. Mekanisme adaptasi sagu
Sebagian besar tumbuhan sagu tumbuh pada lahan yang terendam, baik sifatnya temporer maupun permanen. Pada kondisi habitat yang senantiasa tergenang tersebut memungkinkan kondisi tanah menjadi masam dan miskin oksigen. Hasil penelitian menunjukkan bahwa pH tanah di areal lahan sagu
Habitat TTG T2AT T2AP TPN M. rumphii M. microcanthum M. Sylvestre M. longispinum M. rumphii M. Sylvestre M. longispinum akanaro M. rumphii M. Sylvestre M. longispinum o M. rumphii M. Sylvestre M. longispinum M. sagu M. sagu
berkisar antara 4,47 – 5,63 (pH H2O), dan berpotensi turun lebih rendah lagi
mencapai 4,13 (pH KCl). pH masam pada umumnya bersifat melisis suatu zat sehingga dapat merusak didinding sel. Tanah-tanah masam dengan kandungan logam tinggi seperti Fe dan Al dapat berpengaruh buruk terhadap kehidupan tumbuhan. Syekhfani (1997) mengemukakan logam memiliki kemampuan untuk melisis air sehingga pH tanah dapat semakin masam. Pada saat pH dalam kondisi masam Fe dan Al akan larut sehingga konsentrasinya meningkat. Konsentrasi Fe dan Al yang tinggi dapat meracun perakaran, walaupun Fe merupakan unsur hara esensial, namun termasuk dalam kategori unsur hara mikro, sehingga kelebihannya tidak menguntungkan bagi perakaran.
Separoh atau sebagian besar habitat tumbuhan sagu adalah berupa rawa-rawa yaitu berupa tipe habitat tergenang, baik temporer atau permanen. Pada kondisi habitat seperti itu biasanya sistem perakaran sagu mengalami modifikasi bentuk untuk dapat beradaptasi dengan kondisi habitat tereduksi. Kondisi habitat tergenang atau berupa rawa-rawa, identik dengan kondisi tereduksi. Artinya keadaan dimana terjadi keterbatasan oksigen di dalam tanah karena oksigen atau udara terdesak oleh partikel air (H2O). Pada sisi yang lain untuk menjamin
pertumbuhan diperlukan oksigen untuk proses respirasi akar.
Dalam kaitan dengan kondisi yang tereduksi ini, maka sistem perakaran tumbuhan sagu mengalami modifikasi bentuk dan arah. Biasanya pergerakan akar senantiasa tumbuh ke samping secara horizontal dan vertikal ke lapisan tanah bagian dalam. Namun ketika kondisi tanah tergenang air, maka terdapat sebagian arah pertumbuhan akar sagu berbalik ke atmosfer keluar menembus permukaan air sehingga terjadi kontak langsung dengan udara bebas. Disamping itu jumlah atau volume akar rambut meningkat sehingga luas permukaan kontak bertambah besar. Mekanisme adaptasi sistem perakaran sagu seperti inilah yang ditemukan terjadi untuk memenuhi penyerapan aksigen melalui perluasan kontak permukaan akar dengan udara luar, sehingga kebutuhan oksigen sagu dapat terpenuhi selama terjadi genangan (Gambar 24).
Gambar 24. Modifikasi pertumbuhan akar sagu pada kondisi tergenang Modifikasi sistem perakaran ke arah permukaan air, atau melewati tinggi genangan ini, diduga agar supaya penyerapan oksigen oleh perakaran tumbuhan sagu dapat berlangsung dengan baik, yang dimaksudkan untuk dapat memenuhi kebutuhan oksigen. Mekanisme pergerakan ini selanjutnya disebut sebagai
oxytropisme, yaitu pergerakan akar sagu menuju tempat yang cukup tersedia
oksigen. Menurut Levitt (1980) dikemukakan bahwa lahan yang tergenang dalam tempo cukup lama memunculkan cekaman. Kondisi cekaman tidak menguntungkan bagi banyak jenis tumbuhan. Beberapa tumbuhan dalam menghadapi kondisi cekaman, secara alamiah terjadi pembentukan organ dalam jumlah banyak seperti rhyzome dan memperbanyak jumlah akar. Mekanisme inilah yang terjadi pada tumbuhan sagu untuk mempertahankan kehidupan pada kondisi tergenang. Daubenmire (1974) mengemukakan pula bahwa banyak tumbuhan untuk dapat beradaptasi dengan kondisi lahan yang memiliki aerase jelek, terjadi melalui dua mekanisme adaptasi yaitu : 1) melalui adaptasi morfologi seperti membentuk sistem perakaran dangkal, membentuk jaringan aerase khusus atau organ aerase tertentu, misalnya membentuk sistem ruang udara interseluler yang menghubungkan stomata dengan sistem parakaran, yang disebut