BALAI PENELITIAN TANAMAN KELAPA DAN PALMA LAIN, MANADO
Hama Sexava spp. (Orthoptera : Tettigoniidae) dapat menyebabkan kerusakan serius pada tanaman kelapa. Belalang Sexava terdiri dari empat spesies yaitu Sexava nubila Stal, Sexava coriacea Linnaeus, Sexava karnyi Leefmans dan Sexava novae-guineae Brancsik (Kalshoven, 1981: Lever, 1969; Tjoa,1953). Nimfa dan imago menyerang daun, bunga betina dan buah muda sehingga dapat mempengaruhi produksi kelapa. Beberapa teknik pengendalian sudah diterapkan tetapi sampai sekarang populasi hama ini masih merupakan hambatan utama dalam meningkatkan produksi kelapa di daerah sebaran hama Sexava spp. Penerapan pengendalian hama terpadu (PHT) merupakan salah satu solusi yang tepat untuk mengatasi masalah hama Sexava spp.
Biologi dan ekologi serangga merupakan salah satu unsur dasar PHT sebagai pengetahuan dasar yang harus diketahui, diperhatikan dan dipergunakan untuk menyusunan komponen pengendalian baik secara tunggal, maupun dalam perpaduannya di lapangan dengan komponen lain untuk memperoleh hasil pengendalian yang optimal. Tanpa pengetahuan tentang unsur-unsur dasar maka rekomendasi pengendalian yang disusun tidak akan dapat sesuai dengan prinsip dan tujuan PHT (Watson et al., 1975).
Pemahaman biologi dan ekologi hama Sexava spp. dapat membantu dalam pengambilan keputusan untuk melakukan pengendalian yang efektif dan efisien, untuk itu perlu diketahui deskripsi, daur hidup, daerah penyebaran, tanaman inang lain dan sejarah serangan.
.
Imago Sexava spp., biasanya berwarna hijau, coklat dan hijau kecoklatan (Gambar 1). Dari hasil pengamatan penulis terhadap populasi hama S. nubila di Talaud ternyata yang dominan (>90%) adalah warna hijau sedangkan di Seram dan Papua (Irian Jaya) yang dominan adalah warna coklat. Pada populasi S. coriacea di Maluku Utara, Sangihe Besar dan Bolaang Mongondow, Sulawesi Utara yang dominan adalah warna hijau. Ledakan populasi S. nubila di Talaud lebih sering dibandingkan dengan di Seram dan Papua.
Antena Sexava spp. berbentuk filiform, lebih panjang dari tubuhnya (long-horned
grasshoppers), dan dapat dibedakan dengan belalang dari famili Acrididae yang
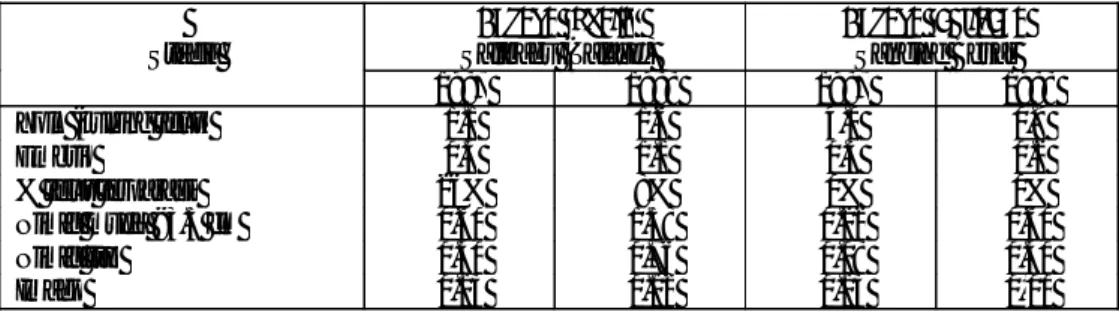
berantena pendek (short-horned grasshoppers). Imago betina memiliki alat peletak telur (ovipositor) yang besar dan berbentuk pedang sehingga hama ini disebut sebagai belalang pedang (Lever, 1969). Deskripsi untuk ketiga spesies Sexava yang ada di Indonesia adalah sebagai berikut:
Bioekologi Hama spp.(Orthoptera: Tettigoniidae)
2
Gambar 1. Variasi warna imago di Kepulauan Talaud: (a) imago betina berwarna hijau, (b) imago betina berwana coklat dan (c) imago jantan berwarna hijau kehitaman
1. Sexava nubila
Hama S. nubila dikenal dengan Belalang Talaud atau boto-boto. Hama ini makan anak daun mulai dari pingggir ke bagian tengah. Kadang-kadang dimakan sebagian atau sampai ke lidi. Bekas gigitan biasanya tidak rata. Serangan berat, terlihat pada pelepah daun bagian bawah tinggal lidi saja.
a
b
Bioekologi Hama spp. (Orthoptera: Tettigoniidae) Telur
Bentuk dan warna telur S. nubila seperti buah padi masak (gabah). Telur yang baru diletakkan sangat tipis dengan alur yang dalam (Gambar 2a, 2b1) kemudian embrio berkembang sehingga membengkak (Gambar 2b3). Telur berumur 2 hari, panjannya 12 mm dan lebarnya 2 mm. Salah satu ujung telur lancip dan lainnya bulat. Telur tua, panjangnya sampai 13 mm dan lebarnya 3 mm. Lama stadium telur di Talaud ± 50 hari (Tjoa, 1953), atau 45 hari (Warouw, 1981).
(a) (b1) (b2) (b3)
Gambar 2. Telur : telur baru diletakkan (a); telur bernas, embrio sudah berkembang dalam telur (b1); telur terparsit terlihat lobang tempat keluar parasitoid (b2); dan telur menetas, terlihat lobang tempat keluar nimfa instar pertama (b3).
Nimfa
Nimfa yang baru ditetaskan, panjangnya 12 mm dan bentuknya sama dengan
S. coriacea. Antenanya halus seperti rambut dan panjangnya sampai 9 cm. Nimfa muda
dan tua berwarna hijau, tetapi kadang-kadang berwarna coklat. Panjang nimfa jantan tua sampai 6 cm dan panjang antena 14 cm dan sudah terlihat bakal sayapnya. Lama stadium nimfa 70 hari (Tjoa, 1953), atau 108 hari (Warouw, 1981).
Belalang dewasa (Imago)
Imago berwarna hijau, antena merah muda dan matanya abu-abu. Bentuknya hampir sama dengan S. coriacea. Alat peletak telur (ovipositor) berwarna hijau pada bagian pangkalnya yaitu sepertiga dari panjang ovipositor, sepertiga lagi berwarna kemerahan dan bagian ujungnya berwarna hitam. Panjang imago betina (kepala + badan + ovipositor) antara 9.5 – 10.5 cm. Panjang ovipositor 3 – 4.5 cm dan panjang antena 16 cm. Panjang imago jantan 6 – 9.5 cm dan antenanya 14-16 cm.
Cara hidup
Imago betina terutama meletakkan telurnya pada malam hari di dalam tanah atau pasir dekat batang kelapa pada kedalaman 1 – 5 cm. Telur-telur diletakkan juga diantara perakaran kelapa, di bawah lumut, di sela-sela batang kelapa, dan di mahkota pohon kelapa yang kotor. Telur yang diletakkan di tanah dapat mencapai 95%. Tanah yang disukai oleh imago betina untuk meletakkan telur adalah tanah liat yang lembab bercampur pasir. Satu ekor imago betina yang dipelihara di laboratorium dapat
Bioekologi Hama spp.(Orthoptera: Tettigoniidae)
4
meletakkan telur sebanyak 53 butir. Pada setiap pohon kelapa terdapat berbagai stadia, mulai dari nimfa yang baru menetas sampai imago.
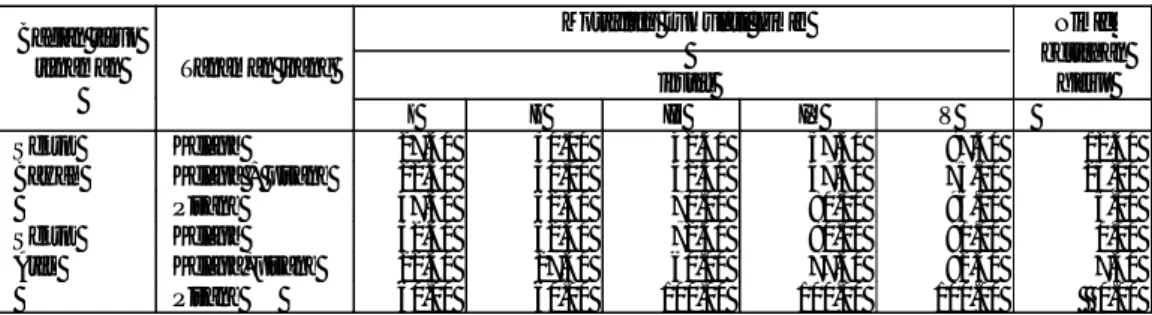
Daur hidup S. nubila, mulai telur diletakkan sampai imago meletakkan telur ± 5 bulan (150 hari) (Tjoa, 1953) atau 183 hari (Warouw, 1981) (Gambar 3). Tahap perkembangan hidup hama S. nubila telah diteliti secara detail oleh Warouw (1981) (Tabel 1). Imago betina turun kebawah pada malam hari untuk bertelur kemudian memanjat lagi pohon kelapa. Imago betina mulai melatakkan telur setelah berumur sekitar satu bulan. Imago Sexava tidak dapat terbang jauh, oleh karena itu serangga tersebut hanya terdapat ditempat itu saja dan hampir tidak berpindah tempat.
Gambar 3. Daur hidup spp.
Tabel 1. Tahap perkembangan S. nubila (Warouw, 1981)
Telur 45.17 Nimfa Instar I 15.38 Instar II 19.56 Instar III 26.38 Instar IV 20.43 Instar V 27.19 108.33 Imago Betina Pra-peneluran 30.13 Peneluran 60.86 Pasca Peneluran 21.50 111.67 Imago Jantan 84.50 Daur hidup 183.63
Periode perkembangan dari Telur sampai imago mati
Imago Betina 265.17
Imago Jantan 238.00
Tjoa (1953) menyatakan bahwa hama ini melakukan aktivitas pada malam hari termasuk aktivitas makan dan berkopulasi. Dalam satu malam, nimfa dan imago dapat memakan daun kelapa sekitar 20 cm. Pada waktu makan, imago jantan mengeluarkan bunyi yang nyaring, demikian juga kalau dipegang imago jantan mengeluarkan bunyi yang sama. Dari hasil pengamatan penulis di laboratorium lapangan (insektarium),
Bioekologi Hama spp. (Orthoptera: Tettigoniidae)
ternyata hama S. nubila dapat berkopulasi pada siang hari antara jam 9.00 – 11.00 pagi. Biasanya imago jantan mendekati imago betina dengan posisi sejajar kemudian mendekatkan abdomennya pada abdomen betina, selanjutnya proses kopulasi berlangsung dengan posisi abdomen jantan berada pada bagian bawah. Jika kopulasi berhasil maka imago jantan mengeluarkan bursa kopulatriks dan menempel pada bagian pangkal ovipositor atau bagian ujung abdomen imago betina (Gambar 4).
Gambar 4. Proses kopulasi hama Abdomen jantan dibengkokkan kearah imago betina (atas), posisi abdomen jantan berada di bawah imago betina (tengah) dan bursa kopulatriks menempel pada bagian ujung abdomen betina (bawah).
Bioekologi Hama spp.(Orthoptera: Tettigoniidae)
6
2
.
Sexava coriacea LinnaeusHama ini dikenal dengan nama belalang Ambon. Bentuk dan warna telur S. coriacea sama dengan S. nubila. Nimfa berwarna hijau, kadang-kadang merah sauh seperti S. nubila. Imagonya sulit dibedakan dengan S. nubila. Panjang imago betina antara 8.5-9.5 cm dan jantan 7-8 cm. Panjang ovipositornya 4-5 cm. Pada dasarnya panjang ovipositor S. coriacea lebih penjang sedikit dari ujung sayapnya.
Cara hidup
Lamanya stadium telur S. coriacea 50 hari dan stadium nimfa 110 hari. Imago betina bertelur pada malam hari sebanyak 1-38 butir per malam. Masa jadi dari telur sampai imago >5 bulan. Imago betina mulai bertelur ± 40 hari setelah mencapai imago. Cara hidup S. coriacea sama dengan S. nubila.
3. Sexava karnyi Leefmans
Hama ini dikenal dengan belalang Togean. Nama setempat: Kuwao (Kepulauan Togean di teluk Tomini) atau Tupako (Ampana). Bentuk dan warna telur S. karnyi seperti buah padi masak/matang (gabah). Panjang telur 9-9.5 cm. Nimfanya hampir sama dengan S. nubila, berwarna hijau atau merah sauh.
Bentuk imago S. karnyi sama dengan imago S. nubila dan S. coriacea, tetapi ukurannya lebih kecil. Tubuh dan antenanya berwarna hijau. Panjang tubuh dari kepala sampai ke ujung sayap 6.5-7 cm. Paha kaki muka dan kaki tengahnya terdapat warna hitam memanjang. Warna hitam itu lebih jelas pada bagian dalam paha kaki belakang.
Cara hidup
Cara hidup S. karnyi hampir sama dengan S. nubila. Telur tidak hanya diletakkan dalam pasir tetapi di sela-sela batang, dan di mahkota pohon kelapa. Diletakkan juga di ketiak daun pandan. Nimfa dan imago hidup dari daun kelapa. Makannya sangat rakus. Hama ini merusak daun secara bersama-sama sehingga tingal lidi saja. Seperti
S. nubila dan S. coriacea, hama ini juga mengeluarkan bunyi pada malam hari.
Belalang Sexava spp. hidup dibagian Timur garis Wallace di Sulawesi, Maluku dan Papua. Hama S. nubila tersebar di Seram, Kei, Aru, Batjan, dan Kepulauan Talaud, Kepulauan Nanusa, Papua, dan Papua Nugini; S. coriacea terdapat di Maluku Utara, Kepulauan Sangihe, di Dumagin Bolaang Mongondow Sulawesi Utara, Sula, Banggai dan Irian bagian Utara; S. karnyi di Kepulauan Una-Una dan Togean (teluk Tomini) dan pesisir pantai Sulawesi Tengah di daerah Poh (1953) dan di daerah antara Ampana dan Poso (1948); dan S. novae-guineae di Papua Nugini (Hosang, 1989; Hosang dan Soekarjoto, 1989; Kalshoven, 1981; Lever, 1969; Tjoa, 1957; Warouw, 1981). Tiga spesies
Bioekologi Hama spp. (Orthoptera: Tettigoniidae)
Sexava nubila Sexava coriacea Sexava karnyi
yang disebutkan pertama merusak tanaman kelapa di Indonesia. Di lapangan hanya dua spesies yang lebih sering menimbulkan masalah yaitu S. nubila dan S. coriacea. Daerah penyebaran dari ketiga spesies yang ada di Indonesia seperti pada Gambar 5.
Gambar 5. Daerah Penyebaran hama spp. di Indonesia
Hama Sexava spp. merupakan serangga polifag karena selain merusak tanaman kelapa dapat juga merusak tanaman pisang, pandan, enau, rumbia, sagu, pinang, salak, jambu air, manggis, Zingiberaceae (misalnya Elettaria) dan lain-lain. Contoh kerusakan tanaman akibat serangan hama Sexava spp. pada tanaman kelapa dan tanaman inang lainnya seperti pada Gambar 6.
Gambar 6. Kerusakan tanaman akibat serangan hama spp. pada kelapa (kiri), pisang (tengah) dan pandan (kanan).
Bioekologi Hama spp.(Orthoptera: Tettigoniidae)
8
Sejarah serangan hama Sexava spp. telah dirangkum dari beberapa peneliti dan didokumentasi dengan baik oleh Warouw (1981). Sampai sekarang Sexava spp. merupakan hama utama perusak tanaman kelapa yang sifatnya kronis di Indonesia Timur dan Papua Nugini. Gangguan pertama dari S. nubila di Talaud dilaporkan terjadi pada tahun 1890. Di Salibabu, Kabupaten Talaud, produksi kopra yang terbaik berakhir pada tahun 1900. Pada saat itu, selama lima dekade yang pertama serangan belalang tersebut berturut-turut terjadi pada tahun 1910, 1911, 1916, 1920, 1923, 1930, 1932, 1937, 1938 dan 1949. Dalam periode 1950 sampai 1964 tidak ada laporan mengenai gangguan belalang itu tetapi dari beberapa wawancara Warouw (1981) dengan penduduk setempat, ternyata pada kurun waktu tersebut masih sering terjadi eksplosi belalang yang usaha pengendaliannya dibiayai oleh Yayasan Kopra.
Ledakan populasi S. nubila di Talaud terjadi lagi pada tahun 1965 dan tahun 1969, 1970. Gangguan sangat mempengaruhi keadaan sosial ekonomi petani sehingga tahun-tahun berikutnya pemerintah melakukan pengendalian secara kimia pada waktu populasi belalang diduga akan naik. Pada tahun 1976 di lokasi tersebut masih terdapat kompleks-kompleks pertanaman kelapa yang padat populasi belalang agak tinggi. Kompleks tersebut menjadi sumber-sumber eksplosi hama sehingga pada bulan Agustus sampai November 1978 terpaksa dilakukan lagi penyemprotan udara pada areal seluas 21 000 hektar untuk aplikasi pertama dan 22 800 hektar untuk aplikasi kedua.
Di tempat lain seperti di Kolepom, Kai dan Aru serta Merauke (Papua) belum pernah terjadi eksplosi S. nubila. Kemungkinan musuh alaminya masih mampu mengendalikan populasi belalang itu atau pengaruh faktor lingkngan.
Di lokasi sebaran gabungan S. nubila dan S. coriacea, gangguan diberitakan terjadi di Bacan pada tahun 1916, 1929, 1959, 1960, 1972; di Waigeo, Tsiof dan Biak pada tahun 1933. Eksplosi hama yang sering terjadi di Bacan kemungkinan disebabkan oleh S.
nubila. Di Seram dan Jayapura yang merupakan daerah sebaran gabungan kedua
spesies tersebut belum pernah terjadi eksplosi hama.
Eksplosi dari S. coriacea terjadi di Ambon pada tahun 1913, di Obi pada tahun 1915 dan 1972, di Halmahera dan Makian pada tahun 1931, 1932, 1963, 1964 dan 1972, di Ternate pada tahun 1960, 1963, di Sangihe, Siau dan Tahulandang pada tahun 1931, 1932 dan 1969, 1970.
Spesies S. karnyi diberitakan merusak pertanaman kelapa di Togean pada tahun 1910, 1933, 1952, 1962, 1972 dan 1975, sedangkan di Ampana (Sulawesi Tengah) terjadi pada tahun 1948.
Gangguan dari S. novae-guineae dan S. nubila di Papua Nugini pernah dilaporkan terjadi pada tahun 1933 dan seterusnya tidak diberitakan lagi meskipun penelitian pengendalian masih berlangsung terus.
Dari uraian sejarah gangguan tersebut terlihat bahwa frekuensi eksplosi paling tinggi terjadi di wilayah sebaran S. nubila di Talaud dan di wilayah sebaran S. karnyi di Togean. Eksplosi hama yang terjadi bukan hanya terbatas pada suatu kompleks pertanaman tetapi meliputi puluhan ribu hektar di seluruh Kepulauan. Selang waktu
Bioekologi Hama spp. (Orthoptera: Tettigoniidae)
terjadinya setiap eksplosi belalang ternyata minimal empat tahun dan maksimal sepuluh tahun bagi S. nubila serta S. karnyi di wilayah sebarannya (Warouw, 1981).
Dari studi ekologi S. nubila di Kepulauan Talaud pada tahun 1997 – 1991 dan tahun 2004 – 2005, ternyata papulasi hama dan tingkat kerusakan tanaman masih tinggi pada lokasi-lokasi tertentu (Zelazny dan Hosang, 1987; 1988, 1991; Hosang et al.; 2004; Balitka, 2005) sehingga perlu diantisipasi supaya tidak terjadi ledakan populasi di lapangan.
Pemahaman biologi dan ekologi hama Sexava spp. sangat diperlukan karena dapat membantu pengambilan keputusan dan penyusunan taktik pengendalian yang tepat untuk menekan populasi hama di lapangan.
Balitka. 2005. Penyempurnaan Pengendalian Hama Sexava Secara Terpadu. Laporan Penelitian. Balai Penelitian Tanaman Kelapa dan Palma Lain. Manado.
Hosang, M.L.A. 1989. Accidental introduction of Sexava coriacea into Dumagin, Pinolosian, Bolaang Mongondow, North Sulawesi. In: UNDP/FAO Integrated Coconut Pest Control Project, Annual Report. Balai Penelitian Kelapa, Manado, North Sulawesi. 126-127.
Hosang, M.L.A. dan Soekarjoto. 1989. Serangan hama Sexava coriacea di desa Dumagin Kabupaten Bolaang Mongondow Sulawesi Utara. Buletin Balitka (9):29-35. Hosang, M.L.A., J. Mawikere, A.A. Lolong, F. Tumewan, S. Sabbatoellah dan J.C.
Alouw. 2004. Interaksi tritrofik antara musuh alami, Hama (Sexava, Oryctes) dan tanaman kelapa. Laporan Penelitian. Balai Penelitian Tanaman Kelapa dan Palma Lain. Manado
Kalshoven, L.G.E. 1981. The Pests of Crops in Indonesia. Revised and translated by P.A. van der Laan with assis tance of G.H.L Rothschild. PT Ichtiar Baru van Hoeve, Jakarta. 701pp.
Lever, R.J.A.W. 1969. Pests of the Coconut Palm. No.18. FAO. Rome, Italy. 190pp. Singh, S.P. and P. Rethinam. 2005. Long-horned grasshoppers and their management in
coconut and oil palm ecosystems. Cococinfo International. 12(2):10-14.
Tjoa Tjien Mo. 1953. Memberantas hama-hama kelapa dan kopra. Noorhoff-holff. Jakarta. 270 pp.
Warouw. J. 1981. Dinamika populasi Sexava nubila Stal (Orthoptera, Tettigoniidae) di Sangihe Talaud dalam hubungan dengan kerusakan tanaman kelapa. Disertasi Doktor IPB.
Watson, T.F., L. Moore, dan G.W. Ware. 1975. Practical Insect Pest Management. A Self-Instruction Manual. W.H. Freeman and Co. San Francisco. 196h.
Bioekologi Hama spp.(Orthoptera: Tettigoniidae)
10
Zelany, B, & M.L.A. Hosang. 1987. Population studies of Sexava spp. In: UNDP/FAO Integrated Coconut Pest Control Project, Annual Report. Balai Penelitian Kelapa, Manado, North Sulawesi. 45-59.
Zelazny, B. and M.L.A. Hosang. 1988. Ecological studies on Sexava spp. and discussion on control with pesticides. In: UNDP/FAO Integrated Coconut Pest Control Project, Annual Report. Balai Penelitian Kelapa, Manado, North Sulawesi. 69-78. Zelazny, B. and M.L.A. Hosang. 1991. Estimating defoliation of coconut pamls by
BALAI PENELITIAN TANAMAN KELAPA DAN PALMA LAIN, MANADO
Sexava spp. (Orthoptera : Tettigoniidae) dapat menyebabkan kerusakan yang
serius pada tanaman kelapa terutama daerah-daerah tertentu di Kawasan Timur Indonesia. Di Indonesia sudah diketahui 3 jenis hama Sexava yang merusak tanaman kelapa, yaitu (a) Sexava nubila Stal di Kepulauan Talaud Sulawesi Utara, di Maluku dan Papua (Irian Jaya), (b) Sexava coriacea L. di Kepulauan Sangihe, di daratan Sulawesi di Desa Dumagin Kecamatan Pinolosian, Bolaang Mongondow, Sulawesi Utara (Hosang, 1989) dan di Maluku Utara, dan (c) Sexava karnyi Leefmans yang merusak tanaman kelapa pada beberapa daerah di Sulawesi Tengah.
Pada kenyataannya di lapangan hanya dua spesies (S. nubila dan S. coriacea) yang lebih banyak menyebabkan kerusakan pada tanaman kelapa. Studi ekologi S. nubila dan S. coriacea telah dilakukan di Kepulauan Sangihe dan Talaud (Zelazny dan Hosang, 1987;1988). Monitoring populasi hama dilaksanakan pada populasi S.
nubila di pulau Salibabu dan populasi S. coriacea di Tahuna (Pulau Sangihe Besar).
Studi kerusakan merupakan salah satu bagian penting dalam pengembangan metode pengendalian hama terpadu dan untuk memprediksi perbandingan biaya dan keuntungan dalam praktek pengendalian. Kerusakan dapat dikelompokkan ke dalam kerusakan langsung dan tidak langsung. Kerusakan langsung adalah kerusakan yang terjadi pada bagian tanaman yang dipanen, sedangkan kerusakan tidak langsung adalah kerusakan pada bagian tanaman yang tidak dipanen (Rauf, 1996). Hama
Sexava spp. menyebabkan dua tipe kerusakan pada tanaman kelapa yaitu (a) langsung
merusak bunga dan buah muda, apabila serangan ringan buah dapat berkembang sampai siap dipanen tetapi serangan berat buah akan gugur, dan (b) merusak daun sehingga kehilangan luas daun secara tidak langsung mempengaruhi produksi pada tanaman dewasa, sedangkan pada tanaman muda pertumbuhannya akan terhambat. Serangan berat dari hama ini dapat menyebabkan tanaman mati (Zelazny dan Hosang, 1988). Tanaman kelapa yang belum berproduksi apabila terserang hama ini secara berkesinambungan maka pertumbuhannya akan terhambat, lambat berproduksi atau tidak berproduksi dan lama kelamaan tanaman akan mati (Gambar 1). Dalam makalah ini dikemukakan tentang tingkat kerusakan dan populasi hama Sexava spp. dalam hubungannya dengan produksi kelapa.
Dampak Kerusakan Tanaman Akibat Serangan spp. Terhadap Penurunan Produksi Kelapa
12
Gambar 1. Serangan hama Sexava spp., tanaman petumbuhan terhambat, lambat berproduksi dan tanaman mati
.
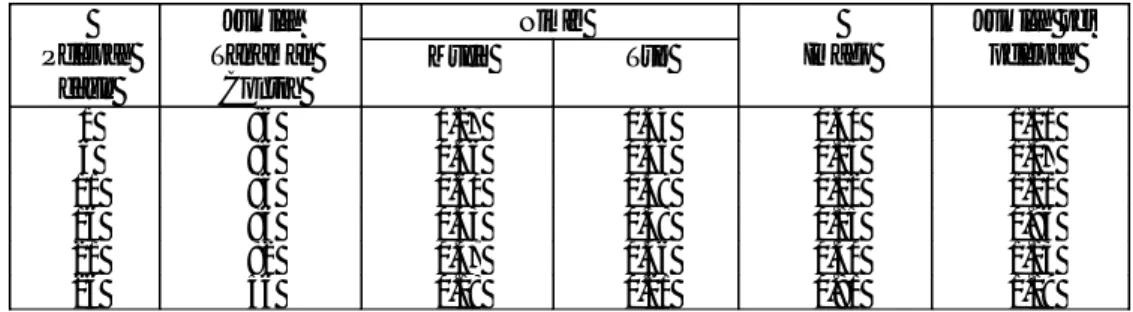
Kerusakan tanaman akibat serangan hama Sexava spp. telah dipelajari secara rinci pada populasi hama S. nubila dan S. coriacea. Data pada Tabel 1 dan 2, memberikan gambaran kerusakan tanaman kelapa dan populasi hama S. nubila dan S.
coriacea di daerah serangan di Kepulauan Talaud dan Sangihe Besar. Serangan hama
S. nubila di Pulau Salibabu Talaud, terlihat ada peningkatan kerusakan tanaman pada
lokasi yang diamati, kecuali satu lokasi yang diaplikasi dengan insektisida sistemik. Pada lokasi tanpa perlakuan insektisida, rata-rata kerusakan tanaman meningkat dari 21.3% menjadi 26.5%. Sebaliknya pada populasi S. coriacea di Sangihe Besar, kerusakan tanaman menurun pada tiga lokasi dari enam lokasi yang diamati (Tabel 1). Perkembangan populasi Sexava spp. (Tabel 2), memperlihatkan adanya peningkatan nimfa pada tahun 1988. Dari data populasi telur, kelihatannya sulit mengambil kesimpulan karena telur yang mengandung kuning telur (yolk) kemungkinan juga tidak fertil atau sudah mati. Hasil ini menunjukkan bahwa tingkat kerusakan hama S. nubila telah melampaui batas ambang kerusakan ekonomi (> 20%) (Balitka, 1990) sehingga perlu dilakukan pengendalian secara terencana supaya populasi hama dapat ditekan, sebaliknya tingkat serangan di Pulau Sangihe Besar masih dapat ditoleransi. Untuk itu perlu dilakukan monitoring secara teratur supaya tidak terjadi ledakan populasi hama di lapangan.
Dampak Kerusakan Tanaman Akibat Serangan spp. Terhadap Penurunan Produksi Kelapa
Tabel 1. Rataan persentase kerusakan daun oleh Sexava spp. (Zelazny dan Hosang, 1988).
Kelompok Tanaman (10 pohon) Salibabu (Talaud) Sangihe Besar 1987 1988 1987 1988 1 28.8 24.1 9.1 8.4 2 27.1 17.3 12.8 6.6 3 21.3 30.4 5.1 2.9 4 15.3 22.9 19.5 11.6 5 14.0 36.0 6.0 5.3 6 31.5 17.4* 0.7 0.8 Total 23.0 25.2 9.2 8.0
Tabel 2. Populasi Sexava spp. pada setiap stadia (Zelezny dan Hosang, 1988).
Stadia Salibabu (Talaud) Sangihe Besar 1987 1988 1987 1988
Yolk (kuning telur) 1.1 1.6 4.0 0.9
Embrio 0.4 1.1 0.3 0.2
% telur terparasit 26% 8% 0% 0%
Nimfa muda <3.5 cm 0.60 1.58 0.22 0.60
Nimfa tua 0.40 0.76 0.08 0.40
Imago 0.13 0.12 0.13 0.00
Pada Tabel 3, terlihat persentase kehilangan luas daun yang dimakan
Sexava spp. pada pelepah dengan tingkat umur berbeda. Kerusakan tanaman
meningkat pada pelepah daun kelapa yang lebih tua, hal ini disebabkan karena pelepah daun tua sudah lebih lama diserang hama dibandingkan dengan pelepah daun muda. Tanaman kelapa menghasilkan pelepah daun dengan laju konstan, apabila Sexava makan secara merata pada seluruh pelepah daun maka akan terjadi peningkatan kerusakan pada mahkota daun dari bagian atas ke bagian bawah. Bagian yang dimakan penting untuk analisis kehilangan hasil, sebab kerusakan pelepah daun muda akan lebih berpengaruh terhadap produksi dibandingkan dengan pelepah tua. Ada indikasi juga bahwa daun yang dipilih sebagai makanan akan berubah tergantung perbedaan tingkat kerusakan. Pada kerusakan berat, peningkatan kerusakan lebih terkonsentrasi pada daun muda, sebab serangga tidak mendapatkan makanan yang cukup pada daun yang tua (Zelazny dan Hosang, 1991) (Gambar 2). Pada beberapa lokasi dengan tingkat kerusakan berat menunjang teori yang dikembangkan oleh Warouw (1981), yang menyatakan bahwa populasi Sexava spp. pada waktu itu menurun karena tidak cukup tersedia daun kelapa yang tertinggal sebagai makanan.
Dampak Kerusakan Tanaman Akibat Serangan spp. Terhadap Penurunan Produksi Kelapa
Dampak Kerusakan Tanaman Akibat Serangan spp. Terhadap Penurunan Produksi Kelapa
Tabel 3. Rataan persentase kerusakan tanaman kelapa oleh serangan Sexava di Sulawesi Utara dan Maluku (Zelazny dan Hosang, 1991).
Pelepah Daun Tingkat Kerusakan Ringan (n=36) Sedang (n=27) Berat (n=23) Total (n=86) Perbedaan 1 0.4 0.8 7.8 3.1 10.7 15.6 12.7 7.3 -0.3 6 4.1 9.6 26.1 13.9 11 9.9 21.6 53.1 29.5 16 15.2 34.0 71.9 42.2 21 22.4 45.2 75.4 49.5 26 28.4 46.4 73.6 49.2
Walaupun jumlah Sexava spp. hampir sama pada semua pelepah daun, terlihat ada perbedaan antara jumlah nimfa muda dan nimfa tua (Tabel 4). Nimfa muda lebih sering ditemukan pada pelepah daun tengah, sedangkan imago lebih sering terdapat pada daun muda dan daun tua. Distribusi nimfa muda ternyata dapat meningkatkan kerusakan daun. Dua faktor yang dapat menyebabkan perbedaan distribusi imago pada mahkota daun adalah :
a. Waktu dilakukan pengamatan pada siang hari, imago dapat berpindah ke pelepah daun muda dan tua.
b. Buah muda diserang oleh imago dan kemungkinan juga oleh nimfa tua. Terdapatnya imago pada pelepah tua memberikan gambaran bahwa mereka lebih dekat ke buah.
Selain daun hama Sexava spp. juga dapat merusak buah Gambar 3). Rataan jumlah buah pada tandan kedua yang langsung dirusak oleh hama S. nubila di Talaud 19%, sedangkan oleh S. coriacea di Sangihe Besar 10% (Hosang et al., 1988; Zelazny dan Hosang, 1988). Pada tingkat serangan berat (Gambar 4) tanaman kelapa tidak dapat berproduksi selama 2 tahun. Untuk itu, perlu dikembangkan konsepsi PHT sehingga populasi hama itu dapat stabil pada aras yang tidak merugikan.
Tabel 4. Rata-rata jumlah nimfa dan imago Sexava spp. yang ditemukan pada pelepah daun yang berbeda umurnya di Talaud, Tahuna dan Seram (Zelazny dan Hosang, 1988).
Pelepah daun Jumlah Tanaman Contoh Nimfa Imago Jumlah per pelepah Muda Tua 1 86 0.27 0.34 0.50 1.11 6 86 0.56 0.36 0.15 1.07 11 86 0.60 0.49 0.12 1.21 16 86 0.43 0.38 0.15 0.96 21 82 0.37 0.46 0.30 1.13 26 56 0.18 0.21 0.81 1.19
Dampak Kerusakan Tanaman Akibat Serangan spp. Terhadap Penurunan Produksi Kelapa
16
Gambar 3. Kerusakan bunga betina dan buah muda
Gambar 4. Serangan berat hama Sexava spp. tanaman tidak berproduksi
Hubungan antara kerusakan dengan kehilangan hasil tergantung pada tingkat kerusakan itu sendiri, fase perkembangan tanaman (fenologi tanaman), dan kondisi lingkungan. Tanaman umumnya mempunyai kemampuan mentoleransi kerusakan, khususnya terhadap kerusakan yang terjadi pada daun. Dengan demikian, hubungan antara tingkat kerusakan dengan kehilangan hasil tidak selalu linier tetapi umumnya bersifat sigmoid. Sampai pada batas kerusakan tertentu, tanaman tidak memperlihatkan penurunan hasil. Batas kerusakan ini dikenal dengan sebagai ambang kehilangan hasil. Tingkat kerusakan yang lebih rendah dari nilai ambang ini tidak berpengaruh terhadap hasil. Petani sering menganggap kerusakan sedikit saja pada
Dampak Kerusakan Tanaman Akibat Serangan spp. Terhadap Penurunan Produksi Kelapa
daun dapat menurunkan hasil. Dalam psikologi sosial fenomena semacam ini dikenal sebagai ”efek hallo”, yaitu memberi kesan yang berlebihan terhadap suatu objek.
Pada kebanyakan tanaman, hubungan antara tingkat kerusakan dengan kehilangan hasil umumnya bervariasi tergantung pada saat kerusakan terjadi dalam kaitannya dengan fase pertumbuhan tanaman. Kerusakan langsung juga sering dapat dikompensasi. Biji yang dirusak hama, sumberdaya tanaman dialihkan ke biji yang sehat sehingga biji tumbuh lebih besar (Rauf, 1996). Hal ini juga dapat terjadi pada tanaman kelapa.
Toleransi tanaman terhadap kerusakan dipengaruhi juga oleh kondisi fisik dan kimiawi. Sudah banyak diketahui bahwa tanaman yang tumbuh merana lebih rentan terhadap infestasi hama. Dalam hal ini, tanaman kekurangan unsur hara atau air kurang memiliki kemampuan untuk mentoleransi atau mengkompensasi kerusakan yang terjadi. Prinsip pengendalian hama terpadu yang menyebutkan ”budidaya tanaman sehat” sebetulnya adalah dalam rangka memperoleh hasil panen yang tinggi serta mengurangi pengaruh buruk dari kerusakan hama. Tanaman yang tumbuh subur memiliki kemampuan kompensasi yang lebih baik (Rauf, 1996).
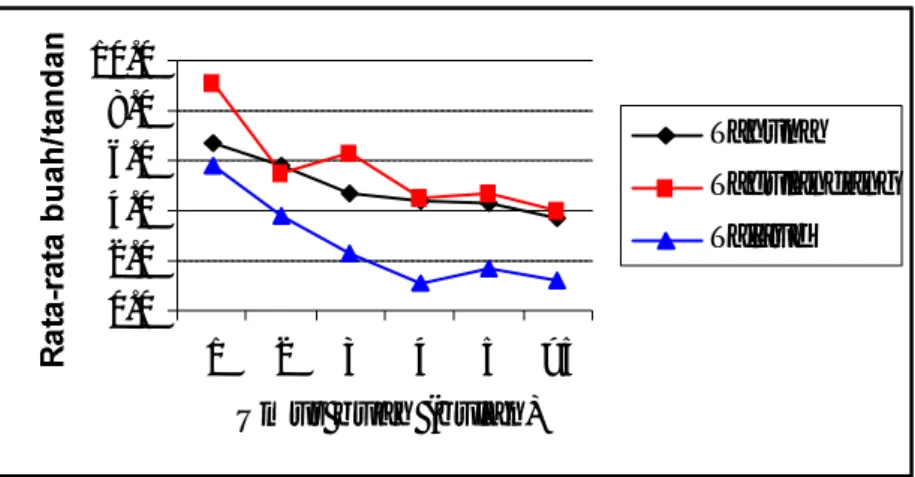
Zelazny dan Hosang (1988) telah mempelajari korelasi antara tingkat kerusakan dan produksi kelapa. Dari hasil penelitian menunjukkan bahwa makin tinggi tingkat kerusakan yang disebabkan oleh hama Sexava spp., makin rendah produksi kelapa (Tabel 5). Hal ini membuktikan bahwa hama Sexava spp. masih merupakan ancaman bagi tanaman kelapa di Kawasan Timur Indonesia, karena dapat menyebabkan kehilangan produksi yang cukup besar. Pada tingkat kersakan 62.5% perkiraan produksi/pohon/ tahun hanya 5.68 butir, hal ini tentunya sangat merugikan petani atau pengusaha kelapa lainnya. Hal ini sejalan dengan penelitian yang dilakukan oleh penulis seperti pada Gambar 5 dan 6.
Tabel 5. Hubungan tingkat kerusakan dan produksi.
Persentase Kerusakan Rataan jumlah buah per tandan (tandan 4-14)
Perkiraan produksi/ pohon/tahun*
11.2 4.26 56.23 24.6 2.55 33.66 38.5 1.41 18.61 52.2 1.31 14.92 62.5 0.43 5.68
Pada Gambar 5, terlihat bahwa produksi kelapa di daerah serangan hama
S. nubila di Kabupaten Talaud lebih rendah dibandingkan dengan produksi kelapa di
daerah serangan hama S. coriacea di Kabupaten Sangihe (Tahuna dan Tagulandang). Hal ini tentunya erat kaitannya dengan tingkat kerusakan yang disebabkan oleh hama
Sexava spp. Rata-rata kerusakan tanaman akibat serangan S. nubila di Kabupaten
Talaud (44.1%) lebih tinggi dibandingkan dengan S. coriacea di Kabupaten Sangihe (3.4% di Tahuna dan 14.41% di Tagulandang).
Dampak Kerusakan Tanaman Akibat Serangan spp. Terhadap Penurunan Produksi Kelapa
18
Gambar 5. Produksi kelapa pada tiga ekosistem pertanaman kelapa yang terserang hama Sexava spp.
Gambar 6. Pengaruh kerusakan daun terhadap produksi kelapa.
Hosang et al. (1992) menyatakan bahwa menurunnya tingkat kerusakan tanaman, ada kecenderungan dapat meningkatkan produksi kelapa. Untuk itu maka tindakan pengendalian perlu dilakukan sebelum terjadi kerusakan ekonomi, walaupun hal ini bukan merupakan satu-satunya kriteria dalam pengendalian, karena selain itu juga harus diperhatikan keadaan musuh alami di lapangan. Pengendalian hama ini sebaiknya diarahkan pada konsepsi pengendalian hama terpadu dengan memanfaatkan semua komponen pengendalian yang sudah tersedia dan sesuai diterapkan di lapangan.
0.0
2.0
4.0
6.0
8.0
10.0
1
2
3
4
5
>5
Umur buah (bulan)
Tahuna
Tagulandang
Talaud
Kerusakan daun (%)
0 20 40 60 80 0 5 10 15 20 Y= 4.86 – 0.07 xY = 4.86 – 0.07 x
Dampak Kerusakan Tanaman Akibat Serangan spp. Terhadap Penurunan Produksi Kelapa
Hama Sexava spp. dapat merusak daun, bunga dan buah muda sehingga dapat mempengaruhi pertumbuhan tanaman dan produksi buah. Makin tinggi tingkat kerusakan tanaman makin rendah produksi kelapa. Pada tingkat kerusakan 62.5% perkiraan produksi/pohon/tahun hanya 5.68 butir.
Tanaman kelapa yang terserang secara berkesinambungan, pertumbuhannya terhambat seperti batang mengecil, lama berproduksi dan lama kelamaan tanaman akan mati.
Balitka. 1990. Pedoman pengendalian hama dan penyakit kelapa. Badan Litbang, Balitka, FAO/UNDP, Dirjenbun, Direktorat Perlintan. 100 pp.
Hosang, M.L.A., 1989. Accidental Introduction of Sexava coreacea into Dumagin, Pinolosian, Bolaang Mongondow, North Sulawesi. In: UNDP/FAO Integrated Cocont Pest Control Project, Annual Reprot. Balai Peneltian Kelapa, Manado, North Sulawesi. 126-127.
Hosang, M.L.A., B. Zelazny and F. Tumewan, 1988. Attempts to Release Gregarines (Protozoa, Gregarine) into Sexava nubila populations of the Talaud Islands. In: UNDP/FAO Integrated Coconut Pest Control Project, Annual Report. Balai Penelitian Kelapa, Manado, North Sulawesi. 62-68.
Hosang, M.L.A. F. Tumewan, Soekarjoto, 1992. Perkembangan populasi hama Sexava
nubila di Pulau Seram, Maluku Tengah. Bull. Balitka, No. 17, 38-43.
Kalshoven, L.G. E. 1981. The pests of crops in Indonesia. Revised and translated by P.A. Van der Laan. PT. Ichtiar Baru van Hoeve. Jakarta. 701 pp.
Rauf, A. 1996. Analisis ekosistem dalam pengendalian hama terpadu. Pelatihan Hama dan Penyakit Tanaman Padi dan Palawija Tingkat Nasionalm, Jatisari 2-19 Januari 1996. 11h.
Warouw, J. 1981. Dinamika Populasi Sexava nubila (Stal) (Orthoptera, Tettigonidae) di Sangihe Talaud dalam Hubungannya dengan Kerusakan Tanaman Kelapa. Tesis Doktor. IPB. 152 hal.
Zelazny, B and A.R. Alfiler, 1987. Ecological Methods for Oryctes rhinoceros (Coleoptera: Scarabaeidae). Ecological Entomology, 12, 227-238.
Zelazny, B. and M.L.A. Hosang, 1987. Population studies on Sexava spp. In: UNDP/FAO Integrated Coconut Pest Control Project, Annual Report. Balai Penelitian Kelapa, Manado, North Sulawesi. 47-59.
Zelazny, B. and M.L.A. Hosang, 1988. Ecologocal Studies on Sexava spp. And Discussion on Control with Pesticides. In: UNDP/FAO Integrated Coconut Pest Control Project, Annual Report. Balai Penelitian Kelapa, Manado, North Sulawesi. 69-78.
Zelazny, B. and M.L.A. Hosang, 1991. Estimating Defoliation of Coconut Palms by Insect Pests. Tropical Pest Management. 37(1) 63-65).
MONOGRAF HAMA DAN PENYAKIT KELAPA
20
BALAI PENELITIAN TANAMAN KELAPA DAN PALMA LAIN, MANADO
Iklim merupakan penyebab penting perubahan populasi hama dalam ekosistem. Serangga sebagai hewan berdarah dingin (poikilotermal) secara fisiologi tidak dapat mengatur suhu tubuhnya sehingga kehidupannya akan sangat dipengaruhi oleh keadaan cuaca dan iklim tempat hidup atau habitatnya. Besarnya pengaruh ini berbeda untuk tiap spesies dan pengaruhnya dapat secara langsung terhadap fisiologi dan tingkah laku (antara lain lokomosi, orientasi dan penyebaran) dari serangga atau secara tidak langsung terhadap populasi serangga melalui pengaruhnya terhadap tanaman inang dan musuh alaminya (Price, 1997). Cuaca dan iklim bekerja sebagai faktor tidak bertautan padat. Dengan kata lain perubahan populasi ditentukan oleh keadaan cuaca dan iklim, tetapi tingkat keseimbangan populasi diatur oleh faktor yang bertautan padat.
Di daerah tropis seperti Indonesia, suhu, kelembaban relatif dan photoperiod tidak terlalu berfluktuasi, tetapi peranan faktor-faktor abiotik ini penting untuk spesies serangga tropis melalui pengaruhnya secara tidak langsung terhadap ketersediaan dan kualitas inang. Dengan variasi suhu yang rendah, maka populasi cenderung lebih rendah selama musim basah dibanding musim kering. Hal ini kemungkinan disebabkan oleh karena pada kelembaban tinggi menghambat faktor lain untuk mempengaruhi kecepatan reproduksi (Schowalter, 2000, Dent dan Walton, 1997, dan Bateman, 1972, Morallo-Rejesus, 2001).
Populasi S. nubila seperti populasi serangga lainnya akan dipengaruhi oleh komponen-komponen lingkungan hidup yang bersifat menyeluruh dan dinamis. Perubahan-perubahan dalam populasi akan terjadi karena pengaruh komponen-komponen tersebut pada seluruh proses perkembangan hidupnya (Warouw, 1981).
Suhu merupakan salah satu faktor pembatas penyebaran hewan. Pengaruhnya dapat terjadi terhadap stadia, daur hidup, pertumbuhan dan perkembangbiakan hewan. Adaptasi suatu spesies terhadap keadaan suhu tinggi dan rendah akan mempengaruhi sebaran geografis spesies tersebut (Krebs, 1978). Serangga hanya dapat berkembang di dalam kisaran suhu terbatas yang menjadi ciri khas dari setiap spesies dan mati pada suhu di luar kisaran tersebut. Secara fisiologi kematian dapat
Pengaruh Iklim Terhadap Populasi spp.
disebabkan karena peningkatan kelarutan lemak dalam membran, denaturasi protein (enzim) atau pembekuan sel (inter/intrasel). Namun serangga memiliki kemampuan adaptasi secara fisiologi terhadap pengaruh suhu tinggi ataupun rendah melalui beberapa proses antara lain penggunaan phosphate dalam respirasi untuk menghasilkan panas, atau kemampuan untuk bertahan dalam udara dingin menggunakan glycerol sebagai “anti-freeze agent” (Nation, 2002).
Pada suhu yang optimal untuk perkembangannya, serangga berkembang sangat cepat atau banyak yang berhasil menyelesaikan perkembangannya. Pada imago, lama hidup dan produksi telur juga tergantung pada suhu optimal. Biasanya imago lebih lama hidup pada suhu yang lebih rendah karena pengeluaran energi minimum (Chapman, 1970). Andrewartha dan Birch (1974) menyatakan bahwa batas kisaran suhu yang nyaman mungkin berhubungan erat dengan karakteristik kisaran suhu tempat suatu spesies biasa hidup. Telah lama diketahui bahwa hewan berdarah dingin termasuk serangga akan lebih cepat menyelesaikan perkembangan stadia hidupnya di daerah panas dari pada daerah dingin.
Kelembaban udara dapat mempengaruhi pembiakan, pertumbuhan,
perkembangan dan keaktifan serangga. Kemampuan serangga bertahan terhadap keadaan udara sekitar sangat berbeda menurut jenisnya. Kisaran toleransi terhadap kelembaban udara berbeda untuk setiap spesies maupun stadia perkembangannya, tetapi kisaran toleransi ini tidak jelas seperti pada suhu. Namun bagi serangga pada umumnya kisaran toleransi terhadap kelembaban udara yang optimum berada di dekat titik maksimum antara 73 – 100 % (Andrewartha dan Birch, 1974).
Faktor suhu dan kelembaban merupakan faktor yang saling bergantung satu sama lain dalam mempengaruhi proses biofisika serangga-serangga di daerah tropis. Kombinasi kedua faktor tersebut lebih berpengaruh terhadap kelimpahan populasi suatu spesies dari pada aksinya secara tunggal (Morallo-Rejesus dan Rejesus, 2001).
Serangga pada umumnya memiliki daya adaptasi untuk menghindari kehilangan air untuk tetap menjaga kelembaban tubuh melalui proses pertukaran gas secara terbatas (restricted gas exchange) dalam trakea, komparmentalisasi air (water
compartmentalization) dan terdapatnya lapisan lemak pada kutikula. Adaptasi tingkah
laku dengan cara bersembunyi di tempat-tempat terlindung atau memilih habitat yang cocok juga dilakukan serangga (Nation, 2002, Klowden, 2002).
Sampai saat ini penyebaran Sexava spp., di Indonesia baru diketahui di kawasan Indonesia Timur, hal ini kemungkinan disebabkan karena faktor lingkungan terutama suhu dan kelembaban sesuai untuk perkembangan populasi hama ini. Walaupun tanaman kelapa tumbuh di Sulawesi Selatan tetapi hama ini belum ditemukan di daerah tersebut. Jelaslah bahwa lingkungan merupakan sesuatu yang khas untuk ditempati spesies tertentu.
Pengaruh Iklim Terhadap Populasi spp.
22
Hujan mempunyai arti penting dalam kehidupan serangga, berpengaruh terutama pada pertumbuhan dan keaktifan serangga. Unsur-unsur yang penting dalam analisis hujan adalah curah hujan, jumlah hari hujan dan kelebatan hujan.
Pengaruh hujan pada kehidupan serangga dapat bersifat langsung secara mekanik atau secara tidak langsung terhadap keadaan kelembaban udara dan tanah. Pengaruh mekanik dimaksudkan sebagai akibat hentakan langsung butir hujan pada serangga atau pada tempat hidupnya. Air merupakan faktor utama bagi perkembangan embrio telur. Embrio telur S. nubila yang berkembang normal akan mengalami penambahan bobot rata-rata 2.40 kali bobot awalnya. Enambahan bobot tersebut disebabkan oleh masuknya air ke dalam telur pada saat terjadi proses perkembangan embrio. Faktor fisik air mempengaruhi penetasa telur (Warouw, 1981).
Sebaran hujan sepanjang tahun di suatu tempat memiliki pola tertentu. Sebaran tersebut akan menunjukkan panjang pendeknya periode hujan dengan curah hujan banyak (bulan basah) dan periode bulan dengan curah hujan sedikit (bulan kering). Ledakan populasi suatu hama mungkin sangat erat hubungannya dengan periodesitas sebaran hujan tersebut. Keadaan kelembaban udara dan tanah yang berbeda antara periode bulan basah dengan bulan kering dapat menghambat atau merangsang keaktifan serangga tersebut.
Angin berperan penting dalam proses perpindahan dan penyebaran serangga untuk mengkolonisasi habitat baru dan/atau menghindari kondisi yang tidak sesuai untuk perkembangannya. Sunjaya (1970) menyatakan bahwa pada keadaan tertentu angin tidak memberikan akibat langsung pada pertumbuhan maupun perkembangan serangga. Angin akan berpengaruh pada proses penguapan dan keadaan lembab udara yang secara tidak langsung memberi akibat pada keseimbangan suhu tubuh maupun kadar air tubuh serangga. Pada beberapa serangga, penyebaran vertikal maupun horizontal seringkali dibantu oleh angin. Hal ini tentunya dapat berlaku pada Sexava, walaupun sudah diketahui bahwa hama ini tidak aktif terbang. Tetapi angin langkisan dapat membantu perpindahan imago dari satu pohon ke pohon lainnya.
Menurut Odum (1971) populasi diartikan sebagai kelompok organisme dari spesies yang sama yang menduduki ruang atau tempat tertentu, memiliki berbagai ciri atau sifat yang merupakan sifat individu di dalam kelompok itu.
Populasi berubah ukurannya, dalam hal ini jumlah individu yang terlibat di dalamnya menurut kondisi lingkungan yang sesuai untuk menghasilkan lebih banyak atau lebih sedikit individu dibandingkan dengan individu yang mati dalam selang waktu tertentu. Perubahan-perubahan ukuran populasi dikenal dengan dinamika populasi (Van den Bosch dan Messenger, 1973). Tujuan mempelajari dinamika
Pengaruh Iklim Terhadap Populasi spp.
populasi ialah untuk menerangkan fluktuasi kepadatan populasi. Berdasarkan pengetahuan itu maka dimungkinkan untuk menduga kepadatan populasi di lapangan (Voute, 1971).
Populasi bergerak sehubungan dengan penyebaran geografis. Populasi cenderung menyebar dalam suatu ruang atau tempat sampai pada kondisi lingkungan yang membatasinya. Faktor penghalang geografis seperti tepi pantai, pegunungan, gurun, atau tidak tersedianya sumber-sumber yang diperlukan seperti makanan atau tanah sebagai habitat. Penelitian tentang dinamika populasi Sexava masih terbatas sehingga dalam tulisan ini pada umumnya didasarkan pada hasil penelitian Warouw (1981).
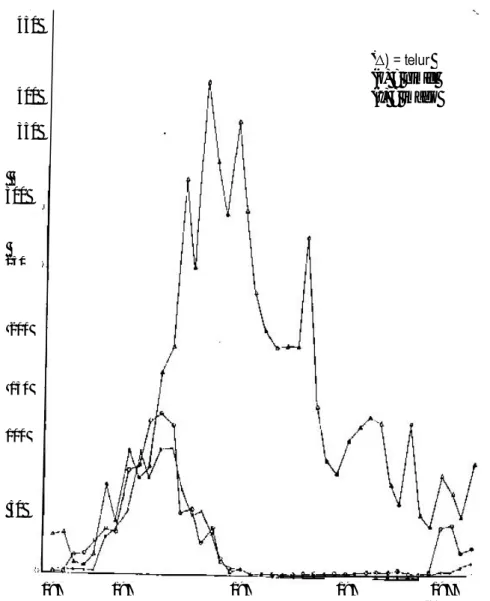
Populasi suatu spesies serangga umumnya mempunyai berbagai atribut yang khas untuk menunjang daya bertahan hidupnya Hasil pengamatan populasi telur, nimfa dan imago S. nubile (Gambar 1), menunjukkan bahwa pada setiap koloni yang diamati selalu ditemukan telur, nimfa dan imago dengan umur individu dalam setiap perkembangannya tidak sama. Populasi telur selalu ditemukan lebih tinggi jumlahnya kemudian diikuti oleh populasi nimfa dan imago. Populasi telur tertinggi yang pernah dicapai rata-rata 406 butir per pohon (Warouw, 1981). Kalshoven (1981) melaporkan bahwa dari 170.000 butir telur S. nubila ternyata hanya 24% yang memperlihatkan perkembangan embrio.
Naik turunnya populasi telur S. nubila (Gambar 1) menandakan adanya pengaruh dari beberapa factor lingkungan tertentu. Persentase mortalitas telur ternyata rendah (+ 11.74%). Hal tersebut menunjukkan bahwa populasi telur S. nubila di lapang mempunyai fertilitas dan kemampuan menetas yang tinggi.
Mortalitas telur Sexava disebabkan antara lain oleh entomopatogen yang berada dalam tanah. Infeksi entomopatogen cendawan diduga memiliki hubungan dengan kelembaban tanah (Warouw, 1981).
Populasi imago dapat mencapai lebih dari 100 ekor per pohon pada bulan Juni, September dan Oktober 1974. Peningkatan populasi imago diikuti pula oleh peningkatan populasi nimfa. Gejala ini menandakan adanya keeratan hubungan antara ketiga tingkat perkembangannya. Populasi nimfa yang maksimum mencapai 132.40 ekor per pohon.
Pengaruh Iklim Terhadap Populasi spp.
24
Gambar 1. Kurva populasi untuk telur, nimfa dan imago dari S. nubila pada tanaman kelapa di Lobo dan Makatara dari November 1973 sampai Maret 1977 (Warouw, 1981).
197
197
197
197
1977
(
(o) = nimfa
(x) = imago
100 150 350 250 50 300 400 200 450Pengaruh Iklim Terhadap Populasi spp.
Sexava spp. merupakan salah satu hama utama yang menyerang tanamankelapa di
wilayah Indonesia Timur.
Air berperan penting dalam perkembangan embrio telur dan mempengaruhi penetasan telur S. nubila.
Suhu dan kelembaban dapat mempengaruhi entomopatogen yang menyerang telur Sexava.
Lingkungan merupakan sesuatu yang khas untuk setiap spesies dan mempengaruhi perkembangan dan pertumbuhan serangga.
Andrewartha, H.G and L.C. Birch. 1974. The distribution and abundance of animal. Sixth Impression. The Univ. of Chicago Press. Chicago, USA. 782 p.
Bateman,M.D. 1972. The ecology of fruit flies. Ann. Rev. Entomol. 17:493-518. Chapman, R.F. 1970. The insects structure and function. American Elsevier Publ
Company, Inc, New York.
Dent, D.R. and M.P. Walton (eds). 1997. Methods in ecological and agricultural ecology. CAB International. 387 p.
Kalshoven, L.G.E. 1981. The pests of crops in Indonesia. PT. Ichtiar Baru-Van Hoeve. Jakarta : 701 p.
Klowden, M.J. 2002. Physiological system in insects. Acad. Press. London.413 pp. Krebs, C.J. 1978. Ecology: the experimental analysis of distribution and abundanc.
2nd ed. Harper and Row Publ. New York. 678 p.
Morallo-Rejesus B and R.S. Rejesus. 2001. Biology and management of stored product and posharvest insect pests. Dept of Entomology, College of Agriculture, UPLB. Philippines. 248 hal.
Nation, J.L. 2002. Insect Physiology Biochemistry. CRC Press. New York. 485 pp. Odum, E.P. 1971. Fundamentals of ecology. 3rd. W.B. Saunders company,
Philadelphia, London, Toronto.
Price, P.W. 1997. Insect ecology. 3rd Ed. Jhon Wiley & Sons, Inc. New York, USA. Schowalter, T.D. 2000. Insect ecology: an ecosystem approach. Academia Press. New
York. 483 p.
Sunjaya, P.I. 1970. Dasar-dasar ekologi serangga. Bagian Ilmu Hama Tanaman Pertanian. IPB. Bogor.
Van den Bosch, R. and P.S. Messenger. 1973. Biological control. Intext Educational Pub. New York.
Voute, A.D. 1971. Presidential address for the advanced study institute on Dynamic of numbers in populations. In. P.J. den Boer and G.R. Gradwel (eds). Dynamics of populations. Proc. Adv. Study Ins. Dynamics numbers Popul. 19-23.
Warouw, J. 1981. Dinamika populasi Sexava nubila (Stal.) (Orthoptera:Tettigonidae) di Sangihe Talaud dalam hubungannya dengan kerusakan tanaman kelapa. Tesis Doktor. IPB. 152 hal.
MONOGRAF HAMA DAN PENYAKIT KELAPA
26
BALAI PENELITIAN TANAMAN KELAPA DAN PALMA LAIN, MANADO
Pengetahuan interaksi antara serangga dan sumber makanannya merupakan dasar untuk mengembangkan pengertian mengenai perilaku, biologi, dan ekologi serangga yang dapat digunakan untuk menyusun strategi pengendalian hama. Interaksi antara serangga dan unsur-unsur pokok yang terkandung dalam makanan termasuk nutrisi dan alelokimia, dapat digunakan untuk membandingkan performans serangga pada tanaman inang yang berbeda. Informasi tersebut dapat digunakan juga untuk mempelajari struktur komunitas, resistensi tanaman inang, dan berguna dalam menyusun model matematika untuk menduga kehilangan produksi tanaman (Farrar et al., 1989).
Ketersediaan makanan menciptakan lingkungan yang kondusif terhadap kecepatan peningkatan suatu spesies serangga. Persediaan energi yang tinggi melalui makanan serta rendahnya kompetisi merupakan kondisi cukup ideal bagi perkembangbiakan suatu spesies serangga. Situasi yang demikian menyebabkan populasi hama berkembang mencapai kepadatan tinggi dalam waktu singkat. Makanan adalah salah satu faktor kunci kelimpahan serangga. Kecepatan perkembangan dipercepat atau diperlambat oleh kandungan nutrisi makanan dan dalam sejumlah kasus oleh jumlah yang tersedia dalam habitat mereka. Pertumbuhan dan reproduksi serangga tidak hanya tergantung pada jumlah nutrisi yang tersedia tetapi juga pada keseimbangan kandungan nutrisi didalamnya (Gordon, 1959 dan House, 1969 dalam Southwood, 1978 ). Distribusi dan kelimpahan sejumlah spesies serangga secara umum digambarkan oleh ketesediaan makanan dalam lingkungan.
Rangsangan makan dan peletakan telur dari serangga dapat dipengaruhi oleh metabolik primer dan sekunder. Semua tanaman mengandung karbohidrat dan asam amino sebagai metabolik primer yang dihasilkan dari aktivitas fotosintesa. Tidak semua serangga menggunakan karbohidrat sebagai rangsangan makan. Kebanyakan spesies serangga yang diteliti menerima rangsangan dari sukrosa, fruktosa dan glukosa. Metabolik tanaman sekunder seperti glukosinolat (minyak glukosida), diantaranya sinigrin pada tanaman Brasica dapat berfungsi sebagai perangsang makan dari hama Pieries (Lepidoptera), Phyllotreta armocaciae (Coleoptera) dan Brevicoryne brassicae, sedangkan glucobrassicin pada Brassica sebagai perangsang peletakan telur dari hama Pieris dan Delio radicum (Diptera) (Schoonhoven et al., 1998).
Studi khusus mengenai interaksi antara hama Sexava dengan tanaman inangnya belum banyak dipelajari. Walaupun demikian, pada dasarnya Sexava berinteraksi dengan tanaman inangnya untuk tujuan peletakan telur, mendapatkan makanan, dan
Interaksi antara Hama spp. dengan Tanaman Inang
sebagai tempat berlindung. Proses ini dapat dipengaruhi oleh kandungan zat perangsang dan keadaan fisik tanaman seperti morfologi permukaan (trichome), tekstur permukaan dan struktur lapisan lilin.
Hasil penelitian Zelazny dan Hosang (1988) pada populasi Sexava di beberapa lokasi yang mengalami kerusakan daun yang berat menunjukkan adanya penurunan populasi Sexava spp. Hal ini kemungkinan disebabkan oleh rendahnya ketersediaan daun kelapa sebagai makanan Sexava. Fenomena ini menunjang teori yang dikemukakan oleh Warouw (1981) bahwa rendahnya populasi Sexava disebabkan oleh tidak cukup tersedia daun kelapa sebagai makanan. Berdasarkan hal tersebut, sehingga dalam makalah ini dibahas tentang interaksi Sexava dengan tanaman inangnya.
Serangga berinteraksi dengan tanaman inang secara spesifik. Agar dapat hidup terus dan berkembangbiak, serangga harus menentukan inangnya. Beberapa serangga pemakan tanaman langsung meletakkan telur pada bahan makanan yang penting untuk perkembangan keturunannya (Brown et al., 1980 dan Richard, 1978).
Banyak tanaman dapat menjadi inang yang cocok untuk serangga tertentu tetapi tidak menarik bagi serangga lain. Hal ini antara lain disebabkan karena tanaman itu tidak mengeluarkan atau menghasilkan rangsangan yang memungkinkan serangga mengenal, berorientasi dan menemukan tanaman tersebut. Pemilihan serangga terhadap tanaman sebagai makanan, tempat bertelur ataupun tempat berlindung sangat ditentukan oleh sifat fisik dan zat-zat yang terkandung dalam tanaman itu sendiri. Sebagian besar kupu-kupu mempunyai inang yang terbatas pada genus dan spesies tanaman yang erat hubungannya. Meskipun kupu-kupu mengisap madu dari banyak tanaman, biasanya telur dan larvanya ditemukan pada beberapa tanaman yang familinya sama (Brown et al., 1980).
Pada tanaman kelapa, salah satu sifat fisik yang mempengaruhi preferensi serangga adalah kandungan trichome pada daun. Davis et al. (1985) menyatakan bahwa kelapa Genjah Kuning Nias (GKN) lebih resisten terhadap serangan hama
Setora nitens (Lepidoptera: Limacodidae) karena memiliki trichome dengan kepadatan
yang tinggi. Hasil penelitian menunjukkan bahwa serangan hama S. nitens pada GKN hanya 12.64% dari 4301 pohon, sedangkan pada kelapa Dalam dapat mencapai 30.50% dari 3347 pohon yang diamati.
Proses pemilihan inang pada serangga fitofag merupakan suatu rangkaian kejadian yang tiap kejadian terdahulu membuka jalan bagi kejadian berikutnya. Ada lima langkah utama dalam proses pemilihan inang yaitu: (a) penemuan habitat inang, (b) penemuan inang, (c) pengenalan inang, (d) penerimaan inang, dan (e) kesusaian inang (Kogan, 1982).
Penemuan habitat inang. Serangga dewasa sampai pada habitat inang melalui mekanisme yang menyangkut fototaksis, anemotaksis, geotaksis, dan pemilihan suhu dan kelembaban. Mekanisme ini mempunyai implikasi ekologi yang penting, dan dalam pengendalian hama perlu diperhatikan. Tahapan ini nampaknya kurang
Interaksi antara Hama spp. dengan Tanaman Inang
28
begitu penting pada Sexava karena sifatnya yang tidak aktif terbang dan biasanya meletakkan telur di tanah atau pada mahkota pohon kelapa, dan nimfa yang menetas langsung mencari pohon kelapa, memanjat dan menetap di sana.
Penemuan inang. Mekanisme jarak jauh, mungkin penglihatan atau penciuman (olfactory), membawa serangga itu dekat dengan tanaman inang. Banyak jenis kutu daun cenderung untuk hinggap pada permukaan berwarna kuning, dan larva dari kumbang tertentu tertarik pada pola vertikal. Apabila bau inang tercium, belalang dan kumbang kentang Leptinotarsa decemlineata cenderung terbang menentang arah angin (anemotaksis positif), sehingga mempertinggi peluang menemukan inangnya. Apabila terjadi kontak antara serangga dan tanaman inang maka masukan rangsangan indera peraba (tactile) dan penciuman menghentikan gerakan serangga itu, dan tetap tinggal pada tanaman tersebut. Reseptor kimia pada antena dan tarsus menerima rangsangan yang memberitahukan bahwa telah hinggap ditempat yang benar.
Pengenalan inang. Meskipun larva mempunyai alat-alat indera yang sampai tingkat tertentu dapat berfungsi dalam pengenalan inang, kerapkali hal ini diambil alih oleh serangga betina. Pada jenis belalang tertentu diketahui bahwa serangga betina menggigit tanaman dahulu sebelum meletakkan telur. Ulat yang menerima rangsangan melakukan uji gigit pada taanman bersangkutan. Gigitan tersebut menyebabkan bahan kimia lain yang terkandung dalam sel merangsang reseptor rasa (gustatory).
Penerimaan inang. Berbagai bahan kimia yang berbeda mengatur tahapan dalam proses makan. Suatu seri bahan kimia diekstrak dari daun murbei mempunyai asosiasi dengan gigitan awal, menelan, makan terus pada ulat sutera (Bombyx mori). Apabila masukan input indera itu tepat maka ulat akan makan terus sampai kenyang.
Kesesuaian inang. Nilai nutrisi tanaman dan tidak adanya bahan beracun menentukan baik tidaknya makanan untuk menunjang berbagai kasus fisiologi yang ada hubungannya dengan pertumbuhan dan perkembangan larva, serta lama hidup dan keperidian serangga dewasa.
Proses pemilihan inang oleh Sexava spp dapat terjadi karena tanaman mengeluarkan bau yang memungkinkan serangga mengenal tanaman inangnya. Oleh sebab itu perlu diteliti kandungan senyawa sekunder pada daun kelapa yang berperan dalam interaksi antara serangga dan tanaman. Seperti telah diidentifikasi beberapa senyawa sekunder pada kulit batang kelapa muda yang dapat memperlihatkan respons dari kumbang Rhyncophorus ferrugineus (Coleoptera: Curculionidae) (Gunawardena, 1994; 1995; Gunawardena et al, 1998).
Beberapa senyawa sekunder sudah dikenal berperan dalam proses pemilihan tanaman inang. Pengaruh glukosida dalam tanaman Brassiceae sebagai perangsang makan bersifat relatif tergantung pada perubahan konsentrasi zat tersebut. Ada empat jenis glukosida dalam tanaman Brassiceae, yaitu glucoiberin, glucoirucin, sinigrin dan progoitrin. Dari keempat glukosida itu yang paling efektif sebagai perangsang adalah sinigrin (Panda dan Kush, 1995).
Interaksi antara Hama spp. dengan Tanaman Inang
Tanaman sebagai makanan serangga herbivora merupakan jantung dari biologi serangga-tanaman. Konsentrasi unsur-unsur utama pada serangga dan tanaman terdapat dalam jumlah yang tidak sama kecuali unsur Mg. Unsur penting adalah Nitrogen karena suplainya singkat dalam bentuk yang dapat digunakan oleh serangga. Kandungan N hewan antara 8-14% dari berat kering, sedang pada tanaman hanya 2-4%. Nilai kalori jaringan serangga 22.8 j/mg sedangkan pada tanaman 18.9 j/mg. Ratio ini menunjukkan bahwa serangga herbivora mengkonsentrasikan N dari tanaman sebagai makanannya. Oleh karena itu nilai nutrisi tanaman bagi serangga terutama ditentukan oleh kandungan N, sedangkan nilai kalorinya kurang penting (Schoonhoven et al., 1998).
Kandungan unsur hara pada daun kelapa dibeberapa lokasi serangan hama Sexava di Sangihe Talaud, Sulawesi Utara adalah sebagai berikut: kandungan unsur N 1.06-1.51%, P 0.08 - 0.11%, K 0.46 - 1.10%, Ca 0.41 - 0.71% dan Mg 0.22 - 0.39%. (Kaat dan Mahmud, 1984) Jika dibandingkan data Schoonhoven et al, 1998 ternyata kandungan unsur hara daun kelapa sebagian besar lebih sedikit dibandingkan kebutuhan hama Sexava spp (jika diasumsi kebutuhannya sama dengan serangga lain). Dengan demikian untuk memenuhi kebutuhan nutrisi maka serangga tersebut dapat mengkonsumsi lebih banyak daun kelapa sebagai sumber makanannnya. Walaupun disadari bahwa kebutuhan nutrisi bagi setiap spesies serangga berbeda.
Warouw (1981) menyatakan bahwa jenis makanan dapat mempengaruhi mortalitas nimfa Sexava. Kelima instar yang makan daun tajuk bawah dari kelapa, kombinasi kelapa – pisang, dan pisang berbeda nyata dengan yang makan daun tajuk atas dari tanaman-tanaman tersebut, kecuali instar pertama. Pada instar pertama tidak terdapat perbedaan mortalitas, karena nimfa tersebut memerlukan makanan yang cukup lunak untuk dikunyah, tetapi makin tua umur nimfa, makin besar pengaruh bagian daun tajuk tanaman inang terhadap mortalitasnya. Nimfa yang makan daun tajuk bawah dan tajuk atas kelapa ternyata persentase mortalitasnya lebih rendah dibandingkan dengan nimfa yang makan daun pisang dari kedua bagian tajuk tanaman itu. Hal tersebut memberikan gambaran bahwa daun kelapa lebih baik pengaruhnya dibandingkan dengan daun pisang. Nimfa belalang yang dapat bertahan hidup pada daun kelapa berkisar antara 10 – 12.5%, sedang pada daun pisang hanya berkisar 0-5% (Tabel 1).
Dari hasil analisis ini ditemukan juga bahwa mortalitas kelima instar nimfa
S. nubila yang makan daun tajuk bawah maupun tajuk atas, kombinasi kelapa–pisang
berbeda nyata dengan yang makan daun kelapa dan daun pisang pada kedua bagian tajuk tanaman itu. Daya bertahan hidup nimfa yang makan daun kombinasi kelapa-pisang dapat mencapai 25%, sedangkan yang makan daun kelapa hanya 10%. Hal ini menunjukkan nutrisi gabungan dari daun kelapa dan pisang sangat cocok bagi perkembangan nimfa S. nubila. Dengan demikian terlihat bahwa perkembangan nimfa S. nubila sangat dipengaruhi oleh kualitas makanan yaitu jenis dan bagian tertentu dari tajuk tanaman inang.
Interaksi antara Hama spp. dengan Tanaman Inang
30
Pengujian pada imago menunjukkan bahwa jenis makanan mempunyai pengaruh yang berbeda terhadap mortalitas imago. Mortalitas imago yang makan daun tajuk bawah kelapa, kombinasi kelapa-pisang, dan daun pisang lebih rendah dibandingkan dengan yang makan daun bagian tajuk atas. Imago yang makan tajuk bawah dan atas kelapa mengalami mortalitas berturut-turut 45 dan 52.5%, sedangkan imago yang makan kombinasi daun kelapa pisang bagian tajuk bawah dan atas beturut-turut 42.5 dan 35% dalam periode peneluran. Didapatkan pula bahwa imago yang makan daun tajuk atas pisang mengalami mortalitas yang tertinggi yakni rata-rata 67. 5 dan 100% (Tabel 2).
Tabel 1. Pengaruh bagian tajuk dari jenis-jenis tanaman inang terhadap mortalitas dan daya bertahan hidup nimfa S. nubila (Warouw, 1981).
Bagian tajuk
tanaman Tanaman inang
Mortalitas kumulatif nimfa instar Nimfa bertahan hidup I II III IV V Sektor Kelapa 27.50 30.00 42.50 57.50 87.50 12.50 Bawah Kelapa + pisang 22.50 30.00 42.50 47.50 75.00 25.00 Pisang 47.50 62.50 70.00 80.00 95.00 5.00 Sektor Kelapa 32.50 62.50 72.50 80.00 90.00 0.00 Atas Kelapa+pisang 22.50 27.50 60.00 77.50 92.50 7.50 Pisang 60.00 60.00 100.00 100.00 100.00 0.00
Tabel 2. Pengaruh bagian tajuk dari jenis-jenis tanaman inang terhadap mortalitas dan daya bertahan hidup imago S. nubila (Warouw, 1981).
Dari penelitian itu terungkap bahwa imago Sexava spp yang dipelihara dengan pemberian daun pisang tidak dapat memproduksi telur. Jumlah telur yang banyak dihasilkan pada kombinasi makanan kelapa-pisang yaitu 110.50-137.75 butir.
Pengujian ini menunjukkan bahwa makanan kombinasi kelapa-pisang sangat cocok bagi daya bertahan hidup imago S. nubila. Makanan kombinasi kelapa-pisang seperti yang disebutkan sebelumnya mempunyai pengaruh yang baik terhadap kepiridian imago S. nubila dan daya bertahan hidup nimfanya. Dengan demikian dapat dinyatakan bahwa makanan kombinasi daun kelapa-pisang merupakan
Bagian tajuk
tanaman Tanaman inang Mortalitas kumulatif imago
Rata-rata Penelusuran Per Perlakuan Pra-penelusuran Penelusuran
...%... ...Telur...
Sektor Kelapa 45.00 67.50 119.25
Bawah Kelapa + Pisang 42.50 57.50 137.75
Pisang 67.50 95.00 17.75
Sektor Kelapa 52.50 80.00 74.50
Atas Kelapa + Pisang 35.00 52.50 110.50
Interaksi antara Hama spp. dengan Tanaman Inang
makanan yag paling cocok untuk perkembangan S. nubila. Berdasarkan hal tersebut ternyata bahwa daya bertahan hidup S. nubila dipengaruhi oleh kualitas makanan tanaman inang. Nutrisi yang cocok dan perbandingan relatif nutrisi yang terkandung pada daun tanaman inang merupakan faktor yang penting bagi perkembangan hama tersebut. Pemberian makanan pada serangga dengan mengkombinasikan dua jenis makanan telah diuji hama Opisina arenosella yang diberi makan daun kelapa dan
Borasus flabelifer menghasilkan reproduksi maksimum, sedang kombinasi inang lain
dengan pinang, reproduksina menurun. Selanjutnya Chamell et al. (1990) meneliti kemungkinan peranan nutrisi pada daun kelapa dihubungkan dengan tingkat serangan O. arenosella. Hasil penelitian menunjukkan jumlah Nitrogen yang tinggi pada pelepah daun menghasilkan tanaman yang rentan terhadap hama tersebut.
Dari hasil penelitian tersebut di atas jelas terlihat bahwa nutrisi sangat berpengaruh terhadap pertumbuhan dan perkembangan serangga. Dengan demikian kualitas makanan yang dikonsumsi serangga dapat menentukan kelangsungan hidupnya.
Southwood (1978) menjelaskan empat tantangan ekologi yang mempengaruhi keberhasilan makan dan kolonisasi oleh spesies serangga pada suatu tanaman:
a. Hambatan nutrisi
Kandungan protein seperti tryptophane dan methionine dan sterol yang rendah dalam tanaman seringkali merupakan suatu hambatan bagi serangga untuk berkembang dengan baik. Kandungan Sterol dalam jumlah sedikit sekali sangat kritis terutama bagi serangga betina dalam proses oviposisi. Disamping itu, factor fisik dan ekologi yang mempengaruhi tanaman juga mempengaruhi status biokimia tanaman.
b. Hambatan pelekatan (attachment)
Hambatan ini dihadapi terutama oleh serangga yang hidup pada bagian atas tanaman. Hambatan ini biasanya diatasi dengan membangun modifikasi struktural atau sekresi substansi untuk meningkatkan kemampuan melekat pada tanaman atau dengan jalan menggulung daun dan hidup didalamnya seperti pada serangga penggulung daun.
c. Hambatan kekeringan
Serangga dapat mengatasi hambatan ini dengan mengambil jumlah air lebih atau istirahat diatas permukaan tanaman atau bagian-bagian tanaman dengan kelembaban relatif tinggi. Oleh sebab itu serangga banyak mengkolonisasi growing