i
EVALUASI KEMURNIAN GENETIK SAPI BALI
MENGGUNAKAN DNA PENCIRI MIKROSATELIT
LOKUS HEL9 DI KABUPATEN BARRU
SKRIPSI MUSDALIFA MANSUR I 111 10 001 FAKULTAS PETERNAKAN UNIVERSITAS HASANUDDIN MAKASSAR 2014
ii
EVALUASI KEMURNIAN GENETIK SAPI BALI
MENGGUNAKAN DNA PENCIRI MIKROSATELIT
LOKUS HEL9 DI KABUPATEN BARRU
SKRIPSI
MUSDALIFA MANSUR I 11110001
Sebagai Salah Satu Syarat untuk Memperoleh Gelar Sarjana pada Fakultas Peternakan Universitas Hasanuddin
FAKULTAS PETERNAKAN UNIVERSITAS HASANUDDIN
MAKASSAR 2014
iii PERNYATAAN KEASLIAN
1. Yang bertanda tangan di bawah ini:
Nama : Musdalifa Mansur
NIM : I 111 10 001
Menyatakan dengan sebenarnya bahwa: a. Karya skripsi yang saya tulis adalah asli
b. Apa bila sebagian atau seluruhnya dari karya skripsi, terutama dalam Bab Hasil dan Pembahasan tidak asli atau plagiasi maka bersedia dibatalkan atau dikenakan sanksi akademik yang berlaku.
2. Demikian pernyataan keaslian ini dibuat untuk dapat dipergunakan sepenuhnya.
Makassar, Juni 2014 TTD
iv HALAMAN PENGESAHAN
Judul Penelitian : Evaluasi Kemurnian Genetik Sapi Bali Menggunakan DNA Penciri Mikrosatelit Lokus HEL9 di Kabupaten Barru
Nama : Musdalifa Mansur
No. Pokok : I 111 10 001 Program Studi : Produksi Ternak Jurusan : Produksi Ternak
Fakultas : Peternakan
Skripsi ini telah diperiksa dan disetujui oleh:
Pembimbing Utama
Prof. Dr. Ir. Lellah Rahim, M,Sc. NIP.19630501 198803 1 004
Dekan Fakultas Peternakan
Prof. Dr. Ir. Syamsuddin Hasan, M.Sc. NIP. 19520923 197903 1 002
Tanggal Lulus: 13 Juni 2014
Pembimbing Anggota
Dr. Muh. Ihsan A. Dagong, S.Pt. M,Si. NIP.19770526 200212 1 003
Ketua Jurusan Produksi Ternak
Prof. Dr. Ir. H. SudirmanBaco, M.Sc. NIP. 19641231 198903 1 025
v ABSTRAK
MUSDALIFA MANSUR (I 111 10 001). Evaluasi Kemurnian Genetik Sapi Bali Menggunakan DNA Penciri Mikrosatelit Lokus HEL9 Di Kabupaten Barru. Dibimbing oleh Lellah Rahim sebagai pembimbing utama dan Muhammad Ihsan Andi Dagong sebagai pembimbing anggota.
Penelitian ini bertujuan untuk mengetahui tingkat kemurnian sapi Bali di Kabupaten Barru berdasarkan identifikasi fenotipe dengan menggunakan DNA penciri mikrosatelit lokus HEL9. Sampel darah di ekstraksi dengan menggunakan Kit DNA ekstraksi Genjet Genomic DNA Extraction (Thermo Scientific), diamplifikasi dengan teknik PCR (Polymerase Chain Reaction). Analisis polimorfisme meliputi frekuensi alel dan genotipe, heterozigositas pengamatan (Ho), heterozigositas harapan (He) dan kesetimbangan Hardy Weinberg. Alel yang ditemukan adalah alel A dan alel B. Dari hasil penelitian ini menunjukkan bentuk tanduk dominan pada jantan adalah silak bajeg sedeangkan bentuk tanduk dominan pada betina adalah silak manggulgangsa. Genotipe yang ditemukan yaitu AB (100%). Pada populasi sapi Bali di kabupaten Barru diperoleh nilai heterozigositas pengamatan (Ho) adalah 1 dan nilai heterozigositas harapan (He) adalah 0.5031. Nilaichi- square pada penelitian ini tidak memenuhi syarat untuk di hitung dan tidak berada dalam keseimbangan Hardy- Weinberg.
vi ABSTRACT
MUSDALIFA MANSUR (I 111 10 001). Evaluation of Genetic Purity Bali Cattle In Barru Province using HEL9 Locus Mikrosatelite Marker. Supervised by Lellah Rahim as the main supervisor and Muhammad Ihsan Andi Dagong as the co supervisor.
The aim of this study to determined the level of genetic purity of Bali cattle in Barru Province based identification of phenotype and used HEL9 locus mikrosatelite marker. The genomic DNA was extracted by using Genomic DNA extraction Kit and then were amplified by Polymerase Chain Reaction machine. The analyzed of the polymorphism were calculated based on genotypic and allelic frequencies, observed heterozygosity (Ho), expected heterozygosity (He) and the Hardy-Weinberg equilibrium. The result of this study showed that the dominant horn shape of the bull was silak bajeg whereas cows was silak manggulgangsa. The genotypes frequencies of AB (100%). In Bali cattle population was found alelle A dan alelle B. In equal percentage (50 : 50). In Bali cattle population the observed heterozygosity value (Ho) was 1 while the expected heterozygosity in (He) was 05031. Chi- square value in this study was not qualified to be counted and were not in Hardy- Weinberg equilibrium.
vii
KATA PENGANTAR
Bismillahirahmanirahim…..
Assalamualaikumwarahmatullahiwabarakatuh
Puji syukur kita panjatkan kehadirat Allah SWT, karena rahmat dan hidayah-Nya sehingga Tugas Akhir / Skripsi ini dapat diselesaikan dengan tepat waktu. Skripsi dengan judul “Evaluasi Kemurnian Genetik Sapi Bali Menggunakan DNA Penciri Mikrosatelit Lokus HEL9 di Kabupaten Barru” Sebagai Salah Satu Syarat untuk memperoleh Gelar Sarjana pada Fakultas Peternakan Universitas Hasanuddin, Makassar.
Ucapan terima kasih dan penghargaan setinggi-tingginya penulis hanturkan dengan penuh rasa hormat kepada :
1. Secara khusus penulis mengucapkan terima kasih yang sebesar-besarnya dengan segenapcinta dan hormat kepada ayahanda tercinta Mansur Gani dan ibunda Gusniaty atas segala doa, motivasi,dan kasih sayang serta materi yang diberikan kepada penulis dan saudara-saudara saya Muslim Mansur, Marwa Mansur, Maghfira Mansur, Marjan Mansur, dan Muhammad Rafli Mansur yang senantiasa membantu dan memberikan motivasi untuk selalu lebih semangat.
2. Prof. Dr.Ir. Lellah Rahim, M.Sc selaku Pembimbing Utama dan Dr. Muhammad Ihsan Andi Dagong, S.Pt., M.Si selaku Pembimbing Anggota, atas segala bantuan dan keikhlasannya untuk memberikan bimbingan, nasehat dan saran-saran sejak awal penelitian sampai selesainya penulisan skripsi ini.
viii 3. Prof. Dr. Ir. H. Sudirman Baco, M.Sc, Prof. Dr. Ir. Djoni Prawira Rahardja, M.Sc, Dr. Hikmah M. Ali, S.Pt., M.Si, Prof. Dr. Ir. Hj. Sahari Banong, M.S dan drh.Hj. Farida Nur Yuliati, M.Si selaku dosen pembahas yang telah memberikan saran-saran dan masukan untuk perbaikan dari skripsi ini.
4. Dr. Hikmah M. Ali, S.Pt.,M.Si, dan drh. Kusumandari Indah Prahesti dan Dr. drh. Dwi Kesuma Sari, M.Sc selaku Penasehat Akademik yang telah memberikan bantuan dan motivasi kepada Penulis.
5. Prof. Dr. Syamsuddin Hasan, M.Sc selaku Dekan Fakultas Peternakan dan seluruh Staf Pegawai Fakultas Peternakan, terima kasih atas segala bantuan kepada penulis selama menjadi mahasiswa.
6. Prof. Dr. Ir. H. SudirmanBaco, M,Sc selaku Ketua Jurusan Produksi Ternak beserta seluruh Dosen dan Staf Jurusan Produksi Ternak atas segala bantuan kepada penulis selama menjadi mahasiswi.
7. Dr. Muhammad Yusuf, S.Pt sebagai Sekertaris Jurusan, terima kasih yang sebesar-besarnya atas bimbingan, dukungan dan bantuannya kepada Penulis. 8. Prof. Dr. Ir. H. Herry Sonjaya, DEA, DES sebagai Koordinator
Laboratorium Fisiologi Ternak, Terima kasih atas bimbingan, nasehat-nasehat, dan dukungannya kepada Penulis.
9. Semua Dosen-Dosen Fakultas Peternakan Universitas Hasanuddin yang telah memberikan ilmunya kepada Penulis.
10.Ricky Triwibowo yang telah memberikan motivasi, semangat dan dukungan selama ini kepada Penulis.
ix 11.Sahabat-sahabat ”FODRIN”, Weny, Inna, Nurmi, Tenri, Uchi, Dian, Rahmi, Evi, Iin, Linda, Putri, Lili, Vivi, Ceceng, Kiki, Maya, Farid, Alam, Rian, David, Iccank, Yafet, Aidil, Herman, Ichwan, Ibnu, Yogi, Aldes, Syahril, April, Munawir, Sudirman terima kasih yang setinggi-tingginya serta penghargaan yang sebesar-besarnya atas segala cinta, pengorbanan, bantuan, pengertian, candatawa serta kebersamaan selama ini, waktu yang dilalui sungguh merupakan pengalaman hidup yang berharga dan tak mungkin untuk terlupakan dan terima kasih telah memberiku sedikit tempat di hatimu untuk menjadikanku sahabat dan teriring dengan doa semoga rekan dan sahabatku sukses selalu.
12.Laboratorium Terpadu Peternakan UNHAS, Kak Nurul Purnomo, Kak Tri terima kasih atas bantuan dan perhatiannya selama penelitian dan penyelesaian skripsi.
10 Kepada Sahabat- Sahabat Penelitian Genetika Molekuler, Weny, Tenri, Kak Hendra, Kak Pymen, Kak Jaidin, Kak Uya, Ibu Ida. Terima kasih atas segala kebaikan serta bantuan yang kalian berikan kepada penulis selama penelitian.
11 Teman-teman HMI Komisariat Peternakan Cabang Makassar Timur, Terima kasih atas Ilmu, Pembelajaran, Nasehat-nasehat, Kebersamaan, Kebaikan, Amanah yang kalian berikan selama penulis berorganisasi.
x 13 Rekan-Rekan Asisten Fisiologi Ternak terima kasih atas bantuan dan canda tawa selama penulis menjadi asisten di Laboratotium Fisiologi Ternak Fakultas Peternakan.
14 Terima Kasih sebesar-besarnya kepada Mama Caya, Dg. Sai, dan Aming atas bantuannya kepada penulis dari masih MABA hingga saat ini.
15 Tanduk 01, Caput 02, Spider 03, Hamster 04, Lebah 05, Colagen 06, Rumput 07, Bakteri 08, Merpati 09, L10N, Matador 10, Situasi 10.
16 Teman- teman KKN Kelurahan Bone-Bone: Jenny, Nelsis, Resty, Anti, Nataly, Vivi, Sakina, Amhy, Mail, dan teman-tenan sekecamatan Bone-Bone Kabupaten Luwu Utara.
17 Jajaran Pemerintahan Kelurahan Bone-Bone Kabupaten Luwu Utara yang telah member banyak membantu Penulis pada saat KKN.
18 Dinas Peternakan Kabupaten Barru yang telah member izin untuk pengambilan sampel darah kepada Penulis.
19 Semua pihak yang tidak dapat penulis sebut satu persatu, terima kasih banyak atas segala bantuannya.
Penulis menyadari bahwa penyusunan skripsi ini masih terdapat kekurangan dan kesalahan. Penulis mengharapkan kritikan dan saran yang sifatnya membangun demi kesempurnaan skripsi ini.
Makassar, Juni 2014
xi DAFTAR ISI
Halaman
HALAMAN SAMPUL ... i
HALAMAN JUDUL ... ii
PERNYATAAN KEASLIAN ... iii
HALAMAN PENGESAHAN... ... iv
ABSTRAK... ... v
ABSTRACT... ... vi
KATA PENGANTAR... ... vii
DAFTAR ISI... ... xi
DAFTAR TABEL .. ... xiii
DAFTAR GAMBAR... ... xiv
DAFTAR LAMPIRAN ... xv
PENDAHULUAN ... 1
TINJAUAN PUSTAKA Asal Usul Sapi Bali dan Penyebarannya ... 4
Karakteristik Spesifik Sapi Bali ... 5
Upaya Pelestarian Sapi Bali ... 7
Perbaikan Mutu Genetik Sapi Bali ... 11
DNA Mikrosatelit ... 12
METODE PENELITIAN Waktu dan Tempat ... 15
Materi Penelitian ... 15
Metode Penelitian ... 16
Analisa Data... 19
HASIL DAN PEMBAHASAN Karakteristik Fenotip Sapi Bali ... 21
xii
Amplifikasi dan Genotip Mikrosatelit Lokus HEL9 pada Sapi Bali ... 25
Frekuensi Genotip, Alel dan Keseimbangan Hardy- Weinberg ... 27
Nilai Heterizigositas ... 29
Hubungan Sifat Fenotip dan Genotip ... 30
PENUTUP Kesimpulan ... ... 31 Saran ... 31 DAFTAR PUSTAKA ... 32 LAMPIRAN ... 36 RIWAYAT HIDUP
xiii DAFTAR TABEL
No. Teks Halaman
1. Urutan dan Ukuran HEL9 ... 18
2. Karakteristik Bentuk Tanduk Sapi Bali ... 21
3. Karakteristik Warna Bulu dan Kaos Kaki... 23
4. Sifat Kuantitatif Sapi Bali ... 24
5. Frekuensi Genotip Miktosatelit Lokus HEL9 ... 27
6. Frekuensi Alel dan Keseimbangan Hardy Weinberg ... 28
7. Nilai HeterozigositasPengamatan (Ho) dan Heterozigositas Harapan (He) ... 29
xiv DAFTAR GAMBAR
No. Teks Halaman 1. Bentuk-Bentuk Tanduk pada Sapi Bali ... 22 2. Visualisasi hasil amplifikasi Mikrosatelit Lokus HEL9 pada mesin
xv DAFTAR LAMPIRAN
No. Teks Halaman
1. Gambar Bentuk Tanduk Sapi Bali di Kabupaten Barru ... 36
2. Pola Warna Bulu Sapi Bali di Kabupaten Barru ... 37
3. Analisis Data dengan Menggunakan Software PopGene32 Versi 1.31 ... 38
1 PENDAHULUAN
Sapi Bali (Bos sondaicus) merupakan salah satu sapi potong asli Indonesia hasil domestikasi dari Banteng (Bos-bibos banteng) dan memiliki potensi yang besar untuk mensuplai kebutuhan protein masyarakat Indonesia. Penyebaran sapi Bali saat ini hampir meliputi seluruh wilayah Indonesia, kecuali Propinsi DKI Jakarta. Empat propinsi yang memiliki jumlah sapi Bali terbesar di Indonesia adalah Propinsi Sulawesi Selatan, NTB, Bali dan NTT.
Sapi Bali merupakan aset nasional yang perlu dilestarikan. Pemerintah telah sejak lama memberikan perhatian yang cukup besar bagi pelestarian plasma nutfah ini dengan menetapkan program nasional pemuliaan untuk sapi Bali. Program nasional tersebut meliputi program pemurnian dan peningkatan mutu genetik sapi Bali. Program pemurnian sapi Bali dilaksanakan dengan penetapan wilayah peternakan murni sapi Bali di Sulawesi Selatan khususnya Kabupaten Barru dan Bone.
Program pelestarian sapi Bali perlu didukung dengan metode identifikasi kemurnian sapi Bali yang cepat, mudah, dan akurat, agar pelestarian sapi Bali pada suatu wilayah tertentu dapat berhasil dilakukan tanpa ada keraguan terhadap kemurniannya. Identifikasi penciri genetik sebagai alat penanda pembeda bangsa merupakan langkah penting sebagai ciri khusus yang bersifat khas pada bangsa ternak tertentu.
Pengujian lokus DNA mikrosatelit HEL9 dan INRA035 dilakukan Handiwirawan (2003) dalam sampel yang lebih besar pada sapi Bali di Propinsi Bali. Hasil identifikasi genotipe sapi Bali dan genotipe Banteng sebagai
2 pembanding menunjukkan bahwa alel A dan alel B merupakan alel yang monomorfik pada lokus mikrosatelit INRA035 pada sapi Bali sehingga dapat digunakan sebagai penciri genetik sapi Bali. Alel A pada lokus mikrosatelit HEL9 merupakan alel dengan frekuensi yang sangat tinggi pada sapi Bali (92,9%) yang dapat dipergunakan sebagai penciri genetik pendukung pada sapi Bali karena semua Banteng yang di uji juga memiliki alel tersebut.
Pada berbagai lingkungan pemeliharaan di Indonesia, sapi Bali memperlihatkan kemampuannya untuk berkembang biak dengan baik yang disebabkan beberapa keunggulan yang dimiliki sapi Bali. Keunggulan sapi Bali dibandingkan sapi lain yaitu memiliki daya adaptasi sangat tinggi terhadap lingkungan yang kurang baik (Masudana, 1990), seperti dapat memanfaatkan pakan dengan kualitas rendah (Sastradipradja, 1990), mempunyai fertilitas dan
conception rate yang sangat baik (Oka dan Darmadja, 1996), persentase karkas yang tinggi yaitu 52-57,7% (Payne dan Rollinson, 1973), memiliki daging berkualitas baik dengan kadar lemak rendah (kurang lebih 4%) (Payne dan Hodges, 1997) dan tahan terhadap parasit internal dan eksternal (National Research Council, 1983). Dengan berbagai keunggulan yang dimiliki tersebut dan mengingat Indonesia merupakan pusat sapi Bali di dunia maka sapi Bali merupakan aset nasional yang perlu dilestarikan.
Berdasarkan uraian tersebut di atas maka perlu dilakukan penelitian mengenai evaluasi kemurniaan genetik sapi Bali menggunakan DNA penciri mikrosatelit lokus HEL 9 di kabupaten Barru.
3 Tujuan dari penelitian ini adalah untuk mengetahui tingkat kemurnian sapi Bali di Kabupaten Barru berdasarkan identifikasi fenotipe (bentuk tanduk, warna bulu, dan kaos kaki) dengan menggunakan DNA penciri mikrosatelit HEL9. Kegunaan dari penelitian ini adalah memberikan informasi kepada stakeholder peternakan tentang kemurnian sapi Bali sehingga dapat mempertahankan kelestarian sapi lokal dan menambah wawasan ipteks peternakan.
4 TINJAUAN PUSTAKA
Asal Usul Sapi Bali dan Penyebarannya
Sapi Bali (Bibos sondaicus) yang ada saat ini diduga berasal dari hasil domestikasi Banteng liar (Bibos banteng). Menurut Rollinson (1984) proses domestikasi sapi Bali itu terjadi sebelum 3.500 SM di Indonesia atau Indochina. Banteng liar saat ini bisa di temukan di Jawa bagian Barat dan bagian Timur, di pulau Kalimantan, serta ditemukan juga di Malaysia (Payne dan Rollinson, 1973). Hardjosubroto dan Astuti (1993) mengemukakan bahwa di Indonesia saat ini, Banteng liar hanya terdapat di hutan lindung Baluran, Jawa Timur dan Ujung Kulon, Jawa Barat, serta di beberapa kebun binatang. Adanya Banteng liar ini memberikan peluang untuk perbaikan mutu sapi Bali atau untuk persilangan dengan jenis sapi lain (National Research Council, 1983).
Tempat dimulainya domestikasi sapi Bali masih terdapat perbedaan pendapat, dimana Meijer (1962) berpendapat proses domestikasi terjadi di Jawa, namun Payne dan Rollinson (1973) menduga asal mula sapi Bali adalah dari pulau Bali mengingat tempat ini merupakan pusat distribusi sapi Bali di Indonesia. Nozawa (1979) menduga gen asli sapi Bali berasal dari pulau Bali yang kemudian menyebar luas ke daerah Asia Tenggara, dengan kata lain bahwa pusat gen sapi Bali adalah di pulau Bali, di samping pusat gen sapi zebu di India dan pusat gen primigenius di Eropa.
Penyebaran sapi Bali di Indonesia dimulai pada tahun 1890 dengan adanya pengiriman ke Sulawesi, pengiriman selanjutnya dilakukan pada tahun 1920 dan 1927 (Herweijer, 1950). Kemudian pada sekitar tahun 1947 dilakukan pengiriman
5 besar-besaran sapi Bali oleh pemerintah Belanda ke Sulawesi Selatan yang langsung didistribusikan kepada petani (Pane, 1991). Sejak saat itu, populasi sapi Bali berkembang dengan cepat sehingga sampai saat ini Propinsi Sulawesi Selatan menjadi propinsi yang memiliki sapi Bali dengan jumlah terbesar di Indonesia. Untuk penyebaran sapi Bali ke Lombok mulai dilakukan pada abad ke-19 yang dibawa oleh raja-raja pada zaman itu (Hardjosubroto dan Astuti, 1993), dan sampai ke Pulau Timor antara tahun 1912 dan 1920 (Herweijer, 1950).
Penyebaran sapi Bali ke banyak wilayah di Indonesia kemudian dilakukan sejak tahun 1962 (Hardjosubroto dan Astuti, 1993) dan saat ini telah menyebar hampir di seluruh wilayah Indonesia. Tidak hanya di wilayah Indonesia, sapi Bali juga telah disebarkan ke berbagai negara. Tercatat sapi Bali telah diintroduksikan ke Semenanjung Cobourg di Australia Utara di antara tahun 1827 dan 1849. Pernah juga dilakukan ekspor secara reguler sapi Bali kastrasi ke Hongkong untuk dipotong. Selain itu, pada masa lalu, sapi Bali juga pernah dikirim ke Philipina, Malaysia dan Hawai (Payne dan Rollinson, 1973), telah juga dikirimkan ke Texas, USA dan New South Wales, Australia sebagai ternak percobaan (National Research Council, 1983).
Karakteristik Spesifik Sapi Bali
Sapi Bali mempunyai ciri-ciri fisik yang seragam, dan hanya mengalami perubahan kecil dibandingkan dengan leluhur liarnya (Banteng). Warna sapi betina dan anak atau muda biasanya coklat muda dengan garis hitam tipis terdapat di sepanjang tengah punggung. Warna sapi jantan adalah coklat ketika muda tetapi kemudian warna ini berubah agak gelap pada umur 12-18 bulan
6 sampai mendekati hitam pada saat dewasa, kecuali sapi jantan yang dikastrasi akan tetap berwarna coklat. Pada kedua jenis kelamin terdapat warna putih pada bagian belakang paha (pantat), bagian bawah (perut), keempat kaki bawah (white stocking) sampai di atas kuku, bagian dalamtelinga, dan pada pinggiran bibir atas (Payne dan Rollinson, 1973; National Research Council, 1983; Hardjosubroto dan Astuti,1993).
Sapi Bali jantan maupun betina mempunyai tanduk, yang berbeda dalam ukuran dan bentuknya dan ada beberapa variasi tipe tanduk pada kedua jenis kelamin tersebut (Payne dan Rollinson, 1973). Panjang tanduk sapi jantan biasanya 20 sampai 25 cm, bentuk tanduk yang ideal pada sapi jantan disebut bentuk tanduk silak conglok yaitu jalannya pertumbuhan tanduk mula-mula dari dasar sedikit keluar (tumbuh ke arah samping), lalu membengkok ke atas dan kemudian pada ujungnya membengkok sedikit ke arah luar. Pada yang betina, bentuk tanduk yang ideal disebut manggul gangsa yaitu jalannya pertumbuhan tanduk satu garis dengan dahi arah ke atas dan pada ujungnya sedikit mengarah ke belakang dan kemudian melengkung ke bawah lagi mengarah ke kepala (ke dalam). Sapi Bali yang tidak bertanduk tidak pernah ditemukan (Payne dan Rollinson, 1973; National Research Council, 1983; Hardjosubroto, 1994). Kepala sapi Bali termasuk panjang tetapi tidak lebar, kedua telinganya tegak dan berukuran sedang (Payne dan Rollinson, 1973).
Terdapat variasi bentuk tanduk pada sapi Bali di Propinsi Bali. Hasil penelitian Handiwirawan (2003) mendapatkan bahwa pada sapi Bali jantan terdapat 7 macam bentuk tanduk sedangkan pada yang betina diperoleh 12 macam
7 bentuk tanduk. Bentuk tanduk yang umum untuk sapi Bali jantan (seperti didefinisikan Payne dan Rollinson, 1973; National Research Council, 1983; Hardjosubroto, 1994) ternyata bukan merupakan bentuk yang umum dimiliki sapi Bali jantan yang teramati. Bentuk tanduk ini hanya dimiliki oleh 6,5% dari sapi Bali jantan yang diamati. Sapi Bali jantan pada umumnya memiliki bentuk tanduk ke samping kemudian ke atas atau ke samping-ke atas kemudian ke belakang, dan proporsi sapi yang bertanduk demikian adalah sebesar 74,5% dari keseluruhan sapi jantan yang diamati. Bentuk umum tanduk sapi Bali betina sesuai dengan bentuk normal yang didefinisikan oleh Payne dan Rollinson (1973), National Research Council (1983) dan Hardjosubroto (1994), yaitu mencapai 31,9%. Ditemukan juga bentuk tanduk yang sedikit berbeda dengan bentuk tanduk yang normal dalam jumlah cukup b esar (29,3%), yaitu mengarah ke atas dan kemudian ke belakang.
Upaya Pelestarian Sapi Bali
Perhatian pemerintah yang lebih besar terhadap upaya pengembangbiakan ternak di Indonesia, selanjutnya diwujudkan dengan dikeluarkannya Undang Undang No. 6 tahun 1967 yang merupakan landasan bagi upaya pengembangbiakan ternak untuk mempertahankan dan meningkatkan mutu bangsa ternak di Indonesia melalui upaya pemurnian atau melalui persilangan antar bangsa ternak (Djarsanto, 1997).
Khusus untuk sapi Bali, telah ditetapkan program nasional yang meliputi program pemurnian dan peningkatan mutu genetik. Sebagai wilayah peternakan murni sapi Bali ditetapkan Pulau Bali, NTB, NTT dan Sulawesi Selatan. Dimulai
8 pada tahun 1976, di pulau Bali telah dilaksanakan program pemuliaan sapi Bali dengan melakukan seleksi dalam bangsa, untuk memperoleh bibit sapi Bali yang baik mutunya melalui Proyek Pengembangan dan Pembibitan Sapi Bali (P3Bali) (Soehadji, 1990). Sementara itu, persilangan hanya dapat dilakukan di luar wilayah peternakan murni.
Dalam perkembangannya di wilayah peternakan murni (sumber bibit) telah terjadi pencemaran genetik sapi Bali dengan bangsa sapi lain (Bos taurus dan zebu), kecuali sapi Bali yang ada di Pulau Bali. Sehubungan dengan itu, daerah di luar Bali yang ditetapkan sebagai daerah pemurnian sapi Bali lebih dipersempit lagi, yaitu Pulau Sumbawa di NTB, Pulau Flores di NTT, Kabupaten Bone di Sulawesi Selatan dan Kabupaten Lampung Selatan di Propinsi Lampung. Walaupun demikian, Pane (1991) mengemukakan bahwa hingga kini hanya sapi Bali yang terdapat di pulau Bali yang masih dapat dipertanggungjawabkan kemurniannya. Upaya penetapan daerah peternakan murni sekaligus dengan meningkatkan produktivitas sapi Bali melalui kegiatan seleksi secara terencana tentunya akan sangat mendukung program pelestarian plasma nutfah ternak asli tersebut.
Pelestarian sapi Bali perlu terus dilakukan dimana kegiatan tersebut harus dipandang sebagai upaya antisipatif penyediaan “bahan baku” bagi perakitan bangsa sapi baru untuk dapat mengantisipasi perubahan selera pasar di masa depan yang tidak mudah untuk diprediksi. Sebagai contoh akhir-akhir ini seiring dengan meningkatnya kesadaran untuk mengkonsumsi pangan sehat dan timbulnya perhatian/pandangan yang negatif terhadap pangan berkolesterol maka
9 terdapat perubahan permintaan pada industri peternakan untuk menghasilkan daging dengan kandungan lemak rendah (lean) dan tentunya dihasilkan dari suatu bangsa ternak tertentu. Melalui program persilangan permintaan pasar tersebut dapat direspon dengan membentuk suatu bangsa baru yang lebih sesuai dengan kondisi lingkungan dan pasar di masa mendatang. Beberapa bangsa sapi terkenal seperti Brangus, Santa Gertrudis, Droughmaster, dan lain-lain merupakan bangsa sapi unggul sebagai hasil persilangan dari beberapa macam bangsa sapi yang masih dipertahankan ada/dilestarikan (Handiwirawan dan Subandriyo, 2004).
Program pelestarian sapi Bali perlu didukung dengan metode identifikasi kemurnian sapi Bali yang cepat, mudah dan akurat, agar pelestarian sapi Bali pada suatu wilayah tertentu dapat berhasil dilakukan tanpa ada keraguan terhadap kemurniannya. Metode identifikasi kemurnian sapi Bali yang andal juga sangat membantu penentuan pemilihan pejantan sapi Bali sebagai pejantan yang dipergunakan di Balai Inseminasi Buatan sehingga pencemaran genetik pada wilayah yang ditentukan sebagai tempat pelestarian sapi Bali murni dapat dihindari (Handiwirawan dan Subandriyo, 2004).
Serangkaian penelitian telah dilakukan untuk mengidentifikasi penciri genetik sapi Bali yang diharapkan dapat dipergunakan sebagai metode untuk menentukan kemurnian sapi Bali. Anonim (2000) telah merintis untuk mulai mencari sifat fisik/fenotip (struktur bulu) maupun dari aspek molekuler (protein darah dan DNA mikrosatelit) yang dapat dipergunakan sebagai penciri bangsa sapi Bali. Walaupun sulit dijustifikasi, terdapat perbedaan struktur bulu rambut sapi Bali dengan struktur bulu bangsa sapi lainnya (Simental Limousin dan
10 Brangus). Namun hasil penelitian ini perlu diuji lagi di lapangan khususnya dalam penurunan alel dari tetua ke progeninya. Hasil pengamatan pada lokus Haemoglobin β menunjukkan bahwa alel haemoglobin βX (Hb βX) adalah khas dan mirip seperti yang dilaporkan Namikawa et al. (1982) bahwa alel haemoglobin βX (Hb βX) adalah alel yang paling umum ditemukan pada sapi Bali dan pada Banteng sebagai leluhur dari sapi Bali. Dari hasil penelitian pada empat bangsa sapi dengan sampel yang terbatas, Winaya (2000) mengemukakan bahwa lokus DNA mikrosatelit HEL9 pada sapi Bali adalah monomorfik, sedangkan pada bangsa sapi lain (Madura, PO dan Brangus) adalah polimorfik. Hasil penelitian Anonim (2000) dengan sampel terbatas pada tiga bangsa sapi lainnya (Simmental, Brangus, dan Limousin) memperkuat hasil penelitian tersebut. Selain itu diperoleh juga hasil bahwa lokus DNA mikrosatelit INRA035 pada seluruh sampel sapi Bali yang diteliti konsisten mempunyai dua alel sehingga kemungkinan dapat dipergunakan sebagai penciri genetik untuk sapi Bali. Handiwirawan (2003) melakukan pengujian lokus DNA mikrosatelit HEL9 dan INRA035 dalam sampel yang lebih besar pada sapi Bali di Propinsi Bali. Hasil identifikasi genotipe sapi Bali dan genotipe banteng sebagai pembanding menunjukkan bahwa alel A dan alel B merupakan alel yang monomorfik pada lokus mikrosatelit INRA035 pada sapi Bali sehingga dapat digunakan sebagai penciri genetik sapi Bali. Alel A pada lokus mikrosatelit HEL9 merupakan alel dengan frekuensi yang sangat tinggi pada sapi Bali (92,9%) yang dapat dipergunakan sebagai penciri genetik pendukung pada sapi Bali karena semua Banteng yang diuji juga memiliki alel tersebut. Dari beberapa hasil penelitin
11 tersebut penggunaan lokus mikrosatelit INRA035 sebagai penciri genetik sapi Bali dapat digunakan, namun sebaiknya dikombinasikan dengan pengujian lokus HEL9 dan identifikasi fenotipe (pola warna tubuh, bentuk tanduk dan struktur bulu) agar diperoleh hasil yang lebih akurat.
Perbaikan Mutu Genetik Sapi Bali
Program pemuliaan khusus untuk sapi Bali telah ditetapkan dan dijalankan pemerintah. Pokok-pokok pemuliaan sapi Bali seperti dikemukakan Soehadji (1990) adalahmeliputi:
1. Menjalankan peternakan murni sapi Bali di Pulau Bali, NTB, Pulau Timor dan beberapa daerah di Sulawesi Selatan sebagai sumber bibit sapi Bali secara nasional,
2. Melakukan uji performans dan uji zuriat di breeding centre P3Bali Pulukan Bali untuk memperoleh pejantan sapi Bali unggul yang digunakan untuk kawin alam atau produksi semen beku,
3. Membentuk populasi dasar sebagai sumber gen yang unggul dan membentuk kelompok sapi Bali betina unggul dan dipelihara di Pusat Pembibitan Sapi Bali di Pulukan, Bali dan Anamina, Dompu-Sumbawa,
4. Melakukan inseminasi buatan berskala nasional untuk mempercepat aliran gen yang unggul dari pejantan sapi Bali unggul.
Proyek Pembibitan dan Pengembangan Sapi Bali (P3Bali) dilaksanakan sebagai upayauntuk memperbaiki mutu genetik sapi Bali diPropinsi Bali melalui seleksi, uji performans dan uji keturunan (progeny test). Dalam kegiatan ini, pejantan elit yang dihasilkan dari uji keturunan akan dipergunakan BIB untuk
12 diambil semennya guna memperbaiki mutugenetik sapi Bali di seluruh Indonesia. Dari kegiatan ini terlihat bahwa performans produksi dan reproduksi sapi Bali di P3Bali dilaporkan lebih baik dibandingkan sapi Bali yang terdapat di Propinsi Bali, NTB, NTT danSulsel (Pane, 1990). Sukmasari (2003)dengan menggunakan metode BLUP (best linear unbiased prediction) mendapatkan hasil bahwa sapi Bali yang dipelihara di breeding center Pulukan mempunyai rataan nilai pemuliaan dugaan lebih tinggi dibandingkan di instalasi populasi dasar. Secara keseluruhan, mulai tahun 1983 sampai 1999 kecenderungan genetik sapi Bali di P3 Bali mengalami fluktuasi dari tahun ke tahun. Dari hasil penelitiannya tersebut, Sukmasari (2003) jugamenyarankan bahwa seleksi agar didasarkanpada nilai pemuliaan agar seleksi dapat dilakukan lebih akurat sehingga kecenderungan genetik sapi Bali di P3 Bali terus meningkat.
DNA Mikrosatelit
Salah satu penanda molekuler yang sangat popular adalah mikrosatelit. Mikrosatelit adalah runutan DNA pendek yang berulang 2 sampai 6 basa nukleotida dan dapat berulang 10-100 kali, runutan yang paling banyak ditemukan pada mamalia adalah (dC – dA) dan (dT - dG) (Moore et al., 1991). Mikrosatelit merupakan salah satu penanda genetic yang sudah diaplikasikan secara luas dalam bidang peternakan. Selain untuk uji test keturunan, dapat pula digunakan untuk mengidentifikasi sejumlah sifat dengan nilai ekonomi tinggi dikarenakan DNA mikrosatelit sangat polimorfik dan terdapat banyak dalan DNA genom (Bawden dan Nicholas, 1999).
13 Mikrosatelit (SSR = Simple Sequence Repeat) merupakan salah satu marka molekuler yang berupa urutan di-nukleotida sampai tetra-nukleotida yang berulang dan berurutan. Mikrosatelit merupakan marka genetik yang bermanfaat karena bersifat kodominan, dapat mendeteksi keragaman alel pada tingkat tinggi, serta mudah dan tidak terlalu mahal untuk dianalisis dengan menggunakan PCR (Moeljopawiro, 2010).
Aplikasi penanda genetik molekuler untuk seleksi dan pemuliaan dapat meningkatkan mutu genetik ternak (Bawden dan Nicholas, 1999). Mikrosatelit merupakan penanda genetik yang sering digunakan untuk mempelajari sistem perkawinan dan struktur populasi (Steffen et al., 1993), pautan (linkage), pemetaan kromosom, dan analisis populasi (Silva et al., 1999). Mikrosatelit banyak digunakan oleh peneliti sebagai marka karena keberadaannya melimpah, bersifat kodominan dan sangat polimorfik (Bennett, 2000).
Marka mikrosatelit merupakan sekuen DNA yang bermotif pendek diulang secara tandem dengan 2 sampai 6 unit nukleotida yang tersebar dan meliputi seluruh genom, terutama pada organisme eukariotik. Pasangan mikrosatelit (forward dan reverse) diaplikasikan dengan PCR berdasarkan hasil konservasi daerah yang diapit (flanking-region) marka untuk suatu gen pada kromosom. Menurut Powell et al (1996), beberapa pertimbangan untuk menggunakan marka mikrosatelit dalam studi genetik diantaranya (1) marka tersebut terdistribusi secara melimpah dan merata dalam genom, variabilitasnya sangat tinggi (banyak alel dalam lokus), sifatnya kodominan dan lokasi genom dapat diketahui; (2) merupakan alat uji yang memiliki reproduksibilitas dan ketepatan yang sangat
14 tinggi; (3) merupakan alat bantu yang sangat akurat untuk membedakan genotip, evaluasi kemurnian benih, pemetaan, dan seleksi genotip untuk karakter yang diinginkan; (4) studi populasi dan analisis diversitas genetik.
15 METODE PENELITIAN
Waktu dan Tempat
Penelitian dilaksanakan selama bulan Februari – Maret 2014. Untuk pengambilan sampel di Kabupaten Barru dan analisis DNA bertempat di Laboratorim Bioteknologi Terpadu, Fakultas Peternakan Universitas Hasanuddin, Makassar.
Materi Penelitian
Bahan utama dari penelitian ini adalah sampel darah sapi Bali sebanyak 80 sampel yang terdiri dari 35 sampel sapi jantan dan 45 sampel sapi betina yang berumur 2 tahun keatas di Kabupaten Barru. Bahan pendukung antara lain: Primer (primer mikrosatelit lokus HEL 9), bahan ekstraksi DNA (Kit DNA ekstraksi (Thermo Scientific), Proteinase K, ethanol 96% ), bahan PCR (dNTP mix, Enzim Taq DNA polymerase, 10x buffer, 10x TBE buffer), bahan elektoforesis (agarose,
Ethidium bromide, Marker DNA 100pb, Loading dye), bahan gel poliakrilamid (acrylamida, bis-akrilamida, APS, TBE, H2O), bahan stainning (AgNO3, NaOH, NH4OH, asam asetat glasial, formalin, gliserol 20%, aquades) tissue dan plastik mika.
Alat yang digunakan yaitu : venoject, tabung vakuttainer, mesin PCR (sensoQuest Germany), centrifuge, alat pendingin, tabung eppendorf besar kecil, gel documention, mikropipet, tip, rak tabung, elektroforesis, autoclave, timbangan, sarung tangan.
16 Metode Penelitian
- Identifikasi Fenotipe
Identifiksi fenotipe sapi Bali dilakukan dengan mengidentifikasi sifat kualitatif khas yang dimiliki sapi Bali. Adapun sifat – sifat kualitatif yang di identifikasi antara lain pola warna bulu, struktur tubuh, bentuk dan ukuran tanduk, serta ciri–ciri fisik khusus pada sapi Bali seperti bulu hitam yang membentuk garis (garis belut) pada bagian punggung dan warna putih pada bagian belakang paha (pantat), bagian bawah (perut), keempat kaki bawah (white stocking) sampai di atas kuku, bagian dalam telinga, dan pada pinggiran bibir atas, sifat kuantitatif seperti bobot badan badan, panjang badan, lingkar dada, dan tinggi badan.
- Evaluasi Kemurnian Genetik Sapi Bali dengan DNA Penciri Mikrosatelit
Pada tahapan ini dilaksanakan dalam beberapa tahap dengan metode yang meliputi :
a. Koleksi Sampel Darah
Materi genetik adalah sampel DNA yang diambil dari darah utuh (whole blood) sapi Bali yang berasal dari daerah sentra pemurnian di Sulawesi Selatan yakni di Kabupaten Barru dengan sejumlah 80 sampel dari 5 kecamatan yang ada yakni: Soppeng Riaja, Tanete Rilau, Barru, Balusu, Tanete Riaja. Pengambilan sampel darah dilakukan dengan mengumpulkan sekitar 5 ml sampel darah dari sapi melalui vena jugularis dengan menggunakan venojet dan tabung vacuntainer yang diberi antikoagulan (heparin atau EDTA). Sampel darah tersebut kemudian disimpan pada suhu 4oC sampai waktu dianaisis (ekstraksi DNA dan PCR).
17 b. Ekstraksi DNA
DNA diisolasi dan dimurnikan dengan menggunakan Kit DNA ekstraksi Genjet Genomic DNA Extraction (Thermo Scientific) dengan mengikuti protokol ekstraksi yang disediakan. Sebanyak 200 μl sampel darah dilisiskan dengan menambahkan 400 μl larutan lysis buffer dan 20 μl Proteinase K (10 mg/ml),
dicampurkan kemudian diinkubasi pada suhu 56 ºC selama 60 menit di dalam
waterbath shaker. Setelah inkubasi larutan kemudian ditambahkan 200 μl Ethanol absolute 96% dan disentrifugasi 6.000 x g selama 1 menit.
Pemurnian DNA kemudian dilakukan dengan metode spin column dengan penambahan 500 μl larutan pencuci wash buffer I yang kemudian dilanjutkan dengan sentrifugasi pada 8.000 x g selama 1 menit. Setelah supernatannya dibuang, DNA kemudian dicuci lagi dengan 500 μl wash buffer II dan disentrifugasi pada 12.000 x g selama 3 menit. Setelah supernatannya dibuang, DNA kemudian dilarutkan dalam 200 μl elution buffer dan disentrifugasi pada 8.000 x g untuk selanjutnya DNA hasil ekstraksi ditampung dan disimpan pada suhu -20 ºC.
c. Analisis PCR dengan Penciri DNA Mikrosatelit HEL 9
Analisis tingkat kemurnian genetik sapi Bali di Kabupaten Barru digunakan dengan menggunakan pendekatan marker mikrosatelit pada lokus HEL 9 sebagai DNA penciri genetik khas sapi Bali (Handiwirawan, 2003 dan Maskur dkk, 2007).
Komposisi reaksi PCR dikondisikan pada volume reaksi 25 μl yang terdiri atas 100 ng DNA, 025 mM masing – masing primer, 150 uM dNTP, 2,5 mM
18 Mg2+, 0,5 U Taq DNA polymerase dan 1x buffer. Kondisi mesin PCR dimulai dengan denaturasi awal pada suhu 94 oC selama 2 menit, diikuti dengan 35 siklus berikutnya masing – masing denaturasi 94 oC selama 45 detik, dengan suhu annealing lokus DNA mikroatelit lokus HEL 9 yaitu 58 oC selama 30 detik yang dilanjutkan dengan ekstensi : 72 oC selama 60 detik, yang kemudian diakhiri dengan satu siklus ekstensi akhir pada suhu 72 oC selama 5 menit dengan menggunakan mesin PCR (SensoQuest, Germany).
Analisis produk PCR dan deteksi terhadap alel mikrosatlit dilakukan dengan elektroforesis pada gel poliakrilamida dan pewarnaan dengan perak mengikuti metode Tegelstrom (1992).
d. Primer Mikrosatelit
Penelitian ini menggunakan DNA penciri mikrosatelit lokus HEL 9 sebagai penanda molekul. Penanda tersebut dipilih berdasarkan dalam Bishop et al., (1994) karena dapat menunjukkan polimorfisme pada sapi Bali.
Tabel 1. Urutan dan Ukuran HEL 9.
Loci Primer sequences (5’ – 3’) Number of allele T0C Size (bp) Sources HEL 9 F : CCCATTCAGTCTTCAGAGGT 12 55 143-165 Bioshop et al.,(1994) R : CACATCCATGTTCTCACCAC F = Forward, R = Reverse
- Elektroporesis pada Gel Poliakrilamid
Komponen gel poliakrilamida terdiri atas campuran 30% acrylamida dan bis-akrilamida sebanyak 6 ml, 10 x TBE sebanyak 6 ml, H2O sampai mencapai volume 30 ml, temed sbanyak 20 μl, 10% APS 200 μl. Sampel DNA tersebut kemudian dimasukkan ke dalam sumur gel setelah gel diletakkan pada tangki
19 elektroforesis yang telah berisi larutan penyangga 1 x TBE. Elektroforesis dilakukan pada voltase konstan 250 V selama 120 menit pada suhu ruang.
- Silver stainning (Pewarnaan Perak)
Pewarnaan dengan perak dilakukan melalui serangkaian proses yaitu pewaraan gel dengan larutan stainning dengan merendam gel dalam larutan yang terdiri atas 0,2 g AgNO3 ; 80 μl NaOH 10 N ; 0,8 ml NH4OH ; 200 ml akuades selama selama 15 menit. Gel kemudian dicuci kembali dengan aquades selama 20 menit sambil digoyang untuk menghilangkan perak yang tidak berikatan dengan DNA. Fragmen DNA yang berikatan dengan perak dapat dideteksi dengan merendam gel dalam larutan NaOH 0,03 g/ml dan formalin yang dipanaskan pada suhu 45 oC sampai fragmen pita DNA tampak. Setelah fragmen DNA tampak, reaksi kemudian dihentikan dengan menggunakan asam asetat glasial (200 μl / 200 ml aquades).
- Penentuan posisi pita DNA
Penentuan posisi pita DNA pada gel poliakrilamida dilakukan secara manual. Pita DNA yang muncul pada gel poliakrilamida diasumsikan sebagai alel mikrosatelit. Ukuran dan jumlah dari alel yang muncul pada gel ditentukan berdasarkan asumsi bahwa semua pita DNA dengan laju migrasi yang sama adalah homolog (Leung et al., 1993), sedangkan alel dengan migrasi paling cepat ditetapkan sebagai alel A, berikutnya adalah alel B dan seterusnya.
Analisis Data
Keragaman alel mikrosatelit ditentukan dari perbedaan migrasi alel pada gel dari masing-masing individu sampel. Karena alel mikrosatelit adalah
20 kodominan maka genotip ditentukan berdasarkan variasi pita alel yang ada. Kemudian dihitung frekuensi masing-masing alel setiap lokus. Keseimbangan Hardy- Weinberg di uji dengan test chi-square (Nei dan Kumar, 2000).
∑ Keterangan:
X2 = nilai uji chi-square
O = jumlah pengamatan genotip ke-i E = jumlah harapan genotip ke-i
- Nilai Heterozigositas Pengamatan (Ho) dan Harapan (He)
Nilai Heterozigositas teramati (Ho) dan heterozigonitas harapan (He) dapat digunakan untuk menduga nilai koefisien inbreeding pada suatu kelompok ternak. Perhitungan Ho dan He dilakukan menurut Hartl (1988) dengan formula sebagai berikut:
∑ Keterangan:
Ho = heterozigositas pengamatan
N1ij = jumlah individu heterozigot pada lokus ke-l N = jumlah individu yang diamati
Nilai heterozigonitas harapan (He) dihitung dengan formula:
∑ Keterangan:
He = heterozigositas harapan
P1i = frekuensi alel ke-1 pada lokus ke-l n = jumlah alel pada lokus ke-l
21 HASIL DAN PEMBAHASAN
Karakteristik Fenotip Sapi Bali
Sifat kualitatif merupakan sifat yang tampak dari luar dan tidak dapat dihitung. Yang termasuk dalam sifat kualitatif, seperti bentuk tanduk, warna bulu, dan warna kaos kaki. Hasil pengamatan karakteristik sapi Bali berdasarkan bentuk tanduk dapat dilihat pada Tabel 2.
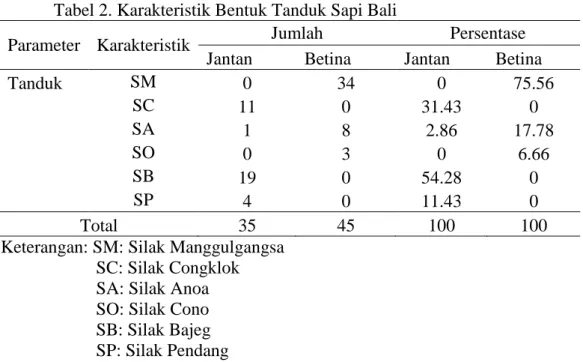
Tabel 2. Karakteristik Bentuk Tanduk Sapi Bali
Parameter Karakteristik Jumlah Persentase
Jantan Betina Jantan Betina
Tanduk SM 0 34 0 75.56 SC 11 0 31.43 0 SA 1 8 2.86 17.78 SO 0 3 0 6.66 SB 19 0 54.28 0 SP 4 0 11.43 0 Total 35 45 100 100
Keterangan: SM: Silak Manggulgangsa SC: Silak Congklok SA: Silak Anoa SO: Silak Cono SB: Silak Bajeg SP: Silak Pendang
Berdasarkan Tabel 2. memperlihatkan bahwa terdapat 6 macam bentuk tanduk yaitu Silak Manggulgangsa, Silak Congklok, Silak Anoa, Silak Cono, Silak Bajeg, Silak Pendang. Bentuk tanduk yang paling dominan pada sapi jantan adalah Silak Bajeg (54,28%) sedangkan pada sapi betina yaitu Silak Manggulgangsa (75,56%). Pada penelitian yang dilakukan Handiwirawan (2003) bahwa bentuk tanduk yang dominan pada jantan adalah Silak Congklok (74,5%) dan pada sapi benita adalah Silak Manggulgangsa (31,9%). Sapi Bali jantan
22 umumnya memiliki bentuk tanduk Silak congklok yaitu jalannya pertumbuhan tanduk mula-mula dari dasar sedikit keluar lalu membengkok keatas, kemudian pada ujungnya membengkok sedikit keluar. Pada sapi betina bentuk tanduk yang ideal disebut manggulgangsa yaitu jalannya pertumbuhan tanduk satu garis dengan dahi arah kebelakang sedikit melengkung kebawah dan pada ujungnya sedikit mengarah kebawah dan kedalam, tanduk ini berwarna hitam (Payne dan Rollinson, 1973; National Research Council, 1983; Hardjosubroto, 1994).
Betina Jantan
Gambar 1. Bentuk-Bentuk Tanduk pada Sapi Bali Silak Manggulgangsa Silak Anoa Silak Cono Silak Bajeg Silak Congklok Silak Pendang
23 Variasi bentuk tanduk sapi Bali betina lebih sedikit daripada sapi Bali jantan. Bradley dan Cunningham (1999) mengemukakan bahwa perbedaan bentuk fenotipe ternak domestikasi saat ini dengan leluhurnya adalah akibat proses domestikasi. Banteng jantan memiliki bentuk tanduk yang sedikit berbeda dibandingkan sapi Bali jantan, National Research Council (1983) mengemukakan bahwa bentuk tanduk banteng jantan adalah mengarah keluar dan kemudian keatas dengan ujung tanduk mengarah kedalam. Hasil analisis karakteristik sapi Bali berdasarkan warna bulu dan warna kaos kaki dapat dilihat pada Tabel 3..
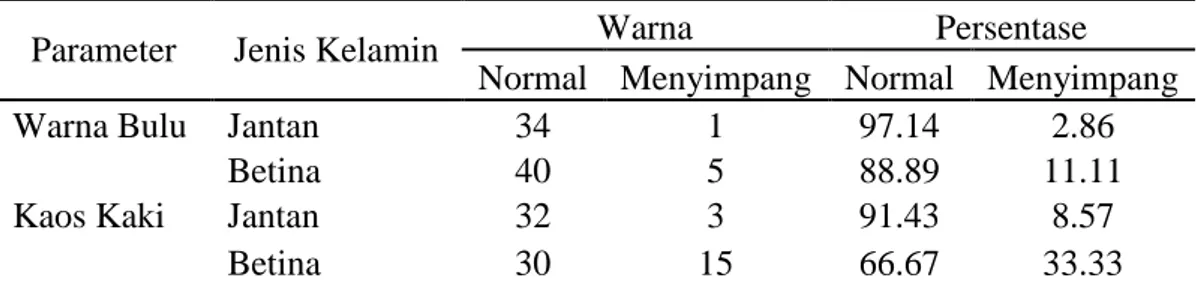
Tabel 3. Karakteristik warna Bulu dan Kaos Kaki
Parameter Jenis Kelamin Warna Persentase
Normal Menyimpang Normal Menyimpang
Warna Bulu Jantan 34 1 97.14 2.86
Betina 40 5 88.89 11.11
Kaos Kaki Jantan 32 3 91.43 8.57
Betina 30 15 66.67 33.33
Tabel 3. memperlihatkan bahwa warna bulu yang normal pada sapi jantan didominasi sekitar 97,14% sedangkan pada sapi Bali betina 88,89%. Sedangkan warna kaos kaki yang normal pada sapi Bali jantan 91,43% begitu pula dengan sapi Bali betina warna kaos kaki yang normal sekitar 66,67%. Sapi Bali mempunyai ciri-ciri khusus antara lain: warna bulu merah bata, tetapi yang jantan dewasa berubah menjadi hitam. Ada tanda-tanda khusus yang harus dipenuhi sebagai sapi bali murni, yaitu warna putih pada bagian belakang paha, pinggiran bibir atas dan pada paha kaki bawah mulai tarsus dan carpus sampai batas pinggir atas kuku, bulu pada ujung ekor hitam, bulu pada bagian dalam telinga putih, terdapat garis belut (garis hitam) yang jelas pada bagian atas punggung, (Hardjosubroto, 1994).
24 Pada penelitian menggunakan sampel darah dari kabupaten Barru ditemukan pola warna yang menyimpang dari normal, pada sapi jantan (2,86%) dan betina (11,11%). Warna menyimpang yang di maksud adalah warna bulu bertutul putih. Hal ini sesuai yang ditemukan Handiwirawan (2003) yang menemukan sapi Bali berwarna tutul putih di provinsi Bali.
Jenis warna menyimpang yang lainnya pada sapi Bali adalah pada warna kaos kaki, pada jantan (8,57%) dan betina (33,33%). Adapun warna yang dimaksud disini adalah warna kaos kaki yang tercampur dengan warna merah bata atau coklat dan bahkan tercampur dengan warna hitam dan putih. Munculnya warna yang menyimpang kemungkinan disebabkan adanya pengaruh genetik dari suatu populasi dan juga di sebabkan oleh faktor lingkungan. Handiwirawan (2003) mengungkapkan munculnya pola warna yang menyimpang kemungkinan ekspresi dari gen homosigot resesif yang terjadi karena tingkat inbreeding yang meningkat.
Sifat Kuantitatif
Sifat kuantitatif dipengaruhi oleh faktor genetik, lingkungan dan
ditemukan pengaruh interaksi keduanya (genetik dan lingkungan). Sifat kuantitatif
sapi Bali yang berada di Kabupaten Barru dapat dilihat pada Tabel 4.
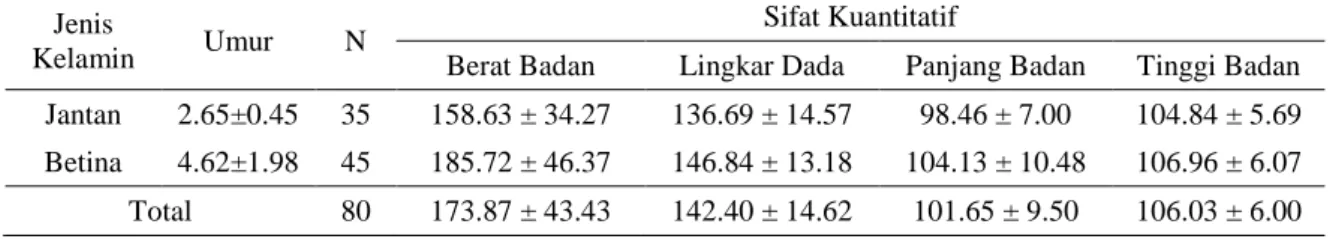
Tabel 4. Sifat Kuantitatif Sapi Bali
Jenis
Kelamin Umur N
Sifat Kuantitatif
Berat Badan Lingkar Dada Panjang Badan Tinggi Badan Jantan 2.65±0.45 35 158.63 ± 34.27 136.69 ± 14.57 98.46 ± 7.00 104.84 ± 5.69 Betina 4.62±1.98 45 185.72 ± 46.37 146.84 ± 13.18 104.13 ± 10.48 106.96 ± 6.07 Total 80 173.87 ± 43.43 142.40 ± 14.62 101.65 ± 9.50 106.03 ± 6.00
25 Berdasarkan hasil penelitian yang dilakukan, pada Tabel 4 menunjukkan bahwa rata-rata berat badan sapi jantan di Kabupaten Barru 158.63 kg, sedangkan pada sapi betina 185.72. Lingkar dada pada sapi jantan yaitu 136.69 cm, sapi betina 146.84 cm. Hasil rata-rata lingkar dan bobot badan yang diperoleh masih dibawah rata-rata lingkar dada dan bobot badan yang dihasilkan oleh Pane (1991) sebesar 158-160 cm dan 266 kg. Rata-rata pengukuran panjang badan sapi Bali jantan berkisar antara 98.46 cm, dan sapi Bali betina 104.13 cm. Tinggi badan sapi Bali jantan mempunyai rata-rata 104.84 cm, sapi Bali betina 106.96 cm. Menurut Hardjosubroto (1994) bahwa ukuran tubuh berkorelasi dan merupakan indikator bobot badan sapi seperti tinggi gumba, lingkar dada, dan panjang badan. Dengan bertambahnya bobot badan maka bertambah pula lingkar dada begitupula sebaliknya, dengan menurunnya bobot badan menurun juga lingkar dada.
Berdasarkan pengamatan yang dilakukan umur rata-rata pada sapi jantan adalah 2,65 tahun dan rata-rata umur sapi betina adalah 4,62 tahun. Umur juga mempengaruhi berat badan hal ini sesuai dengan pendapat Pane (1986) yang menyatakan bahwa pertumbuhan merupakan pertambahan bobot badan atau ukuran tubuh yang sesuai dengan umur.
Amplifikasi dan Genotip Mikrosatelit Lokus HEL9 pada Sapi Bali
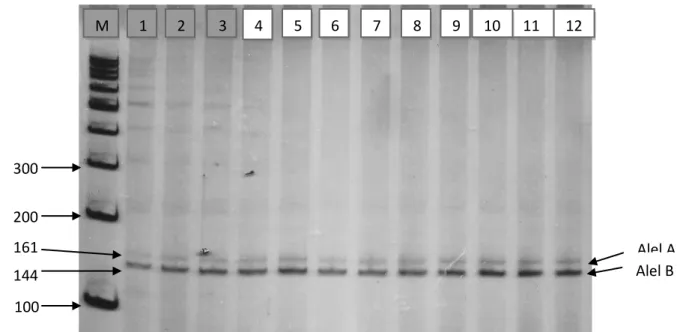
Berdasarkan hasil penelitian yang telah dilakukan bahwa Mikrosatelit Lokus HEL9 berhasil di amplifikasi pada mesin PCR SensoQuest Germany
dengan suhu annealing 55oC. Hasil amplifikasi dapat divisualisasikan pada gel
polyacrylamide 8% dapat dilihat pada Gambar 1. Panjang produk hasil amplifikasi mikrosatelit HEL9 adalah berkisar antara 144-169 pb.
26
Gambar 1. Visualisasi hasil amplifikasi Mikrosatelit Lokus HEL9 pada mesin PCR dalam gel polyacrylamide 8%. M: marker 100 pb, 1-12: sampel sapi Bali
Pada penelitian ini, alel mikrosatelit HEL9 yang berhasil teramplifikasi (ditunjukkan dengan adanya band dengan ukuran yang sesuai). Sekuen alel HEL9 berukuran panjang antara 143-165 pb (Bioshop et al., 1994).
Berdasarkan panjang fragmen hasil PCR dapat ditentukan genotip masing-masing ternak. DNA dengan panjang fragmen ±144 pb di identifikasi sebagai alel A sedangkan panjang fragmen ±161 pb di identifikasi sebagai alel B. Dengan demikian ternak dengan panjang kedua fragmen ±144 pb di identifikasi sebagai genotipe AA, ternak dengan panjang fragmen ±144 pb dan ±161 pb di identifikasi sebagai genotipe AB dan ternak dengan panjang kedua fragmen ±161 pb di identifikasi sebagai genotipe BB. Hal ini sesuai dengan penelitian Handiwirawan
et al. (2003) bahwa alel yang teramplifikasi melalui proses PCR pada lokus 300 200 100 144 161 7 M 1 2 3 4 5 6 8 9 10 11 12 Alel A Alel B
27 HEL9 pada sapi Bali memiliki ukuran panjang yaitu: alel A berukuran ±144 pb, alel B berukuran ±161 pb.
Frekuensi Genotip, Alel dan Keseimbangan Hardy- Weinberg
Hasil analisa frekuensi genotip mikrosatelit lokus HEL9 pada sapi Bali dapat dilihat pada Tabel 5. Polimorfisme dapat ditunjukkan dengan adanya dua alel atau lebih dalam satu populasi. Gen dikatakan polimorfik apabila salah satu alelnya mempunyai frekuensi kurang dari 99% (Nei dan Kumar, 2000).
Tabel 5. Frekuensi Genotip Mikrosatelit Lokus HEL9 Bangsa
Sapi Lokasi
Jumlah Individu
Total Frekuensi Genotipe
AA AB BB AA AB BB
Bali Barru 0 80 0 80 0 100 0
Berdasarkan data pada Tabel 5 dapat dilihat bahwa frekuensi genotip homozigot AA dan BB adalah 0 dan frekuensi heterozigot AB adalah 100. Hal ini terjadi karena hanya satu genotipe yang terdistribusi dan nilai ini muncul karena jumlah sampel yang diteliti sangat sedikit dan tingkat keragaman genetik sangat rendah.
Walaupun sapi Bali merupakan hasil domestikasi dari Banteng, namun hasil pengamatan menunjukkan bahwa alel genotip Banteng dan sapi Bali tidak identik. Pada populasi sapi Bali di kabupaten Barru ditemukan alel lain. Pada lokus HEL9 ditemukan alel B yang berukuran ±161 pb yang tidak ditemukan pada banteng (Handiwirawan et al., 2003). Adanya alel ini diduga karena alel-alel tersebut merupakan alel mutasi sebagai akibat proses replication shappage yang menghasilkan rangkaian yang lebih panjang (Levinson dan Gutman, 1987, Li dan
28 Graur, 1991). Kemungkinan yang lain adalah adanya aliran gen (alel) dari luar atau bangsa sapi lain yang masuk kedalam populasi sapi Bali di Kabupaten Barru.
Frekuensi alel adalah perbandingan keseluruhan fragmen gen yang terdiri dari suatu varian gen tertentu (alel). Keseimbangan Hardy- Weinberg berhubungan eret dengan frekuensi genotip dan frekuensi alel. Frekuensi alel dapat dihitung berdasarkan Nei dan Kumar (2000) dan HWE dengan Uji Chi- Square.
Tabel 6. Frekuensi Alel dan Keseimbangan Hardy- Weinberg
Bangsa Sapi Lokasi N Frekuensi Alel (%) X2 (HWE)
A B
Bali Barru 80 50 50 td
td: tidak memenuhi syarat untuk dihitung
Pada Tabel 6. menunjukkan bahwa frekuensi alel A (50%) hal ini sangat kecil dibandingkan dengan penelitian yang dilaporkan Hardiwirawan (2003) frekuensi alel A (92,9%). Hal ini berarti telah ada aliran gen dari sapi lain yang terdapat pada sapi Bali yang ada di kabupaten Barru.
Pada penelitian ini menunjukkan bahwa frekuensi alel pada lokus HEL9 pada populasi sapi Bali di Kabupaten Barru berada dalam ketidakseimbangan Hardy- Weinberg. Hal- hal yang dapat mempengaruhi keseimbangan Hardy- Weinberg menurut Hardjosubroto (1998) adalah mutasi, gene flow, migrasi, seleksi, dan tidak terjadi perkawinan secara acak.
29 Nilai Heterozigositas
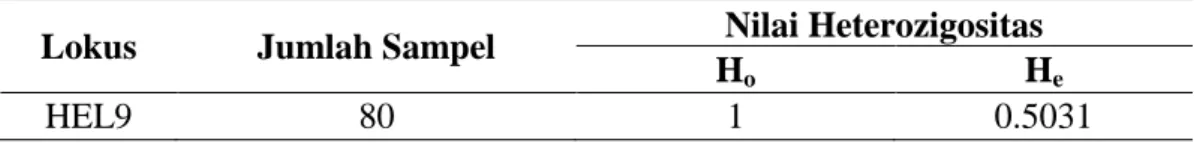
Keragaman genetik suatu populasi dapat diukur dengan nilai heterozigositas. Nilai heterozigositas pengamatan (Ho) dan heterozigositas harapan (He) dihitung berdasarkan rumus Nei dan Kumar (2000) dapat dilihat pada Tabel 7.
Tabel 7. Nilai Heterozigositas Pengamatan (Ho) dan Heterozigositas Harapan (He)
Lokus Jumlah Sampel Nilai Heterozigositas
Ho He
HEL9 80 1 0.5031
Nilai heterozigositas pengamatan (Ho) dan heterozigositas harapan (He) digunakan untuk menduga keragaman genetik. Heterozigositas harapan merupakan penduga keragaman genetik pada populasi ternak lebih tepat karena perhitungannya dilihat berdasarkan frekuensi alel. Nilai Ho adalah 1 dan He 0.5031. Nilai ini lebih tinggi didapatkan dari pada penelitian yang dilakukan oleh Abdullah, dkk (2008) yang hanya menemukan 0,1 dan Handiwirawan (2003) menemukan 0,135. Hal ini disebabkan karena ketidakseimbangan genotipe pada populasi tersebut. Menurut Tambasco et al (2003) perbedaan antara nilai heterozigositas pengamatan (Ho) dan nilai heterozigositas harapan (He) dapat dijadikan sebagai indicator adanya ketidakseimbangan genotipe pada populasi sapi yang diamati yang diindikasikan bahwa sudah ada kegiatan seleksi yang dilakukan dan tidak adanya perkawinan acak.
30 Hubungan Sifat Fenotipe dan Genotipe
Pada penelitian yang telah dilakukan menunjukkan bahwa sapi Bali, tidak diperoleh hubungan antara sifat fenotip dan genotip, karena lokus mikrosatelit HEL9 yang digunakan bukan untuk mendeteksi gen yang membawa sifat warna dan bentuk tanduk pada sapi Bali di Kabupaten Barru. Hal ini sesuai dengan pendapat Handiwirawan (2003) yang menyatakan bahwa lokus mikrosatelit HEL9 tidak berdekatan ataupun berada dalam gen penyandi untuk pola warna tubuh dan bentuk tanduk pada sapi Bali.
31 KESIMPULAN DAN SARAN
Kesimpulan
Berdasarkan penelitan secara fenotipe dan dengan menggunakan mikrosatelit lokus HEL9 sapi Bali di Kabupaten Barru maka dapat disimpulkan bahwa :
- Bentuk tanduk yang paling umum pada sapi jantan adalah Silak Bajeg sebanyak 19 ekor (54,28%) dan pada sapi betina adalah Silak Manggulgangsa sebanyak 34 ekor (75,56%).
- Warna bulu menyimpang 2,28% pada sapi jantan dan pada sapi betina11,11%. Warna kaos kaki yang menyimpang pada sapi Bali jantan 8,57% dan sapi Bali betina warna kaos kaki yang menyimpang sekitar 33,33%. Warna menyimpang pada kaos kaki seperti: terdapat warna hitam dan warna coklat pada kaos kaki sapi tersebut.
- Proporsi alel A 50% dan alel B 50% pada sapi Bali di Kabupaten Barru. - Persentase genotipe AB pada sapi Bali di Kabupaten Barru adalah 100%. Saran
Berdasarkan kesimpulan penelitian ini maka perlu dilakukan identifikasi kemurnian sapi Bali dengan metode lain, seperti: keragaman protein darah dan struktur bulu untuk dikombinasikan dengan yang dilakukan pada penelitian ini.
32 DAFTAR PUSTAKA
Abdullah. M. A. N, R.R. Noor, H. Martojo, dan D.D. Solihin. 2008. Karakterisasi
genetik sapi aceh dengan menggunakan dna mikrosatelit. J.Indon.Trop.Anim.Agric. 33
Anonim. 2000. Uji kemurnian sapi Bali melalui protein, DNA mikrosatelit, struktur bulu dan kromosom. Laporan Penelitian. Bogor: Fakultas Peternakan Institut Pertanian Bogor dan Balai Inseminasi Buatan Singosari.
Bawden, W.S. and K.R, Nicholas. 1999. Molecular genetics of milk production. In The Genetics of Cattle. Edited R. Fries and A. Ruvinsky. CABI Publishing. London. UK. 539-576.
Bennett, G.J. 2000. Update on the neurophysiology of pain transmission and modulation: Focus on The NMDA-Receptor. J Pain Symptom Manage 19: 2–6.
Bishop, M.D., S.M. Kappes, J.W. Keele, R.T. Stone, I.F. Sunden, G.A. Hawkin, S.S. Toldo, R. Fries, M.D. Grosz, J. Voo and C.W. Beattie. 1994. A genetic linkage map for cattle. Genetics. 136: 619-639.
Bradley, D.G. and E.P. Cunningham. 1999. Genetics Aspects of Domestication. In: Fries, R. And Ruvinsky, A., editor. The genetics of Cattle. New York : CABI Publishing. Hlm 15 – 31.
Djarsanto. 1997. Kebijaksanaan Pelestarian Ternak Asli Indonesia dalam Rangka Mendukung Pengembangan Perbibitan Ternak Nasional. Prosiding Seminar Nasional Peternakan dan Veteriner. Bogor, 7-8 Januari 1997. Pusat Penelitian dan Pengembangan Peternakan. Hlm. 182-185.
Handiwirawan, E. 2003. Penggunaan Mikrosatelit HEL9 dan INRA035 sebagai Penciri Khas Sapi Bali. Tesis. Program Pasca Sarjana, Institut Pertanian Bogor.
Handiwirawan, E., R. R. Noor, Muladno and L. Schuler. 2003. The use of HEL9 and INRA035 mikrosatelites as specific markers for Bali cattle. Arch. Tierz., Dummerstorf 46, 503-512.
Handiwirawan, E. dan Subandriyo. 2004. Potensi dan keragaman sumberdaya genetik sapi bali. Lokakarya Nasional Sapi Potong. Pusat Penelitian dan Pengembangan Peternakan. Hlm. 50-60.
33 Hardjosubroto, W. dan J.M. Astuti. 1993. Buku Pintar Peternakan. Jakarta: PT
GramediaWidiasarana Indonesia.
Hardjosubroto, W. 1994. Aplikasi Pemuliabiakan Ternak di Lapangan. Jakarta: PT GramediaWidiasarana Indonesia.
Hardjosubroto, W. 1998. Pengantar Genetika Hewan. Fakultas Peternakan Universitas`Gadjah Mada, Yogyakarta.
Hartl, D. L., 1988 Primer of Population Genetics. 2 ed. Sinauer Associates, Sunderland, Massachusetts. 305 pp.
Herweijer, C.H. 1950. Enkele aantekenigen btreffende de geschiedenis van de runderveeteelt op het Eiland Timor. Hemera Zoa 56: 689.
Leung H., R.J. Nelson & J.E. Leach. 1993. Population structure of plant pathogenic fungi and bacteria. Adv. Plant Pathol. 10: 157 – 205
Levinson, G. and G. A. Gutman. 1987. Slipped-strand mispairing: a major mechanism for DNA sequence evolution. Mol. Biol. Evol. 4: 203-221 Li, W. H. and D. Graur. 1991. Fundamentals of Molecular Evolution Sunderland,
Massachusetts: Sinauer Associates, Inc.
Maskur, Muladno B. Tappa. 2007. Identifikasi genetik menggunakan marker mikrosatelit dan hubungannya dengan sifat kuantitatif pada sapi. Media Peternakan. 30:147-155.
Masudana, I W. 1990. Perkembangan Sapi Bali di Bali dalam Sepuluh Tahun Terakhir (1980- 1990). Proceeding Seminar Nasional Sapi Bali. Denpasar, 20-22 September 1990. Denpasar: Fakultas Peternakan Universitas Udayana. Hlm A-11-A-30.
Meijer, W. C. P. 1962. Das Balirind. A. Ziemsen Verslag, Wittenberg Lutherstandt.
Moeljopawiro, S. 2010. Marka mikrosatelit sebagai alternatif uji buss dalam perlindungan varietas tanaman padi. Buletin Plasmanutfah 16(1): 1-7 Moore, S.S., L.L. Sargeant, J.J. King, J.S. Mattick, M. Georges, and D.J.S Hetzel.
1991. The conservation of dinucleotide microsatellites among mammalian genomes allow the of heterilogous PCR primer pairs in closely related species. Genomics. 10:654-660.
Namikawa, T., T. Amano, B. Pangestu and S. Natasasmita. 1982. Electrophoretic variations of blood proteins and enzymes in indonesian cattle and
34 bantengs. the origin and phylogeny of indonesian native livestock (Part III): Morphological and Genetical Investigations on the Interrelationship between Domestic Animals and their Wild Forms in Indonesia. The Research Group of Overseas Scientific Survey. Hlm 35–42.
National Research Council. 1983. Little-Known Asian Animals with a Promising Economic Future. Washington, D.C.: National AcademicPress.
Nei, M. and S. Kumar. 2000. Molecular Evolution and Phylogenetics. New York: Oxford University Press.
Nozawa, K. 1979. Phylogenetic Studies on The Native Domestic Animals in East and Southeast Asia. Proceeding Workshop Animal Genetic Resources in Asia and Oceania. Tsukuba, 3-7 September 1979. Tsukuba: Society for the Advancement of Breeding Researches in Asia and Oceania (SABRAO). Hlm 23-43.
Oka, I.G.L. and D. Darmadja. 1996. History and development of Bali Cattle. Proceedings seminar on Bali cattle, a special spesies for the dry tropics, held by Indonesia Australia Eastern University Project (IAEUP), 21 September 1996. Udayana University Lodge, Bukit Jimbaran, Bali.
Pane, I. 1986. Pemuliabiakan Ternak Sapi. PT. Gramedia, Jakarta.
Pane, I. 1990. Upaya Peningkatan Mutu Genetik Sapi Bali di P3 Bali. Prosiding Seminar Nasional Sapi Bali.Bali, 20-22 September 1990.
Pane, I. 1991. Produktivitas dan Breeding Sapi Bali. Pros. Seminar Nasional Sapi Bali. 2-3 September 1991. Fakultas Peternakan Universitas Hasanudin. Ujung Pandang.
Payne, W.J.A. and D.H.L. Rollinson. 1973. Bali cattle. World Anim. Rev. 7: 13-21.
Payne, W.J.A. and J. Hodges. 1997. Tropical Cattle: Origin, Breeds and Breeding Policies. Blackwell Science.
Powell, W., G.C. Machray, and J. Provan. 1996. Polymorphism revealed by simple sequence repeats. Trends in Plant Science, Vol.1, No.7, (July 1996), pp. 215-222, ISSN 1360-1385.
Rollinson, D.H.L. 1984. Bali Cattle. In: Evolution of Domesticated Animals. Mason, I.L. (Ed.). New York: Longman.
35 Sastradipradja, D. 1990. Potensi internal sapi Bali sebagai salah satu sumber plasma nutfah untuk menunjang pembangunan peternakan sapi potong dan ternak kerja secara nasional. Pros. Seminar Nasional Sapi Bali. Denpasar, 20-22 September. Denpasar: Fakultas Peternakan Universitas Udayana. Hlm A-47–A54.
Silva, F., L. Gusmao, and A. Amorim. 1999. Segregation analysis of tetra and pentanucleotide short tandem repeat polymorphism : Deviation from Mendelian expectations. Electrophoresis, 20:1697-1701.
Soehadji. 1990. Kebijaksanaan Pemuliaan Ternak (Breeding Policy) Khususnya Sapi Bali, Dalam Pembangunan Peternakan. Pros. Seminar Nasional Sapi Bali. Denpasar, 20-22 September 1990. Denpasar: Fakultas Peternakan Universitas Udayana. Hlm A-1-A- 9.
Steffen, P., A. Eggen, A.B. Dietz, J.E. Womack, G. Stranzinger, and R. Fries. 1993.
Isolation and Mapping of Polymorphic Microsatetlites In Cattle. Anim.
Genet. 24, 121-124.
Sukmasari, A. H. 2003. Pendugaan Nilai Pemuliaan dan Kecenderungan Genetik (Genetic Trends) Bobot Badan Sapi Bali di Proyek Pembibitan dan Pengembangan Sapi Bali (P3 Bali) di Bali. Tesis. Program Pascasarjana, Institut Pertanian Bogor.
Tambasco, D. D., C. C. P. Paz, M. Tambasco-Studart, A. P. Pereira, M. M. Alencar, A. R. Freitas, L. L. Countinho, I. U. Packer and L. C. A. Regitano. 2003. Candidate genes for growth traits in beef cattle Bos Taurus x Bos Indicus. J. Anim. Bred. Genet. 120: 51-60.
Tegelstrom, H. 1992. Mitochondrial DNA in natural population: An improved routine for screening of genetic variation baed on sensitive silver staining. Electrophoresis. 7:226-229.
Winaya, A. 2000. Penggunaan penanda molekuler mikrosatelit untuk deteksi polimorfisme dan analisis filogenetik genom sapi [tesis]. Bogor: Institut Pertanian Bogor. Program Pascasarjana. Program Studi Bioteknologi.
36 Lampiran 1. Gambar Bentuk Tanduk Sapi Bali di Kabupaten Barru
( I ) ( II )
( III ) ( IV )
( V ) ( VI )
Gambar: (I) Silak Anoa ; (II) Silak Manggulgangsa ; (III) Silak Cono ; (IV) Silak Bajeg ; (V) Silak Pendang ; (VI) Silak Congklok.
37 Lampiran 2. Pola Warna Bulu Sapi Bali di Kabupaten Barru
(I) (II)
(III) (IV)
(V)
Gambar : Pola warna tubuh sapi Bali jantan yang normal (I) ; pola warna tubuh sapi Bali betina yang normal (II) ; sapi Bali bintik – bintik putih / sapi tutul (III) ; dan beberapa pola warna tubuh sapi Bali yang menyimpang dari normal, yaitu sapi Bali dengan campuran warna cokelat, putih dan hitam pada bagian kaki bawah (IV) dan (V).
38
Lampiran 3. Analisis Data dengan Menggunakan Software PopGene32 Versi 1.31
****************************************************** POPULATION GENETIC ANALYSIS
****************************************************** Date : 2014/4/28
Time : 11:33:19
Data Description : Test Data Set II: Diploid Data
************************************************************************ Single-Population Descriptive Statistics
************************************************************************ population ID : 1
population name : none
* Population : 1 @ Locus : HEL9*
============================================================ Genotypes Obs. (O) Exp. (E) (O-E)²/E 2*O*Ln(O/E)
============================================================ (A, A) 0 19.8742 19.8742 0.0000
(B, A) 80 40.2516 39.2516 109.9004 (B, B) 0 19.8742 19.8742 0.0000
============================================================ Chi-square test for Hardy-Weinberg equilibrium :
Chi-square : 79.000000
Degree of freedom : 1
Probability : 0.000000
Likelihood ratio test for Hardy-Weinberg equilibrium :
39 Degree of freedom : 1
Probability : 0.000000
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~ Allele Frequency of population 1 :
============================== Allele \ Locus Hel9
============================== Allele A 0.5000
Allele B 0.5000
============================== Summary Statistics of population 1 :
************************************************************************ Summary of Genic Variation Statistics for All Loci
[See Nei (1987) Molecular Evolutionary Genetics (p. 176-187)]
************************************************************************ ==============================
Locus Sample Size na* ============================== Hel9 160 2.0000 Mean 160 2.0000 St. Dev 0.0000 ============================== * na = Observed number of alleles
40 ************************************************************************
Summary of Heterozygosity Statistics for All Loci
************************************************************************ ===============================================================
Locus Sample Size Obs_Hom Obs_Het Exp_Hom* Exp_Het* Nei** Ave_Het ====================================================================== Hel9 160 0.0000 1.0000 0.4969 0.5031 0.5000 0.5000 Mean 160 0.0000 1.0000 0.4969 0.5031 0.5000 0.5000 St. Dev 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000
=============================================================== * Expected homozygosty and heterozygosity were computed using Levene (1949) ** Nei's (1973) expected heterozygosity
The number of polymorphic loci is : 1
41
Lampiran 4. Dokumentasi Penelitian
EKSTRAKSI DNA
42 PROSES PCR