INDUKSI AUXIN TERHADAP AKTIVITAS AUTOTROFIK BIBIT
ANGGREK BOTOL PADA LINGKUNGAN EX-VITRO
I Gede Ketut Adiputra
Program Studi Biologi, FMIPA, Universitas Hindu Indonesia, Jl. Sangalangit, Tembau, Penatih, Denpasar.
ABSTRAK
Auxin adalah zat pengatur tumbuh yang ditemukan pertama kali dan memegang peran kunci dalam pertumbuhan dan perkembangan tanaman. Dalam kultur jaringan, pemberian senyawa ini telah diketahui dapat meningkatkan perkembangan eksplant menjadi tanaman baru yang utuh. Auxin ditambahkan secara eksogen kedalam media yang mengandung sukrosa, mikro dan makro nutrien, vitamin dan mioinositol. Bagi explant, senyawa organik digunakan sebagai substrat untuk biosintesis molekul pertumbuhan. Akan tetapi, pada tanaman yang utuh, senyawa organik tersebut adalah produk dari aktivitas autotrofiknya. Oleh karena itu pertumbuhan explant menjadi callus dan akhirnya menjadi plantlet dapat dikatakan merupakan pertumbuhan heterotrofik, terutama pada fase awal morfogenesis. Dalam teknik kultur jaringan, sukrosa dan auxin tetap diberikan selama tanaman berada dalam botol kultur sehingga aktivitas heterotrofik dapat tetap berlangsung sampai tanaman ditransplantasi ke lingkungan ex-vitro. Kegiatan heterotrofik berkepanjangan tersebut dapat menjadi penyebab rendahnya aktivitas autotrofik plantlet setelah transplantasi ke lingkungan ex-vitro. Untuk mengembalikan pertumbuhan autotrofik, beberapa perlakuan perlu diberikan agar tanaman cukup kuat menghadapi lingkungan baru ex-vitro. Paper ini membahas beberapa perlakuan terutama kemungkinan perlunya pemberian auxin setelah plantlet dipindahkan ke lingkungan baru ex-vitro.
Kata kunci: Auxin, autotrofik, anggrek, ex-vitro.
ABSTRACTS
Auxin is firstly invented growth regulator and plays a central role during plant growth and development. Addition of this growth regulator in cell and tissues culture has been known to enhance cell development. In this culture, the auxin is added exogenously into media containing sucrose, micro and macro nutrient, vitamin and myo-inositol. For the growth of explants, organic compounds added into the growth medium are used as nutrient. So, it is synthesized to produce macromolecules required during morphogenesis. In theory, an autotrophic organism synthesizes organic compounds from inorganic nutrient taken up via the root or leaves system. So, development of explants into callus and eventually plantlet is therefore can be viewed as heterotrophic activity, since it takes up sucrose as nutrient for growth rather than inorganic. In tissues or cell culture, media continuously contain organic compound, even after plantlet has produced chlorophyll. This media enable the plantlet to maintain it heterotrophic activity. However after transplantation, the prolonged heterotrophic activity can make the plants to have a low autotrophic activity. So, in order to resume autotrophic activity and enhanced viability, some treatments are required. This paper discuss some of the treatments particularly the possibility of auxin addition after transplantation of plantlet into a new ex-vitro environment.
PENDAHULUAN
Tumbuhan yang bernilai ekonomi tinggi tetapi sulit berkembang biak secara alami biasanya dikembangkan dengan teknik kultur jaringan. Anggrek adalah salah satu jenis tanaman yang sulit berkembang biak secara alami tetapi memiliki nilai ekonomi tinggi. Pengembangan tanaman ini menjadi bibit melalui tehnik kultur jaringan telah banyak berhasil, tetapi menumbuhkan bibit botol pada lingkungan ex-vitro menjadi tanaman dewasa masih menemukan banyak permasalahan. Bibit yang dikembangkan dengan teknik kultur jaringan ini biasanya memiliki kemampuan autotrofik yang rendah (Daisy dan Wijayani, 1994). Rendahnya kemampuan autotrofik menyebabkan tanaman tidak memiliki bahan organik yang cukup untuk pertumbuhan karena laju sintesis senyawa ini dari senyawa anorganik sangat rendah.
Disamping kelemahan autotrofik, kondisi khusus yang digunakan untuk mengembangkan bibit dengan teknik kultur jaringan ini dapat menghasilkan tanaman yang abnormal, baik morfologi, anatomi maupun fisiologi. Kondisi-kondisi khusus tersebut antara lain, penggunaan botol yang tertutup rapat, pemberian zat pengatur tumbuh dan pemberian karbohidrat (Mineo, 1990, Deasy dan Wijayani, 1994). Menurut Pospisilova (1999), tanaman yang tumbuh dalam botol yang tertutup rapat mengakibatkan daun tidak memiliki lapisan pelindung yang cukup tebal. Apabila tanaman ini kemudian dipindahkan ke lingkungan ex-vitro maka penguapan yang berlebih tidak dapat ditahan. Selanjutnya dikatakan bahwa pemberian hormon untuk menumbuhkan bibit secara invitro dapat menghasilkan tanaman yang abnormal. Tanaman ini akan mudah rusak pada lingkungan ex-vitro. Disamping pemberian hormon, pemberian karbohidrat pada bibit botol juga dapat menghasilkan tanaman yang mudah rusak karena serangan bakteri. Kelemahan-kelemahan tanaman yang dikembangkan dengan teknik
penelitian lanjutan perlu dilakukan agar hasil pengembangan bibit secara invitro dapat ditumbuhkan menjadi tanaman dewasa yang produktif. Hal ini terutama karena regenerasi sebuah sel atau jaringan tanaman menjadi tanaman utuh yang baru hampir tidak mungkin dilakukan tanpa pemberian kondisi-kondisi khusus.
pemberian perlakuan khusus pada lingkungan ex-vitro agar bibit tersebut dapat tumbuh menjadi tanaman yang produktif
PEMBAHASAN
Peran auxin dalam pertumbuhan dan perkembangan tanaman
Auxin adalah hormon tumbuhan yang ditemukan pertama kali. Senyawa ini memegang peran kunci dalam pertumbuhan, perkembangan dan respon tumbuhan terhadap perubahan lingkungan (Tromas & Perrot-Rechenmann, 2010). Menurut Vieten at al. (2007), pada
proses morfogenesis, auxin melakukan pengaturan baik pada fase embryogenesis, organogenesis, differensiasi jaringan pengangkutan, pemeliharaan meristem akar maupun pertumbuhan trofik. Selanjutnya dikatakan bahwa peran yang dimiliki oleh auxin ini dijalankan dengan cara memicu perubahan program pembangunan melalui pemberian informasi vektorial pada jaringan. Secara lebih rinci, mekanisme yang dilakukan oleh auxin untuk mengubah program pembangunan diuraikan oleh Robert & Friml (2009). Menurut peneliti ini, auxin diproduksi pada bagian pucuk tanaman yaitu pada daun muda dan kuncup bunga. Senyawa ini selanjutnya ditransportasikan ke akar atau bagian lain tanaman melalui beberapa jalur translokasi, terutama sistem pembuluh dan jalur transport interselluler. Pada sel tujuan, akumulasi auxin differential kemudian terjadi dan diterima serta diinterpretasi oleh inti yang mengatur ekspresi gen dan perencanaan kembali nasib sel (Robert & Friml, 2009). Kemampuan auksin untuk melakukan perubahan ekspresi gen mengakibatkan senyawa ini menjadi sangat penting dalam upaya mengaktifkan kembali sel yang telah menghentikan proses differensiasi.
Dalam teknik kultur jaringan, sel tanaman yang dikembangkan menjadi tanaman baru umumnya telah menghentikan proses differensiasi. Pada kondisi media yang sesuai, sel eksplant berkembang melalui beberapa tahap. Pada tahap
awal, sel eksplant tumbuh menjadi sel kalus setelah diinduksi oleh hormon pertumbuhan terutama auxin (Siwach et al. 2011). Kumpulan
sel ini kemudian berkembang menjadi plantlet setelah pembentukan plb (protocorm like body).
Plb ini memiliki tingkat perkembangan seperti embrio, sehingga disebut juga embrio somatik (Rianawati et al. 2009). Berbeda dengan embrio
pada biji yang terbentuk melalui fusi sel telur dan sperma dan dirancang untuk berkembang menjadi embrio, embrio somatik ini berkembang dari sel somatik yang memperoleh kompetensi untuk dapat merespon signal embriogenik dan memulai pembangunan embrio (Pasternak et al.
2002). Pada teknik kultur jaringan, pertumbuhan embrio somatik diinduksi dengan penambahan auxin sintetik NAA (Utami et al. 2007). Dengan
pemberian zat pengatur tumbuh BAP (sitokinin) dan 2,4-D (auxin), plb ini kemudian dapat tumbuh menjadi plantlet (Rianawati et al. 2009).
Morphogenesis dalam kultur jaringan ini disebut juga dengan proses 3 langkah (Komal, 2011).
Walaupun individu baru yang terbentuk (plantlet) telah memiliki organ yang lengkap, tetapi tanaman kecil ini masih memiliki aktivitas autotropik yang rendah (Daisy & Ari Wijayani, 1994) dan jaringan pengangkutan lemah (Robinson et al. 2009). Kelemahan sistem yang
terjadi pada tanaman ini merupakan akibat samping dari kondisi yang digunakan untuk menumbuhkan tanaman secara invitro. Kondisi khusus ini sesungguhnya tidak sesuai dengan sistem yang autotrof yang mengubah senyawa anorganik menjadi senyawa organik. Secara skematis, sumber nutrisi untuk pertumbuhan embrio pada biji dan pertumbuhan sel somatik pada kultur jaringan digambarkan pada Gambar 1. Pada gambar ini nampak bahwa pertumbuhan invitro menggunakan sumber organik yang tidak dibuat sendiri (exogenous), sedangkan tanaman
yang dikembangkan dari biji menggunakan senyawa organik yang dibuat sendiri (endogenous) untuk pertumbuhan. Sumber
mekanisme keseimbangan substrat dan produk. Apabila pemberian substrat terlalu banyak maka tanaman utuh yang dihasilkan akan memiliki variasi dari tanaman normal baik anatomi, morfologi maupun fisiologi. Oleh karena itu, bibit tanaman hasil kultur jaringan masih memerlukan penyempurnaan, terutama setelah transplantasi ke lingkungan ex-vitro. Jadi, sebelum tanaman dapat dibudidayakan dalam lingkungan alami ex-vitro, hasil kultur jaringan masih memerlukan perlakuan yang memungkinkan proses fisiologis berjalan normal dalam kondisi yang sangat berbeda dengan lingkungan invitro.
Fisiologi dan morfologi bibit hasil kultur jaringan
Pada fase awal pertumbuhan ex-vitro, tanaman hasil kultur jaringan nampaknya masih bersifat heterotrofit, yaitu lebih menyukai senyawa organik exogenous dari pada memproduksi
Senyawa organik tersebut antara lain; vitamin, hormon, mioinositol dan sukrosa (Daisy & Ari Wijayani 1994). Pemberian senyawa organik eksogenous ini kemungkinan ikut berperan sebagai sinyal untuk mengarahkan kegiatan morfogenesis dan mengakibatkan terjadinya penurunan kemampuan daerah meristematik untuk menumbuhkan jaringan yang berfungsi untuk mengimpor hasil fotosintesis dari perangkat fotosintesis seperti kloroplas. Penurunan pertumbuhan jaringan ini selanjutnya mempengaruhi aktivitas enzim yang terdapat dalam perangkat fotosintesis. Secara teori, apabila produk suatu enzim melebihi titik kesetimbangan, maka aktivitas enzim tersebut akan mengalami hambatan umpan-balik. Mekanisme hambatan inilah yang kemungkinan terjadi pada tanaman ketika masih berada di dalam botol kultur. Oleh karena itu, setelah ditransplantasi ke lingkungan yang alami di luar botol, bibit ini memiliki kemampuan yang sangat rendah untuk menghasilkan senyawa organik secara autotrof.
Masalahnya adalah bagaimana menginduksi tanaman ini agar perangkat fotosintesis dan jaringan pengangkutan cukup banyak tersedia agar biosintesis autotrofik menjadi meningkat. Mengingat pengaturan pertumbuhan ditentukan oleh auxin, yang memiliki peran kunci dalam pertumbuhan dan perkembangan terutama akibat perubahan lingkungan (Tromas & Perrot-Rechenmann, 2010), maka perbaikan aktivitas autotrofik bibit anggrek botol ini nampaknya masih memerlukan auxin eksogenous untuk menyempurnakan pertumbuhan autotrofiknya. Namun demikian, pemberian auxin juga bukan tanpa masalah. Senyawa ini diperlukan dalam jumlah yang sangat rendah dan harus memiliki proporsi yang sesuai dengan senyawa lain agar pertumbuhan autotrofik dapat berlangsung (Albert at al. 1983).
Perbanyakan tanaman anggrek
Tanaman angrek dapat dikembangkan baik secara tradisional (ex-vitro) maupun secara
modern (in-vitro). Pengembangan bibit secara tradisional umumnya tidak menggunakan senyawa organik eksogenous sebagai sumber nutrisi. Sebaliknya, penyediaan nutrisi baik organik, anorganik bahkan hormon pertumbuhan sangat penting jika ingin memperbanyak tanaman menggunakan teknik modern kultur jaringan (Daisy & Wijayani, 1994; Rianawati et al. 2009).
Untuk perbanyakan tanaman anggrek dengan teknik kultur jaringan, bahan tanaman dapat berupa biji atau berupa bagian vegetatif tanaman. Biji dan bagian vegetatif sebagai bahan tanaman yang dikembangkan dengan cara modern ini memiliki perbedaan jalur morfogenesis. Biji telah memiliki embrio sedangkan bagian vegetatif tanaman tidak memiliki embrio. Oleh karena itu, jika bahan tanaman diambil dari bagian sel somatik maka pekerjaan yang diperlukan adalah mulai dari menumbuhkan kalus, embrio somatik dan akhirnya menumbuhkan plantlet (Gambar 1). Walaupun dapat menghasilkan banyak tanaman baru, setiap sel dapat menjadi tanaman baru, tetapi persyaratan teknis yang harus dipenuhi sangat tinggi. Keberhasilan teknik modern ini tergantung tidak hanya dari alat dan bahan laboratorium, tetapi juga ketrampilan teknisi yang menangani.
Biji adalah alat reproduksi, memiliki embrio yang dihasilkan dari fusi sel telur dan spema. Oleh karena sel gamet, telur dan sperma, ini berasal dari dua organisme yang berbeda, maka tanaman yang dihasilkan memiliki sifat yang merupakan gabungan dari sifat induknya. Pada teknik kultur jaringan dikatakan bahwa tanaman yang dihasilkan dari biji ini biasanya tidak seragam (Rianawati et al. 2009). Untuk mendapatkan
auxin, senyawa organik, anorganik dan kondisi yang aseptik.
Kemampuan awal yang harus dimiliki sel somatik ekplant untuk dapat tumbuh menjadi kalus adalah kemampuan menggunakan nutrient yang disediakan dalam media kultur. Untuk tujuan ini, sel somatik tersebut kemungkinan mengarahkan differensiasi untuk pengembangan jaringan yang berfungsi untuk menyerap sumber nutrisi organik eksogenous. Tanpa kemampuan ini, sel tidak dapat tumbuh karena sumber karbon hanya berasal dari lingkungan yaitu media kultur. Akan tetapi, setelah terbentuk kloroplas, yaitu pada fase plb (Utami et al. 2007, Rianawati et al. 2009), morfogenesis mungkin masih tetap
diarahkan untuk pengembangan jaringan yang memiliki fungsi sama, disamping karena jumlah kloroplast masih sedikit, senyawa organik eksogenous masih tersedia dan tidak ada lapisan yang menutup import senyawa organik eksogenous. Hal ini berbeda dengan biji (dikotil) yang dikembangkan secara ex-vitro, embrio berkembang dalam 2 kutub secara berimbang. Senyawa organik yang tersedia secara endogenous disiapkan untuk dapat mengembangkan pertumbuhannya ke kedua kutub tersebut dan ketika persediaan ini habis, tanaman telah memiliki jaringan yang berfungsi untuk menyerap senyawa anorganik dan jaringan yang berfungsi untuk menyalurkan hasil fotosintesis. Sebaliknya, pada tanaman yang dikembangkan secara invitro, kutub pertumbuhan dapat sangat bervariasi (Rao dan Narayanaswami 2006). Menurut peneliti ini, potensi pertumbuhan bervariasi menurut nutrisi yang diberikan, dapat membentuk akar, daun, embrio bipolar dan plantlet. Pada tanaman yang dikembangkan dengan medium MR yang diberi BAP 0.4 mg/l dan 2.4-D 0.2mg/l, sel-sel dapat diinduksi untuk menumbuhkan kotiledon, primordia tunas dan akar (Rianawati et al. 2009).
Induksi tunas juga terjadi pada tanaman yang dikembangkan dengan media NP ditambah 2 mg/
purpurea menemukan bahwa perkecambahan
biji tidak tergantung pada nutrient, tetapi perkembangan lanjutan hanya dapat terjadi pada media Vacint-Went (Dutra et al 2008). Variasi
pertumbuhan tanaman invitro ini kemudian menyebabkan bibit botol memiliki variasi kesiapan jaringan pengangkutan. Hal ini tidak terjadi pada pengembangan bibit secara ex-vitro karena sumber nutrisi organik telah disiapkan pada kotiledon dan redistribusinya diatur secara terprogram dalam DNA. Program ini sangat spesifik menurut spesies dan calon jaringan yang berfungsi untuk menyalurkan nutrisi organik telah terbentuk bahkan ketika masih dalam biji.
Secara teori, jika persediaan makanan cadangan telah habis, tanaman telah memiliki akar yang cukup untuk memperoleh nutrient anorganik. Sebaliknya, perkembangan jaringan pengangkutan pada tanaman yang dikembangkan dengan kultur jaringan, nampaknya jauh lebih lambat dan sangat tergantung pada komposisi nutrient dan posisi explant terhadap nutrient. Mekanisme adaptasi struktur dengan demikian sangat diperlukan oleh bibit tanaman invitro karena pembentukan daun dan akar tidak diimbangi oleh peningkatan jaringan pengangkutan sehingga pengambilan nutrient anorganik untuk disintesa menjadi senyawa organik endogenous menjadi sangat sulit. Kesulitan adaptasi juga dapat terjadi karena pemanfaatan nutrient organik eksogenous tetap berlangsung walaupun akar dan daun telah terbentuk (Gambar 2). Akibatnya adalah akar menjadi tidak memiliki sink strength yang cukup tinggi bagi produks fotosintesis yang dibuat didaun. Produk fotosintesis kemudian tidak dapat mengalami floem loading dan menjadi penghambat umpan-balik bagi enzim fotosintesis. Kesulitan lain adalah bahwa tanaman yang dikembangkan secara invitro biasanya memiliki daun yang lemah secara anatomi (Robinson et al. 2009). Menurut peneliti ini, daun dari tanaman
jaringan floem ini disebabkan oleh lemahnya induksi jalur pertumbuhan jaringan floem dari daun karena sukrosa didapat sebagian besar dari nutrient secara eksogenous (Gambar 2).
Lambatnya pengembangan jaringan floem ini dapat menjadi penyebab lemahnya aktivitas autotrofik pada bibit anggrek botol. Struktur floem yang sedikit tidak memungkinkan terjadinya pengangkutan yang banyak sehingga organ fotosintesis tidak memproduksi hasil fotosintesis dalam jumlah yang besar. Produk fotosintesis yang rendah tidak cukup kuat untuk membantu induksi pertumbuhan floem yang ekstensif. Walaupun faktor yang menjadi penyebab atau menjadi akibat tidak jelas, upaya perbaikan aktivitas autotropik tanaman invitro setelah berada pada lingkungan ex-vitro dapat diupayakan melalui peningkatan aktivitas enzim fotosintesis dan peningkatan pertumbuhan jaringan pengangkutan.
Hubungan Kerja Auxin, Sukrosa dan Unsur Hara Mineral Pada Pertumbuhan Tanaman Tumbuhan utuh adalah organisme autotrof, bekerja sebagai pengubah senyawa anorganik menjadi senyawa organik. Berbeda dengan eksplant yang harus ditumbuhkan dengan pemberian berbagai senyawa organik, pertumbuhan tanaman utuh hanya memerlukan unsur hara anorganik. Pada tanaman utuh ini sumber energi berasal dari matahari, bukan dari nutrient yang diserap dari lingkungan. Unsur hara anorganik ini, akan diserap melalui akar dan
disintesa menjadi senyawa organik sesuai dengan kebutuhan pertumbuhan. Masalah penting pada pemberian senyawa anorganik ini adalah bahwa akumulasi mineral pada jaringan dapat menyebabkan hambatan fisiologis dan pada tingkat tertentu dapat mengakibatkan keracunan. Sebaliknya, apabila unsur hara ini tidak tersedia dalam jumlah yang cukup maka senyawa organik yang dibutuhkan tidak dapat disintesa.
Unsur hara mineral telah lama dikenal mampu mempengaruhi aktivitas autotrofik yaitu menyusun senyawa organik dari senyawa anorganik (Gardner, Pearce dan Mitchell 1991). Penelitian yang dilakukan pada beberapa tanaman (yang dikembangkan secara ex-vitro) menunjukkan bahwa unsur hara sangat menentukan laju fotosinthesis maupun pertumbuhan. Misalnya, defisiensi unsur hara fosfor pada tanaman menyebabkan terjadinya penurunan aktivitas fotosintesis (Sawada et al.
1982, Terry dan Ulrich 1973). Kekurangan unsur hara N mengakibatkan terjadinya penurunan assimilasi carbon (Gastal dan Lemaire 2002). Pada tanaman tomat, Kanai et al. (2007)
tanaman utuh yang dikembangkan secara kultur jaringan dan memiliki variasi perkembangan jaringan pengangkutan, memberi respon yang sama terhadap unsur hara anorganik yang diberikan.
Berbeda dengan tanaman yang dikembangkan secara ex-vitro, bibit tanaman yang dikembangkan secara invitro ditemukan memiliki jaringan floem yang sedikit (Robinson
et al 2009) dan aktivitas autotrofik yang rendah
(Daisy dan Wijayani 1994). Kedua kelemahan yaitu struktur dan fisiologi ini kemungkinan memiliki hubungan sebab-akibat. Pada kondisi invitro, translokasi produk fotosintesis dari daun tidak banyak terjadi karena sumber karbon tersedia secara eksogenous. Sedikitnya ekspor hasil fotosintesis dari daun ini mengakibatkan pertumbuhan jaringan pengangkutan dari daun ke bagian tanaman lainnya tidak perlu memiliki daya angkut yang tinggi. Akibatnya adalah hasil fotosintesis yang diproduksi di daun sulit didistribusikan dan dapat menjadi penghambat umpan-balik bagi enzim fotosintesis. Hambatan, yang berlangsung lama dan menjadi represor bagi gen yang mengkode penghasilan enzim fotosintesis, akan mengubah prilaku morfogenesis. Perubahan prilaku ini terutama terjadi pada sistem penyerapan unsur hara. Misalnya akar pada tanaman invitro yang seharusnya berfungsi untuk menyerap nutrient anorganik, tetapi karena tidak memperoleh penyediaan hasil fotosintesis dari daun maka akan mengubah morfogenesis untuk dapat berfungsi sebagai jaringan penyerap nutrient organik eksogenous. Sumber organik eksogenous ini kemudian disintesa menjadi molekul struktural dan fungsional pada tumbuhan tersebut dan digunakan untuk pertumbuhan. Sebaliknya, daun yang seharusnya menjadi sumber senyawa organik bagi pertumbuhan akar akan berubah fungsi menjadi tempat penyerapan unsur hara karena akar tidak menyediakan unsur hara ini untuk disintesa menjadi senyawa organik di dalam
sukrosa dan hormon auxin yang bekerja sama sebagai signal morfogenesis (Hammond and White 2008).
Tanpa proses ini, sel somatik dari explant tidak akan pernah tumbuh menjadi tanaman baru yang utuh. Secara skematis, perbedaan sumber nutrisi yang digunakan untuk pertumbuhan embrio biji dan embrio somatik (plb) dapat dilihat pada gambar 3.
Gambar 3. A. Pemberian auxin dan sukrosa eksogenous akan mengubah program pembangunan jaringan dari sel somatis ke individu baru. B. Embrio yang tumbuh hanya dari sediaan makanan cadangan tidak mengalami perubahan program pembangunan yang telah dirancang secara genetis melalui fusi sel telur dan sperma.
Persoalannya adalah bagaimana sukrosa dan auxin eksogenous tersebut memberi signal agar akar dan daun yang terbentuk memiliki jaringan penghubung (xylem dan floem) yang kuat. Jika hal ini dapat dilakukan, maka akar dengan mudah menyalurkan unsur hara dan daun juga tidak kesulitan menyalurkan hasil fotosintesis. Akan tetapi, penyediaan auxin dan sukrosa pada media tidak terjadi melalui mekanisme pengaturan produksi dan redistribusi yang terprogram secara genetik sehingga tidak tersedia aturan sampai tahap mana redifferensiasi harus dilakukan. Pada teknik invitro ini, jumlah auxin dan sukrosa yang tersedia hanya tergantung pada formula yang digunakan. Sebagai pembanding dapat dilihat mekanisme produksi dan distribusi auxin pada tanaman autotrof.
dihasilkan tanpa melibatkan gen. Periode peralihan produksi auxin dan sukrosa pada pucuk ini akan berakibat pada perubahan waktu penyediaan auxin untuk pertumbuhan akar. Periode ini nampaknya cukup signifikan untuk mempengaruhi arah pembangunan jaringan dan dikenal sebagai apikal dominan (Suyitno 2006). Pada teknik kultur jaringan, mekanisme dominansi pucuk ini nampaknya belum banyak diperhatikan. Sukrosa dan auxin disediakan secara bersamaan sehingga ketika sel menjadi importir auxin dia juga importir sukrosa. Tanpa mekanisme genetik ini, morfogenesis dapat berlangsung tetapi banyak yang tidak sesuai dengan rencana dasar pertumbuhan dan perkembangan tanaman. Mekanisme penyediaan sukrosa untuk pertumbuhan plb menjadi plantlet juga dapat berpengaruh pada rendahnya kapasitas jaringan transport pada bibit hasil kultur jaringan ini. Pada tanaman yang dikembangkan dengan biji, nutrient organik untuk pertumbuhan embrio diimport atas permintaan embrio itu sendiri. Embrio mengeluarkan hormon gibberelin untuk menginduksi enzim yang dapat memecah bahan makanan cadangan. Setelah menjadi molekul sederhana, bahan makanan diserap untuk pertumbuhan akar atau daun. Tanaman ini baru akan memproduksi auxin setelah pucuk tanaman terbentuk. Bersama sama hasil fotosintesis, auxin disalurkan untuk pengembangan sistem perakaran. Jadi sistem perakaran dikembangkan oleh auxin melalui saluran floem yang telah terbentuk dan fungsional. Jadi pada tanaman ini, tidak terjadi pengembangan akar tanpa saluran pengangkutan dari daun. Pada teknik kultur jaringan, perencanaan pembangunan jaringan ini menjadi agak kacau. Pada komposisi hormon yang diberikan, sel kalus hanya tumbuh menjadi akar dan pada komposisi lain hanya tumbuh menjadi daun (Albert et al. 1983). Sangat besar
kemungkinan pada komposisi lainnya, kalus akan membentuk akar dan daun tetapi tidak memiliki
telah ditemukan oleh Robinson (2009) bahwa jaringan pengakutan pada tanaman yang dikembangkan dengan kultur jaringan sangat lemah. Hal ini diperkuat oleh temuan sebelumnya bahwa orientasi pertumbuhan jaringan dalam kultur sangat ditentukan oleh jenis nutrient (Rao dan Narayanaswami 2006).
pertumbuhan jaringan pengangkutan adalah sangat mungkin karena kedua senyawa ini termasuk molekul kecil dan dapat melakukan transport intercelluler. Dengan tidak tersedianya sukrosa eksogenous, perlakuan ini sangat mungkin dapat meningkatkan aktivitas fotosintesis yang diikuti oleh pengembangan jaringan floem dan diikuti oleh pengembangan jaringan xylem yang mengangkut unsur hara dari akar.
Tergantung pada jenis tanaman dan kondisi lingkungan, perbaikan jaringan penghubung antara akar dan daun atau sebaliknya, perlu dilakukan untuk meningkatkan aktivitas autotrofik bibit dari kultur jaringan. Kajian tentang variasi pemberian senyawa anorganik (tanpa senyawa organik eksogenous) mungkin sangat bermanfaat, demikian juga modifikasi faktor lingkungan lainnya. Pada anggrek epifit, modifikasi fungsi dapat terjadi apabila tanaman menghadapi kondisi kekeringan. Pada tanaman ini terjadi mobilisasi senyawa glucomannan untuk mempertahankan metabolisme sehingga bahaya kekeringan dapat dikurangi (Stancato et al.
2001). Pada tanaman Arabidopsis thaliana,
kekeringan diatasi dengan meningkatkan produksi aquaporin (Sade et al. 2010). Pada
kondisi kekurangan unsur hara, tanaman biasanya merespon dengan peningkatan pertumbuhan akar baru (Cooper and Clarkson 1989, Hammond and White 2008). Pada mekanisme adaptasi ini, tanaman nampaknya meningkatkan pertumbuhan suatu struktur untuk mengatasi kekurangan baik air maupun unsur hara. Adaptasi struktur sangat mungkin juga terjadi pada daun apabila tanaman kekurangan produksi hasil fotosintesis. Tanaman akan meningkatkan pertumbuhan perangkat fotosintesis apabila keperluan akan sukrosa untuk pertumbuhan naik. Akan tetapi, untuk dapat terjadinya mekanisme adaptasi struktur ini, bahan bangunannya harus tersedia dalam jumlah yang cukup. Misalnya, untuk perbanyakan akar ketika unsur hara kurang maka pasokan sukrosa untuk pertumbuhan ini harus cukup. Sebaliknya,
apabila perangkat fotosintesis seperti klorofil harus diperbanyak karena keperluan hasil fotosintesis meningkat maka bahan penyusun klorofil seperti nitrogen harus tersedia dalam jumlah yang cukup. Faktor mana yang pertama harus diperbaiki agar masalah aktivitas autotrofik bibit kultur jaringan dapat diatasi. Apakah meningkatkan produksi sukrosa untuk meningkatkan pertumbuhan akar atau meningkatkan penyediaan unsur hara untuk perbaikan perangkat fotosintesis. Penulis menduga bahwa kedua hal ini dapat dilakukan secara bersamaan. Peningkatan produksi sukrosa untuk pertumbuhan akar dapat dilakukan melalui peningkatan supply air dan CO2. Pada anggrek yang tergolong tanaman CAM, produksi hasil fotosintesis ditemukan naik apabila tanaman ditumbuhkan dengan kadar CO2 yang dinaikkan dan dapat meningkatkan pertumbuhan terutama pertumbuhan akar (Sok Siam Gouk et al. 1999). Kenaikan aktivitas fotosintesis oleh
pemberian CO2 yang tinggi pada anggrek CAM ini hampir sama dengan kenaikan fotosintesis yang terjadi pada tanaman Triticum aestivum
yang ditumbuhkan secara ex-vitro (Mulholland
et al. 1997). Apabila penyediaan unsur hara
dinaikkan maka biosintesis perangkat fotosintesis akan naik. Akan tetapi hal ini baru akan terjadi apabila unsur hara tersebut bisa mencapai daun. Walaupun tidak sesuai dengan fungsi struktur pada organ tanaman, pemberian unsur hara lewat daun adalah alternatif karena jaringan pembuluh dari akar ke daun belum kuat. Pemberian unsur hara pada fase awal pertumbuhan diluar botol tentu harus dilakukan dengan sangat hati-hati karena akumulasi ion yang terlalu tinggi dapat menjadi toksik bagi tanaman (Flower dan Yeo 1986), atau dapat terjadinya induksi abscisc acid yang mengakibatkan tanaman menghentikan pertumbuhan (Bensen et al.1988). Upaya
perbaikan pertumbuhan jaringan pengangkutan dapat dilakukan melaui mekanisme seperti pada gambar 4.
KESIMPULAN
Bibit tanaman yang dihasilkan melalui kultur jaringan memiliki variasi anatomi, morfologi dan fisiologi. Walaupun variasi ini tidak seluruhnya berakibat pada rendahnya viabilitas bibit, penyempurnaan perlu dilakukan baik anatomi maupun fisiologi. Penyempurnaan morfogenesis, sama seperti pertumbuhan invitro, dapat dilakukan melalui pemberian zat pengatur tumbuh auxin, sedangkan penyempurnaan fisiologis dapat dilakukan melalui pengaturan pemberian kondisi lingkungan.
Dengan membaiknya struktur anatomi tanaman maka penyerapan unsur hara dan redistribusi hasil fotosintesis akan makin tinggi yang akhirnya akan mempertinggi aktivitas autotrofik pada tanaman. Hasil fotosintesis tersedia dalam jumlah yang lebih tinggi selanjutnya sangat penting untuk pertumbuhan tanaman sampai fase reproduksi. Akan tetapi karena
spesies adalah spesifik, maka perbaikan kondisi lingkungan hanya berlaku pada spesies tertentu saja.
DAFTAR PUSTAKA
Adiputra IGK and Anderson JW. 1995. Effect of sulphur nutrition on redistribution of sulphur in vegetative barley. Physiol. Plant. 95: 643-650.
Albert B, Bray D, Lewis L, Raff M, Robert K, Watson JD. l983. Molecular Biology of the cell. Garland Publishing, Inc. New
York and London.
Bensen RJ, Boyer JS and Mullet JE. 1988. Water deficit-induced changes in Abscisic acid Growth, Polysomes, and Translatable RNA in Soybean hypocotyls. Plant Physiol 88, 289-294.
root in the regulation of nutrient uptake.
J. Exp. Bot. 40:753-762.
Daisy P. Sriyanti Hendaryono dan Ari Wijayani 1994. Teknik kultur jaringan, pengenalan dan petunjuk perbanyakan tanaman secara vegetatif- modern.
Penerbit Kanisius.
Dutra D, Johnson TR, Kauth PJ, Stewart SL, Kane ME and Richardson L. 2008. Asymbiotic seed germination, in vitro seedling development, and greenhouse acclimatization of the threatened terrestrial orchid Bletia purpurea. Plant Cell, Tissue and Organ Culture Volume 94, Number
1, 11-21, DOI: 10.1007/s11240-008-9382-0
Flower TJ dan Yeo AR. 1986. Ion Relations of Plants Under Drought and Salinity.
Australian Journal of Plant Physiology
13(1) 75. doi:10.1071/PP9860075 Gardner FP, Pearce RB, Mitchell RL. 1991.
Fisiologi Tanaman Budidaya. Penerbit
Universitas Indonesia.
Gastal F and Lemaire G. 2002. N uptake and distribution in crops: an agronomical and ecophysiological perspective. J. Exp. Bot. 53, No. 370: 789-799.
Hammond JP and White PJ. 2008. Sucrose transport in the phloem: integrating root responses to phosphorus starvation.
Journal of Experimental Botany, vol.
59, No.1, pp. 93-109.
Kanai S, Ohkura K, Adu-Gyamfi JJ, Mohapatra, PK, Nguyen NT, Saneoka H and Fujita K. 2007. Depression of sink activity precedes the inhibition of biomass production in tomato plants subjected to potassium deficiency stress. Journal of Experimental Botany
58(11):2917-2928
Komal R 2011. One step method of plantlet regeneration in Trichosanthes dioica Roxb.: An approach towards cost effective and shorter protocol. African Journal of Biotechnology Vol 10 (1), pp. 9-12).
Mineo L. 1990. Plant Tisssue culture techniques. Department of Biology,
Lafayette College, Easton, Pennsylvania 18042.
Mulholland BJ, Craigon J, Black CR, Colls JJ, Atherton J, Landon G. 1997. Impact of elevated atmospheric CO2 and O3 on gas exchange and chlorophyll content in spring wheat (Triticum aestivum L). Journal of experimental Botany, vol. 48, No. 315,
pp. 1853-1863.
Pospisilova J, Ticha I, Kadlecek P, Haisel D and Plzakova S. 1999. Acclimatization of micropropagated plants to ex-vitro condition. Biologia Plantarum 42 (4): 481-497.
Pasternak TP, Prinsen E, Ayaydin F, Miskolczi P, Potters G, Asard H, Onckelen HA, Dudits D, and Fehér A 2002. The role of auxin, pH, and stress in the activation of embryogenic cell division in leaf protoplast-derived cells of alfalfa. Plant Physiol, Vol.
129, pp. 1807-1819.
Rao, P. S. and Narayanaswami, S. 1972. Morphogenetic Investigations in callus cultures of Tylophora indica. Physiologia Plantarum, 27: 271–276. doi: 10.1111/
j.1399-3054.1972.tb03613.x
Rianawati S, Purwito A, Marwoto B, Kurniati R dan Suryanah. 2009. Embriogenesis Somatik dari Eksplan Daun Anggrek Phalaenopsis sp L. J. Agron. Indonesia
37 (3) : 240 – 248.
Robert HS & Friml J 2009. Nature Chemical Biology 5, 325 – 332. doi:10.1038/
nchembio.170
Robinson JP, Britto SJ and Senthilkumar S. 2009. Comparative Anatomical Studies on Emilia zeylanica C. B. Clarke with in vitro Regenerated Plants. Middle-East Journal of Scientific Research 4 (3):
140-143
in improving water use efficiency, hydraulic conductivity, and yield production under salt stress. Plant Physiology, Vol. 152,
pp. 245–254,
Sawada S, Igarashi T and Miyachi S. 1982. Effect of nutritional level of phosphate on photosynthesis and growth studied with single, rooted leaf of dwarf bean. Plant and Cell Physiology 23: 27-33.
Siwach P, Grower K and Gill AR 2011. The influence of plant growth regulator, explant nature and sucrose concentration on in vitro callus growth of Thevetia peruviana.
Asian Journal of Biotechnology 3 (3):
280-292. DOI: 10.3923/ ajbkr.2011.280292.
Sok Siam Gouk, Jie He and Choy Sin Hew 1999. Changes in photosynthetic capability and carbohydrate production in an epiphytic CAM orchid plantlet exposed to super-elevated CO2. Environmental and experimental Botany 41: 219-230.
Stancato GC, Mazzafera P, Buckeridge MS. 2001. Effect of a droght period on the mobolisation of non-structural carbohydrates, photosynthetic efficiency
and water status in an epiphytic orchid.
Plant Physiology and Biochemistry
39:1009-1016. Doi: 10.1016/S0981-9428(01)01321-3.
Suyitno AI.2006. Petunjuk Praktikum Fisiologi Tumbuhan Lanjut. Program
Studi Biologi-Jurdik Biologi, FMIPA, Universitas Negeri Yogyakarta.
Terry N and Ulrich A. 1973.Effect of phosphorus deficiency on the photosynthesis and respiration of leaves of sugar beet.
Tromas A, Perrot-Rechenmann C 2010. Recent progress in auxin biology. C. R. Biologies
333 297–306. Elsevier Masson SAS doi:10.1016/j.crvi.2010.01.005
Utami ESW, Sumardi I, Taryono, Semiarti E 2007. Pengaruh á-naphtaleneacetic acid (NAA) terhadap embriogenesis somatik anggrek bulan Phalaenopsis amabilis (l.) bl. Biodiversitas,volume 8, nomor 4 :
295-299
INVENTARISASI JENIS MOLUSCA DI DANAU TAMBLINGAN, BALI
Ni Made Suartini
Jurusan Biologi FMIPA Universitas Udayana Email: madesuartini_risky@yahoo.com
ABSTRAK
Penelitian ini bertujuan untuk mengetahui jenis-jenis Molusca di danau Tamblingan, Bali. Pengambilan sampel Molusca dilakukan pada bulan Pebruari 2004. Stasiun dibuat mengelilingi danau dan pengambilan sampel dilakukan dari pinggir menuju ke tengah danau sampai pada kedalaman 2 m. Identifikasi sampel dilakukan dengan acuan Butot (1955), Jutting (1956), Pennak (1989) dan membandingkan dengan koleksi yang ada di Laboratorium Malakologi, Puslit Biologi-LIPI Cibinong. Ditemukan enam spesies molusca dimana Anodonta woodiana merupakan spesies dengan kepadatan tertinggi dan dijumpai hampir di setiap stasiun. Kata kunci: jenis, moluska, danau Tamblingan, Anodonta woodiana
ABSTRACT
This research was aimed to investigate Mollusks species in Lake Tamblingan, Bali. For this research, samples were collected in February 2004 from sampling stations located between sites that close to the edge of the lake towards the centre part of the lake which is 2 m in depth. Samples were identified based on procedure by; Butot (1955), Jutting (1956), Pennak (1989) and it were also compared with specimens that available in Malacology Lab, Research Centre for Biology, Indonesian Institute of Science (LIPI), Cibinong. This study found 6 species of Mollusks in the lake. Among the species, Anodonta woodiana was found to have the highest density and was ob-served in almost all sampling stations.
Key words: species, mollusk, Lake Tambingan, Anodonta woodiana
PENDAHULUAN
Danau mempunyai manfaat penting bagi masyarakat yaitu sebagai sumber air untuk keperluan rumah tangga, usaha pertanian, perkebunan, peternakan dan perikanan. Selain itu, danau juga digunakan sebagai obyek wisata alam, tempat dilaksanakannya pendidikan dan penelitian. Bagi masyarakat Bali, selain fungsi tersebut di atas, danau juga mempunyai fungsi sosial budaya yaitu sebagai bagian dari ritus keagamaan yang sudah turun temurun (Bappedalwil II, 1998).
Danau Tamblingan merupakan salah satu sumber perairan tawar yang terletak pada ketinggian 1100 m dari permukaan laut berada
di Kecamatan Banjar Kabupaten Buleleng, Bali. Danau ini merupakan danau vulkano tektonik dengan luas permukaan air 1,15 km2, kedalaman maksimum 40,5 m dan kedalaman rata-rata 23,5 m. Menurut Kayane (1992) dalam Delinom et al. (1997), fluktuasi air danau Tamblingan pada
musim panas adalah 0,69 m dan pada musim hujan 0,3 m.
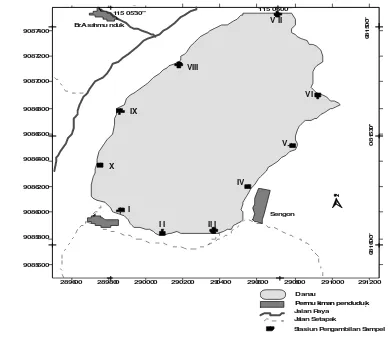
289600 289800 290000 290200 290400 290600 290800 291000 291200
Permu kiman pendudu k lokasi
Berkaitan dengan hal di atas maka untuk menambah informasi keanekaragaman hayati di danau Tamblingan perlu kiranya dilakukan suatu penelitian untuk mengungkap lebih lanjut keberadaan Molusca yang merupakan salah satu sumber keanekaragaman hayati yang terdapat di danau tersebut.
BAHAN DAN METODE Waktu dan Tempat Penelitian
Pengambilan sampel moluska dilakukan di danau Tamblingan Provinsi Bali pada bulan Pebruari 2004. Identifikasi dilakukan di Laboratorium Malakologi, Puslit Biologi-LIPI Cibinong. Analisis kualitas air di Laboratorium Analitik Fakultas MIPA Universitas Udayana sedangkan analisis substrat dilakukan di Laboratorium Tanah, Fakultas Pertanian Univer-sitas Udayana.
Pengambilan Sampel
Stasiun dibuat mengelilingi danau dan pengambilan sampel dilakukan dari pinggir (batas tepi air danau) menuju ke tengah danau sampai pada kedalaman 2 m. Pada daerah pinggir
sampai kedalaman 50 cm, pengambilan sampel dilakukan dengan kwadran ukuran 1 m x 1 m dan kedalaman berikutnya dengan “Ekman grab” (30 cm x 30 cm).
Setiap habitat pada setiap stasiun diidentifikasi kondisi lingkungannya (terbuka atau tertutup), substrat (pasir, lumpur, batu, kerikil dan sebagainya) serta diukur faktor fisika kimiawi dan tekstur substrat lingkungan perairan.
Sampel Molusca yang diambil dipisahkan dari substrat dengan menggunakan saringan bentos kemudian diawetkan dengan alkohol 70%.
Pemilahan dan Pengamatan Morfologi Pemilahan Molusca dilakukan berdasarkan jenis yang diperoleh di setiap stasiun serta dihitung jumlah individunya. Identifikasi sampel dilakukan dengan acuan Butot (1955), Jutting (1956), Pennak (1989) serta membandingkan dengan koleksi yang ada di Laboratorium Malakologi, Puslit Biologi-LIPI Cibinong.
HASIL DAN PEMBAHASAN
Kisaran Parameter Fisika-Kimiawi Perairan
Parameter fisika-kimiawi perairan pada umumnya berada pada kisaran yang diperlukan untuk kehidupan biota perairan (Tabel 1).
Kisaran suhu di danau Tamblingan masih berada pada kisaran suhu untuk kehidupan plankton. Menurut Odum (1993), kisaran suhu untuk kehidupan plankton adalah 200 C-300 C. Secara tidak langsung dapat dikatakan bahwa danau Tamblingan mempunyai suhu yang mendukung kehidupan Molusca karena di danau tersebut plankton masih dapat hidup. Secara umum diketahui bahwa plankton merupakan makanan Molusca.
Kisaran suhu berada pada kisaran dimana Molusca masih dapat hidup tetapi suhu tersebut merupakan batas suhu minimum yang dibutuhkan untuk kelangsungan hidupnya. Pada batas suhu minimum, reproduksi maupun pertumbuhan yang terjadi pada Molusca menjadi rendah. Rendahnya tingkat reproduksi dan pertumbuhan merupakan salah satu faktor yang menyebabkan kepadatan Molusca di danau tersebut menjadi rendah. Hart & Fuller (1974) menyatakan bahwa suhu berpengaruh terhadap kwalitas reproduksi Gastropoda air tawar.
Pada umumnya pH masih berada pada kisaran pH yang disukai oleh biota perairan yaitu 7-8,5 (Effendi, 2003). Anggota kelompok
Molusca umumnya lebih menyukai perairan yang bersifat basa. Keong Lymnaeidae menyukai PH >7 (Pennak 1989), terlihat dari ditemukannya
Lymnaea rubiginosa di danau Tamblingan
sedangkan kerang memerlukan pH 5,6-8,3 (Hart & Fuller, 1974), terlihat dari ditemukannya
Anodonta woodiana.
Nilai kekeruhan dapat dikatakan cukup baik karena nilai kekeruhan 5-30 NTU masih baik untuk kehidupan biota perairan sedangkan nilai kekeruhan kurang dari 5 NTU akan lebih baik bila didukung faktor fisika dan kimiawi lainnya di perairan tersebut.
Kandungan oksigen terlarut terlihat masih mampu mendukung kehidupan Molusca karena beberapa anggotanya seperti A. woodiana
memerlukan kisaran oksigen terlarut untuk kehidupannya sekitar 3,8-12,5 mg/l (Suwignyo, 1975).
Tekstur Substrat
Tekstur substrat terdiri atas tipe lempung, lempung berdebu, lempung berpasir, lempung liat berdebu, lempung liat berpasir, liat, pasir berlempung dan pasir.. Dari semua tipe tekstur substrat tersebut sebagian besar mempunyai fraksi pasir dan debu lebih tinggi dibandingkan dengan fraksi liat.
Kondisi tekstur substrat seperti di atas dapat dikatakan sesuai dengan kehidupan Molusca karena Molusca pada umumnya menyukai Inventarisasi Jenis Molusca Di Danau Tamblingan, Bali Ni Made Suartini
Tabel 1. Kisaran parameter fisika-kimiawi di danau Tamblingan
Parameter Nilai
Suhu (oC) 22 - 24,5
pH 7,99 - 8,30
Kekeruhan (NTU) 0 - 14,86
Oksigen terlarut (mg/l) 7,40 - 8,08
substrat tersebut. Keong Melanoides tuberculata banyak ditemukan di perairan
bagian tepi yang dangkal dan berlumpur (Duggan, 2002). Menurut Suwignyo (1975) A. woodiana
menyukai perairan dengan substrat lumpur dengan sedikit pasir. Adanya substrat lumpur dan juga pasir merupakan salah satu faktor yang mendukung kehadiran A. woodiana.
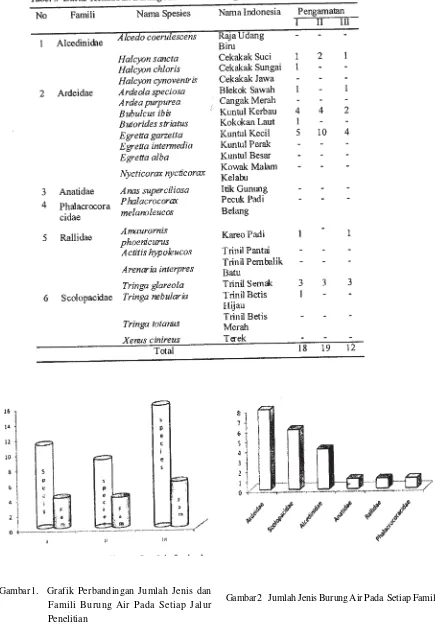
Jenis dan Kepadatan Moluska
Ditemukan 6 jenis Molusca yang termasuk Kelas Gastropoda dan Bivalvia dengan jumlah
individu yang berbeda pada setiap stasium seperti tercantum pada Tabel 2 dan Tabel 3.
Jumlah individu tiap jenis yang diperoleh berkisar antara 1-59 (Tabel 3). StasiunVII jumlahnya paling sedikit karena stasiun tersebut
kurang mendukung untuk kehidupan Molusca terlihat dari kondisi stasiun yang kotor dengan sampah plastik, bekas botol air kemasan dan airnya kelihatan sedikit berbusa. Kondisi stasiun seperti itu karena daerah tersebut merupakan tempat memancing. Disamping itu, stasiun tersebut juga mendapat cukup naungan sehingga suhunya paling rendah dibandingkan dengan stasiun lainnya. Pada suhu rendah tingkat reproduksi serta perkembangan akan menurun. Hart & Fuller (1974) menyatakan bahwa suhu berpengaruh terhadap kualitas reproduksi Gas-tropoda air tawar.
L. rubiginosa ditemukan hanya pada stasiun
ditemukan menempel pada daun teratai dan juga di dasar perairan.
C. javanica hanya ditemukan di stasiun I dan
III yang daerahnya terbuka dengan substrat lumpur. Hal tersebut disebabkan C. javanica menyukai perairan dengan substrat lumpur dan sedikit berpasir ( Djajasasmita, 1993) dan biasanya jenis tersebut hidup dengan cara membenamkan sebagian atau seluruh cangkangnya di dalam lumpur (Marsetiyowati, 1983).
P. canaliculata ditemukan dengan telurnya
yang menempel pada tumbuhan air. Ditemukannya jenis tersebut karena mempunyai adaptasi yang baik dan juga didukung oleh kemampuannya memakan semua yang ada di lingkungan atau bersifat polifagus (Ghesquiere, 2003).
A. woodiana merupakan jenis dengan
jumlah individu terbanyak karena jenis tersebut dapat melangsungkan siklus hidupnya dengan baik. Hal tersebut dipengaruhi oleh kesesuaian faktor lingkungan untuk kelangsungan hidupnya dan adanya ikan nila yang diperlukan sebagai inang untuk penempelan larvanya.
Sistem hidrologi danau juga diduga mempengaruhi jumlah jenis yang ditemukan. Danau Tamblingan merupakan danau tertutup yaitu tidak mempunyai aliran air masuk (inlet) dan keluar (outlet) alami, berbeda dengan danau lain seperti danau Ranau (Sulastri et al., 1999) dan danau Poso (Marwoto, 2000) yaitu dikelilingi banyak sungai yang bermuara ke dalam pearairan danau. Aliran sungai yang masuk ke perairan danau biasanya mengandung detritus yang merupakan sumber makanan bagi hewan pemakan detritus. Dengan tidak adanya inlet di danau Tamblingan maka tidak ada masukan unsur hara dari luar danau. Disamping itu, daerah inlet maupun outlet umumnya lebih dangkal dan umum diketahui bahwa Molusca lebih menyukai perairan yang dangkal. Di danau Poso, genus Melanoides dan Thiara umumnya banyak ditemukan di daerah inlet, outlet dan perairan danau yang dangkal.
KESIMPULAN
Ditemukan enam jenis Molusca yang termasuk kelas Gastropoda dan Bivalvia.
Anodonta woodiana merupakan jenis dengan
kepadatan tertinggi dan dijumpai hampir di setiap stasiun.
SARAN
Diperlukan kajian lebih luas tentang potensi danau sebagai kawasan konservasi maupun ekoturisme termasuk pemanfaatan sumber biota yang ada di danau tersebut.
UCAPAN TERIMAKASIH
Terimakasih diucapkan kepada Dr. Ir. M.F. Rahardjo, DEA dan Dra. Ristiyanti M. Marwoto, MSi yang telah memberikan banyak masukan dalam penelitian ini.
DAFTAR PUSTAKA
[Bappedalwil II] Badan Pengendalian Dampak Lingkungan Wilayah II. 1998. Norma dan Program Danau Lestari Provinsi Dati I Bali.
Butot LJM. 1955. The Mollusc Fauna of Panaitan (Prinseneiland). Land and Frehwater Molluscs. Treubia 23 (1) :
69-135.
Delinom RM, M Djuwansah , MS Siregar. 1997. Hydrological Behaviour of Lake Beratan, Lake Buyan and Lake Tamblingan:A Wa-ter Balance Approach. Di dalam: InWa-terna- Interna-tional Hydrology Programme. Proceed-ing Workshop on Ecosystem Approach to Lake and Reservoir Management;
Kuta-Bali, 22-25 July 1997. Hlm. 37-46 Djajasasmita M. 1993. Catatan Tentang Moluska di Sawah-Sawah Sekitar Bogor: Komposisi Jenis, Potensi dan Peranannya.
Jurnal Biologi Indonesia 1(1): 48-53.
Duggan IC. 2002. First Record of a Wild Popu-lation of the Tropical Snail Melanoides tuberculata in New Zealand Natural
Wa-ters. New Zealand Journal of Marine and Freshwater Research. 36:825-829.
Effendi H. 2003. Telaah Kualitas Air. Bagi Pengelolaan Sumber Daya Alam dan Lingkungan Perairan. Kanisius.
Yogyakarta.
Ghesquiere S. 2003. Apple Snails (Ampullariidae). Ecology. http:// w w w. a p p l e s n a i l s . n e t / c o n t e n t / ecology.php [17 Apr 2004].
Hart Jr CW, SLH Fuller. 1974. Pollution Ecol-ogy of Freshwater Invertebrates.
Aca-demic Press. New York and London. Jutting BWSS. 1956. Systematic Studies on the
Non-Marine Mollusca of the Indo-Aus-tralian Archipelago. Treubia 28 (2) :
259-477
Marwoto RM dan M Djajasasmita. 1986. Moluska di Perairan Tepi Danau Singkarak Sumatra Barat: Komposisi dan Kepadatan Jenisnya. Berita Biologi.
3(6):292-295.
Marsetiyowati R. 1983. Moluska di Kolam-Kolam Kebun Raya Bogor. Buletin Kebun Raya. 6 (2):39-42.
Marwoto RM. 2000. Keong Air Tawar Suku Thiaridae di Danau Poso dan Studi Morfologi, Anatomi Marga Tylomelania dari Danau Poso, Sulawesi Tengah (Moluska:Gastropoda:Caenogastropoda)
[tesis]. Program Studi Biologi, Program Pascasarjana Universitas Indonesia. Jakarta.
Odum EP. 1993. Dasar-Dasar Ekologi. Edisi
Ketiga. Gajah Mada University Press. Yogyakarta.
Pennak RW. 1989. Fresh-Water Invertebrates of the United States. Protozoa to Mol-lusca. Third Edition. John Wiley & Sons,
Inc. New York.
Sulastri, M Badjoeri, Y Sudarso dan MS Stawal. 1999. Kondisi Fisik- Kimia dan Biologi Perairan Danau Ranau Sumatra Selatan.
Limnotek. 6(1): 25-38.
Suwignyo S, B Widigdo, Y Wardianto dan M Krisanti. 1998. Avertebrata Air. Jilid 2.
Institut Pertanian Bogor, Fakultas Perikanan dan Ilmu Kelautan . Bogor. Suwignyo P. 1975. Kijing Taiwan Suatu Sumber
Protein Baru di Indonesia. BIOTROP/TA/
PENGARUH SUPLEMENTASI SOMATOTROPIN TERHADAP
PERUBAHAN BOBOT BADAN TIKUS BETINA USIA
ENAM BULAN DAN SATU TAHUN
Ni Wayan Sudatri
Jurusan Biologi, FMIPA Universitas Udayana, Kampus Bukit Jimbaran Bali Email : wayan_sudatri@yahoo.com
ABSTRAK
Dengan bertambahnya usia, wanita biasanya mengalami perubahan komposisi tubuh yaitu bertambahnya lemak tubuh dan menurunnya masa otot. Hal ini dikarenakan menurunnya sekresi somatotropin yang berperan dalam lipolisis dan anabolisme protein. Rancangan percobaan yang dipergunakan dalam penelitian ini adalah Rancangan Faktorial dengan empat faktor yaitu dosis, lama penyuntikan, umur, dan waktu pembedahan. Dosis somatotropin yang digunakan adalah 0 mg, dan 9 mg/kg bobot badan. Lama penyuntikan adalah 3 minggu dan 6 minggu. Umur tikus terdiri atas enam bulan dan satu tahun, serta waktu pembedahan yaitu setelah akhir periode penyuntikan somatotropin dan 2 minggu setelah penghentian penyuntikan somatotropin. Kombinasi antarfaktor tersebut menjadi 2 x 2 x 2 x 2 = 16. Masing-masing kelompok terdiri atas 3 ulangan sehingga jumlah tikus coba menjadi 3 x 16 = 48 ekor. Hasil penelitian menunjukkan bahwa suplementasi somatotropin dosis 9 mg/kg bobot badan meningkatkan bobot badan tikus betina usia enam bulan dan satu tahun karena terjadinya peningkatan anabolisme protein.
Kata kunci: bobot badan, tikus betina, somatotropin
ABSTRACT
When woman is getting older, chemical composition of her body is changed where fat deposit increased, while muscles masses decreased. These changes is believed to occur because of decreasing in somatotropin secretion. The somatotropin compound plays an important role in lipolysis and protein anabolism. In order to examine the effect of somatotropin, this present study used female rats as experimental organism. In this study, factorial design was employed which consisting of 4 factors, namely doses, duration of injection, age and dissection time. Somatotropin doses were 0 and 9 mg/kg body weight, duration of injection were 3 and 6 weeks, rats age were 6 and 12 months, dissections time were just after the termination of injection using somatotropin and 2 weeks afterward. Combinations of these factors generated 16 combinations and since replicate for each group was 3, total number of rat required for this study become 48 rats. This study showed that supplementation of 9 mg/kg body weight increased body weight of those 6 and 9 month female rat. It is concluded that the increased in body weight is attributed by increasing protein anabolism.
Key words: body weight, female rats, somatotropin
PENDAHULUAN
Pada wanita, semakin bertambahnya usia biasanya ditandai oleh semakin bertambahnya bobot badan dan perubahan komposisi tubuh.
protein dan liposilis lemak. Penurunan produksi hormon ini menyebabkan distribusi lemak di bagian perut pada wanita bertambah seiring dengan meningkatnya umur (Veldhuis et al.
2005).
Hormon pertumbuhan atau growth hormone
(GH) yang juga disebut somatotropin merupakan protein kecil yang mengandung sekitar 191 asam amino dalam satu rantai yang mempunyai berat molekul 22.005. Somatotropin merangsang pertumbuhan semua jaringan tubuh yang mampu tumbuh. Somatotropin meningkatkan pertambahan ukuran sel dan meningkatkan mitosis bersama peningkatan jumlah sel. Sebagai contoh adalah peningkatan pertambahan berat badan tikus yang disuntik dengan hormon pertumbuhan/somatotropin setiap hari, dibanding dengan yang sama sekali tidak mendapat hormon pertumbuhan (Guyton 1995).
Somatotropin dihasilkan oleh pituitari anterior terutama oleh kelompok sel-sel asidofil khususnya sel-sel somatotrof. Pada tikus betina, sel somatotrof dan sel mammotrof jumlahnya berimbang, namun pada tikus jantan jumlah sel somatotrof jauh lebih banyak dari sel mammotrof (6:1). Ratio inilah mungkin yang menyebabkan ukuran jantan jauh lebih besar dari betina pada semua spesies (Bolander 1994).
Pada masa pertumbuhan, ketika pertumbuhan berlangsung cepat, kadar somatotropin sangat tinggi. Sejalan dengan bertambahnya usia, kadar somatotropin dalam tubuh terus menurun. Keberadaan somatotropin ini setelah masa pertumbuhan diperlukan untuk menjaga kesehatan fisik dan mental secara umum seperti untuk perbaikan jaringan, penyembuhan, penggantian sel-sel yang rusak, untuk produksi enzim, untuk menjaga fungsi otak, menjaga kesehatan rambut, kuku, dan lain sebagainya. Setelah umur 20 tahun, produksi somatotropin menurun 14% setiap 10 tahun. Setelah umur 60 tahun, produksi somatotropin menurun 75% atau lebih (Bengtsson 2000).
proses metabolisme tubuh. Somatotropin meningkatkan kecepatan sintesis protein dalam semua sel tubuh, menurunkan penggunaan karbohidrat di seluruh tubuh, meningkatkan mobilisasi lemak dan penggunaan lemak untuk energi serta merangsang produksi IGF-1 di hati untuk memacu pertumbuhan tulang (Guyton 1995).
Pada babi yang disuntik dengan somatotropin, katabolisme protein dan efisiensi makanan meningkat serta daging yang dihasilkan mempunyai kadar lemak yang sangat rendah (Bush et al 2002; Davis et al. 2004).
Sementara itu, pada manusia terapi dengan somatotropin dapat meningkatkan masa otot, kekuatan fisik, menurunkan kelelahan, menurunkan kadar lemak (terutama lemak perut), meningkatkan kekuatan tulang, membuat kulit tampak lebih muda, fungsi seks meningkat, fungsi hati, ginjal, limpa, dan otak menjadi lebih sehat (Klatz & Kahn 1998). Berdasarkan informasi di atas, maka penelitian pengaruh suplementasi sumatotropin pada dosis, waktu pemberian dan umur tikus yang berbeda perlu dipelajari lebih lanjut untuk untuk mengetahui perubahan bobot badan pada tikus betina.
BAHAN DAN METODE
Penelitian dilakukan di kandang percobaan FKH IPB, Laboratorium Fisiologi dan Farmakologi FKH IPB mulai bulan November 2005 sampai dengan Maret 2006. Hewan coba yang digunakan dalam penelitian ini adalah 48 ekor tikus betina strain Wistar yang terdiri atas kelompok umur enam bulan (24 ekor) dan satu tahun (24 ekor) yang dibeli dari Balai Pengujian Obat dan Makanan, Departemen Kesehatan, Jakarta.
terdiri atas enam bulan dan satu tahun serta waktu pengambilan contoh yaitu satu hari dan 2 minggu setelah penyuntikan dihentikan. Kombinasi antarfaktor tersebut menjadi 2 x 2 x 2 x 2 = 16. Masing-masing kelompok terdiri atas 3 ulangan sehingga jumlah tikus coba menjadi :3 x 16 = 48 ekor. Pada kelompok yang disuntik dengan somatotropin selama 3 minggu setelah sinkronisasi disampling sebagian dan dibiarkan sebagian untuk disampling 2 minggu kemudian. Begitu juga dengan kelompok yang disuntik selama 6 minggu, setelah disinkronisasi disampling sebagian, setengahnya lagi disampling 2 minggu kemudian. Pada setiap pembedahan jumlah tikus yang dibedah adalah 12 ekor untuk pengambilan data yang lain.
Sebelum diberi perlakuan, semua hewan coba diaklimatisasikan terlebih dahulu di dalam kandang percobaan selama satu minggu. Selama pemeliharaan hewan coba diberi makan pelet standar dan minum air keran secara ad libitum.
Sebelum diberi perlakuan semua tikus betina ditimbang bobot awalnya. Penyuntikan somatotropin dilakukan secara intramuskuler (im) sesuai dengan rancangan percobaan. Selesai perlakuan semua tikus betina ditimbang bobot akhirnya, sehingga diperoleh perubahan bobot badan dengan mengurangkan bobot akhir dengan bobot awal hewan coba. Untuk
mengetahui perubahan bobot badannya, bobot badan tikus juga ditimbang setiap minggu selama perlakuan maupun yang dibiarkan selama dua minggu tanpa perlakuan baru dibedah.
Data yang didapatkan dianalisis secara statistika dengan ANOVA GLM memakai program Minitab 11 for Windows dan bila terdapat perbedaan yang nyata atau sangat nyata akan dilanjutkan dengan uji Duncan
HASIL DAN PEMBAHASAN
Bobot badan awal tikus betina ini dipengaruhi oleh umur. Tikus betina yang berumur 1 tahun mempunyai bobot badan awal yang lebih besar dari tikus betina umur 6 bulan (Tabel 1).
Peningkatan bobot badan tikus pada kelompok yang disuntik somatotropin baik yang berumur 1 tahun maupun 6 bulan disebabkan oleh peningkatan laju sintesis protein dan meningkatnya penggunaan lemak sebagai sumber energi yang terlihat dari analisis proksimat karkas bahwa kadar protein lebih tinggi dari kadar lemaknya (Eddy 2006). Hal senada juga telah dilaporkan oleh Davis et al. (2004) dan Bush et al. (2002) yang menyatakan bahwa pemberian
somatotropin eksogen meningkatkan sintesis protein. Bartke (2005) juga melaporkan bahwa penurunan masa otot, peningkatan jaringan adiposa, dan tanda-tanda penuaan lainnya disebabkan oleh penurunan sekresi somatotropin dari pituitari. Beberapa perubahan tersebut dapat diperbaiki dengan terapi somatotropin/GH.
KESIMPULAN
Suplementasi somatotropin dosis 9 mg/kg bobot badan meningkatkan bobot badan tikus betina usia enam dan satu tahun karena terjadi peningkatan sintesis protein.
SARAN
Disarankan untuk melakukan penelitian dengan umur tikus lebih dari satu tahun (lebih tua) dan melakukan analisis kadar lemak dan kadar protein untuk mengetahui peningkatan kadar protein atau lemaknya.
UCAPAN TERIMA KASIH
Penulis mengucapkan banyak terima kasih kepada Bapak Prof. Dr. Wasmen Manalu dan Ibu Dr. Nastiti Kusumorini yang telah banyak memberikan masukan sehingga penelitian ini dapat berjalan dengan lancar. Penulis juga mengucapkan banyak terima kasih kepada Lemlit Unud atas bantuan dana yang diberikan melalui Dana Dosen Muda.
DAFTAR PUSTAKA
Bartke A. 2005. Minireview: role of the growth hormone/insulin-like growth factor system in mammalian aging. Endocrinology 146:
3718–3723.
Bengtsson BA, Gudmundur J, Stephen MS, Helen S, Peter HS. 2000 Treatment of growth hormone deficiency in adults . J Clin Endocrinol Metabol 85: 933-942.
Bolander FF. 1994. Molecular Endocrinology. USA: Academic Press, Inc. Hlm 23-30 Bush JA, Guoyao Wu, Suryawan A, Hanh V,
Nguyen,Teresa AD. 2002.Somatotropin induced amino acid conservation in pigs involves differential regulation of liver and gut urea cycle enzyme activity. J Nutr 132:
59-67.
Davis TA, Bush JA, Vann RC, Suryawan A, Kimball SR, Burrin DG. 2004. Somatotropin regulation of protein metabolism in pigs. J Anim Sci 82: 207
– 210.
Eddy L. 2006. Suplementasi somatotropin untuk memperbaiki tampilan fisiologis tikus jantan usia enam bulan dan satu tahun (Tesis). Bogor: Program
Pascasarjana, Institut Pertanian Bogor. Guyton AC.1995. Fisiologi Manusia dan
Mekanisme Penyakit. Edisi III. Petrus A, penerjemah; Jakarta: EGC.Terjemahan dari: Text Book of Medical Physiology.
Klatz R, Kahn C. 1998. Grow Young with hGH.
New York, USA: Harper Perennial. Veldhuis JD , Erickson D, Mielke K, Farty SL,
Daniel M. Keenal, Cyril YB. 2005. Distinctive inhibitory mechanisms of age and relative visceral adiposity on growth hormone secretion in pre and postmenopausal woman studied under hypogonal clamp. J Clin Endocrinol Metabol 87: 5160-5167.
EFEKTIVITAS TANAMAN HIAS Sansevieria lorentii
DALAM MENYERAP POLUTAN TIMBAL (Pb)
Ni Luh Suriani
Jurusan Biologi Fakultas MIPA Unud, Kampus Bukit Jimbaran-Bali niluhsurianisuriani@yahoo.com
ABSTRAK
Tujuan penelitian ini adalah untuk mengetahui efektivitas dan kandungan polutan timbal (Pb) pada tanaman Sanseviera Lorentii yang dipergunakan sebagai tanaman hias jalan.
Penelitian ini dilaksanakan di Laboratorium analitik Universitas Udayana. Penentuan kandungan logam berat Pb pada tanaman Sanseviera lorentii ditentukan dengan teknik
kurva kalibrasi. Hasil penelitian menunjukkan bahwa kandungan logam berat Pb pada
Sanseviera lorentii yang ditaruh pada lokasi yang berbeda selama 1 bulam semuanya
mengalami peningkatan masing-masing daearah. Padangsambian (58.31 µg/g) dengan efektifitas 19,22% kemudian disusul oleh daerah Sesetan ( 56.13 µg/g ) dengan efektifitas 18,63% dan terendah daerah Tabanan (20.5 µg/g) dengan efektifitas 7,72% . Adanya perbedaan kandungan Pb diketiga lokasi disebabkan oleh faktor kepadatan lalu lintas.
Kata kunci:, Sanseviera lorentii, logam berat Pb.lalu lintas
ABSTRACT
This research was aimed to see the effectiveness of Sansevieria plants to take up Pb emitted into the air by vehicles in highways Denpasar-Tuban and Denpasar-Tabanan by placing Sansevieria plants in Sesetan and Padangsambian. Other Sansevieria plants, as control, were placed in a remote area, i.e. Munduk Paku village, Penebel, Tabanan. Concentration of Heavy metal (Pb), in the leaves of Sanseviera plants, were measured in analitical laboratories, Udayana University. This study showed that the highest Pb content was found in plants located in Padangsambian (58.31 µg/g), followed by plants located in Sesetan (56.13µ g/g) and the lowest Pb content was found in plants located in Munduk Paku village. Effectiveness of Sansevieria to take up Pb was 19.22%, 18.63% and 7.72% for Padangsambian, Sesetan and Munduk Paku, respectively. This study concluded that different content of Pb in plants is attributed by the level of traffic density.
Key words, Sanseviera lorentii, heavy metal Pb, traffic
PENDAHULUAN
Di Indonesia, nama sanseviera lebih dikenal
dengan nama lidah mertua. Dulu tanaman hias ini hanya dipandang sebelah mata dan dianggap tidak bermanfaat kecuali sebagai tanaman pagar. Tetapi sejak ditemukan jenis-jenis baru yang
keunggulan yang jarang ditemukan pada tanaman lain, yaitu sangat resisiten terhadap polutan dan bahkan mampu menyerapnya. Hal ini
dikarenakan Sanseviera mengandung bahan
menyerap 107 jenis unsur berbahaya di udara; diantaranya karbon monoksida dan timbal. Timbal merupakan logam berat yang sangat berbahaya. Daya racunnya dapat menyebabkan peradangan pada mulut, diare, anemia, mual,sakit perut serta kelumpuhan (Saeni, 1999). Pb sering ditambahkan pada bensin yang berfungsi sebagai anti letup pada kendaraan bermotor. Kendaraan bermotor sangat besar andilnya didalam menyumbang polutan Pb. Tujuan dilaksanakan penelitian ini adalah untuk mengetahui efektivitas dan kandungan polutan timbal (Pb) pada tanaman
Sansevieralorentii yang dipergunakan sebagai
tanaman hias jalan. Manfaat yang diharapkan dari penelitian ini adalah memberikan informasi kepada masyarakat bahwa tanaman hias
Sansevieralorentii selain digunakan sebagai
tanaman hias juga dapat digunakan sebagai tanaman penyerap polutan Pb
BAHAN DAN METODE
Penelitian ini dilakukan di Laboratorium Analitik Universitas Udayana Kampus Bukit Jimbaran Bali selama 2 bulan. Bahan yang digunakan dalam penelitian ini berupa tanaman
Sansiviera lorentii, Asam sulfat pekat, asam
nitrat, dan aquadest. Alat-alat yang digunakan dalam penelitian ini adalah oven, timbangan analitik, penumbuk, hot plate, labu takar , AAS, dan alat-alat lain yang diperlukan. Sampel diambil di daerah Renon sebanyak 15 pot. Kemudian ditaruh pada tiga lokasi penempatan sampel. masing lokasi ditaruh 5 pot. Masing-masing lokasi tersebut adalah: Lokasi pertama di Desa Munduk Paku , Penebel Tabanan merupakan kontrol karena sedikit kendaraannya. Lokasi kedua di Desa Padang Sambian, Denpasar Barat, yang merupakan daerah dengan lalu lintas kendaran padat, dan lokasi ketiga di daerah Sesetan, Denpasar Selatan yang juga dengan lalu lintas kendaraan padat.
Sebelum sampel tanaman ditaruh di ketiga lokasi, sampel diambil secara komposit untuk dianalisis logam berat Pb-nya di Laboratorium untuk mengetahui kandungan Pb awal (Ho).
Kemudian sampel ditaruh selama 1 bulan di masing-masing lokasi yang telah ditentukan. Setelah satu bulan sampel di masing-masing lokasi dianalisis lagi kandungan logam Pb-nya untuk mengetahui kandungan logam berat Pb akhir. Sampel dipotong kecil-kecil masing-masing sebanyak 0,5 kg dimasukkan ke dalam kantong kertas. Kemudian dikeringkan dalam oven 110
oC, sampai beratnya konstan. Selanjutnya sampel
ditumbuk dan ditimbang sebanyak 0,3 g dan dimasukkan ke dalam labu destruksi. Kemudian ditambahkan asam sulfat secara hati-hati. Larutan dipanasi secara perlahan-lahan sampai larutan berwarna hitam, lalu ditetesi larutan asam nitrat pekat sebanyak 10 sampai 20 tetes sampai larutan berwarna jernih (kekuningan) . Kemudian larutan didinginkan dan diencerkan dengan aquades sampai 25 ml kemudian dikocok. Lalu larutan disaring sehingga diperoleh filtrat. Filtrat tersebut dianalisis dengan AAS.
Data kemudian disajikan dalam bentuk tabel, untuk melihat perbandingan efektifitas kandungan logam berat Pb pada masing-masing lokasi yang ditunjukkan oleh histogram.
HASIL DAN PEMBAHASAN
Hasil penelitian menunjukkan bahwa ada peningkatan kandungan logam berat Pb setelah ditaruh selama sebulan di tiga lokasi yaitu lokasi yang kepadatan lalu lintasnya rendah yang dianggap kontrol (daerah Munduk Paku, Senganan, Penebel, Tabanan) dan lokasi yang kepadatan lalu lintasnya tinggi yaitu daerah Denpasar (Padangsambian dan Sesetan). Secara rinci mengenai hasil rata-rata Pb di tiga lokasi disajikan pada Tabel 1.
Data pada Tabel 1 menunjukkan bahwa terjadi peningkatan kandungan rata-rata Pb. Pb tertinggi terjadi di daerah Padangsambian (58.31 µg/g), kemudian disusul oleh daerah Sesetan ( 56.13 µg/g ) dan terendah daerah Tabanan (20.5 µg/g). Hal ini disebabkan oleh faktor kepadatan lalu lintas. Mengingat daerah Padangsambian dan Sesetan merupakan daerah yang tergolong lalu lintas padat dan daerah Tananan tergolong lalu
lintas tidak padat. Hal ini didukung oleh hasil penelitian Bumi Tunjungan (2011), bahwa pada daerah yang padat lalu lintas kandungan Pb di udara juga tinggi, karena adanya penambahan Pb pada bahan bakar premium yang berfungsi sebagai anti letup pada kendaraan dan sepeda motor.
Tingginya penyerapan logam Pb pada
tanaman Sansiviera lorentii, (Tabel 1)
disebabkan karena tanaman Sansiviera lorentii,
mampu menyerap polutan secara optimal. Kemampuan menyerap ini mungkin disebabkan oleh fakta morfologi ataupun fisiologi. Secara
morfologi, Sansiviera memiliki tekstur daun yang
kasar dan tebal sehingga sangat resisten terhadap polutan (Whika FD, 2010). Secara fisiologi, tanaman ini mengandung bahan aktif pregnane glikosid yang mampu mereduksi polutan menjadi asam organik, gula, dan beberapa senyawa asam amino (Trubus XXXVI, 2006). Namun demikian menurut Ekawati dkk (2009) masing-masing
jenis tanaman Sansiviera memiliki daya serap
terhadap racun yang berbeda-beda, dimana
Sansiviera lorentii memiliki daya serap terhadap
racun paling tinggi dan tidak menunjukkan
kerusakan organ tanaman. Sansiviera juga
mampu menyerap ratusan racun di udara. Badan NASA international menggunakan tanaman
Sansiviera sebagai tanaman peredam radiasi
nuklir dengan cara menanam 100 hektar tanaman
Sansiviera di sekitar badan nuklir (Bumi
Tunjungan, 2011). Tanaman Sansiviera juga
diminati oleh Negara Jepang yang menggunakan tanaman ini sebagai tanaman dalam ruangan yang dapat menetralisir racun-racun yang ada dalam ruangan. Kemampuan sansiviera ini merupakan keuntungan bagi Indonesia karena sampai saat ini Indonesia tercatat sebagai pengekspor
terbesar Sansiviera ke Negara Jepang.
Untuk melihat perbandingan efektifitas penyerapan logam berat Pb pada masing-masing lokasi penelitian maka data disajikan dalam bentuk grafik (Gambar 2).
Data pada Gambar 2 memperlihatkan bahwa efektifitas penyerapan Pb tertinggi terdapat pada sampel yang diletakkan di daerah Padangsambian (19.22%). Karena di daerah ini berdasarkan hasil survey kepadatan lalu lintasnya tinggi dan peletakan sampel dekat sekali dengan jalan raya. Hal ini juga didukung oleh penelitian Ekawati dkk (2010) bahwa efektifitas
sangat dipengaruhi oleh kepadatan lalu lintas. Karena semakin padat lalu lintas maka Pb yang dilepas oleh kendaraan bermotor ke udara semakin tinggi. Sehingga efektifitas penyerapan
sansiviera terhadap logam berat Pb semakin
tinggi. Hal ini didukung oleh Novirina H (2007) dan Faizin (2005) bahwa semakin padat lalu lintas maka logam Pb yang diserap oleh tanaman
dari udara juga semakin tinggi. Sansiviera dapat
dijadikan sebagai filter udara, sehingga udara menjadi bersih dan sehat (Trubus XXXVI,2006). Bumi Tunjungan (2011) menyatakan bahwa 5
helai daun Sansiviera dapat membersihkan
ruangan yang berukuran 100m3.
KESIMPULAN
Dari hasil penelitian ini dapat disimpulkan; terjadi peningkatan kandungan logam berat Pb
pada tanaman hias Sansiviera lorentii setelah
ditaruh selama 1 bulan di lokasi yang padat lalu lintas. Kandungan Pb tertinggi ditemukan di daerah Padangsambian (58.31 µg/g), disusul daerah Sesetan (56,13 µg/g) dan lokasi yang jarang lalu lintas yaitu daerah Tabanan (20,5 µg/ g µg/g ). Efektifitas Penyerapan logam berat Pb
oleh tanaman Sansiviera lorentii tertinggi
terdapat di daerah Padangsambian 19,22 % sisusul oleh daerah Sesetan 18,63 % dan terendah terdapat di daerah Tabanan 7,72 %. Efektifitas penyerapan logam berat Pb oleh
tanaman Sansiviera lorentii sangat dipengaruhi
oleh kepadatan lalu lintas.
DAFTAR PUSTAKA
Almatsier. 2000. Ilmu Gizi.PT. Gramedia.
Bumi Tunjungan. 2011. Sanseivera Tanaman
Penyerap Racun.
Available at : http://www.bumi
tunjungan.blogsport.com/2011/01 Darmono. 1995. Logam Dalam Sistem Makhluk
Hidup. UI-PRES.
Deny. 2011. Kandungan logam berat Pb dan Cd pada Sayuran Wortel dan Sawi Hijau. Hasil Penelitian.
Ekawati dkk.2009. Tanggapan Pertumbuhan
Sanseivera Sanseivera spp Terhadap
Logam berat Pb. Hasil Penelitian.
Faizin. 2005. Kandungan Logam Berat Pb pada
Tanaman Hias Sanseivera Hasil
Penelitian
Kohar dkk. 2004. Studi Kandungan Logam Berat Pada Taaman Kangkung. Jurnal MakaraSain Vol 8 no 3
Novirina.H.2007. Kajian Efektifitas Tanaman Dalam Menyerap Kandungan Pb di Udara. Jurnal Rekayasa Perencanaan. Vol.3.No 2.
Saeni,M. 2000. Kimia Lingkungan.. Diktat. PAU IPB Bogor. Tidak Dipublikasikan. Sudarmaji dkk. 2006. Toksikologi Logam Berat
B-3 dan Dampaknya terhadap
Kesehatan. Jurnal Kesehatan Lingkungan.Vol.2 No 2.
Suriani. 2008. Kualitas Air Mangrove Ditinjau Dari Sitaf Fisik-Kimia Di Hutan
Mangrove Patung Ngurah Rai Tuban Denpasr Selatan Bali. Jurnal Ilmu
Lingkungan. Ecotrophic. ISSN 1907-5626. Unud.
Suriani dan Susun.2010. Uji Kandungan Logam Berat Pb dan Cd pada Buah Apel Lokal Rum Biuty dan Buah Apel Import Fuji yang Beredar di Kota Denpasar. Laporan Penelitian .
Susun dan Suriani. 2010. Bioremediasi Logam Berat Pb dan Cd pada Limbah Pencelupan dengan Tanaman Air. Laporan Peneltian.
Sutyawathi.2011. Kandungan Logam Berat Pada Bayam Cabut. Hasil Penelitian
Trubus XXXVI. 2006. Sisk Building Syndrome. Tanaman Penyerap Racun. Jakarta. Whika FD. 2010. Uji Anatomi dan Molekuler
Sanseivera trifasciata. Jurnal
Matematika, Sains dan Teknologi Vol. 11. No.2.