PENGKLONAN DAN KARAKTERISASI GEN PerL
DARI KEDELAI PEKA CEKAMAN ALUMINIUM
AKHMAD AMIRULLAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2008
ABSTRAK
AKHMAD AMIRULLAH. Pengklonan dan Karakterisasi Gen PerL dari Kedelai Peka Cekaman Aluminium. Dibimbing oleh UTUT WIDYASTUTI SUHARSONO dan ENCE DARMO JAYA SUPENA.
Peroksidase diketahui sebagai salah satu gen yang terinduksi oleh cekaman Aluminium (Al).
Pada kedelai kultivar Lumut yang diberi cekaman pH 4 + Al 1,6 mM, terlihat adanya ekspresi gen peroksidase (PerL). cDNA PerL telah diisolasi dari akar tanaman kedelai kultivar Lumut. Penelitian ini bertujuan untuk mengkarakterisasi gen peroksidase dari tanaman kedelai kultivar Lumut yang memiliki sifat peka terhadap cekaman Al. Keberadaan cDNA PerL pada cDNA total diamplifikasi dengan menggunakan primer spesifik yang didesain berdasarkan Arabidopsis
thaliana (X71794). cDNA PerL disisipkan ke dalam pGEM®-T Easy dan plasmid rekombinan telah berhasil diintroduksi ke dalam Escherichia coli galur DH5α. Pengurutan cDNA PerL dengan DNA sekuencer ABI Prism Model 310 menghasilkan sekitar 1100 pb yang menyandikan 353 asam amino. Analisis kesejajaran lokal nukleotida dengan Per dari beberapa spesies yang terdaftar di
GeneBank menunjukkan bahwa cDNA PerL mempunyai persentase kemiripan sebesar 98%
dengan DNA Per dari A. thaliana (X71794). Hasil penelusuran daerah domain terkonservasi menunjukkan bahwa asam amino PerL diduga termasuk ke dalam tipe secretory peroxidase.
ABSTRACT
AKHMAD AMIRULLAH. Cloning and Characterization of PerL Gene from Aluminum Sensitive-Soybean. Supervised by UTUT WIDYASTUTI SUHARSONO and ENCE DARMO JAYA SUPENA.
Peroxidase is known as one of the genes that induced by Aluminum (Al) stress. PerL gene
was expressed under pH 4 + Al 1,6 mM stress. The PerL cDNA had been isolated from soybean roots cv Lumut. The objective of this research was to characterize PerL gene from Al sensitive-soybean cv Lumut. The PerL cDNA was isolated from total cDNA using specific primers were designed based on Per gene of Arabidopsis thaliana (X71794). The PerL cDNA was inserted into pGEM®-T Easy and introduced into Escherichia coli DH5α strain. Sequencing of PerL cDNA using ABI Prism 310 automated DNA sequencer showed that PerL cDNA has 1100 bp encoding 353 amino acids. Basic local alignment analysis based on nucleotide showed that PerL cDNA has 98% similarity with Per of A. thaliana (X71794). Conserved domain of the PerL amino acids suggested that perL has a secretory peroxidase type.
PENGKLONAN DAN KARAKTERISASI GEN PerL
DARI KEDELAI PEKA CEKAMAN ALUMINIUM
AKHMAD AMIRULLAH
Skripsi
sebagai salah satu syarat memperoleh gelar
Sarjana Sains pada
Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2008
Judul
: Pengklonan dan Karakterisasi Gen PerL dari Kedelai Peka Cekaman
Aluminium
Nama
: Akhmad Amirullah
NRP
: G34101059
Menyetujui:
Pembimbing I
Pembimbing II
Dr. Ir. Utut Widyastuti S, M.Si.
NIP 131 851 279
Dr. Ir. Ence Darmo Jaya Supena, M.Si.
NIP 131 851 278
Mengetahui:
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Dr. drh. Hasim, DEA.
NIP 131 578 806
PRAKATA
Alhamdulillahirobbil ’aalamin. Segala puji bagi Allah SWT atas segala limpahan karunia-Nya, sehingga karya ilmiah ini dapat diselesaikan. Penelitian ini dibiayai oleh Proyek Hibah Bersaing IX Direktorat Jenderal Pendidikan Tinggi (DIKTI) atas nama Dr. Ir. Utut Widyastuti Suharsono, M.Si.
Terima kasih penulis ucapkan kepada Dr. Ir. Utut Widyastuti Suharsono, M.Si dan Dr. Ir. Ence Darmo Jaya Supena, M.Si. sebagai pembimbing atas segala bimbingan, waktu, sarana dan nasihat yang telah diberikan. Terima kasih kepada Dr. Ir. Miftahudin, M.Si. atas saran dan masukannya yang berguna dalam penyempurnaan tulisan ini. Terima kasih kepada Kepala Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB Darmaga beserta seluruh staf dan karyawan atas sarana, prasarana, dan bantuannya selama penulis melakukan penelitian di Laboratorium Biorin (Biotechnology Research Indonesia-the Netherland).
Ucapan terima kasih juga penulis sampaikan kepada Bapak Abdul Mulya, Mbak Pepi Elvafina, Thesiawaty Dian Fauziah, Bapak Syarifin Firdaus, M.Si., Bapak Muzuni, S.Si., M.Si., Bapak Drs. Y. Ulung Anggraito, M.Si., Bapak Yasir, S.Si., Mbak Rina, S.Si., M. Bahrelfi, S.Si., Syahnada Jaya, dan teman-teman seperjuangan di Laboratorium Biorin atas bantuan, kerjasama dan saran yang telah diberikan. Terima kasih juga penulis haturkan kepada teman-teman Biologi 38 yang tidak dapat disebutkan satu-persatu. Secara khusus penulis sampaikan terima kasih kepada orang tua, adik-adik, Nuryanti, dan Zahid Abdurrazzaq, atas kasih sayang dan perhatian kalian yang membuat penulis sangat bersemangat.
Semoga karya ilmiah ini bermanfaat bagi agama, bangsa dan negaraku.
Bogor, Juni 2008
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 20 April 1984 dari ayah bernama Munawar dan ibu bernama Umi Kalsum. Penulis merupakan anak pertama dari tiga bersaudara.
Tahun 2001 penulis lulus dari SMU Negeri 10 Bekasi dan diterima di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam Institut pertanian Bogor melalui jalur Ujian Masuk Perguruan Tinggi Negeri (UMPTN).
Selama mengikuti perkuliahan penulis pernah menjadi asisten praktikum Biologi Dasar pada semester genap tahun ajaran 2004/2005 dan semester ganjil 2005/2006. Penulis aktif di organisasi mahasiswa Dewan Keluarga Masjid Al Ghifari IPB sebagai Wakil Ketua pada tahun 2005/2006. Penulis pernah melaksanakan kegiatan praktik lapang pada tahun 2004 dengan topik Perbanyakan Anggrek secara In Vitro di Kebun Anggrek Parung. Pada tanggal 5 Januari 2007, penulis menikah dengan Nuryanti, dan pada tanggal 7 November 2007 penulis dikaruniai seorang putra diberi nama Zahid Abdurrazzaq.
DAFTAR ISI
Halaman DAFTAR GAMBAR... DAFTAR LAMPIRAN... PENDAHULUAN Tujuan Penelitian... Waktu dan Tempat Penelitian... BAHAN DAN METODEBahan... Metode... Amplifikasi cDNA PerL dari cDNA total kedelai lumut... Pengklonan cDNA PerL ke dalam plasmid pGEM®-T Easy... Seleksi E. coli DH5α yang mengandung plasmid rekombinan... Analisis cDNA sisipan... Sekuensing cDNA PerL dan analisis urutan nukleotida... HASIL DAN PEMBAHASAN
Hasil... Amplifikasi cDNA PerL... Pengklonan cDNA PerL ke dalam plasmid pGEM®-T Easy... Analisis urutan nukleotida... Pembahasan... KESIMPULAN... DAFTAR PUSTAKA... LAMPIRAN... vii vii 1 1 1 1 1 2 2 2 3 3 3 3 4 5 5 5 7
vi
DAFTAR GAMBAR
Halaman 1 2 3 4 5Amplifikasi cDNA PerL menggunakan primer spesifik. (1) 1 kb ladder, (2)
PerL berukuran sekitar 1 100 pb...
Seleksi E. coli DH5α transforman pada media seleksi (ampisilin, X-gal, dan IPTG)... PCR koloni putih E. coli yang membawa PerL. (1) 1 kb ladder, (2), (3), dan (4) adalah koloni putih positif pembawa plasmid rekombinan... Pemotongan plasmid rekombinan. (1) 1 kb ladder, (2) plasmid rekombinan... Daerah domain terkonservasi pada urutan asam amino PerL...
3 4 4 4 5
DAFTAR LAMPIRAN
Halaman 1 2 3 4Peta fisik plasmid pGEM®-T Easy yang digunakan sebagai vektor pengklonan...
Hasil sekuensing dengan menggunakan mesin sekuensing otomatis ABI PRISM 310 Genetic Analyzer... Perbandingan sekuen cDNA PerL dari kedelai kultivar Lumut, DNA Per dari A.
thaliana (X71794), dan Per dari Glycine max (L78163)...
Urutan asam amino PerL dari kedelai kultivar Lumut, Per dari G. max (L78163), dan Per dari A. thaliana (X71794)...
8
9
13
15
PENDAHULUAN
Kedelai (Glycine max) merupakan salah satu komoditi pertanian dengan kandungan protein yang tinggi dan banyak disukai masyarakat. Hal ini ditunjukkan dari besarnya kebutuhan kedelai dalam negeri Indonesia yang mencapai 2 juta ton per tahun, sementara produksi kedelai dalam negeri hanya 650000 ton (Deptan 2008). Sehingga untuk memenuhi kebutuhan kedelai dalam negeri, Indonesia harus mengimpor kedelai dari negara lain.
Upaya untuk meningkatkan produksi kedelai dalam negeri adalah melalui metode intensifikasi yaitu penggunaan kultivar unggul dan perbaikan teknik budidaya, maupun metode ekstensifikasi atau perluasan lahan pertanaman. Namun upaya ekstensifikasi terbentur dengan kondisi lahan di Indonesia. Lahan seluas 18,2 juta hektar di Indonesia memiliki pH rendah (masam), padahal lahan tersebut cukup potensial untuk dikembangkan sebagai lahan pertanian, khususnya untuk kedelai (Mulyani et al. 2003).
Kondisi tanah masam dapat meningkatkan kelarutan ion Aluminium (Al) dalam bentuk ion Al3+ sehingga dapat bersifat toksik bagi tanaman. Pada tanah masam, ion Al3+ yang terakumulasi merupakan faktor penyebab utama yang membatasi tingkat keberhasilan panen (Kochian 1995), karena ion Al3+ menghambat pertumbuhan akar tanaman (Gardner 1998). Cekaman ion Al3+ diketahui dapat menginduksi beberapa gen yang berhubungan dengan sistem pertahanan terhadap oxidative stresses seperti peroksidase (Richards et al. 1998). Cekaman ion Al3+ pada ujung akar tanaman dapat menyebabkan terjadinya proses peroksidasi lipid pada membran sel (Cakmak dan Horst 1991).
Penentuan kadar cekaman Al untuk induksi gen didasarkan pada perbedaan panjang akar antara kontrol dengan perlakuan cekaman minimal sebesar 50% (Ryan et al. 1994). Kedelai kultivar Lumut (peka Al) mengalami penghambatan pertumbuhan panjang akar primer sampai 50% dengan penambahan Al sebesar 0,8 - 1,6 mM (Anwar 1999). Pada kedelai kultivar Lumut yang diberi cekaman pH 4 + Al 1,6 mM, terlihat adanya ekspresi gen
peroksidase, tetapi cekaman Al tidak
menyebabkan gen peroksidase terinduksi (Lubis 2008).
Tujuan Penelitian
Penelitian ini bertujuan untuk mengkarakterisasi gen peroksidase (PerL) dari tanaman kedelai kultivar Lumut yang memiliki sifat peka terhadap cekaman Al.
BAHAN DAN METODE Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan mulai bulan Mei 2007 hingga Mei 2008, bertempat di Laboratorium Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) IPB Darmaga. Pengurutan nukleotida (sekuensing) dilakukan di Laboratorium Balai Pengkajian Bioteknologi, Badan Pengkajian dan Penerapan Teknologi (BPPT) Serpong.
Bahan
cDNA total kedelai kultivar Lumut yang digunakan sebagai cetakan, berasal dari hasil penelitian Lubis (2008). Plasmid pGEM®-T Easy (Lampiran 1) (Promega) digunakan sebagai vektor pengklonan dan Escherichia
coli galur DH5α sebagai inang vektor rekombinan.
Primer AtprxCb-F(1) (5’-TCAACTAG-TTTTGTTTTTCCTCTT-3’) dan primer
AtprxCb-R(1175)
(5’-GTTCTCAAGTAAA-CGTCTTGAGAG-3’) digunakan untuk mengamplifikasi cDNA dari gen peroksidase kedelai kultivar Lumut. Primer AtprxCb-F dan AtprxCb-R didesain berdasarkan urutan nukloetida Arabidopsis thaliana (nomor akses GeneBank X71794). Primer SP6
(5’-ATTTAGGTGACACTATAGAA-3’), T7
(5’-TAATACGACTCACTATAGGG-3’), dan LP (5’-GGGGATTTCCAGTGATTG-AT-3’) digunakan dalam pengurutan urutan nukleotida (sekuensing).
Metode
Penelitian ini terdiri dari beberapa tahap, yaitu amplifikasi cDNA yang menyandikan gen PerL, pengklonan gen
PerL ke dalam plasmid pGEM®-T Easy, pengurutan nukleotida PerL dan analisis urutan nukleotida.
Amplifikasi cDNA PerL dari cDNA total kedelai kultivar Lumut
cDNA total kedelai kultivar Lumut digunakan sebagai cetakan untuk amplifikasi cDNA gen PerL. Komposisi PCR adalah 1 µl cDNA total, 2 µl buffer PCR 10x, 2 µl dNTP 2 mM, 1 µl AtprxCb-F 20 pmol, 1 µl
enzim taq DNA polymerase 1 U (Gen Script Inc.) dan 12 µl ddH2O. Kondisi PCR yang digunakan adalah 95 °C 5 menit untuk pra-PCR, siklus PCR diawali dengan 94 °C 30 detik untuk denaturasi, 56 °C 30 detik untuk penempelan primer, 72 °C 2 menit untuk pemanjangan, selanjutnya siklus diulangi sebanyak 37 siklus, dan pasca PCR pada 72 °C 5 menit diikuti dengan inkubasi pada 20 °C 5 menit.
Pengklonan cDNA PerL ke dalam plasmid pGEM®-T Easy
Pengklonan cDNA PerL ke dalam pGEM®-T Easy menggunakan prosedur Promega (1996) dengan cara mencampurkan 1 µl buffer ligasi 10x, 1 µl vektor pGEM®-T Easy (20 ng/µl), 1 µl T4 DNA ligase (5 U/µl), 3 µl produk PCR dan 4 µl ddH2O, kemudian diinkubasi satu malam pada suhu 4 °C.
Hasil ligasi diintroduksikan ke dalam E.
coli galur DH5α melalui transformasi menggunakan prosedur Suharsono (2002). Transformasi diawali dengan pembuatan bakteri kompeten, yaitu dengan membiakkan satu koloni E. coli DH5α dalam 2 ml LB (bakto tripton 1% (b/v), bakto ekstrak khamir 0.5% (b/v), NaCl 1% (b/v)) dan diinkubasi pada suhu 37 °C dengan
kecepatan bergoyang 250 rpm
(environmental shaker ES-20, Biosan). Sebanyak 100 µl E. coli DH5α disubkultur dalam 30 ml LB kemudian diinkubasi dengan kondisi yang sama hingga mencapai densitas bakteri OD600 = 0.4-0.5 (4-7 x 10
7 sel/ml). Sebanyak 1.5 ml kultur dimasukkan ke dalam tabung ependorf dan diinkubasi di dalam es 10 menit kemudian disentrifugasi pada kecepatan 3000 rpm dengan sentrifugasi (Jouan BR-4i) pada suhu 4 °C selama 10 menit. Selanjutnya supernatan dibuang sedangkan endapan bakteri ditambahkan 495 µl (0.33 x volume kultur) buffer transformasi TB (PIPES 10 mM, CaCl2.2H20 15 mM, KCl 250 mM, MnCl2.4H2O 55 mM, pH 6.7) kemudian disuspensi dan diinkubasi dalam es selama 10 menit, disentrifugasi kembali dengan kecepatan 3000 rpm pada suhu 4 °C selama 10 menit. Supernatan dibuang sedangkan endapan bakteri disuspensi dengan 41,25 µl TB kemudian ditambahkan 3.3 µl DMSO 4% dan diinkubasi dalam es selama 10 menit sehingga diperoleh sel kompeten.
Tahap selanjutnya adalah introduksi plasmid rekombinan, yaitu sebanyak 50 µl bakteri kompeten dicampurkan dengan 10 µl
(50-100 ng) produk ligasi dan diinkubasi di dalam es selama 20 menit. Campuran diberi perlakukan kejutan panas 42 °C selama 45 detik dan segera dimasukan kembali dalam es selama 5 menit. Campuran ditambahkan 100 µl YT (bakto tripton 16 g/l, bakto ekstrak khamir 10 g/l, NaCl 5 g/l, pH 7.0) dan diinkubasi dalam inkubator bergoyang dengan kecepatan 250 rpm pada suhu 37 °C selama 20 menit. Kultur bakteri disebar secara merata pada media LA (LB + agar 15 g/l) yang mengandung ampisilin 100 µg/l,10 µl IPTG 100 mM, dan 50 µl X-gal 2% (b/v), kemudian diinkubasi pada suhu 37 °C selama 16 jam.
Seleksi E. coli DH5α yang mengandung plasmid rekombinan
E. coli DH5α yang membawa plasmid rekombinan diseleksi dengan seleksi putih-biru melalui penambahan X-gal dan IPTG. Koloni putih yang tumbuh pada media yang mengandung ampisilin, X-gal, dan IPTG merupakan koloni yang membawa plasmid rekombinan. Adanya sisipan cDNA PerL di dalam plasmid rekombinan yang terdapat dalam koloni putih dideteksi dengan melakukan PCR koloni. Untuk itu koloni putih diambil dengan menggunakan tusuk gigi steril lalu disuspensikan ke dalam tabung PCR yang telah berisi 6.5 µl ddH2O, kemudian suspensi dipanaskan pada suhu 95 °C selama 10 menit dan diinkubasi pada suhu 15 °C selama 5 menit. Selanjutnya suspensi ditambahkan campuran PCR (komposisi dan kondisi PCR sama dengan tahap amplifikasi cDNA PerL) lalu diamplifikasi menggunakan primer
AtprxCb-F dan AtprxCb-R dengan mesin PCR. Tusuk
gigi yang mengandung koloni putih juga dioleskan pada media LA + ampisilin (100 µg/ml) sebagai replika koloni. Bila produk PCR koloni menunjukkan munculnya pita yang berukuran sekitar 1100 pb, berarti cDNA PerL di dalam plasmid rekombinan terdapat dalam koloni putih, maka selanjutnya replika koloni putih tersebut ditumbuhkan dalam 30 ml LB + ampisilin 100 µg/l dan diinkubasi pada suhu 37 °C dengan kecepatan bergoyang 250 rpm selama 16 jam sebagai persiapan tahap isolasi plasmid.
Analisis cDNA sisipan
Plasmid pGEM®-T Easy rekombinan di dalam E. coli DH5α diisolasi dengan mengikuti prosedur Suharsono (2002). Kultur bakteri yang sudah disiapkan, diambil
2
sebanyak 15 ml dan dimasukan ke dalam tabung falkon 15 ml, kemudian disentrifugasi dengan kecepatan 4000 rpm pada suhu 4 °C selama 10 menit. Supernatan dibuang sedangkan endapan ditambahkan dengan 600 µl larutan suspensi sel (Tris-HCl 50 mM pH 7.5 dan EDTA 10 mM) kemudian divorteks. Selanjutnya campuran ditambahkan dengan 400 µl buffer lisis (NaOH 0.2 M dan SDS 1% (b/v)) dan dibolak-balik secara perlahan beberapa kali. Campuran ditambahkan dengan 400 µl larutan buffer netralisasi (Natrium asetat 1.32 M pH 4.8) kemudian divorteks dan disentrifugasi dengan kecepatan 10000 rpm pada suhu 4 °C selama 10 menit. Supernatan dipindah ke tabung ependorf baru dan selanjutnya diekstrak dengan penambahan PCI (fenol-kloroform-isoamilalkohol dengan perbandingan 25:24:1) sebanyak 1x volume supernatan. Kemudian divorteks dan disentrifugasi dengan kecepatan 10000 rpm pada suhu 20 °C selama 10 menit. Supernatan dipindahkan ke tabung ependorf baru, lalu diendapkan dengan penambahan Natrium asetat 3 M pH 5.2 sebanyak 0.1x volume supernatan dan etanol absolut sebanyak 2x volume supernatan. Larutan diinkubasi pada suhu -20 °C selama 2 jam kemudian disentrifugasi dengan kecepatan 10000 rpm pada suhu 4 °C selama 25 menit. Supernatan dibuang sedangkan endapan DNA plasmid dibilas dengan menggunakan etanol 70% (v/v) dengan bantuan sentrifugasi 10000 rpm pada suhu 4 °C selama 10 menit. Endapan dikeringkan dan disuspensikan dengan ddH2O. Suspensi DNA plasmid ditambahkan RNase (100 µg/ml) sebanyak 0.1x volume suspensi dan diinkubasi pada suhu 37 °C selama 16 jam, kemudian suspensi ditambahkan dengan 500 µl ddH2O. Untuk mengeluarkan cDNA sisipan, DNA plasmid rekombinan dipotong dengan enzim EcoRI (Promega Inc) dengan mencampur 50 ng/µl DNA plasmid, 1 µl enzim restriksi EcoRI 10 U, 2 µl buffer
EcoRI 10x, dan 12 µl ddH2O kemudian diinkubasi pada suhu 37 oC selama 8 jam. Sekuensing cDNA PerL dan analisis urutan nukleotida
Sekuensing cDNA PerL dilakukan dengan DNA sequencer ABI PRISM 310
Genetic Analyzer. Karena ukuran cDNA PerL yang cukup panjang, yaitu 1100 pb,
maka dalam proses ini digunakan primer T7,
SP6, dan LP untuk mendapatkan urutan
nukleotida yang lengkap. Analisis
kesejajaran lokal urutan nukleotida gen PerL menggunakan program BLAST (Basic Local
Alignment Search Tools) (www.ncbi. nlm.nih.gov/blast/Blast.cgi) antara cDNA
PerL dengan DNA Per dari spesies lain
yang terdapat di bank data di GeneBank.
Urutan nukleotida cDNA PerL
diterjemahkan menjadi asam amino dengan menggunakan program expasy translate tool (www.expasy.ch/tools/dna.html). Analisis daerah terkonservasi dan domain pada cDNA PerL dilakukan dengan menggunakan program conserved domain NCBI (www.ncbi.nlm.nih.gov/Structure/
cdd/wrpsb.cgi).
HASIL DAN PEMBAHASAN Hasil
Amplifikasi cDNA PerL
Amplifikasi cDNA PerL pada cDNA hasil penelitian Lubis (2008) menunjukkan hasil positif, ditandai dengan munculnya pita yang berukuran sekitar 1100 pb (Gambar 1).
Pengklonan cDNA PerL ke dalam plasmid pGEM®-T Easy
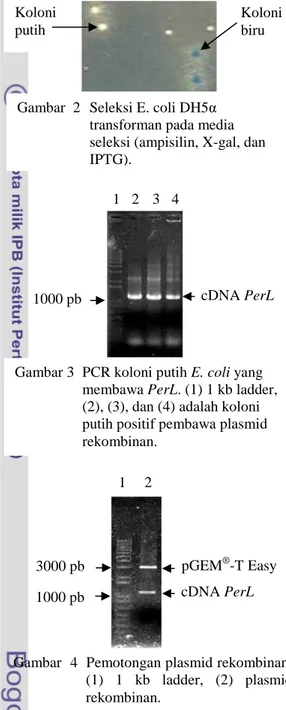
Pada media seleksi yang telah diinkubasi selama 16 jam, muncul koloni E.
coli yang berwarna putih dan biru (Gambar
2).
Dari lima koloni putih yang tumbuh pada media seleksi biru-putih, diambil tiga koloni untuk dilakukan PCR koloni. Hasil PCR koloni berukuran sama dengan hasil amplifikasi cDNA PerL sebelumnya, yaitu sekitar 1100 pb (Gambar 3).
Salah satu koloni putih (nomor 3 pada Gambar 3) dipilih untuk mengeluarkan cDNA PerL dari plasmid rekombinan dengan menggunakan enzim restriksi EcoRI. Hasil pemotongan menunjukkan adanya dua Gambar 1 Amplifikasi cDNA PerL
menggunakan primer spesifik. (1) 1 kb ladder, (2) PerL berukuran sekitar 1100 pb. 1 2 cDNA PerL 1000 pb
3
fragmen yaitu fragmen berukuran sekitar 3000 pb yang merupakan plasmid pGEM®-T Easy, dan fragmen berukuran sekitar 1100 pb yang merupakan PerL (Gambar 4).
Analisis urutan nukleotida
Sekuensing cDNA PerL dengan menggunakan primer T7, SP6, dan LP menghasilkan urutan nukleotida cDNA PerL lengkap (Lampiran 2). Hasil sekuensing kemudian dipisahkan dari nukleotida plasmid dengan memisahkan nukleotida sebelum primer AtprxCb-F dan sesudah
primer AtprxCb-R. Hal ini dilakukan untuk mendapatkan sekuen (urutan nukleotida) cDNA PerL yang diawali dan diakhiri oleh primer spesifik.
Analisis kesejajaran lokal sekuen cDNA
PerL dilakukan untuk melihat kesamaan dan
kekerabatan dengan sekuen DNA Per dari berbagai spesies yang tersedia di GeneBank. Hasil analisis kesejajaran lokal sekuen cDNA PerL menggunakan program BLAST, menunjukkan bahwa sekuen cDNA PerL memiliki nilai kemiripan yang tinggi terhadap sekuen DNA Per dari A. thaliana (X71794). Tingkat keyakinan nilai kemiripan tersebut ditunjukkan oleh nilai persentase kemiripan (Max ident) sebesar 98 %, nilai bit score (BS) sebesar 2071 bits, dan nilai expectation value (E-value) sebesar 0 (nol).
Sekuen DNA Per dari A. thaliana (X71794) dikopi dari GeneBank untuk disejajarkan dengan primer AtprxCb-F,
AtprxCb-R, dan LP. Hal ini dilakukan untuk
membandingkan sekuen DNA Per dari A.
thaliana dengan sekuen cDNA PerL. Hasil
pensejajaran (Lampiran 3) menunjukkan bahwa DNA Per dari A. thaliana (X71794) memiliki sekuen primer AtprxCb-F,
sedangkan pada cDNA PerL terdapat perbedaan beberapa nukleotida primer
AtprxCb-F, yaitu AtprxCb-F =
TCAAC-TAGTTTTGTTTTTCCTCTT sedangkan
cDNA PerL =
TCAACTAGTTTTGT-GTAAAACTT. Sekuen cDNA PerL
diterjemahkan menjadi asam amino dengan menggunakan program expasy translate
tool. Lima kemungkinan urutan asam amino
yang muncul, dipilih satu urutan asam amino yang memiliki banyak asam amino di antara kodon awal (ATG atau M) dan kodon akhir (TGA atau STOP) dan tidak memiliki kodon STOP di tengahnya. Urutan asam amino terpilih ini memiliki 353 asam amino (Lampiran 4). Penelusuran daerah domain terkonservasi pada urutan asam amino PerL dilakukan dengan menggunakan program
conserved domain NCBI. Hasil penelusuran
menunjukkan bahwa urutan asam amino
PerL diduga termasuk ke dalam tipe secretory peroxidase (Gambar 5). Sebagai
pembanding, digunakan hasil penelusuran daerah domain terkonservasi pada urutan asam amino Per dari Glycine max (L78163) dari GeneBank.
Koloni biru Koloni
putih
Gambar 2 Seleksi E. coli DH5α transforman pada media seleksi (ampisilin, X-gal, dan IPTG).
Gambar 3 PCR koloni putih E. coli yang membawa PerL. (1) 1 kb ladder, (2), (3), dan (4) adalah koloni putih positif pembawa plasmid rekombinan.
1000 pb
1 2 3 4
cDNA PerL
Gambar 4 Pemotongan plasmid rekombinan. (1) 1 kb ladder, (2) plasmid rekombinan. 1 2 cDNA PerL pGEM®-T Easy 1000 pb 3000 pb
4
Gambar 5 Perbandingan daerah domain asam amino terkonservasi antara (1) PerL dan (2) Per
G. max (L78163) dari GeneBank.
1 2
Pembahasan
Hasil pensejajaran sekuen cDNA PerL memiliki nilai kemiripan yang tinggi terhadap sekuen DNA Per dari A. thaliana (X71794). Perbedaan terdapat pada sekuen primer AtprxCb-F, namun hal ini tidak mempengaruhi ekspresi gen PerL karena sekuen primer AtprxCb-F terletak pada enam nukleotida sebelum kodon awal (TCAACTAGTTTTGTGTAAAACTTTCA AAAATGCATTTCTC).
Minimnya data tentang peroksidase kedelai di GeneBank menyebabkan tidak ada satupun peroksidase kedelai di GeneBank yang mirip dengan sekuen cDNA PerL. Tentunya hal ini menandakan bahwa sekuen cDNA PerL memiliki ciri khas tersendiri yang berbeda dengan peroksidase kedelai di
GeneBank.
Dari data peroksidase kedelai yang ada di GeneBank, dipilih sekuen DNA Per dari
G. max (L78163) untuk digunakan sebagai
pembanding. Sekuen cDNA PerL sangat berbeda dengan sekuen DNA Per dari G.
max (L78163), seperti yang tersaji pada
Lampiran 3. Tentunya perbedaan juga terlihat pada urutan asam amino yang disandikan oleh kedua sekuen tersebut (Lampiran 4).
Walaupun berbeda sekuen DNA dan urutan asam amino, ternyata PerL dan Per dari G. max (L78163) memiliki kesamaan pada daerah domain terkonservasi (Gambar 5). Urutan asam amino PerL dan Per dari G.
max (L78163) diduga termasuk ke dalam
tipe secretory peroxidase. Namun ada perbedaan letak secretory peroxidase pada kedua urutan asam amino tersebut. Pada
PerL, secretory peroxidase dijumpai pada
asam amino ke-31 sampai urutan ke-331. Sedangkan pada Per dari Glycine max (L78163), secretory peroxidase dijumpai pada asam amino 27 sampai urutan ke-325.
Peroksidase pada tumbuhan, fungi, dan bakteri, dapat dilihat sebagai anggota dari
superfamily yang terdiri atas 3 kelas
berdasarkan kemiripan sekuen (Welinder 1992). Peroksidase kelas I meliputi
cytochrome c peroxidase (CCP) pada ragi, ascorbate peroxidase (AP), dan catalase -peroxidase pada bakteri. Peroksidase kelas II
meliputi secretory peroxidase pada fungi seperti ligninase atau lignin peroxidases (LiPs). Peroksidase kelas III meliputi
secretory peroxidase pada tumbuhan, dengan berbagai fungsi yang spesifik seperti menghilangkan hidrogen peroksida dari kloroplas dan sitosol, mengoksidasi senyawa toksik, biosintesis dinding sel, respon pertahanan terhadap pelukaan, katabolisme
indole 3 acetic acid (IAA), dan biosintesis
etilen. Dengan demikian, PerL diduga termasuk ke dalam Peroksidase kelas III.
KESIMPULAN
Analisis kesejajaran lokal urutan nukleotida menunjukkan bahwa cDNA PerL dari kedelai kultivar Lumut memiliki nilai kemiripan yang tinggi terhadap DNA Per dari A. thaliana (X71794). PerL diduga termasuk ke dalam tipe secretory peroxidase.
DAFTAR PUSTAKA
Anwar S. 1999. Pengklonan Gen-Gen yang Diinduksi oleh Aluminium pada Kedelai [disertasi]. Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor. Cakmak I, Horst WJ. 1991. Effect of
aluminum on lipid peroxidation, superoxide dismutase, catalase and peroxidase activities in root tips of soybean (Glycine max). Physiol Plant 83:463-468.
[Deptan] Departemen Pertanian. 2008. Pemerintah turunkan bea masuk kedelai. Jakarta: Departemen Pertanian.
Gardner RC. 1998. Manipulation of aluminum tolerance by gene transfer [review]. Hayati 5:29-33.
Kochian LV. 1995. Cellular mechanism of aluminum toxicity and resistance in
5
plants. Annu Rev Plan Physiol Plant
Mol Bio 46:237-260.
Lubis RA. 2008. Ekspresi Gen Penyandi Protein Heterotrimerik Gα dan Peroksidase pada Kedelai Kultivar Lumut yang Mendapat Cekaman Aluminium [tesis]. Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor. Mulyani A, Hikmatullah, Subagyo H. 2003
Karakteristik dan potensi tanah masam lahan kering di Indonesia, Prosiding Simposium Nasional Pemberdayaan Tanah Masam, Buku I. Bandar Lampung, 29-30 September 2003. Puslitbang Tanah dan Agroklimat, Balitbangtan Deptan. Bogor.
Promega. 1996. Protocol and application guide. Ed. Ke-3. USA: Promega Co. Richards KD, Schott EJ, Sharma YK, Davis
KR, Gardner RC. 1998. Aluminum induce oxidative stress genes in
Arabidopsis thaliana. Plant Physiol
116:409-418.
Suharsono. 2002. Konstruksi pustaka genom kedelai kultivar Slamet. Hayati 3:67-70. Welinder KG. 1992. Superfamily of plant,
fungal and bacterial peroxidases. Curr
Opin Struct Biol 2:388-393.
Lampiran 1 Peta fisik plasmid pGEM®-T Easy yang digunakan sebagai vektor pengklonan
Lampiran 2 Hasil sekuensing PerL dari kedelai kultivar Lumut dengan menggunakan mesin sekuensing otomatis ABI PRISM 310 Genetic Analyzer (PerL – T7)
Lanjutan Lampiran 2 (PerL – SP6)
Lanjutan Lampiran 2 (PerL – LP)
Lanjutan Lampiran 2 (PerL – LP)
Lampiran 3 Perbandingan cDNA PerL dari kedelai kultivar Lumut, DNA Per dari A. thaliana (X71794), dan Per dari Glycine max (L78163)
> PerL dari kedelai kultivar Lumut
TCAACTAGTT TTGTGTAAAA CTTTCAAAAA TGCATTTCTC TTCGTCTTTA ACATTGTCCA CTTGGACAAT CTTAATCACA TTGGGATGTC TTATGCTTCA TGCATCTTTG TCCGCTGCTC AACTCACCCC TACCTTCTAC GATAGGTCAT GTCCTAATGT CACTAACATC GTACGAGAAA CCATTGTAAA TGAGTTAAGG TCGGACCCTC GTATCGCTGC GAGCATCCTT CGTCTTCACT TCCACGACTG CTTTGTTAAT GGTTGTGACG CATCCATCTT GTTAGACAAC ACGACATCAT TTCGAACAGA GAAAGATGCG TTTGGAAACG CAAATTCGGC TCGGGGATTT CCAGTGATTG ATAGAGTGAA AGCTGCGGTG GAAAGGGCAT GCCCAAGAAC CGTTTCATGC GCAGATATGC TCACCATTGC AGCTCAACAA TCTGACACTT TGGCAGGAGG TCCTTCTTGG AGGGTTCCTT TGGGAAGGAG AGACAGTTTA CAAGCATTCC TGGAACTCGC TAATGCAAAT CTTCCAGCTC CATTCTTTAC ACTTCCACAA CTTAAAGCCA GCTTCAGAAA TGTTGGTCTC GATCGTCCTT CTGATCTCGT TGCTCTCTCC GGTGGTCACA CATTTGGTAA AAATCAATGT CAGTTTATTC TTGACAGATT ATACAATTTC AGCAACACAG GTTTACCCGA CCCTACACTC AACACTACTT ACCTCCAAAC TCTTCGTGGA CTATGCCCCC TTAATGGCAA TCGGAGTGCC TTGGTAGATT TTGATCTACG TACGCCTACG GTTTTCGACA ACAAATACTA CGTGAATCTC AAAGAGCGAA AGGGTCTTAT CCAGAGCGAC CAAGAGTTGT TCTCTAGCCC CAATGCCACT GACACAATCC CCTTGGTGAG AGCATATGCT GATGGCACAC AAACATTCTT CAATGCATTT GTGGAGGCAA TGAATAGGAT GGGAAACATT ACACCAACTA CAGGAACTCA AGGACAAATC AGATTGAACT GTAGAGTTGT GAACTCCAAC TCTCTGCTCC ATGATGTGGT GGATATCGTT GACTTTGTTA GCTCTATGTG AGAATTGTTT ACCCAATATG TGGCTACAAG AATACATATA TATTAATGAA TAAAACTCTC AAGACGTTTA CTTGAGAAC
>Per dari A. thaliana (X71794)
TCAACTAGTT TTGTTTTTCC TCTTTCAAAA ATGCATTTCT CTTCGTCTTC AACATCGTCC ACTTGGACAA TCTTAATCAC ATTGGGATGT CTTATGCTTC ATGCATCTTT GTCCGCTGCT CAACTCACCC CTACCTTCTA CGATAGGTCA TGTCCTAATG TCACTAACAT CGTACGAGAA ACCATTGTAA ATGAGTTAAG GTCGGACCCT CGTATCGCTG CGAGCATCCT TCGTCTTCAC TTCCACGACT GCTTTGTTAA TGGTTGTGAC GCATCCATCT TGTTAGACAA CACGACATCA TTTCGAACAG AGAAAGATCG GTTTGGAAAC GCAAATTCGG CTCGGGGATT TCCAGTGATT GATAGAATGA AAGCTGCGGT GGAGAGGGCA TGCCCAAGAA CCGTTTCATG CGCAGATATG CTCACCATTG CAGCTCAACA ATCTGTCACT TTGGCAGGAG GTCCTTCTTG GAGGGTTCCT TTGGGAAGGA GAGACAGTTT ACAAGCATTC CTGGAACTCG CTAATGCAAA TCTTCCAGCT CCATTCTTTA CACTTCCACA ACTTAAAGCC AGCTTCAGAA ATGTTGGTCT CGATCGTCCT TCTGATCTCG TTGCTCTCTC CGGTGGTCAC ACATTTGGTA AAAATCAATG TCAGTTTATT CTTGACAGAT TCTACAATTT CAGCAACACA GGTTTACCCG ACCCTACACT CAACACTACT TACCTCCAAA CTCTTCGTGG ACTATGCCCC CTTAATGGCA ATCGAAGTGC CTTGGTAGAT TTTGATCTAC GTACGCCTAC GGTTTTCGAC AACAAATACT ACGTGAATCT CAAAGAGCGA AAAGGTCTTA TCCAGAGCGA CCAAGAGTTG TTCTCTAGCC CCAATGCCAC TGACACAATC CCCTTGGTGA GAGCATATGC TGATGGCACA CAAACATTCT TCAATGCATT TGTGGAGGCA ATGAATAGGA TGGGAAACAT TACACCAACT ACAGGAACTC AAGGACAAAT CAGATTGAAC TGTAGAGTTG TGAACTCCAA CTCTCTGCTC CATGATGTGG TGGATATCGT TGACTTTGTT AGCTCTATGT GAGAATTGTT TACCCAATAT GTGGCTACAA GAATACATAT ATATTAATGA ATAAAACTCT CAAGACGTTT ACTTGAGAAC
> Per dari G. max (L78163)
ATGGGTTCCA TGCGTCTATT AGTAGTGGCA TTGTTGTGTG CATTTGCTAT GCATGCAGGT TTTTCAGTCT CTTATGCTCA GCTTACTCCT ACGTTCTACA GAGAAACATG TCCAAATCTG TTCCCTATTG TGTTTGGAGT AATCTTCGAT GCTTCTTTCA CCGATCCCCG AATCGGGGCC AGTCTCATGA GGCTTCATTT TCATGATTGC TTTGTTCAAG GTTGTGATGG ATCAGTTTTG CTGAACAACA CTGATACAAT AGAAAGCGAG CAAGATGCAC TTCCAAATAT CAACTCAATA AGAGGATTGG ACGTTGTCAA TGACATCAAG ACAGCGGTGG AAAATAGTTG TCCAGACACA GTTTCTTGTG CTGATATTCT TGCTATTGCA GCTGAAATAG CTTCTGTTCT GGGAGGAGGT CCAGGATGGC CAGTTCCATT AGGAAGAAGG GACAGCTTAA CAGCAAACCG AACCCTTGCA AATCAAAACC TTCCAGCACC TTTCTTCAAC CTCACTCAAC TTAAAGCTTC CTTTGCTGTT CAAGGTCTCA ACACCCTTGA TTTAGTTACA CTCTCAGGTG GTCATACGTT TGGAAGAGCT
1 61 121 181 241 301 361 421 481 541 601 661 721 781 841 901 961 1021 1081 1141 1 61 121 181 241 301 361 421 481 541 601 661 721 781 841 901 961 1021 1081 1141 1 61 121 181 241 301 361 421 481 541
13
CGGTGCAGTA CATTCATAAA CCGATTATAC AACTTCAGCA ACACTGGAAA CCCTGATCCA ACTCTGAACA CAACATACTT AGAAGTATTG CGTGCAAGAT GCCCCCAGAA TGCAACTGGG GATAACCTCA CCAATTTGGA CCTGAGCACA CCTGATCAAT TTGACAACAG ATACTACTCC AATCTTCTGC AGCTCAATGG CTTACTTCAG AGTGACCAAG AACTTTTCTC CACTCCTGGT GCTGATACCA TTCCCATTGT CAATAGCTTC AGCAGTAACC AGAATACTTT CTTTTCCAAC TTTAGAGTTT CAATGATAAA AATGGGTAAT ATTGGAGTGC TGACTGGGGA TGAAGGAGAA ATTCGCTTGC AATGTAATTT TGTGAATGGA GACTCGTTTG GATTAGCTAG TGTGGCGTCC AAAGATGCTA AACAAAAGCT TGTTGCTCAA TCTAAATAAA CCAATAATTA ATGGGGATGT GCATGCTAGC TAGCATGTAA AGGCAAATTA GGTTGTAAAC CTCTTTGCTA GCTATATTGA AATAAACCAA AGGAGTAGTG TGCATGTCAA TTCGATTTTG CCATGTACCT CTTGGAATAT TATGTAATAA TTATTTGAAT CTCTTTAAGG TACTTAATTA ATCA
601 661 721 781 841 901 961 1021 1081 1141 1201
14
Lampiran 4 Urutan asam amino PerL dari kedelai kultivar Lumut, Per dari G. max (L78163), dan
Per dari A. thaliana (X71794).
....|....| ....|....| ....|....| ....|....|
5 15 25 35
PerL MHFSSSLTLS TWTILITLGC LMLHA--SLS AAQLTPTFYD
Per dari G. max (L78163) ---MGSM RLLVVALLCA FAMHAGFSVS YAQLTPTFYR Per dari A. thaliana (X71794) MHFSSSSTSS TWTILITLGC LMLHA--SLS AAQLTPTFYD
....|....| ....|....| ....|....| ....|....|
45 55 65 75
PerL RSCPNVTNIV RETIVNELRS DPRIAASILR LHFHDCFVNG
Per dari G. max (L78163) ETCPNLFPIV FGVIFDASFT DPRIGASLMR LHFHDCFVQG Per dari A. thaliana (X71794) RSCPNVTNIV RETIVNELRS DPRIAASILR LHFHDCFVNG
....|....| ....|....| ....|....| ....|....|
85 95 105 115
PerL CDASILLDNT TSFRTEKDAF GNANSARGFP VIDRVKAAVE
Per dari G. max (L78163) CDGSVLLNNT DTIESEQDAL PNINSIRGLD VVNDIKTAVE Per dari A. thaliana (X71794) CDASILLDNT TSFRTEKDRF GNANSARGFP VIDRMKAAVE
....|....| ....|....| ....|....| ....|....|
125 135 145 155
PerL RACPRTVSCA DMLTIAAQQS DTLAGGPSWR VPLGRRDSLQ
Per dari G. max (L78163) NSCPDTVSCA DILAIAAEIA SVLGGGPGWP VPLGRRDSLT Per dari A. thaliana (X71794) RACPRTVSCA DMLTIAAQQS VTLAGGPSWR VPLGRRDSLQ
....|....| ....|....| ....|....| ....|....|
165 175 185 195
PerL AFLELANANL PAPFFTLPQL KASFRNVGLD RPSDLVALSG
Per dari G. max (L78163) ANRTLANQNL PAPFFNLTQL KASFAVQGLN -TLDLVTLSG Per dari A. thaliana (X71794) AFLELANANL PAPFFTLPQL KASFRNVGLD RPSDLVALSG
....|....| ....|....| ....|....| ....|....|
205 215 225 235
PerL GHTFGKNQCQ FILDRLYNFS NTGLPDPTLN TTYLQTLRGL
Per dari G. max (L78163) GHTFGRARCS TFINRLYNFS NTGNPDPTLN TTYLEVLRAR Per dari A. thaliana (X71794) GHTFGKNQCQ FILDRFYNFS NTGLPDPTLN TTYLQTLRGL
....|....| ....|....| ....|....| ....|....|
245 255 265 275
PerL CPLNGNRSAL VDFDLRTPTV FDNKYYVNLK ERKGLIQSDQ
Per dari G. max (L78163) CPQNATGDNL TNLDLSTPDQ FDNRYYSNLL QLNGLLQSDQ Per dari A. thaliana (X71794) CPLNGNRSAL VDFDLRTPTV FDNKYYVNLK ERKGLIQSDQ
....|....| ....|....| ....|....| ....|....|
285 295 305 315
PerL ELFSSPNATD TIPLVRAYAD GTQTFFNAFV EAMNRMGNIT
Per dari G. max (L78163) ELFSTP-GAD TIPIVNSFSS NQNTFFSNFR VSMIKMGNIG Per dari A. thaliana (X71794) ELFSSPNATD TIPLVRAYAD GTQTFFNAFV EAMNRMGNIT
....|....| ....|....| ....|....| ....|....|
325 335 345 355
PerL PTTGTQGQIR LNCRVVNSNS LLHDVVDIVD FVSSMS----
Per dari G. max (L78163) VLTGDEGEIR LQCNFVNGDS FGLASVASKD AKQKLVAQSK Per dari A. thaliana (X71794) PTTGTQGQIR LNCRVVNSNS LLHDVVDIVD FVSSM---