Volume
2Nomor
I,
Juni
2015Studi Variasi Tingkat Keanekaragaman Jenis Burung Pada Berbagai Tipe Habitat
DiAreal
Konservasi Perkebunan Sawit PT. MSM, Wilmar Plantation, KaltengBelinda Hastari dan M. Arief
9oendjoto...
1-l I Analisis Pola Agroforestri Pada Kebun PetaniFahruni
12-25Kajian Ekspansi Akasia
DI
Taman Wisata Alam Bukit TangkilingSiti Maimunoh
...
26-34Respon Pertumbuhan Kedelai (Glycine max (L.) Men.) Terhadap Cekaman Kekeringan
Pienyani
Rosowanti
35-44Analisis Kepuasan Petani Terhadap Kegiatan Penyuluhan Pertanian di Kelurahan Kalampangan, Kota Palangka Raya Kalimantan Tengah
Berkat dan Revi
Surwryati..
45-53Pemanfaatan Abu Boiler Kelapa Sawit Terhadap Pertumbuhan Tajuk Tanaman Tomat
Nurul Hidoyati dan Asro' Laelani
Indrayonti
54-65 Analisis Usaha Tani Jagung (Zea moys) di Desa Kuwolu Kecamatan BululawangKabupaten Malang
Pienyani Rosat+'anti, Respon Pertumbuhan Kedelai (Glycine mar (L) Merr.) Terhadap ...
RESPON
PERTUMBUHANKEDELAI
(Glycine mox
(L.\ Merr.)
TERHADAP
CEKAMAN KEKERINGA}I
(Groutth responses of soybean (Glycine mm (L.)
Men.)
to drought stress)Pienyani Rosawanti
Program Studi Agroteknologi Fakultas Pertanian dan Kehutanan Universitas Muhammadiyah Palangkaraya
e-mai I : pieny ani@yahoo.com
Abstract
Crop respotwes to drought stress depend on their gerctypes.The purpose of this research was
to
study the effectof
drought
stress thegrowth
chqractersof
soybean.research wasconducted
in
the
Cikabayan greenhouse-IPB using three genotypesof
soybean (Slamet, Tanggamusand
Wilis)
andPEG
(0%,
20%o). The drought stress wasapplied only
at
a vegetativephose. The result
showedthat the
interaction
between genotypeand
PEG, significant. effect on root-shoot lengthratio.
Drought stress vtith PEG;imulation
causedthe changesin
thegrowth
charactersof
soybean i.e. on shoot length, root-lenght, root-shoot length ratio, leaf dry weight and root dry weight.Key words :genotype, PEG, vegetative phase
Abstrak
Respon tanaman pada kondisi kekeringan terganfLlng pada genotipe.Tujuan penelitian
ini
untuk
mengetahui respon pertumbuhan tanaman kedelai terhadapkondisi
cekamankekeringan.Penelitian dilaksanakan
di
rumah kaca
CikabayanJPB menggunakan tiga genotipe kedelai(Slamet Tanggamus danWilis)
dan PEG (0%,20%). Cekaman kekeringan hanya diberikan pada fase vegetatif.Hasil
penelitian menunjukkan bahwa interaksi antara perlakuan genotipe dan PEG berpengaruh nyata terhadap rasio panjang akar-tajuk. Cekaman kekeringan dengan simulasi PEG menyebabkan perubahan karaher pertumbuhan tanaman kedelai yaitu padatinggi tajulq panjang akar, rasio panjang akar-tajul<, bobot kering daun dan bobot kering akar.Kata kunci: fase vegetatif genotipe, PEG
PENDAIIULUAN
Kedelai (Glycine
mat (L.)
Merr.) merupakan salah satu komoditas tanaman panganyang
penting dalam
mendukungketahanan
pangan
di
Indonesia.Permintaan
kedelai
meningkat
pesat seiring dengan laju pertambahan pendudukdan
meningkatnyakesadaran pentingnya nilai gizi bagi kesehatan.Tahun 2012-2014,rata-rata kebutuhan
kedelai
nasional sebesar 2.59 jutaton
per tahunsedangkanrata-rata
produksi
dan
produktivitas nasionalper
tahun,
berturut-turut hanyasebesar 800.0C
ton/ha
dan
1.5
ton/haJumal Qao4 Vol. 2
No.
1, Juni 2015:3544
(Bappenas 2014).
Hal ini
berarti terdapatkesenjangan
antara produksi
dengankebutuhan.Kondisi
tersebut
mendorong pemerintah untuk meningkatkan produksikedelai
dalam negeri
melalui
strategi peningkatanproduktivitas
dan
perluasan areal tanam terutama melalui pemanfaatanlahan-lahan
marginal
antara
lain
padalahan
kering.
Pemanfaatan lahan-lahanmarginal
di
Indonesia
terutama
lahankering perlu
ditingkatkan
untukpemenuhan
kebutuhan
bahan
panganterutama
kedelai.
Permasalahan yang terjadi pada pemanfaatan lahan keringini
adalah
ketersediaan
air
yang
bisa menyebabkankondisi
kekeringan
atau cekaman kekeringan.Cekaman kekeringan pada tanaman disebabkan karena kurangnya suplai
air
didaerah perakaran atau permintaan air yang
berlebihan
oleh
daun
karena
laju evapotanspirasi melebihilaju
absorbsi airoleh
akar
tanaman, walaupunair
tanahdalam
keadaan
cukup
(Levitt
1980).Ketersediaan
air
yang cukup
sangatdiperlukan
untuk
semua
proses metabolisme dalam tanaman. Pertumbuhan dan perkembangan sel-sel tanaman sangat ditentukan oleh ketersediaan air (Fitter danHay
1981; Gardneret
al.
L99l; Taiz
danZeiger
2002).
Dikemukakan
olehKozlowski
1968, bahwa ketersediaan air dalam tanah merupakan faktor yang sangatpenting dalam proses-proses
fisiologi
padatumbuhan
seperti
perkecambahan benih, penyerapan dan translokasi unsur hara danasimilat
transpirasi
serta
fotosintesis. Rendahnya kandunganair
tanah
dapat membatasi penyerapanunsur
hara
oleh akar tanaman (Marschner 1995).Cekamankekeringan
pada
tanaman
dapat menyebabkan turunnya potensialair
danrendahnya tekanan
turgor.
Pertumbuhan sel, fotosintesis dan produktivitas tanaman sangat dipengaruhi oleh potensial air(Taiz
dan
Ze\ger2002).
Tekananturgor
yangtinggi
dibutuhkan untuk pemanjangan sel,sehingga
adanya cekaman
air
akanmenghambat
pertumbuhan
danperkembangan
sel
tanaman.
Laju pertumbuhan sel-sel tanaman dan efisiensiproses fisiologisnya
mencapai
tingkattertinggi
bila
sel-sel berada pada turgormaksimum
(
Fitter
dan
Hay
1981). Kemampuantanamanuntuk
beradaptasiterhadapcekaman
kekeringantergantungpadaintensitas
danperiode
cekaman, fasepertumbuhandan
genotipe
tanaman(Kalefetoglu dan Ekmekci
2005).Respontanaman terhadap cekaman
kekeringanberbeda-beda
tergantung
pada
lama, intensitas cekaman, spesies tanaman dantahap
pertumbuhantanaman
(Kusvuran 7012).Simulasi
lingkungan
yangmengalami
cekaman kekeringan
dapat#
ft
d
!
PienT'oni Rosawanti, Respon Pertumbuhan Kedelai (Glycine mcu (L) Men.) Terhadap ...
dilakukan
dengan perlakuan
pemberianPEG
(poly-ethyleneglycol).
PenggunaanPEG yang dilarutkan dalam
air
dapatdigunakan
untuk
meniru
besarnya potensial air (Michel dan Kaufmann 1973). Beberapa percobaan dengan menggunakan PEG pada beberapa genotipe kedelai telahbanyak dilakukan
(Sunaryo2002),
juga pada tanamanlain
seperti pada beberapa genotipe kacang tanah lSusilawati 2003). Tujuan percobaanini
adalah untuk melihat responpertumbuhanbeberapa
genotipekedelai
yang
mengalami
cekaman kekeringan pada fase vegetatif.METODOLOGI
Bahan
yang
digunakan
adalah
3genotipe kedelai
yaitu:
Slarnet,Tanggamusdan
Wilis,
PEG6000,
aquadest, pupuk urea, pupuk SPl8
dan pupukKCl,
pasir dan tanah. Sedangkan alat yang digunakanadalah polibag,
meteran,
timbangananalitih
gelas
ukur dan
oven.Penelitianinidilakukan
di
Rumah Kaca
CikabayanIPBdengan menggunakan
RancanganAcak
Kelompok
Lengkap(RAKL)
yang disusun secarafaktorial
dengan2
faktor. Faktor pertama adalah3
genotipe kedelaiyaituSlamet
Tanggamus dan Wilis.Faktor kedua adalahkonsentrasi PEG yang terdiridari
Uohdan
20o/oyang
masing-masing setara dengan potensial osmotik 0 Mpa dan-0,67 Mpa(Mexal 1975).Perlakuan diulang sebanyak lima kali.
Penelitian
ini
dilakukan
densan menggunakan polibag berdiameter20
cmyang
diisi
campuran
tanah
dan
pasir denganperbandingan2:
I
sebanyak 8 kg. Setiap polibag ditanam 2 benih kedelai danpada
umur 3
minggu
dilakukanpenjarangan dengan
meninggalkan
satu tanaman per polibag yang pertumbuhannya paling baik. sebelum penanaman dilakukan pemupukan dasar dengan menggunakanUreq
SP-18 danKCl.
Pemupukan keduadilakukan
setelah tanaman
berumur
3minggu.Penyiangan
dilakukan
secaraberkala
dengan
mencabut
tanamanpengganggu
yang
t'.rmbuh
di
dalampolibag.
Pemeliharaandilakukan
terus menems hingga tanaman dipanen.Pemberian
Perlakuan
CekamanKekeringan
Larutan
PEG
dibuat
denganmelarutkan
kristal PEG 6000
sesuai konsentrasi perlakuan denganair
sampaivolume
mencapaiI
liter.
Larutan
PEGdiberikan pada
tanamansejak
tanamanmemiliki daun
trifoliat
yang
telah berkembang sempurna sebanyak20
ml setiap2
hari sekali sampai tanaman mulai memasukifase reproduktif
(+
umur
30 hari).Jurnal
QanqYoL2
No.
1, Juni2015:3544
Pengamatan
(dilakukan
pada
umur
3l
HST/setelah
pemberian
perlakuan cekaman kekeringan)I
Tinggi
tajuk (cm).
Diukur
daripangkal batang
sampai
pucuk tanaman.2
Panjang
akar (cm).
Diukur
daripangkal
akar
sampai
ujung
akar dengancara
meluruskanakar
yang bergulung.3
Bobot kering daun
(g).
Daun dimasukkanke
dalam oven
dengansuhu
TOoC selama48
jam
sampaiberatnya konstan,
lalu
ditimbang dengan timbangan analitik.4
Bobot
kering
akar
(g).
Akardipisahkan
dari
tajuknya
kemudiandicuci
sampai bersih dan dimasukkanke
dalam oven
dengansuhu
70oCselama
48
jam
sampai
beratnyakonstan,
lalu
ditimbang
dengan timbangan analitik.5
Rasio bobot
kering akar dan
daun.Dihitung
dengan
rumus
sebagaiberikut: rasio bobot kering akar-daun
:
bobot kering akar/bobot keringdaun.6
Rasio panjang akar dan panjang tajuk.Dihitung
dengan
rumus
sebagaiberikut: rasio
panjang akar-tajuk
:
panj ang akarlpanjang taj uk. Analisis Data
Analisis
data
dilakukan
dengan analisis ragam, apabila berpengaruh nyata akan dilakukan analisis lanjutan denganuji
jarak
berganda
atau
DMRT
(Duncan Multiple Range Test)padatanf
5YuIIASIL
DANPEMBAHASAN
Hasil
analisis
sidik
ragammenunjukkan
bahwa PEG
berpengaruh terhadap karakter panjangtajulq
panjang akar primer, bobot kering daun dan bobotkering
akar..Interaksi
terdapat
padakarakter
rasio
panjangakar-tajuk
(Tabell).
Tabel
1
Rekapitulasi sidik ragam karakter3genotipekedelai terhadap perlakuan PEGKarakter Genotipe
(G)
PEG(P)
Interaksi(GxP)Panjang tajuk (cm) Panjang akar primer (cm) Rasio panj ang akar-taj uk Bobot kering daun (g) Bobot kering akar (g)
Rasio bobot kering akar-daun
tn * * tn tn tn tn tn tn tn + *,1. tn tn
**
tn tn tnKeterangan: tn: tidak berbeda,nyata padataraf 5%o. ** : berbeda nyata pada taraf 5%o, +: berbeda nyata pada taraf lo/o.
Pienyani Rosowanti, Respon Pertumbuhan Kedelai (Glycine mar (L) Merr.) Terhadap ...
Panjang
Tajulq
PanjangAkar Primer
dan Rasio PanjangAkar-Tajuk
Cekaman kekeringan menyebabkan penghambatan pembelahan, pembesaran
dan
pemanjangansel
akibat
rendahnyakadar
air
yang
tersedia.
Cekamankekeringan
menyebabkan
menurunnya potensial air tanaman akibat berkurangnyadifusi
air
dari
larutan
tanah
kedalam tanaman sehingga menurunkan turgor sel. Menurunnya tekan turgor sel tanaman jugamenyebabkan terhambatnya
proses pembelahan dan pembesaran sel (Taiz dan T.eiger2002)
sehingga tanaman menjadipendek.Beberapa
penelitian
dengankonsentrasi
PEG yang
bervariasi menunjukkan respon penurunan tajuk yang berbeda-beda(Ashri
2000; Sunaryo 2002;Widoretno 2002) pada
genotipe kedelaiyang
peka
cekaman
kekeringan.Hasil penelitianlain
yang menunjukkan respon penurunantinggi tajuk
akibat
cekamankekeringan ditunjukkan
oleh
tanaman kacangmoth (Soni
et
al.
2011), rumputRaja
(Sinaga
2005)
dan
kacang
tanah(Riduan
et al.
20A5;
Susilawati 2003).Cekamankekeringan
juga
dapatmenghambat
proses
pembelahan
danpembesaran
sel
pada
akar,
akibatnya pertumbuhan akar menjadi terhambat.Tiap
genotipe
memiliki
responpanjang
tajukdan
panjang
akar
primeryang
berbeda
ketika
menghadapi perlaktran cekaman kekeringan (Tabel 2).PEG
0%
mempunyai
respon
tinggitajuktertinggi
(96.044cm)
dan
berbeda nyata dengan PEG 2,0o/o, tajuk mengalami penurunanterhadap kontrol sebesar25.08%o.Pada panjang
akar
primer,
PEG
lYo mempunyai respontertinggi
(15.156 cm) dan berbeda nyata dengan PEG ZAYo.Akarprimer
mengalami
penurunanterhadap kontrol sebesar4O.90%.Tabel2
Pengaruh konsentrasi PEG 6000 terhadap peubah rasio panjangtajuk (cm)
dan panjang akar primer (cm)Perlakuan Panjang tajuk
(cm)
Panjang akar primer (cm) PEG O% PEG 20%96.044
a71.956
b15.156
a8.9s6
b Penurunan relatif terhadap kontrol(%)
25.08 40.90Keterangan: Angka-angka yang diikuti oleh hurulyang sama pada kolom yang sama, tidak berbeda nyata pada
taraf uji 5 9/o (DIr4RT).
Akar
merupakan organ penting pada tanaman terutama untuk menyerapair
dan unsur hara pada raedia tanam. Pada saatkekeringan
dapat
terjadi
perubahananatomi
dan
fisiologi
pada
tanaman terutama padaakar
(Fentaet
al.
2014).Jurnal @ao4 Y ol. 2
No.
l,
Juni 2015 : 35-44Tanaman
lebih
banyak mengembangkansistem
perakaran
dalam
menanggapikekurangan
unsur hara
dan kekeringan(L;rnchdan Brown 2012). Rasiopanjang
akartajuk
dipengaruhi
secaranyata
oleh
genotipe dan PEG. Pada saatcekaman kekeringan, penurunan
rasio panjangakartajuk
terjadi
pada genotipe Tanggamusdan
Wilis
yang
mengalamipenurunan
masing-masing
sebesar 49.61yo, dan 26.56Yo, kecuali pada Slametyang
mengalami peningkatan
rasio panjang akar-tajuk sebesar 37.70% (Tabel3). Hal ini
mengindikasikan
bahwagenotipe
Slamet
mengalami
perubahanrespon terhadap cekaman
kekeringan dengan memperpanjang akar dibandingkan dengan tajuk.Perubahanfisiologi
tersebut dikarenakan akar terus berkembang untuk memperluas daerah penyerapan air.Hasil pada penelitianlain
menunjukkan adanya peningkatan rasio akar-tajuk pada kedelai (Makbul etaI.2011)
dan genotipe kacangtanah yang toleran
terhadap
cekaman kekeringan (Susilawati 2003).Tabel
3
lnteraksi genotipe dankonsentrasi PEG terhadap rasio panjang akar-tajuk kedelaiGenotipe PEG (%) Penurunan
relatif
terhadap
kontrol
(%) 20 Slamet TanggamusWilis
0.12 aA0.lg
aA 0.16 aA 0.17 aA0.i0
aA 0.12 aA37.70
+ 49.61 26.56Keterangan: Angka-angka yang diikuti huruf kecil pada kolom yang sama dan huruf besar pada baris yang sama menunjukkan hasil yang tidak berbeda ni'ata berdasarkan DMRT pada c
:
5Yo; tanda (+) menunjukkan adanya peningkatan.Bobot
Kering
Daun, BobotKering
Akar
dan Rasio BobotKering Akar-Daun
Bobot kering
merupakan
hasil akumulasi fotosintesis tumbuhan selamapertumbuhannya
(Levitt
1980).Cekamankekeringan
pada"
percobaan
ini
menyebabkanpenurunan
bobot
keringdaun
sebesar72.33%(Tabel
4).
Bobot kering daun dipengaruhi, secara nyata olehgenotipe
dan
PEG.
Penurunan
bobotkering daun
diduga
terkait
denganpenurunan
laju
'fotosintesis
selamacekaman kekeringan. Hasil pada penelitian lain menunjukkan penurunan bobot kering
daun
pada
genotipe
kedelai
yangmengalami
cekamankekeringan
(Ashri2000;
Sunaryo
2002),
kacang
tanah(Susilawati
2003)
dan
kacang
hijau (Ranawake etal.201l).
Cekaman kekeringan pada percobaan
ini
menyebabkan penumnan bobot keringakar
sebesarT2. llo/o (Tabel 4).Penurunanbobot kering akar
didugaterkait
denganpenurunan
laju
fotosintesis
selamaPienyani Rosavanti, Respon Pertumbuhan Kedelai (Gly'cine max
(L)
Merr.) Terhadap ...cekaman kekeringan. Hasil pada penelitian lain menunjukkan penurunan bobot kering
akar
pada
kedelai
yang
mengalami cekaman kekeringan (Ashri 2000; Sunaryo 2003) dan kacanghijau
(Ranawake et al. 2011).Tanaman
kedelai
yang
mengalami kekeringan pada fase vegetatif mengalami penurunanpertumbuhan
danperkembangan
yang
sangat
besar(Aboyami 2008),
seperti dengan adanya penurunantinggi
tanaman,jumlah
nodus, panjang akar, bobot kering akar dan tajuk(Riduan 2005).
Ini
merupakan strategikonservatif
untuk
mempertahankan penggunaan energi, dimana tanaman akanmengembangkan
respon ,vang
lebihkomplek
untuk
perbaikan
toleransiterhadap
periode
kekuranganair
lebih lanjut(Kron etal.
2008). Pada saat terjadi cekaman kekeringan, tanamanlebih banyak mengembangkansistem perakaran(Lynch 2007). Sel-sel akar mengalami perubahanantara
lain
dengan
meningkatkan atau mengurangijumlah
maupun ukuran dalammenghadapi
cekaman
kekeringan.Kekeringan
pada fase
vegetatif menyebabkan terjadinya penurunan hasil produksi tanaman kedelai (Kisrnan 2010), sehingga diperlukan genotipe kedelai yangtoleran
kekeringandan
mempunyai dayahasil yang tinggi.
Tabel4
Pengaruh konsentrasi PEG 6000 terhadap peubah bobot kering 'rajuk(g),
bobot kering akar (g) dan rasio bobot kering akar-tajukPerlakuan
Bobot kering
daun
Bobot kering akar(g)
(e)Rasio bobot kering akar-daun PEG OOA PEG 20% 1.312 0.363 0.1 04 0.029 0.079 0.085 a b a b tn tn Penurunan
relatif
terhadap kontrol (%) 7.59Keterangan: Angka-angka yang diikuti oleh hur.rf yang sama pada kolom yang sama- tidak berbeda nyata pada
tarafuji5%(DMRT).
72.33 72,t1
Pada
rasio
bobotkering
akar-daun, perlakuan genotipe dan PEG tidak berbedanyata tetapi
cenderung
terjadipeningkatanrasio
bobot kering
akar-daunsebesar7.59o/o
(Tabel
4).
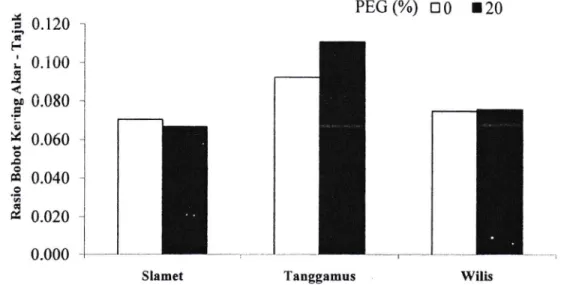
Padagambar
lterdapat
pola
respon
yangcenderung meningkat
pada
genotipeTanggamus
dan
Wilis
kecuali
padagenotipe Slamet yang mempunyai respon cenderung menurun.
41
l*-Jurnal
@ao4Yol.2 No.
l,
Juni 2015:3544
6 F I al,,
E! ,i ov
o€
o tr o ql &PEG(%)
trO
.2a
0.1 20
0.i00
0.080 0.060 0.040 0.020 0.000Slamet
Tanggamus
WilisGambar 1 Rasio bobot kering akar-tajuk pada perlakuan PEG 0o/o dan2lo/o.
Tiap genotipe
memiliki
respon rasiobobot kering akardaun yang
berbeda ketika menghadapi cekaman kekeringan.Peningkatan rasio bobot akar-daun pada genotipe Tanggamus dan Wilis menunjukkan bahwa tanaman melakukanmekanisme
toleransi dengan
carameningkatkan
perkembangan
danpertumbuhan
akar
dibandingkan denganpertumbuhan
dan
perkembangan daun. Sistem perakaran ditingkatkan agar lebihefisien dan etbktif
dalam
memperbaiki serapanair.
Hasil
pada penelitian
lain menunjukkanadanya
peningkatan rasiobobot kering
akar-daunpada
genotipekedelai
yang
mengalami
cekaman kekeringan (Sunaryo 2002; Hanumet
al. 2007;Makbul
et
al.20ll)
dan tanamankacang
moth (Soni
etal. 20Ll),
jagung (Farsiani dan Ghobadi 2009) dan rumput Raja (Sinaga2005).'SIMPT]LAI\{
Interaksi
antara perlakuan genotipe dan PEG berpengaruh nyata tcrhadap rasio panjang akar-tajuk. Cekaman kekeringandengan simulasi
PEG
menyebabkan perubahan karakter perfumbuhan tanamankedelai
yaitu
padatinggi tajulq
panjangakar,
rasio
panjang akar-tajuk,
bobot kering daun dan bobot kering akar.Genotipe
Tanggamus
dan
Wilis
mempunyai
respon
yang
sama
dalam menghadapi cekaman kekeringan terutama pada karakter rasio panjang akar-tajuk dan rasio bobot kering akar-daun.Pierr.v'ani Roscnvanti, Respon Perlumbuhan Kedelai (Glycirw ntru
(L)
Merr.) Terhadap ...DAFTAR PUSTAKA
AboyamiYA.
2008.Comparative growthand grain
yield
responsesof
early and late soybeanmaf.rrit,
group toinCuced
soil
moisture stress
atdifferent
growth
srages.W'orldJ
Agric
Scr.4(1):71-78.Ashri
K. 2006.
Akumulasi
enzim antioksidan dan prolin pada beberapavarietas kedelai
toleran dan
peka cekamankekeringan
[tesis]. Bogor(ID):
Institut Pertanian Bogor.[Bappenas]
Badan
PerencanaanPembangunan
Nasional.
'2014.Rencana
pembangunan
jangka menengah nasionalAPIMI\|)
bidangpangan
dan
pertanian
2015-2019.Direktorat
Pangan
dan
Peftanian,Bappenas.
[Intemet].
413
hlrn;[diunduh
2014
September
22).Tersedia pada:
http://www.baopenas. eo. id/fi les/3 7 I
3 / 93 4 6 I 927 | IRPJMN_B idang_Panga n_dan_Pertanian__20 I 5-20 I 9.pdf. Farsiani
A,
Ghobadi ME (2009). Effectsof
PEG and
NaCl
stress
on
twocultivars
of
corn (Zeo moysl.)
atgermination
and
early
seedling stages. WorldAcad.
Sci.Eng.
Tech.57:382-385.
Fenta
BA,
Beebe SE, Kunert KJ, BurridgeJD, Barlow
KM,
Lynch
JP,
FoyerCH.
2014.
Field
phenotyping
of
soybean
roots
for
droughtstresstolerance. Agronomy.
4:418435.
doi: I 0.3390/agronomy403 04 I 8.
Fitter
AH,
Hay
RKM.
1981. Fisiologi
Lingkungan Tanaman. Andayani S,Purbayanti
ED,
penerjemah;Srigandono
B,
editor.Yogyakarta(ID):
UGM
PR.
Terjemahan dari: Enviromental P hysiologt of P lants. Gardner FP, PearceRB
andMitchell
RL.1991.
Fisiologi
Tanaman Budidaya.Susilo
H,
Subiyanto,
penerjemah. Jakarta(ID): UI
Pr. Terjemahan dari: PkYsiolog,' of CroPPlants-Hanum
C.
Mugnisjah
WQ,
Yahya
S.Sopandy
D.
Idris
K.
SaharA.
2007.Pertumbuhan
akar
kedelai
padacekaman
aluminium,
kekeringan dancekaman gandaaluminium
dan kekeringan. Agrirrop. 26(l): I 3-1 8.Kaleletoglu
T.
Ekmekci
Y.
2005.
Theeffbcts
of
drought
on
plants
andtolerantmechanisms.
J
Sci.t8(1):773-740.
Kisman. 2010. Karakter morfologi sebagai penciri adaptasi kedelai
terhadapcekaman
kekeringan.A gr o t e ks o s. 20 (1'1:23 -29 .
Kozlowski. 1968. Water DeJicits and Plant
Growth.
Ed
ke-3.
New
York(US): Academic Pr.Kron AP,
SouzaGM,
RibeiroRV.
2008.Water
deficiency
at
different developmental stagesof glycine maccan
improve drought
tolerance.B ragantia C ampinas. l(67):43-49 .
Kubis,WieczoreJF,
Jelonek
MA.
2014.Polyamines
induce
adaptive responsesin
water
deficit
stressedcucumber
roots.
J
Plant
Res.127:l5l-158.
Kusvuran,
S.
2012. Influenceof
drought stresson
growth,ion
accumulationand
anti-oxidative enzymes
inokragenotypes.
International JAgricB iol. 1 4:401406.
Lestari
EG.
2005.
Hubungan
antara kerapatan stomata dengan ketahanankekeringan
pada
somaklon
padi Gajahmungkur,Towuti
dan
IR
&.
B iodive rsitas. 7
(l):4448.
Levitt
J.
1980. Responsesof
Plants
toEwironmental
,Srresses.Ed
ke-2. NewYorkOS):
Academic Pr.Lynch
JP,Brown
KM.
2012.New
rootsfor
agriculture:exploiting
the
root phenome.Phil
Trans R SocB.
367:1
598-l
604.Makbul S, Guler NS, Durmus N, Guven S.
2011.
Changesin
anatomical and physiological parametersof
soybeanunder drought
stress.Turk
J
Bot. 35:369-377.Marschner
H.
1995.Mineral Nutrition
of
Higher Plants.
Ed
ke-Z.
London (GB): Academic Pr.Jurnal
@atq
Voi.I
No.
i,
Juni 201 5 :3544
tv{attjik
AA,
Sumertajaya
[Ivi.
2002.Perancangon Percobaan
dengonAplikasi
SASdan Minitab..
Bogor (ID): IPB Pr.Mexal
J,
FisherJT,
OsteryoungJ,
ReidCPP.
1975. Oxygenavailability
in polyethyleneglycol
solution and its implicationsin
plant-water relation. Plant P lrys iol. 5 5 :20-24.Michel
BE,
Kaufmann
MR.
1973. Theosmotic potential
of
polyethyleneglycol
6000.Plant
Physiol. 5 I:914-916.
Ranawake
AL,
Amarasingha
UGS,Rodrigo
WDzu,
Rodrigo
UTD, Dahanayaka N. 201l.
Effect of water stress on growth andyield
of
mung bean(Yigru
radiateL)
Trop AgricRes Extens ion. | 4(4): 7 6-7 9
fuduan
A,
Aswidinnoor
H,
Koswara J,Sudarsono. 2005. Toleransi sejumlah
kultivar
kacang
tanah
terhadap cekaman kekeringan.Hayati.
l2(l):
28-34.
Sinaga
R.
2005.
Tanggap
morfologi, arratomi danfisiologi
rumput gajahdan
rumput
raja
akibat
penurunan ketersediaanair
tanah [tesis]. Bogor(tD):
Institut Pertanian Bogor. Soni P. RizwanM,
BhaffKV,
MohapatraT,
SinghG.
201l. In
vitro
responseof
Vigno aconitifolio
to
drought stress induced by PEG 6000. -rS/ress Physiol Biochem. 7(3): I 08-1 21, Sunaryo W. 2002. Regenerasi dan evaluasivariasi somaklonal kedelai (Glycine
max
(L.)
Merrill) hasil
kulturjaringan
serta
seleksi
terhadap cekaman kekeringan menggunakan simulasi Poly Ethylene Glycol (PEG) [tesis]. Bogor(lD):
Institut Pertanian Bogor.Susilawati
PN.
2003. Respon 16kultivar
kacang
tanah unggul
nasional(Arachis hypogaea
L.)
terhadapkondisi
stress kekeringan
akibatperlakuan penyiraman
PEG
6000 dan evaluasi daya regenerasi embrio somatiknya secarain
vitro
[tesis]. Bogor: lnstitut Pertanian Bogor. TaizL,
ZeigerE.
2002. Plant Plrysiologt.Third Edition. Massachussetts 1US): Sinaue Associaties lnc. Publisher.
Widoretno
W,
Guhardja
E,
Ilyas
S,Sudarsono.
2002.
Efektivitaspolyetlrylene
glycol
untukmengevaluasi tanggapan
ernotipekedelai terhadap
cekamankekeringan pada
fase perkecambahan.Hayati.
9(2):33 -36.