TINJAUAN PUSTAKA
Keratin
Keratin adalah produk pengerasan jaringan epidermal tubuh tersusun dari protein serat yang kaya akan sulfur. Keratin banyak ditemukan pada rambut, kuku, bulu, dan semua produk epidermal. Rantai keratin dikemas dengan kuat dalam bentuk α-heliks (α-keratin) atau β-sheet (β-keratin) menjadi rantai polipeptida superkoil (Parry & North 1998). Banyaknya ikatan disulfida (S-S), ikatan hidrogen dan interaksi hidrofobik pada struktur keratin menyebabkan protein keratin sangat stabil, kaku, dan tidak dapat didegradasi oleh enzim proteolitik yang umum seperti tripsin, pepsin, dan papain (Riffel et al. 2003a). Kandungan sistein pada keratin berkisar 8% dan tidak dimiliki oleh jenis protein lainnya. Jembatan sistein adalah struktur penting keratin dan merupakan penghambat kerja enzin proteolitik dalam memecah keratin (Presland et al. 1989).
Bulu ayam mengandung protein keratin dengan struktur α-helik, material lain yang kaya akan protein α-keratin adalah rambut, wool, sayap, kuku, cakar, duri, sisik, tanduk, kulit penyu, dan lapisan kulit sebelah luar, sedangkan material yang kaya dengan protein β-keratin adalah sutera, bulu, dan jaring laba-laba (Lehninger 1982). Berdasarkan tingkat kemudahan hidrolisis, keratin digolongkan menjadi soft keratin dan hard keratin. Kuku, sisik, bulu, atau wool lebih mudah dihidrolisis dibanding rambut manusia, kemudahan tersebut berkaitan dengan kandungan sistinnya (Kunert 2000).
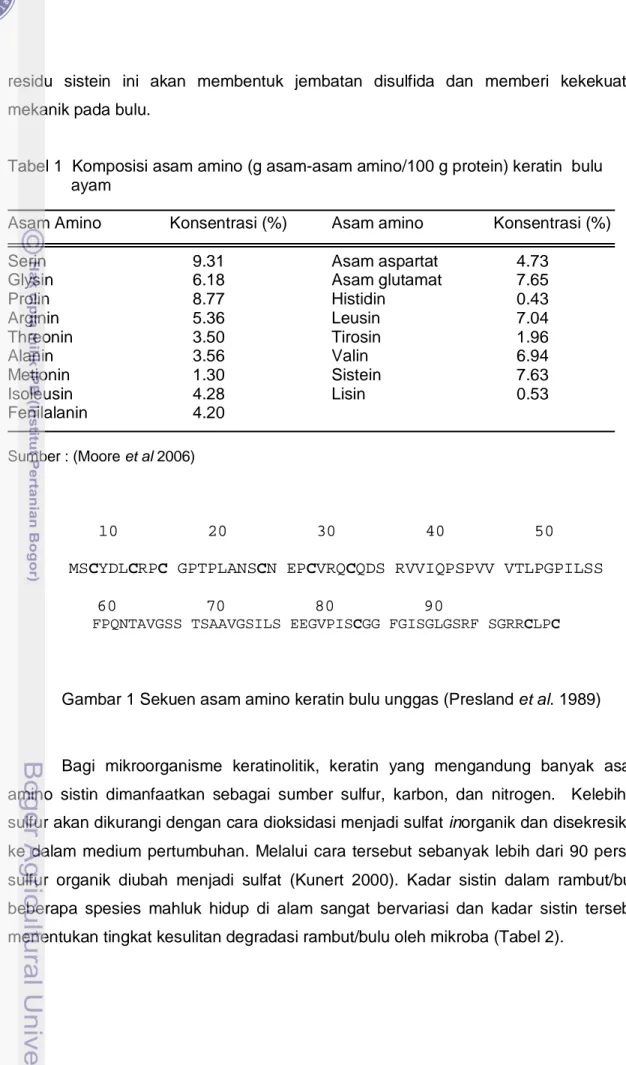
Komposisi kimia bulu ayam adalah 81% protein, 1.2% lemak, 86% bahan kering, dan 1.3% abu (Zerdani et al. 2004), selain itu bulu ayam mengandung mineral kalsium 0.19%, fosfor 0.04%, kalium 0.15%, dan sodium 0.15% (Kim & Patterson 2000). Kandungan asam amino utama pada bulu ayam adalah serin, prolin, glisin, sistein, asam glutamat, leusin, dan valin namun bulu ayam rendah kandungan asam amino histidin, lisin dan metionin (Tabel 1). Bertsch and Coello (2005) melaporkan bahwa tepung bulu ayam yang telah difermentasi oleh Kocuria rosea mengalami peningkatan kadar asam amino lisin, histidin dan metionin bila dibandingkan tepung bulu ayam komersial. Di dalam deret asam amino keratin bulu ayam, terdapat sembilan asam amino sistein (C) dari total 98 residu asam amino (Gambar 1) dan
residu sistein ini akan membentuk jembatan disulfida dan memberi kekekuatan mekanik pada bulu.
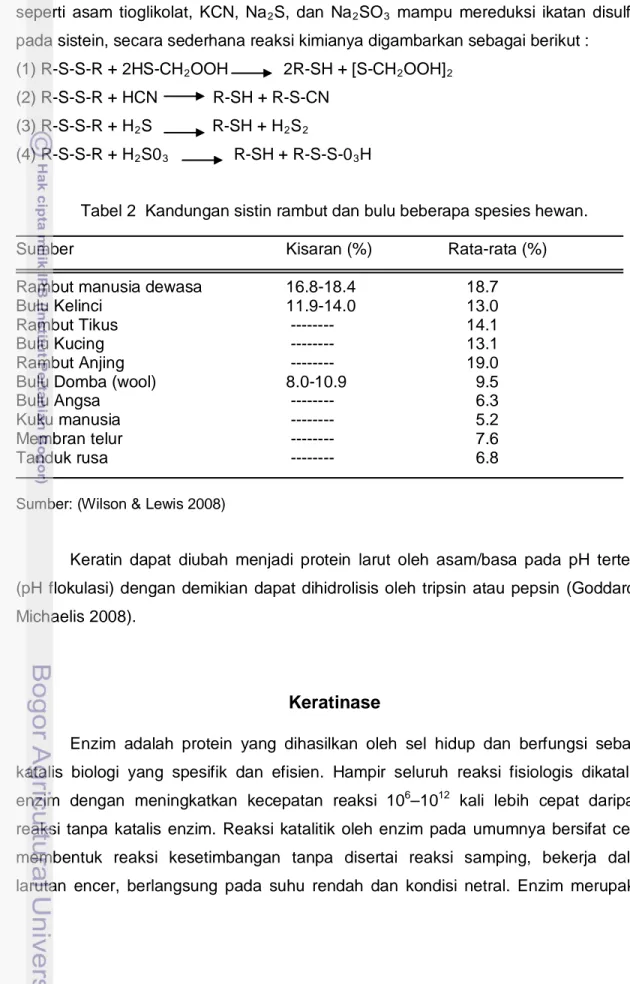
Tabel 1 Komposisi asam amino (g asam-asam amino/100 g protein) keratin bulu ayam
Asam Amino Konsentrasi (%) Asam amino Konsentrasi (%)
Serin 9.31 Asam aspartat 4.73
Glysin 6.18 Asam glutamat 7.65
Prolin 8.77 Histidin 0.43 Arginin 5.36 Leusin 7.04 Threonin 3.50 Tirosin 1.96 Alanin 3.56 Valin 6.94 Metionin 1.30 Sistein 7.63 Isoleusin 4.28 Lisin 0.53 Fenilalanin 4.20 Sumber : (Moore et al 2006) 10 20 30 40 50
MSCYDLCRPC GPTPLANSCN EPCVRQCQDS RVVIQPSPVV VTLPGPILSS
60 70 80 90
FPQNTAVGSS TSAAVGSILS EEGVPISCGG FGISGLGSRF SGRRCLPC
Gambar 1 Sekuen asam amino keratin bulu unggas (Presland et al. 1989)
Bagi mikroorganisme keratinolitik, keratin yang mengandung banyak asam amino sistin dimanfaatkan sebagai sumber sulfur, karbon, dan nitrogen. Kelebihan sulfur akan dikurangi dengan cara dioksidasi menjadi sulfat inorganik dan disekresikan ke dalam medium pertumbuhan. Melalui cara tersebut sebanyak lebih dari 90 persen sulfur organik diubah menjadi sulfat (Kunert 2000). Kadar sistin dalam rambut/bulu beberapa spesies mahluk hidup di alam sangat bervariasi dan kadar sistin tersebut menentukan tingkat kesulitan degradasi rambut/bulu oleh mikroba (Tabel 2).
Goddard dan Michaelis (2008) menyatakan bahwa beberapa senyawa kimia seperti asam tioglikolat, KCN, Na2S, dan Na2SO3
(1) R-S-S-R + 2HS-CH
mampu mereduksi ikatan disulfida pada sistein, secara sederhana reaksi kimianya digambarkan sebagai berikut :
2OOH 2R-SH + [S-CH2OOH] (2) R-S-S-R + HCN R-SH + R-S-CN 2 (3) R-S-S-R + H2S R-SH + H2S (4) R-S-S-R + H 2 2S03 R-SH + R-S-S-03
Sumber Kisaran (%) Rata-rata (%)
H
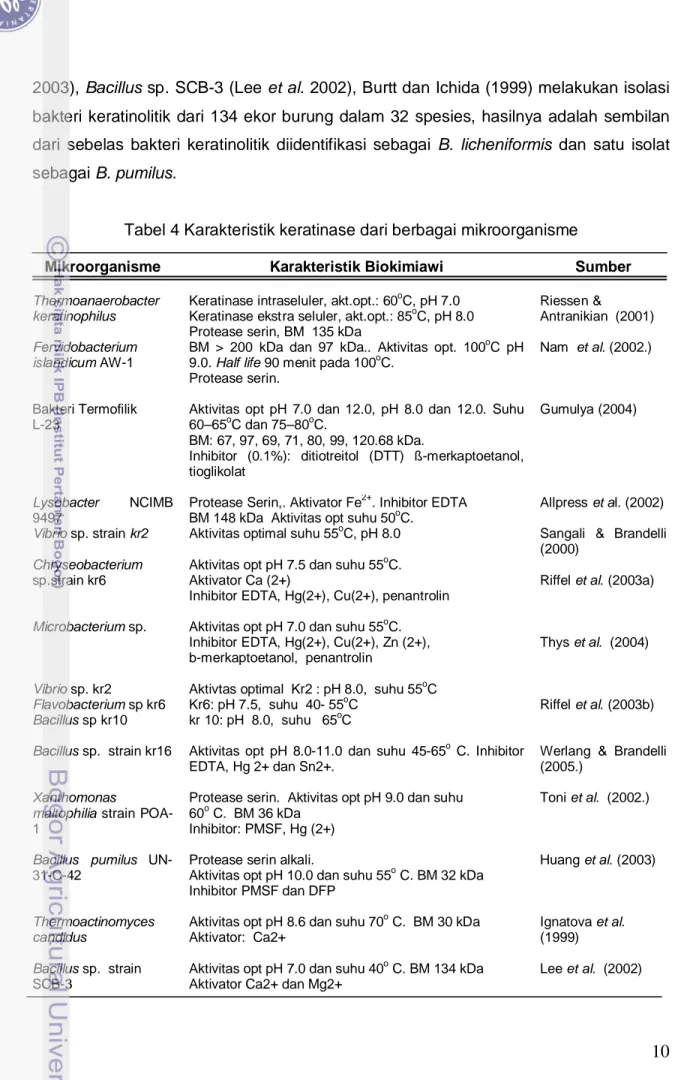
Tabel 2 Kandungan sistin rambut dan bulu beberapa spesies hewan.
Rambut manusia dewasa 16.8-18.4 18.7
Bulu Kelinci 11.9-14.0 13.0
Rambut Tikus --- 14.1
Bulu Kucing --- 13.1
Rambut Anjing --- 19.0
Bulu Domba (wool) 8.0-10.9 9.5
Bulu Angsa --- 6.3
Kuku manusia --- 5.2
Membran telur --- 7.6
Tanduk rusa --- 6.8
Sumber: (Wilson & Lewis 2008)
Keratin dapat diubah menjadi protein larut oleh asam/basa pada pH tertentu (pH flokulasi) dengan demikian dapat dihidrolisis oleh tripsin atau pepsin (Goddard & Michaelis 2008).
Keratinase
Enzim adalah protein yang dihasilkan oleh sel hidup dan berfungsi sebagai katalis biologi yang spesifik dan efisien. Hampir seluruh reaksi fisiologis dikatalisis enzim dengan meningkatkan kecepatan reaksi 106–1012 kali lebih cepat daripada reaksi tanpa katalis enzim. Reaksi katalitik oleh enzim pada umumnya bersifat cepat membentuk reaksi kesetimbangan tanpa disertai reaksi samping, bekerja dalam larutan encer, berlangsung pada suhu rendah dan kondisi netral. Enzim merupakan
protein globular yang terbentuk dari rantai polipeptida yang berlipat secara kompak. Konformasi tersier protein globular merupakan bentuk yang paling stabil karena ditunjang oleh berbagai ikatan yang menstabilkan struktur tersier protein. Jenis-jenis ikatan tersebut adalah: ikatan hidrogen yang terdapat di antara gugus R residu asam amino rantai samping yang berdekatan, ikatan ion di antara gugus R yang berlawanan, interaksi hidrofobik dari gugus R asam amino hidrofobik, dan ikatan kovalen berupa ikatan disulfida dari residu sistin (Copeland 2000; Creighton 1993).
Keratinase atau enzim keratinolitik adalah sebutan bagi enzim protease spesifik yang dapat memecah substrat protease keratin (EC 3.4.21 atau EC 3.4.24 atau EC 3.4.99). Enzim keratinase dihasilkan oleh mikroba baik secara intraseluler maupun ekstraseluler. Sebagian besar protease yang dihasilkan oleh mikroba tergolong protease serin yang memerlukan kofaktor kation Mg dan Ca untuk aktivitasnya dan sebagian lagi merupakan protease alkalin (Toni et al. 2002, Huang et al. 2003, Lee et
al. 2002). Keratinase Bacillus licheniformis PWD-1 adalah protease serin alkali yang
tersusun dari 379 residu asam amino (Gambar 2). Tidak ditemukan residu sistein (C) pada sekuen keratinase dari Bacillus sp. namun pada sekuen keratinase
Streptomyces sp.OWU dan Stenotrophomonas maltophilia ditemukan residu sistein
(Tabel 3).
1 MMRKKSFWLG MLTAFMLVFT MAFSDSASAA QPAKNVEKDY IVGFKSGVKT 51 ASVKKDIIKE SGGKVDKQFR IINAAKAKLD KEALKEVKND PDVAYVEEDH 101 VAHALAQTVP YGIPLIKADK VQAQGFKGAN VKVAVLDTGI QASHPDLNVV 151 GGASFVAGEA YNTDGNGHGT HVAGTVAALD NTTGVLGVAP SVSLYAVKVL 201 NSSGSGSYSG IVSGIEWATT NGMDVINMSL GGASGSTAMK QAVDNAYARG 251 VVVVAAAGNS GSSGNTNTIG YPAKYDSVIA VGAVDSNSNR ASFSSVGAEL 301 EVMAPGAGVY STYPTNTYAT LNGTSMVSPH VAGAAALILS KHPNLSASQV 351 RNRLSSTATY LGSSFYYGKG LINVEAAAQ
Gambar 2 Sekuen asam amino keratinase Bacillus licheniformis PWD-1 (Lin et
al. 1995)
Jenis asam amino pada struktur primer enzim menentukan karakteristik enzim tersebut. Shirai et al. (1997) melaporkan strain alkalifilik Bacillus sp. penghasil
protease serin yang disebut protease, protease ini memiliki pH optimal 12.3. M-protease memiliki jumlah asam amino bermuatan negatif (asam aspartat dan glutamat) dan residu lisin lebih sedikit, serta peningkatan arginin dan asam amino netral hidrofilik (histidin, asparagin, glutamin) lebih banyak dibanding asam amino pada protease subtilisin.
Tabel 3 Total asam amino dan sistein keratinase yang dihasilkan oleh beberapa bakteri
Bakteri Nama Gen ∑ asam amino ∑ Sistein No Akses GenBank
B. licheniformis kerA 379 - AAY82467.1
B. pumilus - 383 - ACM47735.1
B. subtilis - 354 - ABY65723.1
Streptomyces sp.OWU - 268 3 AAU94349.1
Stenotrophomonas kerD 634 5
Kedua residu asam aspartat (Asp/D) dan glutamat (Glu/E) bermuatan negatif, perbedaan keduanya hanya pada gugus metil yang dimiliki. Gugus karboksil pada rantai samping Asp dan Glu mengion pada pH 3.9-4.3 dan menjadi sangat polar. Sementara gugus amin pada rantai samping asparagin (Asn/N) dan glutamin (Gln/Q) tidak mengion dan tidak terlalu reaktif, namun bersifat polar, antara donor dan akseptor ikatan hidrogen. Gugus amin juga labil pada pH sangat ekstrem dan temperatur tinggi, dan residu ini dapat diaminasi menjadi residu Asp dan Glu. Residu lisin (lys/K) dan arginin (Arg/R) bersifat hidrofobik dan bermuatan positif karena adanya gugus amino pada rantai samping Lys mengion pada pH 11.1 dan Arg pH 12.0. Gugus amino pada rantai samping Lys berperan pada berbagai reaksi sehingga memungkinkan Lys berubah menjadi bermuatan positif, negatif, atau netral. Rantai samping imodazole pada residu histidin (His/H) mempunyai beberapa kelengkapan khusus yang membuat His sangat efektif sebagai katalis nukleofilik, yaitu gugus amin yang lebih reaktif dari ion hidroksida (Creighton 1993).
ACN82379.1
maltophilia
Keratinase dihasilkan oleh berbagai spesies basilus yaitu Bacillus FK 28 (Pissuwan & Suntomsuk 2001), Bacillus licheniformis L-25 dan PWD-1 (Lin et al. 2001), Bacillus sp. strain 16 (Werlang & Brandelli 2005), Bacillus pumilus (Huang et al.
2003), Bacillus sp. SCB-3 (Lee et al. 2002), Burtt dan Ichida (1999) melakukan isolasi bakteri keratinolitik dari 134 ekor burung dalam 32 spesies, hasilnya adalah sembilan dari sebelas bakteri keratinolitik diidentifikasi sebagai B. licheniformis dan satu isolat sebagai B. pumilus.
Tabel 4 Karakteristik keratinase dari berbagai mikroorganisme
Mikroorganisme Karakteristik Biokimiawi Sumber
Thermoanaerobacter keratinophilus
Keratinase intraseluler, akt.opt.: 60oC, pH 7.0 Keratinase ekstra seluler, akt.opt.: 85o
Riessen &
Antranikian (2001) C, pH 8.0
Protease serin, BM 135 kDa
Fervidobacterium islandicum AW-1
BM > 200 kDa dan 97 kDa.. Aktivitas opt. 100oC pH 9.0. Half life 90 menit pada 100o
Nam et al. (2002.) C.
Protease serin. Bakteri Termofilik
L-23
Aktivitas opt pH 7.0 dan 12.0, pH 8.0 dan 12.0. Suhu 60–65oC dan 75–80o
Gumulya (2004) C.
BM: 67, 97, 69, 71, 80, 99, 120.68 kDa.
Inhibitor (0.1%): ditiotreitol (DTT) ß-merkaptoetanol, tioglikolat
Lysobacter NCIMB 9497
Protease Serin,. Aktivator Fe2+. Inhibitor EDTA BM 148 kDa Aktivitas opt suhu 50o
Allpress et al. (2002) C.
Vibrio sp. strain kr2 Aktivitas optimal suhu 55o Sangali & Brandelli (2000)
C, pH 8.0
Chryseobacterium
sp.strain kr6
Aktivitas opt pH 7.5 dan suhu 55o
Riffel et al. (2003a) C.
Aktivator Ca (2+)
Inhibitor EDTA, Hg(2+), Cu(2+), penantrolin
Microbacterium sp. Aktivitas opt pH 7.0 dan suhu 55o
Thys et al. (2004) C.
Inhibitor EDTA, Hg(2+), Cu(2+), Zn (2+), b-merkaptoetanol, penantrolin
Vibrio sp. kr2
Flavobacterium sp kr6 Bacillus sp kr10
Aktivtas optimal Kr2 : pH 8.0, suhu 55oC Kr6: pH 7.5, suhu 40- 55oC
kr 10: pH 8.0, suhu 65o
Riffel et al. (2003b) C
Bacillus sp. strain kr16 Aktivitas opt pH 8.0-11.0 dan suhu 45-65o Werlang & Brandelli (2005.)
C. Inhibitor EDTA, Hg 2+ dan Sn2+.
Xanthomonas
maltophilia strain
POA-1
Protease serin. Aktivitas opt pH 9.0 dan suhu 60o
Toni et al. (2002.) C. BM 36 kDa
Inhibitor: PMSF, Hg (2+)
Bacillus pumilus
UN-31-C-42
Protease serin alkali.
Aktivitas opt pH 10.0 dan suhu 55o
Huang et al. (2003) C. BM 32 kDa
Inhibitor PMSF dan DFP
Thermoactinomyces candidus
Aktivitas opt pH 8.6 dan suhu 70o Ignatova et al.
(1999) C. BM 30 kDa
Aktivator: Ca2+
Bacillus sp. strain
SCB-3
Aktivitas opt pH 7.0 dan suhu 40o C. BM 134 kDa Lee et al. (2002) Aktivator Ca2+ dan Mg2+
Keratinase pada umumnya memiliki aktivitas optimal pada pH netral hingga alkali (pH 7.0 – 12). Beberapa spesies bakteri menghasilkan keratinase termostabil dengan aktivitas optimal pada kisaran suhu 60–80o
Disulfida Reduktase
C. Berat molekul keratinase yang dihasilkan mikroba sangat bervariasi, berkisar dari 30 kDa hingga lebih dari 200 kDa. Substrat yang banyak digunakan dalam berbagai pengujian keratinase adalah tepung bulu ayam dan bulu sapi dengan konsentrasi berkisar 0.1–10% (Tabel 4).
Keratinase yang dihasilkan oleh mikroba memiliki spesifisitas luas, keratinase mampu menghidrolisis berbagai protein larut misalnya kasein, gelatin, serum albumin, albumin telur, hemoglobin, mioglobin, dan protein yang tak larut seperti keratin, elastin, kolagen, fibrin, laminin, fibronektin (Letourneau et al. 1998).
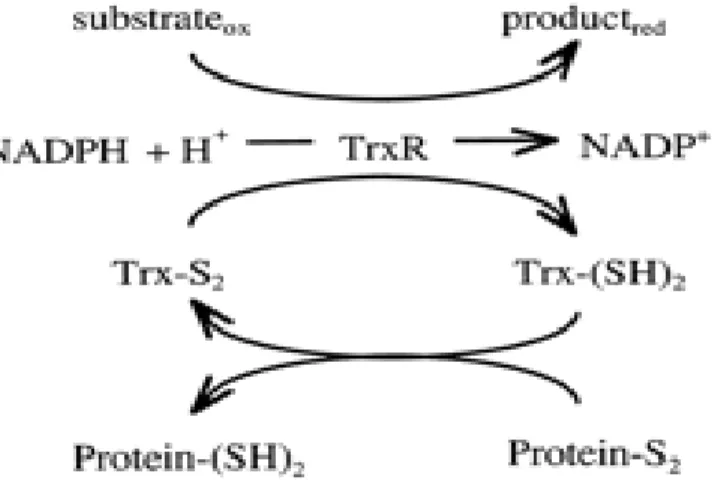
Thioredoxin reduktase (E.C. 1.6.4.5) dan glutathione reduktase (E.C. 1.6.4.2) merupakan dua jenis enzim yang aktif mengkatalisis reduksi ikatan disulfida dan keduanya termasuk golongan oksidoreduktase. Thioredoxin reduktase (TR atau TrxR) merupakan bagian dari sistem thioredoxin (Trx) yang berperan dalam berbagai fungsi biosintesis atau pelipatan protein. Sistem thioredoxin terdiri atas dua protein yaitu thioredoxin reduktase dan thioredoxin. Keduanya adalah protein redox-active dengan dua residu sistein yang terpisah oleh dua asam amino (glisin dan prolin) pada pusat katalitiknya (Rand & Grant 2006). Pada mikroorganisme, thioredoxin reduktase umumnya suatu dimer subunit identik dengan berat molekul 35 kDa, setiap subunit memiliki satu redox-active disulfida dan satu berikatan kuat dengan flavin adenin dinukleotida/FAD. Thioredoxin adalah protein ubikuitus dengan berat molekul 12 kDa dan nilai pI asam. Dua jenis aktivitas oksidoreduktase thioredoxin adalah: (a) sebagai pembawa elektron untuk proses-proses katalitik enzim-enzim biosintetik seperti ribonukleotida reduktase,(b) melindungi protein sel dari agregasi atau inaktivasi melalui pembentukan oksidatif disulfida (Arner & Holmgren 2000).
Motif pada sisi aktif Tiol-disulfida oxidoreduktase adalah Cys-Xxx-Yyy-Cyst dan dua residu sistein tersebut berperan dalam siklus disulfida teroksidasi dan diTiol tereduksi (reaksi redoks). Lokasi Tiol-disulfida oxidoreduktase, misalnya thioredoxin terlibat dalam sitoplasma dan terlibat dalam pemeliharaan lingkungan sitoplasma untuk
menjaga protein/asam amino dalam kondisi tereduksi. Mekanisme kerja thioredoxin tampak pada Gambar 3. Tiol-disulfida oxidoreduktase bakteri yang berada di luar membran sitoplasma terlibat dalam reaksi oksidasi, misalnya E. Coli DsbA, yang mengkatalisis pembentukan ikatan disulfida pada protein yang akan ditranspor melalui membran, reaksi isomerisasi ikatan disulfida misalnya E. Coli DsbC yang berfungsi dalam redistribusi ikatan disulfida di antara residu sistein protein target. ResA merupakan Tiol-disulfida oxidoreduktase yang terlibat dalam perakitan sitokrom c terletak pada sisi luar membran sitoplasma B. subtilis. ResA adalah protein terikat membran dengan domain C-terminal bermotif seperti thioredoxin yang terekspos pada sisi luar membran (Erlendsson et al. 2003).
Gambar 3 Mekanisme aktivitas thioredoxin reduktase (Leichert et al. 2003)
Terdapat dua jenis disulfida reduktase yaitu disulfida reduktase yang bersifat
dependent dan independent NADPH. Secara umum, berat molekul disulfida reduktase
berukuran di bawah seratus kilodalton dan substrat untuk pengujian aktivitas disulfida reduktase antara lain glutation teroksidasi (GSSG), sistin, DTNB dan insulin. Namun informasi terkait suhu optimum reaksi katalitik disulfida reduktase masih sangat terbatas. Disulfida reduktase yang dipelajari sebagian besar diperoleh dari cairan intraseluler, kecuali disulfida reduktase yang berasal dari bakteri gram negatif
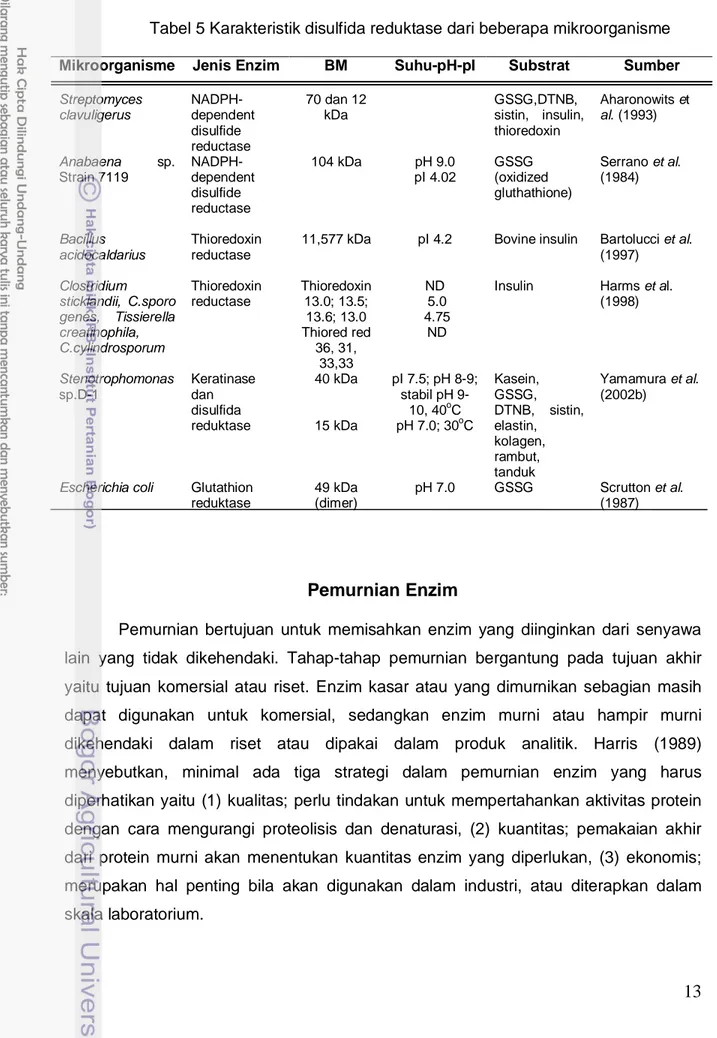
Tabel 5 Karakteristik disulfida reduktase dari beberapa mikroorganisme
Mikroorganisme Jenis Enzim BM Suhu-pH-pI Substrat Sumber
Streptomyces clavuligerus NADPH-dependent disulfide reductase 70 dan 12 kDa GSSG,DTNB, sistin, insulin, thioredoxin Aharonowits et al. (1993) Anabaena sp. Strain 7119 NADPH-dependent disulfide reductase 104 kDa pH 9.0 pI 4.02 GSSG (oxidized gluthathione) Serrano et al. (1984) Bacillus acidocaldarius Thioredoxin reductase
11,577 kDa pI 4.2 Bovine insulin Bartolucci et al. (1997) Clostridium sticklandii, C.sporo genes, Tissierella creatinophila, C.cylindrosporum Thioredoxin reductase Thioredoxin 13.0; 13.5; 13.6; 13.0 Thiored red 36, 31, 33,33 ND 5.0 4.75 ND
Insulin Harms et al. (1998) Stenotrophomonas sp.D-1 Keratinase dan disulfida reduktase 40 kDa 15 kDa pI 7.5; pH 8-9; stabil pH 9-10, 40oC pH 7.0; 30o Kasein, GSSG, DTNB, sistin, elastin, kolagen, rambut, tanduk C Yamamura et al. (2002b)
Escherichia coli Glutathion reduktase 49 kDa (dimer) pH 7.0 GSSG Scrutton et al. (1987)
Pemurnian Enzim
Pemurnian bertujuan untuk memisahkan enzim yang diinginkan dari senyawa lain yang tidak dikehendaki. Tahap-tahap pemurnian bergantung pada tujuan akhir yaitu tujuan komersial atau riset. Enzim kasar atau yang dimurnikan sebagian masih dapat digunakan untuk komersial, sedangkan enzim murni atau hampir murni dikehendaki dalam riset atau dipakai dalam produk analitik. Harris (1989) menyebutkan, minimal ada tiga strategi dalam pemurnian enzim yang harus diperhatikan yaitu (1) kualitas; perlu tindakan untuk mempertahankan aktivitas protein dengan cara mengurangi proteolisis dan denaturasi, (2) kuantitas; pemakaian akhir dari protein murni akan menentukan kuantitas enzim yang diperlukan, (3) ekonomis; merupakan hal penting bila akan digunakan dalam industri, atau diterapkan dalam skala laboratorium.
Pemekatan Enzim
Pemekatan protein enzim merupakan tahap awal prosedur pemurnian enzim sebelum tahap berikutnya atau dapat pula digunakan untuk keperluan analisis enzim (Harris 1989). Ada dua metode pemekatan enzim yaitu analitik dan preparatif (penyiapan). Metode analitik menggunakan pengendapan asam (misalnya asam trikloroasetat), pengendapan organik (misalnya aseton atau etanol), dan imunopresipitasi dapat menyebabkan denaturasi protein. Berbeda dengan metode analitik, maka metode preparatif tetap mempertahankan aktivitas protein. Pemekatan protein dengan metode preparatif misalnya dengan menggunakan garam, pelarut organik, polimer organik, ultrafiltrasi, dan liofilisasi (Bollag & Edelstein 1991).
Prinsip pengendapan dengan garam berdasarkan pada kelarutan protein yang berinteraksi polar dengan molekul air, interaksi ionik protein dengan garam, dan daya tolak menolak protein yang bermuatan sama. Kelarutan protein pada pH dan suhu tertentu akan meningkat saat konsentrasi garam meningkat (salting in). Kenaikan kelarutan protein akan meningkatkan kekuatan ion larutan. Pada saat penambahan garam dengan konsentrasi tertentu kelarutan protein akan mengalami penurunan (salting out). Molekul air yang berikatan dengan ion-ion garam akan semakin banyak sehingga menyebabkan penarikan selubung air yang mengelilingi permukaan protein. Peristiwa ini mengakibatkan protein saling berinteraksi, beragregasi, dan kemudian mengendap (Harris 1989, Scopes 1987). Amonium sulfat merupakan garam yang paling sering digunakan untuk mengendapkan protein karena memiliki sifat kelarutan tinggi di dalam air, relatif tidak mahal, dan kestabilan protein di dalam larutan amonium sulfat (2M – 3M) tahan bertahun-tahun (Scopes 1987).
Pengendapan protein menggunakan pelarut organik berdasarkan pada prinsip pengurangan kelarutan protein dan konstanta dielektrika pelarut. Semakin banyak pelarut organik yang ditambahkan, semakin berkurang daya solvasi air dan muatan pada permukaan molekul protein yang hidrofilik. Hal ini akan menjadikan molekul-molekul protein cenderung berinteraksi dengan sesamanya, hingga akhirnya protein mengendap. Prosedur pengendapan pelarut organik dilakukan pada suhu di bawah 10oC. Pada suhu di atas 10oC, konformasi protein akan segera berubah yang memungkinkan molekul pelarut organik mendapatkan jalan masuk ke bagian dalam
struktur protein, kemudian akan merusak interaksi hidrofobik dan akhirnya akan terjadi denaturasi (Harris 1989, Scopes 1987).
Pengendapan protein dengan polimer organik, misalnya menggunakan polietilen glikol (PEG), memiliki mekanisme yang hampir sama dengan pengendapan pelarut organik, namun pada PEG hanya perlu konsentrasi yang lebih rendah dari 20%. PEG adalah polimer nonionik tidak larut dengan berat molekul berkisar 6000– 20000. Pengendapan dengan PEG tidak mempengaruhi tahap pemurnian berikutnya, misalnya dengan kromatografi penukar ion atau kromatografi afinitas (Harris 1989).
Garam yang berlebihan di dalam larutan enzim, setelah tahap fraksinasi dapat dihilangkan dengan cara dialisis. Pada tahap dialisis, protein ditempatkan di dalam kantung (membran) semipermeabel yang direndam di dalam larutan bufer tertentu. Molekul yang berukuran kecil akan keluar melalui membran dan molekul yang berukuran besar akan tertahan di dalam membran dialisis. Selain dengan dialisis, penghilangan garam dapat dilakukan dengan filtrasi gel. Metode ini biasa diterapkan pada sampel yang sedikit, yaitu tidak lebih dari 25-30% volume kolom untuk mendapatkan resolusi yang memadai antara protein dan garam. Matriks filtrasi gel memiliki pori yang berukuran kecil, misalnya Sephadex G-25. Kekurangan metode ini adalah terjadi pengenceran sampel protein (Harris 1989).
Kromatografi Kolom
Terdapat beberapa cara kromatografi kolom, antara lain kromatografi filtrasi gel, kromatografi penukar ion, kromatografi interaksi hidrofobik, kromatografi afinitas, dan kromatografi cair berkinerja tinggi.
Kromatografi filtrasi gel merupakan teknik pemisahan protein dan makromolekul biologi lain berdasarkan ukuran molekul. Matriks filtrasi gel berupa gel berpori yang dikemas di dalam kolom dan dielusi dengan fase cair-mobil. Pori-pori matriks dapat menampung molekul yang berukuran lebih kecil dan memisahkannya dari molekul yang berberat molekul tinggi. Kromatografi ini biasa digunakan untuk pendugaan berat molekul (Scopes 1987).
Kromatografi penukar ion memanfaatkan perbedaan afinitas antara molekul bermuatan di dalam larutan dengan senyawa yang tidak reaktif yang bermuatan berlawanan sebagai pengisi kolom. Permukaan protein terdiri atas muatan positif dan
negatif tergantung pada rantai samping asam amino. Nilai pH pada saat protein memiliki jumlah muatan positif dan negatif sama disebut titik isoelektrik (pI). Nilai pI sebagian besar protein berada di antara pH 5 dan 9. Protein yang memiliki pH di atas pI akan bermuatan negatif, sedangkan protein dengan pH di bawah pI bermuatan positif.
Pengerjaan kromatografi penukar ion didahului dengan mengelusi protein enzim dengan pH bufer awal yang telah diatur. Protein diharapkan terikat kuat pada kolom, dan protein lain dibiarkan terelusi dulu. Protein yang terikat pada kolom dilepaskan dengan cara mengubah pH bufer atau kekuatan ion pelarut. Matriks penukar ion mengikat secara kovalen gugus fungsional yang bermuatan negatif pada penukar kation, atau gugus fungsional yang bermuatan positif pada penukar anion. Matriks berupa polimer elastis dan mengandung senyawa resin sintetik terbuat dari bahan dekstran, selulosa, atau sephadex. Contoh matriks penukar kation dan anion masing-masing adalah karboksimetil selulosa (CMC) dan dietilaminoetil (DEAE) selulosa (Scopes 1987).
Kromatografi interaksi hidrofobik banyak digunakan untuk pemisahan protein dan peptida. Pada kekuatan ion tinggi, protein akan terikat kuat pada matriks melalui interaksi hidrofobik karena matriks bersifat nonpolar. Campuran protein dialirkan ke dalam kolom dengan bufer yang mengandung konsentrasi garam tinggi. Protein yang tidak terikat keluar lebih dulu, sedangkan protein yang terikat selanjutnya dielusi menggunakan eluen yang polaritasnya diturunkan (konsentrasi garam rendah) (O’Farrel 1998).
Pemisahan protein dengan kromatografi afinitas berdasar pada prinsip interaksi spesifik di antara makromolekul biologi dengan pasangannya (ligan), sebagai contoh enzim dengan substrat atau inhibitornya dan antibodi dengan antigennya. Ligan akan terikat secara kovalen pada matriks sehingga komponen protein yang memiliki afinitas spesifik terhadap ligan akan diikat sedangkan komponen lainnya (protein kontaminan) yang tidak memiliki afinitas akan terelusi lebih dulu. Komponen protein yang terikat dapat dilepaskan dengan mengubah kondisi elusi, misalnya mengubah pH, menambahkan kofaktor atau substrat (Jack 1998).
Kromatografi cair berkinerja tinggi fase balik (reversed-phase high performance
peptida dan protein. Mekanisme teknik kromatografi ini adalah protein akan terikat berdasarkan karakter hidrofobiknya. Interaksi hidrofobik terjadi antara pelarut dengan ligan hidrofobik di pemukaan. Peningkatan muatan ion pelarut pada fase mobil akan mengurangi kekuatan ikatan dan akhirnya senyawa akan terelusi. Teknik kromatografi ini banyak digunakan dalam bioteknologi untuk mengamati tingkat kemurnian dan stabilitas protein (Neville 1998).
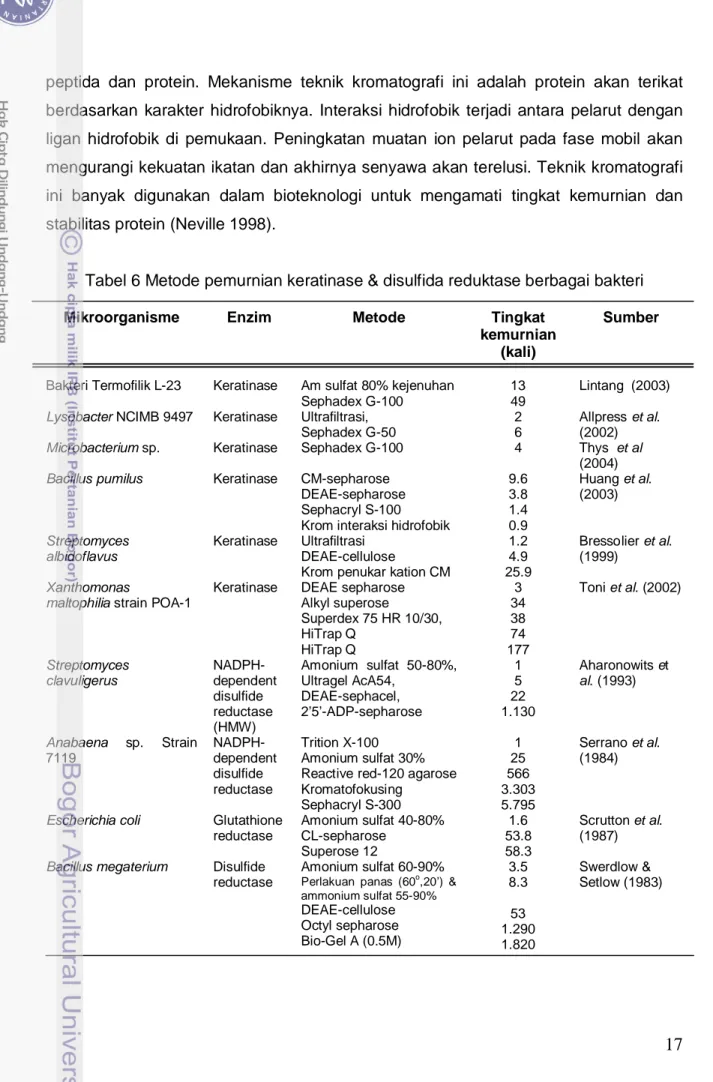
Tabel 6 Metode pemurnian keratinase & disulfida reduktase berbagai bakteri
Mikroorganisme Enzim Metode Tingkat
kemurnian (kali)
Sumber
Bakteri Termofilik L-23 Keratinase Am sulfat 80% kejenuhan Sephadex G-100
13 49
Lintang (2003)
Lysobacter NCIMB 9497 Keratinase Ultrafiltrasi, Sephadex G-50
2 6
Allpress et al. (2002)
Microbacterium sp. Keratinase Sephadex G-100 4 Thys et al
(2004)
Bacillus pumilus Keratinase CM-sepharose DEAE-sepharose Sephacryl S-100 Krom interaksi hidrofobik
9.6 3.8 1.4 0.9 Huang et al. (2003) Streptomyces albidoflavus Keratinase Ultrafiltrasi DEAE-cellulose
Krom penukar kation CM
1.2 4.9 25.9 Bressolier et al. (1999) Xanthomonas
maltophilia strain POA-1
Keratinase DEAE sepharose Alkyl superose Superdex 75 HR 10/30, HiTrap Q HiTrap Q 3 34 38 74 177 Toni et al. (2002) Streptomyces clavuligerus NADPH-dependent disulfide reductase (HMW) Amonium sulfat 50-80%, Ultragel AcA54, DEAE-sephacel, 2’5’-ADP-sepharose 1 5 22 1.130 Aharonowits et al. (1993) Anabaena sp. Strain 7119 NADPH-dependent disulfide reductase Trition X-100 Amonium sulfat 30% Reactive red-120 agarose Kromatofokusing Sephacryl S-300 1 25 566 3.303 5.795 Serrano et al. (1984)
Escherichia coli Glutathione reductase Amonium sulfat 40-80% CL-sepharose Superose 12 1.6 53.8 58.3 Scrutton et al. (1987)
Bacillus megaterium Disulfide reductase Amonium sulfat 60-90% Perlakuan panas (60o 3.5 8.3 53 1.290 1.820 ,20’) & ammonium sulfat 55-90% DEAE-cellulose Octyl sepharose Bio-Gel A (0.5M) Swerdlow & Setlow (1983)
Banyak peneliti telah melaporkan metode pemurnian berbagai jenis keratinase dan disulfida reduktase yang dihasilkan oleh mikroorganisme (Tabel 6). Pemekatan menggunakan amonium sulfat atau ultrafiltrasi banyak dipilih untuk memulai pemurnian, kemudian dilanjutkan dengan kromatografi gel filtrasi atau kromatografi penukar ion.
Elektroforesis dan Zimografi
Elektroforesis pada gel poliakrilamida adalah metode yang paling sering digunakan untuk karakterisasi protein dan campuran protein, karena prosedur ini relatif cepat dan sensitif. Elektroforesis didefinisikan sebagai perpindahan partikel-partikel bermuatan karena pengaruh medan listrik. Mekanisme pada elektroforesis gel poliakrilamida sodium dodesil sulfat (SDS-PAGE) adalah bahwa protein akan bereaksi dengan SDS yang merupakan detergen anionik membentuk kompleks yang bermuatan negatif. Protein akan terdenaturasi dan larut membentuk kompleks berikatan dengan SDS. Kompleks protein yang bermuatan negatif ini kemudian akan terpisahkan berdasar muatan dan ukurannya secara elektroforesis di dalam matriks gel. Berat molekul protein dapat diukur menggunakan protein standar yang telah diketahui berat molekulnya dengan cara membandingkan nilai mobilitas relatifnya (Rf) (Bollag & Edelstein 1991). Berbagai dislfida reduktase asal bakteri berhasil dideteksi menggunakan teknik SDS-PAGE dan hasilnya menginformasikan terdapat disulfida reduktase yang bersifat homodimer (Tabel 7).
Tabel 7 Pendugaan berat molekul beberapa disulfida reduktase asal bakteri menggunakan teknik SDS-PAGE
Mikroorganisme Metode ∑ pita Berat molekul (kDa) Sumber
Streptomyces clavuligerus
SDS-PAGE 2 70 (dimer) dan 12 Aharonowits et al. (1993)
Anabaena sp. Strain
7119
PAGE 1 104 Serrano et al. (1984)
Bacillus acidocaldarius
SDS-PAGE 1 10 Bartolucci et al. (1997)
Escherichia coli SDS-PAGE 1 49 Scrutton et al. (1987)
Bacillus megaterium SDS-PAGE 1 122 Swerdlow & Setlow
(1983)
Elektroforesis gel native (nondenaturasi) memisahkan protein terlarut yang tidak mengendap atau beragregasi selama elektroforesis. Metode ini dapat pula diterapkan untuk protein dengan kelarutan rendah, seperti protein membran, dengan menambahkan detergen nonionik, misalnya 0.5% Triton X-100 ke dalam gel. Tidak seperti pada SDS-PAGE yang molekul-molekul protein yang telah terpisah dapat kehilangan aktivitas biologi dan biokimianya maka pada elektroforesis gel nondenaturasi aktivitas tersebut masih ada (Walker 2002). Tabel 8 menginformasikan teknik mendeteksi molekul protein keratinase yaitu dengan metode SDS-PAGE dan zymografi. Zimografi adalah teknik elektroforesis untuk menetapkan aktivitas enzim secara insitu. Berbeda dengan SDS-PAGE, gel pemisah zimografi mengandung substrat enzim yang akan dihidrolisis oleh enzim selama masa inkubasi. Enzim dipisahkan dalam gel denaturasi (SDS), namun dalam kondisi tidak tereduksi. Penambahan detergen Triton X-100 akan melepaskan SDS sehingga protein kembali melipat (renaturasi). Gel selanjutnya diwarnai sehingga molekul protein yang memiliki aktivitas tampak sebagai pita bening. Metode zimografi bersifat mudah, sensitif, dan kualitatif dalam menganalisis aktivitas enzim (Leber & Balkwill 1997).
Tabel 8 Pendugaan berat molekul keratinase dari beberapa bakteri menggunakan teknik SDS-PAGE dan Zimografi.
Mikroorganisme Metode Jumlah pita Berat molekul
(kDa)
Sumber
Bakteri Termofilik L-23 SDS-PAGE Zimografi 6 2 98.5; 92.6; 82; 64.2; 56.8; 25.6 47 dan 64 Lintang (2003) Streptomyces albidoflavus
SDS-PAGE 1 18 Bressolier et al.
(1999) Fervidobacterium pennavorans SDS-PAGE Zimografi 1 1 130 130 Friedrich & Antranikian (1996)
Bacillus pumilus SDS-PAGE 1 32 Huang et al.
(2003)
Lysobacter NCIMB
9497
SDS-PAGE 1 148 Allpress et al.
(2002.)
Xanthomonas maltophilia strain POA-1
SDS-PAGE 1 36 Toni et al. (2002)
Hingga saat ini, analisis zimografi keratinase masih menggunakan gelatin sebagai substrat. Gelatin adalah protein yang diturunkan dari kolagen. Keratin, elastin, dan kolagen adalah protein fibrosa/serat yang banyak terdapat di alam. Keratin dan gelatin keduanya mengandung banyak residu glisin namun gelatin larut dalam air.
Penggunaan gelatin sebagai substrat dalam teknik zimografi cukup bervariasi, pada bakteri termofilik L-23 (Lintang 2003) dan S. albidoflavus (Bressolier 1999) digunakan 2% gelatin sedangkan pada F. Pennavorans 0.1% (Friedrich & Antranikian 1996).
Degradasi Keratin
Laju dan kesempurnaan degradasi keratin oleh mikroorganisme sangat bergantung pada jenis substratnya, dan hal ini erat berkaitan dengan kandungan sistin. Substrat yang sering digunakan untuk mempelajari aktivitas hidrolisis keratinase adalah bulu ayam (Riffel et al. 2003a, Thys et al. 2004, Toni et al. 2002, Werlang & Brandelli 2005, Zerdani et al. 2004), rambut manusia (Takami et al. 1992, Macedo et
al. 2005), bulu sapi (Riffel et al. 2003b) dan wool (Ignatova et al. 1999, Huang et al.
2003).
Pada fungi keratinolitik, konsentrasi rendah glukosa (0.1%) dan sedikit suplementasi asam amino, pepton atau ekstrak kamir mungkin akan memacu pertumbuhan saat fase awal. Namun nutrisi terlarut dengan konsentrasi tinggi akan menghambat hidrolisis substrat keratin. Degradasi keratin dalam medium cair ditandai dengan dilepaskannya produk-produk hidrolisis ke dalam medium. Produk utama adalah peptida berberat molekul satu hingga dua kilodalton, akan tetapi ditemukan juga asam-asam amino bebas dan protein berberat molekul tinggi. Indikator terbaik terjadinya keratinolisis adalah peningkatan pH medium (sedikitnya mencapai pH 8.0) yang menggambarkan penggunaan protein keratin, deaminasi, dan produksi amonia (Kunert 2000).
Akumulasi produk metabolit yang mengandung sulfur juga ditemukan dalam medium cair. Keratin sangat kaya akan sulfur (0.5% pada soft keratin dan mencapai 4% pada hard keratin). Kelebihan sulfur dikurangi dengan dioksidasi menjadi sulfat dan dilepaskan ke dalam medium. Sulfat merupakan produk akhir dan bersifat inert, konsentrasi sulfat dalam medium dapat menjadi parameter yang baik terjadinya degradasi substrat. Akan tetapi konsentrasi sulfur organik berupa sistin dan peptida mengandung S-sulphocysteine relatif rendah, sedangkan kandungan sistein (Tiol) sangat bervariasi. Tiol, termasuk sistein dalam kondisi alkali akan dioksidasi kembali oleh oksigen menjadi disulfida (Kunert 2000).
Streptomyces memulai aktivitas proteolitiknya dengan proses denaturasi
(reduksi) jembatan disulfida (Bockle & Muller 1997). Namun pada fungi keratinolitik tidak terjadi reduksi keratin oleh enzim ekstraseluler dan atau Tiol. Pada fungi selain protease tampaknya alkalinisasi medium dan sulphitolysis merupakan faktor utama denaturasi keratin (Kunert 2000). Fungi mengeksresikan sulfit dan amonia, dalam kondisi alkali sulfit akan memotong jembatan disulfida substrat. Kemudian secara perlahan substrat akan terdenaturasi dan hal ini mempermudah kerja protease.
Sulphitolysis pada jembatan disulfida menghasilkan produk utama S-sulphocysteine
baik dalam bentuk bebas maupun kombinasi dengan peptida. Pada kultur yang sudah tua, sedikitnya 80% sulfur organik berupa S-sulphocysteine. Sulfit merupakan stimulator yang kuat bagi protease M. gypseum saat menghidrolisis keratin wool (Kunert 1989).
Cys-SS-cys + HSO
3-cys-SH + cys-SSO
3-Sistin Sulfit Sistein S-sulphocysteine
Menurut Kunert (2000), degradasi keratin pada fungi diduga merupakan hasil kerja tiga faktor yaitu “deaminasi” (menghasilkan lingkungan alkali yang dibutuhkan untuk pengembangan substrat, sulphitolysis, dan aksi proteolitik); “sulphitolysis” (denaturasi substrat dengan memutus jembatan disulfida) dan “proteolisis” (pemotongan substrat yang telah terdenaturasi menjadi produk terlarut).
Bacillus licheniformis RG1 menghasilkan enzim keratinolitik ekstraseluler yang
secara sinergis aktivitasnya ditingkatkan oleh disulfida reduktase intraseluler. Terdeteksinya sulfit dalam cairan ekstraseluler selama degradasi mengindikasikan terjadinya sulphitolysis (Ramnani et al. 2005). Ignatova et al. (1999) melaporkan bahwa sebagian besar keratinase yang telah dimurnikan tidak mampu mendegradasi keratin, kecuali keratinase murni B. licheniformis PWD-1 yang dapat menghidrolisis keratin bulu. Degradasi keratin oleh Streptomyces spp. didukung proses reduksi pada ikatan disulfida oleh disulfida reduktase (Bockle & Muller 1997) atau produksi sulfit dan tiosulfat pada Streptomyces fradiae (Kunert 2000). Adanya aktivitas disulfida reduktase juga ditunjukkan oleh Vibrio sp. strain kr6 dengan terdeteksinya senyawa Tiol dalam medium (Riffel et al. 2003a). Yamamura et al. (2002a) melaporkan aksi bersama antara protein serupa disulfida reduktase dan protease yang
dihasilkan oleh Stenotrophomonas sp. dalam degradasi keratin bulu rusa. Mekanisme degradasi keratin yang diusulkan adalah :
K-S-S-K Dis-red like protein K-SH Protease Peptida/asam amino Native keratin Reduced keratin
Penggunaan agens pereduksi (reduktan) untuk meningkatkan degradasi keratin telah dilaporkan oleh beberapa peneliti. Reduktan yang digunakan antara lain 2% sodium azide (Ramnani et al. 2005), 0.05% sodium azide dan 10 mM EDTA ((Bockle & Muller 1997), 10 mM sodium sulfit dan dithioerythritol (Letourneau et al. 1998).
Melekatnya sel/kolonisasi pada substrat merupakan hal yang penting dalam proses degradasi bulu ayam (Bockle & Muller 1997, Onifade et al. 1998). Degradasi lengkap bulu ayam utuh oleh B. Licheniformis RG1 terjadi hanya jika ada sel bakteri hidup, melekatnya sel pada bulu diduga secara kontinu menyediakan reduktan yang digunakan untuk memutus jembatan disulfida (Ramnani et al. 2005). Menurut Bockle dan Muller (1997), pada prokariot sistem redoks cell-bound untuk reduksi ikatan disulfida juga berperan dalam degradasi keratin.
Aplikasi Keratinase
Produksi Tepung Bulu Sebagai Pakan Tenak
Pembuatan tepung bulu sebagai pakan ternak diperoleh dengan cara pemanasan pada suhu dan tekanan tinggi. Perlakuan hidrotermal tersebut menyebabkan kerusakan pada asam-asam amino tertentu seperti metionin, lisin, dan triptofan, serta menghasilkan produk dengan kualitas nutrisi rendah (Wang & Parsons 1997). Penggunaan keratinase mikrobial dapat menjadi alternatif untuk hidrolisis bulu menjadi pakan ternak yang kaya nutrisi (Onifade et al. 1998).
Suplementasi tepung bulu/bulu kasar dengan keratinase PWD-1 mampu mengubah struktur keratin sehingga meningkatkan daya cerna dan pertumbuhan ayam (Odetallah et al. 2003). Tepung bulu relatif lebih murah dan lebih unggul dalam hal kandungan sistein, treonin, dan valinnya bila dibandingkan dengan bungkil kedelai
(Apple et al. 2003) dan tepung bulu hasil hidrolisis mampu menggantikan bungkil kedelai hingga tujuh persen. Enzim kasar juga dapat digunakan sebagai produk
nutriceutical yang secara signifikan dapat meningkatkan performan broiler (Odetallah et al. 2003). Ekstrak kasar keratinase Cunninghamella spp. yang digunakan dalam
fermentasi tepung bulu ayam mampu meningkatkan kualitas protein dan energi metabolisnya (Ramli et al. 2001).
Perbaikan nutrisi bulu/tepung bulu juga dapat dilakukan dengan cara fermentasi menggunakan mikroorganisme keratinolitik. Fermentasi secara signifikan meningkatkan asam-asam amino esensial (metionin, arginin, dan lisin) dan biomassa merupakan sumber protein yang kaya. Percobaan pakan menggunakan biomassa
B.licheniformis PWD-1 mampu meningkatkan kurva pertumbuhan ayam pedaging
sama baiknya dengan standar bungkil kedele (Williams et al. 1991). Guna memenuhi permintaan industri pakan ternak dan meningkatkan produksi keratinase, beberapa peneliti melakukan kloning, over-ekspresi, dan bioimobilisasi (Wang et al. 2003).
Produksi Tepung Bulu Sebagai Pupuk
Kandungan protein yang tinggi pada tepung bulu selain dimanfaatkan sebagai pakan ternak juga dapat diaplikasikan sebagai pupuk nitrogen semi-slow-release pada pertanian organik. Pertanian organik memerlukan pupuk organik yang kaya nitrogen untuk mendukung pertumbuhan tanaman dan aktivitas mikrobial dalam tanah. Guano, secara tradisional telah digunakan luas sebagai pupuk pada pertanian organik, namun harganya sangat mahal sehingga diperlukan upaya untuk mendapatkan alternatf lain (Gupta & Ramnani 2006). Tepung bulu adalah materi yang kaya nitrogen, murah, dan selalu tersedia. Selain dapat menyuplai nitrogen untuk tanaman dan aktivitas mikroba tanah, tepung bulu juga dapat menjaga struktur tanah dan meningkatkan kapasitas retensi air.
Aplikasi Pada Deterjen
Enzim-enzim proteolitik telah lama mendominasi pasar deterjen. Sekitar 89% enzim dalam deterjen adalah protease alkalin dengan pemasok utama Novo Nordisk dan Genencor International. Keratinase memiliki kemampuan mengikat dan
menghidrolisis substrat padatan. Hal ini merupakan sifat utama yang dibutuhkan enzim dalam deterjen karena enzim akan bekerja pada substrat protein yang menempel pada permukaan padat, sehingga enzim diperlukan sebagai aditif untuk pembersih pada permukaan yang keras. Keratinase membantu menghilangkan kotoran berkeratin yang sering ditemukan pada bagian leher kemeja yang tidak dapat dibersihkan oleh sebagian besar protease (Gupta & Ramnani 2006). Aplikasi terbaru keratinase dalam deterjen adalah penggunaan keratinase sebagai aditif untuk menghilangkan sumbatan akibat limbah berkeratin (Farag & Hasan 2004).
Degradasi Protein Prion
Prion adalah partikel protein penyebab penyakit neurodegeneratif yang dikenal sebagai transmissible spongiform encephalopathies (TSE) . Termasuk dalam golongan TSE adalah penyakit sapi gila, scrapie, kuru, dan Creutzfeld-Jacob. Sifat infektif prion berkaitan dengan perubahan protein prion PrPc menjadi protein prion infektif (PrPsc) yang difasilitasi oleh PrPsc itu sendiri (Gupta & Ramnani 2006).
Dewasa ini beberapa peneliti berusaha menggali potensi keratinase bakteri dalam menghancurkan protein prion abnormal pada produk asal ternak. Melalui perlakuan pemanasan di atas 100o
Pada industri kulit sebesar 70% polusi dihasilkan dari proses pre-tanning Polutan utama dari industri kulit adalah sodium sulfida, kromium, asam, dan limbah padat; semua bahan tersebut akan meningkatkan biochemical oxygen demand (BOD),
chemical oxygen demand (COD), dan total padatan terlarut (TDS). Proses biokatalitik
C dan penambahan deterjen Langeveld et al. (2003) berhasil mendegradasi prion abnormal pada jaringan otak sapi dan domba menggunakan keratinase yang dihasilkan oleh B. licheniformis PWD-1. Protein serupa prion yaitu protein Sup35NM-His6 yang dihasilkan oleh Saccharomyces cerevisae diketahui dapat dihancurkan oleh subtilisin, proteinase K dan keratinase dari B.
licheniformis PWD-1 namun tidak oleh kolagenase dan elastase (Wang et al. 2005).
Keratinase juga dapat digunakan untuk dekontaminasi peralatan medis dan laboratorium (Langeveld et al. 2003).
kulit melibatkan enzim campuran antara lain protease, lipase, dan karbohidrase yang biasa digunakan pada berbagai tahap pre-tanning (Gupta & Ramnani 2006).
Protease keratinolitik yang tidak memiliki aktivitas kolagenase namun mempunyai cukup aktivitas elastase dapat membantu proses dehairing dengan memutus secara selektif jaringan keratin pada folikel kulit sehingga rambut akan terlepas tanpa memengaruhi kekuatan kulit. Penggunaan enzim dalam proses
de-hairing kulit selain menghasilkan kulit dengan kualitas baik juga dapat mengurangi
polutan (Macedo et al. 2005). Bacillus subtilis dan Bacillus amiloliquefaciens menghasilkan enzim protease dengan aktivitas depilatory. Berdasar analisis mikroskopis, kulit sapi berambut yang direndam dalam kultur bebas sel selama semalam menunjukkan hilangnya epidermis, bulu, tidak adanya kelenjar kulit dan terbukanya struktur serat kulit (Riffel et al. 2003b). Strain mutan Bacillus pumilus UN-31-C-42 menghasilkan protease kasar dan murni yang memiliki aktivitas de-hairing tinggi dan aktivitas kolagenase rendah sehingga diharapkan dapat menghasilkan kulit dengan kualitas baik (Huang et al. 2003). Dengan demikian proses penghilangan bulu secara kimiawi berbahan dasar sulfida yang dapat merusak lingkungan dapat digantikan dengan proses berbahan dasar enzim keratinase yang lebih ramah lingkungan.
Aplikasi Di Bidang Lain
Potensi aplikasi keratinase yang lain adalah fermentasi anaerobik limbah industri ternak unggas untuk menghasilkan gas alam, modifikasi serat seperti sutera dan wool, di bidang medis dan farmasi untuk menghilangkan jerawat atau psoriasis dan sebagai aditif bahan pencerah kulit dengan mestimulasi degradasi keratin (Gupta & Ramnani 2006).