POTENSI HUTAN RAKYAT MINDI (Melia azedarach L.) SEBAGAI SUMBER BENIH: VARIABILITAS GENETIK POHON INDUK,
MORFOLOGI BENIH DAN PERTUMBUHAN BIBIT
LASWI IRMAYANTI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul “Potensi Hutan Rakyat Mindi (Melia azedarach L.) Sebagai Sumber Benih: Variabilitas Genetik Pohon Induk, Morfologi Benih dan Pertumbuhan Bibit” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
LASWI IRMAYANTI. Potensi Hutan Rakyat Mindi (Melia azedarach L.) Sebagai Sumber Benih: Variabilitas Genetik Pohon Induk, Morfologi Benih dan Pertumbuhan Bibit. Dibimbing oleh ISKANDAR Z. SIREGAR dan PRIJANTO PAMOENGKAS.
Mindi (Melia azedarach L.) merupakan salah satu jenis yang umum ditemukan di hutan rakyat Jawa Barat. Salah satu faktor yang yang mempengaruhi produktivitas hutan rakyat adalah benih yang berkualitas. Akan tetapi, informasi tentang kualitas sumber benih di hutan rakyat saat ini masih kurang, baik mutu fisik, genetik, maupun fisiologi. Tujuan dari penelitian ini adalah untuk: 1) menduga keragaman genetik di dalam dan antar populasi mindi di hutan rakyat Jawa Barat, dan 2) menduga variabilitas dimensi buah mindi yang dikumpulkan dari lima sumber benih yang berbeda, dan pertumbuhan bibit mindi di persemaian pada tujuh media yang berbeda dari campuran top soil, pasir, arang sekam, dan pupuk kandang berupa kotoran sapi.
Hasil penelitian menunjukkan rata-rata alel yang teramati (Ne) sebesar 2.200,
PLP (percentage of polymorphic loci) sebesar 93.33%, dan keragaman genetik dalam populasi (He) mindi sebesar 0.366. Keragaman genetik tertinggi adalah
populasi Padasari (He= 0.454) dan terendah adalah Babakan Rema (He= 0.220).
Rata-rata nilai perbedaan genetik antar populasi (Fst) didapatkan 0.303. Pada
analisis klaster menunjukkan adanya pengelompokan populasi mindi yang terbagi menjadi 3 klaster. Klaster pertama terdiri atas Nagrak dan Padasari, klaster ke dua Babakan Rema, Gambung dan Sukakarya, serta klaster ke tiga hanya populasi Legok Huni. Klaster yang sama menunjukkan kekerabatan yang dekat. Berdasarkan hasil penelitian, keragaman genetik populasi mindi tergolong pada keragaman sedang, yang dapat dijadikan pertimbangan pada waktu seleksi dan pembangunan sumber benih.
Variabilitas dimensi buah mindi secara spasial yang dikumpulkan dari lima sumber benih berbeda, dan pertumbuhan bibit mindi di persemaian tergolong sedang. Buah mindi dari Nagrak mempunyai ukuran, berat, dan seed set yang paling tinggi, serta persentase, nilai dan kecepatan kecambah paling tinggi. Hasil pengamatan pertumbuhan bibit mindi dari sumber benih dan media sapih berbeda menunjukkan bahwa bibit dari Nagrak memiliki pertumbuhan lebih cepat. Media sapih yang mengandung pupuk kandang berupa kotoran sapi memberikan pertumbuhan paling cepat.
SUMMARY
LASWI IRMAYANTI. Potential Mindi (Melia azedarach L.) Community Forest as a Seed source: Genetic Variability Mother Tree, Fruit Morphology and Seedling Growth. Supervised by ISKANDAR Z. SIREGAR and PRIJANTO PAMOENGKAS.
Mindi (Melia azedarach L.) is mostly found in community forests of West Java. One of the factors that affect productivity of mindi plantation is high quality seed that is not easily available. Seed sources of mindi should be established based on the status of genetic variation, preferably the one with high genetic variation. However, information on the status of seed source quality is always lacking. This research was aimed to: 1) estimate genetic variation within and between populations of mindi in West Java community forests based on microsatellites. and 2) determine baseline information on spatial variability of mindi fruits collected from five seed sources, and seedling growths treated in seven different growing media consisting mixtures of soil, rice husk, sand and cattle manure. Six mindi populations were selected in the following villages: Nagrak (Bogor), Babakan Rema (Kuningan), Padasari (Sumedang), Legok Huni (Wanayasa), Sukakarya (Bogor) and Gambung (Bandung).
The results showed that average number of alleles (Ne) was 2.200, PLP
(percentage of polymorphic loci) was 93.33%, and the genetic diversity within populations (He) was 0.366. The highest genetic diversity was found in Padasari (He = 0.454), and the lowest one was in Babakan Rema (He = 0.220). The genetic difference between populations (Fst) was 0.303. Cluster analysis showed three groupings in which the first and second clusterconsisted of Nagrak and Padasari, as well as Babakan Rema,Gambung and Sukakarya, respectively. On the other hand Legok Huni was separated alone. The findings provided information on the moderate status of genetic diversity that may be considered during the selection and establishment of seed sources.
The dimensional variability of mindi fruits collected from different sources of seed and their seedling growth in the nurseries are still relatively moderate. Mindi fruits of Nagrak have the largest size, weight, and seed set and the highest percentage of germination. The observations on the mindi seed growth from different seed sources and seedling media show that the seeds of Nagrak have faster growth. The seedling media containing manure provide the most rapid growth.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Silvikultur Tropika
POTENSI HUTAN RAKYAT MINDI (Melia azedarach L.) SEBAGAI
SUMBER BENIH: VARIABILITAS GENETIK POHON INDUK, MORFOLOGI BENIH DAN PERTUMBUHAN BIBIT
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
Judul Tesis : Potensi Hutan Rakyat Mindi (Melia azedarach L.) Sebagai Sumber Benih: Variabilitas Genetik Pohon Induk, Morfologi Benih dan Pertumbuhan Bibit
Nama : Laswi Irmayanti
NIM : E451120021
Disetujui oleh Komisi Pembimbing
Prof Dr Ir Iskandar Z. Siregar, MForSc Ketua
Dr Ir Prijanto Pamoengkas, MScFTrop Anggota
Diketahui oleh
Ketua Program Studi Silvikultur Tropika
Prof Dr Ir Sri Wilarso Budi R, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji syukur kehadirat Allah SWT atas rahmatNya hingga tesis yang berjudul
“Potensi Hutan Rakyat Mindi (Melia azedarach L.) Sebagai Sumber Benih: Variabilitas Genetik Pohon Induk, Morfologi Benih dan Pertumbuhan Bibit” dapat diselesaikan. Shalawat dan salam semoga selalu tercurah kepada nabi besar Muhammad SAW.
Pada kesempatan ini penulis mengucapkan terimakasih dan penghargaan kepada Prof Dr Ir Iskandar Z. Siregar, MForSc dan Dr Ir Prijanto Pamoengkas, MScFTrop selaku komisi pembimbing, atas arahan dan bimbingannya. Ucapan terima kasih dan penghargaan juga kami sampaikan kepada Direktorat Jenderal Pendidikan Tinggi (DIKTI) yang telah memberikan Beasiswa Unggulan Dikti kepada penulis untuk tahun anggaran 2012-2014 (lampiran surat Dikti No.2460/E4.4/2012). Terimakasih kepada Hastuti, SP, M.Si, Fadly Sonata Siregar, A.Md, dan Alex Yungan S.Hut atas motivasi yang diberikan kepada penulis.
Terimakasih penulis sampaikan kepada Dr.Agus Astho Pramono dan Aam Aminah, S.Hut, M.Si yang telah memberikan ide dan masukan-masukan dalam penelitian. Kepada rekan-rekan di Laboratorium Genetik, Departemen Silvikultur (Arniana Anwar, S.Hut; Asep Mulyadiana, S.Hut; Lily Novianty, S.Pd; Rajjitha Handayani, SP; Jeprianto Manurung, S.Hut dan Arina Nur Faidah) terimakasih atas bantuan dan dukungannya. Terimakasih kepada: Erekso Hadiwijoyo, S.Hut; Asep Hendra Supriatna, S.Hut; Sri Susanti, S.Hut; Indra Alamsyah dan Adi
Setiadi (SVK’48) yang telah membantu penulis dalam pengumpulan sampel penelitian di lapangan. Ungkapan terima kasih juga penulis sampaikan kepada bapak, ibu, serta seluruh keluarga, atas segala do’a dan kasih sayangnya.
Penulis juga mengucapkan terimakasih kepada semua pihak yang telah membantu baik secara langsung maupun tidak langsung selama perencanaan dan pelaksanaan penelitian, sampai tesis ini dapat diselesaikan. Semoga Allah memberi balasan yang berlipat. Amiin.
Penulis menyadari bahwa tesis ini masih jauh dari sempurna. Namun penulis selalu berharap semoga hasil penelitian ini dapat bermanfaat bagi penulis dan para pembaca.
DAFTAR ISI
DAFTAR ISI vi
DAFTAR TEBEL vii
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN vii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 1
Tujuan Penelitian 2
Manfaat Penelitian 2
2 METODE 2
Waktu dan Tempat Penelitian 2
Alat dan Bahan 3
Prosedur 4
Analisis Data 8
3 HASIL DAN PEMBAHASAN 10
Variabilitas Genetik Pohon Induk 10
Variabilitas Dimensi Buah 15
Perkecambahan 17
Pertumbuhan Bibit 19
Potensi Sumber Benih Mindi di Hutan Rakyat Jawa Barat 21
4 SIMPULAN DAN SARAN 22
Simpulan 22
Saran 22
DAFTAR PUSTAKA 22
LAMPIRAN 28
DAFTAR TABEL
2.1 Lokasi pengambilan sampel penelitian 3
2.2 Daftar alat dan bahan penelitian 4
2.3 Primer mikrosatelit mimba untuk amplifikasi silang mindi 5 2.4 Empat komponen bahan yang digunakan dalam reaksi PCR 6
2.5 Komposisi pembuatan gel poliakrilamid 6
2.6 Bahan pewarnaan gel poliakrilamid dan lama perendaman 6 3.1 Nilai parameter keragaman genetik dalam populasi mindi 12 3.2 Jarak genetik dan geografis populasi mindi di hutan rakyat Jawa Barat 12 3.3 Nilai pengujian struktur Hardy-Weinberg (HWE) berdasarkan uji G 14 3.4 Nilai indeks fiksasi setiap lokus pada mikrosatellit populasi mindi 15 3.5 Nilai pengujian struktur inbreeding berdasarkan uji X2 15 3.6 Nilai rata-rata panjang dan lebar buah mindi pada tiap lokasi 16 3.7 Nilai rata-rata massa buah mindi pada tiap lokasi 16 3.8 Rekapitulasi nilai GP, GV, dan GS benih mindi pada tiap lokasi 18 3.9 Analisis varians pengaruh media dan asal benih pada pertumbuhan mindi 19
DAFTAR GAMBAR
2.1 Peta lokasi pengambilan sampel penelitian 3
2.2 Perakitan kaca gel poliakrilamid 6
2.3 Pemilihan pohon induk untuk pengumpulan buah (Mulawarman et al. 2002) 7
2.4 Pengacakan unit percobaan 8
2.5 Cara skoring DNA mikrosatelit 9
2.6 Dendogram pengelompokan/klaster genetik 9
2.7 Tampilan struktur populasi (Purba 2012) 9
3.1 Frekuensi alel mindi di hutan rakyat Jawa Barat pada primer mimba 11 3.2 Korelasi jarak geografis dan jarak genetik populasi mindi 13
3.3 Dendogram mindi di hutan rakyat Jawa Barat 13
3.4 PCA mindi di hutan rakyat Jawa Barat 14
3.5 Tampilan struktur populasi mindi 15
3.6 Penampang dan bagian-bagian buah mindi 17
3.7 Sebaran seed set mindi pada tiap lokasi 17
3.8 Nilai persentase perkecambahan benih mindi dari awal pengamatan 18 3.9 Persentase perkecambahan benih mindi dari lima sumber benih 18 3.10 Pertambahan diameter dan tinggi bibit mindi dari sumber benih berbeda 20 3.11 Pengaruh media sapih pada pertambahan tinggi dan diameter bibit mindi 20
DAFTAR LAMPIRAN
1 Dokumentasi amplifikasi silang primer mimba ke mindi 28
2 Skoring alel pada tiap primer 29
1
PENDAHULUAN
Latar Belakang
Mindi (Melia azedarach L.) merupakan salah satu jenis alternatif pengganti kayu berkualitas yang saat ini sudah mulai sulit ditemukan dan berharga mahal karena permintaan pasar yang semakin meningkat (Syamsuwida et al. 2012). Pada industri kerajinan meubel di Jepara, kayu mindi digunakan sebagai salah satu pengganti kayu jati pada saat pasokannya berkurang. Daun, akar, kulit, dan bunga mindi dapat dimanfaatkan sebagai obat-obatan dan pestisida alami (Karyono & Hariatno 2001).
Pramono (2012) melaporkan bahwa mindi merupakan salah satu jenis tanaman yang memiliki prospek bagus untuk dikembangkan di hutan rakyat. Mindi mampu beradaptasi pada perbedaan kondisi lingkungan tumbuh yang luas (Pramono et al. 2012). Hutan rakyat mindi merupakan salah satu tipe hutan rakyat yang berkembang cukup baik dan menjanjikan khususnya di Jawa Barat (Bramasto 2011). Salah satu faktor yang mempengaruhi produktivitas hutan adalah penggunaan benih yang berkualitas. Namun, benih berkualitas masih sulit didapatkan karena keterbatasan jumlah sumber benih.
Salah satu cara membangun sumber benih adalah membangun tegakan hutan atau memanfaatkan tegakan yang sudah ada. Saat ini keberadaan hutan rakyat dapat berfungsi sebagai alternatif pilihan sumber benih. Asal-usul sumber benih merupakan salah satu penentu kualitas benih yang diharapkan mampu menghasilkan bibit yang berkualitas pula. Akan tetapi, informasi tentang kualitas sumber benih di hutan rakyat saat ini masih kurang. Pengembangan hutan rakyat dengan jenis mindi, perlu ditunjang dengan penyediaan benih (seed procurement) yang bermutu tinggi, baik kualitas fisik, fisiologi maupun genetik (Yulianti et al. 2011). Pada umumnya benih yang digunakan untuk hutan rakyat belum memperhatikan hal tersebut.
Mutu genetik merupakan penampilan benih murni dari spesies tertentu yang menunjukkan identitas genetik dari tanaman induknya (Sutopo 2011). Yulianti et al. (2011) melaporkan bahwa keragaman genetik pohon induk mindi dengan teknik RAPD sebesar 0,1712 dikategori keragaman genetik sedang. Kemudian Rambey (2011) dengan teknik analisis mikrosatelit menyatakan bahwa mindi di daerah Garut memiliki nilai keragaman genetik sebesar 0,373 yang dikategorikan keragaman sedang.
Berkaitan dengan permasalahan di atas, perlu dilakukan penelitian lebih lanjut tentang variabilitas genetik pohon induk dari beberapa sumber benih. Selain itu diperlukan juga penelitian kualitas benih melalui pengamatan variabilitas dimensi buah secara spasial, dan pertumbuhan bibit mindi dari beberapa hutan rakyat di Jawa Barat dan media yang berbeda.
Perumusan Masalah
2
1. Bagaimana variabilitas genetik (dalam dan antar populasi) tegakan mindi pada Hutan Rakyat di Jawa Barat yang dapat dikembangkan menjadi sumber benih dengan marka Mikrosatellit.
2. Bagaimana variabilitas dimensi buah mindi secara spasial yang dikumpulkan dari sumber benih yang berbeda, dan pertumbuhan bibit mindi di persemaian yang dihasilkan dari sumber benih dan media sapih berbeda.
Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Menduga variabilitas genetik (dalam dan antar populasi) tegakan mindi pada Hutan Rakyat di Jawa Barat yang dapat dikembangkan menjadi sumber benih dengan marka Mikrosatellit.
2. Menduga variabilitas dimensi buah mindi secara spasial yang dikumpulkan dari sumber benih yang berbeda, dan pertumbuhan bibit mindi di persemaian yang dihasilkan dari sumber benih dan media sapih berbeda.
Manfaat Penelitian
Manfaat dari penelitian ini yaitu:
1. Memberikan informasi variabilitas genetik (dalam dan antar populasi) tegakan mindi pada Hutan Rakyat di Jawa Barat yang dapat dikembangkan menjadi sumber benih dengan marka Mikrosatellit.
2. Memberikan informasi dimensi buah mindi secara spasial yang dikumpulkan dari sumber benih yang berbeda, dan pertumbuhan bibit mindi di persemaian yang dihasilkan dari sumber benih dan media sapih berbeda.
2
METODE
Waktu dan Tempat Penelitian
3
Tabel 2.1 Lokasi pengambilan sampel penelitian (Yulianti et al. 2011)
No Lokasi Posisi Geografis Ketinggian (m dpl) Kondisi Lapangan
1 Desa Nagrak, Kecamatan
1 250 Berupa tegakan murni mindi, digunakan sebagai peneduh tanaman teh, jarak tanam 4x5 m
Gambar 2.1 Peta lokasi pengambilan sampel penelitian, 1) Nagrak, 2) Sukakarya, 3) Legok Huni, 4) Padasari, 5) Gambung, 6) Babakan Rema
Alat dan Bahan
4
(Kuniyal et al. 2013), 6) pematahan dormansi, perkecambahan, dan pembibitan, dan 7) pengukuran pertumbuhan bibit. Daftar alat dan bahan yang digunakan dalam penelitian disajikan pada Tabel 2.2.
Tabel 2.2 Daftar alat dan bahan penelitian
No Jenis Kegiatan Alat Bahan
2 Elektroforesis cetakan gel agarose,
erlenmeyer, UV
transilluminator, cetakan gel polyakrilamide, kamera
kaliper, timbangan digital Buah mindi
6 Pematahan dormansi, perkecambahan, dan pembibitan
Palu, bak kecambah, polybag,
cangkul, skop pasir
top soil, pasir, arang sekam, pupuk kandang berupa
Sub Penelitian 1. Analisis Keragaman Genetik Pohon Induk
Ekstraksi DNA
Ekstraksi DNA dari daun dilakukan dengan metode CTAB (Cetyl Trimethyl Ammonium Bromide) (Weising et al. 2005 dan Aritonang et al. 2007) yang telah dimodifikasi, dan metode kit (protokol DNA Plant Mini Kit) dari Qiagen (www.qiagen.com) dengan nomor katalog 6235. Langkah pertama dari metode CTAB adalah sampel daun dipotong dengan ukuran 2 cm x 2 cm, kemudian digerus dalam pestel yang bersih dengan penambahan 500 µl buffer ekstrak dan 100 µl PVP 1%. Campuran dimasukkan dalam tube berukuran 1.5 ml.
5 dipisahkan dari fase organik dengan menggunakan mikropipet, kemudian fase air dipindahkan ke dalam tube baru.
Langkah selanjutnya adalah penambahan isopropanol dingin 500 µl dan NaCl 300 µl, lalu disimpan dalam freezer selama 1 jam. Hasil pengendapan disentrifugasi pada kecepatan 10.000 rpm selama 10 menit dan cairan dalam tube dibuang. Kemudian dilakukan proses pencucian DNA dengan penambahan etanol 95% sebanyak 300 µl, lalu disentrifugasi, dan cairan dalam tube dibuang. Proses tersebut dilakukan 2 kali. Pelet DNA yang ada di tube dikeringkan dengan cara disimpan dalam desikator selama 15 menit dengan posisi tube terbalik agar silica gel dalam desikator dapat menyerap cairan yang ada dalam tube.
Uji Kualitas DNA
Langkah pertama pada uji kualitas DNA yaitu menyiapkan gel agarose 1% (0.33 g agarose dalam 33 ml buffer TAE). Untuk proses elektroforesis, ditambahkan buffer TE 50 μl pada pellet DNA lalu disentrifugasi, dan diambil 3 μl DNA ditambahkan 2 μl BJ (Blue Juice) dan dielektroforesis selama 45 menit. Hasil elektroforesis direndam dalam larutan EtBr (Etidium Bromida) selama 15 menit dan difoto pada UV transiluminator model TFX-20.LM (Aritonang et al. 2007).
Mikrosatellit
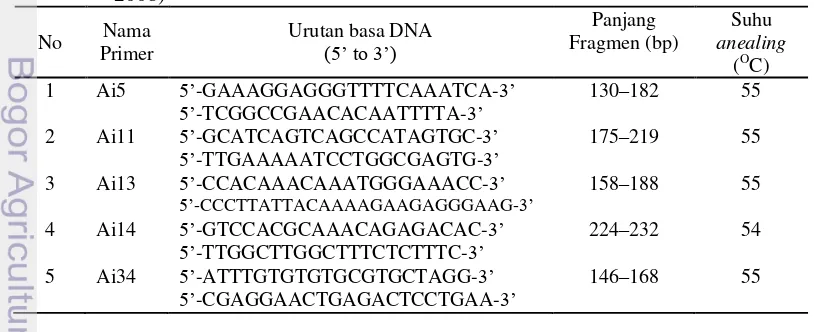
Mikrosatelit merupakan sekuen DNA yang bermotif pendek dan diulang secara tandem dengan 2 sampai 5 unit nukleotida yang tersebar dan meliputi seluruh genom, terutama pada organisme eukariotik (Azrai 2005). PCR mikrosatellit diawali dengan pengenceran DNA. DNA hasil ekstraksi diencerkan 100 kali menggunakan aquabides. Perbandingan antara DNA dan aquabides yaitu 99 μl aquabides dan 1 μl DNA. Primer mikrosatelit untuk jenis mindi belum ada, sehingga dalam penelitian ini menggunakan primer dari jenis lain yang satu famili dengan mindi, yaitu mimba (Azadirachta indica). Primer mikrosatelit yang digunakan dalam penelitian ini disajikan pada Tabel 2.3.
Tabel 2.3 Primer mikrosatelit mimba untuk amplifikasi silang mindi (Boonton et al.
6
VeritiTMThermal Cycler (https://www.appliedbiosystem.com). Komposisi bahan untuk reaksi PCR Mikrosatelit dapat dilihat pada Tabel 2.4.
Elektroforesis mikrosatellit menggunakan gel vertikal poliakrilamid (Wang et al. 2009), dengan komposisi gel seperti pada Tabel 2.5. Akrilamid, bisakrilamid, TBE (1x), dan aquades dicampur dalam tabung erlemeyer dan dikocok selama 15 menit, kemudian pada menit ke 10 dimasukkan temed. Pada menit ke 14 dimasukkan APS, dan pada menit ke 15 larutan dimasukkan ke dalam pasangan kaca akrilamid, kemudian ditunggu sampai larutan memadat menjadi gel (Gambar 2.2).
Tabel 2.4 Empat komponen bahan yang digunakan dalam reaksi PCR
No. Nama Bahan Sampel Reaksi
1 H2O 2.5 μl
2 Green Go Taq Master Mix 7.5 μl
3 Primer F dan R 1.5 μl
4 DNA encer 2 μl
Tabel 2.5 Komposisi pembuatan gel poliakrilamid
Bahan Komposisi
Gambar 2.2 Perakitan kaca gel poliakrilamid
Tabel 2.6 Bahan pewarnaan gel akrilamid dan lama perendaman
Bak Bahan larutan pewarna Lama perendaman gel
7 Kegiatan elektroforesis dilakukan pada 350 V, 40 mA, 80 W selama 75 menit dengan buffer running TBE 1x. Pewarnaan gel dilakukan dengan pewarnaan silver nitrate (Benbouza et al. 2006; Creste et al. 2001). Pewarnaan ini terdiri atas empat bak perendaman. Secara berurutan bak satu sampai empat tertera pada Tabel 2.6. Setelah pewarnaan selesai dilakukan dokumentasi gel poliakrilamid.
Sub Penelitian 2
Variabilitas Dimensi Buah Secara Spasial dan Pertumbuhan Bibit
Dimensi Buah
Secara umum puncak musim buah mindi dijumpai pada bulan Februari hingga April (Pramono 2012). Pengunduhan buah mindi dilaksanakan pada bulan Februari 2014. Buah mindi diunduh pada pohon-pohon yang mempunyai fenotipa baik untuk dijadikan sebagai pohon induk seperti disarankan oleh Mulawarman et al. (2002) (Gambar 2.3). Buah diunduh berdasarkan lokasi (Tabel 2.1) dan nomor pohon induk di lapangan. Semua buah yang sudah terkumpul dilakukan ekstraksi buah sesuai Danu (2005). Total seluruh buah yang terkumpul yaitu 1 644 butir. Setelah buah kering dilakukan pengukuran morfologi buah berupa panjang dan lebar menggunakan kaliper, dan pengukuran massa buah dengan timbangan digital (Kuniyal et al. 2013).
Pengukuran persentase pembentukan benih (seed set) dilakukan dengan memecah buah pada potongan melintang dan membujur dengan palu, kemudian dihitung jumlah ovul dan benih pada tiap buah. Pada masing-masing pohon induk diambil sampel 5 buah, dan diulang 4 kali yang mengacu pada Khan et al. (1999).
Gambar 2.3 Pemilihan pohon induk untuk pengumpulan buah (Mulawarman et al. 2002)
Pematahan Dormansi, Perkecambahan, dan Pembibitan
8
perkecambahan dilaksanakan selama 3 bulan. Setelah semai mempunyai sepasang daun dan relatif seragam dilakukan penyapihan (Sudomo & Santosa 2011).
Rancangan penelitian yang digunakan adalah rancangan petak terpisah (split plot design) yang mengacu pada Mattjik dan Sumertajaya (2013), serta Gomez dan Gomez (2007), dengan perlakuan perbedaan asal benih (faktor utama) dan media penyapihan. Perlakuan media sapih yang digunakan dalam penelitian ini terdiri atas 7 macam media (Danu 2005; Mansur 2010; Sudomo & Santosa 2011), dan asal benih terdiri atas 3 lokasi. Pemilihan 3 lokasi ini didasarkan pada jumlah semai yang cukup untuk semua perlakuan, yaitu dari Nagrak, Babakan Rema, dan Padasari. Masing-masing unit percobaan diulang sebanyak 3 kali, sehingga total unit percobaan ada 63 unit. Desain pengacakan unit percobaan disajikan pada Gambar 2.4.
Pengamatan Pertumbuhan Bibit
Pengukuran variabel pertumbuhan bibit yang dilakukan meliputi tinggi dan diameter (Setyaningsih 2007; Ditjen RLPS 2012). Pengukuran tinggi dan diameter semai dilakukan setiap bulan sampai bibit berumur 4 bulan setelah penyapihan yang mengacu pada Sudomo dan Santosa (2011).
Gambar 2.4 Pengacakan unit percobaan. A menunjukkan asal benih dan P perlakuan media. A1: Nagrak, A2: Babakan Rema, A3: Padasari, P1: tanah, P2: tanah + arang sekam (3:1), P3: tanah + pasir (3:2), P4: tanah + pupuk kandang (3:1), P5: tanah + arang sekam + pasir (3:1:2), P6: tanah + arang sekam + pupuk kandang (3:1:1), P7: tanah + pasir + pupuk kandang (3:2:1).
Analisis Data
Keragaman dalam Populasi
Foto DNA hasil PCR mikrosatelit dianalisis dengan melakukan skoring pola pita yang muncul (Gambar 2.5). Hasil interpretasi foto kemudian dianalisis menggunakan software POPGENE 32 versi 1.31 (Yeh & Yang 1999) dan NTSys versi 2.0 (Rohlf 2008). Dari software tersebut akan ditampilkan beberapa parameter untuk mengetahui keragaman dalam suatu populasi, yaitu nilai PLP (Percentase Locus Polymorphic), Na (Observed number of alleles), Ne (Effective
number of alleles), dan He(Expected Hetegozity).
Keragaman antar Populasi
9 Coordinates Analysis) digunakan software GenAlex Ver 6.5 (Blyton & Nicola 2006).
Gambar 2.5 Cara skoring DNA mikrosatelit (foto pribadi)
Gambar 2.6 Dendogram pengelompokan/klaster genetik
Analisis struktur perbedaan antar populasi digunakan software structure versi 2.3.3 (Evanno et al. 2005; Pritchard et al. 2010; Purba 2012; Dillon et al. 2013). Hasil analisis ditampilkan dalam bentuk grafik seperti disajikan pada Gambar 2.7, warna yang sama menunjukkan bahwa populasi memiliki struktur populasi yang sama.
Populasi 1 2 3 4 5
Gambar 2.7 Tampilan struktur populasi (Purba 2012)
Dimensi Buah, Perkecambahan dan Pertumbuhan Bibit
Persamaan untuk menghitung seed set mengacu pada Hansen dan Molau (1994). Penentuan parameter mutu fisiologi benih menggunakan persamaan Czabator, yaitu: GP (persentase kecambah), GV (nilai kecambah), dan GS
10
(kecepatan kecambah) (Caliskan 2014; Djavanshir & Pourbeik 1976; Gairola et al. 2011).
keterangan:
GV (Germination Value) : nilai kecambah
MDG (Mean Daily Germination) : rata-rata perkecambahan harian PV (Peak Value) : nilai puncak kecambah
DGS (Daily Germination Speed) : kecepatan kecambah harian N (Number of Seed) : jumlah benih yang dikecambahkan GP (Germination Percentage) : persentase kecambah
T (Time) : waktu yang dibutuhkan untuk berkecambah
GS (Germination Speed) : kecepatan kecambah
Analisis yang digunakan untuk menguji pengaruh perlakuan terhadap variabel yang diamati adalah analisis sidik ragam (Mattjik & Sumertajaya 2013). Apabila perlakuan berpengaruh nyata terhadap variabel yang diukur, maka dilanjutkan uji jarak berganda (duncan’s multiple range test-DMRT) dengan bantuan software SAS 9.0 (SAS Institute Inc 2006). Penyajian data berupa boxplot menggunakan software Minitab 15 (Minitab Inc 2007).
3
HASIL DAN PEMBAHASAN
Variabilitas Genetik Pohon Induk
Amplifikasi Silang
Banyak penelitian terkait keragaman genetik yang menggunakan mikrosatelit sebagai penanda genetik (Ng et al. 2009, Lemes et al. 2011, Mantello et al. 2012, Zalapa et al. 2012, Dillon et al. 2013, Goetze et al. 2013). Namun menurut Azrai (2005), mikrosatelit memiliki beberapa kelemahan, misalnya mikrosatelit tidak tersedia pada semua jenis tanaman, sehingga untuk merancang primer baru membutuhkan waktu yang lama dan biaya yang cukup mahal.
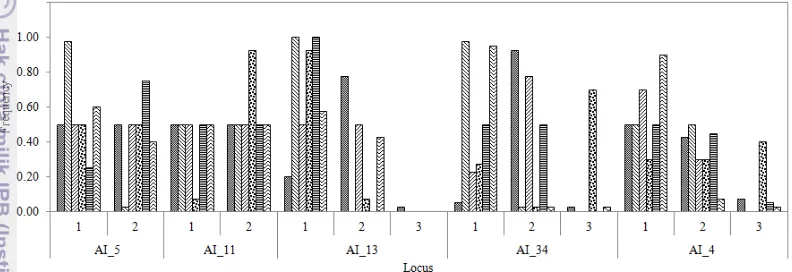
11 DNA target (Aritonang et al. 2007). Hasil amplifikasi silang dari tujuh primer mimba didapatkan lima primer yang polimorfik (Lampiran 1). Frekuensi alel pada tiap primer disajikan pada Gambar 3.1.
Gambar 3.1 menunjukkan bahwa primer Ai-5 dan Ai-11 terdiri atas 2 alel, kemudian primer Ai-13, Ai-34 dan Ai-4 terdiri atas 3 alel. Pada alel ke tiga Ai-13 hanya terdapat alel dari populasi Nagrak, ini disebut private allele (Blyton & Nicola 2006; Kwapata et al. 2013). Hasil skoring alel pada tiap primer disajikan pada Lampiran 2.
Gambar 3.1 Frekuensi alel mindi di hutan rakyat Jawa Barat pada beberapa primer mimba,
Keragaman Genetik dalam Populasi
Kelebihan dari teknik mikrosatelit adalah bersifat polimorfik (Varshney et al. 2007), dengan tingkat reproduksi yang tinggi, dan merupakan penanda kodominan (Rajora dan Rahman 2001; Weising et al. 2005). Perez et al. (2005) melaporkan mikrosatelit merupakan penanda genetik yang sangat baik untuk studi keragaman genetik dalam berbagai tanaman.
Finkeldey (2005) menyatakan bahwa keragaman genetik merupakan perbedaan gen yang terkandung dalam individu suatu populasi dan berhubungan dengan kemampuan beradaptasi suatu individu dalam mengalami perubahan selama proses perkembangan. Keragaman genetik dapat diwariskan kepada keturunannya dan terjadi karena adanya rekombinasi genetik sebagai akibat adanya
persilangan-persilangan dan adanya mutasi. Secara umum variasi genetik dapat diukur dengan
dua parameter, yaitu dalam populasi dan antar populasi.
Peubah yang digunakan untuk mencirikan variasi genetik dalam populasi yaitu Presentase Lokus Polimorfik (PLP), dan rata-rata jumlah alel per lokus (A/L), dan variasi genetik (He) (Finkeldey 2005). Nilai parameter keragaman
genetik dalam populasi mindi disajikan pada Tabel 3.1.
Tabel 3.1 menunjukkan rata-rata alel yang diamati sebasar 2.200, PLP sebesar 93.33%, dan keragaman genetik dalam populasi (He) sebesar 0.3656. Nilai
keragaman genetik tersebut tergolong pada keragaman sedang. Hal ini sesuai dengan Rambey (2011) yang menyatakan bahwa mindi di hutan rakyat Garut memiliki nilai keragaman genetik sebesar 0,3730 dikategorikan pada keragaman sedang.
12
populasi yang diamati keragaman genetik tertinggi adalah populasi Padasari (He=0.4537), dan terendah adalah Babakan Rema (He=0.2195).
Tabel 3.1 Nilai parameter keragaman genetik dalam populasi mindi di hutan
urutan lokasi berdasarkan ketinggian (rendah ke tinggi), N: jumlah individu, Na: jumlah alel yang teramati,
Ne: jumlah alel efektif, He: heterozigositas/keragaman genetik, PLP: Persentase Lokus Polimorfik
Tinggi rendahnya keragaman dalam populasi dapat dipengaruhi oleh pola sebaran tanaman, khususnya di hutan rakyat yang umumnya tanaman tersebar dengan jumlah yang terbatas (Hamid et al. 2008). Tegakan mindi di Babakan rema merupakan areal yang berdekatan dengan perkebunan teh, sehingga tegakan mindi di lahan masyarakat berasal dari area perkebunan yang hanya dikelola satu pengelola (Yulianti 2011). Berbeda dengan tegakan mindi di Padasari yang tersebar pada berbagai tipe dan pemilik lahan dengan sejarah penanaman bervariasi, dan cenderung menghasilkan musim buah yang bervariasi, serta diduga variasi genetiknya lebih tinggi (Pramono 2012). Selain itu, Pramono (2012) juga melaporkan bahwa musim buah tegakan mindi di Babakan Rema tergolong serentak. Musim buah yang serentak ini diduga karena populasi kuningan mempunyai keragaman genetik yang rendah.
Keragaman Genetik antar Populasi
Peubah yang digunakan untuk mencirikan variasi genetik antar populasi menurut Finkeldey (2005) yaitu pembagian variasi genetik (Fst atau Gst), jarak
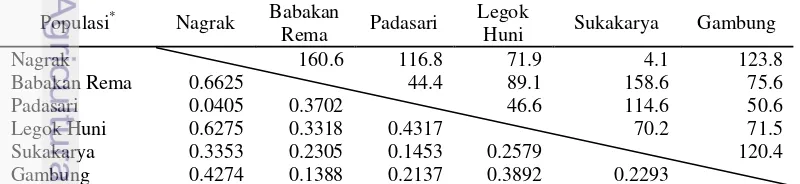
genetik, dan analisis klaster/kelompok. Perbedaan genetik dari dua atau lebih populasi pada umumnya dianalisis dengan sebuah matrik dimana elemen-elemennya berupa jarak genetik dan pasangan kombinasi dari masing-masing populasi (Finkeldey 2005). Jarak genetik mengukur perbedaan struktur genetik antar populasi pada suatu lokus gen tertentu. Jarak genetik antar populasi mindi di hutan rakyat disajikan pada Tabel 3.2.
Tabel 3.2 Jarak genetik dan geografis populasi mindi di hutan rakyat Jawa Barat
13
Tabel 3.2 menunjukkan bahwa jarak genetik terjauh adalah populasi Nagrak dan Babakan Rema, sedangkan jarak genetik terdekat adalah Nagrak dan Padasari. Sehingga Nagrak dan Babakan Rema mempunyai kekerabatan yang jauh, sedangkan Nagrak dan Padasari mempunyai kekerabatan yang dekat. Gambar 3.2 memperlihatkan tidak ada korelasi antara jarak geografis dan jarak genetik, sesuai yang dilaporkan oleh Yulianti (2011).
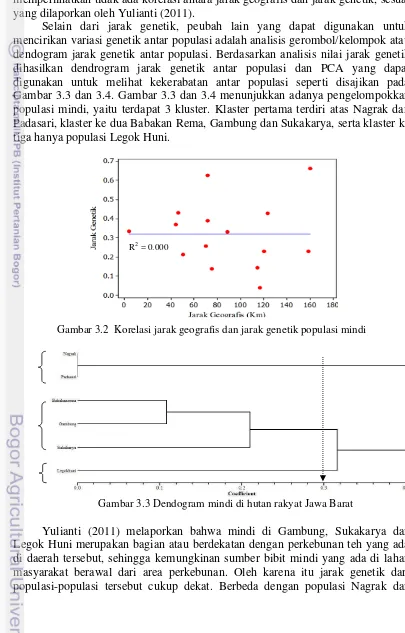
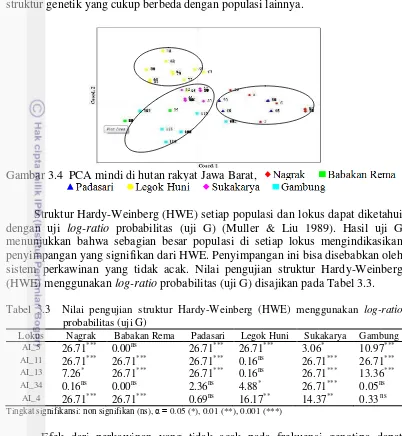
Selain dari jarak genetik, peubah lain yang dapat digunakan untuk mencirikan variasi genetik antar populasi adalah analisis gerombol/kelompok atau dendogram jarak genetik antar populasi. Berdasarkan analisis nilai jarak genetik dihasilkan dendrogram jarak genetik antar populasi dan PCA yang dapat digunakan untuk melihat kekerabatan antar populasi seperti disajikan pada Gambar 3.3 dan 3.4. Gambar 3.3 dan 3.4 menunjukkan adanya pengelompokkan populasi mindi, yaitu terdapat 3 kluster. Klaster pertama terdiri atas Nagrak dan Padasari, klaster ke dua Babakan Rema, Gambung dan Sukakarya, serta klaster ke tiga hanya populasi Legok Huni.
Gambar 3.2 Korelasi jarak geografis dan jarak genetik populasi mindi
Gambar 3.3 Dendogram mindi di hutan rakyat Jawa Barat
Yulianti (2011) melaporkan bahwa mindi di Gambung, Sukakarya dan Legok Huni merupakan bagian atau berdekatan dengan perkebunan teh yang ada di daerah tersebut, sehingga kemungkinan sumber bibit mindi yang ada di lahan masyarakat berawal dari area perkebunan. Oleh karena itu jarak genetik dari populasi-populasi tersebut cukup dekat. Berbeda dengan populasi Nagrak dan
14
Padasari, dimana di daerah ini tidak terdapat perkebunan, sehingga mempunyai struktur genetik yang cukup berbeda dengan populasi lainnya.
Gambar 3.4 PCA mindi di hutan rakyat Jawa Barat,
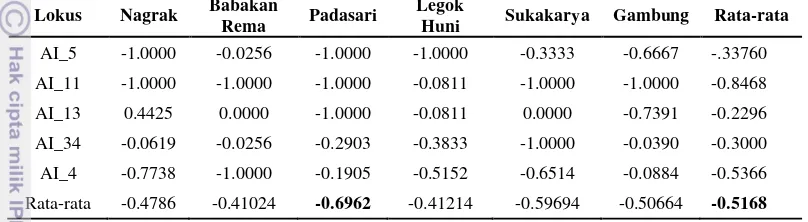
Struktur Hardy-Weinberg (HWE) setiap populasi dan lokus dapat diketahui dengan uji log-ratio probabilitas (uji G) (Muller & Liu 1989). Hasil uji G menunjukkan bahwa sebagian besar populasi di setiap lokus mengindikasikan penyimpangan yang signifikan dari HWE. Penyimpangan ini bisa disebabkan oleh sistem perkawinan yang tidak acak. Nilai pengujian struktur Hardy-Weinberg (HWE) menggunakan log-ratio probabilitas (uji G) disajikan pada Tabel 3.3.
Tabel 3.3 Nilai pengujian struktur Hardy-Weinberg (HWE) menggunakan log-ratio probabilitas (uji G)
Lokus Nagrak Babakan Rema Padasari Legok Huni Sukakarya Gambung
AI_5 26.71*** 0.00ns 26.71*** 26.71*** 3.06* 10.97*** AI_11 26.71*** 26.71*** 26.71*** 0.16ns 26.71*** 26.71*** AI_13 7.26* 26.71*** 26.71*** 0.16ns 26.71*** 13.36*** AI_34 0.16ns 0.00ns 2.36ns 4.88* 26.71*** 0.05ns
AI_4 26.71*** 26.71*** 0.69ns 16.17** 14.37** 0.33 ns Tingkat signifikansi: non signifikan (ns), α = 0.05 (*), 0.01 (**), 0.001 (***)
Efek dari perkawinan yang tidak acak pada frekuensi genotipa dapat diukur menggunakan perbandingan frekuensi heterozigositas harapan HWE, yang diasumsikan kawin acak, dengan frekuensi heterozigositas yang diamati dalam suatu populasi. Besaran tersebut disebut indeks fiksasi (Fis) (Hamilton 2009).
Hasil nilai indeks fiksasi setiap lokus disajikan pada Tabel 3.4.
Tabel 3.4 memperlihatkan rata-rata nilai Fis pada semua lokus dan populasi
15 Populasi Padasari mempunyai nilai Fis paling tinggi (-0.6962), hal ini bisa
disebabkan oleh adanya efek Wahlund (Syamsuwida 2012). Menurut Isoda dan Rimbawanto (2001), efek Wahlund merupakan skema yang dapat meningkatkan heterosigositas. Namun menurut Isik dan Kaya (2010), kekurangan heterozigositas pada jenis pinus (Pinus brutia) merupakan akibat dari efek Wahlund, assortative mating, dan atau seleksi homozigot serta silang dalam.
Tabel 3.4 Nilai indeks fiksasi setiap lokus pada microsatellit populasi mindi
Lokus Nagrak Babakan
Rema Padasari
Legok
Huni Sukakarya Gambung Rata-rata
AI_5 -1.0000 -0.0256 -1.0000 -1.0000 -0.3333 -0.6667 -.33760 AI_11 -1.0000 -1.0000 -1.0000 -0.0811 -1.0000 -1.0000 -0.8468 AI_13 0.4425 0.0000 -1.0000 -0.0811 0.0000 -0.7391 -0.2296 AI_34 -0.0619 -0.0256 -0.2903 -0.3833 -1.0000 -0.0390 -0.3000 AI_4 -0.7738 -1.0000 -0.1905 -0.5152 -0.6514 -0.0884 -0.5366 Rata-rata -0.4786 -0.41024 -0.6962 -0.41214 -0.59694 -0.50664 -0.5168
Tabel 3.5 Nilai pengujian struktur inbreeding menggunakan uji chi square (X2)
Lokus Nagrak Babakan Rema Padasari Legok Huni Sukakarya Gambung
AI_5 19.00*** 0.00ns 19.00*** 19.00*** 1.96ns 8.26** AI_11 19.00*** 19.00*** 19.00*** 0.09ns 19.00*** 19.00*** AI_13 8.30* 19.00*** 19.00*** 0.09ns 19.00*** 10.21** AI_34 0.09ns 0.00ns 1.47ns 3.32ns 19.00*** 0.03ns
AI_4 19.00*** 19.00*** 0.56ns 10.33* 12.09** 0.18ns Tingkat signifikansi: non signifikan (ns), α = 0.05 (*), 0.01 (**), 0.001 (***)
Berdasarkan tampilan struktur populasi (Gambar 3.5), dapat dikatakan hampir semua populasi mempunyai struktur yang sama. Hal ini menunjukkan bahwa gen pool semua populasi mindi dalam penelitian diduga berasal dari satu sumber.
Gambar 3.5 Tampilan struktur populasi mindi
Variabilitas Dimensi Buah
Ukuran Buah
16
yang berasal dari Nagrak dan Padasari. Ukuran panjang dan lebar buah mindi pada tiap lokasi disajikan pada Tabel 3.6.
Secara umum Tabel 3.6 menunjukkan bahwa semakin tinggi tempat, maka ukuran buah akan semakin kecil. Buah dari Sukakarya dan Gambung mempunyai ukuran yang lebih kecil dibandingkan dengan lokasi lainnya. Dilihat dari ketinggian tempat, kedua lokasi tersebut memiliki ketinggian yang lebih tinggi (Tabel 2.1). Orwa et al. (2009) melaporkan bahwa mindi mampu beradaptasi pada ketinggian 0-1 800 m dpl. Kecilnya ukuran buah mindi yang berasal dari Sukakarya dan Gambung bisa disebabkan oleh adaptasi suhu yang rendah pada ketinggian lokasi tersebut.
Tabel 3.6 Nilai rata-rata panjang dan lebar buah mindi pada tiap lokasi
Lokasi*
*) urutan lokasi berdasarkan ketinggian (rendah ke tinggi)
**)huruf di samping angka menunjukkan perbandingan nilai tengah berdasarkan uji Duncan pada α = 0.05
Massa Buah
Rata-rata massa buah mindi pada tiap lokasi didapatkan 0.37-0.40 g. Hasil pengukuran massa buah menunjukkan tidak ada perbedaan yang nyata pada tiap lokasi. Jika massa buah tersebut dikonversi menjadi jumlah buah tiap kg, maka akan didapatkan 2 500 - 2 700 butir. Menurut Danu (2005), jumlah buah mindi kering sebanyak 1 286 butir kg-1. Sedangkan menurut Orwa et al. (2009), jumlah buah mindi tiap kg 470 - 2 800 butir. Sehingga dapat dikatakan hasil pengukuran massa buah mindi dalam studi ini masih dalam kisaran normal. Hasil pengukuran menunjukkan bahwa buah paling berat yaitu buah yang berasal dari Nagrak seperti disajikan pada Tabel 3.7.
Tabel 3.7 Nilai rata-rata massa buah mindi pada tiap lokasi
Lokasi* Kadar Air
urutan lokasi berdasarkan urutan ketinggian (rendah ke tinggi)
**)
huruf di samping angka menunjukkan perbandingan nilai tengah berdasarkan uji Duncan pada taraf nyata 0.05
Persentase pembentukan benih (seed set)
17 benih yang ada pada tiap kg buah. Penampang melintang dan membujur buah mindi disajikan pada Gambar 3.6. Hasil pengamatan menunjukkan bahwa rata-rata seed set tertinggi yaitu pada buah yang berasal dari Nagrak dengan rata-rata 85.47 ± 15.09%. Sebaran seed set mindi pada tiap lokasi disajikan pada Gambar 3.7. Tabel 2 dan 3 menunjukkan bahwa nilai koefisien variasi (CV) pada dimensi buah berkisar 4.82-14.73% yang dikategorikan keragaman sedang (Ginwal et al. 2005).
Gambar 3.6 Penampang dan bagian-bagian buah mindi, (a) penampang membujur dan melintang (foto pribadi), (b) bagian-bagian buah mindi, ovl:ovul, cp:karpel, exc:eksocarp, msc:mesocarp, end:endocarp (Syamsuwida et al. 2012)
Gambar 3.7 Sebaran seed set mindi pada tiap lokasi Perkecambahan
Sutopo (2012) melaporkan bahwa persentase kecambah umumnya digunakan sebagai parameter viabilitas benih. Hasil pengamatan menunjukkan bahwa semua benih pada hari ke-112 tidak mengalami penambahan persentase kecambah yang ditandai garis mulai mendatar (Gambar 3.8).
Gambar 3.8 menunjukkan bahwa benih dari Nagrak dan Padasari pada hari ke-7 sudah mulai berkecambah, kemudian disusul benih dari Babakan Rema yang berkecambah pada hari ke-14. Pengamatan sampai hari ke-56 baru menunjukkan benih dari Sukakarya dan Gambung mulai berkecambah. Pada Tabel 3.6 terlihat bahwa benih dari Sukakarya dan Gambung mempunyai ukuran yang lebih kecil. Khan et al. (1999) dan Kuniyal et al. (2013) melaporkan bahwa benih dari buah yang kecil dan ringan akan mempunyai laju perkecambahan yang lebih lama dibandingkan dengan buah yang lebih besar dan berat.
Persentase kecambah menunjukkan jumlah kecambah normal yang dihasilkan oleh benih pada kondisi lingkungan tertentu (Sutopo 2012). Dari lima sumber benih, persentase perkecambahan tertinggi yaitu benih dari Nagrak, mencapai 65.10% (Gambar 3.9).
(a )
18
Gambar 3.8 Nilai persentase perkecambahan benih mindi dari awal sampai akhir pengamatan,
Gambar 3.9 Persentase perkecambahan benih mindi dari lima sumber benih
Tabel 3.8 Rekapitulasi nilai GP (persentase kecambah), GV (nilai kecambah), dan GS (kecepatan kecambah) benih mindi pada tiap lokasi
Waktu Pengamatan Asal Beniha GP
(%)
urutan lokasi berdasarkan urutan ketinggian (rendah ke tinggi), b persamaan Gzabator (Djavanshir & Pourbeik 1976), c persamaan modifikasi (Caliskan 2014)
19 (2013) melaporkan bahwa benih dari buah yang kecil dan ringan akan memiliki persentase perkecambahan yang rendah. Selain itu, Khan (2004) juga melaporkan bahwa massa buah berkorelasi positif dengan persentase perkecambahan, semakin berat suatu buah maka persentase perkecambahannya akan lebih tinggi.
Hasil penelitian ini menunjukkan bahwa benih dari Nagrak mempunyai GP, GV dan GS yang paling tinggi diantara benih lainnya (Tabel 3.8). Jika dikaitkan dengan ukuran buah (Tabel 3.6), maka secara umum ukuran buah berbanding lurus dengan GP, GV dan GS. Semakin besar ukuran buah maka semakin besar pula nilai GP, GV dan GS suatu benih.
Pertumbuhan Bibit Mindi
Hasil analisis varian menunjukkan bahwa media penyapihan berpengaruh sangat nyata (α = 0.05) terhadap pertumbuhan tinggi dan diameter bibit mindi. Namun interaksi antara media dengan asal benih tidak berpengaruh nyata. Analisis varian asal benih dan kombinasi media pada pertumbuhan tinggi dan diameter bibit mindi disajikan pada Tabel 3.9. Untuk nilai CV pada pengamatan pertumbuhan tinggi dan diameter bibit mindi didapatkan sebesar 11.79-21.77%. Nilai tersebut dikategorikan variasi yang sedang (Ginwal et al. 2005). Dokumentasi pengamatan pertumbuhan bibit mindi di persemaian disajikan pada Lampiran 3.
Tabel 3.9 Analisis varian pengaruh media dan asal benih pada pertumbuhan tinggi dan
diameter bibit mindi pada taraf α = 0.05 dan 0.01
Sumber keragaman p < 0.05
Tinggi Diameter
media 0.0001** 0.0001**
ulangan 0.0820ns 0.8715ns
asal benih 0.0287** 0.0029**
interaksi media dan asal benih 0.3471ns 0.1989ns
**
sangat nyata, ns tidak nyata
Pengaruh Asal Benih pada Pertumbuhan Bibit Mindi
Hasil pengamatan menunjukkan bahwa pertambahan diameter dan tinggi pada bibit mindi paling besar adalah bibit yang sumber benihnya dari Nagrak. Uji lanjut Duncan dengan
α
= 0.05 menunjukkan pertambahan tinggi dan diameter bibit dari Nagrak berbeda nyata dengan bibit dari Padasari dan Babakan Rema (Gambar 3.8).20
Gambar 3.10 Pertambahan diameter dan tinggi bibit mindi umur 4 bulan dari sumber benih yang berbeda
Pengaruh Media Sapih pada Pertumbuhan Bibit Mindi
Hasil penelitian menunjukkan bahwa media P4 (tanah + pupuk kandang) memberikan pertambahan tinggi yang relatif sama baiknya dengan penggunaan media P6 (tanah + arang sekam + pupuk kandang) dan media P7 (tanah + pasir + pupuk kandang). Sedangkan media yang paling baik untuk pertambahan diameter adalah P6 (Gambar 3.11).
Gambar 3.11 Pengaruh media sapih terhadap pertambahan tinggi dan diameter bibit mindi selama 4 bulan, P1: tanah, P2: tanah + arang sekam (3:1), P3: tanah + pasir (3:2), P4: tanah + pupuk kandang (3:1), P5: tanah + arang sekam + pasir (3:1:2), P6: tanah + arang sekam + pupuk kandang (3:1:1), P7: tanah + pasir + pupuk kandang (3:2:1). Huruf-huruf di atas balok menunjukkan pembandingan nilai tengah antar perlakuan berdasarkan uji Duncan pada taraf nyata 0.05
Sudomo dan Santosa (2011) melaporkan bahwa media campuran tanah dan pupuk kandang memberikan pertambahan tinggi bibit mindi yang paling besar,
21 karena pemberian pupuk kandang akan menjamin ketersediaan unsur hara dan perbaikan aerasi media. EPA (2007) melaporkan bahwa kandungan nitrogen dari pupuk kandang biasanya langsung dapat tersedia bagi tanaman. Oleh karena itu, pertumbuhan tinggi dan diameter bibit mindi lebih cepat pada media yang mengandung pupuk kandang.
Potensi Sumber Benih Mindi di Hutan Rakyat Jawa Barat
Sumber benih merupakan areal atau tempat dimana koleksi benih dilakukan. Perbedaan genetik antar sumber benih akan berpengaruh terhadap keberhasilan dalam pembangunan hutan (Na’iem 2011). Menurut Santoso (2011), salah satu permasalahan dalam penanaman adalah sumber benih dengan kualitas genetik tinggi yang masih sangat sedikit. Roshetko et al. (2004), melaporkan bahwa sumber benih hendaknya dibangun dengan mutu setinggi mungkin dan keragaman genetik setinggi mungkin. Mutu genetik perlu diperhatikan karena menyangkut daya adaptasi dengan lingkungan tempat tumbuh. Semakin tinggi keragaman genetiknya semakin besar peluang tanaman untuk beradaptasi dengan lingkungannya. Keragaman genetik yang rendah pada suatu populasi akan menyebabkan inbreeding (silang dalam). Inbreeding yang terjadi terus-menerus akan menyebabkan kepunahan dalam jangka panjang (Hamilton 2009).
Hasil analisis genetik menunjukkan tingkat keragaman genetik yang ada di dalam maupun antar populasi mindi dapat mengindikasikan bagaimana sumberdaya genetik tanaman mindi di hutan rakyat Jawa Barat. Berdasarkan hasil pengolahan data, keragaman genetik tanaman mindi rata-rata sebesar 0.3656. Hal ini menunjukkan keragaman genetik tanaman mindi di hutan rakyat tergolong sedang. Sehingga mindi di enam populasi ini masih dapat direkomendasikan sebagai sumber benih. Pembangunan hutan rakyat sebaiknya juga memperhatikan kekerabatan antar populasi. Populasi yang berkekerabatan dekat hendaknya tidak ditanam berdekatan, misalnya populasi Nagrak dan Padasari. Hal ini dimaksudkan untuk mengurangi kemungkinan inbreeding. Berbeda halnya dengan populasi Nagrak dan Legok Huni yang mempunyai kekerabatan jauh, sumber benih dari kedua populasi tersebut dapat ditanam secara berdekatan.
Keutuhan sumberdaya genetik mindi dapat dilakukan dengan konservasi in situ dan ex situ. Konservasi in situ merupakan metode dan alat untuk melindungi spesies, variasi genetik dan habitat dalam ekosistem aslinya. Sedangkan konservasi ex situ meliputi metode dan alat untuk melindungi spesies dan organisme mikro serta varietas genetik di luar habitat/ekosistem aslinya. Kegiatan yang umum dilakukan antara lain penyimpanan atau pengklonan dengan alasan: (1) habitat mengalami kerusakan akibat konversi, dan (2) materi tersebut dapat digunakan untuk penelitian, percobaan, pengembangan produk baru atau pendidikan lingkungan, misalnya dengan pembangunan kebun raya, koleksi mikologi, museum, bank biji, dan koleksi kultur jaringan.
22
4
SIMPULAN DAN SARAN SimpulanKeragaman genetik dalam populasi (He) mindi dengan penanda mikrosatellit
didapatkan sebesar 0.3656. Nilai keragaman genetik tersebut tergolong pada keragaman sedang, dengan keragaman genetik tertinggi adalah populasi Padasari (He= 0.4537) dan terendah adalah Babakan Rema (He= 0.2195). Berdasarkan nilai
jarak genetik, analisis gerombol (dendogram), dan PCoA menunjukkan ada 3 pengelompokkan populasi mindi. Klaster pertama terdiri atas Nagrak dan Padasari, klaster ke dua Babakan Rema, Gambung dan Sukakarya, serta klaster ke tiga hanya populasi Legok Huni. Jarak genetik terjauh adalah populasi Nagrak dan Babakan Rema, sedangkan jarak genetik terdekat adalah Nagrak dan Padasari. Sehingga Nagrak dan Babakan Rema mempunyai kekerabatan yang jauh, sedangkan Nagrak dan Padasari mempunyai kekerabatan yang dekat. Dilihat dari tampilan struktur populasi hampir semua populasi mempunyai struktur yang sama. Hal ini menunjukkan bahwa gen pool semua populasi mindi dalam penelitian diduga berasal dari satu sumber.
Variabilitas dimensi buah mindi secara spasial yang dikumpulkan dari lima sumber benih berbeda, dan pertumbuhan bibit mindi di persemaian tergolong sedang. Buah mindi dari Nagrak mempunyai ukuran, berat, dan seed set yang paling tinggi, serta persentase, nilai dan kecepatan kecambah paling tinggi. Hasil pengamatan pertumbuhan bibit mindi dari sumber benih dan media sapih berbeda menunjukkan bahwa bibit dari Nagrak memiliki pertumbuhan lebih cepat. Media sapih yang mengandung pupuk kandang berupa kotoran sapi memberikan pertumbuhan paling cepat.
Saran
Pembangunan hutan rakyat sebaiknya memperhatikan kekerabatan genetik. Populasi yang berkekerabatan dekat hendaknya tidak ditanam berdekatan, misalnya populasi Nagrak dan Padasari. Hal ini dimaksudkan untuk mengurangi kemungkinan inbreeding. Berbeda halnya dengan populasi Nagrak dan Legok Huni yang mempunyai kekerabatan jauh, sumber benih dari kedua populasi tersebut dapat ditanam secara berdekatan. Perlu dilakukan infusi genetik untuk memperluas variasi genetik mindi.
DAFTAR PUSTAKA
Abasolo MA, Fernando ES, Borromeo TH, Hautea DM. 2009. Cross-species amplification of shorea microsatellite DNA markers in Parashorea malaanonan (dipterocarpaceae). Philippine J of Science. 138(1):23-28. Aritonang KV, Siregar IZ, Yunanto T. 2007. Manual Analisis Genetik Tanaman
Hutan di Laboratorium Silvikultur Fakultas Kehutanan Institut Pertanian Bogor. Bogor (ID): Fakultas Kehutanan IPB.
23 Benbouza H, Jacquemin JM, Baudoin JP, Mergeai G. 2006. Optimization of a reliable, fast, cheap and sensitive silver staining method to detect SSR markers in polyacrylamide gels. Biotechnol. Agron. Soc. Environ. 10(2):77-81.
Blade C, Vallejo VR. 2008. Seed mass effects on performance of Pinus halepensis Mill. seedlings sown after fire. J Forest Ecology and Management. 255:2362–2372. doi:10.1016/j.foreco.2007.12.039.
Blyton MDJ, Nicola SF. 2006. A Comprehensive Guide to: GenAlEx 6.5. Australia (AU): Australian National University.
Boonton C, Pandey M, Chaangtragon S. 2008. Isolation and characterization of microsatellite markers in Indian neem (Azadirachta indica var. indica A. Juss) and cross-amplification in Thai neem (A. indica var. siamensisValenton). J ConservGenet.doi:10.1007/s10592-008-9610-5. Bramasto RGA. 2011. Hubungan faktor tempat tumbuh dengan produksi buah
mindi (Melia azedarach Linn.) di Hutan Rakyat Jawa Barat [tesis]. Bogor (ID): Institut Pertanian Bogor.
Caliskan S. 2014. Germination and seedling growth of holm oak (Quercus ilex L.): effects of provenance, temperature, and radicle pruning. iForest. 7:103-109. doi.10.3832/ifor0967-007.
Costa BF, Rodrigues LA, Ruas LA, Vieiera BG, Conson ARO, Ruas PM. 2013. Characterization of nine microsatellite loci for the tree species Parapiptadenia rigida (Fabaceae-Mimosoideae) and their transferability. Genetics and Molecular Research. 11(3):2338-2342.
Creste S, Neto AT, Figueira A. 2001. Detection of single sequence repeat polymorphisms in denaturing polyacrylamide sequencing gels by silver staining. Plant Molecular Biology Reporter. 19:299–306.
Danu. 2005. Mindi (Melia azedarach). Di dalam: Rohandi D, Djam’an DF, Aminah A, Sitorus R, editor. Atlas Benih Tanaman Hutan Indonesia. Bogor (ID): Balai Penelitian Teknologi Perbenihan. hlm 70-73.
Dillon NL, Ian SEB, Wright CL, Hucks L, Innes DJ, Dietzgen RG. 2013. Genetic diversity ot the Australian National Mango Genebank. J Scientia Horticulturae. 150:213-226.
[Ditjen RLPS] Direktorat Jenderal Rehabilitasi Lahan dan Perhutanan Sosial. 2012. Standar Mutu Bibit Tanaman Hutan. Jakarta (ID): Direktorat Perbenihan Tanaman Hutan.
Djavanshir K, Pourbeik H. 1976. Germination value: a new formula. Silvae Genetica. 25:79-83.
EPA [Environmental Protection Agency]. 2007. The Use of The Soil Amendments for Remediation, Revitalization, and Reuse. Cincinnati (US): National Service Center for Environmental Publications.
Evanno G, Regnaud S, Goudet J. 2005. Detecting the number of clusters of individuals using the software structure: a simulation study. Molecular Ecology.14:2611–2620.
24
Fox GA. 2003. Assortative mating and plant phenology: evolutionary and practical consequences. Evolutionary Ecology Research. 5:1–18.
Gairola KC, Nautiyal AL, Dwivedi AK. 2011. Effect of temperatures and germination media on seed germination of Jatropha curcas Linn.. Adv.Biores. 2(2):66-71.
Ginwal HS, Phartyal SS, Rawat PS, Srivastava RL. 2005. Seed source variation in morphology, germination and seedling Growth of Jatropha curcas Linn. in Central India. Silvae Genetica. 54(2):76-80.
Goetze M, Louzada RB, Wanderley MGL, Souza LM, Bered F, Silva CP. 2013. Development of microsatellite marker for genetic diversity analysis of Aechmea caudate (Bromeliaceae) and cross-species amplification in other bromeliads. J Biochemical Systematics and Eology. 48:194-198.
Gomez KA, Gomez AA. 2007. Prosedur Statistik untuk Penelitian Pertanian. Sjamsuddin E, Baharsjah JS, penerjemah. Jakarta (ID): UI Press. Terjemahan dari: Statistical Procedures for Agricultural Research.
Gregorius HR. 1989. Characterization and Analysis of Mating Systems. Witzenhausen (DE): Ekopan Verlag.
Hamid AH, Ab-Shukor NA, Senin AL. 2008. Morphometric and genetic variation of six seed sources of Azadirachta excelsa (Jack). Jacobs. J of Biological Science. 8:702-712.
Hamilton MB. 2009. Population Genetics. United State (US): Wiley-Blacwell. Hansen JEL, Molau U. 1994. Pollination biology, mating system, and seed set in a
Danish population of Saxifraga granulata. Nordic J of Botany. 14 (3):257-268.
Hartati D, Rimbawanto A, Taryono, Sulistyaningsih, Widyatmoko. 2007. Pendugaan keragaman genetik di dalam dan antar provenan pulai (Alstonia scholaris (L.) R. Br.) menggunakan penanda RAPD. J Pemuliaan Tanaman Hutan. 2:89-98.
Isik K, Kaya N. 2010. Genetic identification of clones and the genetic structure of seed crops in a Pinus brutia seed orchard. Turk J Agric For. 34:127-134. Isoda K, Rimbawanto A. 2001. Genetic structure of Shorea leprosula in a single
population revealed by microsatellite markers. Di dalam: Thielges B, Satrapradja S, Rimbawanto A, editor. In situ and Ex situ Conservation of Commercial Tropical Trees. Proceedings of the International Conference on ex situ and in situ Conservation of Commercial Tropical Trees; 2001 June 11-13; Yogyakarta, Indonesia. Yokohama (JP): International Tropical Timber Organization (ITTO).
Karyono, Hariatno. 2001. Peluang dan tantangan pemasaran kayu mindi (Melia azedarach L.): studi kasus di Bogor Jawa Barat. J Sosial Ekonomi. 2(2):77-86.
Khan ML, Bhuyan P. Shankar U, Todaria N. 1999. Seed germination and seedling fitness in Mesua ferrea L. in relation to fruit size and seed number per fruit. J Acta Oecologica. 20:599−606.
25 Kuniyal CP, Purohit V, Butola JS, Sundriyal RC. 2013. Seed size correlates seedling emergence in Terminalia bellerica. South African J of Botany. 87:92-94.
Kwapata K, Mwase WF, Bokosi JM, Kwapata MB, Munyenyemb P. 2013. Genetic diversity of Annona senegalensis Pers.populations as revealed by simple sequence repeats (SSRs). African J of Agriculture. 1(4):049-056 Lemes MR, Esashika T, Gaoue OG. 2011. Microsatellite for mahagonies: twelve
new loci for Swietenia macrophylla and its high transferability to Khaya senegalensis. American J of Botani. 5:207-209.doi: 10.3732/ajb.1100074. Mansur I. 2010. Teknik Silvikultur untuk Reklamasi Lahan Bekas Tambang.
Bogor (ID): Seameo Biotrop.
Mantello CC, Suzuki FI, Souza LM, Goncalves PS, Souza AP. 2012. Microsatellite marke development for the rubber tree (Hevea brasiliensis): characterization and cross-amplification in wild Hevea species. BMC Research Notes. 5:329-336.
Mattjik AA, Sumertajaya IM. 2013. Perancangan Percobaan dengan Aplikasi SAS dan Minitab. Bogor (ID): IPB Press.
Minitab Inc. 2007. Meet Minitab 15 for Windows. USA (US): Minitab Inc.
Mulawarman, Roshetko JM, Sasongko SM, Irianto D. 2002. Pengelolaan Benih Pohon, Sumber Benih, Pengumpulan dan Penanganan Benih: Pedoman Lapang untuk Petugas Lapang dan Petani. Bogor (ID): International Centre for Research in Agroforestry (ICRAF).
Muller SG, Liu YQ. 1989. Inferences on the reproductive system of Cunninghamia lanceolata. Forest Ecology and Management. 29:187-198. Na’iem M. 2011. Aspek ilmiah pembangunan sumber benih untuk mendukun
kebijakan penanaman satu milyar pohon. Di dalam: Rimbawanto A, Leksono B, Widyatmoko AYPBC, editor. Peran Sumber Benih Unggul dalam Mendukung Keberhasilan Penanaman Satu Milyar Pohon. Seminar Nasional Pembangunan Sumber Benih; 2011 Jun 30; Yogyakarta, Indonesia. Yogyakarta (ID): Balai Besar Penelitian Bioteknologi dan Pemuliaan Tanaman Hutan.
Ng KKS, Lee SL, Ueno S. 2009. Impact of selective logging on genetic diversity of two tropical tree species with contrasting breeding system using direct comparison and simulation methods. J Forest Ecology and Managemet. 257:107-116.
Nurtjahjaningsih ILG. 2007. Potensi populasi infusi genetik untuk memperluas variasi genetik kebeun benih semai Pinus merkusii di Jember. J Pemuliaan Tanaman Hutan. 3(2):73-81.
Orwa C, Mutua A, Kindt R, Jamnadass R, Simons A. 2009. Agroforestree Database: a tree reference and selection guide version 4.0. Agroforestry Database (4.0) [internet]. [diunduh 2014 Agustus 27]. Tersedia pada: http://www.worldagroforestry.org/treedb/AFTPDFS/Melia_azedarach.pdf. Perez RS, Ruiz D, Dicenta F, Egea J, Gomez PM. 2005. Application of simple
sequence repeat (SSR) markers in apricot breeding: molecular characterization, protection, and genetic relationships. J Scientia Horticulturae. 103:305–315.
26
Pramono AA, Danu, Rohandi. 2012. Zona sebaran populasi mindi (Melia azedarach Lin.) di Jawa Barat dan potensi tegakannya untuk sumber benih. Info Benih. 16(2):55-62.
Pritchard JK, Wena X, Falushb D. 2010. Documentation for structure software: Version 2.3. United States of America (US): University of Oxford.
Purba MP. 2012. Genetic structure and variation of five Shorea leprosula natural populations based on microsatellite marker [tesis]. Bogor (ID): Institut Pertanian Bogor.
Rajora OP, Rahman MH. 2001. Microsatellite DNA Marker and Their Usefulness in Poplar, and Conservation of Microsatellite DNA loci in Salicaceae. Di dalam: Muller SD, Schubert R, Editor. Genetic Response of Forest Systems to Changing Environmental Conditions. Netherlands (NL): Kluwer Academic Publishers.
Rambey R. 2011. Pengetahuan lokal sistem agroforestri mindi (Melia azedarach L.) (Studi Kasus di Desa Selaawi, Kecamatan Talegong, Kabupaten Garut, Propinsi Jawa Barat) [tesis]. Bogor (ID): Institut Pertanian Bogor.
Rohlf FJ. 2008. Numerical Taxonomy and Analysis System (NTSYSpc) Version 2.0. New York (US): University of New York.
Roshetko JM, Mulawarman, Iriantono D. 2004. Kebun benih untuk petani dan LSM-mengapa dan bagaimana?. Di dalam: Iriantono D, Farobi I, Harum F, Ariyanto D, Editor. Benih untuk Rakyat. Suplemen Gedeha. 14:2-6.
SAS Institute Inc. 2006. Base SAS® 9.1.3 Procedures Guide Edisi ke-2. North Carolina (US): SAS Institute Inc.
Setyaningsih L. 2007. Pemanfaatan cendawan mikoriza arbuskula dan kompos aktif untuk meningkatkan pertumbuhan semai mindi (Melia azedarach Linn.) pada media tailing tambang emas pongkor [tesis]. Bogor (ID): Institut Pertanian Bogor.
Santoso H. 2011. Kebijakan sumber benih dan potensi kebutuhan benih untuk mendukung penanaman satu milyar pohon. Di dalam: Rimbawanto A, Leksono B, Widyatmoko AYPBC, editor. Peran Sumber Benih Unggul dalam Mendukung Keberhasilan Penanaman Satu Milyar Pohon. Seminar Nasional Pembangunan Sumber Benih; 2011 Jun 30; Yogyakarta, Indonesia. Yogyakarta (ID): Balai Besar Penelitian Bioteknologi dan Pemuliaan Tanaman Hutan.
Sudomo A, Santosa HB. 2011. Pengaruh media organik dan tanah mineral terhadap pertumbuhan dan indeks mutu bibit mindi (Melia azedarach L.). J Penelitian Hutan dan Konservasi Alam. 8(3):263-271.
Sutopo L. 2012. Teknologi Benih. Jakarta (ID): Rajawali Pers.
Syamsuwida D. 2012. Aspek biologi dan genetika pada sistem reproduksi mindi (Melia azedarach Linn.) [disertasi]. Bogor (ID): Institut Pertanian Bogor. Syamsuwida D, Palupi ER, Siregar IZ, Indrawan A. 2012. Flower initiation,
morphology, and developmental stages of flowering-fruiting of mindi (Melia azedarach L). JMHT. 18(1):10–17.doi: 10.7226/jtfm.18.1.10. Thode VA, Backes A, Mader G, Kriedt R, Bonnato SL, Freitas LB. 2013.
27 Varshney RJ, Chabane K, Hendre PS, Aggarwal RK, Graner A. 2007. Comparative assessment of EST-SSR, EST-SNP and AFLP markers for evaluation of genetic diversity and conservation of genetic resources using wild, cultivated and elite barleys. J Plant Science. 173:638–649.
Wang X, Rinehart TA, Wadl PA, Spiers JM, Hadziabdic D, Windham MT, Trigiano RN. 2009. A new electrophoresis technique to separate microsatellite alleles. African J of Biotechnology. 8(11):2432-2436
Wang HW, Duan JM, Zhang P, Cheng YG, Wu JW, Wang GZ. 2013. Microsatellite markers in Paulownia kawakamii (Scrophulariaceae) and cross-amplification in other paulownia species. Genet. Mol. Res. 12 (3): 3750-3754.doi: http://dx.doi.org/10.4238/2013.
Wee AKS, Takayamaka K, Kajita T, Webb EL. 2013. Microsatellite loci for Avicennia alba (acanthaceae), Sonneratia alba (lythraceae) and Rhizophora mucronata (rhizophoraceae). J of Tropical Forest Science. 25(1):131-136.
Weis AE, Winterer J, Vacher T, Kossler TM, Young CA, Lebuhn GL. 2005. Phenological assortative mating in flowering plants: the nature and consequences of its frequency dependence. Evolutionary Ecology Research. 7:161-181.
Weising K, Nybon H, Wolff K, Kahl G. 2005. DNA Fingerprinting in Plants: Principles, Methods, and Aplications. Amerika Serikat (US): CRC Press Taylor & Francis Group.
Yeh FC, Yang R. 1999. POPGENE Version 1.31: User Guide Centre for International Forestry Research. Alberta (US): University of Alberta. Yulianti. 2011. Strategi pengembangan sumber benih mindi (Melia azedarach L.)
pada hutan rakyat di Provinsi Jawa Barat [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Yulianti, Siregar IZ, Wijayanto N, Darma IGKT, Syamsuwida D. 2011. Genetic variation of Melia azedarach in community forests of West Java assessed by RAPD. J Biodiversitas. 12(2):64-69.
28
LAMPIRAN
Lampiran 1 Dokumentasi amplifikasi silang primer mimba ke mindi
Ai_11
300 bp
200 bp
100 bp
Ai_5
29 Lampiran 2 Skoring alel pada tiap primer
32
Lampiran 3 Dokumentasi pengamatan pertumbuhan bibit mindi di persemaian
Kecambah mindi
Dokumentasi penyapihan
33 RIWAYAT HIDUP
Penulis lahir di Desa Sumurcinde, Kecamatan Soko, Kabupaten Tuban pada tanggal 23 Agustus 1988. Terlahir sebagai anak pertama dari dua bersaudara di keluarga Jayus dan Rupiin. Pada tahun 2007, penulis lulus dari SMA Negeri 2 Tuban, dimana pada tahun yang sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI) pada Departemen Silvikultur. Studi ini diselesaikan pada tahun 2011 dengan skripsi berjudul “Keragaman Genetik Gaharu Budidaya dan Alami Berdasarkan Penanda Mikrosatellit” dibawah bimbingan Prof Dr Ir Iskandar Z. Siregar, MForSc. Pada tahun 2012 penulis terdaftar sebagai mahasiswa Pascasarjana IPB program studi Silvikultur Tropika, dan mendapatkan beasiswa unggulan (BU) Dikti.
Selama menuntut ilmu di IPB, penulis aktif di beberapa organisasi kemahasiswaan yakni pada periode tahun 2007/2008 sebagai pengurus FORCES IPB (Forum for Scientific Studies), aktif di Himpro TGC (Tree Grower Community) divisi Scientific Improvement 2008-2010, dan HMI (Himpunan Mahasiswa Islam) Komisariat Fahutan periode 2009/2010 sebagai bendahara umum, serta HMI Cabang Bogor periode 2012/2013 sebagai wakil bendahara umum. Pada tahun 2012 penulis dipercaya sebagai bendahara Forum Wacana Silvikultur Tropika.
Beberapa kegiatan penulis selama kuliah, diantaranya penulis pernah menjadi asisten program TPB IPB (Tingkat Persiapan Bersama) pada mata kuliah Fisika tahun 2009/2010, untuk program Sarjana pada mata kuliah: Silvikultur (2010-2013), Genetika Hutan (2010-2014), Pemuliaan Pohon (2010-2014) dan Teknik Perbanyakan Tanaman Hutan (2011/2012). Selain itu, penulis juga pernah menjadi asisten PPH (Praktek Pengelolaan Hutan) di Hutan Pendidikan Gunung Walat pada bulan Juli 2011. Untuk program Magister, penulis menjadi asisten praktikum pada mata kuliah Pemanfaatan Sumberdaya Genetik Hutan Tanaman.
Penulis pernah terlibat dalam beberapa penelitian, diantaranya: Analisis Struktur dan Kerapatan Stomata pada Fast Growing dan Slow Growing Species (2010), pada tahun 2012 hasil penelitian tersebut disusun oleh peneliti menjadi buku yang diterbitkan IPB Press dengan judul “Stomata FGS dan SGS”; Master Plant Ruang Terbuka Hijau Kota dan Hutan Kota di Kabupaten Kepulauan Meranti Provinsi Riau (2011); Analisis Carbon pada Tegakan Sengon dan Akasia di Lahan Reklamasi PT. Arutmin Indonesia Tambang Batulicin (2011); Pengembangan Sumber Benih Mindi (Melia azedarach Linn.) untuk Hutan Rakyat di Jawa Barat (2012); Uji klon Jati, Kerjasama antara Koperasi Perumahan Wanabakti Nusantara (KPWN) dan Fakultas Kehutanan IPB (2010-2013), Respon Pertumbuhan Bibit Kayu Afrika (Maesopsis eminii) pada Berbagai Dosis dan Metode Pemberian Pupuk Cair Organik (2014); dan Practical Application of DNA Barcoding in Conservation of Endangered Tree Species (2014).