PEMBENTUKAN GALUR-GALUR DIHAPLOID PADI
SAWAH TADAH HUJAN TOLERAN KEKERINGAN
MELALUI KULTUR ANTERA
CUCU GUNARSIH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Pembentukan Galur-galur Dihaploid Padi Sawah Tadah Hujan Toleran Kekeringan Melalui Kultur Antera adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2015
RINGKASAN
CUCU GUNARSIH. Pembentukan Galur-galur Dihaploid Padi Sawah Tadah Hujan Toleran Kekeringan Melalui Kultur Antera. Dibimbing oleh BAMBANG SAPTA PURWOKO, ISWARI SARASWATI DEWI dan MUHAMAD SYUKUR.
Kekeringan merupakan kendala utama yang menyebabkan rendahnya produksi dan stabilitas produksi padi di ekosistem tadah hujan. Pembentukan padi sawah tadah hujan yang toleran kekeringan dan memiliki hasil tinggi merupakan salah satu solusi untuk mengatasi kendala dalam peningkatan produktivitas padi, dengan cara menyilangkan tetua donor toleran kekeringan dengan tetua padi sawah yang memiliki potensi hasil tinggi. Pembentukan varietas padi sawah tadah hujan toleran kekeringan secara konvensional membutuhkan waktu yang relatif lama. Kultur antera dapat mempercepat diperolehnya galur-galur murni pada generasi pertama. Dengan demikian pemanfaatan kultur antera dapat meningkatkan efisiensi proses seleksi, menghemat biaya, waktu dan tenaga kerja.
kalus dan regenerasi tanaman paling baik. Kedua asal persilangan tersebut juga memberikan jumlah tanaman tertinggi yang berhasil diaklimatisasi sebanyak 108 tanaman dan 102 tanaman. Sebanyak 53 tanaman dihaploid (33.97% dari total tanaman hijau) telah berhasil diidentifikasi.
Penelitian kedua bertujuan untuk mendapatkan informasi tentang parameter genetik terhadap karakter agronomi dan hasil galur-galur dihaploid. Percobaan dilakukan, di rumah kaca BB Biogen pada bulan September 2014 sampai Januari 2015. Percobaan menggunakan rancangan acak lengkap dengan dua ulangan. Percobaan menguji 60 galur dihaploid hasil kultur antera. Hasil penelitian menunjukkan terdapat keragaman karakter antar galur dihaploid hasil kultur antera. Karakter-karakter yang diamati memiliki variabilitas genetik luas, kecuali karakter hasil gabah kering per rumpun. Karakter yang memiliki nilai heritabilitas tinggi yaitu, tinggi tanaman, panjang daun bendera, panjang malai, umur 50% berbunga, umur panen, jumlah gabah hampa per malai, dan bobot 1000 butir. Karakter yang memiliki nilai heritabilitas sedang yaitu, jumlah anakan produktif, jumlah gabah isi per malai, persentase gabah isi per malai, dan hasil gabah kering per rumpun. Berdasarkan nilai heritabilitas, nilai koefisien korelasi, koefisien sidik lintas maka karakter yang dapat digunakan untuk menyusun indeks seleksi bagi hasil gabah kering per rumpun pada galur-galur dihaploid hasil kultur antera adalah karakter persentase gabah isi per malai, jumlah anakan produktif, jumlah gabah isi per malai, bobot 1000 butir, dan tinggi tanaman. Berdasarkan seleksi indeks, setiap peubah yang terpilih diberikan nilai pembobotan. Diperoleh 19 galur dihaploid yang memiliki nilai indeks tertinggi.
Penelitian ketiga bertujuan untuk mendapatkan galur dihaploid yang toleran terhadap cekaman kekeringan. Penelitian dilaksanakan di rumah kaca Muara, BB Padi pada bulan Oktober sampai Desember 2014. Bahan yang digunakan yaitu 60 galur dihaploid (DH) hasil kultur antera, Salumpikit (cek toleran kekeringan), dan IR 20 (cek peka kekeringan). Penelitian menggunakan rancangan acak kelompok dengan 3 ulangan. Berdasarkan seleksi indeks terhadap uji penapisan kekeringan pada fase bibit, terpilih 16 galur dihaploid yang memiliki nilai indeks seleksi tertinggi. Keenambelas galur dihaploid tersebut memiliki toleransi terhadap cekaman kekeringan dan memiliki daya tumbuh kembali yang baik.
Dari ketiga penelitian yang telah dilakukan, terpilih 7 galur dihaploid yang memiliki nilai indeks tertinggi pada masing-masing seleksi indeks. Ketujuh galur dihaploid toleran kekeringan dan memiliki karakter agronomi baik serta potensi hasil yang tinggi. Dengan demikian, ketujuh galur dihaploid hasil kultur antera berpotensi untuk diuji daya hasilnya di sawah tadah hujan.
SUMMARY
CUCU GUNARSIH. Development of Doubled Haploid Rainfed Rice Lines Tolerant to Drought Through Anther Culture. Supervisor by BAMBANG SAPTA PURWOKO, ISWARI SARASWATI DEWI and MUHAMAD SYUKUR.
Drought is a major constraint for productivity and stability of rice in rainfed ecosystems. The rice breeding for drought tolerant and high yielding ability is one solution to solve barriers in increasing rice productivity. This can be done by crossing the donor parent of drought tolerant rice with rice parents that have high yielding ability. Conventional breeding for drought tolerant require a long time relatively. Anther culture can accelerate pure line obtainment as pure line doubled haploid (DH) can be immediately obtained in the first generation. Thereby the use of anther culture can improve the efficiency of the selection process, save costs, time and labor.
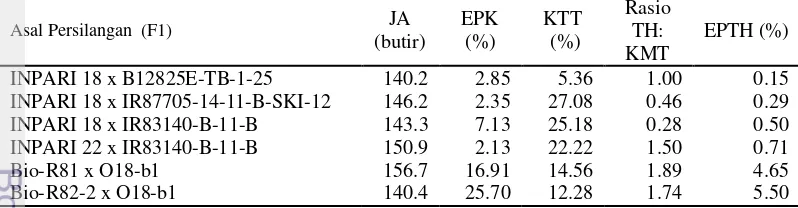
The first research consisted of two experiments i.e. screening of candidate rice parents tolerant to drought at seedling stage, and the development of doubled haploid rice lines through anther culture. The first experiment was aimed at obtaining prospective parents of rice that can tolerate drought stress at seedling stage. It was conducted in Muara greenhouse, Indonesian Center for Rice Research (ICRR) from March to May 2013. The experiment was arranged in randomized complete block design with three replications. Based on weighted index, there were five genotypes have the highest index value namely IR83140-B-11-B, IR87705-14-11-B-SKI-12, INPAGO 8, B12825E-TB-1-25 dan IR87706-215-B-B-B.Three genotypes (IR83140-B-11-B, B12825E-TB-1-25, and IR87705-14-11-B-SKI-12) were selected as candidates for rice parent which had tolerant to drought in seedling stage. The second experiment was aimed at obtaining doubled haploid lines which could be selected further. Crosses were done in Muara green house, ICRR from March to September 2013. The materials used were consisted of six cross combinations. F1 seeds were used as material for anther culture research activities. The experiment was carried out at BB Biogen green house, Bogor from October 2013 until Juny 2014. The experiment was arranged in completely randomized design with 14 replications. The treatments consisted of six F1 genotypes i.e.: INPARI 18 x IR83140-B-11-B, INPARI 18 x B12825E-TB-1-25, INPARI 18 x IR87705-14-11-B-SKI-12, INPARI 22 x IR83140-B-11-B, Bio-R81 x O18b-1, Bio-R82-2 x O18b-1. Callus induction medium was based on N6 medium + 2.0 NAA mg L-1 + 0.5 kinetin mg L-1 + 10-3 M Putrescine, while regeneration medium was based on MS + 0.5 NAA mg L-1 + 2.0 kinetin mg L-1 + 10-3 M putrescine. The result of this study indicated that six F1s derived crosses gave varied responses in anther culture abilities. R82-2 x O18-b1 and Bio-R81 x O18-b1 F1s derived cross had good response of callus induction and plant regeneration. Both F1s derived crosses also gave the highest number of regenerated plants that were successfully acclimatized (108 plants and 102 plants respectively). A total of 53 doubled haploid plants (33.97% total of green plants) were successfully identified.
An experiments were carried out in BB Biogen green house, Bogor from September 2014 to January 2015. The experiment was arranged in completely randomized design with two replications. The experiment evaluated 60 doubled haploid lines obtained from anther culture. The results showed that variation of agronomic characters and yield existed among the doubled haploid lines. Characters observed have broad genetic variability, except for grain yield per hill. Characters that have high heritability values were plant height, leaf blade length, panicle length, days to flowering, days to harvesting, number of unfilled grains per panicle, and 1000 grain weight. Characters that have moderate heritability values were number of productive tillers, number of filled grains per panicle, percentage of filled grains per panicle, grain yield per hill. Characters plant height, number of productive tillers, number of unfilled grains per panicle, percentage of filled grains per panicle, and 1000 grain weight had significant correlation and direct effect on grain yield per hill. Based on heritability, the correlation coefficient, coefficient of path analysis, the characters that can be used to construct index selection for grain yield per hill of the doubled haploid lines were plant height, number of productive tillers, number of filled grains per panicle, percentage of filled grains per panicle, and 1000 grain weight. Based on the weighted index, 19 doubled haploid lines were selected to have the highest index value and potential for further evaluation in rainfed areas.
The objectives of the third research were to obtain the doubled haploid lines which have tolerance to drought. The research was conducted in the Muara greenhouse, ICRR from October to December 2014. The materials used in the research were 60 doubled haploid lines (DH), Salumpikit (drought tolerant check), and IR 20 (drought sensitive check).The experiment was arranged in randomized complete block design with three replications. The experiment evaluated the 60 doubled haploid lines. Based on selection index of variable of screening drought tolerance and recovery, that were select 16 doubled haploid lines which have the highest selection index value above tolerant check Salumpikit. Sixteen of dihaploid lines that were tolerant to drought and had good recovery, which potential to evaluated in rainfed areas.
Of the three research that have been done obtained 7 doubled haploid lines that had the highest index value for each index selection. The seven doubled haploid lines have tolerance to drought and good agronomic characters and high yield. Thus, the seven doubled haploid lines have the potential to be tested in rainfed lowland.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
PEMBENTUKAN GALUR-GALUR DIHAPLOID PADI
SAWAH TADAH HUJAN TOLERAN KEKERINGAN
MELALUI KULTUR ANTERA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Alhamdulillaahirrabbil’aalamiin. Segala puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga tesis ini berhasil diselesaikan. Tema yang dipilih dalam penelitian ini ialah pengembangan galur padi dalam menghadapi fenomena perubahan iklim yaitu cekaman kekeringan. Penelitian dengan judul Pembentukan Galur-Galur Dihaploid Padi Sawah Tadah Hujan Toleran Kekeringan Melalui Kultur Antera.
Pada kesempatan ini penulis menyampaikan penghargaan dan ucapan terima kasih kepada :
1. Prof Dr Ir Bambang S Purwoko, Dr Ir Iswari S Dewi dan Bapak Prof Dr Muhamad Syukur SP MSi selaku pembimbing yang senantiasa memberikan masukan, arahan, bimbingan, dorongan dan motivasi dalam penulisan tesis ini.
2. Dr Ir Trikoesoemaningtyas, MSc dan Dr Ir Yudiwanti Wahyu EK, MS selaku penguji pada ujian thesis atas arahan, kritik, saran, dan masukan untuk perbaikan thesis.
3. Badan Litbang Pertanian atas dukungan dana beasiswa dan bantuan dana penelitiannya, serta kepada Direktorat Jenderal Pendidikan Tinggi atas pendanaan penelitian skema Hibah Penelitian Kompetensi (Prof Dr Ir Bambang S Purwoko sebagai ketua peneliti).
4. Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian, terutama kepada Dr Iswari S Dewi yang telah memfasilitasi penelitian dan bahan penelitian, serta kepada Pak Iman Ridwan, Teh Yeni, dan Deni yang telah membantu penulis selama bekerja di laboratorium dan rumah kaca.
5. Kepala Balai Besar Penelitian Tanaman Padi dan Ketua Kelompok Peneliti Pemuliaan atas pemberian izin menggunakan materi penelitian, fasilitas rumah kaca, bantuan, dukungan, dan masukannya.
6. Rekan-rekan peneliti BB Padi : Heni S, Indrastuti R, Trias S, Yudistira N, Yuni W, Estria FP, Indria, Pak Sunaryo dan tim padi gogo untuk doa, bantuan, dan dukungannya.
7. Rekan-rekan mahasiswa pascasarjana AGH PBT 2012, Forsca dan Forum Petugas Belajar Badan Litbang Pertanian juga disampaikan apresiasi atas kebersamaan dan motivasinya selama studi.
8. Almarhum Wedy Utoro (Suami) dan Avicenna (Ananda) yang senantiasa menjadi inspirasi, dan motivasi penulis untuk menyelesaikan studi.
9. Almarhum M. Somantri (ayahanda) , Ibu Hj Mariam (Ibu), Ibu Hj Waliyah (Ibu Mertua), juga kakak-kakak tersayang (Teh Neni, Teh Arum, Teh Eros dan A Oman) yang senantiasa mendoakan, menguatkan dan mendorong penulis dalam penyelesaian penulisan tesis ini.
10. Teman terbaikku: Zahratul Millah, Arina Saniati, Pak Usman, dan M. Ridha Alfarabi yang selalu siap membantu penulis.
Besar harapan penulis, semoga karya ini bermanfaat dalam pengembangan padi sawah tadah hujan di Indonesia.
DAFTAR ISI
DAFTAR ISI v
DAFTAR TABEL vi
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN vii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
Ruang Lingkup Penelitian 3
2 TINJAUAN PUSTAKA 5
Tanaman Padi dan Ekosistemnya 5
Kekeringan 8
Kultur Antera dalam Pemuliaan Tanaman Padi 11
3 PEMBENTUKAN GALUR DIHAPLOID PADI SAWAH TOLERAN
TERHADAP CEKAMAN KEKERINGAN 14
Pendahuluan 15
Bahan Dan Metode 17
Hasil dan Pembahasan 22
Simpulan 31
4 EVALUASI DAN SELEKSI AGRONOMI GALUR DIHAPLOID
GENERASI PERTAMA (DH0) 32
Pendahuluan 33
Bahan dan Metode 34
Hasil Dan Pembahasan 37
Simpulan 55
5 UJI TOLERANSI GALUR-GALUR DIHAPLOID (DH) HASIL KULTUR
ANTERA TERHADAP CEKAMAN KEKERINGAN 56
Pendahuluan 56
Bahan dan Metode 57
Hasil dan Pembahasan 57
Simpulan 63
6 PEMBAHASAN UMUM 64
7 SIMPULAN DAN SARAN 68
Simpulan 68
Saran 68
LAMPIRAN 80
RIWAYAT HIDUP 84
DAFTAR TABEL
1 Klasifikasi respon tanaman padi terhadap kekeringan 17 2 Klasifikasi respon daya tumbuh kembali (recovery) tanaman padi
setelah perlakuan kekeringan 18
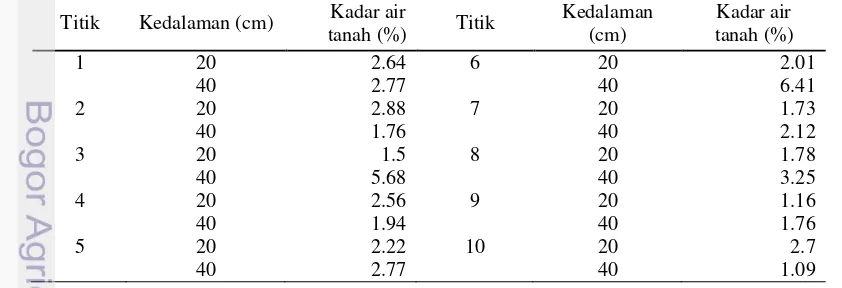
3 Respon genotipe padi terhadap cekaman kekeringan selama 42 HST 23 4 Kadar air tanah saat uji toleransi kekeringan selama 42 HST 23 5 Respon daya tumbuh kembali genotipe padi setelah pengeringan 42
HST 24
6 Indeks seleksi genotipe padi sawah di rumah kaca Muara 25 7 Hasil induksi kalus dari enam asal persilangan (F1) padi sawah hasil
kultur antera 27
8 Regenerasi tanaman dari enam asal persilangan (F1) padi sawah hasil
kultur antera 28
9 Efisiensi pembentukan kalus dan tanaman hijau pada kultur antera padi 29 10 Hasil aklimatisasi dan tanaman dihaploid yang dihasilkan pada kultur
antera padi 31
11 Analisis ragam rancangan acak lengkap 35
12 Skor, indeks penyakit (DI), dan kriteria serangan virus kerdil hampa 39 13 Analisis ragam dari galur-galur dihaploid padi di rumah kaca BB
Biogen 40
14 Tinggi tanaman, jumlah anakan produktif, umur 50% berbunga, umur panen, panjang daun bendera dan panjang malai galur-galur dihaploid
padi generasi pertama (DH0) 42
15 Jumlah gabah isi per malai, jumlah gabah hampa per malai, persentase gabah isi per malai, bobot 1000 butir dan hasil gabah kering per rumpun dari galur-galur dihaploid padi generasi pertama (DH0) 45 16 Warna gabah, warna beras dan eksersi malai dari galur-galur dihaploid
padi generasi pertama (DH0) di rumah kaca BB Biogen 48 17 Nilai komponen ragam dan nilai duga heritabilitas karakter agronomi
dari galur-galur dihaploid tanaman padi 49
18 Nilai koefisien korelasi masing-masing peubah terhadap karakter hasil gabah kering per rumpun galur-galur dihaploid padi sawah 52 19 Pengaruh langsung dan pengaruh tidak langsung beberapa karakter
terhadap bobot per rumpun pada galur-galur dihaploid padi sawah 53 20 Indeks terboboti pada 19 galur dihaploid padi sawah yang terpilih 54 21 Respon galur-galur dihaploid padi terhadap kekeringan setelah 50 HST
di rumah kaca Muara, BB Padi 59
22 Kadar air tanah saat skoring kekeringan (50 HST) 60 23 Respon daya tumbuh kembali (recovery) galur-galur dihaploid padi
terhadap kekeringan di rumah kaca Muara, BB Padi 61 24 Indeks seleksi galur-galur dihaploid padi berdasarkan respon
25 Indeks seleksi untuk ketiga calon tetua padi yang terpilih 64 26 Tujuh galur dihaploid padi yang terpilih berdasarkan seleksi indeks 67
DAFTAR GAMBAR
1 Diagram alir penelitian 4
2 Keragaan genotipe padi sawah saat penapisan di rumah kaca Muara 22 3 Keragaan kemampuan tumbuh kembali genotipe padi sawah setelah
diuji toleransi kekeringannya 24
4 Kultur antera padi 26
5 Proses aklimatisasi dan pertumbuhan tanaman hijau hasil kultur antera
padi di rumah kaca 30
6 Keragaan pertanaman galur dihaploid tanaman padi 38 7 Kondisi pertanaman galur dihaploid padi yang normal dan tidak normal
pada saat fase reproduktif 38
8 Keragaan galur dihaploid padi (CG12-30-1-1) 46
9 Karakter bentuk dan warna gabah padi dan beras 47 10 Eksersi malai dari galur-galur dihaploid hasil kultur antera padi 47 11 Diagram lintasan beberapa karakter dengan hasil gabah kering padi per
rumpun 54
12 Keragaan galur-galur dihaploid padi pada saat skrining kekeringan di
rumah kaca Muara BB Padi 58
13 Penampilan tanaman padi setelah skrining kekeringan 50 HST 58
DAFTAR LAMPIRAN
1 Luas panen, Produktivitas, dan Produksi dari tahun 2011-2014 81 2 Galur-galur dihaploid hasil kultur antera yang dievaluasi di rumah kaca
1
1
PENDAHULUAN
Latar Belakang
Beras merupakan makanan pokok utama bagi masyarakat Indonesia sehingga beras merupakan komoditas pangan yang sangat strategis. Seiring dengan laju pertambahan penduduk Indonesia sebesar 1.49% per tahun dengan konsumsi beras per kapita sebesar 139.15 kg-1 tahun, maka diperkirakan kebutuhan beras nasional pada tahun 2014 sekitar 33 juta ton (BPS 2010). Ada tiga tantangan utama dalam penyediaan beras di Indonesia, diantaranya adalah (i) meningkatnya permintaan beras sesuai dengan peningkatan jumlah penduduk, (ii) terbatasnya ketersediaan beras dunia dan (iii) kecenderungan meningkatnya harga pangan (Deptan 2010). Menurut data angka sementara (ASEM) (BPS 2015) produksi padi tahun 2014 sebanyak 70.83 juta ton gabah kering giling atau mengalami penurunan sebesar 0.45 juta ton (0.63%) dibandingkan tahun 2013. Penurunan produksi padi terjadi di Pulau Jawa sebesar 0.83 juta ton, sedangkan produksi padi di luar Pulau Jawa mengalami kenaikan sebanyak 0.39 juta ton. Penurunan produksi diperkirakan terjadi karena penurunan luas panen seluas 41.61 ribu ha (0.30%) dan penurunan produktivitas sebesar 0.017 ton ha-1 (0.33%) (Lampiran 1). Untuk itu peningkatan produksi padi perlu diupayakan melalui peningkatan produktivitas tanaman, dan peningkatan luas areal panen sehingga dapat mendukung ketahanan pangan nasional.
Luas lahan sawah tadah hujan sekitar 2.02 juta ha atau 24 % dari total luas lahan di Indonesia dengan kisaran hasil 1.8–3.1 ton ha-1 (Pane et al. 2009). Dilihat dari potensi lahan, lahan sawah tadah hujan merupakan lumbung padi kedua nasional setelah lahan sawah irigasi. Potensi lahan sawah tadah hujan yang cukup luas tersebar di Provinsi Banten, Jawa Barat, Jawa Tengah, Lampung, Sulawesi Selatan, dan Nusa Tenggara Barat (Badan Litbang 2008). Peningkatan produksi padi melalui penggunaan varietas unggul yang toleran kekeringan mempunyai peluang yang besar untuk menunjang peningkatan padi secara nasional. Dengan peningkatan hasil mencapai 1 ton ha-1, maka 50% dari total luasan 2.02 juta ha mampu memberikan kontribusi padi nasional 1.0 juta ton.
2
dengan tetua yang memiliki potensi hasil tinggi (Atlin et al. 2006; Bernier et al. 2008; Babu 2010).
Kekeringan adalah fenomena yang sering terjadi dan merupakan kendala utama yang penting untuk produksi padi tadah hujan di Asia. Hal ini menjadi ancaman terbesar bagi ketahanan pangan, kesehatan manusia, dan kehidupan jutaan orang miskin. Paling sedikit 23 juta ha lahan sawah (20% dari luas total lahan sawah) di Asia mengalami kekeringan dengan intensitas yang berbeda-beda. Cekaman kekeringan menjadi salah satu faktor utama yang berkontribusi atas rendahnya dan tidak stabilnya produksi padi di wilayah Asia (Pandey dan Bhandari 2008).
Pembentukan varietas unggul padi sawah tadah hujan yang toleran kekeringan secara konvensional membutuhkan waktu yang relatif lama. Pada tanaman padi, penggaluran membutuhkan waktu 6 sampai 8 generasi dari mulai proses menyilangkan dilanjutkan dengan seleksi, sehingga jangka waktu sampai mendapatkan galur murni dengan sifat-sifat yang unggul membutuhkan waktu lebih dari 5 tahun (Dewi et al. 1996). Teknik kultur antera dapat mempercepat perolehan galur-galur murni karena galur murni dihaploid (doubled haploid/DH) dapat segera diperoleh pada generasi pertama. Dengan demikian pemanfaatan kultur antera dapat meningkatkan efisiensi proses seleksi, serta menghemat biaya, waktu dan tenaga kerja (Dewi dan Purwoko 2012). Teknik ini telah banyak dimanfaatkan oleh para peneliti padi dengan berbagai macam tujuan, antara lain untuk mendapatkan galur-galur toleran naungan (Sasmita et al. 2002), toleran aluminium (Dewi et al. 2009a), perakitan tetua padi hibrida (Dewi et al. 2005; Lestari dan Nugraha 2006), dan padi gogo dengan sifat-sifat tipe baru (Herawati et al. 2008; Safitri et al. 2010).
Perumusan Masalah
Perubahan iklim global menyebabkan kelangkaan air yang sering dihadapi di lahan padi sawah tadah hujan. Kekeringan merupakan kendala utama yang menyebabkan rendahnya produksi dan stabilitas produksi padi di ekosistem tadah hujan. Perakitan padi sawah tadah hujan yang toleran kekeringan serta memiliki hasil tinggi merupakan salah satu solusi untuk mengatasi kendala dalam peningkatan produktivitas padi. Salah satu caranya dengan memanfaatkan tetua donor yang toleran kekeringan yang disilangkan dengan tetua yang memiliki potensi hasil tinggi. Pembentukan varietas unggul padi sawah tadah hujan yang toleran kekeringan secara konvensional membutuhkan waktu yang relatif lama. Teknik kultur antera dapat mempercepat perolehan galur-galur murni karena galur murni dihaploid dapat segera diperoleh pada generasi pertama. Dengan demikian pemanfaatan kultur antera dapat meningkatkan efisiensi proses seleksi, serta menghemat biaya, waktu dan tenaga kerja.
3 dilakukan uji toleransi galur dihaploid hasil kultur antera terhadap cekaman kekeringan pada fase bibit.
Tujuan Penelitian
Tujuan umum penelitian ini adalah untuk mendapatkan galur-galur padi sawah tadah hujan dengan sifat agronomi baik dan toleran kekeringan. Tujuan khusus penelitian ini adalah untuk :
1. Mendapatkan calon tetua persilangan yang memiliki toleransi terhadap cekaman kekeringan pada fase bibit.
2. Mendapatkan individu galur-galur dihaploid yang dapat diseleksi lebih lanjut.
3. Mendapatkan informasi tentang parameter genetik terhadap karakter agronomi yang menunjang pembentukan padi sawah tadah hujan yang toleran kekeringan.
4. Mendapatkan galur dihaploid dengan sifat agronomi baik yang toleran terhadap cekaman kekeringan.
Manfaat Penelitian
Hasil penelitian yang telah dilaksanakan ini diharapkan:
1. Sebagai bahan pertimbangan bagi pemulia tanaman dalam perakitan genotipe padi sawah tadah hujan yang memiliki toleransi terhadap cekaman kekeringan.
2. Sebagai bahan rujukan dan pengkajian lebih lanjut bagi perkembangan ilmu pengetahuan dalam pengembangan pertanian.
3. Bermanfaat bagi masyarakat dalam mengembangkan varietas padi sawah tadah hujan yang memiliki kemampuan toleransi terhadap cekaman kekeringan
Ruang Lingkup Penelitian
4
5
2
TINJAUAN PUSTAKA
Tanaman Padi dan Ekosistemnya
Tanaman padi (Oryza sativa L.) dalam sistematika tumbuhan diklasifikasikan ke dalam divisio Spermatophyta, sub divisio Angiospermae, kelas Monocotyledoneae, Ordo Poales, famili Graminae, dan genus Oryza Linn (Watanabe 1997, Watanabe 1993). Hal ini berdasarkan metode klasifikasi tanaman padi yang digunakan Kato (1928, 1930) dalam Watanabe (1993) berdasarkan pada berbagai macam kriteria, yaitu karakteristik/bentuk tanaman padi, pembentukan biji di bawah kondisi penyerbukan sendiri, fertilitas tanaman F1, pemasakan butir polen pada tanaman F1, fertilitas tanaman F2, serta adanya perbedaan serologis.
Tanaman padi terbagi menjadi dua sub spesies, yaitu tipe japonica dan tipe indica. Namun demikian, beberapa ilmuwan mengungkapkan adanya tipe javanica yang mengindikasikan tipe gabungan antara tipe japonica dan tipe indica (Katayama 1993). Subspesies japonica mempunyai karakteristik: daun berukuran sempit-lebar dan berwarna hijau muda, ukuran gabah pendek sampai panjang, umumnya gabah tidak mempunyai ekor (awn), bulu sekam jarang dan pendek, gabah mudah rontok, jumlah anakan banyak, postur anakan tegak, jumlah malai banyak, bobot malainya berat, batang sedang sampai tinggi, jaringan lunak, kepekaan yang bervariasi terhadap panjang hari, dan kandungan amilosa pada biji 23-31%. Subspesies indica memiliki karakteristik: daun berukuran sempit berwarna hijau tua, bentuk gabah pendek dan agak bulat panjang, pada umumnya gabah tidak mempunyai ekor (awn), bulu sekam lebat dan panjang, gabah agak mudah rontok, jumlah anakan sedang, postur anakan menyebar, jumlah malai banyak, bobot malainya ringan, batang pendek sampai sedang, jaringan keras, tidak peka atau agak peka terhadap panjang hari, dan kandungan amilosa pada biji 10-24%. Subspesies javanica mempunyai karakteristik: daun berukuran lebar, kaku dan berwarna hijau muda, gabah panjang lebar dan tebal, gabah mempunyai ekor (awn) panjang atau tidak ada, bulu sekam panjang, gabah sulit rontok, jumlah anakan sedikit, postur anakan tegak, jumlah malai sedikit, bobot malainya berat, batang tinggi, jaringan keras, agak peka terhadap panjang hari, dan kandungan amilosa pada biji 20-25% (Matsuo 1952 dalam Katayama 1993; Gupta dan O’toole 1986). Distribusi wilayah penyebaran padi subspecies japonica yaitu Jepang, Korea, China Utara, Afrika dan Cina bagian tengah; subspecies javanica di Jawa, Sumatra, Philipina, Eropa dan Amerika; subspecies indica di India, Indochina, Cina bagian Selatan dan bagian barat, Taiwan (Katayama 1993).
6
yang mudah sampai sedang, peka terhadap kerebahan, memiliki adaptasi yang tinggi terhadap lahan suboptimal, hasil gabah tinggi, sensitif terhadap panjang hari, persilangan alaminya jarang; padi bulu memiliki karakter ketebalan batang yang tebal, jumlah batang yang sedikit, daun bendera yang lebar dan kaku, memiliki rambut pada daun bendera yang jarang dan kasar, gabah berekor, bobot gabah yang ringan, bentuk gabah yang lebar dan tebal, kerontokan gabah yang sedang sampai sukar, tahan rebah, memiliki adaptasi yang rendah terhadap lahan suboptimal, hasil gabah rendah, tidak sensitif terhadap panjang hari, persilangan alaminya moderat sampai luas (Katayama 1993).
Bunga padi merupakan bunga terminal yang berbentuk malai yang terdiri atas bunga-bunga tunggal (spikelet). Tiap spikelet terdiri atas 2 lemma steril, sekam besar (lemma), palea (sekam kecil), 6 buah benang sari yang masing-masing memiliki 2 kotak sari dan sebuah putik. Kepala putik berjumlah dua buah dengan bulu-bulu halus. Lodikula terdapat di dasar bunga yang memiliki peran yang sangat penting pada saat bunga mekar. Pada saat tanaman padi akan berbunga, lodikula mengembang dan mendorong lemma dan palea sehingga keduanya terpisah dan akhirnya membuka. Mekarnya bunga diikuti dengan pecahnya kotak sari, selanjutnya lemma dan palea menutup kembali sehingga memungkinkan tepung sari menempel pada kepala putik di bunga yang sama (Fehr dan Hadley 1980).
Pertumbuhan tanaman padi dikelompokkan ke dalam tiga fase, yaitu fase vegetatif, fase reproduktif, dan fase pematangan. Fase vegetatif dimulai dari awal pertumbuhan sampai pembentukan malai, fase reproduktif dari pembentukan malai sampai pembungaan dan fase pematangan dari pembungaan sampai gabah matang. Di daerah tropis, fase generatif berlangsung 35 hari dan fase pematangan 30 hari. Perbedaan masa pertumbuhan dibedakan berdasar lamanya fase vegetatif (Taslim dan Fagi 1989). IRRI (2004) mengelompokkan pertumbuhan tanaman padi dalam sembilan fase, yaitu: (0) perkecambahan (germination), (1) fase bibit (seedling), (2) fase pembentukan anakan (tillering), (3) fase pemanjangan batang (stem elongation), (4) fase inisiasi bunga sampai bunting (panicle initiation to booting), (5) fase heading, (6) fase berbunga (flowering), (7) fase masak susu (milk grain), (8) fase masak tepung (dough grain), (9) fase biji masak (mature grain). Fase 0-3 termasuk fase vegetatif, fase 4-6 termasuk fase generatif, dan fase 7-9 termasuk fase pematangan (ripening).
7 Badan Litbang Pertanian (2008) menegaskan bahwa revolusi hijau lestari akan lebih mengarah kepada pengembangan lahan sub optimal yang meliputi lahan kering, sawah tadah hujan, sawah pasang surut dan lahan rawa lebak. Beratnya tantangan yang dihadapi untuk pemenuhan kebutuhan pangan di masa depan disebabkan laju tingkat produktivitas lahan sawah irigasi yang cenderung menurun dan populasi penduduk semakin bertambah. Faktor-faktor penyebab rendahnya produktivitas padi sawah tadah hujan di tingkat petani yaitu, belum diterapkannya teknik budidaya yang baik, varietas yang ditanam sebagian besar masih varietas lokal atau varietas unggul lama, kurang intensifnya pengendalian gulma, gangguan hama dan penyakit, dan tingkat kesuburan lahan yang relatif rendah. Selain itu, cekaman kekeringan yang hampir terjadi setiap tahun disebabkan jumlah curah hujan yang rendah dan musim hujan yang pendek (Serraj et al. 2009).
Luas lahan padi sawah tadah hujan di Indonesia sekitar 2.08 juta ha, dengan sistem produksi yang diterapkan sebagian besar petani Jawa Tengah (Rembang dan Pati) berupa teknologi budidaya padi gogo rancah (Fagi 1995). Masalah yang menjadi kendala dalam produksi tanaman padi di lahan tadah hujan lebih komplek dibandingkan di sawah irigasi. Selain itu, wilayah ekosistem lahan tadah hujan umumnya dihuni oleh petani miskin dengan infrastruktur terbatas. Teknologi yang diterapkan masih didominasi oleh teknologi tradisional. Begitupun dengan adanya fenomena El-nino yang menjadi ancaman dalam sistem produksi padi dan ketahanan pangan nasional (Balitpa 2002).
Beberapa kendala lainnya terkait budidaya tanaman padi di sawah tadah hujan yaitu, lahan sawah tadah hujan dengan iklim kering memiliki periode hujan yang singkat, jumlah dan intensitas curah hujan yang rendah, distribusi curah hujan yang tidak merata sepanjang musim, dan kondisi biofisik–kimia tanah yang beragam (Quisenberry 1982; Serraj et al. 2009).
Salah satu upaya untuk menanggulangi permasalahan tersebut adalah mengembangkan genotipe padi sawah tadah hujan yang selain memiliki potensi hasil tinggi juga memiliki toleransi terhadap cekaman kekeringan. Skrining dengan karakter fertilitas tinggi, pada kondisi optimum dan kondisi cekaman moderat pada fase reproduktif dibutuhkan untuk membentuk kultivar yang mengkombinasikan potensi hasil tinggi dengan toleran kekeringan (Atlin et al. 2006). Hasil penelitian Atlin et al. (2006) mengungkapkan bahwa kultivar indica yang terseleksi memiliki fertilitas tinggi telah diuji di sawah tadah hujan dataran tinggi. Beberapa kultivar seperti IR55423-01 menunjukkan hasil tertinggi (23-69 % lebih tinggi dibandingkan kultivar japonica tropis dan varietas-varietas untuk padi sawah irigasi) walaupun ditanam di bawah beberapa kondisi cekaman (cekaman moderat di lahan dataran tinggi, cekaman air yang parah dan lokasi dataran tinggi yang kurang subur). Hal ini mengindikasikan bahwa hasil tinggi dan toleran cekaman air yang moderat mampu dikombinasikan. Ini merupakan suatu pendekatan baru untuk meningkatkan produksi dan mengurangi kerugian pada sistem padi sawah tadah hujan di Asia.
8
malai), dan eksersi malai yang baik (skor 7-9) (Nanda dan Chofman 1979). Galur-galur dihaploid yang dihasilkan dari penelitian ini diharapkan memiliki karakteristik yang mendukung karakteristik agronomi padi sawah tadah hujan yang toleran kekeringan.
Perakitan varietas unggul di Indonesia yang sesuai untuk ditanam di sawah tadah hujan telah dilakukan BB Padi dengan melepas varietas Dodokan tahun 1987 dan Silugonggo tahun 2001, yang memiliki umur panen sekitar 96-100 hari. Namun kedua varietas tersebut masih memiliki kelemahan karena produktivitasnya masih rendah, juga memiliki tekstur nasi yang pera. Untuk itu, BB Padi telah mengevaluasi dan melepas kembali varietas unggul lainnya yang memiliki umur sangat genjah dan potensial untuk dikembangkan di daerah yang rawan kekeringan. BB Padi secara resmi telah melepas 3 VUB yang dianjurkan untuk ditanam di lahan sawah tadah hujan yang memiliki potensi hasil tinggi, memiliki ketahanan terhadap hama dan penyakit, serta bermutu sesuai preferensi petani pada tahun 2010, yaitu INPARI 11, INPARI 12 dan INPARI 13 (Suprihatno et al. 2010). Tahun berikutnya BB Padi juga telah melepas VUB INPARI 18 dan INPARI 19 dengan umur sangat genjah, potensi hasil tinggi dan tahan hama dan penyakit yang direkomendasikan cocok untuk ditanam di lahan irigasi dan tadah hujan (BB Padi 2013).
Penelitian ini memanfaatkan tetua donor padi sawah tadah hujan yang toleran kekeringan disilangkan dengan tetua padi sawah yang memiliki potensi hasil tinggi. Tetua-tetua yang toleran kekeringan tersebut yaitu galur IR83140-B-11-B, B12825E-TB-1-25, IR87705-14-11-B-SKI-12, dan IR87706-215-B-B-B berasal dari BB Padi. Galur-galur tersebut termasuk galur generasi lanjut yang sedang dievaluasi potensi hasil dan daya adaptasinya di beberapa sentra padi sawah tadah hujan. Selain itu, dimanfaatkan juga galur O18b-1 yang memiliki keunggulan diantaranya toleran terhadap kekeringan, agak toleran cekaman aluminium dan toleran terhadap naungan, walaupun potensi hasilnya tergolong rendah (Mara 2014).
Kekeringan
Definisi kekeringan berbeda-beda sesuai dengan lokasi geografi, jumlah dan durasi curah hujan. Definisi kekeringan menurut perspektif pemulia tanaman adalah suatu keadaan kurangnya air tersedia yang menyebabkan kehilangan hasil atau suatu periode dimana tidak ada hujan atau irigasi yang mengakibatkan tidak cukupnya kadar air tanah sehingga menurunkan pertumbuhan tanaman dan hasil (Fukai dan Cooper 1995; Blum 2011). Kekeringan menurut perspektif meteorologi adalah kurangnya curah hujan dari rata-rata curah hujan normal pada suatu periode waktu tertentu. Kekeringan menurut perspektif hidrologi adalah keadaan ketika jumlah air yang tersedia terbatas dibandingkan jumlah air yang dibutuhkan. Perspektif pertanian, kekeringan didefinisikan merupakan jumlah air yang tidak mencukupi tanaman untuk tumbuh optimal (Wilhite dan Glantz 1985; Jodo 1995); atau merupakan kadar air tanah yang tidak mencukupi untuk memenuhi kebutuhan air tanaman (evapotranspirasi) sehingga mengakibatkan kehilangan produksi.
9 tersedianya sumber irigasi dalam semusim dan antar musim, dan fase pertumbuhan tanaman. Hal ini menyulitkan untuk membuat kondisi cekaman kekeringan yang seragam di lapangan untuk mendapatkan metode skrining kekeringan yang efektif (White dan Singh 1991). Lanceras et al. (2004) melaporkan bahwa pengujian di rumah kaca dengan mengontrol kekeringan menggunakan pot dapat membuat cekaman kekeringan yang seragam, namun fasilitas tersebut mahal dan tidak sesuai untuk skrining dalam skala luas, serta studi cekaman kekeringan pada pot jauh berbeda dengan cekaman kekeringan alami di lapangan. Afa (2013) melaporkan bahwa percobaan di pot dengan perlakuan cekaman kekeringan pada kadar air 60% kapasitas lapangan mengakibatkan penurunan hasil yang drastis rata-rata mencapai 78.4 persen. Hal ini disebabkan, umumnya media di pot menyebabkan hambatan untuk penetrasi dan perkembangan akar karena terbatasnya ukuran pot.
Padi setiap saat (fase vegetatif atau fase reproduktif) dapat mengalami cekaman kekeringan. Kekurangan air pada saat fase vegetatif mengakibatkan terhambatnya pertumbuhan. Kekeringan yang terjadi pada saat fase berbunga dan pengisian biji merupakan fase yang kritis bagi tanaman padi (Fukai dan Lilley 1994; Sikuku et al. 2010). Beberapa akibat kekeringan pada fase tersebut diantaranya adalah tanaman tidak berkembang (Lubis et al. 1993), menurunnya jumlah gabah isi (Sikuku et al. 2010), dan menurunnya hasil gabah (Cruz dan O’Toole 1984).
Cekaman kekeringan pada fase reproduktif telah diidentifikasi sebagai fase yang paling menyebabkan penurunan hasil gabah (O’Toole et al. 1982). Pengurangan hasil gabah dapat mencapai 34-53% di bawah cekaman kekeringan moderat dan mencapai 65-88% di bawah cekaman kekeringan berat jika dibandingkan dengan kontrol padi sawah irigasi (Verulkar et al. 2010).
Terbatasnya ketersediaan air pada saat pembungaan, menyebabkan pengurangan hasil gabah terutama dengan meningkatnya kehampaan gabah (Jongdee et al. 2002; Yamin et al. 2012; Afa 2013). Tanaman padi dengan umur panen yang lebih pendek memiliki peluang yang lebih besar dibandingkan padi yang berumur panjang dalam hal tingkat toleransi terhadap kekeringan dan produksi hasil gabah (O’Toole dan De Datta 1986).
Kekeringan merupakan kendala paling penting yang mempengaruhi hasil padi sawah tadah hujan. Kekeringan dapat mengurangi produktivitas baik melalui dampak langsung terhadap produksi biomassa dan jumlah butiran gabah dan juga melalui terlambatnya pengelolaan tanaman seperti jadwal tanam, pemupukan dan penyiangan (Atlin et al. 2008).
10
Pemahaman genetik terhadap karakter toleran kekeringan merupakan prasyarat keberhasilan program pemuliaan. Hal ini akan membantu memprediksikan penampilan tanaman di lapangan dan juga membantu memodifikasi level toleransi untuk meningkatkan stabilitas hasil di bawah kondisi cekaman kekeringan. Toleransi kekeringan merupakan karakter yang kompleks yang dikendalikan secara poligenik dengan heritabilitas rendah sampai medium (Ekanayake et al. 1985). Analisis genetik dari karakter morfologi, hasil dan komponen hasil, serta beberapa karakter akar di bawah kondisi cekaman kekeringan menunjukkan pewarisan poligenik dengan aditif yang signifikan dan efek genetik dominan, serta interaksinya, dan juga menunjukkan interaksi yang signifikan dengan lingkungan (Asfaq et al. 2012; Panwar 2005).
Beberapa upaya khusus dilakukan untuk pengembangan varietas padi yang toleran kekeringan untuk mengantisipasi kendala kekeringan. Diantaranya adalah seleksi pada karakter sekunder pada kondisi cekaman kekeringan seperti arsitektur akar, potensial air daun, potensial air malai, penyesuaian osmotik, dan kadar air relatif (Fukai et al. 1999; Jongdee et al. 2002; Pantuwan et al. 2002) namun tidak memberikan hasil yang diharapkan untuk meningkatkan produksi di bawah cekaman kekeringan (Kumar 2009).
Selain itu, penelitian terbaru di IRRI telah menunjukkan bahwa heritabilitas karakter hasil gabah di bawah cekaman kekeringan adalah moderat sampai tinggi (Bernier et al. 2007; Venuprasad et al. 2007; Kumar 2009), sehingga membuka peluang untuk seleksi langsung terhadap hasil gabah dibandingkan dengan seleksi yang dilakukan pada karakter sekunder.
Seleksi langsung untuk hasil gabah di bawah cekaman kekeringan telah dilaporkan cukup efektif (Kumar et al. 2008; Venuprasad et al. 2008) dan telah dibuktikan dengan menggabungkan potensi hasil tinggi dengan hasil yang baik di bawah cekaman kekeringan (Kumar et al. 2008; Venuprasad et al. 2008). Hasil penelitian IRRI menunjukkan bahwa seleksi secara langsung untuk karakter hasil di bawah cekaman kekeringan, dikombinasikan dengan seleksi untuk potensi hasil di bawah kondisi optimum, merupakan cara yang efektif dalam pembentukan varietas (Atlin et al. 2008).
11 toleransi padi hibrida terhadap cekaman kekeringan adalah dengan pemberian larutan PEG 6000 konsentrasi 25%. Genotipe-genotipe yang terseleksi dari hasil pengujian berdasarkan metode seleksi dini pada fase perkecambahan, fase bibit dan metode pot menunjukkan konsistensinya sebagai genotipe-genotipe yang toleran cekaman kekeringan di lapangan (Afa 2013).
Kombinasi penggunaan varietas unggul dan teknologi pengelolaan tanaman untuk meningkatkan produktifitas dan produksi padi di sawah tadah hujan sangat penting. Ludlow (1989) membagi mekanisme adaptasi tanaman terhadap cekaman kekeringan menjadi : (1) drought escape, di mana tanaman melengkapi siklus hidupnya tepat waktu bila tidak terjadi cekaman kekeringan (2) drought avoidance, di mana tanaman memaksimalkan serapan air dan meminimalkan kehilangan air saat terjadi cekaman kekeringan; (3) drought tolerance, di mana tanaman terus tumbuh dan berfungsi pada saat cekaman kekeringan. Selain itu, Fukai dan Cooper (1995) menambahkan mekanisme adaptasi lain yaitu drought recovery, yang menunjukkan daya pulih tanaman dari cekaman kekeringan berhubungan dengan kemampuannya untuk mempertahankan daun hijau selama periode tersebut.
Mekanisme penghindaran dari kekeringan (avoidance) dengan mengurangi kehilangan air banyak berhubungan dengan respon secara morfologi, seperti perubahan struktural stomata, mengurangi luas daun, meningkatkan ketebalan daun, pertumbuhan akar yang lebih dalam, dan perubahan ketebalan lilin daun (Galle dan Feller 2007; Agbicodo et al. 2009). Selain itu, meningkatnya panjang perakaran tanaman dan rasio akar tajuk (Farooq et al. 2008); penggulungan daun juga merupakan karakter sekunder yang dimiliki tanaman yang toleran terhadap kekeringan (Banziger et al. 2000).
Kultur Antera dalam Pemuliaan Tanaman Padi
Pemuliaan pada tanaman menyerbuk sendiri seperti padi ditujukan untuk memperoleh galur-galur murni yang memiliki sifat-sifat unggul. Galur-galur murni tersebut umumnya diperoleh dengan cara persilangan yang diikuti dengan sejumlah proses seleksi pada setiap generasi. Pada tanaman padi, proses tersebut membutuhkan waktu selama 6 sampai 8 generasi (Fehr 1987; Oono 1997; Dewi dan Purwoko 2012). Dengan memanfaatkan teknik kultur antera, waktu yang dibutuhkan untuk perakitan tanaman padi menjadi lebih singkat, karena tanaman dihaploid adalah galur murni yang dapat diperoleh dari generasi pertama. Dalam hal ini, keseragaman genetik adalah homozigositas pada individu dihaploid hasil kultur antera akan tercapai dalam waktu singkat setelah hibridisasi, karena tanaman yang diperoleh berasal dari mikrospora (haploid) yang terdapat di dalam antera F1 yang dikulturkan. Oleh karena itu, strategi pemuliaan dihaploid memiliki keunggulan kompetitif dibandingkan dengan metode konvensional (Kao 1996).
12
untuk mendapatkan galur murni tanaman transgenik dengan cepat pada padi, jagung dan gandum (Brisibe et al. 2000; Otani et al. 2005). Penggandaan kromosom sel haploid pada tahap tertentu selama tahap regenerasi tanaman dalam teknik kultur antera pada tanaman serealia merupakan langkah penting dalam memproduksi tanaman dihaploid (Obert dan Barnabás 2004). Hal ini mendorong Indonesia untuk memanfaatkan teknik kultur antera dalam program perbaikan tanaman padi. Induksi haploid melalui kultur antera yang mengandung mikrospora merupakan metode yang paling sederhana dan mudah dilaksanakan dibandingkan dengan kultur mikrospora langsung (Datta 2005).
Aplikasi bioteknologi melalui kultur antera dapat mempersingkat siklus pemuliaan tanaman karena kegiatan seleksi langsung dilakukan pada tanaman yang homozigos. Selain itu, perolehan galur-galur murni yang diinginkan terseleksi berdasarkan keunggulan dari karakter-karakter agronomi pada populasi dihaploid yang homozigos. Dengan demikian, salah satu keuntungan penggunaan kultur antera antara lain jumlah populasi tanaman yang diseleksi menjadi lebih sedikit. Namun ada ukuran minimum untuk melakukan seleksi pada populasi dihaploid, hal ini tergantung pada variasi jumlah gen yang mengendalikan karakter yang ditargetkan. Dewi dan Purwoko (2001) menyatakan bahwa sejumlah n gen merupakan perbedaan pada tetua persilangan dengan diasumsikan tidak mengalami pautan, maka ukuran minimum sejumlah 2n tanaman disyaratkan untuk ditanam dengan tujuan agar semua genotipe homozigos dapat terwakili. Hal ini berbeda dengan pendekatan pemuliaan konvensional yang membutuhkan sejumlah 4n tanaman. Somantri et al. (2003) menyatakan bahwa semakin banyak jumlah gen yang mengendalikan suatu karakter maka semakin besar jumlah populasi yang harus ditanam.
Beberapa faktor yang mempengaruhi keberhasilan kultur antera, yaitu genotipe tanaman, komposisi media, praperlakuan antera sebelum dikulturkan, fase pembentukan mikrospora pada saat antera dikulturkan, kondisi lingkungan tanaman yang akan diambil anteranya dan waktu pada saat pengambilan malai (Chu 1978; Gupta dan Borthakur 1987; Lee et al. 2004).
Tanaman padi yang diregenerasikan melalui kultur antera menghasilkan tanaman hijau serta tanaman albino (mutan kloroplas) (Sato 1997; Dewi et al. 2004). Albinisme merupakan fenomena sangat penting dalam kultur in vitro pada famili Gramineae. Hal ini merupakan salah satu kendala yang dihadapi para peneliti kultur antera dan kultur mikrospora. Tanaman albino mengalami defisiensi kandungan klorofil, yang disebabkan plastid tidak dapat berkembang menjadi kloroplas dan tidak adanya sintesis klorofil (Zhou 1996; Zapata et al. 1983). Faktor-faktor penyebab terjadinya albino antara lain yaitu faktor genotipe, fase perkembangan mikrospora, suhu kultur dan pra perlakuan sebelum antera dikulturkan. Masalah tersebut dapat diatasi dengan dengan penggunaan poliamin (diantaranya yaitu pemberian 10-3 M Putresin) dapat meningkatkan induksi kalus dan meningkatkan regenerasi tanaman hijau (Dewi et al. 2003; Dewi et al. 2004; Dewi dan Purwoko 2008; Dewi dan Purwoko 2012).
13 membedakan tanaman dihaploid dan diploid dengan cara ditanam secara bersamaan, selanjutnya dibandingkan penampilan fenotipenya terutama karakter kualitatif.
Penampilan fenotipik yang dapat membedakan antara tanaman padi haploid dengan dihaploid pada generasi awal (DH0), yaitu melalui perbedaan dalam bentuk tanaman (stature), warna daun, bentuk daun, perkembangan anakan dan akar, pembentukan biji, serta jumlah dan ukuran stomata (Dewi et al. 2011). Selain itu tanaman padi haploid ditandai dengan tidak adanya ligula dan aurikula, jumlah anakan yang hampir dua kali lebih banyak, ukuran tinggi tanaman, panjang malai, dan panjang daun sekitar 60-70% dari tanaman diploid, serta tidak dapat menghasilkan biji (Gosal et al. 1997).
Beberapa keuntungan yang diperoleh dari pemanfaatan teknik kultur antera selain memperpendek siklus pemuliaan dengan diperolehnya homozigositas secara cepat, serta meningkatkan efisiensi seleksi, memperluas variabilitas genetik melalui produksi variasi gametoklonal, mempercepat terekspresinya gen resesif, menyediakan sumber benih yang homozigos, dan menghemat waktu, biaya dan tenaga (Fehr 1987; Zapata 1990; Dewi et al. 1996; Masyhudi et al. 1997; Kim dan Baenziger 2005).
14
3
PEMBENTUKAN GALUR DIHAPLOID PADI SAWAH
TOLERAN TERHADAP CEKAMAN KEKERINGAN
Abstrak
Pembentukan tanaman dihaploid padi sawah tadah hujan yang memiliki toleransi terhadap kekeringan dapat dilakukan dengan menggunakan kultur antera. Penelitian ini terdiri atas 2 kegiatan yaitu, penapisan calon tetua padi sawah yang toleran terhadap cekaman kekeringan pada fase bibit, dan pembentukan galur dihaploid padi sawah melalui kultur antera. Percobaan pertama bertujuan untuk mendapatkan calon tetua persilangan yang memiliki toleransi terhadap cekaman kekeringan pada fase bibit. Percobaan kedua bertujuan untuk mendapatkan individu galur-galur dihaploid yang dapat diseleksi lebih lanjut. Percobaan pertama dilakukan di rumah kaca (RK) Muara, Balai Besar Penelitian Padi, Bogor pada bulan Maret sampai Mei 2013. Percobaan dilakukan dengan menggunakan rancangan acak kelompok dengan 3 ulangan. Percobaan kedua terdiri atas dua kegiatan, yaitu persilangan dan kultur antera F1. Persilangan dilakukan di RK Muara BB Padi pada bulan Maret sampai September 2013. Bahan yang digunakan pada kegiatan ini yaitu 6 kombinasi persilangan. Hasil persilangan berupa benih-benih F1 tersebut digunakan sebagai bahan kegiatan kultur antera. Penelitian kultur antera dilaksanakan menggunakan rancangan acak lengkap dengan 14 ulangan. Perlakuan terdiri atas enam genotipe F1 yaitu: INPARI 18 x IR83140-B-11-B, INPARI 18 x B12825E-TB-1-25, INPARI 18 x IR87705-14-11-B-SKI-12, INPARI 22 x IR83140-B-11-B, Bio-R81 x O18b-1, Bio-R82-2 x O18b-1. Media induksi kalus adalah media N6 + 2.0 NAA mg L-1 + 0.5 kinetin mg L-1 + 10-3 M Putresin, sedangkan media regenerasi kalus adalah media MS + 0.5 NAA mg L-1 + 2.0 kinetin mg L-1 + 10-3 M putresin. Hasil seleksi indeks berdasarkan respon toleransi terhadap kekeringan pada fase bibit selama 42 HST dan respon daya tumbuh kembali diperoleh lima genotipe terbaik yaitu IR83140-B-11-B, IR87705-14-11-B-SKI-12, INPAGO 8, B12825E-TB-1-25 dan IR87706-215-B-B-B. Terpilih tiga genotipe (IR83140-B-11-B, B12825E-TB-1-25, dan IR87705-14-11-B-SKI-12) untuk dijadikan calon tetua padi sawah yang memiliki toleransi terhadap cekaman kekeringan pada fase bibit. Hasil penelitian kultur antera menunjukkan bahwa keenam galur F1 yang diuji pada penelitian ini memberikan respon yang bervariasi terhadap kultur antera padi. Populasi asal persilangan (F1) Bio-R82-2 x O18-b1 dan Bio-R8 x O18-b1 menghasilkan respon induksi kalus dan regenerasi tanaman paling baik. Kedua populasi asal persilangan tersebut juga memberikan jumlah tanaman tertinggi yang berhasil diaklimatisasi berturut-turut sebanyak 108 tanaman dan 102 tanaman. Sebanyak 53 tanaman dihaploid (33.97% dari total tanaman hijau) telah berhasil diidentifikasi.
15 Abstract
Breeding of rainfed rice tolerant to drought can be accomplished using anther culture. This study consisted of two experiments, namely screening of candidate parent which tolerance to drought at seedling stage, and the development of doubled haploid rice lines through anther culture. The first experiment was aimed at obtaining candidate parents of rice that tolerate drought at seedling stage. The objectives of the second experiment was to obtain doubled haploid lines which could be selected further. The first experiment was arranged in completely randomized design with 14 replications. This research was conducted in Muara Greenhouse, ICRR in March-May 2013. The first experiment was arranged in randomized block design with three replications. The second experiment consisted of two activities, namely crosses and anther culture of F1. Crosses were done in Muara greenhouse, ICRR in March-September 2013. The materials used in this activity were 6 cross combinations which then were used as material for anther culture. The anther culture studies were arranged in completely randomized design with 14 replications. The treatments consisted of six F1 genotypes i.e.: INPARI 18 x IR83140-B-11-B, INPARI 18 x B12825E-TB-1-25, INPARI 18 x IR87705-14-11-B-SKI-12, INPARI 22 x IR83140-B-11-B, Bio-R81 x O18b-1, Bio-R82-2 x O18b-1. Callus induction medium was based on N6 medium + 2.0 NAA mg L-1 + 0.5 kinetin mg L-1 + 10-3 M Putrescine, while regeneration medium was based on MS + 0.5 NAA mg L-1 + 2.0 kinetin mg L-1 + 10-3 M Putrescine. The results of selection indices based on the response to the drought tolerance of seedlings phase for 42 DAS and recovery responses obtained five best genotypes with the highest weight namely IR83140-B-11-B, IR87705-14-11-B-SKI-12, INPAGO 8, B12825E-TB-1-25 dan IR87706-215-B-B-B. Three genotypes (IR83140-B-11-B, B12825E-TB-1-25, and IR87705-14-11-B-SKI-12) were selected as candidates for rainfed rice parent which had tolerance to drought in seedling stage. The result of anther culture study indicated that six populations gave varied responses in anther culture abilities. R82-2 x O18-b1 and Bio-R81 x O18-b1 F1s derived cross had good response of callus induction and plant regeneration. In this research, both F1s derived cross also gave the highest number of regenerated plants that were successfully acclimatized (108 plants and 102 plants respectively). A total of 53 doubled haploid plants (33.97% total of green plants) were successfully identified.
Keywords: Callus induction, doubled haploid, rainfed, regeneration, tolerance to drought
Pendahuluan
16
Metode pemuliaan tanaman berbasis teknologi, mempertimbangkan level pengetahuan yang mendasari mekanisme dan mengontrol proses perbanyakan dan seleksi terhadap tipe tanaman superior. Terdapat 3 periode utama dalam pemuliaan tanaman yang telah teridentifikasi, yaitu (1) pemuliaan tanaman berdasarkan pengamatan terhadap variasi dengan menyeleksi tanaman berdasarkan variasi yang tampak alami atau di dalam varietas tradisional; (2) pemuliaan tanaman berdasarkan persilangan/perkawinan yang dikontrol dengan cara menyeleksi tanaman yang menampilkan adanya rekombinasi dari gen-gen yang diinginkan dari tetua-tetua yang berbeda, (3) pemuliaan tanaman berdasarkan monitoring rekombinasi dengan menyeleksi gen spesifik atau profil marka, menggunakan peralatan molekuler untuk melacak variasi dalam genom (Breseghello dan Coelho 2013). Keragaman genetik pada penelitian ini diperoleh melalui persilangan yang telah didesain untuk memperoleh galur padi sawah tadah hujan yang toleran kekeringan serta memiliki hasil tinggi. Dengan memanfaatkan tetua donor toleran kekeringan yang disilangkan dengan tetua yang memiliki potensi hasil tinggi (Atlin et al. 2006; Bernier et al. 2008; Babu 2010).
Pembentukan varietas unggul padi sawah tadah hujan yang toleran kekeringan secara konvensional memerlukan waktu yang relatif lama. Penggaluran tanaman padi memerlukan waktu 6 sampai 8 generasi dari mulai proses menyilangkan dilanjutkan dengan seleksi, sehingga jangka waktu sampai mendapatkan galur murni dengan sifat-sifat yang unggul membutuhkan waktu lebih dari 5 tahun. Teknik kultur antera dapat mempercepat perolehan galur-galur murni karena galur murni dihaploid (DH) dapat segera diperoleh pada generasi pertama. Dengan demikian, pemanfaatan kultur antera dapat meningkatkan efisiensi proses seleksi, serta menghemat biaya, waktu dan tenaga kerja (Dewi dan Purwoko 2012).
Umumnya tanaman haploid pada kultur antera padi diperoleh melalui dua tahap, yaitu tahap induksi butir tepung sari menjadi kalus (embrioid), dan tahap diferensiasi menjadi tanaman kecil (plantlet) (Dewi dan Purwoko 2001). Teknik kultur antera telah banyak digunakan dalam kegiatan pemuliaan, namun keberhasilannya masih dibatasi oleh beberapa faktor, yaitu genotipe tanaman, komposisi media, praperlakuan antera sebelum dikulturkan, fase pembentukan mikrospora pada saat antera dikulturkan, kondisi lingkungan tanaman yang akan diambil anteranya dan waktu pada saat pengambilan malai (Lee et al. 2004; Datta 2005). Di samping itu, interaksi antara faktor-faktor tersebut juga mempengaruhi respon dalam kultur antera (Özgen et al. 2005). Kaushal et al. (2014) melaporkan bahwa genotipe, suhu saat praperlakuan, durasi praperlakuan dan interaksinya menunjukkan pengaruh yang nyata terhadap induksi kalus dan regenerasi tanaman hijau. Bagheri dan Jelodar (2008) menunjukkan bahwa faktor genetik sangat mempengaruhi proses kultur antera, baik dalam induksi kalus maupun regenerasi tanaman hijau karena keduanya diwariskan secara kuantitatif.
17 (ratun) dan pemberian perlakuan kolkisin 0.1–0.3% (Dewi et al. 2007; Fu et al. 2008).
Program pemuliaan padi sawah tadah hujan yang toleran kekeringan melalui teknik kultur antera berpeluang mempersingkat perolehan galur-galur murni yang homozigot. Penelitian ini terdiri atas 2 percobaan, yaitu penapisan calon tetua padi sawah yang toleran terhadap cekaman kekeringan pada fase bibit, dan pembentukan galur dihaploid padi sawah melalui kultur antera. Percobaan pertama bertujuan untuk mendapatkan calon tetua persilangan yang memiliki toleransi terhadap cekaman kekeringan pada fase bibit, sedangkan percobaan kedua bertujuan untuk mendapatkan individu galur-galur dihaploid yang dapat diseleksi lebih lanjut.
Bahan Dan Metode
A. Penapisan Calon Tetua Padi Sawah yang Toleran terhadap Cekaman Kekeringan pada Fase Bibit
Percobaan dilaksanakan pada bulan Maret sampai Mei 2013 di rumah kaca Muara, Balai Besar Penelitian Padi, Bogor. Bahan yang digunakan untuk penapisan calon tetua terdiri atas 16 genotipe yaitu genotipe B12497E-MR-45, B12825E-TB-1-25, IR87705-14-11-B-SKI-12, SMD9-15D-MR-4, IR83140-B-11-B, BP3672-2E-KN-17-3-3*IR83140-B-11-B, IR87706-215-B-B-IR83140-B-11-B, BP10620F-BB4-13-BB8, IR 87707-118-B-B-B, BP13990-4B-GRT-3-1-2-2-4, INPARI 13, INPARI 18, INPARI 22, INPAGO 8, Salumpikit (cek toleran kekeringan), dan IR 20 (cek peka kekeringan). Sepuluh benih per genotipe uji ditanam dengan jarak 20 cm x 20 cm dalam bak skrining berukuran (panjang x lebar x tinggi) 5.5 m x 1.3 m x 0.6 m diisi tanah. Penanaman benih dalam bak penapisan ditata sesuai dengan tata letak rancangan acak kelompok dengan 3 ulangan.
Pemeliharaan tanaman dilakukan secara intensif, penyiraman air hanya dilakukan sampai tanaman berumur 14 Hari Setelah Tanam (HST). Setelah fase tersebut tanaman dalam bak penapisan dibiarkan tumbuh tanpa disiram air lagi. Penilaian toleransi terhadap kekeringan dilakukan pada saat varietas pembanding Peka (IR20) mati atau semua daunnya telah mengering (skor 9) sesuai Standard Evaluation System (SES) dari IRRI (2002) pada Tabel 1.
Tabel 1 Klasifikasi respon tanaman padi terhadap kekeringan
Nilai Kriteria Keterangan
0 Sangat Toleran Tidak ada gejala
1 Toleran Ujung daun mengering
3 Agak Toleran ¼ ujung daun kering
5 Moderat ¼ sampai ½ ujung daun ada yang kering
7 Agak Peka ½ sampai 2/3 ujung daun ada yang kering
9 Peka Semua daun kering
Sumber: IRRI (2002)
18
dilakukan penilaian daya tumbuh kembali (recovery) pada setiap genotipe yang diuji dengan menggunakan skala pengamatan sesuai Standard Evaluation System (SES) dari IRRI (2002) pada Tabel 2.
Tabel 2 Klasifikasi respon daya tumbuh kembali (recovery) tanaman padi setelah perlakuan kekeringan
Nilai Kriteria Persentase tanaman tumbuh kembali (%)
1 Toleran 90-100
3 Agak Toleran 70-89
5 Moderat 40-69
7 Agak Peka 20 – 39
9 Peka 0 -19
Sumber: IRRI (2002)
Indeks Seleksi Terboboti
Indeks seleksi terboboti digunakan untuk menentukkan genotipe yang direkomendasikan sebagai calon tetua padi sawah yang toleran kekeringan pada fase bibit. Peubah yang digunakan pada indeks seleksi terboboti adalah peubah respon tanaman padi terhadap kekeringan dan respon daya tumbuh kembali (recovery) tanaman padi setelah perlakuan kekeringan. Menurut Falconer dan Mackay (1996), penentuan indeks seleksi terboboti berdasarkan:
Indeks seleksi : I = b1X1 + b2X2 + b3X3 + ... + bnXn.
Dimana; I adalah indeks seleksi; bn adalah bobot dari peubah ke-n; Xn adalah nilai fenotipe yang telah distandardisasi untuk peubah ke-n
Keterangan: x = Nilai tengah peubah tiap genotipe = Nilai tengah peubah
= Ragam peubah
B. Pembentukan Galur Dihaploid Padi Sawah melalui Kultur Antera
Persilangan
Pembentukan materi genetik terdiri atas dua kegiatan yaitu penanaman tetua dan persilangan di antara tetua untuk mendapatkan benih F1. Kedua kegiatan tersebut dilakukan di kebun percobaan Muara, Balai Besar Penelitian Padi, Bogor pada bulan Maret sampai September 2013.
19 toleran cekaman aluminium dan toleran terhadap naungan, walaupun potensi hasilnya tergolong rendah (Mara 2014).
Pembentukan populasi F1 diawali dengan penanaman tetua persilangan. Penyemaian benih tetua persilangan menggunakan bak yang berukuran 30 cm x 50 cm, kemudian bibit dipindahtanamkan ke lapangan setelah berumur 21 hari. Bibit tetua persilangan ditanam di lapangan dengan jarak tanam 25 cm x 25 cm, satu bibit per lubang tanam. Benih tetua persilangan ditanam dengan interval waktu 7-10 hari yang diulang tiga kali, hal ini bertujuan untuk sinkronisasi pembungaan pada saat persilangan. Aplikasi pemupukan pada pertanaman menggunakan 300 kg.ha-1 Phonska dan 95 kg.ha-1 urea. Pada saat tanaman memasuki fase berbunga, tanaman yang akan dijadikan tetua betina, dipindahkan ke dalam pot dan dibawa ke rumah kaca untuk dikastrasi (dibuang bunga jantannya). Stadia bunga yang baik untuk diemaskulasi adalah pada saat benang sari berada pada pertengahan bunga. Stadia ini menunjukkan bahwa bunga akan mekar dalam 1 - 2 hari. Bunga digunting dengan kemiringan 600, kemudian benang sari dikeluarkan dengan cara dihisap menggunakan pompa penghisap. Malai yang telah diemaskulasi ditutup dengan kertas minyak dan diberi label.
Persilangan dilakukan pada hari berikutnya pada pukul 10.00–12.00. Bunga jantan diambil dari pertanaman di lapangan dengan cara memotong malai yang sebagian bunganya sudah mekar. Pengambilan malai dilakukan pada pukul 08.00–09.00. Malai dari lapangan ini kemudian diletakkan dalam gelas berisi air dalam ruang persilangan. Ruang persilangan merupakan ruang tertutup yang diberi lampu tembak (4 x 100 watt) dengan jarak ± 2.5 m dari malai sehingga suhunya lebih tinggi dibandingkan suhu lapangan (350–400C). Hal ini bertujuan untuk mempercepat mekarnya bunga. Penyerbukan dilakukan dengan cara menggoyang-goyangkan malai tetua jantan di atas malai tetua betina yang sudah dikastrasi. Bunga yang sudah diserbuki ditutup dengan kertas minyak dan diberi label. Hasil persilangan (benih F1) dapat dipanen 3 minggu setelah persilangan dilakukan. Setelah dijemur 2 hari dan dibuang sekamnya, benih dikeringkan dengan cara dioven dalam kantong kertas pada suhu ± 450C selama 3 hari. Benih F1 ini dapat ditanam 15 hari setelah panen.
Kultur Antera F1
Penanaman tanaman F1 donor eksplan dilakukan di rumah kaca. Kegiatan kultur antera dilakukan di Laboratorium Kultur Jaringan, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian, pada bulan Oktober 2013 sampai dengan Juni 2014.
20
benar hasil persilangan (F1), tetapi jika penampilan fenotipiknya mirip 100% tetua betina, maka genotipe tersebut berasal dari hasil selfing.
Media kultur antera yaitu N6 (Chu 1978) untuk induksi kalus dan MS (Murashige dan Skoog 1962) untuk regenerasi dan perakaran yang masing-masing diberi NAA dan Kinetin. Jenis poliamin yang digunakan adalah Putresin 10-3 M dan sukrosa yang ditambahkan pada media induksi kalus dan media regenerasi. Zat pengatur tumbuh lainnya yaitu IBA, Bahan untuk sterilisasi malai menggunakan 20% Clorox dan air steril. Bahan untuk sterilisasi alat yaitu alkohol 90%, dan alkohol 70%. Phytagel TM 3 g L-1, media disesuaikan pada pH 5.8 digunakan sebagai pemadat.
Percobaan kultur antera dilakukan dengan menggunakan rancangan acak lengkap dengan 14 ulangan. Perlakuan yang digunakan adalah 6 genotipe padi yang terdiri atas enam genotipe F1. Setiap satuan percobaan merupakan satu cawan petri yang berisi antera dari 25 buah bulir bunga (spikelet) dari satu tanaman yang berasal dari satu genotipe F1. Sumber eksplan antera adalah enam genotipe F1, yang ditanam dalam pot masing-masing genotipe terdiri atas 20–30 tanaman, dengan 3 tanaman per pot. Pelaksanaan kultur antera mengikuti metode Dewi et al. (2004).
a. Pembuatan media
Media induksi kalus adalah media N6 yang ditambahkan 2.0 NAA mg L-1 dan 0.5 kinetin mg L-1, sedangkan media regenerasi kalus adalah media MS yang ditambahkan 0.5 NAA mg L-1 dan 2.0 kinetin mg L-1. Putresin 10-3 M dan sukrosa berturut-turut sebanyak 60 g L-1 dan 40 g L-1 ditambahkan ke dalam media induksi kalus dan media regenerasi. Media perakaran adalah media MS ditambah 0.5 IBA mg L-1dan 30 sukrosa g L-1. Pemadat yang digunakan adalah agar phytagel TM 3 g L-1 dengan pH media 5.8. Media disterilisasi dalam autoklaf selama 20 menit pada suhu 120 0C tekanan 18-20 psi.
b. Pemilihan dan inkubasi eksplan
Malai yang diambil anteranya adalah malai yang masih dalam keadaan bunting dengan jarak aurikel daun bendera dengan aurikel daun di bawahnya 7–10 cm. Malai yang masih terselubung dicuci bersih kemudian dibungkus dengan kertas tissue yang telah dibasahi. Selanjutnya malai disimpan dalam ruang dingin bersuhu 50C selama 7-10 hari. Perlakuan suhu dingin berguna untuk menyeragamkan stadia polen sehingga lebih banyak polen pada stadia uninukleat yang dapat digunakan.
c. Sterilisasi eksplan
21 d. Penanaman atau inokulasi eksplan
Spikelet–spikelet tersebut diketuk-ketukkan menggunakan pinset pada cawan petri yang berisi 25 mL media induksi kalus sehingga antera akan keluar dan jatuh ke media. Setiap cawan petri berisi antera yang berasal dari 25-30 spikelet atau berisi ± 150 butir antera. Kegiatan ini dilakukan di LAF.
e. Inkubasi kultur antera
Antera diinkubasi dalam ruang gelap bersuhu 25±2 0C yang bertujuan untuk menginduksi keluarnya kalus yang berasal dari butir sari di dalam antera. Kalus biasanya muncul sekitar 3-4 minggu setelah inokulasi.
f. Regenerasi tanaman dari kalus
Kalus yang memiliki tekstur kompak berukuran 1–2 mm dipindahkan ke dalam botol yang berisi 25 mL media regenerasi untuk merangsang keluarnya tunas. Tanaman hijau (planlet) yang tumbuh mencapai 3–5 cm pada media regenerasi dipindahkan ke tabung kultur yang berisi 15 mL media perakaran. Kultur ditempatkan dalam kondisi terang (cahaya 1600– 1800 lux) dengan suhu 25±2 0C selama regenerasi dan perakaran. Sekelompok (cluster) tanaman yang tumbuh dari satu kalus tidak dipisahkan. Setelah akar tumbuh sempurna, maka tanaman siap untuk diaklimatisasi.
g. Aklimatisasi
Planlet hasil kultur antera diaklimatisasi dalam tabung reaksi berisi air steril selama ±1 minggu. Selanjutnya tanaman dipindahkan ke dalam bak semai berisi tanah berlumpur selama 1 minggu. Tanaman diperlakukan pada kondisi cahaya dengan intensitas yang secara bertahap meningkat selama proses aklimatisasi. Hal ini bertujuan supaya tanaman mampu beradaptasi dengan kondisi lapangan. Bibit padi hasil kultur antera dipindahkan dari bak ke pot di rumah kaca dan dipelihara sampai tanaman menghasilkan biji.
h. Pengamatan
Pengamatan dilakukan terhadap jumlah antera dalam petri, jumlah antera menghasilkan kalus, jumlah kalus yang terbentuk, jumlah kalus yang menghasilkan tanaman, jumlah tanaman hijau, jumlah tanaman albino, dan jumlah galur dihaploid yang dihasilkan.
i. Analisis Data